

Abstract
Repeated cold stress (RCS) is known to transiently induce functional disorders associated with hypotension and hyperalgesia. In this study, we investigated the effects of RCS (24 and 4 °C alternately at 30-min intervals during the day and 4 °C at night for 2 days, followed by 4 °C on the next 2 consecutive nights) on the thresholds for cutaneous mechanical pain responses and on peripheral expression of “pain-related genes” in SHRSP5/Dmcr rats, which are derived from stroke-prone spontaneously hypertensive rats. To define genes peripherally regulated by RCS, we detected changes in the expression of pain-related genes in dorsal root ganglion cells by PCR-based cDNA subtraction analysis or DNA microarray analysis, and confirmed the changes by RT-PCR. We found significantly changed expression in eight pain-related genes (upregulated: Fyn, St8sia1, and Tac 1; downregulated: Ctsb, Fstl1, Itpr1, Npy, S100a10). At least some of these genes may play key roles in hyperalgesia induced by RCS.
Keywords: Repeated cold stress, Mechanical hyperalgesia, Peripheral pain mechanisms, Pain-related genes
Introduction
When repeatedly exposed to a cold environment, animals show functional disorders associated with hypotension [1, 2] and facilitated responses to noxious stimuli, namely, transient decreases in the nociceptive threshold to chemical, heat and pressure stimuli [3–7]. This stress model, called repeated cold stress (RCS) or specific alteration of rhythm in environmental temperature (SART) stress, is considered to be a chronic pain model. The hyperalgesia in this model lasts for several days after cessation of RCS [6, 7]. This hyperalgesia is thought to be induced by hypofunction of endogenous pain inhibitory systems that modulate pain sensation in the central nervous system [8]. However, some peripheral mechanisms may also be responsible for this hyperalgesia. Studies focusing on peripheral regulation of pain are needed to clarify the mechanisms underlying the hyperalgesia induced by RCS.
Spontaneously hypertensive rats (SHR) and stroke-prone spontaneously hypertensive rats (SHRSP) are established as parallel lines from outbred Wistar-Kyoto rats (WKY). SHR is reported to be more insensitive to mechanical pain than WKY [9]. SHRSP5/Dmcr rats are a substrain of SHRSP [10], so we expected that obvious hyperalgesia would be induced by RCS in SHRSP5/Dmcr rats, owing to their high pain threshold.
In this study, we investigated the effects of RCS on the thresholds of cutaneous mechanical pain responses and on changes in expression of pain-related genes in dorsal root ganglion cells (DRGs) in SHRSP5/Dmcr rats. We focused on the “pain-related genes” contained in the Pain Genes Database [11]. To determine the gene expression changes accompanied by RCS, we performed PCR-based cDNA subtraction, “suppression subtractive hybridization” and DNA microarray analysis of mRNAs from DRGs of SHRSP5/Dmcr rats. The gene expression changes detected by these analyses were confirmed by reverse transcription polymerase chain reaction (RT-PCR).
There are no reported investigations of pain responses in SHRSP5/Dmcr rats. Thus, we compared the cutaneous mechanical pain thresholds and blood pressures of SHRSP5/Dmcr rats with those of Wistar and Sprague-Dawley (SD) rats.
A preliminary account of this experiment has been reported elsewhere [12].
Materials and methods
Animals
Adult male SHRSP5/Dmcr rats (10–24 weeks old at the start of the experiments, n = 28), adult male Slc:Wistar rats (8–15 weeks old, n = 8) and adult male SD rats (7–14 weeks old, n = 8) were used. They were kept on a 12-h alternating light-dark cycle (light between 8:00 and 20:00). The animals were housed in groups of three at 24 °C with food and water available ad libitum. All rats were fed with a normal diet (CE-2, CLEA Japan, Inc.) throughout the experiment. All animal procedures were approved by the Experimental Animal Research Committee of Kinjo Gakuin University and conducted in accordance with its Animal Experiment Guidelines.
Measurement of pain threshold
Animals were placed in wire-bottom cages, allowing access to the underside of their paws. The cutaneous mechanical pain threshold was measured by the hind paw withdrawal response to the von Frey hair (VFH) test. VFHs (diameter: 0.5 mm) of different strengths were applied in ascending order of force (bending forces 21.9–348 g/mm2 in quasi-logarithmic order). Each VFH was applied five times perpendicularly to the mid-plantar surface of the one hind paw for 1–2 s, and the number of withdrawal responses immediately upon application of the stimulus was noted. The pain threshold was defined as the minimum force at which the animal showed at least four withdrawal responses in response to five stimuli.
Repeated cold stress exposure
RCS was applied to SHRSP5/Dmcr rats according to a modified method, reducing one cold night, based upon the protocol described by Satoh et al. [6]. Briefly, the rats were kept at 4 °C from 16:30 on the first day (day 0) to 10:00 the following morning and then alternately exposed to room temperature (24 °C) and cold temperature 4 °C at 30-min intervals from 10:00 to 16:30 (day 1). These procedures were repeated on day 2. From the evening of day 2 until the morning of day 4, the rats were exposed to 4 °C between 16:30 and 10:00 (Fig. 1). The stress exposures were carried out in separate rooms. Initially, we observed the effects of RCS on the pain threshold and blood pressure 2 or 3 days a week from 7 days before to 18 days after starting RCS (n = 4) (long-term follow-up study). Then we observed the effects of RCS on the pain threshold and blood pressure from 7 days before to 4 days after starting RCS (n = 12) and conducted a comparison with an age-matched control rat group kept at 24 °C throughout the experiment (n = 12) (short-term follow-up study). To blind the evaluators as to which group an animal belonged to in the short-term follow-up study, control rats were mixed with the RCS rat group and measured at the same time.
Fig. 1.
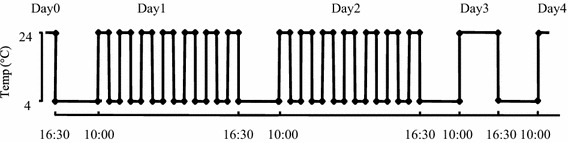
Schematic representation of RCS exposure protocols. Rats were kept at 4 °C from 16:30 on the first day (day 0) to 10:00 the following morning and then alternately exposed to room temperature (24 °C) and cold temperature 4 °C at 30-min intervals from 10:00 to 16:30 (day 1). These procedures were repeated on day 2. From the evening of day 2 until the morning of day 4, the rats were exposed to 4 °C between 16:30 and 10:00. The pain threshold and blood pressure were measured before the start of RCS stress and at day 3 and day 4 after RCS
Measurement of blood pressure
Blood pressure was measured using an indirect blood pressure meter (BP-98A, Softron, Japan) according to the tail pulse method. Changes in systolic blood pressure appeared to be in parallel directions with diastolic blood pressure. Calculated mean arterial blood pressure (MBP) data were recorded.
Tissue dissection and extraction of total RNAs
At the end of the short-term follow-up study, the rats were deeply anesthetized with sodium pentobarbital (100 mg/kg, i.p.) and transcardially perfused with diethylpyrocarbonate (DEPC)-treated saline. The DRGs (L3–L6) were dissected. These tissues were immediately frozen in liquid nitrogen and stored at −80 °C until RNA extraction.
Total RNAs were extracted from DRGs with RNeasy™ Plus Mini Kit (Qiagen, Germany). RNA purity was routinely assessed by measuring the 260/280 nm ratio of the sample. These samples were analyzed by DNA microarray analysis, PCR-based cDNA subtraction and RT-PCR.
Pain Genes Database
We focused on the “pain-related genes” after filtering against the Pain Genes Database [11], which is a comprehensive catalog of genes implicated in pain sensitivity through mouse transgenic knockout studies. The database contains over 250 pain-related genes. The data of PCR-based cDNA subtraction and DNA microarray analysis were filtered using this database.
PCR-based cDNA subtraction
The total RNA samples (1 μg each) taken from two animals’ DRGs in the control group and the RCS group, respectively, were combined. The first-strand cDNAs were synthesized using a SMARTer™ PCR cDNA Synthesis Kit (Clontech, CA, USA). PCR-based cDNA subtraction was conducted using a PCR-Select™ cDNA Subtraction Kit (Clontech, CA, USA), which is based on the suppressive subtractive hybridization method. The suppressive hybridization method is able to detect small changes by the normalization of the abundance of cDNAs within the target population and the subtraction of the common sequences between the test and control populations. The unhybridized sequences were amplified by PCR and cloned into the vector pGem-T Easy™ (Promega, WI, USA). The resultant plasmids were transformed into Escherichia coli strain JM109 and amplified, then sequenced. Sequencing primers were as follows: (sense) M13FW, 5′-GTA AAA CGA CGG CCA GT-3′; (antisense) M13RV, 5′-GGA AAC AGC TAT GAC CAT G-3′.
DNA microarray analysis
The total RNA samples (2 μg each) taken from two animals' DRGs of the control group and the RCS group, respectively, were combined. The cDNA microarray assay was done by Filgen Inc. (Aichi, Japan). The assays were performed on the platform of CodeLink™ Rat Whole Genome Bioarray (Applied Microarray, Inc. AZ, USA), in which 33,849 oligonucleotide probes were spotted. The Pain Genes Database included 251 genes when we searched it. After excluding overlapping genes, the number of genes was 128. This microarray analysis could detect 113 out of 128 pain-related genes. The expression level of each gene was compared between the RCS and control groups. Differences were considered significant if the changes were smaller than four-tenths or greater than two-fold.
RT-PCR
The target mRNAs of RT-PCRs were those with expression changes detected by PCR-based cDNA subtraction or cDNA microarrays and were selected with the Pain Genes Database. Total RNAs (1 μg) were reverse transcribed using oligo (dT)16 primer and M-MuLV reverse transcriptase (NEB, MA, USA), and the transcripts were amplified using sequence-specific primers for the genes and Taq DNA polymerase (NEB, MA, USA). Potions of the PCR product mixtures were electrophoresed in an agarose gel and stained with ethidium bromide. The densities of the bands were quantified using the Image J image analysis software (version 1.47d) [13]. The expression levels were normalized to the expression levels of cyclophilin A (CypA, a housekeeping gene) measured as an internal standard in each sample. Subsequently, amplicons were digested with restriction enzymes, and the size of fragments was checked (data not shown). The sequences of PCR primers, PCR condition and product sizes are shown in Table 1.
Table 1.
PCR primers, conditions and sizes of PCR products
| Gene name | Protein name | Accession no. | Primer | Temp (°C)a | Cycleb | Size (bp)c |
|---|---|---|---|---|---|---|
| Sense Antisense | ||||||
| Ctsb | CathepsinB | BC072490 |
5’-GAAGCCATGTCTGACCGAAT-3’ 5’-CCACTGGGCCATTTTTGTAG-3’ |
60 | 28 | 406 |
| Fstl1 | Follistatin-like 1 | NM_024369 |
5’-GCCATGGAGACCTAACCAAA-3’ 5’-TGAGGGACCTCTTGTGCTTT-3’ |
56 | 32 | 386 |
| Fyn | Src kinase p59 | NM_012755 |
5’-GCTTGGATAATGGGCTGTGT-3’ 5’-CATCTTCTGTCCGTGCTTCA-3’ |
60 | 34 | 306 |
| Grin2b | Glutamate ionotropic receptor | NM_012574 |
5’-CCCAAGTTCTGGTTGGTGTT-3’ 5’-GATCTGAGCGATGGCTTCTT-3’ |
62 | 40 | 303 |
| Itpr1 | Inositol1,4,5-triphosphate | NM_001007235 |
5’-GGTTGTGAACTTAGCCAGGAAC-3’ 5’-CTGGTACATTACTTGGCCCTTC-3’ |
58 | 36 | 481 |
| Map2k1 | Mitogen-activated protein kinase | NM_031643 |
5’-GCAAGATGCCCAAGAAGAAG-3’ 5’-GATGATCTGGTTCCGGATTG-3’ |
60 | 34 | 339 |
| Npy | Neuropeptide Y | NM_012614 |
5’-GCTAGGTAACAAACGAATGGGG-3’ 5’-CACATGGAAGGGTCTTCAAGC-3’ |
55 | 37 | 288 |
| S100a10 | Protein S100-A10 (p11) | BC091565 |
5’-TGCTCATGGAAAGGGAGTTC-3’ 5’-GCTTGTCGAATTGGAGTTGG-3’ |
60 | 30 | 330 |
| St8sia1 | GD3 synthase | NM_012813 |
5’-GGAAAGCCAAAGGTGTGTGT-3’ 5’-GGTTGCAGCAGTCTTCCATT-3’ |
56 | 34 | 373 |
| Tac 1 | Substance P | NM_012666 |
5’-CGAGCAGTGAGCGGTAAAAT-3’ 5’-CGCAAGACACACAGGAGTTT-3’ |
60 | 27 | 305 |
| CypA | Peptidylprolyl isomerase A (cyclophilin A) (internal standard) | BC059141 |
5’- ACGCCGCTGTCTCTTTTC -3’ 5’- TGCCTTCTTTCACCTTCC -3’ |
55 | 24 | 440 |
aTemp: annealing temperature
bCycle: PCR cycles
cSize: sizes of PCR products
Statistical analysis
The data are presented as mean ± SE and were analyzed using the following statistical analyses or tests: Student’s t test for unpaired observations, one-way repeated measures analysis of variance (ANOVA) followed by Bonferroni’s multiple comparison test or Tukey’s multiple comparison test, and two-way repeated measures ANOVA followed by Tukey’s multiple comparison test. Differences were considered significant if p < 0.05.
Results
Comparison of cutaneous mechanical pain thresholds and blood pressures of SHRSP5/Dmcr rats with those of Wistar and SD rats
The mechanical pain thresholds and blood pressures of SHRSP5/Dmcr rats were compared with those of Wistar and SD rats (Fig. 2). The mechanical pain thresholds of SHRSP5/Dmcr rats (183.9 ± 18.5 g/mm2) were significantly higher than those of Wistar rats (82.6 ± 15.6 g/mm2) (p < 0.05 by one-way repeated measures ANOVA followed by Tukey’s multiple comparison test) and similar to those of SD rats (179.4 ± 20.6 g/mm2). On the other hand, the mean blood pressures of SHRSP5/Dmcr rats (160.5 ± 2.6 mmHg) were significantly higher than those of Wistar (103.5 ± 2.2 mmHg) and SD rats (111.3 ± 1.93 mmHg) (p < 0.05 by one-way repeated measures ANOVA followed by Tukey’s multiple comparison test). We found that both the pain threshold and blood pressure of SHRSP5/Dmcr rats were high.
Fig. 2.

Pain thresholds and blood pressures of Slc:Wistar, SD and SHRSP5/Dmcr rats. Each column and bar represent the mean and SE (n = 8, respectively). *p < 0.05 between strains by one-way repeated measures ANOVA followed by Tukey’s multiple comparison test
Effects of repeated cold stress on the cutaneous mechanical pain threshold and blood pressure
In the long-term follow-up study, the exposure of SHRSP5/Dmcr rats to RCS markedly decreased the mechanical pain threshold and blood pressure (n = 4). Compared with day 0, the pain threshold was significantly decreased at day 4 and day 7 (p < 0.001 by one-way repeated measures ANOVA followed by Bonferroni’s multiple comparison test), while the blood pressure was significantly decreased at day 4 (p < 0.01 by one-way repeated measures ANOVA followed by Bonferroni’s multiple comparison test) (Fig. 3). The recovery of the RCS-induced decrease in pain threshold was slower than that of blood pressure. The greatest decreases in the pain threshold and blood pressure were seen at day 4. Afterwards, the pain thresholds and blood pressure gradually returned to near the baseline level.
Fig. 3.

Changes in the pain threshold and blood pressure by RCS in the long-term follow-up study. The pain threshold (solid circles) and blood pressure (open circles) were measured from 7 days before to 18 days after the start of the RCS, 2 or 3 days a week (n = 4). Obvious decreases in the pain threshold and blood pressure were induced by RCS. The recovery rate of the pain threshold was slower than that of blood pressure. *p < 0.05, **p < 0.01, ***p < 0.001 vs. pre-RCS (day 0) by one-way repeated measures ANOVA followed by Bonferroni’s multiple comparison test
To investigate changes in gene expression induced by RCS, we performed a short-term follow-up study in which we observed the effect of RCS on the pain threshold up to day 4, after which the DRGs were dissected. In this short-term follow-up study, the pain thresholds were significantly decreased at day 3 and day 4 compared with the control (p < 0.05 vs. corresponding control by two-way repeated measures ANOVA followed by Tukey’s multiple comparison test, n = 12) (Fig. 4).
Fig. 4.
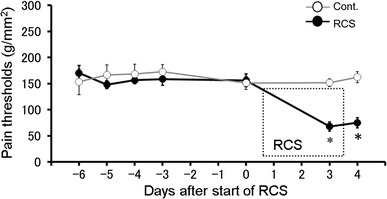
Changes in pain threshold from RCS in the short-term follow-up study. The pain threshold was measured from 6 days before to 4 days after the start of RCS (solid circles) and compared with the controls (non-RCS, open circles) (n = 12, respectively). Obvious decreases in pain threshold were induced by RCS. Each circle and bar represent the mean and SE. *p < 0.05 vs. corresponding control by two-way repeated measures ANOVA followed by Tukey’s multiple comparison test
PCR-based cDNA subtraction
We compiled both up- and downregulated cDNA sequences in the DRGs of RCS rats relative to those in the DRGs of control rats. The number of up- and downregulated genes in the RCS rats was 20 and 44, respectively. In the upregulated group, there was one known pain-related gene, Tac 1 (substance P), among the 20 upregulated genes. In the downregulated group, there were three known pain-related genes, Ctsb (cathepsin B), Fstl1 (follistatin-like 1) and S100a10 (annexin 2 light), among the 44 downregulated genes. These genes are listed in Table 2.
Table 2.
RT-PCR of the pain-related genes whose expression changes were detected by PCR-based cDNA subtraction or DNA microarray analysis
| Gene name | Protein name | Assay | Direction | RT-PCR | |
|---|---|---|---|---|---|
| (Fold of increase or decrease) | Direction | Fold of changes | |||
| Ctsb | CathepsinB | Subtraction | Down | Down | 0.9 |
| Fstl1 | Follistatin-like 1 | Subtraction | Down | Down | 0.8 |
| Fyn | Src kinase p59 | Microarray | Up (2.7) | Up | 1.7 |
| Grin2b | Glutamate ionotropic receptor | Microarray | Up (2.6) | non | – |
| Itpr1 | Inositol 1,4,5-triphosphate receptor, type 1 | Microarray | Down (0.5) | Down | 0.6 |
| Map2k1 | Mitogen-activated protein kinase | Microarray | Up (2.6) | non | – |
| Npy | Neuropeptide Y | Microarray | Down (0.5) | Down | 0.7 |
| S100a10 | Annexin 2 light | Subtraction | Down | Down | 0.5 |
| St8sia1 | GD3 synthase | Microarray | Up (2.4) | Up | 1.3 |
| Tac 1 | Substance P | Subtraction | Up | Up | 3 |
Microarray, DNA microarray analysis; subtraction, PCR-based cDNA subtraction
DNA microarray analysis
Using DNA microarray analysis, we detected changes in gene expression to identify genes affected by RCS. We compiled both up- and downregulated cDNA sequences in the DRGs of RCS rats that showed greater than twofold changes in expression relative to those of DRGs of control rats. The number of up- and downregulated genes in the RCS rats was 381 and 314, respectively. In the upregulated genes, there were four pain-related genes, Fyn (Src kinase p59), Grin2b (glutamate ionotropic receptor), Map2k1 (mitogen-activated protein kinase) and St8sia1 (GD3 synthase). In the downregulated group, there were two pain-related genes, Itpr1 (inositol 1, 4, 5-triphosphate) and Npy (neuropeptide Y). These genes are listed in Table 2. The changes in expression of Ctsb, Fstl1, S100a10 and Tac 1 were not significant in this analysis with changes detected by PCR-based cDNA subtraction.
Changes in gene expression levels detected by RT-PCR
PCR-based cDNA subtraction analysis and DNA microarray analysis detected up- or downregulation of ten pain-related genes (Ctsb, Fstl1, Fyn, Grin2b, Itpr1, Map2k1, Npy, S100a10, St8sia1 and Tac 1). Among these genes, the expression changes were confirmed by RT-PCR. The expression levels of Fyn, St8sia1 and Tac 1 at day 4 were significantly increased, and those of Ctsb, Fstl1, Itpr1, Npy and S100a10 were significantly decreased, as shown in Fig. 5 (p < 0.05, p < 0.01 or p < 0.001 vs. control by Student’s t test, n = 12, respectively). The fold changes of gene expression in Grin2b and Map2k1 were 0.91 ± 0.12 and 1.03 ± 0.09, respectively, and these differences were not significant.
Fig. 5.
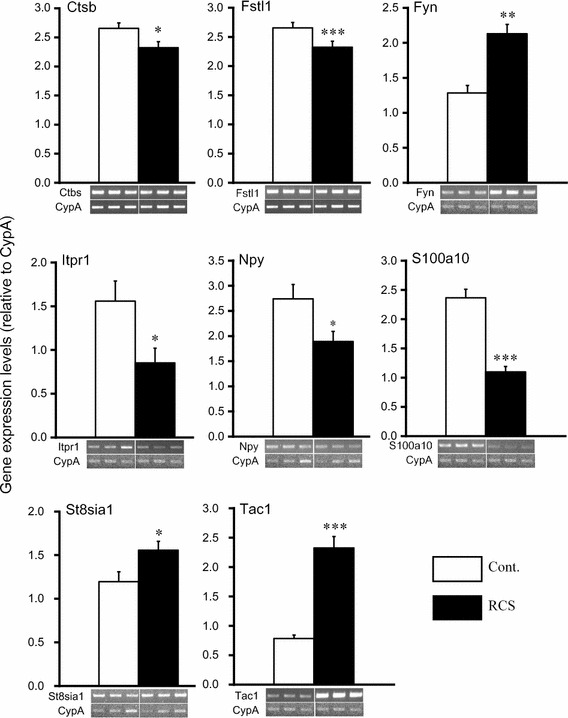
Changes in gene expression of pain-related genes in DRGs induced by RCS. Analysis of the pain-related gene expression levels in the control group (white bars) and RCS group (black bars) was performed using RT-PCR. Significant changes in the expression of eight pain-related genes were observed. The values were normalized by the level of CypA. Each column and bar represent the mean and SE (n = 12, respectively). *p < 0.05, **p < 0.01 and ***p < 0.001 vs. control by Student’s t test. Three representative images of each RT-PCR amplification in the control (left hand side) and RCS group (right hand side) are also shown, respectively
Discussion
The pain thresholds in SHRSP5/Dmcr rats were similar to those in SD rats, while the mean blood pressures in SHRSP5/Dmcr rats were significantly higher than those in SD rats. It is thought that there is no direct relationship between pain thresholds and blood pressures. In SHRSP5/Dmcr rats hyperalgesia was also shown to be induced by RCS in a manner similar to that in SD rats [6]. SHRSP5/Dmcr rats were shown to develop hypercholesterolemia in response to a high-fat diet over 2 weeks [10]. However, in this study, all SHRSP5/Dmcr rats were fed a normal diet before and during the experiment. We consider the pain response in SHRSP5/Dmcr rats fed a normal diet to be similar to that in SD rats. We tried to apply RCS to SHRSP5/Dmcr rats under the modified condition reported by Satoh et al. As expected, obvious hyperalgesia and hypotension were induced by RCS in SHRSP5/Dmcr rats.
RCS is considered to be a chronic pain model caused by imbalance in the autonomic nervous system, specifically vagotonia or decreased activity of the sympathetic nervous system [4, 14]. This hyperalgesia is thought to be induced by hypofunction of endogenous pain inhibitory systems, which modulate the pain sensation in the central nervous system [8]. The central nervous system functions associated with “pain perception and processing” are complex [15, 16]; therefore, many regions, including the hypothalamus, are speculated to be involved in hyperalgesia from RCS. However, some peripheral mechanisms may be also responsible for this hyperalgesia. In this study, we focused on RCS-induced peripheral regulation of pain sensation and analyzed several pain-related genes in DRGs using cDNA subtraction and DNA microarray analysis. We observed for the first time changes in the expression of pain-related genes induced by RCS in DRG using DNA microarray and/or cDNA subtraction analysis.
RCS model animals have been produced by changing the temperature from 24 to 4 °C (RCS at 4 °C) in ddY mice [17] and from 24 to −3 °C (RCS at −3 °C) in Wistar rats [1]. Mechanical hyperalgesia has been induced by RCS in Wistar rats [5] and in SD rats (RCS at 4 °C) [6, 7]. In this study, we observed obvious hyperalgesia and hypotension in SHRSP5/Dmcr rats after the end of RCS, which was alternately 24 and 4 °C at 30-min intervals during the day and 4 °C at night for 2 days, followed by 4 °C on 2 consecutive nights.
Immediately after RCS, the mechanical pain threshold was significantly decreased. Obvious decreases in pain threshold and blood pressure were induced by RCS. Afterwards, the pain threshold and blood pressure gradually returned to near the baseline level. The recovery rate of the pain threshold was slower than that of blood pressure. Pain sensation might be strongly influenced by RCS.
PCR-based cDNA subtraction is able to detect even slight changes in gene expression, but is not quantitative. Using PCR-based cDNA subtraction and RT-PCR, we detected upregulation of Tac 1 (substance P) and downregulation of Ctsb (cathepsin B), Fstl1 (follistatin-like 1) and S100a10 (annexin 2 light) (Table 2; Fig. 4).
Substance P, coded by Tac 1, is known to be released from both central and peripheral terminals of primary afferent sensory neurons, transmitting nociceptive information. Satoh et al. [6] suggested that enhanced synaptic transmission mediated by substance P and calcitonin gene-related peptide in the spinal dorsal horn is, at least in part, involved in hyperalgesia due to RCS. In this study, changes in the expression of calcitonin gene-related peptide were not detected.
Follistatin-like 1, coded by Fstl1, is secreted from sensory afferent axons and activates presynaptic Na+/K+-ATPase, leading to membrane hyperpolarization. Deletion of Fstl1 in DRG neurons induces enhanced afferent synaptic transmission and sensory hypersensitivity [18]. Fstl1 plays an important role in the suppression of nociceptive afferent transmission and its reduction in primary afferent neurons, resulting in pain hypersensitivity.
Cathepsin B, coded by Ctsb, is thought to be an essential enzyme for the induction of chronic inflammatory pain, inducing the maturation and secretion of IL-1β and IL-18 by spinal microglia [19]. The reduced expression of Ctsb may provide a protective adaptation to RCS.
S100a10, coding annexin 2 light chain, is involved in the trafficking of the voltage-gated sodium channel Nav1.8, which is a sensory neuron-specific voltage-gated sodium channel [20, 21]. Reduced expression of S100a10 may therefore also provide a protective adaptation to RCS.
Furthermore, using DNA microarray analysis and RT-PCR, we detected upregulation of St8sia1 (GD3 synthase) and Fyn (Src kinase p59) and downregulation of Itpr1 (inositol 1,4,5-triphosphate) and Npy (neuropeptide Y) (Table 2; Fig. 4).
Src-family protein tyrosine kinases coded by the Fyn phosphorylate NR2B subunit of the NMDA receptor increase synaptic currents [22], leading to central sensitization in the spinal dorsal horn.
Knockdown of NPY produces rapid and reversible increases in the intensity and duration of tactile and thermal hypersensitivity [23]. NPY is thought to inhibit the spinal transmission of sensory signals. RCS induces a decrease in NPY expression in DRG and subsequent excitation of DRG, leading to hyperalgesia.
Inositol 1,4,5-triphosphate receptor (IP3R1) coded by Itpr1 is a specific intracellular calcium release channel protein, which is observed in the central and peripheral nervous systems [24]. Increased calcium ion influx induces hyperexcitability of nociceptive sensory neurons. Calcium released from IP3R1-gated stores may participate in modulating presynaptic function [25], and reduced expression of IP3R1 may induce hyperalgesia.
GD3 synthase, coded by St8sia1, produces ganglioside, which is important as a component of lipid rafts and serves as a key for cell-cell interactions and signal transduction complexes in peripheral nociceptor endings [26].
LaCroix-Fralish et al. [27] performed a meta-analysis of 20 microarray studies of neuropathic or inflammatory pain. They found that Reg3b (regenerating islet-derived 3 beta; pancreatitis-associated protein) and Ccl2 (chemokine [C–C motif] ligand 2) were significantly upregulated in every neuropathic or inflammatory pain condition in the rat and proposed these two genes as potential biomarkers of chronic pain. Of these two genes, only Ccl2 is listed in the Pain Genes Database [11]. In this study, there was no significant change in expression of Ccl2 with RCS. Here, we look at Tac 1 and Npy, which were up- and downregulated, respectively, by RCS in this study. In LaCroix-Fralish’s paper, Tac 1 and Npy, along with many other genes, are listed as being upregulated mainly by neuropathy. That is, Tac 1 was upregulated by RCS or neuropathy, while Npy was downregulated by RCS and upregulated by neuropathy. It is thought that participating pain-related genes depend on the type of pain. Fyn, St8sia1, Ctsb, Fstl1, Itpr1 and S100a10, which were up-/downregulated by RCS, are not listed in LaCroix-Fralish’s paper.
Some genes are reported to be differentially expressed between different strains (SHR and WKY), and even between substrains from different sources [28]. Therefore, gene expression profiles in SHRSP5/Dmcr rats may be different from those in SD rats and WKY rats. In the present study, we revealed that Fyn, St8sia1 and Tac 1 were upregulated and that Ctsb, Fstl1, Itpr1, Npy and S100a10 were downregulated by RCS in SHRSP5/Dmcr rats. We conclude that hyperalgesia induced by RCS is peripherally regulated by at least some of these genes, coordinated through modulation of signal transduction in peripheral nociceptor endings and synaptic transmission. Some of them may also be related to protective adaptation mechanisms to RCS.
Acknowledgments
This work was supported in part by a Grant-in-Aid (23590724) for Scientific Research (C) from the Ministry of Education, Science, Sports, and Culture, Japan.
Conflict of interest
There is no conflict of interest.
References
- 1.Kita T, Hata T, Yoneda R, Okage T. Stress state caused by alteration of rhythm in environmental temperature, and the functional disorders in mice and rats. Folia Pharmacol Jpn. 1975;71:195–210. doi: 10.1254/fpj.71.195. [DOI] [PubMed] [Google Scholar]
- 2.Hata T, Kita T, Namimatsu A, Ito E. Changes of blood pressure and regional blood flow in SART rat and drug actions on these changes. Folia Pharmacol Jpn. 1982;79:335–342. doi: 10.1254/fpj.79.335. [DOI] [PubMed] [Google Scholar]
- 3.Kita T, Hata T, Iida J, Yoneda R, Isida S. Decrease in pain threshold in SART stressed mice. Jpn J Pharmacol. 1979;29:479–482. doi: 10.1254/jjp.29.479. [DOI] [PubMed] [Google Scholar]
- 4.Hata T, Kita T, Itoh E, Kawabata A. The relationship of hyperalgesia in SART (repeated cold)-stressed animals to the autonomic nervous system. J Auton Pharmac. 1988;8:45–52. doi: 10.1111/j.1474-8673.1988.tb00168.x. [DOI] [PubMed] [Google Scholar]
- 5.Kawanishi C, Fukuda M, Tamura R, Nishijo H, Ono T. Effects of repeated cold stress on feeding, avoidance behavior, and pain-related nerve fiber activity. Physiol Behav. 1997;62:849–855. doi: 10.1016/S0031-9384(97)00248-5. [DOI] [PubMed] [Google Scholar]
- 6.Satoh M, Kuraishi Y, Kawamura M. Effects of intrathecal antibodies to substance P, calcitonin gene-related peptide and galanin on repeated cold stress-induced hyperalgesia: comparison with carrageenan-induced hyperalgesia. Pain. 1992;49:273–278. doi: 10.1016/0304-3959(92)90151-Z. [DOI] [PubMed] [Google Scholar]
- 7.Nasu T, Taguchi T, Mizumura K. Persistent deep mechanical hyperalgesia induced by repeated cold stress in rats. Eur J Pain. 2010;14:236–244. doi: 10.1016/j.ejpain.2009.05.009. [DOI] [PubMed] [Google Scholar]
- 8.Ohara H, Kawamura M, Namimatsu A, Miura T, Yoneda R, Hata T. Mechanism of hyperalgesia in SART stressed (repeated cold stress) mice: antinociceptive effect of neurotropin. Jpn J Pharmacol. 1991;57:243–250. doi: 10.1254/jjp.57.243. [DOI] [PubMed] [Google Scholar]
- 9.Taylor BK, Roderick RE, Lezin EST, Basbaum AI. Hypoalgesia and hyperalgesia with inherited hypertension in the rat. Am J Physiol Regulatory Integrative Comp Physiol. 2001;280:R345–R354. doi: 10.1152/ajpregu.2001.280.2.R345. [DOI] [PubMed] [Google Scholar]
- 10.Kitamori K, Naito H, Tamada H, Kobayashi M, Miyazawa D, Yasui Y, Sonoda K, Tsuchikura S, Yasui N, Ikeda K, Moriya T, Yamori Y, Nakajima T. Development of novel rat model for high-fat and high-cholesterol diet-induced steatohepatitis and severe fibrosis progression in SHRSP5/Dmcr. Environ Health Prev Med. 2012;17:173–182. doi: 10.1007/s12199-011-0235-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.LaCroix-Fralish ML, Ledoux JB, Mogil JS (2007) The pain genes database: an interactive web browser of pain-related transgenic knockout studies. Pain 131:3.e1–3.e4. http://www.jbldesign.com/jmogil/enter.html. Accessed 8 May 2015 [DOI] [PubMed]
- 12.Kozaki Y, Umetsu R, Mizukami Y, Yamamura A, Kitamori K. Mechanical hyperalgesia induced by repeated cold stress in SHRSP5/Dmcr rats: PCR-based cDNA subtraction analysis for peripheral expression changes in pain related genes. J Physiol Sci. 2014;64(Suppl. 1):S157. [Google Scholar]
- 13.Rasband WS (1997–2012) Image J. US National Institutes of Health, Bethesda, Maryland, USA. http://imagej.nih.gov/ij/. Accessed 8 May 2015
- 14.Kita T, Hata T, Itoh E, Namimatsu A. Testing methods for vegetative syndrome in the rat and effects of neurotropin and other drugs. Shinshin-Igaku. 1983;23:61–68. [Google Scholar]
- 15.Apkarian AV, Bushnell MC, Treede R-D, Zubieta J-K. Human brain mechanisms of pain perception and regulation in health and disease. Eur J Pain. 2005;9:463–484. doi: 10.1016/j.ejpain.2004.11.001. [DOI] [PubMed] [Google Scholar]
- 16.Latremoliere A, Woolf CJ. Central sensitization: a generator of pain hypersensitivity by central neurol plasticity. J Pain. 2009;10:895–926. doi: 10.1016/j.jpain.2009.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hata T, Kita T, Itoh E, Harada N. Experimental studies on optimal conditions of loading SART stress (repeated cold stress) upon animals. Shinshin-Igaku. 1984;24:257–266. [Google Scholar]
- 18.Li K-C, Zhang F-X, Li C-L, Wang F, Yu M-Y, Zhong Y-Q, Zhang K-H, Lu Y-J, Wang Q, Ma X-L, Yao J-R, Wang J-Y, Lin L-B, Han M, Zhang Y-Q, Kuner R, Xiao H-S, Bao L, Gao X, Zhang X. Follistatin-like 1 suppresses sensory afferent transmission by activating Na+, K+ -ATPase. Neuron. 2011;69:974–987. doi: 10.1016/j.neuron.2011.01.022. [DOI] [PubMed] [Google Scholar]
- 19.Sun L, Wu Z, Hayashi Y, Peters C, Tsuda M, Inoue K, Nakanishi H. Microglial cathepsin B contributes to the initiation of peripheral inflammation-induced chronic pain. J Neurosci. 2012;32:11330–11342. doi: 10.1523/JNEUROSCI.0677-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Okuse K, Mlik-Hall M, Baker MD, Poon W-YL, Kong H, Chao MV, Wood JN. Annexin II light chain regulates sensory neuron-specific sodium channel expression. Nature. 2002;417:653–656. doi: 10.1038/nature00781. [DOI] [PubMed] [Google Scholar]
- 21.Foulkes T, Nassar MA, Lane T, Matthews EA, Baker MD, Gerke V, Okuse K, Dickenson AH, Wood JN. Deletion of annexin 2 light chain p11 in nociceptors causes deficits in somatosensory coding and pain behavior. J Neurosci. 2006;26:10499–10507. doi: 10.1523/JNEUROSCI.1997-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yaka R, He D-Y, Phamluong K, Ron D. Pituitary adenylate cyclase-activating polypeptide (PACAP(1-38)) enhances N-methyl-D-aspartate receptor function and brain-derived neurotrophic factor expression via RACK1. J Biol Chem. 2003;278:9630–9638. doi: 10.1074/jbc.M209141200. [DOI] [PubMed] [Google Scholar]
- 23.Solway B, Bose SC, Corder G, Donahue RR, Taylor BK. Tonic inhibition of chronic pain by neuropeptide Y. Proc Natl Acad Sci USA. 2011;108:7224–7229. doi: 10.1073/pnas.1017719108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dent MAR, Raisman G, Lai A. Expression of type 1 inositol 1,4,5-trisphosphate receptor during axogenesis and synaptic contact in the central and peripheral nervous system of developing rat. Development. 1996;122:1029–1039. doi: 10.1242/dev.122.3.1029. [DOI] [PubMed] [Google Scholar]
- 25.Luo C, Gangadharan V, Bali KK, Xie R-G, Agarwal N, Kurejova M, Tappe-Theodor A, Tegeder I, Feil S, Lewin G, Polgar E, Todd AJ, Schlossmann J, Hofmann F, Liu D-L, Hu S-J, Feil R, Kuner T, Kuner R. Presynaptically localized cyclic GMP-dependent protein kinase 1 is a key determinant of spinal synaptic potentiation and pain hypersensitivity. PLoS Biol. 2012;10:e1001283. doi: 10.1371/journal.pbio.1001283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Watanabe S, Tan-No K, Tadano T, Higashi H. Intraplantar injection of gangliosides produces nociceptive hehavior and hyperalgesia via a glutamate signaling mechanism. Pain. 2011;152:327–334. doi: 10.1016/j.pain.2010.10.036. [DOI] [PubMed] [Google Scholar]
- 27.LaCroix-Fralish ML, Austin J-S, Zheng FY, Levitin DJ, Mogil JS. Patterns of pain: meta-analysis of microarray studies of pain. Pain. 2011;152:1888–1898. doi: 10.1016/j.pain.2011.04.014. [DOI] [PubMed] [Google Scholar]
- 28.Okuda T, Sumiya T, Iwai N, Miyata T. Difference of gene expression profiles in spontaneous hypertensive rats and Wistar-Kyoto rats from two sources. Biochem Biophys Res Commun. 2002;296:537–543. doi: 10.1016/S0006-291X(02)00902-6. [DOI] [PubMed] [Google Scholar]