

Abstract
The circadian nature of physiology and behavior is regulated by a circadian clock that generates intrinsic rhythms with a periodicity of approximately 24 h. The mammalian circadian system is composed of a hierarchical multi-oscillator structure, with the central clock located in the suprachiasmatic nucleus (SCN) of the hypothalamus regulating the peripheral clocks found throughout the body. In the past two decades, key clock genes have been discovered in mammals and shown to be interlocked in transcriptional and translational feedback loops. At the cellular level, each cell is governed by its own independent clock; and yet, these cellular circadian clocks in the SCN form regional oscillators that are further coupled to one another to generate a single rhythm for the tissue. The oscillatory coupling within and between the regional oscillators appears to be critical for the extraordinary stability and the wide range of adaptability of the circadian clock, the mechanism of which is now being elucidated with newly advanced molecular tools.
Keywords: Circadian clock, Suprachiasmatic nucleus, Oscillatory coupling, Clock gene, Luciferase reporter
Introduction
Robust circadian rhythms in physiological functions and behaviors are conserved across all organisms, from cyanobacteria to humans. These rhythms are driven by an intrinsic machinery called the circadian clock, which generates self-sustaining rhythms with a periodicity of about (circa) 1 day (dies). In the last two decades, we have come to appreciate the underlying mechanisms of the mammalian circadian clock at the molecular, cellular, network, and organismal levels. Indeed, the 2017 Nobel Prize for Physiology and Medicine was awarded to three chronobiologists who first cloned the Drosophila clock gene, Period (Per) in 1984 [1, 2]. The first mammalian clock gene, clock, was successfully cloned in 1997 [3] and, owing to the remarkable similarities of the circadian pacemaker system between mammals and Drosophila, our understanding of the clock mechanism has advanced rapidly.
Characteristics of circadian rhythms
For a biological process to be considered circadian, it must exhibit the following three characteristics: an endogenous free-running cycle, entrainability, and temperature compensation. Under constant environmental conditions, such as continuous darkness (DD) under constant temperature, a circadian rhythm exhibits intrinsic, self-sustaining cycles with a period close to but slightly deviated from 24 h [4]. This cycle is called the free-running rhythm, and distinguishes the process from one that is merely a reaction to external signals. Free-running rhythms are observed not only at the organism level, e.g., sleep-wakefulness and behavior (Fig. 1a), but also at the cellular level, such as in fluctuations of gene/protein expression and the release of hormones and neurotransmitters (Fig. 1c) [4].
Fig. 1.
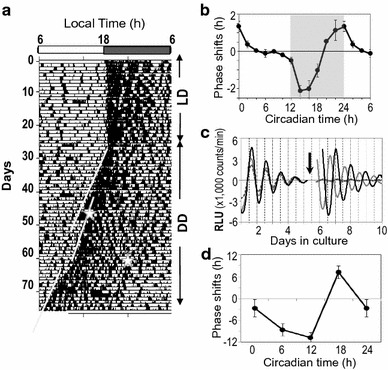
Phase–response curves (PRCs) of the behavior rhythm and the peripheral clock of mice. a Behavioral activity of a C57BL/6 J mouse monitored by an area sensor under light–dark (LD) and continuous darkness (DD) cycles. Under DD, the mouse showed a stable free-running rhythm that phase-dependently shifted by light pulses (30 min, 300 lx) at circadian times (CT) 14 (on day 48) and CT22 (on day 68). The onset phase of behavior activity is designated as CT12. White stars in the actogram indicate the timing of the light pulses. Regression lines fitted to the onset of consecutive activity before and after the light pulse demonstrate that the light induced a phase-delay at CT14 and a phase-advance at CT22. b PRC of behavioral rhythm after light pulses (mean and standard error) demonstrates that light pulses (30 min, 300 lx) during early subjective night induce a phase-delay (negative number of the ordinate), while light pulses during the subjective late night to early morning induce phase-advance shifts (positive number of the ordinate). No shift is observed when a light pulse is administered during subjective day. Shaded area, subjective night. c Cultured nasal mucosa of a mouse carrying a luciferase reporter for PER2 exhibits robust circadian rhythm in PER2::LUCIFERASE (PER2::LUC) levels, which is reset by dexamethasone (Dex). Data was collected using a photomultiplier tube and is expressed in relative light units (RLU). An arrow indicates the time of Dex or vehicle treatment. Original data was detrended by subtracting the 24-h moving average. Solid lines indicate cultures treated with Dex at different phases, and the dashed line indicates cultures treated with vehicle. d PRC of the nasal clock shows a large phase-advance (approximately 10 h) and phase-delay shifts (approximately − 12 h) (see Ref. [114])
Although circadian rhythms are intrinsic, living organisms are also under the influence of cyclic changes of environments, such as day–night cycles and seasonal changes in photoperiods and temperature. The alignment of the circadian rhythm to such environmental cues or “Zeitgebers” to maintain periodicity is referred to as entrainment. For all organisms, light is the strongest Zeitgeber. The circadian clock entrains to a 24-h light–dark (LD) cycle resulting in diurnal, nocturnal, or crepuscular behavior.
The circadian clock can be phase-shifted by any of these time cues, and the extent and direction (advance or delay) of the shifts depend on the phase of the circadian clock at which the cues are introduced. The phase–response curve (PRC) summarizes the relationship between the time when the cue, such as light (Fig. 1b) and dexamethasone (Fig. 1d), is introduced and the induced phase shifts [5]. Using the PRC, we can predict how the clock will respond to the environment, and we can calculate the time course of re-entrainment after changes in the LD cycle, as well as the limit of entrainment [6]. For example, a single light pulse at the subjective early night induces a phase-delay shift, while a pulse during the subjective late night to morning results in a phase-advance shift. No shift is observed when a pulse is given during the middle of subjective day. Interestingly, the shape of the PRC has been shown to be remarkably similar across all living organisms recorded thus far [7], which strongly suggests that the circadian clock developed very early during evolution. Experiments using cyanobacteria mutants with different circadian periodicities have demonstrated the evolutionary advantage of having a circadian clock [8]. Without competitors, cyanobacteria mutants with a short or prolonged periodicity proliferated at the same speed as wild type cyanobacteria. However, when populations with different periodicities were mixed, the colony with a circadian period most similar to the environmental periodicity eventually outpaced their unsynchronized counterparts, highlighting the importance of matching environmental cues to life itself.
Finally, a circadian rhythm requires the ability to compensate for changes in temperature to maintain periodicity. Typically, a circadian oscillation would have a Q10 coefficient (a measure of the rate of change in a biological system dependent upon temperature) of approximately 1.0, meaning that the periodicity is not affected by changes in increasing temperatures that can affect the kinetics of cellular processes [9]. In homothermic animals like mammals and birds, the significance of temperature compensation was initially overlooked as the central circadian clock is located within the central nervous system where the temperature is held constant. However, the discovery of peripheral clocks has reignited the field to research the importance of temperature compensation. While the mechanism of how the circadian clock compensates for the change in temperature is not completely understood, it has been shown that the structure of a clock protein KaiC in cyanobacteria is critical for temperature compensation by slowing down the speed of oscillation and reducing energy expenditure [10]. Furthermore, casein kinase 1 ε was demonstrated to play an important role in temperature compensation in mammals [11].
The mammalian circadian system is composed of a central clock located in the suprachiasmatic nucleus (SCN) of the hypothalamus that regulates additional circadian rhythms found in non-SCN brain regions and in peripheral organs throughout the body [12–14]. The SCN is the only clock that is reset by the light from the environment; the photic signals are transferred to the SCN via the retinohypothalamic tract (RHT). Once the SCN pacemaker is light-entrained, it synchronizes the peripheral clocks to the same 24-h cycle through mechanisms that remain largely unknown.
The circadian clock allows us to prepare and adapt our physiology to cyclic changes in our environment. Notably, ablation of the circadian clock does not directly terminate the life of organisms, but without a fully functioning clock, the organisms appear to become vulnerable when placed in a naturally or socially competitive environment. The circadian clock thus plays a critical role in the survival of individuals and of species as a whole.
Molecular machinery that generates circadian rhythm at a cellular level
Clock genes and molecular feedback
The first known mammalian clock gene, Clock, encoding a basic helix-loop-helix (bHLH) Period-Arnt-Sim (PAS)-type transcription factor, was cloned in 1997 [3], which was soon followed by the cloning of the clock genes Period 1 (Per1) [15] and Bmal1 [16, 17], encoding a PAS protein and a bHLH-PAS transcription factor, respectively. BMAL1 heterodimerizes with CLOCK and binds to an E-box enhancer (CANNTG) site upstream of the Per gene, inducing Per transcription. The PER protein then translocates to the nucleus with cryptochrome (CRY) 1 and 2 and binds to the CLOCK/BMAL1 heterodimer to inhibit enhancer activity [18]. One cycle of this negative feedback loop takes about 24 h, thus generating circadian rhythmicity (Fig. 2). CRY was initially identified as a photoreceptor in fungi and plants [19, 20], although in mammals, CRY1 and CRY2 have no such photoreceptor function [21]. In mammals, the Cry1 and 2 genes are also regulated by an E-box enhancer.
Fig. 2.
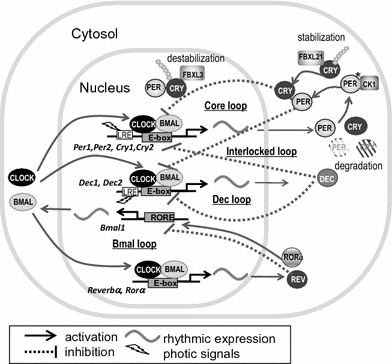
Molecular clock of a mammalian cell. Molecular machinery of the circadian clock is composed of interlocked molecular feedback loops. The core loop is composed of positive elements (CLOCK and BMAL1) and negative elements (PERs and CRYs). The loop is interlocked antiphasic to the Bmal1 loop (positive element: ROR; negative element: REB-ERBα) and in phase with the Dec loop (positive elements: CLOCK and BMAL1; negative elements: DECs). Light-responsive elements such as cAMP response element (CRE), to which phosphorylated CREB binds to induce transcription, exists upstream of Per1, Per2, and Dec1 transcription start sites. Post-transcriptional and post-translational modifications such as ubiquitination and phosphorylation are important for regulating the circadian period
Due to the similarities between the mammalian and Drosophila clock [1, 2], key members of transcriptional and translational feedbacks were quickly identified in mammals. And in the case of Clock and Bmal1 (called Cycle in Drosophila), Drosophila orthologs were cloned after the cloning of mammalian genes [22, 23]. However, it is important to note that there are also a few differences between mammals and Drosophila. For example, mammals have three Per homologs, Per1, Per2, and Per3, each of them playing a specific role [24]. Furthermore, in Drosophila, TIMELESS protein exhibits photosensitivity and plays a role in regulating the circadian rhythm. In contrast, the mammalian TIMELESS has been shown to regulate cell cycle and carcinogenesis [25], and its role in the circadian rhythm has remained elusive.
In addition to the core molecular feedback loop consisting of PER1, PER2, CRY1, CRY2, CLOCK, and BMAL1, interlocking feedback loops have also been identified to regulate circadian rhythms. One example is the Bmal1 loop, which interlocks with the core loop [26] and exhibits an antiphasic gene expression pattern [17]. Bmal1 is regulated by a retinoic acid receptor-related orphan receptor (ROR) enhancer site located upstream of the Bmal1 gene; ROR binding activates gene expression, while REB-ERBα binding inhibits transcription [27]. REB-ERBα itself is regulated by an E-box enhancer located upstream of its transcription start site, resulting in an expression pattern that subsequently puts Bmal1 completely out of phase with Per1 and Per2 [28]. Dec1 and Dec2 are two other elements of an interlocking loop found to negatively affect circadian rhythms [29]. Both Dec1 and Dec2 have E-box enhancers and are positively regulated by CLOCK and BMAL1, and negatively by PER1, PER2, CRY1, and CRY2. Furthermore, the transcription of Dec1 and Dec2 is self-regulated by their encoded proteins. DECs also form a mutually interlocked molecular loop with the Per loop. These interlocked multiple molecular loops are thought to be advantageous by providing stability and fine tuning to the circadian periodicity [30].
Molecular mechanisms of light entrainment
The circadian clock in the SCN is reset by photic signals that are received by melanopsin-expressing, intrinsically photosensitive retinal ganglion cells (ipRGCs) in the retina [31–33] and are transmitted via the RHT [34]. Among the five subtypes of ipRGCs (M1–M5), the M1 ipRGCs predominantly (and, to a lesser extent, the M2 ipRGCs) project photic signals to the SCN to entrain the circadian clock [35]. ipRGCs are also involved in light-induced pupil constriction, masking behavior in response to light, and even certain kinds of image-forming visual pathways [36–39].
The molecular mechanisms of light entrainment have been intensively studied. Photic signals are projected to the SCN core where glutamate, the neurotransmitter of the RHT, induces phosphorylation of Ca2+-cAMP response element-binding (CREB) protein, resulting in the transcription of some key clock genes (Fig. 2). Photic induction of Per1 and Per2 [40, 41], but not Per3 [24], is detected only during the photosensitive zone of PRC (Fig. 1). Dec1, but not Dec2, expression is also induced by phase-resetting light stimuli [29]. Photic signals administered during the dead-zone of PRC do not induce Per or Dec expression and do not lead to shifts in behavioral rhythms. Interestingly, light induction of these gene expressions is observed at the core of the SCN where the circadian clock gene expression level is low, whereas circadian gene expression is high in the shell region of the SCN [40]. Among these genes, Per1 and Dec1 expressions are induced by lights with the kinetics comparable to those of an immediately early gene. Furthermore, Per1 antisense injection into the SCN blocked light-induced phase shifts, suggesting a critical role of Per1 in mediating light entrainment. Per and Dec genes exhibit robust circadian expression rhythms in the SCN with a peak located during the subjective day, i.e., the dead-zone of PRC (Fig. 2). When light is given in subjective evening when Per expression is decreasing, light-induced gene expression may elongate this part of the cycle, resulting in a phase-delay of the molecular clock [40]. In contrast, when light is given in the subjective morning hours when Per expression is increasing, light-induced gene expression makes an early start of the rising phase of the cycle, resulting in the advancement of the molecular clock. Since the gene expression would already be high during the subjective day at the dead-zone of PRC, we would not expect a light-induced shift of the molecular clock.
Although this mechanism explains light-induced phase shifts and light entrainment, Per1, Per2, or Dec1 knockout mice show LD-entrained behavioral rhythms similar to those of wild type mice, suggesting redundancy in gene functions or alternative mechanisms. Among the light-inducible clock genes, Per1 and Dec1 are expressed immediately following light exposure, and their mRNAs have a relatively short half-life [29, 40]. Subsequently, Per2 expression is observed, and its high mRNA level is maintained for a longer period than for Per1 [41, 42]. Thus, it appears that Per1 and Per2 have non-redundant roles in light entrainment.
Post-transcriptional and post-translational regulation of molecular oscillation
Recent studies have demonstrated that 70–80% of mRNA demonstrating circadian rhythms in their content did not exhibit circadian rhythm in their de novo transcription [43], indicating a critical role of post-transcriptional regulation in the molecular circadian clock. For example, an RNA editing enzyme, Adarb1, has been shown to have a circadian expression pattern. As a result, its downstream targets also exhibited post-transcriptionally regulated oscillations [44].
Post-translational modifications also affect the circadian rhythm, especially the period length. Tau was the first mammalian clock gene identified in a mutant hamster that displayed a short circadian period [45]. The cloning of tau was delayed compared to that of Clock [3] and Per [15] for many years because it was found in hamsters in which genome information was limited. Tau mutation is a missense mutation in the phosphorylation site of casein kinase 1 ε (CK1ε) [46], a kinase which phosphorylates many clock proteins, including PER and CRY. Soon after the finding of the Tau mutation in hamsters, patients with advanced sleep-phase syndrome (ASPS) with short circadian periods, were shown to possess a missense mutation in the Per2 gene at the CK1ε phosphorylation site [47]. Interestingly, further studies revealed the Tau mutation to be a gain-of-function mutation, which instead induced hyperphosphorylation in other regions of the PER2 protein, resulting in shortening of the molecular oscillation period [48]. Other kinases were also found to affect the periodicity, such as glycogen synthase kinase 3 β [49].
In addition to phosphorylation, other post-translational modifications such as ubiquitination [50, 51], acetylation [52], and SUMOylation [53] have been shown to play important roles in the fine-tuning of the circadian period. For example, FBXL3, an F-box-type E3 ligase, ubiquitinates CRY1 and CRY2 in the nucleus, which mediates their degradation and shortens the circadian period (Fig. 2). In contrast, FBXL21 ubiquitinates CRYs in the cytosol and stabilizes them. Thus, FBXL21 in the cytosol counteracts the period-shortening effects of FBXL3 in the nucleus [54, 55]. In addition to the familiar ASPS due to Per2 mutation [47], various mutations have already been identified in different types of sleep–wake rhythm disorders in humans [56–58]. Clock gene functions have also become research targets for mood [59–61] and neurodevelopmental [62–64] disorders that result in disrupted circadian rhythms.
The multiple interacting molecular loops, post-transcriptional, and post-translational modifications of the clock genes increase the stability as well as adaptability of the system. Further study is needed to fully understand the mechanisms that regulate the clock at the molecular and cellular levels.
Technical advances in long-term monitoring of physiological functions
Canonical clock genes such as Per1, Per2, and Bmal1 exhibit robust circadian rhythms in their transcriptional activity and protein levels [15, 17, 41, 65]. To monitor their circadian rhythms in living cells for prolonged periods of time, bioluminescent and fluorescent reporters have been introduced. Among them, luciferase reporters have been widely used to analyze clock gene expression and circadian rhythms because detection of luminescence produced by the luciferin–luciferase reaction requires no excitation. This low phototoxicity is advantageous for long-term, continuous recording of the reporters. To detect these signals, a sensitive photon counting device called a photomultiplier tube (PMT) can be used; however, PMTs lack spatial resolution [14, 66]. To overcome this, a charge coupled device (CCD) camera can be used to image bioluminescence to monitor circadian rhythms at the cellular level. To detect very dim luminescence, CCD and electron multiplying CCD cameras can be cooled to − 60 to −80 ℃ to reduce background noise [67, 68].
Since 1972, it was thought that the SCN was the only site of a circadian clock in mammals [34, 69]. However, recent studies using the luciferase reporter technique revealed that every tissue of the body actually has its own circadian clock, called the peripheral clocks (Fig. 3a) [14, 65]. Furthermore, every single cell in culture, as well as the ones that were frozen and stored in the laboratory for many generations, maintain their cellular clock (Fig. 3b) [70]. Cellular rhythms are synchronized and amplified by dexamethasone treatment, suggesting the glucocorticoids as time cues for peripheral clocks. Moreover, continuous in vivo bioluminescence monitoring (Fig. 3c) in freely moving mice is now available [71], which allows us to examine the responses of the circadian clock to various external stimuli [72, 73]. How such techniques are contributing to the research of mammalian circadian clocks, especially in the SCN, will be described in the following sections.
Fig. 3.

Circadian rhythm of clock gene expression monitored using luciferase reporters. a Bioluminescence rhythms of cultured explants (300-μm-thick slices of the SCN, lung, and liver) and an eye from transgenic mice carrying a Bmal1 promoter-driven luciferase reporter (Bmal1-luc) (see Ref. [115]). Data was monitored by a PMT and expressed in relative light units (RLU). Horizontal gray and black bars on the abscissa indicate the light–dark (LD) cycles to which animals were entrained. b Bmal1-luc rhythm of a single Rat-1 fibroblast cell before and during dexamethasone treatment. c In vivo recording of bioluminescence using fiber optics situated above the SCN (gray line). Behavioral activity (black histogram) was monitored simultaneously from a freely moving mouse carrying a Per1-luc reporter. Horizontal gray and black bars on the abscissa indicate subjective light and dark phases, respectively. Bioluminescence data was smoothed using the 4-h moving average method [72]
The central circadian pacemaker, SCN
Architecture of the SCN
The SCN is a pair of oval nuclei in the anterior hypothalamus, located just above the optic chiasm, and its structure is conserved from rodents to primates. The SCN is cytochemically and functionally classified into two regions, the core (also called the ventrolateral region) and the shell (dorsomedial region) (Fig. 4a, b) [74–76]. The core predominantly consists of vasoactive intestinal polypeptide (VIP) and gastrin-releasing peptide (GRP), and receives major afferents. The RHT from the retina is the most important input and its neurotransmitters are glutamate and PCAP. The core also receives serotonergic inputs from the raphe nuclei and GABA/NPY inputs from the thalamic intergeniculate leaflet. In contrast, the shell consists of primarily arginine vasopressin (AVP) neurons, while a small number of somatostatin neurons are also found. The SCN shell is considered to play an important role in rhythm output. In addition, neuromedin S-containing neurons are distributed both in the core and the shell [77]. Neuromedin S is expressed almost exclusively in the SCN in the brain, although its physiological role in the SCN is still unknown [78]. Interestingly, almost all SCN neurons contain GABA [79, 80]; however, it is also not understood why the central circadian pacemaker would be composed almost entirely of GABAergic neurons.
Fig. 4.
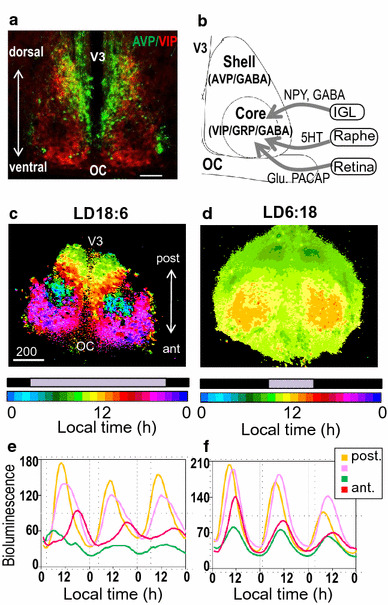
Photoperiodic clock in the SCN. a A coronal section of a mouse SCN immunolabeled with anti-AVP (green) and anti-VIP (red). Scale bar: 200 μm; OC, optic chiasm; V3, third ventricle. b Schematic drawing showing the cytoarchitecture of the SCN with major afferents and their neurotransmitters. Glu, glutamate; NPY, neuropeptide Y. c, d Phase maps for Per1-luc rhythms of horizontal SCN slices from mice housed under light–dark (LD) cycles 18:6 (L:3:00–21:00) (c) and 6:18 (L:9:00–15:00) (d). Pseudocolored bioluminescent images of peak phases of Per1-luc rhythms (each pixel = 3.7 × 3.7 μm). The peak phases are widely distributed in LD18:6 (from approximately 3:00 in the central SCN to 22:00 in the anterior SCN), and are consolidated between 9:00–14:00 in LD6:18. (see Ref. [116]). e, f, Bioluminescence from single pixels of horizontal SCN slices indicate a large regional phase distribution from the posterior (post) to anterior (ant) SCN in LD18:6, but are synchronized across the SCN as a whole in LD18:6 (see Ref. [93])
Regional oscillators and networks
Dispersed SCN neurons exhibit circadian rhythms in their clock gene expression and spontaneous firing [81–83]. Importantly, SCN neurons in wild type animals exhibit independent circadian rhythms, with cell-specific periods with a Gaussian distribution covering a wide range of 20–30 h. In contrast, the circadian periods of SCN neurons in a slice culture are synchronized to one another, and the slice-to-slice variability is much smaller than that observed in dissociated cell culture [84]. Free-running periods of behavior were distributed in an even narrower range than in slices [85]. These findings suggest that in an intact SCN, individual neurons communicate to achieve synchronized periodicity representative of the majority of constitutional cellular oscillators. The coupling of the oscillator network in the SCN is critical for generating a coherent output from the SCN and, subsequently, for the regulation of the peripheral clocks.
Bioluminescence imaging of coronal SCN slices of either transgenic mice expressing a Per1-promoter-driven luciferase reporter (Per1-luc) or knock-in mice carrying a PER2::LUCIFERASE fusion reporter (PER2::LUC) demonstrated robust cellular circadian rhythms with synchronized periodicity. However, the phase distribution within the SCN was specific to topology and changed dynamically in response to a number of perturbing signals, in particular, photic signals [86, 87]. Abrupt phase shifts in an LD cycle induced phase shifts in behavioral rhythm with a transient period of approximately 1 week. This transient period is regarded as the lag between the immediate phase shift of the central clock and the gradual shifts of the peripheral clocks involved in overt rhythm expression. Interestingly, the rate of change in clock gene expression rhythms appears to be region-dependent; a shift was immediately observed in the SCN core following the LD perturbation, while a more gradual change was recorded over several days in the shell [86]. This finding supports the presence of regional oscillators in the SCN, which are synchronized under steady-state conditions, but become distinct in the presence of external stimuli. Previously, we reported the separation of the core and shell rhythms by measuring AVP and VIP releases from an organotypic SCN slice culture [88]. We found that the circadian rhythms of the release of the two peptides were initially synchronized; however, if they were treated with antimitotic drugs at the beginning of the culture, the two systems became gradually desynchronized over time. This demonstrated that the synchrony was kept among the identical peptidergic cells, but AVP and VIP neurons could be separated, suggesting the presence of two types of cellular networks within the SCN.
Another type of regional oscillator was reported in hamsters exhibiting split rhythms (two activity components that were 180 degrees out of phase) under constant light (LL). When rhythm splitting was observed in their behavior, gene expression rhythms of clock genes such as Per1, Per2, and Bmal1 were desynchronized and exhibited anti-phasic rhythms between the right and left SCN [89], even though the phase relation among these clock genes was intact unilaterally. Additionally, each SCN independently regulated their output functions, such as the release of gonadotrophin-releasing hormone [90]. This result demonstrates that under certain circumstances, the circadian clocks of the right and left SCN can also be separated.
Oscillators in the SCN that regulate seasonal changes in behavior
Another set of regional oscillations were found to change their phase relations when animals were exposed to different photoperiods. Nocturnal rodents are known to entrain to a wide range of photoperiods, from an LD cycle of light 0.5 h and dark 23.5 h (LD 0.5:23.5) to an LD 20:4. In order to explain this wide range of adaptability, the two-oscillator model was proposed in 1976 [91]. In this model, the evening (E) oscillator is reset by a light-off signal and regulates the onset of activity, whereas the morning (M) oscillator is reset by a light-on signal and regulates the end of activity. In order to study the localization and the molecular mechanisms of the E and M oscillators, we exposed the Per1-luc mice to either LD18:6 (long day) or LD 6:18 (short day) for 3 weeks. On the last day of the experiment, we collected two consecutive coronal SCN slices and found that the Per1-luc rhythm of the posterior SCN phase-led the anterior counterpart without exception [92]. Notably, the phase difference between the two SCN slices increased with the length of the light phase, and in mice exposed to LD18:6, the Per1-luc signal peaked twice a day in the anterior SCN slice, in the morning and in the evening. Furthermore, we found that the Per1-luc rhythm in the anterior SCN was phase-locked to the onset of behavior activity, while that of the posterior SCN coincided with the end of behavior activity, indicating the anterior and posterior SCN as sites of the E and M oscillators, respectively. We also observed a third oscillator, which exhibited a peak in the early morning in the anterior SCN only under LD18:6; the function of this was unknown.
To identify the precise location of the three oscillators in the SCN, we made horizontal SCN slices that contained both the anterior and posterior SCNs in a single slice. A geometrical transformation technique was applied to set the shape of the cultured SCNs to the same shape as the arbitrary-selected template SCN so that the bioluminescence image data could be statistically evaluated [93]. We found that the oscillator cells located in the posterior tip corresponded to the M oscillator; cells of the open-ring shape covering the anterior SCN corresponded to the E oscillator; and cells in the center appeared to respond to light. The light-responsive area may transfer the lights-on and -off signals to the M and E oscillators, respectively.
We also analyzed the rhythms of PER2::LUC mice to compare the roles of Per1 and Per2 in photoperiodic responses. Intriguingly, the peak phases of Per1-luc rhythms changed almost proportionally to the changes in photoperiods (Fig. 4c–f), whereas those of PER2:LUC exhibited minimal changes in the circadian phase within a slice. The distinct localization of the E and M oscillator cells were not detected in the PER2::LUC mice. Per1 and Per2 were shown to have non-redundant roles in the photoperiodic response.
Clock gene mutation revealed clock mechanisms at cellular and network levels
Gene mutations of clock and clock-related genes can induce rhythm disturbances at multiple levels in the organism. In most mutant mice that exhibited altered circadian periods in behavior, similar deviations were also observed in their dispersed SCN cells [94, 95]. However, behavior rhythms are also affected by the mutation affecting rhythm coupling and output. Despite no known role of VIP in the generation of circadian rhythm in the SCN, mice lacking its receptor, VIP Receptor 2 (VIPR2), often exhibit behavioral arrhythmicity in DD [96]. They also display a variety of abnormalities such as splitting and relative coordination [97]. VIPR2 is widely expressed in neurons of both the SCN core and shell. In cultured SCN slices, individual SCN cells exhibited significant circadian rhythms, but their circadian periods were distributed over a wider range of time compared to that of wild type mice [98]. Collectively, these results demonstrate the importance of coupling in vivo, and that the mechanism that translates circadian rhythm in the SCN to behavior is far more complicated than we had previously expected.
Cry1/Cry2-double-deficient (Cry1−/−/Cry2−/−) mice become behaviorally arrhythmic immediately after they are exposed to DD [99]. Since CRYs are critical molecules for inhibiting CLOCK and BMAL1 heterodimer-mediated activation of Per, the arrhythmic behavior in Cry1−/−/Cry2−/− mice has been regarded as a result of the termination of molecular oscillation. In addition, nocturnal behavior of Cry1−/−/Cry2−/− mice under an LD cycle has been regarded as the masking of behavior by light and darkness [99]. However, we found that the onset phase of nocturnal behavior in Cry1−/−/Cry2−/− mice systematically changed according to the period length of the LD cycle (T cycle) [100], which further indicated that a light-entrainable circadian oscillator was present even in mice lacking CRYs. We predicted that if individual cellular clocks with a wide range of periods were unable to synchronize, but still retained the ability to entrain to an LD cycle, then coherent rhythm outputs observed under LD would be immediately lost due to desynchronization of constitutional cellular clocks. As expected, adult SCN slices of Cry1−/−/Cry2−/− lacked circadian rhythm in a whole SCN tissue; however, sloppy but statistically significant circadian rhythms were observed in individual SCN cells. Surprisingly, Cry1−/−/Cry2−/− SCNs of neonatal mice exhibited robust and synchronous circadian rhythms in their clock gene expression (Fig. 5a). The ability of the cellular oscillators to synchronize decreased over time, and the SCN tissue became totally arrhythmic by postnatal week 3 [101]. Furthermore, when the SCN neurons from postnatal day 6 mice (when synchronized PER2::LUC rhythms were still observed in Cry1−/−/Cry2−/− SCN) were dissociated or when those in a cultured SCN slice were treated with tetrodotoxin (TTX), individual cellular rhythms were desynchronized and period lengths were distributed over an extremely wide range, compared to the narrow distribution of periodicity observed in wild type SCN cells. Desynchronization of cellular rhythms in Cry1−/−/Cry2−/− mice resulted in arrhythmicity of the SCN tissue and behavior. Interestingly, their clock gene expression rhythms in the SCN tissue were rescued upon co-culturing with wild type SCN [101], suggesting that a diffusible factor(s) was involved in the synchronization of cellular clocks. Furthermore, only neonatal wild type SCN were able to rescue the arrhythmic adult Cry1−/−/Cry2−/− SCN, indicating that the release of this diffusible factor was age-dependent. Although the wild type SCN exhibits robust circadian rhythms with synchronized cellular oscillations throughout their life, these results suggest that the mechanism for rhythm synchronization changes from a CRY-independent to a CRY-dependent coupling one during postnatal development.
Fig. 5.

Development of cellular networks in the suprachiasmatic nucleus (SCN). a PER2::LUC rhythms in SCN slices at postnatal day (P) 7, P14, and P21 were monitored using a photomultiplier tube (PMT). The SCNs of Cry1−/−/Cry2−/− mice exhibit a robust circadian rhythm at P7; the rhythm is gradually dampened and completely undetectable by P21. Wild type SCN exhibits robust rhythms in both neonatal and adult cultures. Data from three littermates are shown for each panel. Vertical lines in the graph indicate 0:00 local time. Numbers on the right margin indicate the age of each SCN culture. b PER2::LUC rhythm of an adult Vipr2−/−/Cry1−/−/Cry2−/− SCN is rescued by co-culturing with neonatal SCN of wild type mice carrying no luciferase reporter on the 5th day of culture (indicated by an arrow). The antagonists for V1a and V1b AVP receptors suppressed the rhythm (gray line), indicating that AVP from the wild type SCN is critical for the coupling of cellular rhythm. Black, vehicle treatment (see Ref. [98])
Using triple knockout mice lacking VIPR2, CRY1, and CRY2 (VIPR2−/−/Cry1−/−/Cry2−/−), we demonstrated that the CRY-independent coupling found during the neonatal period was dependent on VIP signaling, and that AVP and VIP had redundant roles in the coupling of cellular rhythms [98]. AVP expression was attenuated in the Cry1−/−/Cry2−/− SCN and VIP signaling naturally lost its ability to regulate rhythm coupling over time, thus resulting in the loss of synchronous cellular rhythms in adult Cry1−/−/Cry2−/− SCN slices. Recovery of AVP signaling from the neonatal wild type SCN rescued circadian rhythms in both the adult VIPR2−/−/Cry1−/−/Cry2−/− (Fig. 5b) and Cry1−/−/Cry2−/− SCNs. It appears that peptidergic signaling is critical for generating coordinated rhythm expression from the SCN, and may be advantageous over the fast classical neurotransmitter signaling, which exerts its effect in the order of milliseconds.
Circadian rhythms in calcium levels and membrane potential
Intracellular Ca2+ mediates input signals to the molecular clock of the cell (e.g., phase-resetting stimuli to the SCN) and output signals from the molecular clock (e.g., neurotransmitter release). Taking advantage of the improvements in long-term and large-scale Ca2+ imaging, we were able to monitor circadian Ca2+ rhythms across all neurons of cultured SCN slices [102, 103]. In every section of the SCN neuron examined so far — including the regions with low clock gene expression — a robust circadian Ca2+ rhythm with a topologically specific phase distribution was observed [104]. Circadian Ca2+ rhythms of the dorsal SCN neurons phase-led those of the ventral neurons, similarly to the phase-distribution pattern observed for Per1 and Per2 [67]. The gap junction blocker carbenoxolone had minimal effects on the Ca2+ rhythm at the single cell as well as at the network levels, while the Na+ channel blocker TTX uncoupled the Ca2+ rhythms between the dorsal and ventral SCNs. Under TTX treatment, Ca2+ rhythms continued although the amplitude was reduced by approximately 30% [104]. These findings suggest that neural firing may be necessary to establish the hierarchy among the multiple regional oscillators in the SCN, even though the synchronization within the regional oscillator are independent of the cellular transmission via Na+ channels or gap junctions. The 30% reduction of amplitude under TTX treatment is thought to reflect the Ca2+ increase due to input signals in the overall Ca2+ rhythms.
The SCN outputs circadian signals to multiple brain areas and peripheral clocks to orchestrate the circadian rhythms throughout the body. In addition to the multielectrode array dish (MED) to record spontaneous firing from multiple neurons of the SCN, we found that the genetically encoded voltage sensor ArchLightD is a powerful tool to monitor circadian rhythm of the membrane potential from entire SCN slices. Unexpectedly, the circadian voltage rhythms were synchronous throughout the coronal SCN slice [105, 106]. Circadian rhythms of spontaneous firing, simultaneously monitored by the MED system, were also synchronous between the dorsal and ventral SCN, despite the topologically specific phase distribution of Ca2+ rhythms within the SCN. The similarity in phase distribution between the Per1-luc/PER2::LUC and the Ca2+ rhythms indicates that the synchronous rhythm of the neural outputs at the network level is independent of the molecular clock of individual neurons. The differences in regulation of the firing and clock gene expression rhythms [86, 87] may allow for the precise phase regulation of rhythm outputs.
Perspectives
In this review, I have highlighted the recent progress made in the understanding of the mammalian circadian clock, with a special focus on cellular oscillation mechanisms and oscillator networks in the SCN. We now know that the circadian system is composed of the central clock in the SCN and peripheral clocks found throughout in the body (Fig. 6). The mechanisms remain to be elucidated as to how the SCN orchestrates these peripheral clocks and how these clocks cross their rhythm signals. In addition, mammals have at least two extra-SCN circadian oscillators in the central nervous system, namely, the food entrainable oscillator [107, 108] and methamphetamine-induced oscillator [109]. These oscillators drive rhythms of circadian periodicity in various physiological functions including sleep-wakefulness, feeding, drinking, body temperature and autonomic nervous functions [110, 111] even in SCN lesioned animals. The hierarchical multi-oscillator system composed of the SCN central clock, extra SCN brain oscillators and peripheral clocks throughout the body highlight the complicated nature of the circadian system.
Fig. 6.
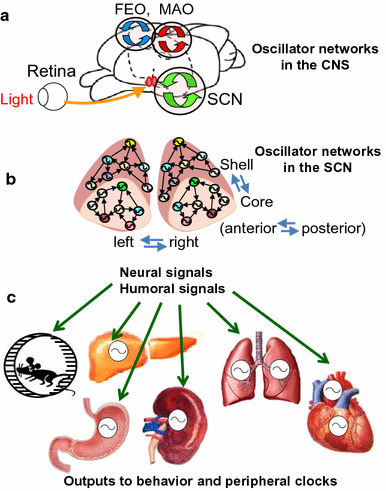
Hierarchical multi-oscillator circadian system. a In addition to the central circadian clock in the SCN, two extra-SCN oscillators, food-entrainable oscillator (FEO) and methamphetamine-induced oscillator (MAO), are known to regulate various physiological functions, including sleep–wake rhythms. These are under the control of the SCN, but can be dissociated when food is restricted at certain hours in daytime (FEO) or methamphetamine is administered (MAO). b The SCN is composed of regional oscillators such as those in the core and shell, right and left, and anterior and posterior, each of which is comprised of multiple cellular oscillators. Regional oscillators are dissociable under specific conditions such as LD shifts, constant light, and photoperiod changes. c Peripheral clocks and behavioral rhythms are thought to be orchestrated by the SCN via neural signals (e.g., sympathetic nerves) and humoral signals (e.g., hormones and cytokines)
The circadian clock regulates neuronal, metabolic, and hormonal functions, and many clock gene mutants exhibit disturbances in sleep–wake rhythms and metabolisms [112, 113]. Further research is needed to reveal how the SCN orchestrates all of the peripheral clocks, how these clocks communicate their circadian rhythm, and how the hierarchical multi-oscillator circadian system is established. Finally, the structure and functions of the extra-SCN oscillators would be fascinating research areas to target.
Compliance with ethical standards
Conflict of interest
The author declares no conflicts of interest.
Research involving human participants and/or animals
All applicable international, national, and institutional guidelines for the care and use of animals were followed. This article did not contain any studies involving human participants.
Informed consent
Not applicable.
References
- 1.Zehring WA, Wheeler DA, Reddy P, Konopka RJ, Kyriacou CP, Rosbash M, Hall JC. P-element transformation with period locus DNA restores rhythmicity to mutant, arrhythmic Drosophila melanogaster. Cell. 1984;39:369–376. doi: 10.1016/0092-8674(84)90015-1. [DOI] [PubMed] [Google Scholar]
- 2.Bargiello TA, Jackson FR, Young MW. Restoration of circadian behavioural rhythms by gene transfer in Drosophila. Nature. 1984;312:752–754. doi: 10.1038/312752a0. [DOI] [PubMed] [Google Scholar]
- 3.Gekakis N, Staknis D, Nguyen HB, Davis FC, Wilsbacher LD, King DP, Takahashi JS, Weitz CJ. Role of the CLOCK protein in the mammalian circadian mechanism. Science. 1998;280:1564–1569. doi: 10.1126/science.280.5369.1564. [DOI] [PubMed] [Google Scholar]
- 4.Aschoff J. Freerunning and entrained circadian rhythms. In: Aschoff J, editor. Handbook of behavioral neurobiology. 4. Biological rhythms. New York: Plenum Press; 1981. [Google Scholar]
- 5.Daan S, Pittendrigh CS. A functional analysis of circadian pacemakers in nocturnal rodents. II. The variability of phase response curves. J Comp Biol. 1976;106:253–266. [Google Scholar]
- 6.Pittendrigh CS, Daan S. A functional analysis of circadian pacemakers in nocturnal rodents. IV. Entrainment: pacemaker as clock. J Comp Biol. 1976;106:291–331. [Google Scholar]
- 7.Johnson CH. An atlas of phase responses curves for circadian and circatidal rhythm. Nashville: Vanderbilt University, Nashville; 1990. [Google Scholar]
- 8.Ma P, Woelfle MA, Johnson CH. An evolutionary fitness enhancement conferred by the circadian system in cyanobacteria. Chaos, Solitons Fractals. 2013;50:65–74. doi: 10.1016/j.chaos.2012.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Narasimamurthy R, Virshup DM. Molecular mechanisms regulating temperature compensation of the circadian clock. Front Neurol. 2017;8:161. doi: 10.3389/fneur.2017.00161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Abe J, Hiyama TB, Mukaiyama A, Son S, Mori T, Saito S, Osako M, Wolanin J, Yamashita E, Kondo T, Akiyama S. Atomic-scale origins of slowness in the cyanobacterial circadian clock. Science. 2015;349:312–316. doi: 10.1126/science.1261040. [DOI] [PubMed] [Google Scholar]
- 11.Shinohara Y, Koyama YM, Ukai-Tadenuma M, Hirokawa T, Kikuchi M, Yamada RG, Ukai H, Fujishima H, Umehara T, Tainaka K, Ueda H. Temperature-sensitive substrate and product binding underlie temperature-compensated phosphorylation in the clock. Mol Cell. 2017;67:783–798. doi: 10.1016/j.molcel.2017.08.009. [DOI] [PubMed] [Google Scholar]
- 12.Weaver DR, Reppert SM. Coordination of circadian timing in mammals. Nature. 2002;418:935–941. doi: 10.1038/nature00965. [DOI] [PubMed] [Google Scholar]
- 13.Mohawk JA, Green CB, Takahashi JS. Central and peripheral circadian clocks in mammals. Ann Rev Neurosci. 2012;35:445–462. doi: 10.1146/annurev-neuro-060909-153128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yamazaki S, Numano R, Abe M, Hida A, Takahashi R, Ueda M, Block GD, Sakaki Y, Menaker M, Tei H. Resetting central and peripheral circadian oscillators in transgenic rats. Science. 2000;288:682–695. doi: 10.1126/science.288.5466.682. [DOI] [PubMed] [Google Scholar]
- 15.Tei H, Okamura H, Shigeyoshi Y, Fukuhara C, Ozawa R, Hirose M, Sakaki Y. Circadian oscillation of a mammalian homologue of the Drosophila period gene. Nature. 1997;389:512–516. doi: 10.1038/39086. [DOI] [PubMed] [Google Scholar]
- 16.Ikeda M, Nomura M. cDNA cloning and tissue-specific expression of a novel basic helix-loop-helix/PAS protein (BMAL1) and identification of alternatively spliced variants with alternative translation initiation site usage. Biochem Biophys Res Commun. 1997;233:258–264. doi: 10.1006/bbrc.1997.6371. [DOI] [PubMed] [Google Scholar]
- 17.Honma S, Ikeda M, Abe H, Tanahashi Y, Namihira M, Honma K, Nomura Circadian oscillation of BMAL1, a partner of a mammalian clock gene Clock, in rat suprachiasmatic nucleus. Biochem Biophys Res Commun. 1998;250:83–87. doi: 10.1006/bbrc.1998.9275. [DOI] [PubMed] [Google Scholar]
- 18.Kume K, Zylka MJ, Sriram S, Shearman LP, Weaver DR, Jin X, Maywood ES, Hastings MH, Reppert SM. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell. 1999;98:193–205. doi: 10.1016/S0092-8674(00)81014-4. [DOI] [PubMed] [Google Scholar]
- 19.Horwitz BA, Gressel J, Malkin S, Epel BL. Modified cryptochrome in vivo absorption in dim photosporulation mutants of Trichoderma. Proc Natl Acad Sci U S A. 1985;82:2736–2740. doi: 10.1073/pnas.82.9.2736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sponga F, Deitzer GF, Mancinelli AL. Cryptochrome, phytochrome, and the photoregulation of anthocyanin production under blue light. Plant Physiol. 1986;82:952–955. doi: 10.1104/pp.82.4.952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Griffin EA, Jr, Staknis D, Weitz CJ. Light-independent role of CRY1 and CRY2 in the mammalian circadian clock. Science. 1999;286:768–771. doi: 10.1126/science.286.5440.768. [DOI] [PubMed] [Google Scholar]
- 22.Alada R, White NE, So WV, Hall JC, Roshach MA. A mutant Drosophila homolog of mammalian clock disrupts circadian rhythms and transcription of period and timeless. Cell. 1998;93:791–804. doi: 10.1016/S0092-8674(00)81440-3. [DOI] [PubMed] [Google Scholar]
- 23.Rutila JE, Suri V, Le M, So WV, Rosbash M, Hall JC. CYCLE is a second bHLH-PAS clock protein essential for circadian rhythmicity and transcription of Drosophila period and timeless. Cell. 1998;93:805–814. doi: 10.1016/S0092-8674(00)81441-5. [DOI] [PubMed] [Google Scholar]
- 24.Takumi T, Taguchi K, Miyake S, Sakakida Y, Takashima N, Matsubara C, Maebayashi Y, Okumura K, Takekida S, Yamamoto S, Yagita K, Yan L, Young MW, Okamura HA. Light-independent oscillatory gene mPer3 in mouse SCN and OVL. EMBO. 1998;17:4753–4759. doi: 10.1093/emboj/17.16.4753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Unsal-Kaçmaz K, Mullen TE, Kaufmann WK, Sancar A. Coupling of human circadian and cell cycles by the timeless protein. Mol Cell Biol. 2005;25:3109–3116. doi: 10.1128/MCB.25.8.3109-3116.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Shearman LP, Sriram S, Weaver DR, Maywood ES, Chaves I, Zheng B, Kume K, Lee CC, van der Horst GT, Hastings MH, Reppert SM. Interacting molecular loops in the mammalian circadian clock. Science. 2000;288:1013–1019. doi: 10.1126/science.288.5468.1013. [DOI] [PubMed] [Google Scholar]
- 27.SatoTK Panda S, Miraglia LJ, Reyes TM, Rudic RD, McNamara P, Naik KA, FitzGerald GA, Kay SA, Hogenesch JB. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron. 2004;43:527–537. doi: 10.1016/j.neuron.2004.07.018. [DOI] [PubMed] [Google Scholar]
- 28.Preitner N, Damiola F, Lopez-Molina L, Zakany J, Duboule D, Albrecht U, Schibler U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell. 2002;110:251–260. doi: 10.1016/S0092-8674(02)00825-5. [DOI] [PubMed] [Google Scholar]
- 29.Honma S, Kawamoto T, Takagi Y, Fujimoto K, Sato F, Noshiro M, Kato Y, Honma K. Dec1 and Dec2 are regulators of the mammalian molecular clock. Nature. 2002;419:841–844. doi: 10.1038/nature01123. [DOI] [PubMed] [Google Scholar]
- 30.Ueda HR, Hayashi S, Chen W, Sano M, Machida M, Shigeyoshi Y, Iino M, Hashimoto S. System-level identification of transcriptional circuits underlying mammalian circadian clocks. Nat Genet. 2005;37:187–192. doi: 10.1038/ng1504. [DOI] [PubMed] [Google Scholar]
- 31.Provencio I, Jiang G, De Grip WJ, Hayes WP, Rollag MD. Melanopsin: an opsin in melanophores, brain, and eye. Proc Natl Acad Sci U S A. 1998;95:340–345. doi: 10.1073/pnas.95.1.340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Freedman MS, Lucas RJ, Soni B, von Schantz M, Munoz M, David-Gray Z, et al. Regulation of mammalian circadian behavior by non-rod, non-cone, ocular photoreceptors. Science. 1999;284:502–504. doi: 10.1126/science.284.5413.502. [DOI] [PubMed] [Google Scholar]
- 33.Berson DM, Dunn FA, Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science. 2002;295:1070–1073. doi: 10.1126/science.1067262. [DOI] [PubMed] [Google Scholar]
- 34.Moore RY, Eichler VB. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 1972;42:201–206. doi: 10.1016/0006-8993(72)90054-6. [DOI] [PubMed] [Google Scholar]
- 35.Schmidt TM, Do MT, Dacey D, Lucas R, Hattar S, Matynia A. Melanopsin-positive intrinsically photosensitive retinal ganglion cells: from form to function. J Neurosci. 2011;31:16094–16101. doi: 10.1523/JNEUROSCI.4132-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lucas RJ, Freedman MS, Munoz M, Garcia-Fernandez JM, Foster RG. Regulation of the mammalian pineal by non-rod, non-cone, ocular photoreceptors. Science. 1999;284:505–507. doi: 10.1126/science.284.5413.505. [DOI] [PubMed] [Google Scholar]
- 37.Lucas RJ, Douglas RH, Foster RG. Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Nat Neurosci. 2001;4:621–626. doi: 10.1038/88443. [DOI] [PubMed] [Google Scholar]
- 38.Barnard AR, Hattar S, Hankins MW, Lucas RJ. Melanopsin regulates visual processing in the mouse retina. Curr Biol. 2006;16:389–395. doi: 10.1016/j.cub.2005.12.045. [DOI] [PubMed] [Google Scholar]
- 39.Altimus CM, Guler AD, Villa KL, McNeill DS, Legates TA, Hattar S. Rods-cones and melanopsin detect light and dark to modulate sleep independent of image formation. Proc Natl Acad Sci USA. 2008;105:19998–20003. doi: 10.1073/pnas.0808312105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shigeyoshi Y, Taguchi K, Yamamoto S, Takekida S, Yan L, Tei H, Moriya T, Shibata S, Loros JJ, Dunlap JC, Okamura H. Light-induced resetting of a mammalian circadian clock is associated with rapid induction of the mPer1 transcript. Cell. 1997;91:1043–1053. doi: 10.1016/S0092-8674(00)80494-8. [DOI] [PubMed] [Google Scholar]
- 41.Shearman LP, Zylka MJ, Weaver DR, Kolakowski LF, Jr, Reppert SM. Two period homologs: circadian expression and photic regulation in the suprachiasmatic nuclei. Neuron. 1997;19:1261–1269. doi: 10.1016/S0896-6273(00)80417-1. [DOI] [PubMed] [Google Scholar]
- 42.Yan L, Takekida S, Shigeyoshi Y, Okamura H. Per1 and Per2 gene expression in the rat suprachiasmatic nucleus: circadian profile and the compartment-specific response to light. Neuroscience. 1999;94:141–150. doi: 10.1016/S0306-4522(99)00223-7. [DOI] [PubMed] [Google Scholar]
- 43.Koike N, Yoo SH, Huang HC, Kumar V, Lee C, Kim TK, Takahashi JS. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science. 2012;338:349–354. doi: 10.1126/science.1226339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Terajima H, Yoshitane H, Ozaki H, Suzuki Y, Shimba S, Kuroda S, Iwasaki W, Fukada Y. ADARB1 catalyzes circadian A-to-I editing and regulates RNA rhythm. Nat Genet. 2017;49:146–151. doi: 10.1038/ng.3731. [DOI] [PubMed] [Google Scholar]
- 45.Ralph MR, Menaker M. A mutation of the circadian system in golden hamsters. Science. 1988;241:1225–1227. doi: 10.1126/science.3413487. [DOI] [PubMed] [Google Scholar]
- 46.Lowrey PL, Shimomura K, Antoch MP, Yamazaki S, Zemenides PD, Ralph MR, Menaker M, Takahashi JS. Positional syntenic cloning and functional characterization of the mammalian circadian mutation tau. Science. 2000;288:483–492. doi: 10.1126/science.288.5465.483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Toh KL, Jones CR, He Y, Eide EJ, Hinz WA, Virshup DM, Ptácek LJ, Fu YH. An hPer2 phosphorylation site mutation in familial advanced sleep phase syndrome. Science. 2001;291:1040–1043. doi: 10.1126/science.1057499. [DOI] [PubMed] [Google Scholar]
- 48.Gallego M, Virshup DM. Post-translational modifications regulate the ticking of the circadian clock. Nat Rev Mol Cell Biol. 2007;8:139–148. doi: 10.1038/nrm2106. [DOI] [PubMed] [Google Scholar]
- 49.Hirota T, Lewis WG, Liu AC, Lee JW, Schultz PG, Kay SA. A chemical biology approach reveals period shortening of the mammalian circadian clock by specific inhibition of GSK-3. Proc Natl Acad Sci USA. 2008;105:20746–20751. doi: 10.1073/pnas.0811410106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Siepka SM, Yoo S-H, Park J, Song W, Kumar V, Hu Y, Lee C, Takahashi JS. Circadian mutant overtime reveals F-box protein FBXL3 regulation of Cryptochrome and Period gene expression. Cell. 2007;129:1011–1023. doi: 10.1016/j.cell.2007.04.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Godinho SI, Maywood ES, Shaw L, Tucci V, Barnard AR, Busino L, Pagano M, Kendall R, Quwailid MM, Romero MR, O’neill J, Chesham JE, Brooker D, Lalanne Z, Hastings MH, Nolan PM. The after-hours mutant reveals a role for Fbxl3 in determining mammalian circadian period. Science. 2007;316:897–900. doi: 10.1126/science.1141138. [DOI] [PubMed] [Google Scholar]
- 52.Belden WJ, Dunlap JC. SIRT1 is a circadian deacetylase for core clock components. Cell. 2008;134:212–214. doi: 10.1016/j.cell.2008.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Cardone L, Hirayama J, Giordano F, Tamaru T, Palvimo JJ, Sassone-Corsi P. Circadian clock control by SUMOylation of BMAL1. Science. 2005;309:1390–1394. doi: 10.1126/science.1110689. [DOI] [PubMed] [Google Scholar]
- 54.Yoo SH, Mohawk JA, Siepka SM, Shan Y, Huh SK, Hong HK, Kornblum I, Kumar V, Koike N, Xu M, Nussbaum J, Liu X, Chen Z, Chen ZJ, Green CB, Takahashi JS. Competing E3 ubiquitin ligases govern circadian periodicity by degradation of CRY in nucleus and cytoplasm. Cell. 2013;152:1091–1105. doi: 10.1016/j.cell.2013.01.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hirano A, Yumimoto K, Tsunematsu R, Matsumoto M, Oyama M, Kozuka-Hata H, Nakagawa T, Lanjakornsiripan D, Nakayama KI, Fukada Y. FBXL21 regulates oscillation of the circadian clock through ubiquitination and stabilization of cryptochromes. Cell. 2013;152:1106–1118. doi: 10.1016/j.cell.2013.01.054. [DOI] [PubMed] [Google Scholar]
- 56.Xu Y, Padiath QS, Shapiro RE, Jones CR, Wu SC, Saigoh N, Saigoh K, Ptácek LJ. Fu YH (2005) Functional consequences of a CKI delta mutation causing familial advanced sleep phase syndrome. Nature. 2009;434:640–644. doi: 10.1038/nature03453. [DOI] [PubMed] [Google Scholar]
- 57.He Y, Jones CR, Fujiki N, Xu Y, Guo B, Holder JL, Jr, Rossner MJ, Nishino S, Fu YH. The transcriptional repressor DEC2 regulates sleep length in mammals. Science. 2009;325:866–870. doi: 10.1126/science.1174443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ebisawa T, Uchiyama M, Kajimura N, Mishima K, Kamei Y, Katoh M, Watanabe T, Sekimoto M, Shibui K, Kim K, Kudo Y, Ozeki Y, Sugishita M, Toyoshima R, Inoue Y, Yamada N, Nagase T, Ozaki N, Ohara O, Ishida N, Okawa M, Takahashi K, Yamauchi T. Association of structural polymorphisms in the human period3 gene with delayed sleep phase syndrome. EMBO Rep. 2001;2:342–346. doi: 10.1093/embo-reports/kve070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Partonen T. Clock gene variants in mood and anxiety disorders. J Neural Transm. 2012;119:1133–1145. doi: 10.1007/s00702-012-0810-2. [DOI] [PubMed] [Google Scholar]
- 60.Albrecht U. Molecular mechanisms in mood regulation involving the circadian clock. Front Neurol. 2017;8:30. doi: 10.3389/fneur.2017.00030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.De Leersnyder H, Claustrat B, Munnich A, Verloes A. Circadian rhythm disorder in a rare disease: smith-Magenis syndrome. Mol Cell Endocrinol. 2006;252:88–91. doi: 10.1016/j.mce.2006.03.043. [DOI] [PubMed] [Google Scholar]
- 62.Shi SQ, Bichell TJ, Ihrie RA, Johnson CH. Ube3a imprinting impairs circadian robustness in Angelman syndrome models. Curr Biol. 2015;25:537–545. doi: 10.1016/j.cub.2014.12.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Tsuchiya Y, Minami Y, Umemura Y, Watanabe H, Ono D, Nakamura W, Takahashi T, Honma S, Kondoh G, Matsuishi T, Yagita K. Disruption of MeCP2 attenuates circadian rhythm in CRISPR/Cas9-based Rett syndrome model mouse. Genes Cells. 2015;20:992–1005. doi: 10.1111/gtc.12305. [DOI] [PubMed] [Google Scholar]
- 64.Field MD, Maywood ES, O’Brien JA, Weaver DR, Reppert SM, Hastings MH. Analysis of clock proteins in mouse SCN demonstrates phylogenetic divergence of the circadian clockwork and resetting mechanisms. Neuron. 2000;25:437–447. doi: 10.1016/S0896-6273(00)80906-X. [DOI] [PubMed] [Google Scholar]
- 65.Yoo SH, Yamazaki S, Lowrey PL, Shimomura K, Ko CH, Buhr ED, Siepka SM, Hong HK, Oh WJ, Yoo OJ, Menaker M, Takahashi JS. PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc Natl Acad Sci USA. 2004;101:5339–5346. doi: 10.1073/pnas.0308709101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Nishide S, Honma S, Nakajima Y, Ikeda M, Baba K, Ohmiya Y, Honma K. New reporter system for Per1 and Bmal1 expressions revealed self-sustained circadian rhythms in peripheral tissues. Genes Cells. 2006;11:1173–1182. doi: 10.1111/j.1365-2443.2006.01015.x. [DOI] [PubMed] [Google Scholar]
- 67.Yamaguchi S, Isejima H, Matsuo T, Okura R, Yagita K, Kobayashi M, Okamura H. Synchronization of cellular clocks in the suprachiasmatic nucleus. Science. 2003;302:1408–1412. doi: 10.1126/science.1089287. [DOI] [PubMed] [Google Scholar]
- 68.Honma S. Unveiling functions of the central circadian clock by imaging “clock time”. J Physiol Sci. 2016;66:S13. [Google Scholar]
- 69.Stephan FK, Zucker I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc Natl Acad Sci U S A. 1972;69:1583–1586. doi: 10.1073/pnas.69.6.1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nagoshi E, Saini C, Bauer C, Laroche T, Naef F, Schibler U. Circadian gene expression in individual fibroblasts: cell-autonomous and self-sustained oscillators pass time to daughter cells. Cell. 2004;119:693–705. doi: 10.1016/j.cell.2004.11.015. [DOI] [PubMed] [Google Scholar]
- 71.Yamaguchi S, Kobayashi M, Mitsui S, Ishida Y, van der Horst GT, Suzuki M, Shibata S, Okamura H. View of a mouse clock gene ticking. Nature. 2001;409:684. doi: 10.1038/35055628. [DOI] [PubMed] [Google Scholar]
- 72.Ono D, Honma K, Honma S. Circadian and ultradian rhythms of clock gene expression in the suprachiasmatic nucleus of freely moving mice. Sci Rep. 2015;21:12310. doi: 10.1038/srep12310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hamada T, Sutherland K, Ishikawa M, Miyamoto N, Honma S, Shirato H, Honma K. In vivo imaging of clock gene expression in multiple tissues of freely moving mice. Nat Commun. 2016;7:11705. doi: 10.1038/ncomms11705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Van del Pol AN. The hypothalamic suprachiasmatic nucleus of the rat: intrisic anatomy. J Comp Neurol. 1980;191:661–702. doi: 10.1002/cne.901910410. [DOI] [PubMed] [Google Scholar]
- 75.Card JP, Brecha N, Karten HJ, Moore RY. Immunocytochemical localization of vasoactive intestinal polypeptide-containing cells and processes in the suprachiasmatic nucleus of the rat: light and electron microscopic analysis. J Neurosci. 1981;1:1289–1303. doi: 10.1523/JNEUROSCI.01-11-01289.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Moore RY. Entrainment pathways and the functional organization of the circadian timing system. In: Buijs RM, Kalsbeek A, Romijn HJ, Pennartz CMA, Mirmiran M, editors. Hypothalamic integration of circadian rhythms. Amsterdam: Elsevier; 1996. pp. 101–117. [Google Scholar]
- 77.Mori K, Miyazato M, Ida T, Murakami N, Serino R, Ueta Y, Kojima M, Kangawa K. Identification of neuromedin S and its possible role in the mammalian circadian oscillator system. EMBO J. 2005;24:325–335. doi: 10.1038/sj.emboj.7600526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lee IT, Chang AS, Manandhar M, Shan Y, Fan J, Izumo M, Ikeda Y, Motoike T, Dixon S, Seinfeld JE, Takahashi JS, Yanagisawa M. Neuromedin s-producing neurons act as essential pacemakers in the suprachiasmatic nucleus to couple clock neurons and dictate circadian rhythms. Neuron. 2015;85:1086–1102. doi: 10.1016/j.neuron.2015.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Moore RY, Speh JC. GABA is the principal neurotransmitter of the circadian system. Neurosci Lett. 1993;150:112–116. doi: 10.1016/0304-3940(93)90120-A. [DOI] [PubMed] [Google Scholar]
- 80.Abrahamson EE, Moore RY. Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res. 2001;916:172–191. doi: 10.1016/S0006-8993(01)02890-6. [DOI] [PubMed] [Google Scholar]
- 81.Welsh DK, Logothetis DE, Meister M, Reppert SM. Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms. Neuron. 1995;14:697–706. doi: 10.1016/0896-6273(95)90214-7. [DOI] [PubMed] [Google Scholar]
- 82.Honma S, Shirakawa T, Katsuno Y, Namihira M, Honma K. Circadian periods of single suprachiasmatic neurons in rats. Neurosci Lett. 1998;250:157–160. doi: 10.1016/S0304-3940(98)00464-9. [DOI] [PubMed] [Google Scholar]
- 83.Herzog ED, Kiss IZ, Mazusk C. Measuring synchrony in the mammalian central circadian circuit. Methods Enzymol. 2015;552:3–22. doi: 10.1016/bs.mie.2014.10.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Honma S, Nakamura W, Shirakawa T, Honma K. Diversity in the circadian periods of single neurons of the rat suprachiasmatic nucleus depends on nuclear structure and intrinsic period. Neurosci Lett. 2004;358:173–176. doi: 10.1016/j.neulet.2004.01.022. [DOI] [PubMed] [Google Scholar]
- 85.Honma S, Ono D, Suzuki Y, Inagaki N, Yoshikawa T, Nakamura W, Honma K. Suprachiasmatic nucleus: cellular clocks and networks. Prog Brain Res. 2012;199:129–141. doi: 10.1016/B978-0-444-59427-3.00029-0. [DOI] [PubMed] [Google Scholar]
- 86.Nagano M, Adachi A, Nakahama K, Nakamura T, Tamada M, Meyer-Bernstein E, Sehgal A, Shigeyoshi Y. An abrupt shift in the day/night cycle causes desynchrony in the mammalian circadian center. J Neurosci. 2003;23:6141–6651. doi: 10.1523/JNEUROSCI.23-14-06141.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Nakamura W, Yamazaki S, Takasu NN, Mishima K, Block GD. Differential response of Period 1 expression within the suprachiasmatic nucleus. J Neurosci. 2005;25:5481–5487. doi: 10.1523/JNEUROSCI.0889-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Shinohara K, Honma S, Katsuno Y, Abe H, Honma K. Two distinct oscillators in the rat suprachiasmatic nucleus in vitro. Porc Natl Acad Sci USA. 1995;92:7396–7400. doi: 10.1073/pnas.92.16.7396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.de la Iglesia HO, Meyer J, Carpino A, Jr, Schwartz WJ. Antiphase oscillation of the left and right suprachiasmatic nuclei. Science. 2000;290:799–801. doi: 10.1126/science.290.5492.799. [DOI] [PubMed] [Google Scholar]
- 90.de la Iglesia HO, Meyer J, Schwartz WJ. Lateralization of circadian pacemaker output: activation of left- and right-sided luteinizing hormone-releasing hormone neurons involves a neural rather than a humoral pathway. J Neurosci. 2003;23(19):7412–7414. doi: 10.1523/JNEUROSCI.23-19-07412.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Pittendrigh CS, Daan S. A functional analysis of circadian pacemakers in nocturnal rodents. V. Pacemaker structure: a clock for all seasons. J Comp Biol. 1976;106:333–355. [Google Scholar]
- 92.Inagaki N, Honma S, Ono D, Tanahashi Y, Honma K. Separate oscillating cell groups in mouse suprachiasmatic nucleus couple photoperiodically to the onset and end of daily activity. Proc Natl Acad Sci USA. 2007;104:7664–7669. doi: 10.1073/pnas.0607713104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Yoshikawa T, Inagaki N, Takagi S, Kuroda S, Yamasaki M, Watanabe M, Honma S, Honma K. Localization of photoperiod responsive circadian oscillators in the mouse suprachiasmatic nucleus. Sci Rep. 2017;7:820. doi: 10.1038/s41598-017-00991-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Nakamura W, Honma S, Shirakawa T, Honma K. Clock mutation lengthens the circadian period without damping rhythms in individual SCN neurons. Nat Neurosci. 2002;5:399–400. doi: 10.1038/nn843. [DOI] [PubMed] [Google Scholar]
- 95.Liu AC, Welsh DK, Ko CH, Tran HG, Zhang EE, Priest AA, Buhr ED, Singer O, Meeker K, Verma IM, Doyle FJ, 3rd, Takahashi JS, Kay SA. Intercellular coupling confers robustness against mutations in the SCN circadian clock network. Cell. 2007;129:605–616. doi: 10.1016/j.cell.2007.02.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Harmar AJ, Marston HM, Shen S, Spratt C, West KM, Sheward WJ, Morrison CF, Dorin JR, Piggins HD, Reubi JC, Kelly JS, Maywood ES, Hastings MH. The VPAC2 receptor is essential for circadian function in the mouse suprachiasmatic nuclei. Cell. 2002;109:497–508. doi: 10.1016/S0092-8674(02)00736-5. [DOI] [PubMed] [Google Scholar]
- 97.Aton SJ, Colwell CS, Harmar AJ, Waschek J, Herzog ED. Vasoactive intestinal polypeptide mediates circadian rhythmicity and synchrony in mammalian clock neurons. Nat Neurosci. 2005;8:476–483. doi: 10.1038/nn1419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ono D, Honma S, Honma K. Differential roles of AVP and VIP signaling in the postnatal changes of neural networks for coherent circadian rhythms in the SCN. Sci Adv. 2016;2:e1600960. doi: 10.1126/sciadv.1600960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.van der Horst GT, Muijtjens M, Kobayashi K, Takano R, Kanno S, Takao M, de Wit J, Verkerk A, Eker AP, van Leenen D, Buijs R, Bootsma D, Hoeijmakers JH, Yasui A. Mammalian Cry1 and Cry2 are essential for maintenance of circadian rhythms. Nature. 1999;398:627–630. doi: 10.1038/19323. [DOI] [PubMed] [Google Scholar]
- 100.Honma S, Ono D, Honma K. Cellular oscillators in the suprachiasmatic nucleus for behavior rhythm expression in the mouse lacking CRYPTOCHROME. In: Honma K, editor. Dynamics of circadian oscillation in the SCN. Sapporo: Hokkaido University Press; 2014. pp. 85–95. [Google Scholar]
- 101.Ono D, Honma S, Honma K. Cryptochromes are critical for the development of coherent circadian rhythms in the mouse suprachiasmatic nucleus. Nat Comm. 2013;4:1666. doi: 10.1038/ncomms2670. [DOI] [PubMed] [Google Scholar]
- 102.Enoki R, Ono D, Hasan MT, Honma S. Honma K (2012) Single-cell resolution fluorescence imaging of circadian rhythms detected with a Nipkow spinning disk confocal system. J Neurosci Methods. 2012;202:72–79. doi: 10.1016/j.jneumeth.2012.03.004. [DOI] [PubMed] [Google Scholar]
- 103.Oda Y, Enoki R, Honma K, Honma S. Simultaneous imaging of circadian Ca2 + rhythms and fast Ca2+ activities in the suprachiasmatic nucleus. J Physiol Sci. 2017;67:S165. doi: 10.1007/s12576-016-0480-1. [DOI] [Google Scholar]
- 104.Enoki R, Kuroda S, Ono C, Hasan MT, Ueda T, Honma S, Honma K. Topological specificity and hierarchical network of the circadian calcium rhythm in the suprachiasmatic nucleus. Proc Natl Acad Sci USA. 2012;109:21498–21503. doi: 10.1073/pnas.1214415110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Enoki R, Oda Y, Mieda M, Ono D, Honma S, Honma K. Optical detection of circadian voltage rhythms in the suprachiasmatic nucleus. J Physiol Sci. 2017;67:S73. doi: 10.1073/pnas.1616815114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Enoki R, Oda Y, Mieda M, Ono D, Honma S, Honma K. Synchronous circadian voltage rhythms with asynchronous calcium rhythms in the suprachiasmatic nucleus. Proc Natl Acad Sci USA. 2017;114:E2476–E2485. doi: 10.1073/pnas.1616815114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Mistlberger RE. Circadian food-anticipatory activity: formal models and physiological mechanisms. Neurosci Biobehav Rev. 1994;18:171–195. doi: 10.1016/0149-7634(94)90023-X. [DOI] [PubMed] [Google Scholar]
- 108.Stephan FK. The “other” circadian system: food as a Zeitgeber. J Biol Rhythms. 2003;17:284–289. doi: 10.1177/074873002129002591. [DOI] [PubMed] [Google Scholar]
- 109.Natsubori A, Honma K, Honma S. Dual regulation of clock gene Per2 expression in discrete brain areas by the circadian pacemaker and methamphetamine-induced oscillator in rats. Eur J Neurosci. 2014;39:229–240. doi: 10.1111/ejn.12400. [DOI] [PubMed] [Google Scholar]
- 110.Honma K, Honma S. The SCN-independent clocks, methamphetamine and food restriction. Eur J Neurosci. 2009;30:1707–1717. doi: 10.1111/j.1460-9568.2009.06976.x. [DOI] [PubMed] [Google Scholar]
- 111.Pezuk P, Mohawk JA, Yoshikawa T, Sellix MT, Menaker M. Circadian organization is governed by extra-SCN pacemakers. J Biol Rhythms. 2010;25:432–441. doi: 10.1177/0748730410385204. [DOI] [PubMed] [Google Scholar]
- 112.Sassone-Corsi P, Christen Y (2016) A Time for metabolism and hormones. Springer Cham Heidelberg New York Dordrecht London, eBook. https://www.ncbi.nlm.nih.gov/books/NBK453176/pdf/Bookshelf_NBK453176.pdf [PubMed]
- 113.Takahashi JS. Transcriptional architecture of the mammalian circadian clock. Nat Genet Review. 2017;18:164–179. doi: 10.1038/nrg.2016.150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Honma A, Yamada Y, Nakamaru Y, Fukuda S. Glucocorticoids reset the nasal circadian clock in mice. Endocrinology. 2015;156:4302–4311. doi: 10.1210/en.2015-1490. [DOI] [PubMed] [Google Scholar]
- 115.Nishide SY, Honma S, Nakajima Y, Ikeda M, Baba K, Ohmiya Y, Honma K. New reporter system for Per1 and Bmal1 expressions revealed self-sustained circadian rhythms in peripheral tissues. Gene Cells. 2006;11:1173–1182. doi: 10.1111/j.1365-2443.2006.01015.x. [DOI] [PubMed] [Google Scholar]
- 116.Yoshikawa T, Honma S, K-i Honma. Photoperiodic response of multiple circadian oscillators in mouse SCN. J Physiol Sci. 2012;62(S1):215. [Google Scholar]