

Abstract
Reaction time variability (RTV), reflecting fluctuations in response time on cognitive tasks, has been proposed as an endophenotype for many neuropsychiatric disorders. There have been no large-scale genome-wide association studies (GWAS) of RTV and little is known about its genetic underpinnings. Here, we used data from the UK Biobank to conduct a GWAS of RTV in participants of white British ancestry (n = 404,302) as well as a trans-ancestry GWAS meta-analysis (n = 44,873) to assess replication. We found 161 genome-wide significant single nucleotide polymorphisms (SNPs) distributed across 7 genomic loci in our discovery GWAS. Functional annotation of the variants implicated genes involved in synaptic function and neural development. The SNP-based heritability (h2SNP) estimate for RTV was 3%. We investigated genetic correlations between RTV and selected neuropsychological traits using linkage disequilibrium score regression, and found significant correlations with several traits, including a positive correlation with mean reaction time and schizophrenia. Despite the high genetic correlation between RTV and mean reaction time, we demonstrate distinctions in the genetic underpinnings of these traits. Lastly, we assessed the predictive ability of a polygenic score (PGS) for RTV, calculated using PRSice and PRS-CS, and found that the RTV-PGS significantly predicted RTV in independent cohorts, but that the generalisability to other ancestry groups was poor. These results identify genetic underpinnings of RTV, and support the use of RTV as an endophenotype for neurological and psychiatric disorders.
Subject terms: Genetics, Neuroscience, Psychology
Introduction
Elevated intra-individual variability in reaction time, namely increased trial-by-trial fluctuations in response time on cognitive tasks, has been associated with neurodevelopmental and neurodegenerative disorders [1]. Increased variability in reaction time is thought to reflect disruptions in attentional control and executive function and it has been associated with abnormalities in brain structure and function [1–4]. Increased reaction time variability (RTV) has been demonstrated in attention-deficit hyperactivity disorder (ADHD) [5, 6], schizophrenia [4, 7], bipolar disorder [8], and major neurocognitive disorders [9]. The heritability of RTV has been established in twin and family studies (h2 = 0.28–0.5) [10, 11] and consequently, RTV has been proposed as an endophenotype for some of these disorders.
Measures of RTV and measures of central tendency, such as mean reaction time, are known to be correlated however, it has been suggested that RTV may provide insight into cognitive function over and above mean performance metrics [1, 12, 13]. Abnormalities in RTV have been detected in people classified as at risk mental state for psychosis when mean reaction times are normal [12]. In studies of age-related changes in cognitive performance, RTV has been a better predictor of subsequent cognitive decline than measures of central tendency [14, 15]. Additionally, a systematic review of longitudinal changes in RTV found that the association between elevated RTV at baseline and accelerated cognitive decline, mild neurocognitive disorders, dementia, and mortality remained after controlling for mean reaction time [9]. Thus, RTV may offer unique predictive power beyond the mean and may be useful in detecting early neuropathological changes prior to the onset of more severe cognitive dysfunction. Given the proposed utility of measures of RTV, a better understanding of its neurobiological underpinnings is required.
Despite increasing interest in the biological basis of RTV, its genetic architecture remains poorly understood. A limited number of candidate gene studies have provided evidence for an association of RTV with catecholamine system genes [16–18]. However, candidate gene studies have largely failed to identify replicable genes associated with behavioural traits, including RTV, and there has been a shift towards genome-wide association studies (GWAS) to identify genotype-phenotype associations using a hypothesis-free approach [19]. There has only been one GWAS of RTV to date (n = 857), which identified one genome-wide significant SNP, rs62182100 [20]. The significant SNP is an intronic variant located within the HDAC4 gene, which plays a role in transcriptional regulation and has been implicated in synaptic plasticity, learning and memory [21]. However, due to the small sample size and the lack of independent replication in this GWAS, insight into the genetic underpinnings of reaction time variability remains limited. A GWAS with a larger sample size may facilitate the identification of more significant loci and provide the power needed for a more comprehensive investigation of the genetic architecture of this trait.
Here, we conduct the largest GWAS of RTV to date with a sample size of 404,302 individuals using data from the UK Biobank. We aim to identify common genetic variants and genes associated with RTV, and we calculate the first SNP-based heritability (h2SNP) estimate for the trait. We also calculate estimates of genetic correlations with neurodevelopmental disorders and other phenotypes that have been previously associated with RTV. Lastly, we test the external validity of our results by performing polygenic prediction of RTV in independent samples of European and African ancestries.
Materials and methods
Participants and phenotype definition
This study used data from the UK Biobank (UKB), obtained under accession number 27412. The UKB is a large-scale biomedical database with genotype and phenotype data for approximately 500,000 individuals [22]. At baseline, a brief cognitive assessment, including a custom-made reaction time test, was administered to participants (aged 40–70 years) as part of the fully automated questionnaire. The UKB reaction time test is based on a Go/NoGo test and is designed to measure processing speed [23]. Participants were shown 2 cards with symbols on them and asked to push a button as quickly as possible when the symbols on the card matched. The test consisted of 12 trials, 9 of which contained matching cards. The UKB reaction time test has demonstrated good internal consistency (Cronbach α = 0.85) [24], moderate test-retest reliability (Pearson r12 = 0.55) [23], and good concurrent validity with well-validated tests of reaction time [23]. In the discovery and replication GWAS, RTV was operationalized as the intra-individual standard deviation (ISD) of reaction times across correct trials. Prior to calculating the ISD, trials with a reaction time <50 ms (suggesting anticipation instead of reaction), and >200 ms (indicating a response after the cards had disappeared) were excluded. ISD scores were calculated for participants with ≥3 correct trials. As RTV was non-normally distributed, RTV values were rank-based inverse normal transformed. Since longer reaction times may result in an increased ISD for an individual [25], we also calculated the intra-individual coefficient of variation (ICV) for reaction times for all participants. The ICV is calculated by dividing the ISD by the mean reaction time for an individual. For the discovery dataset, we included 405,022 individuals with “white British” ancestry (54% females; mean age 56.88 years), classified according to self-declared ethnicity and genetic principal component analysis. We used all other ancestry groups from the UKB for replication analysis - this included participants who completed the UKB reaction time test with a self-reported ethnicity of “white non-British” (n = 28,600), “Asian or Asian British” (n = 8904), or “Black or Black British” (n = 7415), totalling to 44 919 individuals (55% female, mean age 54.27 years) for inclusion in the replication GWAS.
Genome-wide association analysis
GWAS was conducted using version 3 of the UKB genetic data. Genotyping, imputation, and central quality control procedures for the UKB genotypes are described in detail elsewhere [26]. The REGENIE method was used and involves 2 steps. In step 1, polygenic predictors are constructed by fitting a whole genome regression model to the UKB genotype data. Additional quality control filters were applied to the UKB genotype calls using PLINK 2.0 [27] prior to conducting step 1 of REGENIE. Quality control steps included removing: (1) individuals with >10% missing genotype data, (2) SNPs with >10% genotype missingness, (3) SNPs failing the Hard-Weinberg equilibrium tests at p = 1 × 10−15, and (4) SNPs with a minor allele frequency (MAF) < 1% or minor allele count (MAC) < 50. After quality control, 582,052 variants and 405,019 samples were included in step 1 of REGENIE. In step 2 of REGENIE, a linear regression model was used to test for phenotype-genotype associations using imputed UKB genotype data, conditional upon the predictions of the model from step 1. The association model in step 2 included age, sex and the first 10 genetic principal components as covariates. Variants with an INFO score <0.8 and MAC < 20 were excluded in step 2 leaving 19,963,755 SNPs and 404,302 samples for inclusion in the GWAS.
Replication cohort and meta-analysis
We sought to replicate the lead SNPs from the discovery GWAS in an independent association analysis. First, we used REGENIE to conduct association analysis within all other ancestry groups (“White non-British”, “Asian or Asian British”, and “Black or Black British”) from the UKB separately. Quality control procedures were identical to those used for the discovery analysis. Following GWAS, the summary statistics for 28 396 731 SNPs (n = 44,873 after quality control) were meta-analysed using an inverse variance based approach implemented in METAL [28]. To assess for replication, we determined whether lead SNPs from the discovery GWAS reached significance in the replication GWAS (α = 0.05/7; p < 0.0071). Additionally, we examined if the effect directions of the A1 allele of lead SNPs from the discovery GWAS were concordant across the discovery and replication GWAS. A binomial test was performed using R v4.1.0 [29] to assess for an excess or deficit of concordant SNPs than would be expected by chance. Lastly, we used METAL to conduct an inverse variance-weighted meta-analysis of the discovery and replication GWAS.
Genomic risk loci characterisation
Genomic risk loci for RTV were characterised from the GWAS results using Functional Mapping and Annotation of Genome-Wide Association Studies (FUMA) [30]. First, the SNP2GENE function was used to identify independent significant SNPs, defined as SNPs with a p-value ≤ 5 × 10−8 and independent of other genome-wide significant SNPs at r2 < 0.6. The correlation estimates were calculated using the 1000 Genomes Project Phase 3 release European reference panel [31]. A genomic risk locus included all SNPs, including those from the reference panel, that were in linkage disequilibrium of r2 ≥ 0.6 with an independent significant SNP. Genomic risk loci that were within 250 kilobases (kb) of each other were merged into one locus. Lead SNPs were defined as independent significant SNPs that were independent of each other at r2 < 0.1. Regional visualisation plots were produced using LocusZoom [32].
Functional mapping and annotation
The independent significant SNPs and SNPs in LD (r2 > 0.6) with the independent significant SNPs (henceforth referred to as candidate SNPs) were functionally annotated using ANNOVAR [33], combined dependent depletion (CADD) [34], RegulomeDB (RDB) [35], and 15-core chromatin states [36]. The NHGRI-EBI GWAS catalogue was searched to assess for previous associations of the candidate SNPs. eQTL mapping for significant SNP-gene pairs (FDR q < 0.05) was performed using GTEx v8 whole blood and brain tissue (http://www.gtexportal.org/home/datasets), RNAseq data from the CommonMind Consortium [37], and the BRAINEAC database (http://www.braineac.org/).
Identified lead SNPs were mapped to likely target genes using The OpenTargets Variant-to-Gene pipeline which integrates a positional score (based on distance to the canonical transcription start site) with data from quantitative trait loci and chromatin interaction experiments and in silico functional predictions [38, 39]. For each lead SNP, we also report the nearest gene identified through positional mapping using FUMA. Gene-based analysis of 19,129 protein coding genes was performed using MAGMA [40] as implemented in FUMA, with an SNP-wise mean model and the 1000 genomes project phase 3 release European reference panel. To control for multiple testing, a Bonferroni corrected p-value was used (α = 0.05/19 129 genes tested; p < 2.61 × 10−6). Additionally, gene-set enrichment analysis was conducted using: (1) significant genes from MAGMA gene-based analysis, (2) genes identified through the OpentTargets’ Variant-to-Gene pipeline, and (3) genes identified through positional mapping in FUMA. Hypergeometric tests were applied through the GENE2FUNC function in FUMA to assess if the identified genes were over-represented in 15,496 gene sets obtained from MsigDB v7.0 [41]. Bonferroni correction for multiple testing was applied and gene sets with p < 3.23 × 10−6 were considered significant.
Heritability, polygenicity, and discoverability
We used univariate GCTA-GREML analysis [42] and MiXeR [43] to estimate the proportion of variance explained by common genetic factors, i.e. h2SNP. The covariates included in the GCTA-GREML analysis were the same as those included in the GWAS. The proportion of causal variants (polygenicity) and the average explained variance per causal variant (discoverability) were estimated using MiXeR v1.2 [43]. The univariate mixture model considers MAF, sample size, LD structure, and genomic inflation to derive estimates of heritability, polygenicity, and discoverability using maximum likelihood estimation.
Genetic correlation and phenotypic associations
Genetic correlations between RTV and phenotypes known to be associated with RTV were calculated using linkage disequilibrium score regression (LDSC) [44, 45]. Summary statistics for general cognitive ability (GCA), educational attainment, Alzheimer’s disease, post-traumatic stress disorder (PTSD), ADHD, schizophrenia, neuroticism, intracranial volume, cortical surface area, cortical thickness, and 7 subcortical brain volumes (nucleus accumbens, amygdala, brainstem, caudate nucleus, pallidum, putamen, and thalamus) were used to calculate genetic correlation estimates. Supplementary Table 1 provides further details on the sources of the GWAS summary statistics. Using data from the UKB, the relationships between RTV and the same phenotypes as listed above were assessed using linear regression (Supplementary Note; Supplementary Table 2).
Comparison with other measures of RTV and mean reaction time
First, we conducted a GWAS of ICV, an alternative measure of RTV, using the UKB reaction time test and the same participants and analysis pipeline as the discovery RTV-GWAS (n = 404,302). Next, we assessed the significance and effect direction of lead SNPs from the discovery RTV-GWAS in the ICV-GWAS as well as a publicly available GWAS of mean reaction time in the UKB [46]. We estimated the genetic correlation between RTV (measured by ISD), mean reaction time, and ICV using LDSC. Lastly, we estimated the genetic correlation between mean reaction time and the same 17 phenotypes from the genetic correlation analyses with RTV (Supplementary Table 1). We tested whether the genetic correlations' estimates for the 17 traits were different for RTV compared to mean reaction time (Supplementary Note).
Polygenic score validation
For polygenic score validation we used controls from two independent cohorts of European and African ancestry, The Thematically Organised Psychosis (TOP) Study [47] and The Genomics of Schizophrenia in the South African Xhosa People (SAX) Study [48], respectively. RTV on a continuous performance test was calculated for 182 healthy controls from the TOP study and 563 controls (people without psychotic disorders) from the SAX study. Additional information on the TOP and SAX study can be found in Supplementary Note 1. We also assessed the predictive ability of an RTV polygenic score (PGS) for RTV in the “White non-British”, “Asian or Asian British”, and “Black or Black British” ancestry groups from the UKB.
The RTV-PGS were calculated from Z-score effect size estimates from the discovery of RTV-GWAS using a pruning and thresholding approach implemented in PRSice [49]. Prior to PGS calculation, SNPs with MAF < 0.05 were excluded and pruning was performed using an r2 < 0.1 within a 250 kb window. We calculated PGS across 10 p-value thresholds (1, 0.1, 0.05, 0.01, 1 × 10−3, 1 × 10−4, 1 × 10−5, 1 × 10−6, 1 × 10−7, 5 × 10−8) in the white non-British participants from the UKB and linear regression models were used to test the association between RTV and PGS at each threshold. The best performing PGS was used to determine the p-value threshold for PGS calculation in all other ancestry groups. Sex, age and the first ten principal components were included covariates. For comparison, we calculated an RTV-PGS in each target cohort using PRS-CS [50], which uses a Bayesian regression framework and places a continuous shrinkage prior on SNP effect sizes. The 1000 Genomes Phase 3 release European sample [31] was used as the LD reference panel for PRS-CS. The Bonferroni correction was applied to account for multiple testing (α = 0.05/19 polygenic scores; p < 2.63 × 10−3).
Results
Genome-wide associations
Genome-wide association tests for RTV in the discovery analysis identified 161 genome-wide significant SNPs (p < 5 × 10−8) (Fig. 1; Supplementary Table 3). There were 13 independent significant SNPs distributed across 7 genomic loci (Table 1). Regional visualisation plots for the significant loci are depicted in Fig. 2 and Supplementary Fig. 3. Four of the seven genome-wide significant loci have been reported as significant in previous GWAS of general cognitive ability and intelligence (Supplementary Table 4). The linkage disequilibrium score regression intercept was 1 (SE = 0.01), consistent with minimal inflation of the test statistic due to population stratification.
Fig. 1. Manhattan plot of discovery GWAS for RTV in the UK Biobank.

Manhattan plot for the observed -log10 p-values for an association with RTV in the discovery GWAS. The dotted line indicates a genome-wide significance threshold of 5 × 10-8. The lead SNPs from the GWAS are outlined in black and the candidate SNPs are shown in bold.
Table 1.
Genome-wide significant loci for the discovery GWAS of RTV in 404,302 individuals.
| Genomic risk locus | Lead variant | Chr | Position | A1/A2 | Discovery GWAS | Replication GWAS | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 Freq | Beta | P-value | Top assigned gene | Nearest gene | Beta | P-value | |||||
| 1 | rs183204237 | 2 | 162188514 | C/G | 9.79 × 10–4 | −0.21 | 2.34 × 10–8 | PSMD14 | PSMD14 | −0.088 | 0.46 |
| 2 | rs35206686 | 4 | 152762934 | T/TC | 0.736 | −0.015 | 6.09 × 10–9 | FHIP1A | RP11-424M21 | −0.004 | 0.615 |
| 3 | rs17167210 | 7 | 133339343 | A/G | 0.435 | 0.013 | 1.5 × 10–9 | EXOC4 | EXOC4 | −0.01 | 0.141 |
| 4 | rs35859241 | 15 | 51736660 | G/A | 0.254 | −0.014 | 3.69 × 10–8 | SCG3 | RP11-707P17.1 | −0.001 | 0.864 |
| 5 | rs912243472 | 15 | 74096786 | G/GA | 0.135 | 0.018 | 1.97 × 10–8 | TBC1D21 | INSYN1 | 0.005 | 0.575 |
| 6 | rs1863115 | 17 | 44625928 | A/C | 0.74 | 0.017 | 2.47 × 10-10 | LRRC37A2 | LRRC37A2 | 0.003 | 0.755 |
| 7 | rs11697176 | 20 | 3831629 | T/C | 0.112 | −0.021 | 1.9 × 10–8 | PANK2 | MAVS | 0.008 | 0.58 |
Fig. 2. Regional association plots for three genome-wide significant loci in the discovery RTV-GWAS.

Regional plots for rs17167210 (A), rs35859241 (B) and rs1863115 (C). The dotted line denotes a genome-wide significance threshold of 5 × 10–8. SNPs in the genomic risk loci are colour-coded as a function of their linkage disequilibrium r2 to the lead SNP in the region.
None of the lead SNPs from the discovery GWAS reached significance in the replication GWAS (Table 1; Supplementary Fig. 1). Due to the limited number of lead SNPs at a genome-wide significant threshold, binomial tests for concordance were performed using lead SNPs from the discovery GWAS at a suggestive threshold of p of ≤5 × 10−5. There were 261 lead SNPs at the suggestive level in the discovery GWAS, with 156 of them having concordant direction of effects (binomial test p = 0.047) in the replication GWAS.
In the meta-analysis of the discovery and replication GWAS (n = 449,175), there were 41 genome-wide significant SNPs (p < 5 × 10−8) distributed across 6 genomic loci (Supplementary Fig. 1; Supplementary Table 5). Thirty-six of the genome-wide significant SNPs were also significant in the discovery GWAS. Inspection of the quantile-quantile plot for the meta-analysis shows greater test-statistic inflation above the null for moderately significant p-values than in the discovery GWAS (Supplementary Fig. 2). The linkage disequilibrium score regression intercept for the meta-analysis was 1 (SE = 0.01), suggesting that the inflation of the test statistic reflects true associations with RTV.
Integration with functional genomic data
Each lead SNP from the discovery GWAS was mapped to one gene using the OpenTargets Variant-to-Gene pipeline, resulting in 7 mapped genes (Table 1). MAGMA gene-based analysis identified 5 genes significantly associated with RTV: EXOC4 (p = 6.3 × 10−7), TBC1D21 (p = 2.57 × 10−6), CNTNAP4 (p = 2.59 × 10−7), LRRC37A (p = 9.81 × 10−10), and NSF (p = 4.97 × 10−7) (Supplementary Table 6). An additional 17 genes were mapped to candidate SNPs from the discovery GWAS using FUMA positional mapping; resulting in a total of 27 input genes (Supplementary Table 7) for gene-set enrichment analysis. Gene-set enrichment analysis did not identify any significant gene sets associated with RTV.
The lead variant for the GWAS, rs1863115 (p = 2.47 × 10−10) (Fig. 2C), is a non-synonymous exonic variant for LRRC37A2 and an intronic variant for ARL17A. The CADD score for rs1863115 is 18.32, suggestive of variant deleteriousness. Based on annotation by the OpenTargets genetic platform, the most likely gene affected by this variant is LRRC37A2, a gene that encodes an integral component of the cellular membrane. LRRC37A2 has been associated with intelligence, and mean reaction time in previous GWAS [46, 51].
There was evidence of functionality for variants in genomic risk loci 3 and 4 (Table 1). The lead variant for locus 3, rs17167210 (p = 1.5 × 10−9), is located in an intron of EXOC4 and is an eQTL for EXOC4 and LRGUK in brain tissue (CommonMind Consortium) (Fig. 2A). A nearby intronic variant, rs11768150 (R2 = 0.88, p = 1.71 × 10−7), has a CADD score of 13.5, suggestive of variant deleterious, and a RegulomeDB score of 3a, indicating that the variant is likely to be involved in gene regulation. The lead variant for locus 4, rs35859241 (p = 3.69 × 10−8), is an eQTL for SCG3 and GLDN in brain tissue (CommonMind Consortium, GTEx Brain). This variant is in LD with rs2606134 (R2 = 0.81, p = 1.14 × 10−4), which is located within the 5′ untranslated region of SCG3 (Fig. 2B). The SNP, rs2606134, has a CADD score of 13.15 and a RegulomeDB score of 2b, suggesting that this variant may be biologically relevant.
Estimating heritability, polygenicity, and discoverability
The h2SNP for RTV was estimated at 0.029 (SE = 0.002) using GCTA-GREML. MiXeR analysis suggested that RTV is highly polygenic with an estimated 6800 causal variants explaining the h2SNP for RTV. As expected for a trait with a low h2SNP and high polygenicity, discoverability was low (σ2β = 5.38 × 10−6, SD = 2.85 × 10−7) indicating that most SNP-associations have a weak effect. Akaike’s Information Criteria (AIC) for MiXeR analysis was 18.39 indicating reliable model fit.
Genetic correlations and phenotypic associations
We assessed the genetic correlations and phenotypic relationships between RTV and 17 traits that have been posited to be associated with RTV using LDSC and linear regression respectively (Fig. 3; Supplementary Tables 8 and 9). After Bonferroni correction, we found significant genetic correlations (α = 0.05/19 traits; p < 2.63 × 10−3) between RTV and general cognitive ability (rg = −0.44, SE = 0.03), educational attainment (rg = −0.23, SE = 0.03), schizophrenia (rg = 0.26, SE = 0.03), and neuroticism (rg = 0.13, SE = 0.03). The analysis of phenotypic data from the UKB revealed a significant relationship between RTV and several traits, including those that showed significant genetic correlations with RTV (Fig. 3, Supplementary Table 9).
Fig. 3. Genetic correlations and phenotypic associations between RTV and 19 selected traits.

A Genetic correlations were calculated with LD score regression using SNP summary statistics from discovery RTV-GWAS and publicly available summary statistics for other traits (Supplementary Table 1). B Associations between RTV and the same 19 traits were calculated using phenotypic data from the UK Biobank (Supplementary Methods). Point estimates for correlations and beta coefficients are shown with 95% confidence intervals. Dark blue dots indicate nominally significant p-values and light blue dots indicate significant p-values after Bonferroni correction.
Comparison with other measures of RTV and mean reaction time
We sought replication of the 7 lead SNPs from the discovery RTV-GWAS in the GWAS of ICV and found that all lead SNPs were significant (α = 0.05/7; p < 0.0071) in the ICV-GWAS (Supplementary Table 10). Similar to the replication analyses described earlier, we used the 261 lead SNPs from the discovery GWAS at a suggestive threshold of p ≤ 5 × 10−5 to conduct binomial tests for concordance. All lead SNPs from the discovery RTV-GWAS had a concordant direction of effect in the ICV-GWAS. We found significant genetic correlations between RTV and ICV (rg = 0.89, SE = 0.01) as well as RTV and mean reaction time (rg = 0.69, SE = 0.02). Consistent with the relatively high genetic correlation between RTV and mean RT, the number of lead SNPs from the RTV-GWAS that showed a concordant direction of effect in the mean reaction time GWAS was greater than expected by chance (binomial test p < 2.2 × 10−16). However, only one of the lead SNPs from the discovery RTV-GWAS reached significance (p < 0.0071) in the GWAS of mean reaction time. Further, we found that 7 (ARL17A, ARL17B, LRRC37A2, NSF, WNT3, TBC1D21, CDC27) of the 27 genes identified in the RTV-GWAS had a documented association with mean reaction time in the GWAS catalogue [52]. Lastly, we found that the genetic correlations between RTV and 17 selected traits and mean reaction time and the same 17 traits were similar for most traits. Notably, we found significant differences in the genetic correlations between RTV and educational attainment, general cognitive ability, and ADHD when compared to the genetic correlations between mean reaction time and the same traits (Supplementary Table 8). We show that ADHD has a nominally significant positive genetic correlation with RTV (rg = 0.1, SE = 0.02, p = 5.1 × 10−3) and a nominally significant negative correlation with mean reaction time (rg = −0.06, SE = 0.03, p = 0.03) (Fig. 3; Supplementary Fig. 4; Supplementary Table 8). The magnitude of genetic correlations was significantly greater for RTV compared to mean reaction time for educational attainment and general cognitive ability (Supplementary Fig. 4; Supplementary Table 8).
Polygenic prediction of RTV
To evaluate the replicability and predictive ability of the results from our discovery GWAS, we calculated a PGS for RTV in the five independent target samples using PRSice and PRS-CS. For the PGS calculated using PRSice, the most significant association between the RTV-PGS and RTV in non-British European participants from the UKB was achieved when all SNPs surviving LD pruning (n = 166,662) were included in the PGS calculation (p-value threshold = 1) (Supplementary Table 11). The variance explained by this PGS was r2 = 0.0027 (p = 6.09 × 10−18). The PRSice RTV-PGS performed poorly in the other ancestry groups from the UKB and there were no significant associations between the PGS and RTV in the South Asian or African ancestry groups (Fig. 4). There was an improvement in predictive power when using PRS-CS to calculate the PGS and there was a significant association between the RTV-PGS and RTV in the non-British European (r2 = 0.0048, p = 3.08 × 10−31) and South Asian ancestry groups (r2 = 0.0012, p = 1.73 × 10−3) from the UKB (Fig. 4). The PRSice and PRS-CS RTV-PGS did not predict RTV in controls from the TOP and SAX study (Supplementary Table 11; Supplementary Fig. 5).
Fig. 4. Bar chart showing the predictive accuracy of the RTV-PGS in three independent cohorts.
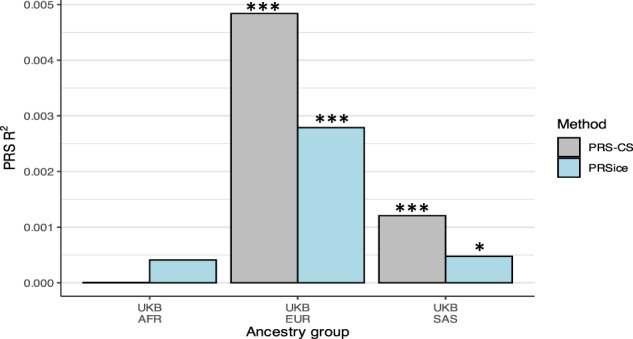
Prediction of RTV by polygenic score (PGS) in the African, non-British European, and South Asian ancestry groups from the UK Biobank. The predictive accuracy of the PGS (R2) was assessed in each cohort for a PGS calculated using two methodologies, PRSice and PRS-CS. PRSice PGS were calculated using all single nucleotide polymorphisms surviving LD pruning from the discovery GWAS (p-value threshold of 1). *p < 0.05, ***p < 2.63 × 10-3.
Discussion
Using UKB data, we have performed the largest GWAS of RTV to date and have made several contributions to our understanding of the genetic basis of this cognitive trait. We identified 161 genome-wide significant SNPs for RTV distributed across 7 genomic loci, all of which are novel for RTV. We identified several genes that may play a role in RTV, many of which have been associated with cognitive traits previously. We provide the first SNP-based heritability estimate for RTV, and the first estimates for genetic correlations between RTV and several neuropsychiatric traits. We demonstrate that RTV-PGS derived from the discovery GWAS can significantly predict RTV in an independent cohort, but that the predictive ability declines if the discovery and target populations are of different ancestries.
The genes identified by the GWAS may provide insight into the biological underpinnings of RTV. Although the exact role that many of the identified genes may play in RTV is unclear, several are worthy of further investigation. For example, two of the significant genes, contactin associated protein family member 4 (CNTNAP4) and N-ethylmaleimide sensitive factor, vesicle fusing ATPase (NSF) encode proteins that play a role in synaptic function. CNTNAP4 is involved in the synaptic transmission of dopamine and GABA [53] and NSF regulates glutamate receptor binding activity [54, 55]. Alterations in dopaminergic, glutaminergic and GABAergic activity have been associated with RTV [1, 56–58] and thus, further exploration of the association between RTV and CNTNAP4 and NSF may be warranted. Variants in EXOC4 and SCG3 showed evidence of regulatory functionality and variant deleteriousness. Both genes are highly expressed in the brain and have been associated with cognitive traits in previous GWAS [51, 59]. EXOC4 encodes a component of the exocyst complex which plays a role in multiple physiological processes, including neuronal development [60, 61]. SCG3 encodes a member of the granin family of neuroendocrine secretory proteins and is involved in secretory granule biosynthesis and the storage and transport of neurotransmitters [62, 63]. Another identified gene, inhibitory synaptic factor 1 (INSYN1), is involved in post-synaptic inhibition in the central nervous system [64] and may be considered for further study. INSYN1 is a novel association for a cognitive trait but it has been associated with psychiatric disorders, including ADHD [65], PTSD [66], and Tourette syndrome [67]. Many of the identified genes play a role in neural development and synaptic functioning, suggesting an important role for these processes in the biology of reaction time variability.
We reported an h2SNP of 3%, high polygenicity, and low discoverability for RTV. It is possible that the high polygenicity, despite the relatively low heritability, may be explained by the range of exogenous factors, such as age, sex, handedness, and treatment effects [5, 68, 69], that influence RTV. We hypothesise that a large proportion of the identified 6800 causal variants may be associated with these exogenous factors and thus, only have indirect and weak effects on RTV.
We found that a PGS, derived from the RTV-GWAS in white British participants from the UKB, was significantly predictive of RTV in the UKB white non-British participants, explaining 0.5% of the variance in the measure, which is expected with a h2SNP of 3% [70]. The predictive accuracy of the PGS was substantially lower in non-European ancestry populations. This is in keeping with prior work on the generalisability of PGS across ancestrally diverse populations with the predictive accuracy of the PGS decreasing as the genetic distance between the discovery and target populations increases [71]. These results further emphasise the need to increase the representation of ancestrally diverse populations in genomic studies.
We found significant positive genetic correlations between RTV and schizophrenia, and neuroticism. The result for schizophrenia is consistent with previous findings of increased RTV in people with schizophrenia [4, 7]. It is hypothesised that the elevations in RTV reflect cognitive control deficits that occur in the disorder [4]. The positive genetic correlation between RTV and neuroticism is supported by our phenotype analysis, which demonstrate a positive relationship between the two phenotypes. To our knowledge, the association between these two traits has not been studied and future research is needed to explore the mechanisms that contribute to a relationship between RTV and neuroticism. There were significant negative genetic correlations between educational attainment, and general cognitive ability. This result is in keeping with the negative relationship between these two traits and RTV on the phenotypic level reported in previous literature [72, 73].
As our primary measure of RTV (ISD) is often highly correlated with mean reaction time [25], we conducted an additional GWAS of another measure of RTV, the ICV. ICV is the ratio of a participant’s ISD to their mean reaction time and provides a certain degree of control for mean reaction time. We found the genetic basis of both measures of RTV, ISD and ICV, to be similar. All lead SNPs from the discovery RTV-GWAS reached significance in the ICV-GWAS and showed a consistent direction of effect. The significant high genetic correlation between ISD and ICV provides additional support for consistency between the common genetic determinants of both measures of RTV. Consistent with the strong phenotypic association between RTV and mean reaction time, we found evidence of similarities in the common genetic determinants of the two traits. However, we also demonstrate differences in the genetic basis of RTV and mean reaction time and show that most of the lead SNPs and identified genes from the RTV-GWAS are not associated with mean reaction time. Additionally, the results from our genetic correlation analyses show that while the patterns of correlation with the 17 selected phenotypes are similar for RTV and mean reaction time, there are significant differences in the magnitude and direction of correlation for certain phenotypes (e.g. educational attainment, general cognitive ability and ADHD). These analyses demonstrate distinctions in the common genetic variants associated with RTV and mean reaction time and provide support for our approach of studying RTV separately to mean reaction time.
There are some limitations to this study. First, the UKB reaction time test is brief and consists of fewer trials than are typically used in simple reaction time tests. This paucity of trials may have reduced the reliability of the measurement thereby affecting our ability to accurately capture RTV for participants, contributing towards the low estimate for h2SNP. While the associations between RTV and other mental health and cognitive phenotypes in the UKB are in keeping with the associations observed in previous studies of RTV using validated reaction time tests, future studies should consider using a more comprehensive assessment of reaction time. Second, the assessment of reaction time variability differed between the UKB, SAX, and TOP study and heterogeneity in the phenotype may have affected comparisons of RTV among studies. Third, there is a lack of well-powered studies with which to conduct a replication GWAS. The moderate sample size of the replication study and limitations pertaining to the trans-ancestry replicability of risk variants may account for the non-replication of the lead SNPs from our discovery GWAS. Fourth, the low h2SNP of RTV may have affected the accuracy and predictive power of the RTV-PGS. While this low h2SNP limits the potential use of the PGS to predict RTV, we were still able to fulfil the aim of the polygenic score analyses, which was to evaluate the replicability of the results from the discovery GWAS. Lastly, we used self-reported ethnicity as a population descriptor for participants from the UKB. While using the ethnic groups provided by the UKB facilitates comparability with other studies using the same data, future work should consider alternative population descriptors that are better able to capture genetic variation between groups.
In summary, we have conducted the first large-scale GWAS of RTV using 404,302 samples and identified 7 independent associated loci. Several of the implicated genes are involved in neural development and synaptic function and are known to be associated with other cognitive traits. These findings suggest that disruptions to these processes may affect shared biological mechanisms responsible for maintaining the integrity of various aspects of cognitive function. Despite the relatively low SNP-based heritability of RTV observed in our study, it provides evidence that there is a genetic contribution to the trait. Future studies may leverage these findings to improve our understanding of the genetic mechanisms contributing to RTV and gain novel insight into the biological underpinnings of related complex disorders, like schizophrenia.
Supplementary information
Acknowledgements
We would like to thank the participants and members of the research teams involved in the UK Biobank, SAX Study and TOP Study. We would like to express our gratitude to Torill Ueland and Beathe Haatveit for their support of this work and for facilitating access to the cognitive data from the TOP Study. The current study was supported by National Institute of Mental Health (NIMH: Grant number U01MH125053), and The Research Council of Norway (275054). This research has been conducted using data from UK Biobank, a major biomedical database (Project ID number 27412; www.ukbiobank.ac.uk). This work was partly performed on the TSD (Tjeneste for Sensitive Data) facilities, owned by the University of Oslo, operated and developed by the TSD service group at the University of Oslo, IT-Department (USIT) (tsd-drift@usit.uio.no). Computations for data from the SAX study were performed using facilities provided by the University of Cape Town’s ICTS High Performance Computing team: hpc.uct.ac.za. The SAX study was funded by the NIMH (Grant number: 5UO1MH096754). AS was supported by the EEA and Norway grant (#EEA-RO-NO-2018-0573) and the Research Council of Norway (#326813). OA receives funding from the Research Council of Norway (#223273, #275054, #324252, #324499). DS, RR and SD are supported by the South African Medical Research Council. The content is solely the responsibility of the authors and does not necessarily represent the official views of the funders.
Author contributions
OW conceptualised the research question, performed the data analysis, and drafted the manuscript with input from AAS and SD. All authors contributed to data interpretation and editing of the manuscript. All authors approved the final version of the manuscript.
Funding
Open access funding provided by University of Cape Town.
Code availability
The present study applied previously published approaches, of which codes are shared on public repositories: regenie v3.1 (https://github.com/rgcgithub/regenie), METAL (https://github.com/statgen/METAL/tree/master/metal), MiXeR v1.2 (https://github.com/precimed/mixer), GCTA v1.94 (https://github.com/jianyangqt/gcta), LDSC v1.0.1 (https://github.com/bulik/ldsc), PRSice v2.3.5 (https://github.com/choishingwan/PRSice), and PRS-CS v1.0.0 (https://github.com/getian107/PRScs/tree/v1.0.0). All code used for carrying out the described analysis is available upon request from the corresponding author.
Competing interests
The authors declare the following competing interest: OA has received speaker’s honorarium from Lundbeck and is a consultant for Healthlytix. The remaining authors declare no competing interests.
Ethics approval
This work was approved by the University of Cape Town Human Research Ethics Committee (reference number - 734/2021). The UKB has ethical approval (REC reference number - 11/NW/0382) and is overseen by an Independent Ethics and Governance council. The SAX Study was approved by the University of Cape Town Human Research Ethics Committee (reference number - 049/2013). The TOP Study was approved by the Norwegian Scientific Ethical Committee and the Norwegian Data Protection Agency. Informed consent was obtained from participants in the UKB, SAX and TOP Study.
Footnotes
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
The online version contains supplementary material available at 10.1038/s41380-023-02292-9.
References
- 1.MacDonald SW, Li SC, Bäckman L. Neural underpinnings of within-person variability in cognitive functioning. Psychol Aging. 2009;24:792–808. doi: 10.1037/a0017798. [DOI] [PubMed] [Google Scholar]
- 2.Haynes BI, Bunce D, Kochan NA, Wen W, Brodaty H, Sachdev PS. Associations between reaction time measures and white matter hyperintensities in very old age. Neuropsychologia. 2017;96:249–55. doi: 10.1016/j.neuropsychologia.2017.01.021. [DOI] [PubMed] [Google Scholar]
- 3.Bunce D, Bielak AA, Cherbuin N, Batterham PJ, Wen W, Sachdev P, et al. Utility of intraindividual reaction time variability to predict white matter hyperintensities: a potential assessment tool for clinical contexts? J Int Neuropsychol Soc. 2013;19:971–6. doi: 10.1017/S1355617713000830. [DOI] [PubMed] [Google Scholar]
- 4.Fassbender C, Scangos K, Lesh TA, Carter CS. RT distributional analysis of cognitive-control-related brain activity in first-episode schizophrenia. Cogn Affect Behav Neurosci. 2014;14:175–88. doi: 10.3758/s13415-014-0252-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kofler MJ, Rapport MD, Sarver DE, Raiker JS, Orban SA, Friedman LM, et al. Reaction time variability in ADHD: a meta-analytic review of 319 studies. Clin Psychol Rev. 2013;33:795–811. doi: 10.1016/j.cpr.2013.06.001. [DOI] [PubMed] [Google Scholar]
- 6.Kuntsi J, Klein C. Intraindividual variability in ADHD and its implications for research of causal links. Curr Top Behav Neurosci. 2012;9:67–91. doi: 10.1007/7854_2011_145. [DOI] [PubMed] [Google Scholar]
- 7.Kaiser S, Roth A, Rentrop M, Friederich HC, Bender S, Weisbrod M. Intra-individual reaction time variability in schizophrenia, depression and borderline personality disorder. Brain Cogn. 2008;66:73–82. doi: 10.1016/j.bandc.2007.05.007. [DOI] [PubMed] [Google Scholar]
- 8.Brotman MA, Rooney MH, Skup M, Pine DS, Leibenluft E. Increased intrasubject variability in response time in youths with bipolar disorder and at-risk family members. J Am Acad Child Adolesc Psychiatry. 2009;48:628–35. doi: 10.1097/CHI.0b013e3181a27527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Haynes BI, Bauermeister S, Bunce D. A systematic review of longitudinal associations between reaction time intraindividual variability and age-related cognitive decline or impairment, dementia, and mortality. J Int Neuropsychol Soc. 2017;23:431–45. doi: 10.1017/S1355617717000236. [DOI] [PubMed] [Google Scholar]
- 10.Crosbie J, Arnold P, Paterson A, Swanson J, Dupuis A, Li X, et al. Response inhibition and ADHD traits: correlates and heritability in a community sample. J Abnorm Child Psychol. 2013;41:497–507. doi: 10.1007/s10802-012-9693-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kuntsi J, Rogers H, Swinard G, BÖRger N, Meere J, Rijsdijk F, et al. Reaction time, inhibition, working memory and ‘delay aversion’ performance: genetic influences and their interpretation. Psychol Med. 2006;36:1613–24. doi: 10.1017/S0033291706008580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shin YS, Kim SN, Shin NY, Jung WH, Hur JW, Byun MS, et al. Increased intra-individual variability of cognitive processing in subjects at risk mental state and schizophrenia patients. PLoS ONE. 2013;8:e78354. doi: 10.1371/journal.pone.0078354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rentrop M, Rodewald K, Roth A, Simon J, Walther S, Fiedler P, et al. Intra-individual variability in high-functioning patients with schizophrenia. Psychiatry Res. 2010;178:27–32. doi: 10.1016/j.psychres.2010.04.009. [DOI] [PubMed] [Google Scholar]
- 14.Bielak AA, Hultsch DF, Strauss E, Macdonald SW, Hunter MA. Intraindividual variability in reaction time predicts cognitive outcomes 5 years later. Neuropsychology. 2010;24:731–41. doi: 10.1037/a0019802. [DOI] [PubMed] [Google Scholar]
- 15.Lövdén M, Li S-C, Shing YL, Lindenberger U. Within-person trial-to-trial variability precedes and predicts cognitive decline in old and very old age: longitudinal data from the Berlin Aging Study. Neuropsychologia. 2007;45:2827–38. doi: 10.1016/j.neuropsychologia.2007.05.005. [DOI] [PubMed] [Google Scholar]
- 16.Cummins TDR, Jacoby O, Hawi Z, Nandam LS, Byrne MAV, Kim BN, et al. Alpha-2A adrenergic receptor gene variants are associated with increased intra-individual variability in response time. Mol Psychiatry. 2014;19:1031–6. doi: 10.1038/mp.2013.140. [DOI] [PubMed] [Google Scholar]
- 17.Krabbendam L, Isusi P, Galdos P, Echevarria E, Bilbao JR, Martin-Pagola A, et al. Associations between COMTVal158Met polymorphism and cognition: direct or indirect effects? Eur Psychiatry. 2006;21:338–42. doi: 10.1016/j.eurpsy.2006.03.007. [DOI] [PubMed] [Google Scholar]
- 18.Grant P, Kuepper Y, Wielpuetz C, Hennig J. Differential associations of dopamine-related polymorphisms with discrete components of reaction time variability: relevance for attention deficit/hyperactivity disorder. Neuropsychobiology. 2014;69:220–6. doi: 10.1159/000360367. [DOI] [PubMed] [Google Scholar]
- 19.Chabris CF, Hebert BM, Benjamin DJ, Beauchamp J, Cesarini D, van der Loos M, et al. Most reported genetic associations with general intelligence are probably false positives. Psychol Sci. 2012;23:1314–23. doi: 10.1177/0956797611435528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pinar A, Hawi Z, Cummins T, Johnson B, Pauper M, Tong J, et al. Genome-wide association study reveals novel genetic locus associated with intra-individual variability in response time. Transl Psychiatry. 2018;8:207. doi: 10.1038/s41398-018-0262-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim M-S, Akhtar MW, Adachi M, Mahgoub M, Bassel-Duby R, Kavalali ET, et al. An essential role for histone deacetylase 4 in synaptic plasticity and memory formation. J Neurosci. 2012;32:10879–86. doi: 10.1523/JNEUROSCI.2089-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sudlow C, Gallacher J, Allen N, Beral V, Burton P, Danesh J, et al. UK Biobank: an open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 2015;12:e1001779. doi: 10.1371/journal.pmed.1001779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fawns-Ritchie C, Deary IJ. Reliability and validity of the UK Biobank cognitive tests. PLoS ONE. 2020;15:e0231627. doi: 10.1371/journal.pone.0231627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hagenaars SP, Harris SE, Davies G, Hill WD, Liewald DCM, Ritchie SJ, et al. Shared genetic aetiology between cognitive functions and physical and mental health in UK Biobank (N=112 151) and 24 GWAS consortia. Mol Psychiatry. 2016;21:1624–32. doi: 10.1038/mp.2015.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Stuss DT, Murphy KJ, Binns MA, Alexander MP. Staying on the job: the frontal lobes control individual performance variability. Brain. 2003;126:2363–80. doi: 10.1093/brain/awg237. [DOI] [PubMed] [Google Scholar]
- 26.Bycroft C, Freeman C, Petkova D, Band G, Elliott LT, Sharp K, et al. The UK Biobank resource with deep phenotyping and genomic data. Nature. 2018;562:203–9. doi: 10.1038/s41586-018-0579-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chang CC, Chow CC, Tellier LC, Vattikuti S, Purcell SM, Lee JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience. 2015;4:7. doi: 10.1186/s13742-015-0047-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Willer CJ, Li Y, Abecasis GR. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics. 2010;26:2190–1. doi: 10.1093/bioinformatics/btq340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.R Core Team. R: a language and environment for statistical computing. 4.1.0. ed. Vienna, Austria: R Foundation for Statistical Computing; 2021.
- 30.Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. 2017;8:1826. doi: 10.1038/s41467-017-01261-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Abecasis GR, Altshuler D, Auton A, Brooks LD, Durbin RM, Gibbs RA, et al. A map of human genome variation from population-scale sequencing. Nature. 2010;467:1061–73. doi: 10.1038/nature09534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pruim RJ, Welch RP, Sanna S, Teslovich TM, Chines PS, Gliedt TP, et al. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 2010;26:2336–7. doi: 10.1093/bioinformatics/btq419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wang K, Li M, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010;38:e164. doi: 10.1093/nar/gkq603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kircher M, Witten DM, Jain P, O’Roak BJ, Cooper GM, Shendure J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat Genet. 2014;46:310–5. doi: 10.1038/ng.2892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Boyle AP, Hong EL, Hariharan M, Cheng Y, Schaub MA, Kasowski M, et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 2012;22:1790–7. doi: 10.1101/gr.137323.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ernst J, Kellis M. Chromatin-state discovery and genome annotation with ChromHMM. Nat Protoc. 2017;12:2478–92. doi: 10.1038/nprot.2017.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fromer M, Roussos P, Sieberts SK, Johnson JS, Kavanagh DH, Perumal TM, et al. Gene expression elucidates functional impact of polygenic risk for schizophrenia. Nat Neurosci. 2016;19:1442–53. doi: 10.1038/nn.4399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ghoussaini M, Mountjoy E, Carmona M, Peat G, Schmidt EM, Hercules A, et al. Open targets genetics: systematic identification of trait-associated genes using large-scale genetics and functional genomics. Nucleic Acids Res. 2021;49:D1311–20. doi: 10.1093/nar/gkaa840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mountjoy E, Schmidt EM, Carmona M, Schwartzentruber J, Peat G, Miranda A, et al. An open approach to systematically prioritize causal variants and genes at all published human GWAS trait-associated loci. Nat Genet. 2021;53:1527–33. doi: 10.1038/s41588-021-00945-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.de Leeuw CA, Mooij JM, Heskes T, Posthuma D. MAGMA: generalized gene-set analysis of GWAS data. PLoS Comput Biol. 2015;11:e1004219. doi: 10.1371/journal.pcbi.1004219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Liberzon A, Birger C, Thorvaldsdóttir H, Ghandi M, Mesirov JP, Tamayo P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015;1:417–25. doi: 10.1016/j.cels.2015.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yang J, Lee SH, Goddard ME, Visscher PM. GCTA: a tool for genome-wide complex trait analysis. Am J Hum Genet. 2011;88:76–82. doi: 10.1016/j.ajhg.2010.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Holland D, Frei O, Desikan R, Fan CC, Shadrin AA, Smeland OB, et al. Beyond SNP heritability: polygenicity and discoverability of phenotypes estimated with a univariate Gaussian mixture model. PLoS Genet. 2020;16:e1008612. doi: 10.1371/journal.pgen.1008612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bulik-Sullivan BK, Loh PR, Finucane HK, Ripke S, Yang J, Schizophrenia Working Group of the Psychiatric Genomics Consortium, et al. LD score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat Genet. 2015;47:291–5. doi: 10.1038/ng.3211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bulik-Sullivan B, Finucane HK, Anttila V, Gusev A, Day FR, Loh PR, et al. An atlas of genetic correlations across human diseases and traits. Nat Genet. 2015;47:1236–41. doi: 10.1038/ng.3406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Davies G, Lam M, Harris SE, Trampush JW, Luciano M, Hill WD, et al. Study of 300,486 individuals identifies 148 independent genetic loci influencing general cognitive function. Nat. Commun. 2018;9:2098. doi: 10.1038/s41467-018-04362-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Engh JA, Friis S, Birkenaes AB, Jónsdóttir H, Klungsøyr O, Ringen PA, et al. Delusions are associated with poor cognitive insight in schizophrenia. Schizophr Bull. 2010;36:830–5. doi: 10.1093/schbul/sbn193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gulsuner S, Stein DJ, Susser ES, Sibeko G, Pretorius A, Walsh T, et al. Genetics of schizophrenia in the South African Xhosa. Science. 2020;367:569–73. doi: 10.1126/science.aay8833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Choi SW, O’Reilly PF. PRSice-2: Polygenic Risk Score software for biobank-scale data. GigaScience. 2019;8:giz082. doi: 10.1093/gigascience/giz082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ge T, Chen C-Y, Ni Y, Feng Y-CA, Smoller JW. Polygenic prediction via Bayesian regression and continuous shrinkage priors. Nat Commun. 2019;10:1776. doi: 10.1038/s41467-019-09718-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Savage JE, Jansen PR, Stringer S, Watanabe K, Bryois J, de Leeuw CA, et al. Genome-wide association meta-analysis in 269,867 individuals identifies new genetic and functional links to intelligence. Nat Genet. 2018;50:912–9. doi: 10.1038/s41588-018-0152-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sollis E, Mosaku A, Abid A, Buniello A, Cerezo M, Gil L, et al. The NHGRI-EBI GWAS Catalog: knowledgebase and deposition resource. Nucleic Acids Res. 2023;51:D977–d985. doi: 10.1093/nar/gkac1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Karayannis T, Au E, Patel JC, Kruglikov I, Markx S, Delorme R, et al. Cntnap4 differentially contributes to GABAergic and dopaminergic synaptic transmission. Nature. 2014;511:236–40. doi: 10.1038/nature13248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nishimune A, Isaac JT, Molnar E, Noel J, Nash SR, Tagaya M, et al. NSF binding to GluR2 regulates synaptic transmission. Neuron. 1998;21:87–97. doi: 10.1016/s0896-6273(00)80517-6. [DOI] [PubMed] [Google Scholar]
- 55.Hanley JG. NSF binds calcium to regulate its interaction with AMPA receptor subunit GluR2. J Neurochem. 2007;101:1644–50. doi: 10.1111/j.1471-4159.2007.04455.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Russell VA, Oades RD, Tannock R, Killeen PR, Auerbach JG, Johansen EB, et al. Response variability in attention-deficit/hyperactivity disorder: a neuronal and glial energetics hypothesis. Behav Brain Funct. 2006;2:30. doi: 10.1186/1744-9081-2-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.MacDonald SW, Cervenka S, Farde L, Nyberg L, Bäckman L. Extrastriatal dopamine D2 receptor binding modulates intraindividual variability in episodic recognition and executive functioning. Neuropsychologia. 2009;47:2299–304. doi: 10.1016/j.neuropsychologia.2009.01.016. [DOI] [PubMed] [Google Scholar]
- 58.Pouget P, Wattiez N, Rivaud-Péchoux S, Gaymard B. A fragile balance: perturbation of GABA mediated circuit in prefrontal cortex generates high intraindividual performance variability. PLoS ONE. 2009;4:e5208. doi: 10.1371/journal.pone.0005208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lam M, Trampush JW, Yu J, Knowles E, Davies G, Liewald DC, et al. Large-scale cognitive GWAS meta-analysis reveals tissue-specific neural expression and potential nootropic drug targets. Cell Rep. 2017;21:2597–613. doi: 10.1016/j.celrep.2017.11.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mei K, Guo W. The exocyst complex. Curr Biol. 2018;28:R922–5. doi: 10.1016/j.cub.2018.06.042. [DOI] [PubMed] [Google Scholar]
- 61.Martin-Urdiroz M, Deeks MJ, Horton CG, Dawe HR, Jourdain I. The exocyst complex in health and disease. Front Cell Dev Biol. 2016;4:24. doi: 10.3389/fcell.2016.00024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hosaka M, Watanabe T. Secretogranin III: a bridge between core hormone aggregates and the secretory granule membrane. Endocr J. 2010;57:275–86. doi: 10.1507/endocrj.k10e-038. [DOI] [PubMed] [Google Scholar]
- 63.Taupenot L, Harper KL, O’Connor DT. The chromogranin–secretogranin family. N Engl J Med. 2003;348:1134–49. doi: 10.1056/NEJMra021405. [DOI] [PubMed] [Google Scholar]
- 64.Uezu A, Kanak DJ, Bradshaw TW, Soderblom EJ, Catavero CM, Burette AC, et al. Identification of an elaborate complex mediating postsynaptic inhibition. Science. 2016;353:1123–9. doi: 10.1126/science.aag0821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Karlsson Linnér R, Mallard TT, Barr PB, Sanchez-Roige S, Madole JW, Driver MN, et al. Multivariate analysis of 1.5 million people identifies genetic associations with traits related to self-regulation and addiction. Nat Neurosci. 2021;24:1367–76. doi: 10.1038/s41593-021-00908-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wendt FR, Pathak GA, Deak JD, De Angelis F, Koller D, Cabrera-Mendoza B, et al. Using phenotype risk scores to enhance gene discovery for generalized anxiety disorder and posttraumatic stress disorder. Mol Psychiatry. 2022;27:2206–15. doi: 10.1038/s41380-022-01469-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Yu D, Sul JH, Tsetsos F, Nawaz MS, Huang AY, Zelaya I, et al. Interrogating the genetic determinants of Tourette’s syndrome and other tic disorders through genome-wide association studies. Am J Psychiatry. 2019;176:217–27. doi: 10.1176/appi.ajp.2018.18070857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Finkel D, McGue M. Genetic and environmental influences on intraindividual variability in reaction time. Exp Aging Res. 2007;33:13–35. doi: 10.1080/03610730601006222. [DOI] [PubMed] [Google Scholar]
- 69.Woods DL, Wyma JM, Yund EW, Herron TJ, Reed B. Factors influencing the latency of simple reaction time. Front Hum Neurosci. 2015;9:131. doi: 10.3389/fnhum.2015.00131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Choi SW, Mak TS, O’Reilly PF. Tutorial: a guide to performing polygenic risk score analyses. Nat Protoc. 2020;15:2759–72. doi: 10.1038/s41596-020-0353-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Majara L, Kalungi A, Koen N, Tsuo K, Wang Y, Gupta R, et al. Low and differential polygenic score generalizability among African populations due largely to genetic diversity. HGG Adv. 2023;4:100184. doi: 10.1016/j.xhgg.2023.100184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Der G, Deary IJ. Reaction times match IQ for major causes of mortality: evidence from a population based prospective cohort study. Intelligence. 2018;69:134–45. doi: 10.1016/j.intell.2018.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Rabbitt P, Osman P, Moore B, Stollery B. There are stable individual differences in performance variability, both from moment to moment and from day to day. Q J Exp Psychol A. 2001;54:981–1003. doi: 10.1080/713756013. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The present study applied previously published approaches, of which codes are shared on public repositories: regenie v3.1 (https://github.com/rgcgithub/regenie), METAL (https://github.com/statgen/METAL/tree/master/metal), MiXeR v1.2 (https://github.com/precimed/mixer), GCTA v1.94 (https://github.com/jianyangqt/gcta), LDSC v1.0.1 (https://github.com/bulik/ldsc), PRSice v2.3.5 (https://github.com/choishingwan/PRSice), and PRS-CS v1.0.0 (https://github.com/getian107/PRScs/tree/v1.0.0). All code used for carrying out the described analysis is available upon request from the corresponding author.