

Abstract
We evaluated the effects of moderate-intensity continuous training (MICT) and high-intensity interval training (HIIT) protocols on the alterations in oxidative stress parameters caused by a high-fat diet (HFD), in the blood and liver of rats. The HFD enhanced thiobarbituric acid reactive substances (TBA-RS) and protein carbonyl content, while reducing total sulfhydryl content and catalase (CAT) and glutathione peroxidase (GSH-Px) activities in the blood. Both training protocols prevented an increase in TBA-RS and protein carbonyl content, and prevented a reduction in CAT. HIIT protocol enhanced SOD activity. In the liver, HFD didn’t alter TBA-RS, total sulfhydryl content or SOD, but increased protein carbonyl content and CAT and decreased GSH-Px. The exercise protocols prevented the increase in protein carbonyl content and the MICT protocol prevented an alteration in CAT. In conclusion, HFD elicits oxidative stress in the blood and liver and both protocols prevented most of the alterations in the oxidative stress parameters.
Keywords: Aerobic exercise training protocols, Oxidative stress, Blood, Liver
Introduction
Obesity is considered to be a polygenic and multifactorial disease, due to an interplay between genetic susceptibility and dietary, environmental and metabolic variables [1]. The disease is characterized by excessive body fat, resulting from the deposit of triacylglycerol and free fatty acids, which consequently lead to weight gain and increased body mass index (BMI) [2, 3].
Increasing BMI can lead to the respiratory, metabolic and vascular systems suffering stress, due to the greater than usual work load, and induces a decrease in physiological reserve with significant/negative changes to the homeostasis of an organism [2]. Studies have shown that the prevalence of obesity in association with other comorbidities is rapidly increasing, and is closely related to the adoption of a sedentary lifestyle [4, 5].
Another important aspect of obesity is that it is associated with a chronic inflammatory state, characterized by abnormalities in the production of proinflammatory cytokines, such as leptin, whose function is to encode genes involved in the regulation of body weight [6]. The presence of obesity-related chronic inflammation predisposes individuals to many conditions, including type 2 diabetes mellitus, dyslipidemia and hypertension, reduces immune capacity, and increases metabolic and cardiovascular risks [7, 8]. Evidence suggests that the imbalance between pro-oxidants and antioxidant defense mechanisms leads to oxidative stress, which is involved in the etiopathogenesis of various diseases, including obesity [9].
Physical exercise is an important non-drug therapy that can be used to prevent and treat obesity and its physical and metabolic cellular complications [8, 10]. Numerous studies have since shown that other tissues respond to exercise by improving the antioxidant defense system and by increasing resistance against the actions of free radicals [11–14]. Considering the high global rates of obesity, in association with sedentary lifestyles, this study aimed to evaluate the effects of two aerobic exercise training protocols (AETP), moderate-intensity continuous training (MICT) and high-intensity interval training (HIIT), on the alterations in oxidative stress parameters caused by a high-fat diet (HFD) in the blood and the liver of rats.
Materials and methods
Animal model
Adult male Wistar rats (60 days old) were obtained from the UNIVALI (Universidade do Vale de Itajai, Itajai, Brazil) breeding colony, and were randomly assigned into HFD (protein, 20%; carbohydrate, 20%; lipid, 60%: Prag Soluções Biosciências, Jaú-SP-Brazil) or normal diet (ND) (protein, 20%; carbohydrate, 70%; lipid, 10%: Nuvital Nutrientes, Curitiba-PR-Brazil) for 8 weeks. The rats were maintained on a light–dark inverted cycle (12:12-h light–dark cycle), with a temperature-controlled environment (22 ± 1 °C) and free access to respective diets and tap water. Upon completion of 8 weeks of the diet, animals from the HFD were randomly assigned to an untrained group (HFD-UNT; n = 6) or to two groups that trained with different AETPs, as follows: HFD-MICT, n = 6 and HFD-HIIT, n = 6. Animals from the ND were assigned to an untrained group (ND-UNT; n = 6). After this phase, these 4 experimental groups underwent 9 more weeks of diet and training. The AETPs are described in detail below. All rats were euthanized at 2 days after the last day of training. This study was conducted in accordance with the Principles of Laboratory Animal Care (NIH publication 85–23, revised 1985), and was approved by the UNIVILLE (Universidade da Região de Joinville, Joinville, Brazil) Ethics Committee (Protocol No. 002/213–COEA). All chemicals used in the analysis of oxidative stress parameters were purchased from Sigma Chemical Co., St Louis, MO, USA.
Experimental design
Aerobic exercise training protocols (AETP): (1) moderate-intensity continuous training (MICT) and (2) high-intensity interval training (HIIT)
Before the AETP, rats were conditioned to treadmill exercises over a period of a week (10 min of exercise per session), and at the end of this period, they were subjected to a graded treadmill exercise test. During the test, rats were placed on the treadmill and allowed to acclimatize for at least 10 min. Rats ran on a graded treadmill until exhaustion at 20° inclination; the speed was started at 6 m/min and was increased by 3 m/min every 3 min until rats were unable to run [15]. The graded treadmill exercise test was performed prior to exercise training and then during the 4th and 8th weeks of exercise training.
Both AETPs consisted of an 8-week program running on a motorized treadmill (KT-4000 model INBRAMED, RS, Brazil), at a 20° inclination, 5 days a week.
(1) MICT: this protocol was performed at a treadmill speed corresponding to 60% of the maximum running speed obtained from the graded treadmill test, which was kept unchanged throughout the entire session. (2) HIIT was performed in such a way that rats ran for 3 min at 60% of the maximum running speed, followed by 4-min intervals at 85% of the maximum running speed, which was repeated 7 times, so each HIIT session lasted for 49 min. MICT and HIIT protocols were of matched volume, meaning that the total running distances in each session of either MICT or HIIT were identical; therefore, the MICT session duration was adjusted to match HIIT distance [16]. A 5-min warm up at 40% of the maximum running speed was performed by both trained groups before each AETP session.
HFD-UNT and ND-UNT animals were placed on the treadmill twice a week for 10 min on each day at 40% of the maximum running speed to maintain running skills.
Experimental protocols
Erythrocyte and plasma preparation
Erythrocytes and plasma were prepared from whole blood samples obtained from rats. Whole blood was collected and transferred to heparinized tubes for erythrocyte separation. Blood samples were centrifuged at 1000×g, plasma was then removed by aspiration and frozen at − 80 °C until use in assays. Erythrocytes were washed 3 times with cold saline solution (0.153 mol/l sodium chloride). Lysates were prepared by the addition of 1 ml of distilled water to 100 μl of washed erythrocytes and frozen at − 80 °C until determination of the antioxidant enzyme activities.
For antioxidant enzyme activity determination, erythrocytes were frozen and thawed 3 times, and centrifuged at 13,500×g for 10 min. The supernatant was diluted in order to achieve an approximate concentration of 0.5 mg/ml of protein.
Tissue preparation
After decapitation of rats, the livers were removed and kept on ice-cold buffered sodium phosphate solution (containing sodium phosphate mono and dibasic at a concentration of 20 mM, 140 mM KCl, pH 7.4). The liver was homogenized in ten volumes (1:10 w/v) of appropriate buffer, according to the technique to be performed. Homogenates were prepared using a Potter-Elvehejem homogenizer (Remi motors, Mumbai, India) by passing 5 pulses and centrifuging at 800×g for 10 min at 4 °C before discarding nuclei and cell debris. The pellet was discarded and the supernatant was divided into aliquots and stored at − 80 °C for assaying the activity of antioxidant enzymes, damage to proteins and estimation of lipid peroxidation [17].
Thiobarbituric acid reactive substances (TBA-RS)
TBA-RS was determined according to the method described by Ohkawa et al. [18]. TBA-RS methodology measures malondialdehyde (MDA), a product of lipoperoxidation, caused mainly by hydroxyl free radicals. Liver and plasma in 1.15% KCl were mixed with 20% trichloroacetic acid and 0.8% thiobarbituric acid and heated in a boiling water bath for 60 min. TBA-RS were measured by determining the absorbance at 535 nm. A calibration curve was obtained using 1,1,3,3-tetramethoxypropane as the MDA precursor and each curve point was subjected to the same treatment as that of the supernatants. TBA-RS content was calculated as nanomoles of MDA formed per milligram of protein.
Total sulfhydryl content
The total thiol group concentration was determined by the method of Aksenov and Markesbery [19]. Briefly, 50 µl of homogenate and plasma were added to 1 ml of phosphate-buffered saline (PBS), pH 7.4, containing 1 mM ethylenediaminetetraacetic acid (EDTA). The reaction was started by the addition of 30 µl of 10 mM 5,5´-dithiobis-(2-nitrobenzoic acid) (DTNB) and incubated for 30 min at room temperature in a dark room. Total sulfhydryl content was determined by measuring the absorbance at 412 nm. Analyses of a blank (DTNB absorbance) was also performed. Results are reported as nmol 3-thio-2-nitrobenzoic acid (TNB)/mg protein.
Protein carbonyl content
Carbonyl content was assayed by a method described by Reznick and Packer [20], based on the reaction of protein carbonyls with dinitrophenylhydrazine to form dinitrophenylhydrazone, a yellow compound, measured spectrophotometrically at 370 nm. Briefly, 200 µl of homogenate and plasma were added to plastic tubes containing 400 µl of 10 mM dinitrophenylhydrazine (prepared in 2 M HCl). Samples were kept in the dark for 1 h and vortexed every 15 min. Subsequently, 500 µl of 20% trichloroacetic acid were added to each tube. The mixture was vortexed and centrifuged at 14,000×g for 3 min and the supernatant obtained was discarded. The pellet was washed with 1 ml ethanol/ethyl acetate (1:1 v/v), vortexed and centrifuged at 14,000×g for 3 min. The supernatant was then discarded and the pellet re-suspended in 600 µl of 6 M guanidine (prepared in a 20-mM potassium phosphate solution, pH 2.3), before vortexing and incubating at 60°C for 15 min. Samples were then centrifuged at 14,000×g for 3 min and the supernatant was used to measure absorbance at 370 nm (UV) in a quartz cuvette. Results were reported as carbonyl content (nmol/mg protein).
Catalase assay (CAT)
CAT activity was assayed by the method of Aebi [21] using a UV–visible Shimadzu spectrophotometer. The method used is based on the disappearance of H2O2 at 240 nm in a reaction medium containing 20 mM H2O2, 0.1% Triton X-100, 10 mM potassium phosphate buffer, pH 7.0, and 0.1–0.3 mg protein/ml. One CAT unit is defined as 1 μmol of H2O2 consumed per minute and the specific activity is calculated as CAT units/mg protein.
Glutathione peroxidase assay (GSH-Px)
GSH-Px activity was measured by the method of Wendel [22] using tert-butyl-hydroperoxide as substrate. NADPH disappearance was monitored at 340 nm using a UV–visible Shimadzu spectrophotometer. The medium contained 2 mM GSH, 0.15 U/ml GSH reductase, 0.4 mM azide, 0.5 mM tert-butyl-hydroperoxide and 0.1 mM NADPH. One GSH-Px unit is defined as 1 μmol of NADPH consumed per minute and the specific activity is presented as GSH-Px units/mg protein.
Superoxide dismutase assay (SOD)
The method used to assay SOD activity is based on the capacity of pyrogallol to autoxidize, a process highly dependent on superoxide (O•−2), which is a substrate for SOD [23]. Briefly, to 15 μl of each sample, 215 μl of a mixture containing 50 μM Tris buffer, pH 8.2, 1 μM EDTA and 30 μM CAT were added. Subsequently, 20 μl of pyrogallol were added and the absorbance was immediately recorded every 30 s for 3 min at 420 nm using a UV–visible Shimadzu spectrophotometer. The inhibition of autoxidation of pyrogallol occurs in the presence of SOD, whose activity can be indirectly assayed spectrophotometrically. A calibration curve was performed with purified SOD as reference, to calculate the activity of SOD present in the samples. One SOD unit is defined as the amount of SOD necessary to inhibit 50% of pyrogallol autoxidation and the specific activity is reported as SOD units/mg protein.
Protein determination
Protein was measured by the Lowry [24] method, using serum bovine albumin as standard.
Statistical analysis
The data are presented as means and standard error of the means (mean ± SEM). One-way ANOVA with post hoc testing by Duncan was used to compare the effects of training in all analyses. Results were considered to be statistically significant when the p value was < 0.05. We used the statistical program SPSS for Windows, version 12 (SPSS, Chicago, IL, USA). The Kolmogorov–Smirnov normality test and Levene homogeneity test were performed before ANOVA analysis.
Results
First, we investigated the effect of two AETPs, MICT and HIIT, on TBA-RS (an important parameter of lipid peroxidation), total sulfhydryl content and total carbonyl content (important parameters of protein damage) and on the activity of antioxidant enzymes (CAT, SOD and GSH-Px) in the erythrocytes/plasma of rats subjected to HFD.
Figure 1 shows that HFD significantly enhanced TBA-RS (Fig. 1a) (p < 0.001) and protein carbonyl content (Fig. 1c) (p < 0.05) in the HFD-UNT group, when compared to the ND-UNT group, while the MICT and HIIT protocols prevented the increase in TBA-RS levels (p < 0.001) and abolished the increase in protein carbonyl content in the HFD-MICT and HFD-HIIT groups, respectively. Furthermore, Fig. 1b shows that HFD significantly reduced total sulfhydryl content in the HFD-UNT group, when compared to the ND-UNT group (p < 0.01); however, this parameter was not prevented by the different training protocols (MICT and HIIT).
Fig. 1.
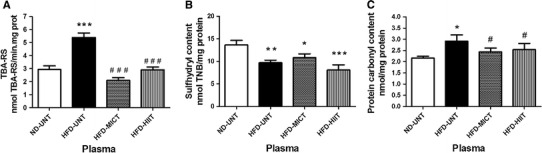
Oxidative stress parameters, a thiobarbituric acid reactive substances (TBA-RS); b total sulfhydryl content; and c protein carbonyl content in the plasma. The experimental animals were divided into four groups: ND-UNT (normal diet untrained; n = 6), HFD-UNT (high-fat diet untrained; n = 6), HFD-MICT (high-fat diet + moderate-intensity continuous training; n = 6) and HFD-HIIT (high-fat diet + high-intensity interval training; n = 6). The data are presented as mean ± SEM and were compared between groups by one-way analysis of variance (ANOVA) with post hoc Duncan. ***p < 0.001; **p < 0.01; *p < 0.05 vs ND-UNT; ### p < 0.001; # p < 0.05 vs HFD-UNT
As can be seen in Fig. 2, HFD significantly diminished the activity of CAT (Fig. 2b) (p < 0.05) and GSH-Px (Fig. 2c) (p < 0.001) in the HFD-UNT group, when compared to the ND-UNT group, but did not alter SOD (Fig. 2a) activity (p > 0.05). With regard to SOD activity, the HIIT protocol significantly enhanced the activity of this enzyme in the HFD-HIIT group, when compared to the HFD-UNT and ND-UNT groups (p < 0.05). The training protocols prevented the reduction in CAT activity observed in the HFD-UNT group (p < 0.001) and the HIIT protocol significantly increased the activity of this enzyme (HFD-HIIT group), when compared to the ND-UNT group (p < 0.01); however, the different AETPs did not prevent the reduction in GSH-Px activity that was observed in the HFD-UNT rats.
Fig. 2 .
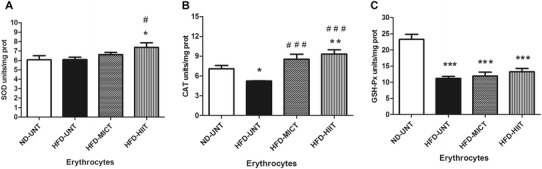
Antioxidant enzymes, a superoxide dismutase (SOD); b catalase (CAT); and c glutathione peroxidase assay (GSH-Px) in the erythrocytes. The experimental animals were divided into four groups: ND-UNT (normal diet untrained; n = 6), HFD-UNT (high-fat diet untrained; n = 6), HFD-MICT (high-fat diet + moderate-intensity continuous training; n = 6), and HFD-HIIT (high-fat diet + high-intensity interval training; n = 6). The data are presented as mean ± SEM and were compared between groups by one-way analysis of variance (ANOVA) with post hoc Duncan. ***p < 0.001; **p < 0.01; *p < 0.05 vs ND-UNT; ### p < 0.001; # p < 0.05 vs HFD-UNT
Subsequently, we investigated the effects of the two AETPs, MICT and HIIT, on the same parameters of oxidative stress in the liver of rats subjected to HFD. Figure 3 shows that HFD did not alter TBA-RS (Fig. 3a) (p > 0.05) and total sulfhydryl content (Fig. 3b) (p > 0.05) in the HFD-UNT group, when compared to the ND-UNT group in the liver of rats. However, protein carbonyl content (Fig. 3c) was significantly increased in the HFD-UNT group, when compared to the ND-UNT group (p < 0.05). Furthermore, the MICT and HIIT protocols prevented the increase in protein carbonyl content in the liver of HFD rats (p < 0.05).
Fig. 3.
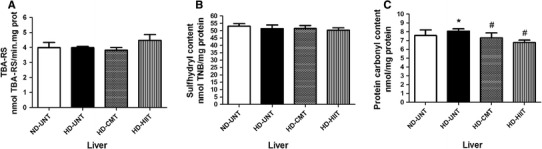
Oxidative stress parameters, a thiobarbituric acid reactive substances (TBA-RS); b total sulfhydryl content; and c protein carbonyl content in the liver. The experimental animals were divided into four groups: ND-UNT (normal diet untrained; n = 6), HFD-UNT (high-fat diet untrained; n = 6), HFD-MICT (high-fat diet + moderate-intensity continuous training; n = 6) and HFD-HIIT (high-fat diet + high-intensity interval training; n = 6). The data are presented as mean ± SEM and were compared between groups by one-way analysis of variance (ANOVA) with post hoc Duncan. *p < 0.05 vs ND-UNT; # p < 0.05 vs HFD-UNT
With regard to SOD activity, Fig. 4a shows that HFD did not alter this enzyme’s activity in the HFD-UNT group, when compared to the ND-UNT group (p > 0.05). Likewise, AETPs did not alter SOD activity, when comparing the HFD-MICT and HFD-HIIT groups to the HFD-UNT and ND-UNT groups. In contrast, Fig. 4b shows that HFD significantly increased CAT activity in the HFD-UNT group, when compared to the ND-UNT group (p < 0.01). The MICT protocol prevented this alteration (HFD-MICT group), when compared to the HFD-UNT group (p < 0.01), while the HIIT protocol (HFD-HIIT group) was unable to prevent this alteration. With regard to GSH-Px activity, Fig. 4c demonstrates that HFD significantly diminished the activity of this enzyme in the HFD-UNT group, when compared to the ND-UNT group (p < 0.001). Furthermore, the two training protocols (MICT and HIIT) were unable to prevent this alteration.
Fig. 4.
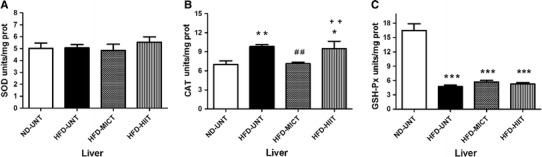
Antioxidant enzymes, a superoxide dismutase (SOD); b catalase (CAT); and c glutathione peroxidase assay (GSH-Px) in the liver. The experimental animals were divided into four groups: ND-UNT (normal diet untrained; n = 6), HFD-UNT (high-fat diet untrained; n = 6), HFD-MICT (high-fat diet + moderate-intensity continuous training; n = 6) and HFD-HIIT (high-fat diet + high-intensity interval training; n = 6). The data are presented as mean ± SEM and were compared between groups by one-way analysis of variance (ANOVA) with post hoc Duncan. ***p < 0.001; **p < 0.01; *p < 0.05 vs ND-UNT; ## p < 0.01 vs HFD-UNT; ++ p < 0.01 vs HFD-MICT
Discussion
Oxidative stress is considered to be an imbalance between pro-oxidant and antioxidant factors, inducing cellular injury as a consequence. This process contributes significantly to the pathophysiology of several diseases, due to the increased production of reactive oxygen species (ROS) and reactive nitrogen species (RNS), which are associated with inflammatory processes in obesity and its comorbidities, including hypertension, dyslipidemias, diabetes, cardiovascular disease, sleep apnea, osteoarthritis and some cancers [25].
The link between obesity, inflammation and oxidative stress occurs via various physicochemical mechanisms, such as the elevation of circulating glucose levels, an increase in the production and storage of lipids, stimulation of mitochondrial fatty acid oxidation and also the elevation of proinflammatory cytokines, such as tumor necrosis factor alpha (TNF-α) and interleukin-6 (IL-6), leading to the oxidative stress that is present in the obese condition [26, 27]. As such, the aim of this study was to investigate the effects of two AETPs, MICT and HIIT, on parameters of oxidative stress, such as TBA-RS, total sulfhydryl content and total carbonyl content, to correlate with pro-oxidant factors, and on the activity of antioxidant enzymes (CAT, SOD and GSH-Px), in the erythrocytes/plasma and liver of rats subjected to HFD, with the intention of minimizing the oxidative stress caused in this condition.
This study revealed that HFD significantly enhanced TBA-RS and protein carbonyl content and reduced total sulfhydryl content in the HFD-UNT group, when compared to the ND-UNT group in the plasma of rats. These data corroborate those of Yida [28], where HFD feeding caused inflammation and oxidative stress in the liver and kidney of rats, and with Charradi [29], who found an increase in MDA and carbonyl content in the plasma of rats that had followed a 6-week HFD.
In regard to AETPs, we decided to investigate the effects of MICT versus HIIT because there is still controversy about these two protocols [30]. Based on the hormesis hypothesis, strenuous exercise should increase oxidative stress [31], but there are, in fact, studies reporting similar beneficial outcomes with both protocols or even reporting the HIIT protocol as more effective in preventing oxidative stress [32].
In the present study, the MICT and HIIT protocols prevented an increase in TBA-RS, and partially abolished the alteration in protein carbonyl content; however, the training protocols did not prevent a reduction in total sulfhydryl content in the HFD group. TBA-RS reflects the content of malondialdehyde, the most abundant individual aldehyde that results from the lipid peroxidation processes [33]. In contrast, the level of protein carbonyl and sulfhydryl content is widely used as a marker of oxidative protein damage. Thus, our data indicate that HFD elicits oxidative damage to lipids and proteins in the blood of obese rats, and that the training protocols were more effective at protecting against damage caused to lipids, and therefore lipid peroxidation.
The antioxidant enzymes are important endogenous defenses required to inhibit the formation of ROS or to promote the removal of free radicals and their precursors. The main antioxidant enzymes are CAT, SOD, GSH-Px and glutathione reductase. CAT is a ferric heme protein that directly catalyzes the decomposition of hydrogen peroxide (H2O2). SOD removes the superoxide anion by accelerating the rate of its dismutation to H2O2 [34]. GSH-Px catalyzes the decomposition of H2O2 or organic peroxides, using the glutathione (GSH) cofactor, which is oxidized (GSSG). To complete the redox cycle, glutathione reductase reduces GSSG to GSH [35].
With respect to the antioxidant enzymes, HFD diminished the activity of CAT and GSH-Px in the HFD-UNT group, when compared to the ND-UNT group, but did not alter SOD activity in the erythrocytes of obese rats. Considering the AETPs, the HIIT protocol enhanced the activity of SOD in the HFD-HIIT group, suggesting that this protocol modulates this enzyme’s activity. Both protocols were able to prevent the reduction in CAT activity observed in the HFD-UNT group, and increased the activity of this enzyme, when compared to the ND-UNT group, suggesting that the protocols modulate the activity of CAT. Additionally, neither of the training protocols was able to prevent the reduction in GSH-Px activity observed in the HFD-UNT rats, in agreement with the finding that neither of the protocols were able to prevent the reduction in total sulfhydryl content, since glutathione (GSH) is a cofactor for this enzyme.
The HIIT protocol increased the activity of SOD in erythrocytes, signifying greater removal of superoxide and the formation of H2O2, which could be detoxified by the increasing CAT activity in both AETPs (MICT and HIIT), thereby preventing the formation of harmful hydroxyl radicals. The increase in CAT activity observed using the two training protocols may have occurred as a consequence to the reduction in GSH-Px activity, since neither of the AETPs were able to prevent this alteration.
Since oxidative stress results from an imbalance between the total antioxidant defense of the tissue and the reactive species generated, our present data strongly indicate that obesity provokes oxidative stress in the blood of rats, which induces oxidation of lipids and proteins and changes in CAT and GSH-Px activities. Our data also indicate that exercise influences these parameters in a positive way, minimizing oxidative stress and modulating antioxidant activity. Corroborating our results, Bogdanis [36] reported an increase in CAT activity in subjects following a 3-week HIIT protocol. Shing [37] reported increased resting plasma total antioxidant status concentration and reduced post-exercise increases in plasma MDA concentrations after 3 days of high-intensity exercise in eight highly trained male cyclists. Furthermore, Vezzoli [38] found amelioration in the oxidative stress biomarkers after MICT and HIIT protocols, concluding that both methods produced similar beneficial effects on redox homeostasis in master runners.
Furthermore, this study also showed that HFD increased protein carbonyl content, but did not alter TBA-RS, total sulfhydryl content and the activity of SOD in the HFD-UNT group, when compared to the ND-UNT group in the liver of rats. In addition, both the AETPs (MICT and HIIT) prevented an increase in protein carbonyl content, but did not alter the other parameters analyzed.
In relation to CAT and GSH-Px activities, HFD increased CAT and decreased GSH-Px activities in the HFD-UNT group, when compared to the ND-UNT group. The increase in CAT activity probably occurred to compensate the reduction in GSH-Px activity caused by the HFD, with the intention of preventing an increase in the levels of H2O2 and hydroxyl radical formation (which can cause damage to proteins, lipids and DNA). The MICT protocol (HFD- MICT) prevented the alteration in CAT activity, when compared to the HFD-UNT group, while the HIIT protocol did not prevent this alteration. Furthermore, neither of the AETPs (MICT and HIIT) prevented the reduction in GSH-Px activity in the liver of obese rats.
Our results corroborate those of Pimenta [39], who showed that while HFD exacerbates the deleterious effects of free radicals, the HIIT protocol can increase immune defenses, causing increased resistance to infectious diseases. Steinbacher [40] suggested that the performance of aerobic exercise training increases antioxidant defenses which, in association with morphological and functional adaptations, can provide the organism with protection against oxidative stress.
In conclusion, our study showed that HFD elicits oxidative stress in the blood and liver of rats. Moreover, both AETPs (MICT and HIIT) were able to prevent most of the alterations caused by HFD with regard to the oxidative stress parameters tested, inducing similar beneficial effects on redox homeostasis. However, further studies are necessary to evaluate which configuration of physical exercise session (type, duration, intensity) could be useful as a potential adjuvant for the prevention of obesity and the oxidative stress induced by obesity in humans.
Acknowledgements
This work was supported by grants from Universidade da Região de Joinville.
Compliance with ethical standards
Conflict of interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
Ethical statement
This study was conducted in accordance with the national and institutional guidelines for the care and use of animals.
References
- 1.Rao KR, Lal N, Giridharan NV. Genetic & epigenetic approach to human obesity. Indian J Med Res. 2014;140(5):589–603. [PMC free article] [PubMed] [Google Scholar]
- 2.González-Muniesa P, Mártinez-González M-A, Hu FB, Després J-P, Matsuzawa Y, Loos RJF, et al. Obesity. Nat Rev Dis Prim. 2017;3:17034. doi: 10.1038/nrdp.2017.34. [DOI] [PubMed] [Google Scholar]
- 3.Oliveira SA, Jr, Pai-Silva MD, Martinez PF, Lima-Leopoldo AP, Campos DHS, Leopoldo AS, Okoshi MP, Okoshi K, Padovani CR, Cicogna AC, Oliveira Junior SA, dal Pai-Silva M. Diet-induced obesity causes metabolic, endocrine and cardiac alterations in spontaneously hypertensive rats. Med Sci Monit. 2010;16(12):367–373. [PubMed] [Google Scholar]
- 4.Gómez-Cabello A. Sitting time increases the overweight and obesity risk independently of walking time in elderly people from Spain. Maturitas. 2012;73(4):337–343. doi: 10.1016/j.maturitas.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 5.Donaldson L, Rutter P (2017) Healthier, fairer, safer: the global health journey, 2007-2017. World Health Organization, Geneva. Licence: CC BY-NC- SA 3.0 IGO
- 6.Qatanani M, Lazar MA. Mechanisms of obesity-associated insulin resistance: many choices on the menu. Genes Dev. 2007;21:1443–1455. doi: 10.1101/gad.1550907. [DOI] [PubMed] [Google Scholar]
- 7.Vegiopoulos A, Rohm M, Herzig S. Adipose tissue: between the extremes. EMBO J. 2017;36(14):1999–2017. doi: 10.15252/embj.201696206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sassi F. Obesity and the economics of prevention. Fit not fat. Paris: OECD; 2010. pp. 24–44. [Google Scholar]
- 9.Kehrer JP, Klotz LO. Free radicals and related reactive species as mediators of tissue injury and disease: implications for health. Crit Rev Toxicol. 2015;45(9):765–798. doi: 10.3109/10408444.2015.1074159. [DOI] [PubMed] [Google Scholar]
- 10.Harman D. The free radical theory of aging. Antioxid Redox Signal. 2003;5(5):557–561. doi: 10.1089/152308603770310202. [DOI] [PubMed] [Google Scholar]
- 11.Brinkmann C, Brixius K. Peroxiredoxins and sports: new insights on the antioxidative defense. J Physiol Sci. 2013;63(1):1–5. doi: 10.1007/s12576-012-0237-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Somani SM, Ravi R, Rybak LP. Effect of exercise training on antioxidant system in brain regions of rat. Pharmacol Biochem Behav. 1995;50(4):635–639. doi: 10.1016/0091-3057(94)00357-2. [DOI] [PubMed] [Google Scholar]
- 13.Polidori MC, Mecocci P, Cherubini A, Senin U. Physical activity and oxidative stress during aging. Int J Sports Med. 2000;21:154–157. doi: 10.1055/s-2000-8881. [DOI] [PubMed] [Google Scholar]
- 14.Sugama K, Suzuki K, Yoshitani K, Shiraishi K, Miura S, Yoshioka H, Mori Y, Kometani T. Changes of thioredoxin, oxidative stress markers, inflammation and muscle/renal damage following intensive endurance exercise. Exerc Immunol Rev. 2015;21:130–142. [PubMed] [Google Scholar]
- 15.Ferreira JCB, Rolim NPL, Batholomeu JB, Gobatto CA, Kokubun E, Brum PC. Maximal lactate steady state in running mice: effects of exercise training. Clin Exp Pharmacol Physiol. 2007;34(8):760–765. doi: 10.1111/j.1440-1681.2007.04635.x. [DOI] [PubMed] [Google Scholar]
- 16.Haram PM, Kemi OJ, Lee SJ, Bendheim MO, Al-Share QY, Waldum HL, Gilligan LJ, Kock LG, Britton SL, Najjar SM, Wisloff U. Aerobic interval training vs. continuous moderate exercise in the metabolic syndrome of rats artificially selected for low aerobic capacity. Cardiovasc Res. 2008;81:723–732. doi: 10.1093/cvr/cvn332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ferreira AGK, Cunha AA, Machado FR, Pederzolli CD, Dalazen GR, Assis AM, Lamers ML, Santos MF, Dutra-Filho CS, Wyse ATS. Experimental hyperprolinemia induces mild oxidative stress, metabolic changes, and tissue adaptation in rat liver. J Cel Biochem. 2012;113(1):174–183. doi: 10.1002/jcb.23342. [DOI] [PubMed] [Google Scholar]
- 18.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem. 1979;95:351–358. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 19.Aksenov MY, Markesbery WR. Changes in thiol content and expression of glutathione redox system genes in the hippocampus and cerebellum in Alzheimer’s disease. Neurosci Lett. 2001;302:141–145. doi: 10.1016/S0304-3940(01)01636-6. [DOI] [PubMed] [Google Scholar]
- 20.Reznick AZ, Packer L. Oxidative damage to proteins: spectrophotometric method for carbonyl assay. Methods Enzymol. 1994;233:357–363. doi: 10.1016/S0076-6879(94)33041-7. [DOI] [PubMed] [Google Scholar]
- 21.Aebi H. Catalase in vitro. Methods Enzymol. 1984;105:121–126. doi: 10.1016/S0076-6879(84)05016-3. [DOI] [PubMed] [Google Scholar]
- 22.Wendel A. Glutathione peroxidase. Methods Enzymol. 1981;77:325–333. doi: 10.1016/S0076-6879(81)77046-0. [DOI] [PubMed] [Google Scholar]
- 23.Marklund S. Pyrogallol autooxidation. Handbook of methods for oxygen radical. Boca Raton: CRC Press; 1985. pp. 243–247. [Google Scholar]
- 24.Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the folin phenol reagent. J Biol Chem. 1951;193:265–275. [PubMed] [Google Scholar]
- 25.Tomao F, Papa A, Zaccarelli E, Rossi L, Caruso D, Minozzi M, Vici P, Frati L, Tomao S. Triple-negative breast cancer: new perspectives for targeted therapies. Onco Targets Ther. 2015;8:177–193. doi: 10.2147/OTT.S67673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Huang CJ, McAllister MJ, Slusher AL, Webb HE, Mock JT, Acevedo EO. Obesity-related oxidative stress: the impact of physical activity and diet manipulation. Sports Med Open. 2015;1:32. doi: 10.1186/s40798-015-0031-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Higa TS. Comparison between cafeteria and high-fat diets in the induction of metabolic dysfunction in mice. Int J Physiol Pathophysiol Pharmacol. 2014;6(1):47–54. [PMC free article] [PubMed] [Google Scholar]
- 28.Yida Z, Imam MU, Ismail M, Ismail N, Ideris A, Abdullah MA. High fat diet-induced inflammation and oxidative stress are attenuated by N-acetylneuraminic acid in rats. J Biomed Sci. 2015;22(1):96. doi: 10.1186/s12929-015-0211-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Charradi K, Elkahoui S, Limam F, Aouani E. High-fat diet induced an oxidative stress in white adipose tissue and disturbed plasma transition metals in rat: prevention by grape seed and skin extract. J Physiol Sci. 2013;63(6):445–455. doi: 10.1007/s12576-013-0283-6. [DOI] [PubMed] [Google Scholar]
- 30.Vassalle C, Pingitore A, De Giuseppe R et al (2015) Biomarkers part II: biomarkers to estimate bioefficacy of dietary/supplemental antioxidants in sport. In: Lamprecht M (ed) Antioxidants in sport nutrition, chap 16. CRC Press/Taylor & Francis, Boca Raton, FL [PubMed]
- 31.Pingitore A, Lima GPP, Mastorci F, Quinones A, Iervasi G, Vassalle C. Exercise and oxidative stress: potential effects of antioxidant dietary strategies in sports. Nutrition. 2015;31(7–8):916–922. doi: 10.1016/j.nut.2015.02.005. [DOI] [PubMed] [Google Scholar]
- 32.Aro CEP, Guzmán JAR, Muñoz MES, González BEV. Effects of high intensity interval training versus moderate intensity continuous training on the reduction of oxidative stress in type 2 diabetic adult patients: CAT. Medwave. 2015;15(7):1–13. doi: 10.5867/medwave.2015.07.6212. [DOI] [PubMed] [Google Scholar]
- 33.Halliwell B, Whiteman M. Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean? Br J Pharmacol. 2004;142:231–255. doi: 10.1038/sj.bjp.0705776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Halliwell B. Cell culture, oxidative stress, and antioxidants: avoiding pitfalls. Biomed J. 2014;37(3):99–105. doi: 10.4103/2319-4170.128725. [DOI] [PubMed] [Google Scholar]
- 35.Young IS, Woodside JV. Antioxidants in health and disease. J Clin Pathol. 2001;54:176–186. doi: 10.1136/jcp.54.3.176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bogdanis GC, Stavrinou P, Fatouros IG, Philippou A, Chatzinikolaou A, Draganidis D, Ermidis G, Maridaki M. Short-term high-intensity interval exercise training attenuates oxidative stress responses and improves antioxidant status in healthy humans. Food Chem Toxicol. 2013;61:171–177. doi: 10.1016/j.fct.2013.05.046. [DOI] [PubMed] [Google Scholar]
- 37.Shing CM, Peake JM, Ahern SM, Strobel NA, Wilson G, Jenkins DG, Coombes JS. The effect of consecutive days of exercise on markers of oxidative stress. Appl Physiol Nutr Metab. 2007;32:677–685. doi: 10.1139/H07-051. [DOI] [PubMed] [Google Scholar]
- 38.Vezzoli A, Pugliese L, Marzorati M, Serpiello FR, La Torre A, Porcelli S. Time-course changes of oxidative stress response to high-intensity discontinuous training versus moderate-intensity continuous training in master runners. PLoS One. 2014;9(1):1–9. doi: 10.1371/journal.pone.0087506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pimenta M, Bringhentii S, Mello VS, Mendes IKS, Aguila MB, Lacerda CAM. High-intensity interval training beneficial effects on body mass, blood pressure, and oxidative stress in diet-induced obesity in ovariectomized mice. Life Sci. 2015;139:75–78. doi: 10.1016/j.lfs.2015.08.004. [DOI] [PubMed] [Google Scholar]
- 40.Steinbacher P, Eckl P. Impact of oxidative stress on exercising skeletal muscle. Biomolecules. 2015;5(2):356–377. doi: 10.3390/biom5020356. [DOI] [PMC free article] [PubMed] [Google Scholar]