

Abstract
Alzheimer’s disease is a progressive neurodegenerative disorder with decline in memory. The role of oxidative stress is well known in the pathogenesis of the disease. The purpose of this study was to evaluate pretreatment effects of oleuropein on oxidative status and cognitive dysfunction induced by colchicine in the hippocampal CA1 area. Male Wistar rats were pretreated orally once daily for 10 days with oleuropein at doses of 10, 15 and 20 mg/kg. Thereafter, colchicine (15 μg/rat) was administered into the CA1 area of the hippocampus to induce cognitive dysfunction. The Morris water maze was used to assess learning and memory. Biochemical parameters such as glutathione peroxidase and catalase activities, nitric oxide and malondialdehyde concentrations were measured to evaluate the antioxidant status in the rat hippocampus. Our results indicated that colchicine significantly impaired spatial memory and induced oxidative stress; in contrast, oleuropein pretreatment significantly improved learning and memory retention, and attenuated the oxidative damage. The results clearly indicate that oleuropein has neuroprotective effects against colchicine-induced cognitive dysfunction and oxidative damage in rats.
Keywords: Oleuropein, Colchicine, Alzheimer’s disease, Oxidative stress, Hippocampus
Introduction
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder that induces decline in memory and other cognitive functions in the elderly [8]. The neuropathological signs of AD include deposits of amyloid β fibrils in senile plaques and the presence of abnormal tau protein filaments in the form of neurofibrillary tangles [7]. It has been recognized that β-amyloid aggregates and iron accumulation both synergistically cause oxidative damage by free radical generation [41]. Also, it has been proposed that oxidative stress has an important role in the pathogenesis of AD [27, 41]. The cortex, the limbic system and the hippocampus are parts of the brain which are destroyed in Alzheimer’s disease [28].
Alzheimer’s disease is shown to be associated with microtubule dysfunction and characterized by the appearance of specific cytoskeletal cellular abnormalities, which are associated with cognitive impairment [17]. Colchicine is a plant alkaloid which binds to its site on tubulin and destroys microtubule assembly. This microtubule destroying agent affects the shape and development of the cell and neuronal function like axoplasmic transport, and also increases free radical generation and oxidative damage [15, 40]. It also reduces cholinergic function and is known as a potent inhibitor of behavioral processes, and impairs cognitive function [40]. In this regard, central administration of colchicine to induce cognitive dysfunction is an animal model of senile dementia of Alzheimer’s type [17, 23].
The beneficial effects of antioxidants against oxidative stress are well known, which fortify the immune system [35]. There are various antioxidants such as curcumine and rosmarinic acid with protective effects against Alzheimer’s disease [6, 22]. In this context, olive leaf extract contains antioxidant polyphenols such as oleuropein that has been reported to be a free radical scavenger [42, 43]. Recent studies indicate that oleuropein is a phenolic compound which possesses diverse pharmacological characteristics, e.g., it has vasodilatory, hypotensive, anti-inflammatory, neuroprotective and antioxidant effects [9, 14, 18, 43]. Also, it has been shown that the administration of oleuropein before ischemia–reperfusion in the CA1 area reduces oxidative stress [12]. Therefore, the purpose of this study was to evaluate the pretreatment effects of oleuropein on oxidative status and cognitive dysfunction induced by colchicine in the hippocampal CA1 area in rats.
Materials and methods
Animals
This study was carried out on 35 adult male Wistar rats (weighing 200–220 g). The animals were housed under standard laboratory conditions with a natural 12:12 h light/dark photocycle with food and tap water supplied ad libitum. All rats were treated humanely and in compliance with the recommendations of the Animal Care Committee of the Lorestan University of Medical Sciences.
Drugs and treatments
Colchicine (SDFCL, India) and oleuropein (Dana Kasian, Iran) solutions were made fresh at the beginning of each experiment. Colchicine was prepared in physiological saline as 15 µg/µl and injected intra CA1 hippocampus. Oleuropein was administered in doses of 10, 15 and 20 mg/kg orally once daily for 10 consecutive days before the administration of colchicine. The rats were divided randomly into five groups (n = 7 in each group), as follows: (1) physiological saline + colchicine (colchicine group), (2) oleuropein (10 mg/kg, p.o.) + colchicine, (3) oleuropein (15 mg/kg, p.o.) + colchicine, (4) oleuropein (20 mg/kg, p.o.) + colchicine, and (5) the control group which received equivalent volumes of vehicle for oleuropein (physiological saline) + vehicle for colchicine (physiological saline). All groups except the control group received colchicine with a dosage of 15 µg/rat, once.
Surgery and intra-hippocampal CA1 administration of colchicine
On the 10th day of oleuropein treatment, all animals were anesthetized with chloral hydrate (400 mg/kg, i.p.) (Merck company, Germany) and positioned in a stereotaxic apparatus (Narishige Company, Japan); their skulls were exposed. Two holes were drilled in the skull for bilateral placement of the guide cannula into the hippocampus. Coordinates for the intra hippocampal CA1 cannula implantation were 3.8 mm posterior to bregma, 1.8 mm lateral to the sagittal suture and 2.2 mm ventral to outer skull surface, according to the atlas of Paxinos and Watson [32]. The intracerebral injection was administered through the guide cannula (21-gauge) using injection needles (27-gauge) connected by polyethylene tubing to a 5.0-µl Hamilton microsyringe. The injection needle was inserted 0.5 mm beyond the tip of the cannula. Then 0.5 µl of vehicle or colchicine was injected into each side of CA1 region during 1–2 min and the needle was left in place for an additional 60 s to allow for diffusion of the solution away from the needle tip. Then, the animals were allowed to recover for 1 week before behavioral testing.
Spatial memory assessment
To assess spatial memory, the Morris water maze (MWM) apparatus was used. The MWM consists of a black circular tank with a diameter of 140 cm and a height of 80 cm that is placed in the center of the room with visual cues on the walls and filled with water (temperature around 23 °C) [30, 38]. A hidden black platform, 10 cm in diameter, was located in the water (2 cm below the water surface) and the tank was conceptually divided into four quadrants and had four points designed as starting positions (N, S, W or E). The rats performed four trials per day for four consecutive days. In the swimming trials, each individual rat was placed into the water at a randomly chosen quadrant. During each trial, each rat was given 60 s to find the hidden platform. If a rat found the platform it was allowed to stay on the platform for 15 s and then taken back into the cage. If they could not escape to the platform within 60 s by themselves, the rats were placed on the platform by hand for 15 s and their escape latency was accepted as 60 s. A semi-automatic video tracking system (Radiab-1 software, made by Nomirei, Iran) was used to measure the time to reach the platform (latency), the length of swim path (distance) and the swim speed. On the 5th day, subjects were tested on a probe trial, during which the escape platform was removed and the time spent in the correct quadrant was measured for a 60-s trial. All testing began at 13:00 h.
Biochemical tests
Tissue preparation for assessment of antioxidant enzyme activities and MDA concentration
At the end of the behavioral experiments, the animals were decapitated and the hippocampi were removed quickly, rinsed with saline, and then frozen in a freezer (−80 °C) for 2 months. Then the samples were thawed and homogenized in a cold phosphate buffer (0.1 M, pH 7.4, containing 5 mmol/l EDTA) on liquid nitrogen [5], and debris were removed by centrifugation (Centrifuge 5415 R; Rotofix 32A, Germany). Supernatants were used for the measurement of protein concentration, antioxidant enzyme activities and MDA concentration as lipid peroxidation marker. Protein contents of brain tissue homogenates were determined using a colorimetric method as described by Lowry [24].
The measurement of glutathione peroxidase (GPx) activity
GPx activity was measured by the method described by Rotruck et al. [36]. Briefly, the reaction mixture contained 0.2 ml 0.4 M phosphate buffer (pH = 7), 0.1 ml 10 mmol/l sodium azide, 0.2 ml tissue homogenized in 0.4 M phosphate buffer (pH = 7), 0.2 ml glutathione, and 0.1 ml 0.2 mmol/l H2O2. The contents were incubated for 10 min at 37 °C, 0.4 ml 10 % TCA was added to stop the reaction, and centrifuged at 3200 rpm for 20 min. The supernatant was assayed for glutathione content using Ellman’s reagent [19.5 g 5, 5′-dithiobisnitrobenzoic acid (DTNB) in 100 ml 0.1 % sodium citrate]. The activity was expressed as U/mg protein.
Measurement of catalase (CAT) activity
Catalase activity was estimated using the colorimetric method. The reaction was started by the addition of 20 μl of the sample in 2 ml of 30 mmol/l hydrogen peroxide (H2O2) in 50 mmol/l potassium phosphate buffer (pH 7.0). Each unit of CAT is mmol/l of consumed H2O2 per min-gram protein or milliliter solution. The results are expressed as units per mg of protein (U/mg of protein) [1].
NO assay (nitrite content)
Tissue levels of NO were measured by measuring the accumulation of nitrite in tissue using the Griess reaction with sodium nitrate as the standard. Briefly, 50-μl samples of tissue were mixed with equal volumes of 1 % sulphanilamide and 0.1 % N-1-naphthylethylene diaminedihydrochloride in 0.5 % H3PO4. After 10 min, at room temperature, the absorbance at 540 nm was measured in a microplate reader. Nitrite concentrations were calculated through comparison with a standard calibration curve with sodium nitrite (NaNO2: 0–110 µmol/l) [33].
Levels of malondialdehyde (MDA)
The amount of lipid peroxidation was indicated by the content of thiobarbituric acid reactive substances (TBARS) in the brain homogenates as shown by MDA. Tissue MDA was determined by following the production of thiobarbituric acid reactive substances as described previously [39], which was reported by Alirezaei et al. [5]. In short, 40 µl of homogenate was added to 40 µl of 0.9 % NaCl and 40 µl of deionized H2O, resulting in a total reaction volume of 120 µl. The reaction was incubated at 37 °C for 20 min and stopped by the addition of 600 µl of cold 0.8 M hydrochloride acid, containing 12.5 % trichloroacetic acid. Following the addition of 780 µl of 1 % TBA, the reaction was boiled for 20 min and then cooled at 4 °C for 1 h. In order to measure the amount of TBARS produced by the homogenate, the cooled reaction was spun at 1500×g in a microcentrifuge for 20 min and the absorbance of the supernatant was spectrophotometrically (S2000 UV model; WPA, Cambridge, UK) read at 532 nm, using an extinction coefficient of 1.56 × 105/M cm. The blanks for all of the TBARS assays contained an additional 40 µl of 0.9 % NaCl, instead of homogenate as just described. MDA results were expressed as nanomoles per milligram of tissue protein (nmol/mg protein).
Determination of caspase-3 activity
Caspase-3 activity was measured using a caspase-3 colorimetric assay kit according to the manufacturer’s instructions by a spectrophotometer (Jenway, UV/Vis, Staffordshire, UK). The caspase-3 colorimetric assay is based on the hydrolysis of the peptide substrate acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) by caspase-3, resulting in the release of the p-nitroaniline (pNA) moiety. p-Nitroaniline absorbance is at 405 nm (εmmol/l = 10.5). The concentration of the pNA released from the substrate is calculated from the absorbance values at 405 nm. Enzyme activity was defined as produced micromole pNA/min/milligram tissue protein and expressed as U/mg protein.
Histological verification
Histological verification of the injection sites was carried out in five rats that were randomly chosen from the five groups, one from each. After behavioral testing, those animals were killed with an overdose of chloroform. Each animal received 0.5 µl/site injection of ink (methylene blue solution). The experimental animals’ brains were removed and fixed in formalin solution 10 % for 48 h before sectioning. Sections were examined to determine location of the cannula aimed for the CA1. The cannula placements were verified using the atlas of Paxinos and Watson [32].
Statistics
Results were expressed as mean ± SEM. The behavioral assessment data in the MWM test were determined by one way ANOVA. The biochemical estimations were separately analyzed by one way ANOVA. Post-hoc comparisons between groups were made using an LSD test. The value p < 0.05 was considered significant.
Results
Histology
Figure 1a shows location of cannula position and drug injection sites in the CA1 regions of the hippocampus. The approximate spread of drug assessed by dye (methylene blue solution) injection was verified histologically. Figure 1b also shows a representative section taken from the rat brain atlas of Paxinos and Watson [32].
Fig. 1.
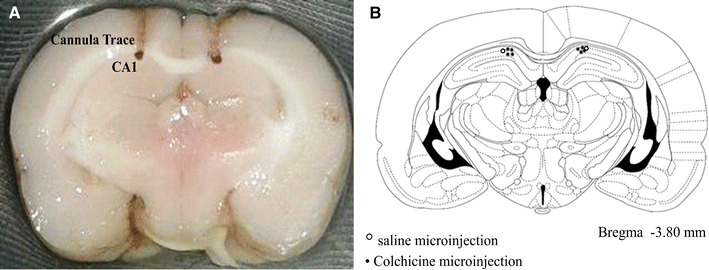
Histological verification of cannula position and drug injection sites in the CA1 area of the hippocampus (a) and verified section taken from the atlas of Paxinos and Watson (b). Open circle saline microinjection, filled circle colchicine microinjection
Effect of oleuropein on spatial navigation task in colchicine-injected rats
Figure 2 illustrates the reduction of latency time and traveled distance to find the hidden platform in all groups during the 4-day training trial. Group comparisons revealed that colchicine-lesioned animals presented a higher latency time and traveled distance than the control group (p < 0.001), showing a poorer learning performance due to CA1-colchicine infusion. Pretreatment with oleuropein significantly improved learning performance in the oleuropein group as compared to the colchicine group (10, 15 mg/kg, p < 0.05, and 20 mg/kg, p < 0.001).
Fig. 2.
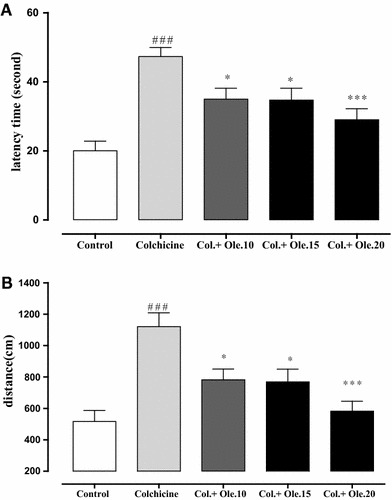
Effects of oleuropein on escape latency (a) and traveled distance (b) in MWM test. Oleuropein at the dosages of 10, 15, and 20 mg/kg significantly decreased the escape latency and traveled distance. Data are the mean ± SEM (n = 7). *p < 0.05 and ***p < 0.001 indicate significant difference versus colchicine group. ### p < 0.001 indicates significant difference versus control group
According to Fig. 3, which illustrates the probe trial test, colchicine-lesioned animals failed to remember the location of the platform, spending less time in the target quadrant than the control group (p < 0.01). However, the time spent in the target quadrant was significantly increased as a result of the administration of oleuropein as compared to the colchicine group (10 mg/kg, p < 0.05; 15, 20 mg/kg, p < 0.01). In addition, there was no significant difference among the swim speeds of all five groups (Fig. 3, p > 0.05).
Fig. 3.

Effects of oleuropein on percentage of time spent in target zone during the probe trials (a) and swimming speed (b) in MWM test. Oleuropein at the dosages of 10, 15, and 20 mg/kg significantly increased time spent in the target zone but there were no significant effects on swimming speed. Data are the mean ± SEM (n = 7). *p < 0.05 and **p < 0.01 indicate significant difference versus colchicine group, ## p < 0.01 indicates significant difference versus control group, NS, there were no significant differences for swim speed between groups (p > 0.05)
Antioxidant activities estimation
MDA
Free radical damage following colchicine injection was assessed using lipid peroxidation (LPO), which was measured as MDA levels. According to Fig. 4, there was an increase in MDA levels of the colchicine group (p < 0.001) as compared to the control group. The oral administration of oleuropein 10, 15, 20 mg/kg resulted in a significant reduction of MDA levels in the brain tissue of colchicine + oleuropein animals as compared to the colchicine group (p < 0.05, p < 0.01).
Fig. 4.

Effects of oleuropein on MDA concentration in brain of rats. Oleuropein at the dosages of 10, 15, and 20 mg/kg significantly decreased MDA concentration. Data are the mean ± SEM (n = 7). *p < 0.05 and **p < 0.01 indicate significant difference versus colchicine group, ### p < 0.001 indicates significant difference versus control group
Glutathione peroxidase (GPx) activity
According to Fig. 5, the GPx activity was significantly (p < 0.001) decreased in the colchicine group as compared to the control group. However, the decrease in the GPx activity was significantly restored by oleuropein treatment (10, 15 mg/kg, p < 0.05 and 20 mg/kg, p < 0.01 ).
Fig. 5.

Effects of oleuropein on GPx activity in brain of rats. Oleuropein at the dosages of 10, 15, 20 mg/kg increased GPx activity. Data are the mean ± SEM (n = 7). *p < 0.05 and **p < 0.01 indicate significant difference versus colchicine group, ### p < 0.001 indicates significant difference versus control group
Catalase (CAT) activity
According to Fig. 6, the CAT activity was significantly (p < 0.05) decreased in the colchicine group as compared to the control group. Administration of oleuropein 20 mg/kg resulted in a significant increase in catalase activity compared to the colchicine group (p < 0.01).
Fig. 6.

Effects of oleuropein on CAT activity in brain of rats. Oleuropein at the dose of 20 mg/kg increased CAT activity. Data are the mean ± SEM (n = 7). **p < 0.01 indicates significant difference versus colchicine group, # p < 0.05 indicates significant difference versus control group
Nitrite estimation
According to Fig. 7, administration of colchicine significantly increased tissue nitrite level when compared to the control group (p < 0.001). Treatment with oleuropein 20 mg/kg significantly attenuated the increase in tissue nitrite induced by colchicine (p < 0.001).
Fig. 7.
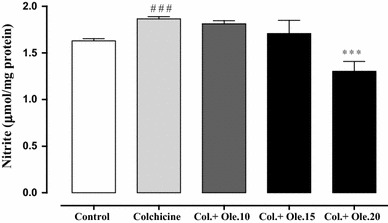
Effects of oleuropein on nitrite levels in brain of rats. Oleuropein at the dose of 20 mg/kg decreased nitrite levels. Data are the mean ± SEM (n = 7). ***p < 0.001 indicates significant difference versus colchicine group, ### p < 0.001 indicates significant difference versus control group
Effect of oleuropein on caspase-3 activity
The protective effect of oleuropein was confirmed by assaying the apoptotic response, in terms of caspase-3 activation, in rats treated with colchicine. According to Fig. 8, administration of colchicine significantly (p < 0.001) increased the activity of caspase-3 when compared to the control group. The groups of animals treated with oleuropein 10, 15 and 20 mg/kg showed a significant decrease (p < 0.001) in caspase-3 activity compared to the colchicine group.
Fig. 8.

Effects of oleuropein on caspase-3 activity in brain of rats. Oleuropein at the doses of 10, 15 and 20 mg/kg decreased caspase-3 activity. Data are the mean ± SEM (n = 7). ***p < 0.001 indicates significant difference versus colchicine group, ### p < 0.001 indicates significant difference versus control group
These results indicate that oleuropein has protective antioxidant effects against colchicine-induced neuron damage. The neuroprotective effects of oleuropein are well correlated with its antioxidant and anti-apoptotic properties.
Discussion
Alzheimer’s disease (AD) is a fatal neurodegenerative disorder manifested by early cognitive dysfunction associated later by behavioral and social deterioration. The hippocampus is the primary neuronal injury region involved in the pathophysiology of the disease [28]. Earlier studies indicated that centrally administered colchicine induced an animal model of senile dementia of Alzheimer’s type in humans [17]. Hence, the goal of the present study was to clarify the possible neuroprotective role of oleuropein in an animal model of AD induced by colchicine. The present study showed that the injection of colchicine into CA1 hippocampus resulted in significant memory deficits in MWM tasks, increased the escape latency time and traveled distance, indicating that colchicine could diminish memory in rats. The results of the current study also showed that injection of colchicine causes an increase in free radical generation leading to oxidative stress that induces cognitive dysfunction. The effects of colchicine on cognitive function have been investigated in previous studies [40, 43]. These findings are in agreement with our previously described harmful effects of colchicine on hippocampal CA1 in treated rats [34].
Colchicine is also a well-known alkaloid derivative which binds tightly to microtubules, causing their depolymerization and inhibiting their assembly. This leads to reduced intracellular trafficking, and in turn leads to the death of nerve cells [26]. It induces neurofibrillary degeneration which is associated with loss of cholinergic neurons in the brain, resulting in impairment of learning function [23]. These effects of colchicine can be induced by its ability to cause apoptosis in neuronal populations by activating the c-Jun N-terminal kinase (JNK) pathway [31].
Experimentally, it has been shown that central administration of colchicine increases the expression of inducible nitric oxide synthase which increases nitric oxide (NO) levels, the peroxynitrite precursor, that results in neuronal damage. Colchicine also elevates the glutamate/GABA ratio in the cerebral cortex of mice [14, 43]; an increase in glutamate leads to free radical-induced neurotoxicity [9]. Thus, nitrite content measured in this study indicates nitrosative stress in the colchicine group when compared to the oleuropein-pretreated rats.
On the other hand, the present study revealed that pretreatment with oleuropein led to a significant improvement of the retention performance of the spatial navigation task in MWM, as evident by the significant decrease in latency time and increase in the time spent in the target quadrant (Figs. 2, 3) and attenuated colchicine-induced oxidative stress. The neuroprotective effects of the oleuropein against oxidative stress were also evaluated by measuring the MDA concentration, nitrite level and CAT and GPx activities. Results of the present study showed that the administration of oleuropein for 10 consecutive days significantly inhibited lipid peroxidation and decreased MDA and nitrite levels in the colchicine treated rats, indicating an antinitrosative stress effect of oleuropein (Figs. 4, 7), and also elevated the CAT and GPx activities compared to the colchicine treated group (Figs. 5, 6), which is in agreement with the results of Sarbishegi et al. [37]. Earlier studies suggested that oleuropein has neuroprotective effects which could be linked to its potent antioxidant actions [18, 29]. Results of the present study also indicated that oleuropein acted as a suppressor of oxidative stress, which is in accordance with previous studies [2–4]. Daccache et al. proposed that this polyphenol acts as a tau aggregation inhibitor and other studies also revealed anti-amyloidogenic and anti-inflammatory effects of oleuropein that confirm its beneficial properties against neurodegeneration [11, 20, 25]. It has also been shown that olive leaf extract decreases age-induced oxidative stress in major organs of aged rats [10]. Another study has shown that oleuropein is capable of interfering with the toxic amyloid aggregation of Aβ in the C. elegans model of AD [13].
Increased oxidative damage triggers various intracellular signals that lead to apoptosis. In vivo and in vitro studies have suggested that colchicine induces caspase-3-dependent apoptotic cell death of granule cells in the dentate gyrus and upregulates cyclooxygenase-2 expression [19]. In this study, it has been shown that colchicine induces apoptosis through increasing the caspase-3 activity in CA1 neurons. The mitochondria are well known to trigger an apoptotic signaling pathway that leads to the release of mitochondrial constituents such as cytochrome c, which in fact stimulates caspase-3 activity [21]. It has previously been demonstrated that colchicine increases the release of cytochrome c in primary cultures of cerebellar granule cells [16]. Therefore, these cytoskeleton disruptors may directly alter mitochondrial functions and thereby initiate the apoptosis pathway in dentate gyrus neurons. The findings of our study showed that treatment with oleuropein reduced caspase-3 activity and provided protection against colchicine-induced apoptosis (Fig. 8). This, thereby, provides the evidence for the neuroprotective effects of oleuropein.
Therefore, we conclude that oleuropein protects neurons against colchicine-induced oxidative damage and appears to be a good candidate in the prevention of colchicine-induced injuries in the hippocampus. Further investigation to elucidate the neurochemical and molecular mechanisms involved in the neuroprotective effects of oleuropein is recommended.
Acknowledgments
This research was financially supported by Razi Herbal Medicines Research Center. We are most grateful to Dr. B. Rasoulian for his kind cooperation in Razi Herbal Medicines Research Center, Lorestan University of Medical Sciences and to Mr. M. Almasian for proofreading the manuscript.
References
- 1.Ahmadvand H, Tavafi M, Khosrowbeygi A. Amelioration of altered antioxidant enzymes activity and glomerulosclerosis by coenzyme Q10 in alloxan-induced diabetic rats. J Diabetes Complicat. 2012;26(6):476–482. doi: 10.1016/j.jdiacomp.2012.06.004. [DOI] [PubMed] [Google Scholar]
- 2.Alirezaei M, Dezfoulian O, Kheradmand A, Neamati S, Khonsari A, Pirzadeh A. Hepatoprotective effects of purified oleuropein from olive leaf extract against ethanol-induced damages in the rat. Iran J Vet Res. 2012;13:218–226. [Google Scholar]
- 3.Alirezaei M, Dezfoulian O, Neamati S, Rashidipour M, Tanideh N, Kheradmand A. Oleuropein prevents ethanol-induced gastric ulcers via elevation of antioxidant enzyme activities in rats. J Physiol Biochem. 2012;68(4):583–592. doi: 10.1007/s13105-012-0177-8. [DOI] [PubMed] [Google Scholar]
- 4.Alirezaei M, Dezfoulian O, Sookhtehzari A, Asadian P, Khoshdel Z. Antioxidant effects of oleuropein versus oxidative stress induced by ethanol in the rat intestine. Comp Clin Pathol. 2014;23:1359–1365. doi: 10.1007/s00580-013-1791-8. [DOI] [Google Scholar]
- 5.Alirezaei M, Khoshdel Z, Dezfoulian O, Rashidipour M, Taghadosi V. Beneficial antioxidant properties of betaine against oxidative stress mediated by levodopa/benserazide in the brain of rats. J Physiol Sci. 2015;65(3):243–252. doi: 10.1007/s12576-015-0360-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Alkam T, Nitta A, Mizoguchi H, Itoh A, Nabeshima T. A natural scavenger of peroxynitrites, rosmarinic acid, protects against impairment of memory induced by Aβ 25–35. Behav Brain Res. 2007;180(2):139–145. doi: 10.1016/j.bbr.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 7.Anand R, Gill KD, Mahdi AA. Therapeutics of Alzheimer’s disease: past, present and future. Neuropharmacology. 2014;76:27–50. doi: 10.1016/j.neuropharm.2013.07.004. [DOI] [PubMed] [Google Scholar]
- 8.Auld DS, Kornecook TJ, Bastianetto S, Quirion R. Alzheimer’s disease and the basal forebrain cholinergic system: relations to β-amyloid peptides, cognition, and treatment strategies. Prog Neurobiol. 2002;68(3):209–245. doi: 10.1016/S0301-0082(02)00079-5. [DOI] [PubMed] [Google Scholar]
- 9.Bondy SC. The relation of oxidative stress and hyperexcitation to neurological disease. Proc Soc Exp Biol Med. 1995;208(4):337–345. doi: 10.3181/00379727-208-43862. [DOI] [PubMed] [Google Scholar]
- 10.Çoban J, Öztezcan S, Doğru-Abbasoğlu S, Bingül I, Yeşil-Mizrak K, Uysal M. Olive leaf extract decreases age-induced oxidative stress in major organs of aged rats. Geriatr Gerontol Int. 2014;14(4):996–1002. doi: 10.1111/ggi.12192. [DOI] [PubMed] [Google Scholar]
- 11.Daccache A, Lion C, Sibille N, Gerard M, Slomianny C, Lippens G, Cotelle P. Oleuropein and derivatives from olives as tau aggregation inhibitors. Neurochem Int. 2011;58(6):700–707. doi: 10.1016/j.neuint.2011.02.010. [DOI] [PubMed] [Google Scholar]
- 12.Dekanski D, Selaković V, Piperski V, Radulović Ž, Korenić A, Radenović L. Protective effect of olive leaf extract on hippocampal injury induced by transient global cerebral ischemia and reperfusion in Mongolian gerbils. Phytomedicine. 2011;18(13):1137–1143. doi: 10.1016/j.phymed.2011.05.010. [DOI] [PubMed] [Google Scholar]
- 13.Diomede L, Rigacci S, Romeo M, Stefani M, Salmona M. Oleuropein aglycone protects transgenic C. elegans strains expressing Aβ42 by reducing plaque load and motor deficit. PLoS One. 2013;8(3):e58893. doi: 10.1371/journal.pone.0058893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dufourny L, Leroy D, Warembourg M. Differential effects of colchicine on the induction of nitric oxide synthase in neurons containing progesterone receptors of the guinea pig hypothalamus. Brain Res Bull. 2000;52(5):435–443. doi: 10.1016/S0361-9230(00)00286-0. [DOI] [PubMed] [Google Scholar]
- 15.Goldschmidt RB, Steward O. Neurotoxic effects of colchicine: differential susceptibility of CNS neuronal populations. Neuroscience. 1982;7(3):695–714. doi: 10.1016/0306-4522(82)90075-6. [DOI] [PubMed] [Google Scholar]
- 16.Gorman AM, Bonfoco E, Zhivotovsky B, Orrenius S, Ceccatelli S. Cytochrome c release and caspase-3 activation during colchicine-induced apoptosis of cerebellar granule cells. Eur J Neurosci. 1999;11(3):1067–1072. doi: 10.1046/j.1460-9568.1999.00512.x. [DOI] [PubMed] [Google Scholar]
- 17.Iqbal K, Zaidi T, Wen GY, Grundke-Iqbal I, Merz PA, Shaikh SS, Wisniewski HM, Alafuzoff I, Winblad B. Defective brain microtubule assembly in Alzheimer’s disease. Lancet. 1986;328:421–426. doi: 10.1016/S0140-6736(86)92134-3. [DOI] [PubMed] [Google Scholar]
- 18.Khalatbary AR, Ahmadvand H. Neuroprotective effect of oleuropein following spinal cord injury in rats. Neurol Res. 2012;34(1):44–51. doi: 10.1179/1743132811Y.0000000058. [DOI] [PubMed] [Google Scholar]
- 19.Kim JA, Mitsukawa K, Yamada MK, Nishiyama N, Matsuki N, Ikegaya Y. Cytoskeleton disruption causes apoptotic degeneration of dentate granule cells in hippocampal slice cultures. Neuropharmacology. 2002;42(8):1109–1118. doi: 10.1016/S0028-3908(02)00052-7. [DOI] [PubMed] [Google Scholar]
- 20.Kostomoiri M, Fragkouli A, Sagnou M, Skaltsounis LA, Pelecanou M, Tsilibary EC, Tzinia AK. Oleuropein, an anti-oxidant polyphenol constituent of olive promotes α-secretase cleavage of the amyloid precursor protein (AβPP) Cell Mol Neurobiol. 2013;33(1):147–154. doi: 10.1007/s10571-012-9880-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kroemer G, Dallaporta B, Resche-Rigon M. The mitochondrial death/life regulator in apoptosis and necrosis. Annu Rev Physiol. 1998;60:619–642. doi: 10.1146/annurev.physiol.60.1.619. [DOI] [PubMed] [Google Scholar]
- 22.Kumar A, Naidu PS, Seghal N, Padi SS. Effect of curcumin on intracerebroventricular colchicine-induced cognitive impairment and oxidative stress in rats. J Med Food. 2007;10(3):486–494. doi: 10.1089/jmf.2006.076. [DOI] [PubMed] [Google Scholar]
- 23.Kumar A, Seghal N, Naidu PS, Padi SS, Goyal R. Colchicine-induced neurotoxicity as an animal model of sporadic dementia of Alzheimer’s type. Pharmacol Rep. 2007;59(3):274–283. [PubMed] [Google Scholar]
- 24.Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 25.Luccarini I, Dami T, Grossi C, Rigacci S, Stefani M, Casamenti F. Oleuropein aglycone counteracts Aβ42 toxicity in the rat brain. Neurosci Lett. 2014;558:67–72. doi: 10.1016/j.neulet.2013.10.062. [DOI] [PubMed] [Google Scholar]
- 26.Mandelkow EM, Stamer K, Vogel R, Thies E, Mandelkow E. Clogging of axons by tau, inhibition of axonal traffic and starvation of synapses. Neurobiol Aging. 2003;24(8):1079–1085. doi: 10.1016/j.neurobiolaging.2003.04.007. [DOI] [PubMed] [Google Scholar]
- 27.Markesbery WR. Oxidative stress hypothesis in Alzheimer’s disease. Free Radical Biol Med. 1997;23:134–147. doi: 10.1016/S0891-5849(96)00629-6. [DOI] [PubMed] [Google Scholar]
- 28.McIlroy S, Craig D. Neurobiology and genetics of behavioural syndromes of Alzheimer’s disease. Curr Alzheimer Res. 2004;1(2):135–142. doi: 10.2174/1567205043332180. [DOI] [PubMed] [Google Scholar]
- 29.Mohagheghi F, Bigdeli MR, Rasoulian B, Hashemi P, Pour MR. The neuroprotective effect of olive leaf extract is related to improved blood–brain barrier permeability and brain edema in rat with experimental focal cerebral ischemia. Phytomedicine. 2011;18:170–175. doi: 10.1016/j.phymed.2010.06.007. [DOI] [PubMed] [Google Scholar]
- 30.Morris R, Garrud P, Rawlins JN, O’Keefe J. Place navigation impaired in rats with hippocampal lesions. Nature. 1982;297(5868):681–683. doi: 10.1038/297681a0. [DOI] [PubMed] [Google Scholar]
- 31.Müller GJ, Geist MA, Veng LM, Willesen MG, Johansen FF, Leist M, Vaudano E. A role for mixed lineage kinases in granule cell apoptosis induced by cytoskeletal disruption. J Neurochem. 2006;96(5):1242–1252. doi: 10.1111/j.1471-4159.2005.03590.x. [DOI] [PubMed] [Google Scholar]
- 32.Paxinos G, Watson C. The rat brain in stereotaxic coordinates. New York: Academic; 1986. [DOI] [PubMed] [Google Scholar]
- 33.Petrola MJ, de Castro AJ, Pitombeira MH, Barbosa MC, Quixadá AT, Duarte FB, Goncalves RP. Serum concentrations of nitrite and malondialdehyde as markers of oxidative stress in chronic myeloid leukemia patients treated with tyrosine kinase inhibitors. Rev Bras Hematol Hemoter. 2012;34(5):352–355. doi: 10.5581/1516-8484.20120091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Porkhodadad S, Karami M, Jalali MR. Positive effect of nitric oxide on morphine-induced place conditioning in Wistar rats treated by colchicine intra-hippocampal CA1. J Clin Toxicol. 2011 [Google Scholar]
- 35.Rajadurai M, Stanely Mainzen Prince P. Preventive effect of naringin on lipid peroxides and antioxidants in isoproterenol-induced cardiotoxicity in Wistar rats: biochemical and histopathological evidences. Toxicology. 2006;228:259–268. doi: 10.1016/j.tox.2006.09.005. [DOI] [PubMed] [Google Scholar]
- 36.Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, Hoekstra WG. Selenium: biochemical role as a component of glutathione peroxidase. Science. 1973;179(4073):588–590. doi: 10.1126/science.179.4073.588. [DOI] [PubMed] [Google Scholar]
- 37.Sarbishegi M, Mehraein F, Soleimani M. Antioxidant role of oleuropein on midbrain and dopaminergic neurons of substantia nigra in aged rats. Iran Biomed J. 2014;18(1):16–22. doi: 10.6091/ibj.1274.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Stackman RW, Hammond RS, Linardatos E, Gerlach A, Maylie J, Adelman JP, Tzounopoulos T. Small conductance Ca2+-activated K+ channels modulate synaptic plasticity and memory encoding. J Neurosci. 2002;22(23):10163–10171. doi: 10.1523/JNEUROSCI.22-23-10163.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Subbarao KV, Richardson JS, Ang LC. Autopsy samples of Alzheimer’s cortex show increased peroxidation in vitro. J Neurochem. 1990;55(1):342–345. doi: 10.1111/j.1471-4159.1990.tb08858.x. [DOI] [PubMed] [Google Scholar]
- 40.Veerendra Kumar MH, Gupta YK. Intracerebroventricular administration of colchicine produces cognitive impairment associated with oxidative stress in rats. Pharmacol Biochem Behav. 2002;73(3):565–571. doi: 10.1016/S0091-3057(02)00838-9. [DOI] [PubMed] [Google Scholar]
- 41.Viña J, Lloret A, Ortí R, Alonso D. Molecular bases of the treatment of Alzheimer’s disease with antioxidants: prevention of oxidative stress. Mol Aspects Med. 2004;25:117–123. doi: 10.1016/j.mam.2004.02.013. [DOI] [PubMed] [Google Scholar]
- 42.Waterman E, Lockwood B. Active components and clinical applications of olive oil. Altern Med Rev. 2007;12(4):331–342. [PubMed] [Google Scholar]
- 43.Yu Z, Cheng G, Hu B. Mechanism of colchicine impairment on learning and memory, and protective effect of CGP36742 in mice. Brain Res. 1997;750:53–58. doi: 10.1016/S0006-8993(96)01158-4. [DOI] [PubMed] [Google Scholar]