

Abstract
Early-life stress may cause several neuropsychological disorders in adulthood. Such disorders may be induced as a result of instability of neuronal circuits and/or synaptic formation. However, the mechanisms underlying such instability have not yet been clearly understood. We previously reported that the mushroom spine in the somatosensory cortex (SSC) is unstable in early-life stressed mice not only in the juvenile stage but also in adulthood. In this study, we measured the number and motility of microglial processes in early-life stressed mice to understand the mechanism further. We found that the number and motility of filopodia-like protrusions of microglial processes tended to increase in the SSC of early-life stressed mice. Interestingly, the motility of protrusions correlated significantly with the nociceptive threshold level measured by the von Frey test. These results indicated that the activity of microglia affected the neuronal function in early-life stressed mice.
Keywords: Maternal deprivation, In vivo imaging, In vivo microdialysis, Somatosensory cortex
Introduction
Early-life stress during perinatal development affects brain development and causes various neuropsychological disorders throughout life. However, how exposure to stress during childhood leads to functional alteration in adulthood has not yet been clearly understood [1]. To study the mechanisms, maternal separation/deprivation models have been used (see [2] for review). In these models, early-life stress induces various disorders in adulthood, i.e., an enhancement of anxiety-related behaviors [2–6]. Early-life stressed rodents show neuronal changes such as an increase in synaptic spine density in the infralimbic cortex and somatosensory cortex (SSC) [7, 8]. We also reported previously that the mushroom spine in the SSC becomes unstable in an early-life stress model, and the nociceptive threshold level decreases in this model [9]. We have also reported that maternal deprivation (MD) increases the glutamate (Glu) release level in the SSC. Such an increase augments the electrophysiological activity in the SSC [10, 11]. Because the activity of the neuronal network defines brain functions, the impairment of neuronal circuit formation by early-life stress in rodents may lead to psychiatric disorders and behavioral alterations in adulthood.
Recently, the role of microglia, the resident immune cells in the central nervous system, in the maintenance of synaptic function has been well studied [12–15]. In response to chronic or acute brain damage such as cerebral ischemia or application of bacterial lipopolysaccharide, microglia are activated, producing structural changes such as elongation and retraction of processes [16, 17]. Previous studies showed that microglial processes come in contact with synapses in an activity-dependent manner, possibly to modify synaptic function [17, 18]. These results indicate that the activity of microglia also changes in MD mice, and it induces instability of the spine.
In this study, using an in vivo imaging technique we found that the motility of microglial processes in the SSC increased in MD mice, particularly after vibrotactile limb stimulation. Interestingly, this increase significantly correlated with the decrease in the nociceptive threshold level studied by the von Frey test. These results indicated that the increase in activity of SSC increases the motility of microglia in MD mice.
Materials and methods
This study was performed in accordance with the guidelines and protocols approved by the Animal Care and Experimentation Committee, Gunma University, and the National Institutes of Natural Sciences, according to EU Directive 2010/63/EU on the protection of animals used for scientific purposes. All efforts were made to minimize the suffering and number of animals used.
Animals
Adult male Iba1-EGFP transgenic mice derived from the C57BL/6J strain (2–2.5 months of age) [19] and wild-type C57BL/6J mice (2–2.5 months of age; purchased from SLC Japan) were used in this study. All the mice were housed with food and water provided ad libitum under controlled temperature (25 ± 5 °C), humidity, and illumination (12:12 light:dark cycle; lights on at 7:00 a.m.) conditions. Cages were changed once a week.
MD mice were separated for 3 h (usually from 9:00 a.m. to 12:00 a.m.) every day from their mother from postnatal day (P) 2 to P14. Separated MD mice were individually placed in a locally made incubator at regulated humidity (50–70 %) and temperature (37 °C). The control mice were naïve so that they did not experience MD. All experiments except MD were performed in males at 2–2.5 months of age in the present study. There is some possibility that the separation paradigm induces nutritional disorder, which affected the anatomical structure of synapses [20]. However, the body weights were not different between the control and MD mice examined at several stages of development [9]. Thus, separation from the mother did not affect the body weight, an index of the nutritional status of mice, as found in our experiments following the MD protocol.
The von Frey hair test
Mice were individually placed on an elevated metal wire mesh table [9]. A von Frey filament was applied to the middle plantar surface of a hindpaw until the filament was bent. The filament was held in this position for 1 s. Only robust and immediate withdrawal responses to the stimulation were counted as withdrawal responses. Each trial consisted of the filament being applied eight times at 30-s intervals. The mechanical nociceptive threshold was considered as the lowest force that evoked a paw withdrawal in at least 50 % of trials.
In vivo imaging
In vivo imaging was conducted 12–24 h after the von Frey test. Mice were anesthetized by injection of urethane (1.7 g/kg body weight, i.p.) and atropine (0.4 mg/kg body weight, i.p.), as previously reported [21]. The body temperature was maintained at 37 ± 0.5 °C using a heating pad and feedback regulation from a rectal temperature probe. Then, in vivo imaging was performed using a two-photon laser microscopy system customized for in vivo imaging (FV1000 confocal microscope, Olympus; MaiTaiHP, Spectra-Physics Inc.) using a 20× water objective lens (NA 0.90). The laser power was adjusted below 15 mW (7–12 mW) at 950 nm ([9]; see also Fig. 1). To avoid the changes in microglial phenotype due to the surgical procedure, images of the microglia were taken using the thinned-skull imaging window, as previously reported [22]. Briefly, the skull of a mouse was carefully thinned using a high-speed drill (Narishige) and a microsurgical blade (Surgistar no. 6400). Then, the first imaging was performed. One image stack consisted of 30–50 horizontal planes, covering an area of 210 × 210 μm2 (512 × 512 pixels) with 0.75-μm axial spacing. The imaging depths were from 50 to 100 μm from the surface. Since one optical imaging takes 1.644 s, the total duration of imaging of each stack was from 50 to 80 s. Thus, ten stacks can be imaged in 20 min, and each projected image was used for analysis. All analysis of imaging was performed by ImageJ. The number of newly generated processes over 20 min (a process that was not found in the first imaging stack and did not appear until the ninth imaging stack) was counted and expressed as the number of processes/cell. The change in length (elongation or retraction) of each process from one stack to the next was also measured and expressed as motility (μm/min). Furthermore, the duration of the transient stop between elongation and retraction was measured and expressed as an interval (min). Before limb stimulation, images were taken for 20 min. The number of processes of microglia, motility of processes, and duration of the transient stop were determined as shown above. Then, vibrotactile somatosensory stimulations (500 ms stimulation duration, 0.1 Hz for 20 min) were applied to the right fore- and hindlimbs using a custom-made mechanical limb manipulator equipped with a vibration motor (8400 rpm; SE-4F-A3A1-X0, Shicoh Engineering Co., Ltd.). During and after the stimulation, in vivo imaging was continuously performed. The number of newly generated processes, motility of each process of the microglia, and duration of each interval were measured as described above.
Fig. 1.
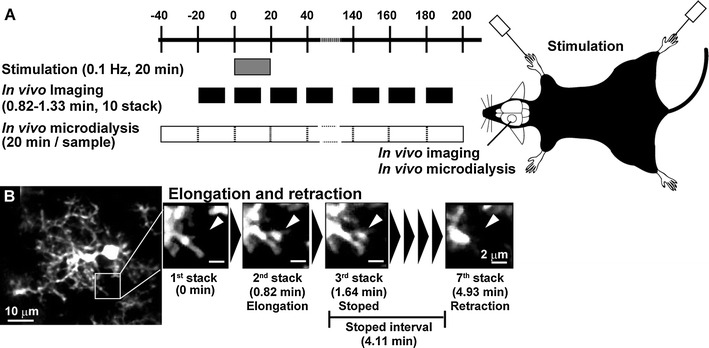
a Schematic drawing of the experimental protocol. In vivo imaging and in vivo microdialysis sampling were carried out every 20 min before and after vibrotactile limb stimulation (0.1 Hz, 20 min). Imaging and microdialysis sampling were carried out on the left somatosensory cortex. b In vivo images of a single microglia (left panel) and representative images of a process showing elongation, retraction, and temporary stop (right panels)
In vivo microdialysis
A guide cannula was inserted into the SSC through the cranial window (ϕ 0.5 mm) located 1.5–2 mm lateral to the sagittal suture and −1.5 to 1 mm anterior to the bregma under ketamine/xylazine (22.5 mg/ml ketamine and 1 mg/ml xylazine in 0.9 % NaCl, i.p.; 5 ml/kg) anesthesia, as previously reported [21]. Mice were allowed to recover from the surgery for at least 2–3 days before the von Frey test. Approximately 12–24 h after the von Frey test, microdialysis was conducted. Each microdialysis was conducted in the light phase (7:00–19:00) under anesthesia induced with urethane (1.7 g/kg body weight, i.p.) and atropine (0.4 mg/kg body weight, i.p.). A microdialysis probe (0.37 mm in diameter and 1.5 mm in membrane length) was inserted into the guide cannula at 4–6 h before dialysis. The probe was perfused at a rate of 2 μl/min with HEPES buffer (in mM: NaCl, 140.0; KCl, 2.5; CaCl2, 2.0; MgCl2, 1.0; HEPES, 5.0; glucose, 10.0; pH 7.4). After 40 min (20 min × 2 samples) of preperfusion, vibrotactile limb stimulation was performed as described above. The obtained microdialysate was sampled until 3 h after stimulation (Fig. 1). Then, Glu, glutamine (Gln), glycine (Gly), and gamma-aminobutyric acid (GABA) levels were immediately measured by high-performance liquid chromatography (EICOM) [21]. The concentrations of Glu, Gln, Gly, and GABA were normalized using a calibration curve from 100 μM to 10 nM.
Statistical analyses
All the data were compared by one- or two-way ANOVA. Post hoc comparison was performed by the Bonferroni test, Student’s t test, or Scheffe’s group comparison using Excel statistics (Esumi). Correlation was calculated by Pearson correlation. The difference was considered significant at p < 0.05. All values are presented as mean ± SEM.
Results
Motility of microglia was changed in MD mice
As reported previously [9], MD mice showed a decrease in the threshold level for nociceptive stimulation (1.32 ± 0.07 g in control mice, 0.57 ± 0.05 g in MD mice). These values were found to be statistically significantly different by Student’s t test; t (9) = 8.63, p = 0.00001). The number (density) of microglia was not changed in early-life stressed mice. In control mice, 3.8 ± 0.6 microglia (313.7 ± 26.2 microglia/mm3) were detected in imaging area, whereas 4.0 ± 0.3 microglia (319.4 ± 17.1 microglia/mm3) were detected in early-life stressed mice (no statistical difference). As shown in Fig. 1b, each microglial cell had highly ramified processes. Even without movement of the cell soma, each process was highly motile, showing elongation and retraction with temporary stops of movement (Fig. 1b right panels). As shown in Fig. 2a, the number of newly generated filopodia-like processes during a 20-min imaging session was 5.1 ± 1.2/cell in control mice (n = 5 mice including 13 cells). The number of such processes tended to increase in MD mice (9.1 ± 1.3/cell, n = 5 mice including 14 cells; t (8) = 2.27, p = 0.053 by Student’s t test). As shown in Fig. 2b, in control mice, the extents of change in the length of processes during a 20-min imaging session were 1.80 ± 0.20 μm/min for elongation and −1.77 ± 0.13 μm/min for retraction. In MD mice, the extents of elongation and retraction increased significantly (elongation, 2.70 ± 0.22 μm/min, by Student’s t test. t (8) = 3.03, p = 0.016; retraction, −2.73 ± 0.20 μm/min, t (8) = 4.04, p = 0.004). As shown in Fig. 2c, the duration of the temporary stop between elongation and retraction (interval) was 1.19 ± 0.16 min in control mice. The duration did not significantly change in MD mice (1.60 ± 0.16 min) (by Student’s t test. t (8) = 1.80, p = 0.109). Interestingly, the motility negatively/positively correlated with the nociceptive threshold level (Fig. 2b; elongation, R = −0.654, p = 0.04; retraction, R = 0.736, p = 0.015).
Fig. 2.
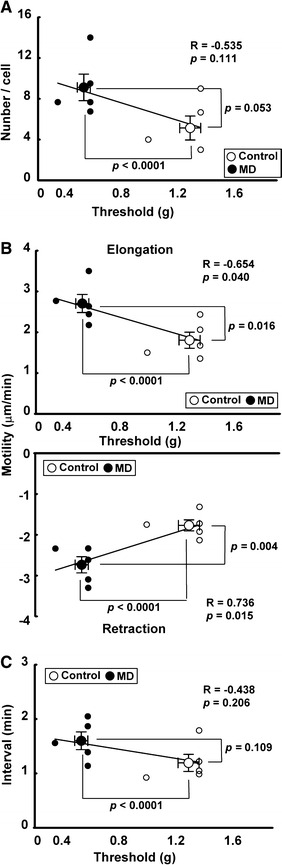
Effect of maternal deprivation on the nociceptive threshold level of the von Frey hair test, number and motility of processes, and interval of transient stop (interval). a Change in the nociceptive threshold level by maternal deprivation. As previously reported, MD mice show a low threshold level in the von Frey hair test. The number of filopodia-like processes tended to increase in MD mice. b Motility of the processes significantly increased in MD mice. Interestingly, the value was significantly correlated with the value of nociceptive threshold level. c The interval of transient stop slightly increased in MD mice. However, the value was not statistically significant
We next carried out the vibrotactile limb stimulation to activate the SSC. As shown in Fig. 3a, the number of processes increased significantly (two-way ANOVA; group × time; F (1,10) = 3.71, p = 0.0004) particularly 20–40 min after the onset of stimulation in control mice (11.8 ± 1.7/cell; by the Bonferroni test; F (21,87) = 18.85, p < 0.000001; t (8) = 3.97, p = 0.0001). After this time period, however, the number decreased to the prestimulation level (6.5 ± 0.9/cell 60–80 min after stimulation. 5.3 ± 0.4/cell 180–200 min after stimulation). In MD mice, although the number also increased for the first 20 min similarly to those in control, the increase lasted longer. The number (17.3 ± 0.9/cell) in MD mice was significantly higher than that in control mice even 3 h after stimulation (14.9 ± 1.1/cell 180–200 min after stimulation; t (8) = 6.93, p < 0.000001).
Fig. 3.
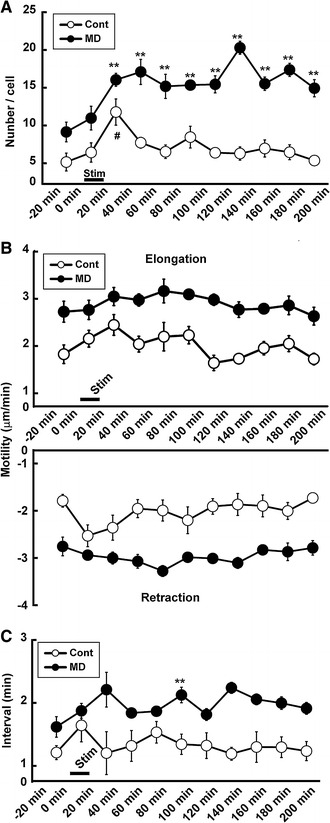
Effects of vibrotactile limb stimulation on the number and motility of processes and duration of transient stop. a Change in number of processes. The number of processes increased significantly only during 0–20 min after somatosensory stimulation in control mice. In other hands, the number continuously increased even 160 min after somatosensory stimulation in MD mice. b Change in motility of processes. Note that although the motility was higher in MD mice throughout the study, limb stimulation did not markedly alter the motility in control and MD mice. c Duration of transient stop of processes. Note that the duration of transient stop increased in control mice after somatosensory stimulation, whereas it was continuously higher in MD mice after stimulation. Hash indicates α < 0.05 compared between pre- and poststimulation in control mice; double asterisk indicates α < 0.01 compared between control and MD mice by the Bonferroni test
As shown in Fig. 3b, the extent of elongation of processes did not significantly increase following vibrotactile limb stimulation in either the control or MD group (two-way ANOVA; group × time; F (1,10) = 0.61, p = 0.779), although the values were significantly different between the control and MD groups throughout the experiment (by group Scheffe’s test, t (21) = 7.85, p < 0.000001). Like for elongation, the motility of processes during retraction did not significantly dramatically increase following vibrotactile limb stimulation in both the control and MD groups (two-way ANOVA; group × time; F (1,10) = 1.12, p = 0.345), although the values were significantly different between the control and MD groups throughout the experiment (by group Scheffe’s test, t (21) = 9.22, p < 0.000001).
As shown in Fig. 3c, the duration of the transient stop between elongation and retraction changed significantly (two-way ANOVA; group × time; F (1,10) = 2.12, p = 0.031). In control mice, the value was significantly increased during the stimulation (1.97 ± 0.13 min during stimulation; by Student’s t test; t (8) = 3.24, p = 0.012). However, the duration decreased to the prestimulation level immediately after the termination of stimulation. In MD mice, on the other hand, the duration did not significantly change after stimulation, although the value was higher than in control mice during the whole experiment (by group Scheffe’s test, t (21) = 5.42, p < 0.000001).
Glutamate level in the SSC kept high in MD mice even under anesthesia
We next examined the concentration of Glu in the SSC under anesthesia using in vivo microdialysis. As shown in Fig. 4a, the levels before stimulation were 56.4 ± 11.6 nM in control mice (n = 7) and 266.1 ± 70.9 nM in MD mice (n = 10) (Student’s t test; t (16) = 1.83, p = 0.017). Interestingly, these levels correlated negatively with the nociceptive threshold level (R = −0.576, p = 0.016). As shown in Fig. 4b, the Glu level was continuously higher in MD mice than in control mice throughout the study (Scheffe’s group comparison; t (198) = 61.43, p < 0.000001), although the level did not significantly change during/after the stimulation in either group.
Fig. 4.
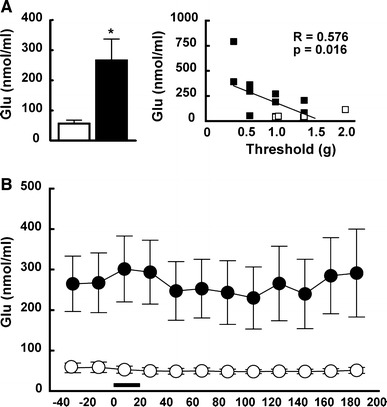
Glu level before and following somatosensory stimulation. a The basal level of Glu increased significantly in MD mice. The level was negatively correlated with the threshold level in the von Frey hair test. b The Glu level did not significantly change following somatosensory stimulation in both the control and MD mice. However, the level was continuously high in MD mice
We also measured the concentration of Gln, Gly, and GABA. The levels were not significantly different between the control and MD mice before stimulation (Gln: 59.5 ± 29.3 μM in control mice and 40.0 ± 10.4 μM in MD mice; Gly: 121.3 ± 68.1 nM in control mice and 104.6 ± 48.4 nM in MD mice; GABA: 45.0 ± 1.2 nM in control mice and 48.8 ± 2.3 nM in MD mice). Like Glu, the levels did not change significantly during/after the stimulation in both groups (data not shown).
Discussion
In this study, we found that the number of processes of microglia, which is not significantly different under basal conditions, increased significantly in MD mice particularly after limb stimulation (Fig. 3a). The motility of processes also increased significantly in MD mice under basal conditions and remained higher than those of the control mice during and after limb stimulation, although limb stimulation did not markedly increase the motility. Interestingly, the duration of temporary stop increased for at least 3 h after somatosensory stimulation in MD mice (Fig. 3c). Furthermore, the Glu level continuously increased in MD mice (Fig. 4). Since a previous study has shown that an enhancement of synaptic regeneration may increase the duration of contact of microglial processes with synapses [17], these results indicate the possibility that the change in motility of microglia potentially induces instability of the spine, leading to a decrease in the nociceptive threshold level in MD mice.
As reported previously, different types of environmental stress enhance glutamate release and reduce glial cell-mediated Glu cycling [23], producing an increase in Glu in the interstitial space. Regarding the mechanism, acute exposure to stress or administration of glucocorticoids rapidly increases the glutamate release in the hippocampus and other brain regions [24–26]. Glucocorticoid increases the expression of presynaptic soluble n-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) protein complexes via the glucocorticoid receptor in the presynaptic membrane [27]. Chronic stress also decreases the number of glial fibrillary acid protein (GFAP)-expressing cells and impairs clearance of synaptic glutamate through excitatory amino acid transporters [28]. These changes increase the concentration of glutamate and basal activity in the brain. Nevertheless, the effects of chronic stress on glutamate release have not yet been fully understood [29].
Microglia play important roles not only in the developing brain but also in the adult brain. In neurodegenerative disorders, such as Alzheimer’s disease, Parkinson’s disease, and cerebral ischemia, microglia are activated and may contribute to pathogenesis by inducing neuronal death or removing synapses, although they may also play an important role in the defense against the development of many neurodegenerative disorders [30, 31]. Recently, it has been reported that nonactivated microglia may also contribute to the synaptic formation/disruption [14, 17, 18, 32–34]. In an ischemic penumbra in the SSC, microglia comes in contact with synapses for a markedly longer period. The duration of synaptic contact correlates to an increased turnover of presynaptic boutons [17]. In this study, we found that the motility of microglial processes increased significantly in MD mice (Fig. 2). These changes were enhanced by somatosensory stimulation. On the other hand, the duration of the transient stop also increased after stimulation, indicating the potential increase in microglia-synapse interaction, which may lead to the removal of synapses. Such changes may induce the instability of neural transmission in MD mice [9].
As shown in Fig. 4, the concentration of Glu was 10–50 % under anesthesia compared with those in free-moving conditions [11]. Our stimulation method, however, was strong enough to induce neuronal and glial cell responses under anesthesia [21]. In this study, although limb stimulation did not increase the Glu concentration, the level was continuously higher in MD mice. Furthermore, the nociceptive threshold level significantly correlated with the motility of processes and Glu level (Figs. 2b, 3b). Based on these results, we hypothesized that the motility may have increased as a consequence of microglial activation by Glu. To confirm this hypothesis further, we should examine (1) the precise change in the local Glu concentration and (2) the effect of inhibition of the Glu receptor by applying antagonists. Such treatment requires insertion of a microdialysis probe or electrode, or intracerebral application of drugs. These procedures may produce mechanical insults that may alter the microglial motility. In the present study, we applied the thin-skull method to avoid such insults so that we could observe the intact microglia [35]. In addition, the effect of anesthesia may need to be considered. Changing the method of anesthesia (e.g., inhalation anesthesia) and/or combining it with the free-moving in vivo imaging method [36], the release of Glu could be altered, leading to a change in microglial motility particularly after somatosensory stimulation. However, alteration of the method of anesthesia may potentially change the condition of not only microglia but also other subsets of cells (neuron and/or astrocyte), making the interpretation of findings rather difficult. Thus, although it may not be appropriate to alter the experimental conditions in the present study, it is of interest to design a new experimental procedure with different approaches.
Our previous study [9] showed that MD induced an increase in mushroom spine turnover in the SSC in adulthood. Using an electrophysiological approach, we have also shown that neuronal activity in the SSC is increased together with an increase in glutamate release [11]. These results indicate that alteration of neuronal activity, particularly that of glutamatergic neurons, may alter the somatosensory threshold. However, it is still not clear whether the increase in the somatosensory threshold is a direct consequence of the change in spine turnover, because, although the turnover is significantly altered, the change is only 3–5 % of total mushroom spines. There is a possibility that the increase in neuronal activity may alter the somatosensory threshold and turnover of the synaptic spine separately. To clarify the mechanism further, more study is necessary.
Conclusions
In summary, the number and motility of microglia processes in the SSC increased in MD mice with a decreasing somatosensory threshold level. Furthermore, the duration of the transient stop between elongation and retraction also increased in MD mice after somatosensory stimulation. The activation of microglia may alter the synaptic plasticity, which may in turn alter the somatosensory threshold. Thus, the neuropsychological disorders in adulthood induced by early-life stress may at least in part be due to the alteration of the plasticity of microglia.
Acknowledgments
This work was supported by a Grant-in-Aid for Scientific Research on Innovative Areas (23111504) from the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) to YT, a Grant-in-Aid for Scientific Research (B) (21390065) from the Japan Society for the Scientific of Sciences (JSPS) to NK, and Core Research for Evolutional Science and Technology from Japan Science and Technology Agency and Grant-in-Aid for Scientific Research (B) (19390055) and (A) (22240042) from the Japan Society for the Scientific of Sciences (JSPS) to JN.
Conflict of interest
There is no conflict of interest in the present study.
References
- 1.Romeo RD, McEwen BS. Stress and the adolescent brain. Ann N Y Acad Sci. 2006;1094:202–214. doi: 10.1196/annals.1376.022. [DOI] [PubMed] [Google Scholar]
- 2.Shair HN. Acquisition and expression of a socially mediated separation response. Behav Brain Res. 2007;182:180–192. doi: 10.1016/j.bbr.2007.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kosten TA, Lee HJ, Kim JJ. Early life stress impairs fear conditioning in adult male and female rats. Brain Res. 2006;1087:142–150. doi: 10.1016/j.brainres.2006.03.009. [DOI] [PubMed] [Google Scholar]
- 4.Parfitt DB, Levin JK, Saltstein KP, Klayman AS, Greer LM, Helmreich DL. Differential early rearing environments can accentuate or attenuate the responses to stress in male C57BL/6 mice. Brain Res. 2004;1016:111–118. doi: 10.1016/j.brainres.2004.04.077. [DOI] [PubMed] [Google Scholar]
- 5.Slotten HA, Kalinichev M, Hagan JJ, Marsden CA, Fone KC. Long-lasting changes in behavioural and neuroendocrine indices in the rat following neonatal maternal separation: gender-dependent effects. Brain Res. 2006;1097:123–132. doi: 10.1016/j.brainres.2006.04.066. [DOI] [PubMed] [Google Scholar]
- 6.Wigger A, Neumann ID. Periodic maternal deprivation induces gender-dependent alterations in behavioral and neuroendocrine responses to emotional stress in adult rats. Physiol Behav. 1999;66:293–302. doi: 10.1016/S0031-9384(98)00300-X. [DOI] [PubMed] [Google Scholar]
- 7.Bock J, Gruss M, Becker S, Braun K. Experience-induced changes of dendritic spine densities in the prefrontal and sensory cortex: correlation with developmental time windows. Cereb Cortex. 2005;15:802–808. doi: 10.1093/cercor/bhh181. [DOI] [PubMed] [Google Scholar]
- 8.Ovtscharoff W, Jr, Braun K. Maternal separation and social isolation modulate the postnatal development of synaptic composition in the infralimbic cortex of Octodon degus. Neuroscience. 2001;104:33–40. doi: 10.1016/S0306-4522(01)00059-8. [DOI] [PubMed] [Google Scholar]
- 9.Takatsuru Y, Yoshitomo M, Nemoto T, Eto K, Nabekura J. Maternal separation decreases the stability of mushroom spines in adult mice somatosensory cortex. Brain Res. 2009;1294:45–51. doi: 10.1016/j.brainres.2009.07.092. [DOI] [PubMed] [Google Scholar]
- 10.Toya S, Takatsuru Y, Amano I, Koibuchi N. The molecular mechanism of early life stress for synaptic instability in somatosensory cortex. J Physiol Sci. 2013;63:S152. [Google Scholar]
- 11.Toya S, Takatsuru Y, Kokubo M, Amano I, Shimokawa N, Koibuchi N. Early-life-stress affects the homeostasis of glutamatergic synapses. Eur J Neurosci. 2014;40:3627–3634. doi: 10.1111/ejn.12728. [DOI] [PubMed] [Google Scholar]
- 12.Eyo UB, Wu LJ. Bidirectional microglia-neuron communication in the healthy brain. Neural Plast. 2013;2013:456857. doi: 10.1155/2013/456857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Miyamoto A, Wake H, Moorhouse AJ, Nabekura J. Microglia and synapse interactions: fine tuning neural circuits and candidate molecules. Front Cell Neurosci. 2013;7:70. doi: 10.3389/fncel.2013.00070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Morris GP, Clark IA, Zinn R, Vissel B. Microglia: a new frontier for synaptic plasticity, learning and memory, and neurodegenerative disease research. Neurobiol Learn Mem. 2013;105:40–53. doi: 10.1016/j.nlm.2013.07.002. [DOI] [PubMed] [Google Scholar]
- 15.Wake H, Moorhouse AJ, Miyamoto A, Nabekura J. Microglia: actively surveying and shaping neuronal circuit structure and function. Trends Neurosci. 2013;36:209–217. doi: 10.1016/j.tins.2012.11.007. [DOI] [PubMed] [Google Scholar]
- 16.Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo . Science. 2005;308:1314–1318. doi: 10.1126/science.1110647. [DOI] [PubMed] [Google Scholar]
- 17.Wake H, Moorhouse AJ, Jinno S, Kohsaka S, Nabekura J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J Neurosci. 2009;29:3974–3980. doi: 10.1523/JNEUROSCI.4363-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tremblay M-È, Lowery RL, Majewska AK. Microglial interactions with synapses are modulated by visual experience. PLoS Biol. 2010;8:e1000527. doi: 10.1371/journal.pbio.1000527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hirasawa T, Ohsawa K, Imai Y, Ondo Y, Akazawa C, Uchino S, Kohsaka S. Visualization of microglia in living tissues using Iba1-EGFP transgenic mice. J Neurosci Res. 2005;81:357–362. doi: 10.1002/jnr.20480. [DOI] [PubMed] [Google Scholar]
- 20.Crawford MA, Doyle W, Leaf A, Leighfield M, Ghebremeskel K, Phylactos A. Nutrition and neurodevelopmental disorders. Nutr Health. 1993;9:81–97. doi: 10.1177/026010609300900205. [DOI] [PubMed] [Google Scholar]
- 21.Takatsuru Y, Eto K, Kaneko R, Masuda H, Shimokawa N, Koibuchi N, Nabekura J. Critical role of the astrocyte for functional remodeling in contralateral hemisphere of somatosensory cortex after stroke. J Neurosci. 2013;33:4683–4692. doi: 10.1523/JNEUROSCI.2657-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grutzendler J, Kasthuri N, Gan WB. Long-term dendritic spine stability in the adult cortex. Nature. 2002;420:812–816. doi: 10.1038/nature01276. [DOI] [PubMed] [Google Scholar]
- 23.Sanacora G, Treccani G, Popoli M. Towards a glutamate hypothesis of depression: an emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology. 2012;62:63–77. doi: 10.1016/j.neuropharm.2011.07.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lowy MT, Gault L, Yamamoto BK. Adrenalectomy attenuates stress-induced elevations in extracellular glutamate concentrations in the hippocampus. J Neurochem. 1993;61:1957–1960. doi: 10.1111/j.1471-4159.1993.tb09839.x. [DOI] [PubMed] [Google Scholar]
- 25.Moghaddam B. Stress preferentially increases extraneuronal levels of excitatory amino acids in the prefrontal cortex: comparison to hippocampus and basal ganglia. J Neurochem. 1993;60:1650–1657. doi: 10.1111/j.1471-4159.1993.tb13387.x. [DOI] [PubMed] [Google Scholar]
- 26.Reznikov LR, Grillo CA, Piroli GG, Pasumarthi RK, Reagan LP, Fadel J. Acute stress-mediated increases in extracellular glutamate levels in the rat amygdala: differential effects of antidepressant treatment. Eur J Neurosci. 2007;25:3109–3114. doi: 10.1111/j.1460-9568.2007.05560.x. [DOI] [PubMed] [Google Scholar]
- 27.Musazzi L, Milanese M, Farisello P, Zappettini S, Tardito D, Barbiero VS, Bonifacino T, Mallei A, Baldelli P, Racagni G, Raiteri M, Benfenati F, Bonanno G, Popoli M. Acute stress increases depolarization-evoked glutamate release in the rat prefrontal/frontal cortex: the dampening action of antidepressants. PLoS ONE. 2010;5:e8566. doi: 10.1371/journal.pone.0008566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Banasr M, Duman RS. Glial loss in the prefrontal cortex is sufficient to induce depressive-like behaviors. Biol Psychiatry. 2008;64:863–870. doi: 10.1016/j.biopsych.2008.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Popoli M, Yan Z, McEwen BS, Sanacora G. The stressed synapse: the impact of stress and glucocorticoids on glutamate transmission. Nat Rev Neurosci. 2011;13:22–37. doi: 10.1038/nrn3138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Aguzzi A, Barres BA, Bennett ML. Microglia: scapegoat, saboteur, or something else? Science. 2013;339:156–161. doi: 10.1126/science.1227901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Perry VH, O’Connor V. The role of microglia in synaptic stripping and synaptic degeneration: a revised perspective. ASN Neuro. 2010;2:e00047. doi: 10.1042/AN20100024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Paolicelli RC, Bolasco G, Pagani F, Maggi L, Scianni M, Panzanelli P, Giustetto M, Ferreira TA, Guiducci E, Dumas L, Ragozzino D, Gross CT. Synaptic pruning by microglia is necessary for normal brain development. Science. 2011;333:1456–1458. doi: 10.1126/science.1202529. [DOI] [PubMed] [Google Scholar]
- 33.Parkhurst CN, Yang G, Ninan I, Savas JN, Yates JR, 3rd, Lafaille JJ, Hempstead BL, Littman, Gan WB. Microglia promote learning-dependent synapse formation through brain-derived neurotrophic factor. Cell. 2013;155:1596–1609. doi: 10.1016/j.cell.2013.11.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Schafer DP, Lehrman EK, Kautzman AG, Koyama R, Mardinly AR, Yamasaki R, Ransohoff RM, Greenberg ME, Barres BA, Stevens B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron. 2012;74:691–705. doi: 10.1016/j.neuron.2012.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Xu HT, Pan F, Yang G, Gan WB. Choice of cranial window type for in vivo imaging affects dendritic spine turnover in the cortex. Nat Neurosci. 2007;10:549–551. doi: 10.1038/nn1883. [DOI] [PubMed] [Google Scholar]
- 36.Dombeck DA, Harvey CD, Tian L, Looger LL, Tank DW. Functional imaging of hippocampal place cells at cellular resolution during virtual navigation. Nat Neurosci. 2010;13:1433–1440. doi: 10.1038/nn.2648. [DOI] [PMC free article] [PubMed] [Google Scholar]