

Abstract
This investigation was conducted to evaluate the effect of maternal diabetes on fetal testicular structure and function, and reproductive hormones levels. Sixteen female rats were divided into two groups. Diabetes was induced in one group by alloxan. Blood was collected from 90-day-old male offspring of both groups, and the level of blood glucose, testosterone, FSH and LH in their serum was measured. Weight, volume and various histological parameters of testes were determined. A significant increase in blood glucose and decrease in LH, FSH and testosterone in sera of offspring of diabetic mothers (ODM) were observed. The weight and volume of testes in the ODM were 22.7 and 22.9% higher, respectively, than those of the control group (P < 0.05), while the ratio of testes to body weight did not change significantly. The number of seminiferous tubules increased (+21%) significantly (P < 0.05), while thickness of the testicular capsule (−25%), number of Leydig cells (−15.6%), number of Sertoli cells (−14.9%), number of spermatogonia (−26.3%) and diameter of seminiferous tubules (−11%) showed significantly reduced values in the ODM compare to the control. In conclusion, maternal hyperglycemia has a deleterious effect on testicular parameters during fetal life, which will affect reproductive endocrine during postpuberty.
Keywords: Maternal diabetes, Testis, Rat, Histomorphometry, Endocrine
Introduction
In diabetic mothers during pregnancy, placental transport of glucose and other nutrients increases because of an increased availability at the maternal site, resulting in fetal and neonatal macrosomia [1, 2]. The elevated glucose concentration in the mother, accompanying hyperglycemia in the fetus, leads to degranulation of the fetal B-cells, resulting in fetal hypoinsulinaemia [3]. The pancreas of a newborn of a badly controlled diabetic mother (blood glucose >16.7 mmol l−1) shows degranulation of the majority of B-cells [3].
The consequences of diabetes during pregnancy are not confined to fetal and neonatal life. Several epidemiological data show that consequences extend to adult life and even to the next generation through the maternal line. Knowler et al. [4] showed that the risk for diabetes is significantly higher in the offspring of mothers who have non-insulin-dependent diabetes.
Data indicate that pre-gestational maternal diabetes is associated with strong teratogenic effects on the kidney, urinary tract and heart, and is strongly associated with multiple congenital abnormalities [5]. The malformations often involve the heart and central nervous system, and are potentially lethal. Major congenital anomalies are two to four times more common in diabetic pregnancies than in normal pregnancies [6].
One of the mammalian systems that is clearly impaired in diabetes is the male reproductive functioning. Diabetes-induced alterations of Leydig cell functions include a decrease in androgen synthesis and in the total number of these cells. Together, these effects cause an impairment of male libido [7]. Disturbances in sexual function, including a decrease in libido, impotence and infertility, were reported to be due to testicular dysfunction associated with sustained hyperglycemia in diabetic rats [8]. Spontaneously diabetic BB rats also show a decrease in testicular testosterone production, indicating that this process is inherent to this disease. Moreover, this alteration is responsible for the diabetes-related effects on libido [9]. The diabetic BB rat also shows altered spermatogenesis [10]. Lower pregnancy rates have been associated with the basic fertility potential in diabetic men, and variations of semen quality have been reported in these patients [11, 12]. Diabetes-related effects on testicular function have been attributed to the lack of insulin. The regulatory action of this hormone is known, and observations of a direct effect on both Leydig cells [13, 14] and Sertoli cells [15, 16] have been reported. Testicular function is primarily controlled by pituitary hormones. The follicle-stimulating hormone (FSH) regulates spermatogenesis, whereas the luteinizing hormone (LH) controls Leydig cell function. Decreases in the serum levels of FSH, LH, prolactin and growth hormone have been reported in diabetes [17, 18]. The fetuses of diabetic mothers also experience hyperglycemia and hypoinsulinemia, which may have an adverse effect on their reproductive system (endocrine and testicular structure) during fetal development. The aim of this investigation was to evaluate the effect of maternal diabetes on fetal testicular structure and function, and reproductive hormones levels (gonadotropins and testosterone), which may contribute to further complications and infertility.
Materials and methods
Animals
Sixteen adult female Sprague Dawley rats (200–230 g and 4–5 months old) were housed in an air-conditioned room (22 ± 2°C) and supplied with standard pellet food with tap water ad libitum. Animals were divided into two groups, diabetics and normal (control). The animals were cared for and treated in accordance with the guidelines for laboratory animals established by the National Institute of Health as well as by the local ethical committee.
Induction of diabetes mellitus
Diabetes was induced in eight rats by a single intraperitoneal injection of alloxan tetrahydrate (Sigma, St. Louis, MO) (150 mg/kg) according to our previous experience. The animals were fasted 12 h before and after alloxan injection. Rats with blood glucose above 200 mg/dl, as well as with polydipsia, polyuria and polyphagia for at least 1 week, were considered to be diabetic and selected for the experiment.
Experimental design
Female animals of both groups in estrus stage of the reproductive cycle were caged with male rats for mating. Mating was confirmed by observation of a vaginal plug. Male offspring of both groups were reared in similar conditions in an animal house for 90 days. At the end of the experiment, the animals were anesthetized with diethyl ether and killed by whole blood collection through a heart puncture. Blood serum was isolated and used for endocrine analysis. The volume of fresh isolated testes was obtained, using Archimedes' principle, by immersion in distilled water [19]; after that, they were fixed with 10% buffered formalin solution.
Histomorphometric study
Formaldehyde-fixed samples were embedded in paraffin and then sliced (slice thickness, 4–5 μm). They were further deparaffined with xylol, and histologic observations were performed after staining by the hematoxylin–eosin or Green Masson’s trichrome method [20]. For histomorphological and histomorphometric study, the sections were observed under a light microscope, and the following factors were evaluated in testes of both control and test groups: (1) thickness of the testicular capsule, (2) number of seminiferous tubules/mm2, (3) number of Leydig cells/mm2, (4) average number of Sertoli and spermatogonia cells in seminiferous tubules and (5) diameter of seminiferous tubules.
Testicular capsule thickness
Thickness of the testicular capsule was measured at ×100 magnification using Olysia software (Olysia soft imaging system provided by Olympus 2000) and an Olympus BX51 light microscope. At least six points of the capsule sections were chosen randomly and measured for each tests.
Counting of seminiferous tubules
Seminiferous tubules were counted at ×40 magnification using a 441-intersection grid placed in the ocular of the light microscope (Olympus BX51). Ten sections were chosen at random from each testis, and the number of round or nearly round seminiferous tubuli in square millimeters (mm2) was obtained.
Leydig cells
The number of Leydig cells per square millimeter was determined at ×1,000 magnification using a 441-intersection grid placed in the ocular of the light microscope (Olympus BX51). Ten sections were randomly chosen for each testis, the total number of Leydig cell nuclei was counted, and the mean of Leydig cells per square millimeter was scored.
Spermatogonia and sertoli cells
All spermatogonia and Sertoli cells were counted at ×1,000 magnification, and at least ten round or nearly round seminiferous tubule cross-sections were chosen at random for each testis. The total number of spermatogonia and Sertoli cells per seminiferous tubule was determined from the corrected counts of spermatogonia cell nuclei and Sertoli cell nucleoli per seminiferous tubule cross section [21].
Analytic procedures in serum samples
Serum testosterone, FSH and LH levels were measured by radioimmunoassay (RIA) technique, commercial kit (Immunotech-Radiova, Prague, Czech Republic). The intra-assay and inter-assay coefficient for FSH was below or equal to 2.6 and 6.3%, respectively. The intra-assay and inter-assay for LH was below or equal to 3.5 and 8.8%, respectively. For testosterone the intra-assay and inter-assay was below or equal to 14.8 and 15%, respectively.
Statistical analysis
Morphometric data are presented as the mean ± SD, and to analyze these data, Student’s t test was performed using the SPSS program. Significant differences were considered when P < 0.05.
Results
Comparison of body weights
Changes in body weights of newborns in both groups from birth to day 90 of age are presented in Table 1. Significant (P < 0.05) changes in body weights were noticed on day 1 (33%), 30 (42%) and 60 (25%), while no significant change was observed on day 90.
Table 1.
Comparison of body weight (mean ± SD) between two groups (n = 10)
| Age (day) | Weight of control (g) | Weight of ODM (g) | Percent of changes |
|---|---|---|---|
| 1 | 4.5 ± 0.5 | 6.0 ± 0.5* | 33 |
| 30 | 49.6 ± 12.8 | 70.8 ± 7.3* | 42 |
| 60 | 133.9 ± 4.3 | 167.7 ± 5.6* | 25 |
| 90 | 215.8 ± 11.2 | 232.8 ± 10.8 | 7.8 |
ODM Offspring of diabetic mother
* Parameters showed significant difference between two groups (P < 0.05)
Blood glucose and endocrine levels
Fasting blood glucose in the ODM showed a higher value (125.8 ± 12.2 vs. 90.1 ± 6.1) compared to the control group (Table 2) (P < 0.05).
Table 2.
Comparison of blood glucose and hormones level (mean ± SD) in controls and ODM (n = 10)
| Groups | Blood glucose (mg/dl) | FSH (IU/l) | LH (IU/l) | Testosterone (ng/ml) |
|---|---|---|---|---|
| Control | 90.1 ± 6.1 | 4.22 ± 0.22 | 0.31 ± 0.06 | 4.04 ± 1.34 |
| ODM | 125.8 ± 12.2* | 1.84 ± 0.51* | 0.15 ± 0.04* | 1.28 ± 0.44* |
ODM Offspring of diabetic mothers
* Significant difference observed between two groups (P < 0.05)
Level of serum testosterone in the ODM was significantly lower than that in the control group (1.28 ± 0.44 vs. 4.04 ± 1.34 ng/ml). As shown in Table 2, serum gonadotropins, FSH (4.22 ± 0.22 vs. 1.84 ± 0.51 IU/ml) and LH (0.31 ± 0.06 vs. 0.15 ± 0.04 IU/ml) also showed a significant decrease in the ODM when compared with control values (P < 0.05).
Testicular histomorphology
The values obtained for various testicular parameters are presented in Table 3. In the ODM group average weight and volume of testes were about 22.7% (1.35 ± 0.02 vs. 1.10 ± 0.06 g) and 22.9% (1,256.5 ± 42.1 vs. 1,022.5 ± 51.8 mm3), respectively, higher than those of the control group (P < 0.05). Histological sections from testes of control and ODM are presented in Figs. 1 and 2. All histomorphic testicular parameters, except the number of seminiferous tubules per square millimeter (+21%), showed a significantly lower value in ODM compared to controls (P < 0.05): thickness of the testicular capsule (−25%), number of Leydig cells per square millimeter (−15.6%), number of Sertoli cells in each seminiferous tubule (−14.9%), number of spermatogonia in the seminiferous tubule (−26.3%) and the diameter of the seminiferous tubules (−11%).
Table 3.
Comparison of testicular parameters (mean ± SD) between two groups (n = 10)
| Parameter | Control | ODM | % of changes |
|---|---|---|---|
| Testicular volume (mm3) | 1,022.5 ± 51.8 | 1,256.5 ± 42.1* | 22.9 |
| Testicular weight (g) | 1.10 ± 0.06 | 1.35 ± 0 .21* | 22.7 |
| Testes weight/body weight (%) | 0.51 | 0.58 | 0.07 |
| Spermatogonia/ST | 60.2 ± 11.5 | 44.4 ± 10.7* | 26.3 |
| Sertoli cell/ST | 19.1 ± 3.2 | 16.2 ± 1.4* | 14.9 |
| ST/mm2 | 7.38 ± 1.07 | 8.94 ± 1.23* | 21 |
| Leydig cell/mm2 | 990.2 ± 80.5 | 835.1 ± 99.0* | 15.6 |
| ST diameter (μm) | 271.86 ± 23.65 | 241.39 ± 18.44* | 11 |
| Thickness of capsule (μm) | 24.56 ± 3.64 | 18.42 ± 2.10* | 25 |
ST Seminiferous tubule, ODM offspring of diabetic mother
* Parameters showed a significant difference between the two groups (P < 0.05)
Fig. 1.
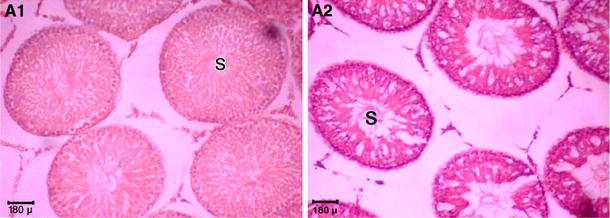
Section of testes showing seminiferous tubules of 90-day-old rats. H&E stain. A1 Offspring from normal mothers, A2 offspring from diabetic mothers, ×200
Fig. 2.
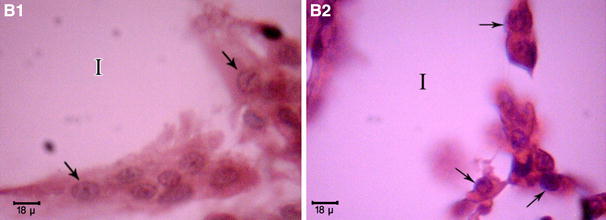
Section of testes showing interstitial tissue (Leydig cells) of 90-day-old rat. H&E staining. B1 Offspring from normal mother, B2 offspring from diabetic mother, ×2000. Arrows indicate the location of Leydig cells. Leydig cells in offspring from normal mother rats presented a characteristic appearance, with a very active nucleus, as shown by the presence of a decondensed chromatin and several nucleoli (B1). In contrast, in offspring from diabetic mother rats, B2 Leydig cells were scarce and showed a highly vacuolated cytoplasm. Furthermore, the nuclei of the cells were of irregular shape, and the chromatin content had a distinct appearance
Discussion
During first 2 months of life, the average body weight of ODM was significantly higher than that in control group. The difference was more remarkable during the 1st month, which could be due to increased placental transport of glucose and nutrients during fetal life [2]. This declined afterward so that it became non-significant on day 90. This condition, accompanied by a moderate increase in fasting blood glucose in ODM, may be due to maternal hyperglycemia leading to fetal hyperglycemia and hypoinsulinemia [22, 23].
Levels of FSH, LH and testosterone in ODM were significantly lower than that of the control. A decrease in levels of FSH, LH, prolactin and growth hormone has been reported in diabetic rats [17, 18]. Diabetes induces a decrease in the serum levels of LH, which is responsible for normal Leydig cell function (testosterone secretion) [18]. The pituitary of diabetic rats has a blunted response, with a diminished stimuli-induced secretion of FSH and LH [24]. Moreover, a relationship between insulin/glucose and LH/FSH levels in serum has been proposed, although the mechanisms by which insulin, glucose or both control these two hormones are unclear [25]. A decrease in testosterone level may be related to a decrease in LH, a decrease in the number of Leydig cells or both. This situation was reported in male diabetic rats [25].
Although testes weight in ODM was slightly increased, the ratio of testes to body weight did not change significantly. Hence, the increase in the size of testes is correlated to an increase in whole body size. Our results showed that, in testes of ODM, the number of Leydig cells, Sertoli cells and the diameter of seminiferous tubules were reduced when compared with the control group. To our knowledge, this is the first report on endocrine profile and histomorphological study of gonads in offspring from diabetic mothers. Existing data suggest that the altered maternal glucose metabolism at a critical time during embryogenesis is a major determinant for the risk of anomalies in pregnancies complicated by diabetes [26].
Severe hyperglycemia in the maternal rat results in hyperglycemia and hypoinsulinemia of the fetuses and fetal growth retardation [22, 23]. We have seen decreased live litter size in diabetic mothers, and some dead fetuses showed abnormality in the limbs. Moreover, some degenerative masses were also observed in the dissected uterus of diabetic mothers before delivery (data not presented). Interplay between maternal and fetal metabolism during the preimplantation period was proposed to influence the establishment of pregnancy and fertility problems in diabetic mothers [27].
The decreased numbers of Leydig cells observed in the ODM group may be due to hypoinsulinemia that occurred during fetal life. Induced diabetes was reported to cause a decrease in total Leydig cell number and impairment in cell function due to hypoinsulinemia [25]. The remarkable decrease in the expression of interstitial tissue insulin receptors in diabetic rats, together with the virtual absence of serum insulin, is expected to lead to a total loss of the insulin-mediated stimulation of androgen biosynthesis and cell proliferation, which is consistent with the morphologic alterations in the interstitial tissue observed.
Two parallel mechanisms were proposed to cause diabetes-related alterations in Leydig cells: (1) the effect of hypoinsulinemia on serum LH levels and (2) the combined effects of the decreased levels of LH and insulin on Leydig cells [25]. Moreover, diabetes mellitus was reported to alter the activity of arginase (an enzyme important in cell proliferation) in the reproductive system, which may impair Leydig cell proliferation [28].
In the ODM group, the number of seminiferous tubules per square millimeter increased, while the diameter of each tube decreased. This condition was accompanied by a decrease in FSH level in the serum. A decrease in FSH in diabetic rats will decrease tubular FSH receptors, which in turn will diminish the response of the epithelium of seminiferous tubules to FSH stimulation significantly [25]. Additionally, a decrease in the expression of insulin receptors could lead to a loss of insulin-mediated cell proliferation in seminiferous tubules [25]; both conditions will decrease the number of spermatogonia in seminiferous tubules, leading to a decrease in seminiferous tubule diameter. A decrease in seminiferous diameter was reported in alloxan diabetic rats [29]. An increased number of seminiferous tubules per millimeter could be due to a decrease in the diameter of seminiferous tubules.
In conclusion, maternal hyperglycaemia has significant deleterious effects on the structure and function of both the reproductive endocrine and testicular structures. This detrimental change is likely to have occurred during fetal life and remains during postnatal life; it shows a similar pattern of change as previously reported in adult male diabetic rats.
References
- 1.Pedersen J. The pregnant diabetic and her newborn: problems and management. 2. Baltimore: Williams & Wilkins; 1977. pp. 91–197. [Google Scholar]
- 2.Persson B, Hanson U. Neonatal morbidities in gestational diabetes mellitus. Diabetes Care. 1998;21:79–84. [PubMed] [Google Scholar]
- 3.Van Assche FA, Aerts L, De Prins FA. Degranulation of the insulin-producing B cells in an infant of a diabetic mother. Case report. Br J Obstet Gynecol. 1983;90:182–185. doi: 10.1111/j.1471-0528.1983.tb08906.x. [DOI] [PubMed] [Google Scholar]
- 4.Knowler W, Pettitt DJ, Kunzelman CL, Everhart J. Genetic and environment determinants of non-insulin dependent diabetes mellitus. Diabetes Res Clin Pract. 1985;1:309. [Google Scholar]
- 5.Chung CS, Myrianthopoulos NC. Factors affecting risks of congenital malformations. II. Effect of maternal diabetes on congenital malformations. Birth Defects. 1975;11:23–38. [PubMed] [Google Scholar]
- 6.Drazancic A, Blajic J, Djelmis J (1993) Early spontaneous abortions and congenital malformations in relation to diabetes regulation in very early pregnancy. In: 24th annual meeting of diabetic pregnancy Denmark. Abstract 3
- 7.Foglia VG, Rosner JM, Ramos M, Lema BE. Sexual disturbances in the male diabetic rat. Horm Metab Res. 1969;1:72–77. doi: 10.1055/s-0028-1095155. [DOI] [PubMed] [Google Scholar]
- 8.Cameron DF, Rountree J, Schultz RE, Repetta D, Murray FT. Sustained hyperglycemia results in testicular dysfunction and reduced fertility potential in BBWOR diabetic rats. Am J Physiol. 1990;259:881–889. doi: 10.1152/ajpendo.1990.259.6.E881. [DOI] [PubMed] [Google Scholar]
- 9.Hassan AA, Hassouna MM, Taketo T, Gagnon C, Elhilali MM. The effect of diabetes on sexual behavior and reproductive tract function in male rats. J Urol. 1993;149:148–154. doi: 10.1016/s0022-5347(17)36028-7. [DOI] [PubMed] [Google Scholar]
- 10.Murray FT, Cameron DF, Orth JM. Gonadal dysfunction in the spontaneously diabetic BB rat. Metabolism. 1983;32:141–147. doi: 10.1016/S0026-0495(83)80028-6. [DOI] [PubMed] [Google Scholar]
- 11.Fairburn CG, McCulloch DK, Wu FC. The effects of diabetes on male sexual function. Clin Endocrinol Metab. 1982;11:749–765. doi: 10.1016/S0300-595X(82)80011-X. [DOI] [PubMed] [Google Scholar]
- 12.Vignon F, Lefaou A, Montagnon D. Comparative study of semen in diabetic and healthy men. Diabetes Metab. 1991;17:350–354. [PubMed] [Google Scholar]
- 13.Khan S, Teerds K, Dorrington J. Growth factor requirements for DNA synthesis by Leydig cells from the immature rat. Biol Reprod. 1992;46:335–341. doi: 10.1095/biolreprod46.3.335. [DOI] [PubMed] [Google Scholar]
- 14.Hurtado de Catalfo G, Nelva I, De Gómez Dumm T. Lipid dismetabolism in Leydig and Sertoli cells isolated from streptozotocin-diabetic rats. Int J Biochem Cell Biol. 1998;30:1001–1010. doi: 10.1016/S1357-2725(98)00055-7. [DOI] [PubMed] [Google Scholar]
- 15.Borland K, Mita M, Oppenheimer CL, Blinderman LA, Massagué J, Hall PF, Czech MP. The actions of insulin-like growth factor-I and -II on cultured Sertoli cells. Endocrinology. 1984;114:240–246. doi: 10.1210/endo-114-1-240. [DOI] [PubMed] [Google Scholar]
- 16.Mita M, Borland K, Price JM, Hall PF. The influence of insulin and insulin-like growth factor-I on hexose transport by Sertoli cells. Endocrinology. 1985;116:987–992. doi: 10.1210/endo-116-3-987. [DOI] [PubMed] [Google Scholar]
- 17.Hutson JC, Stocco DM, Campbell GT, Wagoner J. Sertoli cell function in diabetic, insulin-treated diabetic, and semi-starved rats. Diabetes. 1983;32:112–116. doi: 10.2337/diabetes.32.2.112. [DOI] [PubMed] [Google Scholar]
- 18.Benítez A, Pérez Díaz J. Effect of streptozotocin-diabetes and insulin treatment on regulation of Leydig cell function in the rat. Horm Metab Res. 1985;17:5–7. doi: 10.1055/s-2007-1013433. [DOI] [PubMed] [Google Scholar]
- 19.Elias H, Hyde DM. An elementary introduction to stereology (quantitative microscopy) Am J Anat. 1980;159:411–417. doi: 10.1002/aja.1001590407. [DOI] [PubMed] [Google Scholar]
- 20.Luna LG (1968) Manual of histologic staining methods of the Armed forces institute of pathology, 3rd edn. American registry of pathology. McGraw Hill, USA, pp 34–77
- 21.Franca LR, Suescun MO, Miranda JR, Giovambattista A, Perello M, Spinedi E, et al. Testis structure and function in a non-genetic hyperadipose rat model at prepubertal and adult ages. Endocrinology. 2005;147:1556–1563. doi: 10.1210/en.2005-0640. [DOI] [PubMed] [Google Scholar]
- 22.Aerts L, Van Assche FA. Rat fetal endocrine pancreas in experimental diabetes. J Endocrinol. 1977;73:339–346. doi: 10.1677/joe.0.0730339. [DOI] [PubMed] [Google Scholar]
- 23.Devaskar S, McMenamy K, Holtzclaw L, Sadiq F. Long-term maternal-fetal exposure to high-low insulin concentrations alter liver but not brain insulin receptors. Am J Obstet Gynecol. 1990;163:1350–1356. doi: 10.1016/0002-9378(90)90718-m. [DOI] [PubMed] [Google Scholar]
- 24.Seethalakshmi L, Menon M, Diamond D. The effect of streptozotocin induced diabetes on the neuroendocrine-male reproductive tract axis of the adult rat. J Urol. 1987;138:190–194. doi: 10.1016/s0022-5347(17)43042-4. [DOI] [PubMed] [Google Scholar]
- 25.Ballester J, Munoz MC, Dominguez J, Rigau T, Guinovart JJ, Rodriguez-Gil JE. Insulin-dependent diabetes affects testicular function by FSH and LH-linked mechanisms. J Androl. 2004;25:706–719. doi: 10.1002/j.1939-4640.2004.tb02845.x. [DOI] [PubMed] [Google Scholar]
- 26.Schaefer-Graf UM, Buchanan TA, Xiang A, Songster G, Montoro M, Kjos SL. Patterns of congenital anomalies and relationship to initial maternal fasting glucose levels in pregnancies complicated by type 2 and gestational diabetes. Am J Obstet Gynecol. 2000;182:313–320. doi: 10.1016/S0002-9378(00)70217-1. [DOI] [PubMed] [Google Scholar]
- 27.Neill JD. Knobil and Neill’s Physiology of Reproduction. 3. New York: Elsevier/Academic Press; 2006. pp. 291–292. [Google Scholar]
- 28.Jelodar GA, Razmi N, Golampour V. Arginase alteration in reproductive system of alloxan diabetic dogs. J Reprod Dev. 2007;53:317–321. doi: 10.1262/jrd.18049. [DOI] [PubMed] [Google Scholar]
- 29.Hassen NS, EIRoubi NM, Omara EA. Evaluation of the influence of each of melatonin and chromium against diabetes-induced alteration in the testis of Albino rats using light and electron microscopies. Egypt J Hospital Med. 2007;27:143–162. [Google Scholar]