

Abstract
Immobilization induces skeletal muscle fibrosis characterized by increasing collagen synthesis in the perimysium and endomysium. Transforming growth factor-β1 (TGF-β1) is associated with this lesion via promoting differentiation of fibroblasts into myofibroblasts. In addition, reactive oxygen species (ROS) are shown to mediate TGF-β1-induced fibrosis in tissues. These reports suggest the importance of ROS reduction for attenuating skeletal muscle fibrosis. Astaxanthin, a powerful antioxidant, has been shown to reduce ROS production in disused muscle. Therefore, we investigated the effects of astaxanthin supplementation on muscle fibrosis under immobilization. In the present study, immobilization increased the collagen fiber area, the expression levels of TGF-β1, α-smooth muscle actin, and superoxide dismutase-1 protein and ROS production. However, these changes induced by immobilization were attenuated by astaxanthin supplementation. These results indicate the effectiveness of astaxanthin supplementation on skeletal muscle fibrosis induced by ankle joint immobilization.
Keywords: Skeletal muscle fibrosis, Transforming growth factor-β1, Reactive oxygen species, Astaxanthin, Immobilization
Introduction
Joint immobilization induces muscle contracture, limiting daily activities, and needs rehabilitation during and/or after immobilization. Although the prevention of this condition is very important, it is difficult to continue physical therapies (e.g., passive stretching, joint mobilization) during immobilization such as casting. In previous studies [1, 2], muscle fibrosis in the extracellular matrix was strongly associated with reduced muscle extensibility and decreased range of motion during immobilization. Moreover, overexpressed collagen in extracellular matrix could not be reduced after remobilization for the same period of immobilization [3]. Recently, Honda et al. [4] implied that the mechanism underlying immobilization-induced muscle fibrosis was strongly associated with transforming growth factor-β1 (TGF-β1), the most potent and ubiquitous profibrogenic cytokine [5], by promoting activation and differentiation of fibroblasts into myofibroblasts, which have a higher activity for collagen production than fibroblasts [6], in the same way as the mechanism in the other fibrotic organs [4]. Furthermore, reactive oxygen species (ROS) mediates the TGF-β1-related development of fibrosis in lung [5, 7, 8].
ROS has been recognized as signaling molecules to regulate multiple redox-sensitive signaling pathways that play a critical role in cellular processes including gene expression and protein modification [9]. The overproduction of ROS from mitochondria has been increased by hindlimb immobilization [10–12]. In addition, ROS has been shown to mediate TGF-β1-induced fibroblast activation and myofibroblast differentiation [13]. Therefore, the prevention of ROS overproduction would be important for attenuating skeletal muscle fibrosis induced by immobilization.
Astaxanthin, a pinkish-orange pigment naturally found in many marine organisms such as crustaceans and fishes (especially salmon), effectively scavenges ROS and reduces oxidative stress, and has been widely used in many experiments as a powerful antioxidant [14, 15]. Several previous studies have shown that astaxanthin possesses higher antioxidant activity than the other antioxidants such as carotenoids and coenzyme Q10 under a variety of stress conditions [16–19]. In addition, astaxanthin does not become pro-oxidant in the body so it cannot cause harmful oxidation [20, 21]. Therefore, there would be a possibility that astaxanthin supplementation has an effective impact on skeletal muscle fibrosis by suppressing oxidative stress induced by immobilization. However, little information is available about the efficacy of astaxanthin on muscle fibrosis along with immobilization. If the efficacy is verified, it could establish an effective preventive intervention for muscle contracture during immobilization, and contribute to rehabilitation. Thus, the aim of the present study was to investigate the effects of astaxanthin supplementation on skeletal muscle fibrosis induced by TGF-β1, via modulation of ROS production induced by ankle joint immobilization.
Materials and methods
Animals
Twenty-eight male Wistar rats (age, 7 weeks; body weight, 159.3 ± 1.0 g; Japan SLC, Shizuoka, Japan) were used. Rats were randomly divided into two groups and provided with oil either with or without astaxanthin (Fuji Chemical Industry, Toyama, Japan) (n = 14/group). Animals were further divided into the following four groups: control (CON, n = 7), control with astaxanthin supplementation (CON + Ax, n = 7), joint immobilization (IM, n = 7), and joint immobilization with astaxanthin supplementation (IM + Ax, n = 7). Rats in the IM and IM + Ax group were immobilized for 14 days. All animals were housed at a temperature of 22 ± 2 °C with a 12:12 h light:dark cycle and provided with standard rodent chow and water ad libitum. This study was approved by the Institution Animal Care and Use Committee and followed the Kobe University Animal Experimentation Regulations (Kobe, Japan). All experimental and animal care procedures were conducted in accordance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication no. 85-23, revised 1996).
Ankle joint immobilization
Rats in the IM and IM + Ax groups were subject to ankle joint immobilization for 14 days using the modified method previously described [4]. Briefly, these animals were anesthetized with pentobarbital sodium (40 mg/kg, i.p.), and both ankle joints were fixed in maximum plantar flexion position with plaster casts (Pliton-100; Alcare, Tokyo, Japan) to immobilize the soleus muscle in a shortened position. The plaster cast, which fits the frontal side of the leg directly below the ankle joint to the toe, was rolled once every 4 days because of loosening consequent to muscle atrophy. During this period, the rats were able to freely eat food and drink water. After 14 days, the animals in IM and IM + Ax groups were removed from the immobilization and immediately sacrificed.
Astaxanthin supplementation
Astaxanthin supplementation was performed by the method previously described [22, 23]. The rats in the CON + Ax and IM + Ax groups were administrated with astaxanthin oil (Fuji Chemical Industry) orally at 100 mg/kg/day by a feeding tube for 3 weeks (for 1 week before ankle joint immobilization and for 2 weeks during joint immobilization). In a previous study, Aoi et al. [24] reported that astaxanthin shifted and was accumulated in skeletal muscle after oral ingestion, even though most dietary carotenoids accumulate mainly in the liver and show relatively little distribution to skeletal muscle. Therefore, we started supplementation before 1 week of immobilization in this study. The rats in the CON and IM groups were administered medium chain triglyceride (MCT), which is the medium of astaxanthin oil, as the placebo at the same frequency and dose of the astaxanthin groups during experimental period.
Tissue extraction
At the end of the experimental period, animals were anaesthetized with sodium pentobarbital (50 mg/kg, i.p.) and injected heparin (5000 IU/kg, i.p.). Animals were killed and the soleus muscles were rapidly removed and weighed. The muscle tissues were mounted on cork, rapidly frozen in an isopentane cooled in dry ice and stored at −80 °C in a freezer until histological and biochemical analyses.
Histological analyses
In the present study, we histologically analyzed both right and left soleus muscles of each rat, and then averaged these data as an individual. Serial transverse 12-μm sections were cut using a cryostat from the middle part of the muscle belly for histological analysis. The sections were subjected to hematoxylin and eosin (H&E) staining to measure the fiber cross-sectional area (FCSA) of the soleus muscle. The other sections were subjected to Elastica van Gieson (EVG) staining to observe the collagen fiber in the extracellular matrix of the soleus muscle as previously described [25]. In this staining, portions of the collagen fibers area were stained red. The sections stained with H&E and EVG were observed in an optical microscope (Cx41; Olympus, Tokyo, Japan) and then chromatic figures were taken by a CCD camera (Moticam 1000; Shimadzu Rika, Tokyo, Japan), which was connected to the microscope. To examine both the deep (near the bone) and superficial regions of each muscle, we acquired more than 6 microscopic images from each transverse tissue section in the present study. FCSA and collagen fiber area as a percent of total area of the transverse tissue section was measured using Image J software (NIH, Bethesda, MD, USA).
Western blotting
The expression levels of TGF-β1, α-smooth muscle actin (SMA), a myofibroblast marker, and superoxide dismutase (SOD)-1, one of antioxidant enzymes, were quantified by western blotting. A portion (~20 mg) of the left soleus muscle was homogenized in ice-cold homogenizing buffer (Ez RIPA Lysis buffer; ATTO, Tokyo, Japan) and supplemented with Sigma protease inhibitor mixture (P8340; Sigma-Aldrich, St. Louis, MO, USA). The homogenates were centrifuged at 15,000 rpm for 15 min at 4 °C, and the resulting supernatants were collected. Total protein content of the samples was measured using the Bradford method [26]. The supernatants were solubilized in sodium dodecyl sulfate (SDS) sample buffer (62.5 mM Tris–HCl, 2 % SDS, 10 % glycerol, 5 % 2-mercaptoethanol and 0.02 % bromophenol blue) and boiled for 15 min at 70 °C. Sample proteins were electrophoresed on 10–15 % SDS-polyacrylamide gels and transferred to a polyvinylidene difluoride (PVDF) membrane. After blocking the membranes in 5 % skimmed milk in phosphate-buffered saline with Tween 20 (PBST), the membranes were incubated using a solution with antibodies against TGF-β1 (1:50 in PBST, sc-13067; Santa Cruz Biotechnology, Santa Cruz, CA, USA), α-SMA (1:500 in PBST, M0851; Dako Denmark, Denmark) or SOD-1 (1:200 in PBST, sc-8637; Santa Cruz Biotechnology), overnight at 4 °C and then incubated in a solution with anti-rabbit, mouse or goat secondary antibody (1:1000 in PBST) for 1 h, respectively. The proteins were detected using the ECL Westlumi One Western Blotting Detection System (Amersham Life Science, Arlington Heights, IL, USA). Images were analyzed with the LAS-1000 (Fujifilm, Tokyo, Japan) using a chemiluminescent image analyzer. The digitized signals were quantified using the Multi-Gauge Image Analysis Software program (Fujifilm) against the relative concentration of GAPDH as an internal control.
Detection of ROS production
The measurement of ROS production was evaluated using oxidative fluorescent dihydroethidium (DHE) as previously described [22, 23]. DHE is a cell-permeable agent and it interacts with nucleic acids. DHE produces red fluorescence when oxidized to ethidium bromide by ROS, including superoxide anion [27]. We detected the red light by fluorescence microscopy (BX51; Olympus) with a rhodamine filter (excitation 490 nm, emission 590 nm). Briefly, the sections were incubated with 0.1 % DHE (Wako Pure Chemicals, Osaka, Japan) solution for 30 min at 37 °C in a dark box, rinsed with PBS, and viewed using a fluorescence microscope (filter with excitation at 545 nm). Densitometric analysis of DHE fluorescence was performed using the ImageJ software program (NIH) in three images per muscle and reported as a percentage of the control group. The individuals performing the densitometric analyses of fluorescence were blinded to the experimental groups.
Statistical analysis
All data are presented as mean ± SEM. Significant differences were determined by two-way analysis of variance (ANOVA). Bonferroni post hoc analyses were subsequently performed between significant means when the ANOVA demonstrated a significant difference. P < 0.05 was considered to be statistically significant.
Results
Body and muscle weight
Mean body weight, soleus muscle wet weight, and muscle to body weight ratio were shown in Table 1. The main effect of immobilization was significant (P < 0.01); however, the main effect of supplementation (P = 0.95, 0.52, 0.40) and interaction (P = 0.73, 0.89, 0.70) were not significant in the two-way ANOVA. When compared with the control groups providing the same supplementation by post hoc test, the body weight, soleus muscle wet weight, and muscle to body weight ratio significantly decreased in IM and IM + Ax groups.
Table 1.
Body mass, soleus muscle wet weight, muscle to body weight ratio and fiber cross-sectional area (FCSA)
| CON | IM | ANOVA P values | |||||
|---|---|---|---|---|---|---|---|
| Placebo | Astaxanthin | Placebo | Astaxanthin | Food | Immobilization | F × I | |
| Body weight (g) | 252 ± 3 | 251 ± 3 | 207 ± 3a | 208 ± 5a | P = 0.95 | P < 0.01 | P = 0.73 |
| Muscle wet weight (mg) | 94 ± 2 | 97 ± 2 | 38 ± 1a | 39 ± 1a | P = 0.52 | P < 0.01 | P = 0.89 |
| Muscle to body weight ratio (mg/100 g) | 38 ± 1 | 38 ± 1 | 19 ± 1a | 19 ± 1a | P = 0.40 | P < 0.01 | P = 0.70 |
| FCSA (µm2) | 2014 ± 47 | 2002 ± 46 | 872 ± 54a | 994 ± 45a | P = 0.26 | P < 0.01 | P = 0.18 |
Data are expressed as the mean (±SEM)
aVersus CON with same supplementation (P < 0.05)
Muscle fiber cross-sectional area
Muscle fiber cross-sectional area (FCSA) was shown in Table 1. The main effect of immobilization was significant (P < 0.01); however, the main effect of supplementation (P = 0.26) and interaction (P = 0.18) were not significant in the two-way ANOVA. When compared with the control groups providing the same supplementation by post hoc test, FCSA significantly decreased in the IM and IM + Ax groups.
Change of collagen fiber area
Collagen fiber deposition in skeletal muscle was measured by EVG stain (Fig. 1a). The areas of collagen fibers as percent of the total field are shown in Fig. 1b. The main effect of immobilization, supplementation and interaction were significant in the two-way ANOVA (P < 0.01, <0.05, and <0.05, respectively). Moreover, the collagen fiber deposition significantly increased in both groups of immobilized rats compared with the control groups provided the same supplementation, whereas the collagen fiber area in the IM + Ax group was significantly lower than that in the IM group.
Fig. 1.
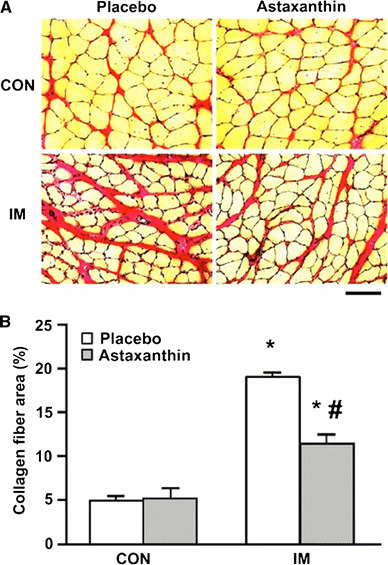
Collagen fiber area. Cross-sectional images of Elastica van Gieson staining of the soleus muscle (a). Scale bar 100 μm. Also shown is the collagen fiber area as a percent of the total field area (b). * vs CON with same supplementation, # Placebo vs astaxanthin (P < 0.05)
Profibrotic factor expression level
TGF-β1 and α-SMA expression levels were determined by western blotting (Fig. 2). In the TGF-β1 expression levels, the main effect of supplementation was not significant; however, the main effects of immobilization and interaction were significant in the two-way ANOVA (P = 0.26, <0.01, <0.05). However, also in the α-SMA expression levels, the main effect of supplementation was not significant, although the main effects of immobilization and interaction were significant in the two-way ANOVA (P = 0.08, <0.01, <0.05). Therefore, in the immobilized muscle, these fibrotic factors were especially attenuated by astaxanthin supplementation. As a result of the post hoc test, the expression levels of TGF-β1 and α-SMA significantly increased in both groups of immobilized rats compared with the control groups providing the same supplementation, whereas TGF-β1 and α-SMA expression levels in the IM + Ax group were significantly lower than those in the IM group, respectively.
Fig. 2.

Protein levels of TGF-β1 and α-SMA. Protein levels of TGF-β1 (a) and α-SMA (b) in all experimental groups. Each protein was quantified in relation to the levels of GAPDH proteins from each gel. The expression levels are calculated as the fold change (in a.u.) and indicated as mean ± SEM. *Versus CON with same supplementation, #placebo vs. astaxanthin (P < 0.05)
Production of ROS
ROS production in skeletal muscle was measured by DHE stain (Fig. 3a) as DHE fluorescence intensity in the nucleus (Fig. 3b). The main effect of immobilization, supplementation and interaction were all significant in the two-way ANOVA (P < 0.05, <0.01, <0.05). Therefore, DHE fluorescence intensity was increased by immobilization and decreased by astaxanthin supplementation. Moreover, especially in the immobilized muscle, the increase of DHE fluorescent intensity was prevented by astaxanthin supplementation. In addition, as a result of post hoc testing, the DHE fluorescence intensity in the IM group was higher than that in the CON group. Meanwhile, the DHE fluorescence intensities in the CON + Ax group and the IM + Ax group were equivalent, and that in the IM + Ax group was significantly lower than that in the IM group.
Fig. 3.

Factors of oxidative stress. Cross-sectional images of dihydroethidium (DHE) staining of the soleus muscle (a). Scale bar 100 μm. The DHE fluorescence intensity (b), and protein levels of SOD-1 (c) are shown in all experimental groups. SOD-1 protein was quantified in relation to the levels of GAPDH proteins from each gel. The expression levels are calculated as the fold change (in a.u.) and indicated as mean ± SEM. *Versus CON with same supplementation, #placebo vs. astaxanthin (P < 0.05)
Anti-oxidative enzyme expression level
SOD-1 expression level was determined by western blotting (Fig. 3c). In the SOD-1 expression levels, the main effects of immobilization, supplementation and interaction were all significant in the two-way ANOVA (P < 0.01, <0.01, <0.05). Therefore, SOD-1 expression level was increased by immobilization and decreased by astaxanthin supplementation. Moreover, especially in the immobilized muscle, the increase of SOD-1 was prevented by astaxanthin supplementation. In addition, as a result of post hoc testing, the expression levels of SOD-1 protein increased in both groups of immobilized rats compared with the control groups providing the same supplementation, whereas the levels of SOD-1 expression in the IM + Ax group were significantly lower than those in the IM group.
Discussion
To our knowledge, this is the first study to report the effects of astaxanthin supplementation on skeletal muscle fibrosis induced by ROS production and profibrotic factor expression in ankle joint immobilization. In the present study, astaxanthin prevented the overproduction of ROS induced by immobilization. In addition, astaxanthin supplementation attenuated increases in collagen fiber, TGF-β1, α-SMA, and SOD-1 expression levels. These novel data demonstrate that astaxanthin supplementation is effective as a nutritional regimen to attenuate muscle fibrosis during joint immobilization.
In the present study, joint immobilization increased collagen fibers and the profibrotic factor (TGF-β1 and α-SMA) protein expression levels. In addition, the DHE fluorescent intensity and SOD-1 protein expression levels were also increased by immobilization. Honda et al. [4] reported that immobilization resulted in the overproduction of the collagen fiber area in the perimysium and endomysium, up-regulation of TGF-β1 mRNA, and increases in α-SMA-positive cells in the soleus muscle. Meanwhile, Kondo et al. [28] reported that immobilization induced overproduction of ROS in skeletal muscle. Therefore, the expression levels of fibrotic factors and oxidative stress were observed in the present study. These results are in agreement with those of previous studies. TGF-β1 is a potent profibrotic cytokine, which plays a pivotal role in the pathogenesis of organ fibrosis [29, 30]. TGF-β1 mRNA upregulation in skeletal muscle affected the differentiation of fibroblasts into myofibroblasts, which possess a high capacity for collagen production [6], early in the immobilization period, and these alterations were associated with the incidence of immobilization-induced muscle fibrosis [4]. Moreover, TGF-β1 has been shown to increase mitochondrial ROS production in different types of cells, which mediate TGF-β-induced α-SMA expression resulting in fibroblast differentiation into myofibroblasts [31, 32]. In contrast, ROS increases TGF-β1 transcription [33] and promotes TGF-β1 activity [34]. Therefore, the overproduced ROS and highly expressed TGF-β1 induced by immobilization would act synergistically to promote α-SMA expression and deteriorate skeletal muscle fibrosis in the present study.
The present study showed the attenuating effects of astaxanthin supplementation on increases in collagen fiber area and profibrotic factors (TGF-β1 and α-SMA) protein expression levels induced by immobilization for the first time (Fig. 4). In addition, the DHE fluorescent intensity and SOD-1 expression levels were also decreased compared with the immobilized group with astaxanthin supplementation. Dihydroethidium produces red fluorescence when oxidized to ethidium bromide by ROS [27]. Meanwhile, SOD-1, one of the cellular antioxidant enzymes, is upregulated in response to increases in oxidative stress [35]. In the present study, ROS production and SOD-1 expression induced by immobilization was prevented by astaxathin supplementation. Astaxanthin quenched the overproduction of ROS, and reduced oxidative stress in previous studies [16, 36]. In addition, astaxanthin actually showed the effects on skeletal muscle by anti-oxidative capacity in various situations, e.g., accelerating lipid utilization during exercise [37], attenuating exercise-induced damage [24], and delaying physical exhaustion [14]. Moreover, astaxanthin supplementation prevented capillary regression via overproduction of ROS and SOD-1 in skeletal muscle during hindlimb unloading in our studies [22, 23]. Therefore, these results in the present study indicated that astaxanthin supplementation could prevent the overproduction of ROS resulting in the relief of oxidative stress in skeletal muscle during immobilization as well as other stress conditions.
Fig. 4.
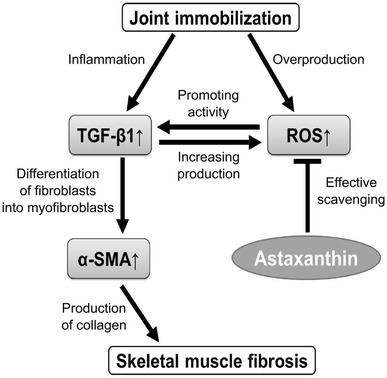
Putative effect of astaxanthin supplementation on immobilization-induced muscle fibrosis. Astaxanthin quenchs the overproduction of ROS and reduces oxidative stress. Astaxanthin prevents the collagen fiber area and profibrotic factors (TGF-β1 and α-SMA) protein expression levels induced by immobilization via inhibiting ROS. As a result, astaxanthin can prevent skeletal muscle fibrosis during immobilization as well as other stress conditions
In numerous types of cells (hepatic satellite cells [38]; lung fibroblasts [31]) with fibrotic situation, ROS production is increased by TGF-β1, which promotes collagen production via activation of fibroblast or differentiation of fibroblasts into myofibroblasts [39]. In contrast, ROS play a central role in the fibrogenic activity of TGF-β1 [40]. Jaffer et al. [41] reported that ROS from mitochondria activated TGF-β1 expression and activity in airway epithelium. In addition, Liu et al. [40] indicated that therapeutic targeting of TGF-β-mediated ROS production might provide a beneficial effect for the treatment of fibrotic diseases. Actually, astaxanthin supplementation prevented fibrosis induced by TGF-β1 in other organs (lung [42, 43]; liver [44]; kidney [45]). Thus, these studies indicate that astaxanthin has preventive effects on TGF-β1-mediated fibrosis through mediating ROS production. In the present study, the preventive effect of ROS overproduction and the attenuating effect of collagen expression was obtained by astaxanthin supplementation. Therefore, astaxanthin supplementation would effectively attenuate immobilization-induced collagen expression in extracellular matrix of skeletal muscle through prevention of ROS overproduction.
We focused on TGF-β1 as a therapeutic target for muscle fibrosis, because it is an essential role for fibrosis development in almost all organs including skeletal muscle [4, 40, 46]. However, the underlying mechanism in fibrosis development is associated not only with TGF-β1 but also connective tissue growth factor (CTGF) [47], which has also been shown to be induced by overproduction of ROS [48, 49]. Thus, immobilization-induced ROS production observed in the present study would promote the expression of CTGF in immobilized muscle. Therefore, the suppression of overproduced ROS by astaxanthin might attenuate CTGF expression in immobilized muscle, leading to amelioration of developing muscle fibrosis.
In the present study, astaxanthin supplementation failed to attenuate immobilization-induced muscle atrophy as observed in the results for muscle wet weight and FCSA (Table 1). In fact, the effects of antioxidants on muscle atrophy is still debated [11, 50–54]. With respect to astaxanthin, Yoshihara et al. and Shibaguchi et al. suggested that the supplementation could attenuate disuse-induced atrophy [55, 56], but Kanazashi et al. showed that supplementation failed to attenuate immobilization-induced muscle atrophy in spite of successful suppression of oxidative stress in disused muscle [22, 23]. These results suggest that the efficacy of astaxanthin on muscle atrophy depends on experimental conditions, e.g., duration, timing, and dosage of astaxanthin administration, and applied animal model of muscle atrophy. Therefore, our results may be partly explained by the fact that we performed astaxanthin supplementation using the same protocol, i.e., supplemdose (100 mg/kg body weight/day) as previously described by Kanazashi et al. However, the molecular mechanisms were not assessed in the present study, so further investigations are needed to determine the effect of astaxanthin on muscle atrophy.
Clinically, joint immobilization-induced muscle contracture results in permanent impairment and disability [57]. Unfortunately, medical professionals are still struggling to find definitive therapies for the prevention and management of joint contracture [58]. In the present study, we revealed the protective effects of astaxanthin on muscle fibrosis via attenuating immobilization-induced activation of profibrogenic TGF-β1 signaling. We believe that astaxanthin supplementation could be an effective preventive intervention without cast removal for muscle contracture during joint immobilization, and contributes to rehabilitation after remobilization.
Conclusion
In conclusion, we find that astaxanthin supplementation is effective to attenuate skeletal muscle fibrosis induced by ankle joint immobilization. This study indicated that astaxanthin supplementation could be an effective preventive intervention on skeletal muscle contracture induced by joint immobilization.
Acknowledgment
This study was supported in part by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant (Grant number 25560065, 25282165, 15K16516, 16K12934).
Compliance with ethical standards
Conflict of interest
The authors declare that there are no conflicts of interest.
References
- 1.Okita M, Yoshimura T, Nakano J, Motomura M, Eguchi K. Effects of reduced joint mobility on sarcomere length, collagen fibril arrangement in the endomysium, and hyaluronan in rat soleus muscle. J Muscle Res Cell Motil. 2004;25(2):159–166. doi: 10.1023/B:JURE.0000035851.12800.39. [DOI] [PubMed] [Google Scholar]
- 2.Hibino I, Okita M, Inoue T, Banno Y, Hosono M. Effect of immobilization on insoluble collagen concentration and type I and type III collagen isoforms of rat soleus muscle. J Jpn Phys Ther Assoc. 2008;11(1):1–6. doi: 10.1298/jjpta.11.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Slimani L, Micol D, Amat J, Delocros G, Meunier B, Taillandier D, Polge C, Bechet D, Dardevet D, Picard B, Attaix D, Listrat A, Combaret L. The worsening of tibialis anterior muscle atrophy during recovery post-immobilization correlates with enhanced connective tissue area, proteolysis, and apoptosis. Am J Physiol Endocrinol Metab. 2012;303(11):E1335–E1347. doi: 10.1152/ajpendo.00379.2012. [DOI] [PubMed] [Google Scholar]
- 4.Honda Y, Sakamoto J, Nakano J, Kataoka H, Sasabe R, Goto K, Tanaka M, Origuchi T, Yoshimura T, Okita M. Upregulation of interleukin-1β/transforming growth factor-β1 and hypoxia relate to molecular mechanisms underlying immobilization-induced muscle contracture. Muscle Nerve. 2015;52(3):419–427. doi: 10.1002/mus.24558. [DOI] [PubMed] [Google Scholar]
- 5.Manoury B, Nenan S, Leclerc O, Guenon I, Boichot E, Planquois JM, Bertrand CP, Lagente V. The absence of reactive oxygen species production protects mice against bleomycin-induced pulmonary fibrosis. Respir Res. 2005;6(1):11. doi: 10.1186/1465-9921-6-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Petrov VV, Fagard RH, Lijnen PJ. Stimulation of collagen production by transforming growth factor-beta1 during differentiation of cardiac fibroblasts to myofibroblasts. Hypertension. 2002;39(2):258–263. doi: 10.1161/hy0202.103268. [DOI] [PubMed] [Google Scholar]
- 7.Lenz AG, Costabel U, Maier KL. Oxidized BAL fluid proteins in patients with interstitial lung diseases. Eur Respir J. 1996;9(2):307–312. doi: 10.1183/09031936.96.09020307. [DOI] [PubMed] [Google Scholar]
- 8.Rottoli P, Magi B, Cianti R, Bargagli E, Vagaggini C, Nikiforakis N, Pallini V, Bini L. Carbonylated proteins in bronchoalveolar lavage of patients with sarcoidosis, pulmonary fibrosis associated with systemic sclerosis and idiopathic pulmonary fibrosis. Proteomics. 2005;5(10):2612–2618. doi: 10.1002/pmic.200401206. [DOI] [PubMed] [Google Scholar]
- 9.Zuo L, Pannell BK. Redox Characterization of Functioning Skeletal Muscle. Front Physiol. 2015;6:338. doi: 10.3389/fphys.2015.00338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Min K, Smuder AJ, Kwon OS, Kavazis AN, Szeto HH, Powers SK. Mitochondrial-targeted antioxidants protect skeletal muscle against immobilization-induced muscle atrophy. J Appl Physiol. 2011;111(5):1459–1466. doi: 10.1152/japplphysiol.00591.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Talbert EE, Smuder AJ, Min K, Kwon OS, Szeto HH, Powers SK. Immobilization-induced activation of key proteolytic systems in skeletal muscles is prevented by a mitochondria-targeted antioxidant. J Appl Physiol. 2013;115(4):529–538. doi: 10.1152/japplphysiol.00471.2013. [DOI] [PubMed] [Google Scholar]
- 12.Gram M, Vigelsø A, Yokota T, Helge JW, Dela F, Mogensen MH. Skeletal muscle mitochondrial H2O2 emission increases with immobilization and decreases after aerobic training in young and older men. J Physiol. 2015;593(17):4011–4027. doi: 10.1113/JP270211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu RM, Desai LP. Reciprocal regulation of TGF-β and reactive oxygen species: a perverse cycle for fibrosis. Redox Biol. 2015;6:565–577. doi: 10.1016/j.redox.2015.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Polotow GW, Vardaris CV, Mihaliuc AR, Gonçalves MS, Pereira B, Ganini D, Barros MP. Astaxanthin supplementation delays physical exhaustion and prevents redox imbalances in plasma and soleus muscles of Wistar rats. Nutrients. 2014;6(12):5819–5838. doi: 10.3390/nu6125819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Liu PH, Aoi W, Takami M, Terajima H, Tanimura Y, Naito Y, Itoh Y, Yoshikawa T. The astaxanthin-induced improvement in lipid metabolism during exercise is mediated by a PGC-1α increase in skeletal muscle. J Clin Biochem Nutr. 2014;54(2):86–89. doi: 10.3164/jcbn.13-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Guerin M, Huntley ME, Olaizola M. Haematococcus astaxanthin: applications for human health and nutrition. Trends Biotechnol. 2003;21(5):210–216. doi: 10.1016/S0167-7799(03)00078-7. [DOI] [PubMed] [Google Scholar]
- 17.Shibata A, Kiba Y, Akati N, Fukuzawa K, Terada H. Molecular characteristics of astaxanthin and beta-carotene in the phospholipid monolayer and their distributions in the phospholipid bilayer. Chem Phys Lipids. 2001;113(1–2):11–22. doi: 10.1016/S0009-3084(01)00136-0. [DOI] [PubMed] [Google Scholar]
- 18.Nishida Y, Yamashita E, Miki W. Quenching activities of common hydrophilic and lipophilic antioxidants against singlet oxygen using chemiluminescence detection system. Carotenoid Sci. 2007;11:16–20. [Google Scholar]
- 19.Rodrigues E, Mariutti LR, Mercadante AZ. Scavenging capacity of marine carotenoids against reactive oxygen and nitrogen species in a membrane-mimicking system. Mar Drugs. 2012;10(8):1784–1798. doi: 10.3390/md10081784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kidd P. Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Altern Med Rev. 2011;16(4):355–364. [PubMed] [Google Scholar]
- 21.McNulty HP, Byun J, Lockwood SF, Jacob RF, Mason RP. Differential effects of carotenoids on lipid peroxidation due to membrane interactions: X-ray diffraction analysis. Biochim Biophys Acta. 2007;1768(1):167–174. doi: 10.1016/j.bbamem.2006.09.010. [DOI] [PubMed] [Google Scholar]
- 22.Kanazashi M, Okumura Y, Al-Nassan S, Murakami S, Kondo H, Nagatomo F, Fujita N, Ishihara A, Roy RR, Fujino H. Protective effects of astaxanthin on capillary regression in atrophied soleus muscle of rats. Acta Physiol (Oxf) 2013;207(2):405–415. doi: 10.1111/apha.12018. [DOI] [PubMed] [Google Scholar]
- 23.Kanazashi M, Tanaka M, Murakami S, Kondo H, Nagatomo F, Ishihara A, Roy RR, Fujino H. Amelioration of capillary regression and atrophy of the soleus muscle in hindlimb-unloaded rats by astaxanthin supplementation and intermittent loading. Exp Physiol. 2014;99(8):1065–1077. doi: 10.1113/expphysiol.2014.079988. [DOI] [PubMed] [Google Scholar]
- 24.Aoi W, Naito Y, Sakuma K, Kuchide M, Tokuda H, Maoka T, Toyokuni S, Oka S, Yasuhara M, Yoshikawa T. Astaxanthin limits exercise-induced skeletal and cardiac muscle damage in mice. Antioxid Redox Signal. 2003;5(1):139–144. doi: 10.1089/152308603321223630. [DOI] [PubMed] [Google Scholar]
- 25.Takeuchi K, Hatade T, Wakamiya S, Fujita N, Arakawa T, Miki A. Heat stress promotes skeletal muscle regeneration after crush injury in rats. Acta Histochem. 2014;116(2):327–334. doi: 10.1016/j.acthis.2013.08.010. [DOI] [PubMed] [Google Scholar]
- 26.Tanaka M, Kanazashi M, Maezawa T, Kondo H, Fujino H. Abnormalities in three-dimensional capillary architecture and imbalance between vascular endothelial growth factor-A and thrombospondin-1 in soleus muscle of ovariectomized rat. Acta Histochem. 2015;117(7):605–611. doi: 10.1016/j.acthis.2015.06.001. [DOI] [PubMed] [Google Scholar]
- 27.Mülsch A, Oelze M, Klöss S, Mollnau H, Töpfer A, Smolenski A, Walter U, Stasch JP, Warnholtz A, Hink U, Meinertz T, Münzel T. Effects of in vivo nitroglycerin treatment on activity and expression of the guanylyl cyclase and cGMP-dependent protein kinase and their downstream target vasodilator-stimulated phosphoprotein in aorta. Circulation. 2001;103:2188–2194. doi: 10.1161/01.CIR.103.17.2188. [DOI] [PubMed] [Google Scholar]
- 28.Kondo H, Miura M, Itokawa Y. Oxidative stress in skeletal muscle atrophied by immobilization. Acta Physiol Scand. 1991;142(4):527–528. doi: 10.1111/j.1748-1716.1991.tb09191.x. [DOI] [PubMed] [Google Scholar]
- 29.Border WA, Nobble NA. Transforming growth factor beta in tissue fibrosis. N Engl J Med. 1994;331(19):1287–1292. doi: 10.1056/NEJM199411103311907. [DOI] [PubMed] [Google Scholar]
- 30.Williams E, Iredale J. Hepatic regeneration and TGF-beta: growing to a prosperous perfection. Gut. 2000;46(5):593–594. doi: 10.1136/gut.46.5.593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jain M, Rivera S, Monclus EA, Synenki L, Zirk A, Eisenbart J, Feghali-Bostwick C, Mutlu GM, Budinger GR, Chandel NS. Mitochondrial reactive oxygen species regulate transforming growth factor-β signaling. J Biol Chem. 2013;288(2):770–777. doi: 10.1074/jbc.M112.431973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bracey NA, Gershkovich B, Chun J, Vilaysane A, Meijndert HC, Wright JR, Jr, Fedak PW, Beck PL, Muruve DA, Duff HJ. Mitochondrial NLRP3 protein induces reactive oxygen species to promote Smad protein signaling and fibrosis independent from the inflammasome. J Biol Chem. 2014;289(28):19571–19584. doi: 10.1074/jbc.M114.550624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bellocq A, Azoulay E, Marullo S, Flahault A, Fouquwray B, Philippe C, Cardranel J, Baud L. Reactive oxygen and nitrogen intermediates increase transforming growth factor-beta1 release from human epithelial alveolar cells through two different mechanisms. Am J Respir Cell Mol Biol. 1999;21(1):128–136. doi: 10.1165/ajrcmb.21.1.3379. [DOI] [PubMed] [Google Scholar]
- 34.Qi S, den Hartoq GJ, Bast A. Superoxide radicals increase transforming growth factor-beta1 and collagen release from human lung fibroblasts via cellular influx through chloride channels. Toxicol Appl Pharmacol. 2009;237(1):111–118. doi: 10.1016/j.taap.2009.02.019. [DOI] [PubMed] [Google Scholar]
- 35.Chan KM, Decker EA. Endogenous skeletal muscle antioxidants. Crit Rev Food Sci Nutr. 1994;34(4):403–426. doi: 10.1080/10408399409527669. [DOI] [PubMed] [Google Scholar]
- 36.Hussein G, Sankawa U, Goto H, Matsumoto K, Watanabe H. Astaxanthin, a carotenoid with potential in human health and nutrition. J Nat Prod. 2006;69(3):443–449. doi: 10.1021/np050354+. [DOI] [PubMed] [Google Scholar]
- 37.Aoi W, Naito Y, Takanami Y, Ishii T, Kawai Y, Akagiri S, Kato Y, Osawa T, Yoshikawa T. Astaxanthin improves muscle lipid metabolism in exercise via inhibitory effect of oxidative CPT I modification. Biochem Biophys Res Commun. 2008;366(4):892–897. doi: 10.1016/j.bbrc.2007.12.019. [DOI] [PubMed] [Google Scholar]
- 38.Yang Y, Kim B, Park YK, Koo SI, Lee JY. Astaxanthin prevents TGFβ1-induced pro-fibrogenic gene expression by inhibiting Smad3 activation in hepatic stellate cells. Biochim Biophys Acta. 2015;1850(1):178–185. doi: 10.1016/j.bbagen.2014.10.014. [DOI] [PubMed] [Google Scholar]
- 39.Desmoulie`re A, Geinoz A, Gabbiani F, Gabbiani G. Transforming growth factor-b1 induces a-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J Cell Biol. 1993;122:103–111. doi: 10.1083/jcb.122.1.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Liu RM, Gaston Pravia KA. Oxidative stress and glutathione in TGF-beta-mediated fibrogenesis. Free Radic Biol Med. 2010;48(1):1–15. doi: 10.1016/j.freeradbiomed.2009.09.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jaffer OA, Carter AB, Sanders PN, Dibbern ME, Winters CJ, Murthy S, Ryan AJ, Rokita AG, Prasad AM, Zabner J, Kline JN, Grumbach IM, Anderson ME. Mitochondrial-targeted antioxidant therapy decreases transforming growth factor-β-mediated collagen production in a murine asthma model. Am J Respir Cell Mol Biol. 2015;52(1):106–115. doi: 10.1165/rcmb.2013-0519OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhang J, Xu P, Wang Y, Wang M, Li H, Lin S, Mao C, Wang B, Song X, Lv C. Astaxanthin prevents pulmonary fibrosis by promoting myofibroblast apoptosis dependent on Drp1-mediated mitochondrial fission. J Cell Mol Med. 2015;19(9):2215–2231. doi: 10.1111/jcmm.12609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wang M, Zhang J, Song X, Liu W, Zhang L, Wang X, Lv C. Astaxanthin ameliorates lung fibrosis in vivo and in vitro by preventing transdifferentiation, inhibiting proliferation, and promoting apoptosis of activated cells. Food Chem Toxicol. 2013;56:450–458. doi: 10.1016/j.fct.2013.03.004. [DOI] [PubMed] [Google Scholar]
- 44.Shen M, Chen K, Lu J, Cheng P, Xu L, Dai W, Wang F, He L, Zhang Y, Chengfen W, Li J, Yang J, Zhu R, Zhang H, Zheng Y, Zhou Y, Guo C. Protective effect of astaxanthin on liver fibrosis through modulation of TGF-β1 expression and autophagy. Mediators Inflamm. 2014 doi: 10.1155/2014/954502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Alex M, Sauganth Paul MV, Abhilash M, Mathews VV, Anilkumar TV, Nair RH. Astaxanthin modulates osteopontin and transforming growth factor β1 expression levels in a rat model of nephrolithiasis: a comparison with citrate administration. BJU Int. 2014;114(3):458–466. doi: 10.1111/bju.12537. [DOI] [PubMed] [Google Scholar]
- 46.McCormick LL, Zhang Y, Tootell E, Gilliam AC. Anti-TGF-beta treatment prevents skin and lung fibrosis in murine sclerodermatous graft-versus-host disease: a model for human scleroderma. J Immunol. 1999;163(10):5693–5699. [PubMed] [Google Scholar]
- 47.Iyer P, Maddala R, Pattabiraman PP, Rao PV. Connective tissue growth factor-mediated upregulation of neuromedin U expression in trabecular meshwork cells and its role in homeostasis of aqueous humor outflow. Invest Ophthalmol Vis Sci 26. 2012;53(8):4952–4962. doi: 10.1167/iovs.12-9681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Park SK, Kim J, Seomun Y, Choi J, Kim DH, Han IO, Lee EH, Chung SK, Joo CK. Hydrogen peroxide is a novel inducer of connective tissue growth factor. Biochem Biophys Res Commun 22. 2001;284(4):966–971. doi: 10.1006/bbrc.2001.5058. [DOI] [PubMed] [Google Scholar]
- 49.Tsai CC, Wu SB, Chang PC, Wei YH. Alteration of connective tissue growth factor (CTGF) expression in orbital fibroblasts from patients with Graves’ ophthalmopathy. PLoS ONE 24. 2015;10(11):e0143514. doi: 10.1371/journal.pone.0143514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.McClung JM, Whidden MA, Kavazis AN, Falk DJ, Deruisseau KC, Powers SK. Redox regulation of diaphragm proteolysis during mechanical ventilation. Am J Physiol Regul Integr Comp Physiol. 2008;294(5):R1608–R1617. doi: 10.1152/ajpregu.00044.2008. [DOI] [PubMed] [Google Scholar]
- 51.Desaphy JF, Pierno S, Liantonio A, Giannuzzi V, Digennaro C, Dinardo MM, Camerino GM, Ricciuti P, Brocca L, Pellegrino MA, Bottinelli R, Camerino DC. Antioxidant treatment of hindlimb-unloaded mouse counteracts fiber type transition but not atrophy of disused muscles. Pharmacol Res. 2010;61(6):553–563. doi: 10.1016/j.phrs.2010.01.012. [DOI] [PubMed] [Google Scholar]
- 52.Brocca L, Pellegrino MA, Desaphy JF, Pierno S, Camerino DC, Bottinelli R. Is oxidative stress a cause or consequence of disuse muscle atrophy in mice? A proteomic approach in hindlimb-unloaded mice. Exp Physiol. 2010;95(2):331–350. doi: 10.1113/expphysiol.2009.050245. [DOI] [PubMed] [Google Scholar]
- 53.Powers SK, Smuder AJ, Criswell DS. Mechanistic links between oxidative stress and disuse muscle atrophy. Antioxid Redox Signal 1. 2011;15(9):2519–2528. doi: 10.1089/ars.2011.3973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bennett BT, Mohamed JS, Alway SE. Effects of resveratrol on the recovery of muscle mass following disuse in the plantaris muscle of aged rats. PLoS ONE 12. 2013;8(12):e83518. doi: 10.1371/journal.pone.0083518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Yoshihara T, Yamamoto Y, Shibaguchi T, Miyaji N, Kakigi R, Naito H, Goto K, Ohmori D, Yoshioka T, Sugiura T. Dietary astaxanthin supplementation attenuates disuse-induced muscle atrophy and myonuclear apoptosis in the rat soleus muscle. J Physiol Sci. 2016 doi: 10.1007/s12576-016-0453-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Shibaguchi T, Yamaguchi Y, Miyaji N, Yoshihara T, Naito H, Goto K, Ohmori D, Yoshioka T, Sugiura T (2016) Astaxanthin intake attenuates muscle atrophy caused by immobilization in rats. Physiol Rep 4(15). doi:10.14814/phy2.12885 [DOI] [PMC free article] [PubMed]
- 57.Trudel G, Jabi M, Uhthoff HK. Localized and adaptive synoviocyte proliferation characteristics in rat knee joint contractures secondary to immobility. Arch Phys Med Rehabil. 2003;84(9):1350–1356. doi: 10.1016/S0003-9993(03)00233-8. [DOI] [PubMed] [Google Scholar]
- 58.Kojima S, Hoso M, Watanabe M, Matsuzaki T, Hibino I, Sasaki K. Experimental Joint immobilization and remobilization in the rats. J Phys Ther Sci. 2014;26(6):865–887. doi: 10.1589/jpts.26.865. [DOI] [PMC free article] [PubMed] [Google Scholar]