

Abstract
Afterimages are illusory, conscious visual perseverations commonly induced by preceding light stimulation. A retinal centric view on the physiological source of afterimages is dominant. In addition, post-retinal mechanisms have been considered in the formation and modulation of afterimage perception, including cortical processes. A cortical role in afterimage perception posits possible shared neural mechanisms between afterimages and other conscious perceptions that emerge completely from central neural sources (e.g., imagery, hallucination, and dreams). To examine this hypothesis, we tested a perceptual link between afterimages and visual imagery. Framing the current experiment, we review more than a century of literature that evidences post-retinal processes in afterimage perception. Subsequently, we present an innovative afterimage perception reporting paradigm, validated on image stimuli, that allowed participants to indicate the perceived sharpness, contrast, and duration of their afterimages. From these perceptual reports, we discovered a novel category of evidence for cortical mechanisms in afterimage perception: the vividness of visual imagery positively correlates with afterimage brightness and sharpness. This result motivates future investigations on the neural mechanisms of afterimage perception and encourages implementing afterimages as a model perception to interrogate other kinds of conscious experience with known cortical origin.
Introduction
Afterimages are visual perseverations – lasting seconds to minutes – that often follow the cessation of intense light stimulation (e.g., viewing the Sun) but absent the original inducing light source. Analogous perceptual perseverations are also reported in other human senses, including auditory afterimages or aftersounds1–3. Afterimages have been a source of intrigue for centuries because of their apparent ubiquity, including among non-human animals (e.g., macaques, cats, and pigeons) and its unique insight on the physiological mechanisms of vision4–7. In fact, afterimages helped debunk emission theories of vision that explained conscious sight by the projection of light or aether rays from the eyes. Afterimages share perceptual characteristics with aftereffects (e.g., the McCollough effect8) and filling-in illusions (e.g., Kanizsa or occluded stimuli illusions9). However, afterimages are distinct because they do not require concurrent visual input (i.e., they can appear in total darkness), while aftereffects and illusory filling-in necessarily act on visual sensory signals10. Accordingly, afterimages can be framed as a kind of hallucination – a conscious vision without a simultaneous real world light source.
A motivating interest in afterimage perception is its unique perceptual effect for interrogating the neural mechanisms of conscious perception11. For example, a curious observation in afterimage perception that is used to interpret the cortical mechanisms of visual conscious perception is that the adaptor or inducer stimulus (i.e., the light stimulation that may subsequently produce an afterimage) can be made unconscious (e.g., continuous flash suppression), yet still the afterimage is consciously perceived11–14. In an analogous dissociation between the conscious perception of inducer stimuli and afterimages, some people with cortical blindness can perceive afterimages in their blinded field but without perceiving the preceding inducer stimulus15,16. Thus, afterimages provide a unique probe to interrogate the mechanisms of conscious perception in healthy and disordered neurophysiology.
Also encouraging the study of afterimages is a major unresolved query on its physiological source: to what degree are afterimages retinal versus cortical phenomena17? Initial theories on the physiological mechanisms of afterimages argued a pure retinal process. Likewise, Hermann von Helmholtz (1821–1894) described afterimages as a “photograph on the retina”18. This early, retinal-centric view of afterimages was evidenced by the observation that afterimages follow eye movements, although this property is also attributable to central processes19. Later, evidence emerged that afterimages result by the fatigue or bleaching of retinal photoreceptors that persistently signal in the absence of physical light stimulation7,10,20–22. A similar process is suggested to form the basis of the opponent-process theory for chromatic afterimages, whereby complementary color afterimages are perceived according to the inducer color (e.g., a yellow inducer forms a blue afterimage), predicted by opponent visual pairs – black-white, red-green, and blue-yellow – so that adaptation to one half of an opponent pair will drive its opposite hue in the subsequent afterimage10,23.
In support of the retinal view, the duration of afterimages is predicted by the extent of dark adaption and the contrast and duration of the inducer stimulus24–26. Also, the color of afterimages corresponds with the concentration of cone receptors in the retina, for example, negative afterimages induced by a blue adaptor image highlight the foveal blue scotoma (i.e., the lack of short-wave-sensitive cones in the human foveola)27. Moreover, direct recordings from retinal ganglion cells find a latent postreceptor rebound response following inducer stimulation that may originate from photoreceptor signaling28.
If retinal physiology fully explains afterimage perception, its mechanism is best described as a bottom-up process – emergent from the visual periphery. However, a retinal explanation does not fully account for all findings on the physiological source of afterimages. In fact, there is more than a century of research and clinical reports that evidence post-retinal processes in afterimage perception (this literature is summarized below). Therefore, it remains an open question to what degree non-retinal mechanisms, particularly the cortex, contributes to forming and modifying afterimage perception.
Determining the degree of retinal versus cortical mechanisms in afterimage perception is significant because it helps to define the perceptual category of afterimages. If afterimages are totally retinal then they should be considered a type of sensory vision – a retinal sourced visual perception but without simultaneous light stimulation. Alternatively, if afterimages emerge by cortical mechanisms, these perceptions could be linked with visual conscious experiences that originate directly from cortical processing (i.e., no sensory input), including imagination, hallucination, and dreams – a central vision independent of peripheral physiology. Accordingly, the current study examined a possible perceptual relationship between afterimages and visual imagery. Establishing the context in which this investigation was pursued are experimental and clinical examples suggesting post-retinal neural processes in afterimage perception. Below is a brief review of this evidence.
An initial indication of post-retinal mechanisms in afterimage perception is that the bleaching of photoreceptors is not a necessary condition for the formation of afterimages6,29,30. In fact, afterimages can emerge without previous photoreceptor stimulation, as in afterimages by illusory vision (e.g., a perceptually filled-in image)31. Likewise, studies find that color spreading in afterimages can extend beyond the boundary of the preceding inducer stimulus32,33. Furthermore, there are reports of afterimages evoked by dreams, imagery, and hallucination17,34–36. Similarly, conditioned afterimages are reported by pairing tones and inducer stimuli and then withholding the anticipated inducer, yet participants still report seeing afterimages without the preceding visual stimulation37.
Post-retinal mechanisms of afterimages are also suggested by the observation of afterimage interocular transfer or grouping effects, although the strength of this evidence for central neural involvement has been criticized38. For example, when an afterimage is induced in one eye, the visual input to the non-induced eye can influence the afterimage perception (e.g., duration and contrast)39,40. Moreover, interocular suppression effects are manifest by reduced afterimage duration in a binocular rivalry paradigm (i.e., the inducer stimulus is shown in one eye and the distractor stimulus is shown in the other eye)41. Another experiment found that when each eye is presented with a separate image with unique form-color pairings (e.g., pink-vertical gratings and green-horizontal gratings), the subsequent afterimage might show a misbinding of the original form-color pairs (e.g., pink-horizontal gratings), hinting at cortical processes with access to binocular input42. Yet another experiment found that two unique images shown in either eye that are by themselves incoherent can be combined in the afterimage perception to form a coherent vision43. Similarly, when two patterned images of perpendicular gratings are shown in either eye, the most commonly reported afterimage perception was one that required interocular grouping44. Together, these results support the role of post-retinal mechanisms that act to integrate preceding sensory input from each eye in the formation of afterimages, just as binocular fusion that occurs in visual cortex for normal sensory vision.
An additional observation that disputes a pure retinal explanation of afterimages is that the low-level features of the inducer stimulus that predicts photoreceptor bleaching does not perfectly determine the perceptual qualities of the afterimage. For example, the duration of afterimages are not identical for equally bright grating inducers, instead partly modulated according to the grating orientation45. Thus, afterimages can be influenced by higher-order visual features of the inducer that are represented in cortex. Likewise, there is evidence of numerous top-down influences on afterimage perception. For example, there are multiple reports that attention and awareness influence afterimage perception13,41,46–50. Moreover, priming and priors modulate afterimages (e.g., the perceived gender of a face, color, and vividness)51,52. In a linked result, afterimages disappear when they are positioned at eccentricities that extends beyond the typical boundary of the visual field, an effect present even in total darkness, indicating the influence of kinesthetic central priors (e.g., eye position) that act to degrade afterimages when moved outside the typical visual field boundary53.
Further evidence of post-retinal neural mechanisms in afterimage perception are examples of contextual influences. For example, afterimages will appear, disappear, and reappear depending on if the content of the afterimage registers or is congruent with the concurrent visual sensory input54. In another contextual case, the visual spatial boundary, for example, the outline of an open shape, acts to contain the subsequent afterimage that perceptually fills within this boundary, and this visual boundary can even dictate the afterimage color based on the inducer characteristics, thus a single inducer resulting in multiple types of afterimages depending on the contextual constraints33,55. Contextual influences are also found in the instances of the Ebbinghaus-Titchener illusion (i.e., perceived image size is influenced by its perceived visual surround) and Emmert’s law (i.e., perceived image size is influenced by its perceived visual distance). In these instances, afterimages will appear larger or smaller depending on its surrounding context or the perceived distance of the surface on which the afterimage is projected, even though the size of the retinal stimulation is fixed according to the initial inducer56–60. Importantly, the perceived distance, even if illusory, predicts afterimage size58. In contrast, a visual scotoma caused by retinal damage does not change size according to viewing distance19. A functional magnetic resonance imaging (fMRI) study corroborates these behavioral results, showing signal increases in more eccentric sites on primary visual cortex (V1) corresponding with the perceived size of the afterimage, modulated by viewing distance (i.e., larger afterimages according to viewing distance predicts fMRI signal located at greater V1 eccentricity, while the inducer stimulus viewing distance is fixed)61. Interestingly, in the Ebbinghaus-Titchener illusion, when the contextual images are shown as an afterimage, they too can influence the perceived size of physical images, demonstrating that afterimages can also act as a contextual modulator62.
Another instance of contextual modulation in afterimage perception is the warping of the afterimage shape by the surface on which it is viewed. For example, an afterimage induced by the image of a circle, nonetheless, appears oval when the afterimage is seen on a slanted surface – the afterimage stretched along the dimension of the perceived slant63. However, if a physically slanted surface is perceived as flat (e.g., the illusory perception that a physically angled wall is flat in an Ames room), the afterimage is not warped, thus this contextual modulation is perceptual63. Together, these examples of contextual effects support that afterimages are more than static imprints on the retina, instead interacting with higher-order surround features, an integrative process attributed to the cortex (e.g.,64).
Multisensory influences on afterimage perception extends support of post-retinal processes because multisensory signal integration largely occurs in cortex65. Already, the previously discussed report of conditioned afterimages by pairing tones and images suggest an instance of a multisensory influence on afterimage perception37. In addition, sound stimulation is found to enhance the brightness and size of afterimages66. Furthermore, proprioceptive feedback alters afterimage perception. For example, the afterimage of hands or held objects will selectively disappear or “crumble” when a movement is made in the dark (i.e., without visual feedback; only sensory-motor input of movement), in fact, this effect scaling with the strength of proprioceptive input67–70. These results suggest that afterimages are inhibited when proprioceptive feedback indicates that the afterimage no longer concurs with the current body or object location in space.
Furthermore, moving the head back and forward or moving the hand that is captured in an afterimage nearer or further away while in complete darkness can increase and decrease the size of the afterimages in accordance with Emmert’s law67. This result corroborates the rare reports in healthy people who experience their afterimages as fixed to real world coordinates during head and eye movements71. Surprisingly, in the same individuals, afterimages of their hands or held objects updated their perceived position in or near real time according to hand movements, all without visual sensory feedback (i.e., with eyes closed)71. Perplexing still, it was observed that the afterimage of objects grasped in each hand would switch between hands in the afterimage when the hands of the participant were crossed at the midline71. In summary, these results suggest that the neural mechanisms involved in forming a body schema and tracking objects in space – processes with known cortical bases – are also acting to modulate afterimage perception but without subsequent visual input.
Finally, support of a cortical role in afterimage perception comes by clinical cases of sensory perseveration that suggests the same kind of sensory persistence as in afterimages but the result of cortical aberrance. For example, palinopsia is the experience of exaggerated afterimages, including prolonged movement traces that typically result from cortical lesion in parietal-occipital sites72–74. Curiously, the same kind of visual motion trace afterimages are reported by healthy people while under the influence of psychedelic substances – a chemically evoked palinopsia that can persist for years after drug use.75 Similarly, in posterior cortical atrophy – a variant of Alzheimer’s disease that typical involves the temporal, parietal, and occipital lobes – prolonged color afterimages are reported76,77.
Clinical groups that suggest cortical aberrance as the source of chronic and extreme sensory perseveration is corroborated by neurophysiological studies of afterimages in healthy cortical physiology. For example, an early experiment in cats found visual cortical neurons that persist in their activity after the offset of a visual stimulus corresponding with the duration of the afterimage, although this neural response could also be explained by persistent retinal input following photoreceptor bleaching5. Moreover, afterimage-linked V1 fMRI signals have been recorded in humans61. Likewise, cortical responses have been measured during aftersounds78. Lastly, an experiment with transcranial magnetic stimulation of the visual cortex found that disrupting occipital cortical electrophysiology influenced the perceptual fading of afterimages79.
The totality of the reviewed evidence tells that a purely retinal explanation of afterimages is incomplete. Instead, afterimages are likely the result of both retinal and cortical processes, and the balance between retinal versus cortical neural mechanisms may differ depending on how the afterimage was induced (e.g., weak versus strong photoreceptor bleaching inducers, or the apparent instances of afterimages absent retinal stimulation altogether)35,80.
If some categories of afterimages involve post-retinal processes, particularly cortical mechanisms, a possible implication is that afterimages share neural bases with visual conscious perceptions that have central neural origins (e.g., imagery and hallucination). Accordingly, we speculated that afterimage perception may share perceptual features to those conscious experiences of known cortical origin. Specifically, we tested if the vividness of visual imagery (i.e., the ability to evoke lifelike visual perception by imagination) correlates with the vividness of afterimages. We hypothesized that the perceptual vividness of visual imagery and afterimages are positively linked (i.e., people with more vivid visual imagery ability also experience more vivid afterimages). Interrogating the perceptual relationship between visual imagery and afterimages is significant because this examination may supply novel behavioral evidence of cortical mechanisms in the emergence and modulation of afterimage perception. Moreover, a relationship in the perception of visual imagery and afterimages supports future investigations on afterimages towards interrogating those conscious perceptions that arrive absent simultaneous sensory input (i.e., sensory-independent perceptions), including hallucinations and dreams.
Results
Afterimage Perception Rate and VVIQ Score
The mean afterimage perception rate following the inducer stimulus presentation was 90.79% (standard deviation [SD] = 14.84%; minimum participant afterimage perception rate = 38.89%; maximum participant afterimage perception rate = 100%; Figure 2A). The mean Vividness of Visual Imagery Questionnaire (VVIQ) score was 60.55 (SD = 10.78; minimum participant VVIQ score = 24; maximum participant VVIQ score = 80; Figure 2B).
Figure 2. Afterimage perception rate, VVIQ score, and relative image and observed afterimage sharpness, contrast, and duration.

(A) Afterimage perception rate calculated as the percentage of inducers where a subsequent afterimage was reported across all trials of the afterimage perception matching tasks (90 trials total). The bar graph indicates the mean afterimage perception percentage across participants (90.79%) and the error bar displays standard deviation (SD; 14.84%). (B) The Vividness of Visual Imagery Questionnaire (VVIQ) score calculated as the sum of scores across all questionnaire items within participant (score range: 16–80; larger values indicating more vivid visual imagery). The bar graph indicates the mean VVIQ score (60.55) and the error bars display the SD (10.78). (C) Relative image and observed afterimage reported maximum sharpness in pixels (px). The relative image sharpness is compared on a trial level against the true image sharpness (true values: 10, 15, or 20 px). The bar height indicates the group mean (Relative = 0.033 px; Observed = 14.27 px) and the error bars display SD (Relative = 1.60 px; Observed = 3.88 px). (D) Relative image and observed afterimage reported maximum contrast. The relative image contrast is compared against the true image maximum contrast (0.25). The bar height indicates the group mean (Relative = 0.01; Observed = 0.21) and the error bars display SD (Relative = 0.04; Observed = 0.068). (E) Relative image and observed afterimage reported duration in seconds (s). The relative image contrast is compared against the true image duration (4 s). The bar height indicates the group mean (Relative = −0.33 s; Observed = 5.35 s) and the error bars display SD (Relative = 0.93 s; Observed = 1.60 s). Comparing the relative image contrast, sharpness, and duration values from zero was not statistically significant (Wilcoxon Rank Sum tests, p > 0.05). In all subplots, the open circles represent individual participants (N = 62).
Image and Afterimage Perceptual Features
The mean image and afterimage maximum sharpness values were 14.87 pixels (SD = 1.91 pixels) and 14.27 pixels (SD = 3.88 pixels), respectively (Figure 2C Observed; image observed not shown). The mean image and afterimage maximum contrast values were 0.26 (SD = 0.04) and 0.21 (SD = 0.068), respectively (Figure 2D Observed; image observed not shown). The mean image and afterimage duration values were 3.67 seconds (SD = 0.93 seconds) and 5.35 seconds (SD = 1.60 seconds), respectively (Figure 2E Observed; image observed not shown).
Accuracy of the Reported Image Sharpness, Contrast, and Duration
There was no statistically significant difference (p > 0.05) between 0 (i.e., a perfect match between the reported and true image sharpness, contrast, and duration) and the reported sharpness, contrast, and duration minus the true image sharpness, contrast, and duration (Figure 2C, D, E Relative; see Methods section for image features and statistical testing details).
VVIQ Score Versus Image Sharpness, Contrast, and Duration
There was no statistically significant correlation for VVIQ score versus image contrast (Pearson correlation coefficient [r] = −0.088; p = 0.50; Figure 3A), VVIQ score versus image sharpness (r = 0.027; p = 0.84; Figure 3C), nor VVIQ score versus image duration (r = −0.045; p = 0.73; Figure 3E).
Figure 3. VVIQ score versus image and afterimage contrast, sharpness, and duration.

All subplots display the Vividness of Visual Imagery Questionnaire (VVIQ) score along the horizontal axis (score range: 16–80; larger values indicating more vivid visual imagery). Larger contrast, sharpness, and duration values indicate brighter, crispier, and prolonged image and afterimage perception, respectively. The thicker grey and red lines draw the linear regression fit of VVIQ score versus image or afterimage contrast, sharpness, and duration. The thin grey and red lines on either side of the main trend line is the 95% confidence interval of the linear regression fit. (A) VVIQ score versus image contrast (Pearson correlation is not statistically significant; Pearson correlation coefficient [r] = −0.088; p = 0.50). (B) VVIQ score versus afterimage contrast (Pearson correlation is statistically significant **; r = 0.34; p = 0.007). (C) VVIQ score versus image sharpness (Pearson correlation is not statistically significant; r = 0.027; p = 0.84). (D) VVIQ score versus afterimage sharpness (Pearson correlation is statistically significant *; r = 0.28; p = 0.028). (E) VVIQ score versus image duration (Pearson correlation is not statistically significant; r = −0.045; p = 0.73). (F) VVIQ score versus afterimage duration (Pearson correlation is not statistically significant; r = 0.23; p = 0.068). In all subplots, the open circles represent individual participants (N = 62).
VVIQ Score Versus Afterimage Sharpness, Contrast, and Duration
A statistically significant positive correlation was found between VVIQ score and afterimage contrast (r = 0.34; p = 0.007; linear regression fit trend line equation: Y = 0.0021*X + 0.083; Figure 3B). The estimated contrast value according to the linear regression fit trend line for a low VVIQ score (minimum participant VVIQ score = 24) was 0.13 and a high VVIQ score (maximum participant VVIQ score = 80) was 0.25 (Figure 4B, C). A statistically significant positive correlation was found between VVIQ score and afterimage sharpness (r = 0.28; p = 0.028; linear regression fit trend line equation: Y = 0.10*X + 8.19; Figure 3D). The estimated sharpness value according to the linear regression fit trend line for a low VVIQ score (minimum participant VVIQ score = 24) was 10.60 pixels and a high VVIQ score (maximum participant VVIQ score = 80) was 16.23 pixels (Figure 4A, C). There was no statistically significant correlation between VVIQ score and afterimage duration (r = 0.23; p = 0.068; Figure 3F).
Figure 4. Image reconstruction of the estimated perceived afterimage sharpness and contrast for low and high VVIQ score participants.
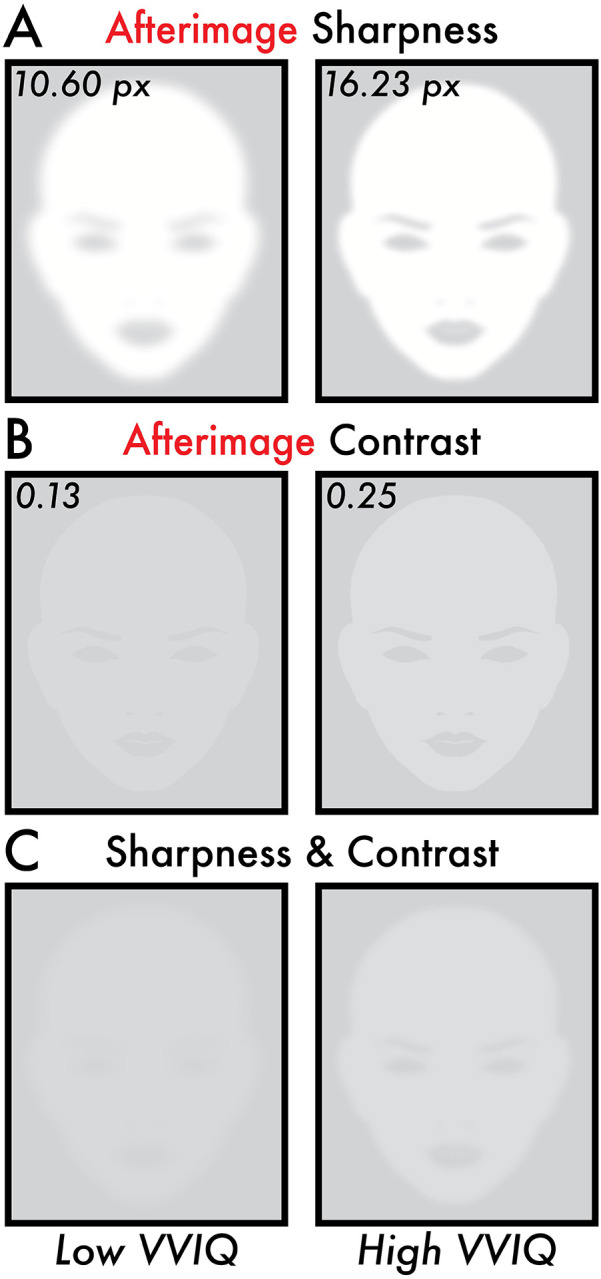
All subplots display image reconstructions of the estimated perceived afterimage sharpness and contrast values according to the fitted linear regression trend lines (Figure 3B, D) for low and high Vividness of Visual Imagery Questionnaire (VVIQ) scores. The low and high VVIQ scores are the minimum (24) and maximum (80) VVIQ scores reported among participants (Figure 2B). (A) Image reconstruction of the estimated perceived afterimage sharpness for low (10.60 pixels [px]) and high (16.23 px) VVIQ scores. (B) Image reconstruction of the estimated perceived afterimage contrast for low (0.13) and high (0.25) VVIQ scores. (C) Image reconstruction combining the estimated perceived afterimage sharpness (A) and contrast (B) for low (10.60 px and 0.13) and high (16.23 px and 0.25) VVIQ scores. Image reconstructions show visually apparent differences in the estimated perceived sharpness and contrast of afterimages between low and high VVIQ score participants.
Discussion
In the current investigation, we introduce a novel source of behavioral evidence for a cortical role in afterimage perception: a link in the perceived vividness of visual imagery and afterimages. To interrogate this relationship, we developed a perception reporting paradigm where participants manipulated the appearance of on-screen, controllable images to match the perceived sharpness, contrast, and duration of image stimuli and afterimages. The efficacy of this reporting method was confirmed by the participant reporting accuracy for the known sharpness, contrast, and duration values of image stimuli (Figures 2C, D, E Relative). Participants used the identical reporting method to indicate the sharpness, contrast, and duration of their afterimages. Future studies can consider using a similar method for reporting other perceptual features (e.g., pattern, form, and color). By these perceptual reports, we discovered a statistically significant, positive correlation between visual imagery vividness and the perceived contrast and sharpness but not duration of afterimages (Figure 3B, D, F). In short, people who reported more vivid visual imagery also tended to report brighter and sharper afterimages.
The correlation effect size between the vividness of imagery and afterimages was small. Nonetheless, reconstructions of the estimated afterimage perceptions for low and high VVIQ scores are visibly distinct – the high VVIQ score afterimage reconstruction revealing an apparently brighter image with sufficient sharpness to discern facial features that are absent in the low VVIQ afterimage reconstruction (Figure 4C Low versus High VVIQ). The main result is further bolstered by the specificity of this relationship to afterimage perception, as in the same participants no relationship was found between visual imagery vividness and image contrast, sharpness, and duration (Figure 3A, C, E).
According to our review of the previous research on afterimages, there is no previous study comparing the perceptual qualities of afterimages and imagery. The nearest instances include the previously highlighted reports of imagery inducing afterimages, particularly among people with vivid imagination (e.g.,34), another study where the author noted that people could experience an afterimage of an image that was otherwise challenging to imagine, and a note by William James (1842–1910) that his visual imagery could be subliminally driven by afterimages, only realized after the fact when shutting his eyes revealed an afterimage that shared perceptual features with the thought that he previously believed was evoked spontaneously71,81. Therefore, the current study is the first direct examination relating afterimage perception and imagery. Likewise, the relationship between the vividness of afterimages and visual imagery is a novel source of evidence that afterimages and visual imagery share neural mechanisms that could explain their linked perceptual experience.
An alternative explanation of these findings is the influence of mediating sensory, cognitive, or behavioral variables required by the perception reporting task (e.g., reaction time and sensory sensitivity) and reporting on visual imagery vividness. The veracity of this account is dampened because indicating the perceptual vividness of images and afterimages versus visual imagery involved orthogonal tasks (see Image and Afterimage Perceptual Vividness Methods section). Specifically, perceptual image and afterimage vividness required adjusting a controllable image with key presses. Meanwhile, visual imagery vividness was inquired using a self-paced questionnaire (i.e., the VVIQ) that involved marking responses with a mouse click. Therefore, sensory, cognitive, and behavioral ability are excluded as likely factors influencing the current findings. Indeed, if such mediating variables explained these results, we would also expect a relationship between the perceptual vividness of images and visual imagery, as identical reporting procedures were involved in both image and afterimage perception, but only for afterimages was a relationship found with VVIQ score. Thus, the reporting produce itself is unlikely to explain the current findings. Still, we cannot rule out the influence of other unknown factors that may be shared across reporting methods and unique to afterimages (e.g., metacognitive or introspection ability).
A limitation of the current investigation is that the recruited participants do not represent the lowest range of the VVIQ score scale – approximately the bottom third – that is estimated to account for a minority (<5%) of the general population82. Only one participant in our data set breached the ~30-score threshold that is commonly used to designate aphantasia – the near or total inability to form visual imagery. Notably, this participant was accurate in reporting the sharpness, contrast, and duration of the image stimuli, suggesting they were not impaired in reporting on visual sensory perceptions. Meanwhile, their reported afterimage contrast and duration values were near to the group average. However, they indicated the least sharp (i.e., blurriest) afterimages of all participants. When this participant is removed from the data set, the correlation between VVIQ score and afterimage contrast and VVIQ score and afterimage duration become stronger, while the correlation for VVIQ score and afterimage sharpness maintains its positive trend but is no longer statistically significant (p > 0.05).
If subsequent experimentation finds that people with low imagery vividness or aphantasia also experience less vivid afterimages, this result would corroborate previous experiments that show sensory sensitivity and imagery are linked to cortical excitability, which may be reduced in aphantasia83,84. Therefore, reduced afterimage vividness in aphantasia would hint at a relationship between cortical excitability and afterimage perception. This result is anticipated by previous findings that show similarity in the modification and character of image and afterimage perception (e.g., corresponding contextual and size constancy effects).
An additional set of future directions to extend these findings and explore the possible cortical mechanisms of afterimages is to interrogate the relationship between afterimages and other forms of centrally emergent conscious perceptions. For example, are people who hallucinate more susceptible to afterimages (i.e., afterimage perception rate) and more vivid afterimages? Hinting at an answer, previous studies find afterimages and aftereffects are altered in people with schizophrenia, a psychiatric disorder partly characterized by hallucinations85,86. A robust confirmatory finding could offer afterimages as a predictive measure in psychiatric disorders and possibly other neurological conditions, as previously suggested in posterior cortical atrophy and Parkinson’s disease76,87. Finally, as a behavioral experiment, the current results only indirectly support shared neural mechanisms between afterimages and visual imagery. In fact, the precise neural mechanisms of afterimage perception are unknown. Future investigations on the neural mechanisms of afterimage perception and other forms of sensory perseveration should involve direct recordings from peripheral and central neurophysiology.
Conclusion
Afterimages have long been a source of curiosity and implemented as a perceptual tool to interrogate vision and the neural mechanism of consciousness. However, the number of publications each year on afterimages has declined since peaks in the 1970s and early 2000s (search term: “afterimage”; PubMed: https://pubmed.ncbi.nlm.nih.gov). In the current investigation, two major contributions are made that may help to revitalize the implementation and expand the implications of research on afterimages. First, we developed a reporting method that allows for accurate acquisition of various perceptual features of image and afterimage perceptions. Second, our results forward a novel finding that the perceptual vividness of visual imagery and afterimages are correlated. This represents a new source of evidence for a cortical basis of afterimage perception and links afterimages to conscious perceptions of central neurophysiological origin. Accordingly, afterimages may be implemented as a model perception for interrogating the neural bases of conscious perceptions that emerge independent of concurrent sensory input, including imagery, hallucination, illusory filling-in, and dreams. The long-term vision for current and future research on afterimage perception should be to contribute to elucidating the neural mechanisms of consciousness and forward approaches for treating people with aberrant conscious perceptions, including palinopsia and other forms of chronic sensory perseveration.
Methods
Participants
Healthy, adult participants (N = 62; males = 22; mean age: 28.90 years; age SD: 10.31 years; mean education = 16.34 years; education SD: 1.91 years) were recruited from the local Bethesda, Maryland community. Two additional participants who completed the study were excluded from analyses because of poor behavioral performance or a corrupted behavioral file. All participants were recruited and consented following protocols approved by the Institutional Review Board of the National Institute of Mental Health. Inclusion criteria included: (1) being between the ages of 18 and 65 years old at the time of experimentation, (2) a healthy physical examine completed by a nurse practitioner within a year of the study session, and (3) ability to give informed consent. Exclusion criteria included: (1) no previous nor current histories of neurologic or psychiatric disorder, (2) low vision (corrected normal vision was acceptable), and (3) no head injuries (e.g., loss of consciousness for >30 minutes and three or more concussive injuries). Prior to each testing session, a nurse practitioner completed a health exam for each participant, including recording temperature, vitals, and assessment for Covid-19 symptoms.
Afterimage Induction
Afterimages were elicited using an inducer stimulus: a black silhouette image of a human face in frontal view (presentation duration = 4 seconds; visual angle: 4.60 × 8.47 degrees; https://creazilla.com/nodes/2524-face-silhouette; Figure 1C Inducer Stimulus; Supplementary Movie 1). The inducer stimulus resulted in negative afterimages that appeared as white or light grey versions of the inducer and maintained a face-like identity. In pilot testing, it was observed that some participants perceived an instantaneous, illusory crisp white version of the inducer stimulus at the moment of its disappearance. This experience was sometimes confused with the subsequent negative afterimage that was typically delayed from the offset of the inducer, less sharp than the inducer, and lasted for several seconds. To limit the occurrence of this flashbulb-like perceptual effect at the sudden offset of the full contrast inducer, in the first and last second of the inducer presentation, the inducer contrast was gradually ramped up to full contrast and down to no contrast, respectively. Thus, the inducer appeared at full contrast for a total of 2 seconds. In pilot testing (data not shown), this contrast ramping adjustment did not impact the occurrence of the afterimages, while succeed in suppressing the perceived inducer offset flash.
Figure 1. Target perceptual features, session sequence, and perception matching paradigms.

(A) The target image and afterimage perceptual features were: (1) sharpness, (2) contrast, and (3) duration. (B) Participants completed four task phases in the following order: (1) image and (2) afterimage sharpness perception matching and (3) image and (4) afterimage contrast and duration perception matching. (C) The stimuli and controllable images presented in the perception matching tasks. The afterimage perception is depicted as a dashed outline because no image was presented – the afterimage is an illusory visual perception. Depending on the task phase, the controllable image allowed participants to manually adjust its sharpness or contrast. The controllable image is depicted with a hand icon to indicate that participants manually adjusted these images with key presses, although this symbol was not physically present during the task. (D) The main trial events of the image perception matching task (Supplementary Movies 2 and 4). Each trial began with a fixation interval (6–8 seconds [s]). When the image stimulus appeared (4 s) on either the left or right side of the central fixation, participants were instructed to immediately present and adjust the controllable image using key presses to match with the image stimuli according to the target perceptual quality (i.e., sharpness and contrast/duration; see Sharpness and Contrast and Duration Perception Matching Paradigm Methods sections). A subsequent fixation interval (10–12 s) followed the Image & Report stage prior to initiating the next trial. (E) The main trial events of the afterimage perception matching task (Supplementary Movies 3 and 5). Each trial began with a jittered fixation interval (6–8 s). Next, the inducer stimulus was shown (4 s) on either the left or right side of the central fixation and, subsequently, an afterimage might appear. If an afterimage was perceived, participants were instructed to immediately display and adjust the controllable image to match with the target perceptual feature of their afterimage perception (i.e., sharpness and contrast/duration; see Sharpness and Contrast and Duration Perception Matching Paradigm Methods sections). The Afterimage & Report stage completed when the participant no longer perceived their afterimage, and the remaining duration of time (10–12 s) was a fixation interval prior to initiating the next trial.
During initial task instructions, participants were shown the inducer repeatedly to determine their susceptibility for perceiving afterimages. If there was any confusion regarding what parts of their visual experience constituted the afterimage, as some participants were initially naïve to afterimages, clarifying instructions were provided by the experimenter to guide when and what parts of their visual perception following the inducer constituted the afterimage perception.
Image and Afterimage Perceptual Vividness
Participants were asked to report on three target perceptual features that contribute to the overall perceived vividness of conscious vision: (1) sharpness (i.e., crisp versus blurry), (2) contrast (i.e., bright versus dim), and (3) duration (Figure 1A). Sharpness, contrast, and duration are previously interrogated as markers of the vividness of afterimages (e.g.,21,45). Here, participants made judgements on these perceptual features for both image and afterimage perceptions (Figure 1D, E). These perceptual reports were achieved using a paradigm whereby participants adjusted the appearance of an on-screen image – a so-called controllable image (Figure 1C Controllable Images) – to match in real time with the perceived sharpness, contrast, and duration of images and afterimages. Note that the contrast and duration reports were acquired simultaneously (see Contrast and Duration Perception Matching section). The current approach builds on previous methods for reporting on the perceptual features of afterimages (e.g.,24,52,66,80). Before completing the perception matching tasks, participants were administered instructions and a practice session (see Sharpness and Contrast and Duration Perception Matching sections). Subsequently, participants completed the image and afterimage perception matching tasks (Figure 1B, D, E).
Sharpness Perception Matching Paradigm
In the sharpness matching phase, participants were asked to notice and report on the maximum perceived sharpness of images and afterimages. In the image sharpness matching phase, the image stimulus was a white version of the inducer stimulus, otherwise, identical in size and presentation duration as the inducer (Figure 1C Image Stimulus). The image stimuli initially appeared blurrier, then gradually reached a maximum sharpness after 1 second from its onset, maintained this maximum sharpness for 2 seconds, and then gradually became blurrier again before the stimulus entirely disappeared from the screen, 4 seconds after its onset (Supplementary Movie 2). This dynamic of increasing and decreasing sharpness was programmed according to pilot testing (data not shown) that suggested the general trend in the perception of afterimage sharpness was to cycle from blurrier, to sharper, and blurrier again before the afterimage perception disappeared.
The sharpness values applied to the image stimulus ranged from 0 to 25 in increments of 1, each value representing the number of pixels in the radius of a gaussian kernel used to blur the image stimulus (blurred image size = 600 × 800 pixels; gaussian blur; Illustrator, Adobe, Inc.). Therefore, 0 indicated no blurring of the original image stimulus (i.e., maximum sharpness), while 25 indicated maximum blurring. In the analyses and figures (Figure 2C; Figure 3C, D; Figure 4A), the sharpness values were inverted so that 0 pixels indicated the blurriest perception and 25 pixels the sharpest. Inverting the sharpness pixel scale was implemented because it corresponded with the contrast and duration scales where larger numbers indicate more vivid images and afterimages. Thus, all sharpness values and accompanying figures are reported along the inverted pixel scale.
Participants were instructed to report the maximum sharpness of the image stimulus in real time. This was achieved in the following steps within each sharpness matching task trial (Supplementary Movie 2): (1) participants fixated at a central plus sign inside an open circle (1.33 × 1.33 degrees) on a blank grey screen for a jittered pre-stimulus interval (6–8 seconds; Figure 1D Fixation phase). (2) An image stimulus appeared for 4 seconds (Figure 1D Image & Report phase). The image would appear at random, but in equal proportion, either to the left or right of the fixation point along the midline (image stimulus location from central fixation = 5.88 degrees) and among one of three sharpness values: 10, 15, and 20 pixels. (3) When participants saw the image stimulus, they were instructed to immediately select a key to display the controllable image that was previously absent from the screen (Figure 1C Controllable Images - Sharpness). The controllable image appeared at a random initial sharpness value (0–25 pixels) and shown on the opposite side of the screen from where the on-screen stimulus appeared, so not to confuse between the image stimulus that initially appeared on one side of the screen and the subsequent controllable image stimulus that participants voluntarily summoned in the mirrored screen location. Using two dedicated keys, participants manually adjusted the sharpness of the controllable stimulus to match with the perceived maximum sharpness of the image stimulus: one key increasing and the other key decreasing the controllable image sharpness in increments of 1 pixel (Figure 1D Image & Report phase). (4) Once participants completed adjusting the sharpness of the controllable stimulus, they were instructed to press a third key to record their selection. While participants were encouraged to report the maximum sharpness of the image while it was still present on-screen or soon after its disappearance, participants had a minimum of 10 seconds and a maximum of 12 seconds from the image stimulus offset (i.e., 10–12 seconds jittered post-image interval) to continue adjusting the controllable image and make their selection, otherwise, the trial was automatically aborted and no response was logged for that trial (Figure 1D post-Image & Report Fixation phase). Across all trial phases, the participants were instructed to maintain fixation. A total of 20 trials of the image sharpness matching task was completed for each participant.
Participants completed a similar afterimage sharpness perception matching task (Figure 1E; Supplementary Movie 3). Here, the goal was for participants to report the maximum sharpness of their perceived afterimages. The reporting method was identical to that used for the image stimulus (i.e., the use of a controllable image and manually updating its sharpness to match with the perceived afterimage maximum sharpness). The key difference between the image and afterimage sharpness matching task phases was that in the afterimage condition, participants were first shown the inducer stimulus (see Afterimage Induction section; Figure 1E Inducer phase). When the inducer disappeared, the participants might see an afterimage and were instructed to immediately display and adjust the controllable image with two keys to match with their perceived afterimage maximum sharpness in real time (Figure 1E Afterimage & Report phase). The controllable image appeared on the opposite side of the screen from the perceived afterimage while centrally fixating. Participants pressed a third key to record their selection. Participants had a minimum of 10 seconds and a maximum of 12 seconds from the inducer stimulus offset (i.e., 10–12 seconds jittered post-inducer interval) to make their selection, otherwise, the trial was automatically aborted and no response was logged for that trial. When the participant no longer perceived an afterimage, the remaining duration of the jittered post-inducer interval was a passive fixation period prior to initiating the next trial. If participants did not see an afterimage, they were instructed to not press any keys and wait until the next trial began automatically. A total of 30 trials of the afterimage sharpness matching task was completed for each participant.
Contrast and Duration Perception Matching Paradigm
Previous studies have used perceptual cancellation to assess afterimage contrast (i.e., overlaying a physical image over the afterimage location and having participants adjust that physical image until the afterimage percept disappears; e.g.,24). As in the sharpness matching phase, the current method used a side-by-side contrast and duration perception matching procedure. Participants were asked to notice and report on the brightness of images and afterimages overtime (i.e., instead of matching to a single contrast value, participants were asked to follow the change in the image and afterimage contrast throughout its perception). This was achieved by the following steps within each trial (Supplementary Movie 4): (1) participants fixated at a central plus sign inside an open circle (1.33 × 1.33 degrees) on a blank grey screen for a jittered pre-stimulus interval (6–8 seconds; Figure 1D Fixation phase). (2) An image stimulus would appear for 4 seconds (Figure 1D Image & Report phase). The image would appear at random but in equal proportion either to the left or right of the fixation point along the midline (image stimulus location from central fixation = 5.88 degrees). In the first second of the image presentation, the stimulus was shown gradually increasing its contrast to a maximum contrast of 0.25, where 1 is full contrast, and then gradually decreasing its contrast until the stimulus disappeared (i.e., a contrast of 0). The maximum contrast value (0.25) was selected according to pilot testing (data not shown) that suggested this contrast was similar to the maximum brightness of afterimages that appeared following the inducer stimulus. The image maintained this maximum contrast for a varied interval of time (0.5, 1, 1.5, 2, and 2.5 seconds). There were three increasing contrast intervals (1, 1.5, and 2 seconds from the image stimulus onset until the image reached maximum contrast) and three decreasing contrast intervals (2.5, 3, and 3.5 seconds from the image stimulus onset until the image contrast began decreasing until it disappeared at 4 seconds from the initial image stimulus onset). This ramping contrast intervals were selected to approximate the contrast change dynamic of afterimage perceptions reported in pilot testing (data not shown).
(3) Upon perceiving the image, participants were instructed to immediately begin pressing a key that would increase the contrast of the controllable image in increments of 0.025, which was initially set to a contrast of 0 (Figure 1C Controllable Images - Contrast). The controllable image appeared on the opposite side of the screen from the image stimulus. When the image stimulus began to reduce in contrast, participants used a second key to decrease the contrast of the controllable image in increments of 0.025. Participants could also use a third key that would immediately disappear the controllable image from whatever its current contrast value (i.e., set the image contrast to 0), thus offering the option to report the perception of an immediate disappearance rather than a gradual reduction of contrast. Critically, participants were instructed to manipulate the controllable image to match with the image stimulus contrast throughout its presentation, so that at any given moment both the image and controllable image appeared with identical contrast. Thereby, the reported duration of the images was acquired by measuring the length of time participants manipulated the controllable image (i.e., the time when participants first reported a perceived image with greater than 0 contrast and its subsequent disappearance time; see the Statistical Analyses Duration subsection). While participants were encouraged to report in real time with the image presentation, participants had a minimum of 10 seconds and a maximum of 12 seconds after the image stimulus offset (i.e., 10–12 seconds jittered post-image interval) to continue adjusting the controllable image, otherwise, the trial was automatically aborted, and the responses made in the preceding interval were logged for that trial (Figure 1D post-Image & Report Fixation phase). A total of 18 trials of the image contrast and duration matching task was completed for each participant.
Participants completed a similar afterimage contrast and duration perception matching task (Figure 1E; Supplementary Movie 5). The goal was for participants to report the change in contrast overtime of their afterimage perceptions. The reporting method was identical to that used for the image stimulus (i.e., the use of a controllable image and manually updating its contrast to match with the afterimage overtime). The key difference between the image and afterimage contrast matching task phases was that in the afterimage condition, participants were first shown the inducer stimulus (see Afterimage Induction section; Figure 1E Inducer phase). When the inducer disappeared, the participants might see an afterimage and were instructed to immediately display and adjust the controllable image with two keys to match with their perceived afterimage contrast in real time (Figure 1E Afterimage & Report phase). The controllable image appeared on the opposite side of the screen from the perceived afterimage while centrally fixating. Participants had a minimum of 10 seconds and a maximum of 12 seconds from the inducer stimulus offset (i.e., 10–12 seconds jittered post-inducer interval) to report the duration and contrast of their afterimage. When the participant no longer perceived an afterimage, the remaining duration of the jittered post-inducer interval was a passive fixation period prior to initiating the next trial. If participants did not see an afterimage, they were instructed to not press any keys and wait until the next trial began automatically. A total of 60 trials of the afterimage contrast and duration perception matching task was completed for each participant.
Visual Imagery Vividness
Acquiring details about imagery perceptual vividness is challenging due to the notorious problem of introspection (i.e., the inaccessibility of subjective conscious perception except by the conscious experiencer). Recent attempts of developing covert markers of imagery vividness (e.g., pupil size) are promising and validate self-report metrics of imagery88. In the current investigation, visual imagery vividness was acquired with the 16-item, self-reported Vividness of Visual Imagery Questionnaire (VVIQ)89. The questionnaire asks participants to imagine people, objects, and scenes and then introspect on how vivid that imagined content appears in their visual imagery on a 5-point scale between “no image at all” to “perfectly clear and as vivid as normal vision”. Participants were instructed to complete the VVIQ with their eyes open and were given no time constraint in completing the questionnaire. The VVIQ was displayed on a computer monitor and participants used a mouse click to select their answers for each questionnaire item. The VVIQ was administered at either the beginning or end of the study session.
Equipment, Software, and Facility
The behavioral study was completed in a single 2-hour study session in a windowless behavioral testing room. The room lighting was set to a consistent brightness level for all participants. The experimenter was present in the testing room but positioned out of sight of the participant to monitor behavior and deliver task instructions. The behavioral paradigm was coded in Python and run with PsychoPy (v2022.2.4; Open Science Tools Ltd.) on a behavioral laptop (MacBook Pro; 13-inch; 2560 × 1600 pixels, 2019; Mac OS Catalina v10.15.7; Apple, Inc)90. The behavioral laptop monitor was mirrored by DVI cable to a VIEWPixx monitor (1920 × 1200 pixels; VPixx Technologies, Inc.) on which the participants viewed the experimental paradigms and the VVIQ. The participants were positioned approximately 56cm from the center of the display monitor. The viewing distance was fixed using a table mounted head-chin rest. All participants used their right hand (regardless of handedness) to make key presses during the task with a keyboard positioned on a table in front of the participant.
Statistical Analyses
All analyses were completed in MATLAB v2022b (MathWorks, Inc.) and Prism v10 (Graphpad, Inc.). Figures where generated and edited in Prism (Graphpad, Inc.) and Illustrator (Adobe, Inc.).
Afterimage Perception Rate
Afterimage perception rate measures how often afterimages were perceived by each participant following the inducer stimulus. The perception rate was calculated by finding the percentage of inducer presentations that an afterimage was perceived across the sharpness and contrast and duration perception matching tasks – a total of 90 trials (i.e., the number of perceived afterimage trials in the sharpness matching task plus the number of perceived afterimage trials in the contrast and duration matching task divided by the total number of trials across all tasks). Perception rate values were multiplied by 100 to convert from units of fraction to percentage.
VVIQ Score
The VVIQ score for each participant was calculated by taking the sum of all scores across the questionnaire items. Each item was scored on a scale from 1 (no image) to 5 (perfectly clear). Therefore, the minimum and maximum VVIQ score was 16 and 80, respectively, where larger values indicate more vivid visual imagery.
Sharpness
Calculating reported sharpness.
Participants reported the perceived maximum sharpness of images and afterimages (see Sharpness Perception Matching Paradigm section). The participant image and afterimage sharpness values were calculated by averaging all trial sharpness values within participant and image and afterimage sharpness matching tasks. Trials without a sharpness value (e.g., response timeout or afterimage was not perceived) were excluded from consideration in calculating the participant mean sharpness value. The sharpness value scale was inverted, so that larger values correspond with a sharper perception. This scale inversion was achieved by taking the absolute value of the participant mean sharpness value minus the maximum sharpness value (25; i.e., the largest pixel radius of the blurring gaussian kernel).
Calculating reported image sharpness accuracy.
Participant reported image sharpness accuracy was calculated by comparing the reported maximum sharpness with the true image maximum sharpness on a trial-by-trial basis. This was achieved by subtracting the reported maximum image sharpness from the known image maximum sharpness (10, 15, or 20 pixels) across trials. Next, all subtracted or relative sharpness trial values were averaged within participant. A positive relative sharpness value indicates the image was reported as sharper than its true maximum sharpness, while a negative relative sharpness indicates the image was reported as blurrier than its true maximum sharpness, where a value of 0 indicates a perfect match between the participant reports and the true image maximum sharpness (Figure 2C Relative). To statistically test the reporting accuracy of the image maximum sharpness, a Wilcoxon Rank Sum test (p < 0.05) was applied on the relative sharpness values and tested against 0. If the relative image sharpness is found no different from 0, then participants were accurate in reporting on the maximum sharpness of the image stimulus.
Calculating the correlation between VVIQ score and sharpness.
The relationship between VVIQ scores and the reported image and afterimage maximum sharpness were statistically tested using a two-tailed, Pearson correlation test (p < 0.05; Figure 3C, D). Correlation analyses were applied in two comparisons: (1) VVIQ score versus image sharpness and (2) VVIQ score versus afterimage sharpness. A linear regression fit was applied to model the trend and 95% confidence interval for each of the comparisons.
Image reconstruction of the afterimage sharpness.
The perceived maximum sharpness of the afterimage was reconstructed for low (24) and high (80) VVIQ scores, representing the minimum and maximum VVIQ score recorded among participants (Figure 2B). Reconstruction was achieved by finding the sharpness value for the low and high VVIQ scores along the VVIQ score versus afterimage sharpness linear regression fit trend line and creating images (gaussian blur; Illustrator; Adobe, Inc.) that matched with these estimated sharpness values (Figure 4A, C).
Contrast
Calculating reported contrast.
Participants reported the perceived contrast of images and afterimages overtime (see Contrast and Duration Perception Matching Paradigm section). The participant image and afterimage contrast values were calculated by finding the maximum contrast value reported in each trial and averaging the maximum contrast values across trials within participant and image and afterimage contrast matching tasks. Any trial with less than two reported contrast time points or a maximum contrast value of 0 (e.g., an afterimage was not perceived) was ignored from calculating the participant image and afterimage contrast value.
Calculating reported image contrast accuracy.
Participant reported image contrast accuracy was calculated by subtracting the reported maximum image contrast values from the known image maximum contrast value (0.25) across trials. Next, all subtracted or relative contrast trial values were averaged within participant. A positive relative contrast indicates the image was reported as brighter than its true maximum contrast, while a negative relative contrast indicates the image was reported as dimmer than its true maximum contrast, where a value of 0 indicates a perfect match between the participant reports and the true image maximum contrast (Figure 2D Relative). To statistically test the reporting accuracy of the image maximum contrast, a Wilcoxon Rank Sum test (p < 0.05) was applied on the relative contrast values and tested against 0. If the relative image contrast is found no different from 0, then participants were accurate in reporting on the contrast of the image stimulus.
Calculating the correlation between VVIQ score and contrast.
The relationship between VVIQ scores and the reported image and afterimage maximum contrast were statistically tested using a two-tailed, Pearson correlation test (p < 0.05; Figure 3A, B). Correlation analyses were applied in two comparisons: (1) VVIQ score versus image contrast and (2) VVIQ score versus afterimage contrast. A linear regression fit was applied to model the trend and 95% confidence interval for each of the comparisons.
Image reconstruction of the afterimage contrast.
The perceived maximum contrast of the afterimage was reconstructed for low (24) and high (80) VVIQ scores, representing the minimum and maximum VVIQ score recorded among participants (Figure 2B). Reconstruction was achieved by finding the contrast value for the low and high VVIQ scores along the VVIQ score versus afterimage contrast linear regression fit trend line and creating images (Illustrator; Adobe, Inc.) that matched with these estimated contrast values (Figure 4B, C).
Duration
Calculating reported duration.
Image and afterimage durations were calculated from the contrast and duration perception matching task (see Contrast and Duration Perception Matching Paradigm section). Contrast and duration perception matching task trials were considered valid by the same criteria for calculating the reported maximum contrast of images and afterimages (see Contrast Statistical Analyses section). The duration was measured as the time between the initial and final key press participants made to adjust the controllable image to match with the perceived contrast of the images and afterimages overtime or when the participant reported the image or afterimage had a contrast of zero, whichever occurred first.
Calculating reported image duration accuracy.
Participant reported image duration accuracy was calculated by subtracting the reported image duration across trials within participant from the known image duration (4 seconds). A positive relative duration indicates participants reported on average that the image was presented longer than its true duration, while a negative relative duration suggests that participants reported on average that the image was briefer (Figure 2E Relative). A value of 0 indicates a perfect match between the participant reports and the true image duration. To statistically test how accurate participants were in reporting the image duration, a Wilcoxon Rank Sum test (p < 0.05) was applied on the relative duration values and tested against 0. If the relative image duration is found no different from 0, then participants were accurate in reporting on the duration of the image stimulus.
Calculating the correlation between VVIQ score and duration.
The relationship between VVIQ scores and the reported image and afterimage duration were statistically tested using a two-tailed, Pearson correlation test (p < 0.05; Figure 3E, F). Correlation analyses were applied in two comparisons: (1) VVIQ score versus image duration and (2) VVIQ score versus afterimage duration. A linear regression fit was applied to model the trend and 95% confidence interval for each of the comparisons.
Supplementary Material
Acknowledgements
This research was made possible by the support of the National Institute of Mental Health Intramural Research Program (ZIAMH002783). The study was completed in compliance with the National Institutes of Health Clinical Center protocol ID 93-M-0170 (ClinicalTrials.gov ID: NCT00001360). We thank members of the Section on Functional Imaging Methods for their constructive feedback.
Data and Code Availability
All data and scripts will be made available prior to publication.
References
- 1.Rosenblith W. A., Miller G. A., Egan J. P., Hirsh I. J. & Thomas G. J. An Auditory Afterimage? Science 106, 333–335 (1947). 10.1126/science.106.2754.333 [DOI] [PubMed] [Google Scholar]
- 2.Wiegrebe L., Kössl M. & Schmidt S. Auditory sensitization during the perception of acoustical negative afterimages: Analogies to visual processing? Naturwissenschaften 82, 387–389 (1995). 10.1007/BF01134569 [DOI] [PubMed] [Google Scholar]
- 3.Zwicker E. “Negative afterimage” in hearing. The Journal of the Acoustical Society of America 36, 2413–2415 (1964). 10.1121/1.1919373 [DOI] [PubMed] [Google Scholar]
- 4.Williams J. L. Evidence of complementary afterimages in the pigeon. J Exp Anal Behav 21, 421–424 (1974). 10.1901/jeab.1974.21-421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Duysens J., Orban G. A., Cremieux J. & Maes H. Visual cortical correlates of visible persistence. Vision Res 25, 171–178 (1985). 10.1016/0042-6989(85)90110-5 [DOI] [PubMed] [Google Scholar]
- 6.McLelland D., Ahmed B. & Bair W. Responses to static visual images in macaque lateral geniculate nucleus: Implications for adaptation, negative afterimages, and visual fading. J Neurosci 29, 8996–9001 (2009). 10.1523/JNEUROSCI.0467-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Goethe J. W. v. Theory of Colours. (The MIT Press, 1970). [Google Scholar]
- 8.McCollough C. Color Adaptation of Edge-Detectors in the Human Visual System. Science 149, 1115–1116 (1965). 10.1126/science.149.3688.1115 [DOI] [PubMed] [Google Scholar]
- 9.Kanizsa G. Subjective Contours. Sci Am 234, 48–53 (1976). [DOI] [PubMed] [Google Scholar]
- 10.Favreau O. E. & Corballis M. C. Negative aftereffects in visual perception. Sci Am 235, 42–48 (1976). 10.1038/scientificamerican1276-42 [DOI] [PubMed] [Google Scholar]
- 11.Kirschfeld K. Afterimages: A tool for defining the neural correlates of visual consciousness. Conscious Cogn 8, 462–483 (1999). 10.1006/ccog.1999.0388 [DOI] [PubMed] [Google Scholar]
- 12.Anstis S., Geier J. & Hudak M. Afterimages from unseen stimuli. Iperception 3, 499–502 (2012). 10.1068/i0547sas [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tsuchiya N. & Koch C. Continuous flash suppression reduces negative afterimages. Nat Neurosci 8, 1096–1101 (2005). 10.1038/nn1500 [DOI] [PubMed] [Google Scholar]
- 14.Craik K. J. W. Origin of visual after-images. Nature 145, 512 (1940). [Google Scholar]
- 15.Weiskrantz L., Cowey A. & Hodinott-Hill I. Prime-sight in a blindsight subject. Nat Neurosci 5, 101–102 (2002). 10.1038/nn793 [DOI] [PubMed] [Google Scholar]
- 16.Mazzi C., Savazzi S. & Silvanto J. On the “blindness” of blindsight: What is the evidence for phenomenal awareness in the absence of primary visual cortex (V1)? Neuropsychologia 128, 103–108 (2019). 10.1016/j.neuropsychologia.2017.10.029 [DOI] [PubMed] [Google Scholar]
- 17.Oswald I. After-images from retina and brain. Q J Exp Psychol 9 (1957). 10.1080/17470215708416225 [DOI] [Google Scholar]
- 18.Helmholtz H. v. in Helmholtz on perception: Its physiology and development (eds Warren R. M. & Warren R. P.) (John Wiley + Sons, 1968). [Google Scholar]
- 19.Davies P. Visual scotoma and visual afterimages: Some evidence that the perceived visual afterimage may not be a purely retinal phenomenon: A single case study. Percept Mot Skills 81, 849–850 (1995). 10.2466/pms.1995.81.3.849 [DOI] [PubMed] [Google Scholar]
- 20.Rushton W. A. H. & Henry G. H. Bleaching and regeneration of cone pigments in man. Vision Res 8, 617–631 (1968). 10.1016/0042-6989(68)90040-0 [DOI] [PubMed] [Google Scholar]
- 21.Brindley G. S. Two new properties of foveal after-images and a photochemical hypothesis to explain them. J Physiol 164, 168–179 (1962). 10.1113/jphysiol.1962.sp007011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.MacLeod D. I. A. & Hayhoe M. Rod origin of prolonged afterimages. Science 185, 1171–1172 (1974). 10.1126/science.185.4157.1171 [DOI] [PubMed] [Google Scholar]
- 23.Hurvich L. M. & Jameson D. An opponent-process theory of color vision. Psychol Rev 64, 384–404 (1957). 10.1037/h0041403 [DOI] [PubMed] [Google Scholar]
- 24.Georgeson M. A. & Turner R. S. E. Afterimages of sinusoidal, square-wave and compound gratings. Vision Res 25, 1709–1720 (1985). 10.1016/0042-6989(85)90143-9 [DOI] [PubMed] [Google Scholar]
- 25.Miller N. D. Positive afterimage following brief high-intensity flashes. J Opt Soc Am 56, 802–806 (1966). 10.1364/josa.56.000802 [DOI] [PubMed] [Google Scholar]
- 26.Feinbloom W. A Quantitative Study of the Visual After-image. Columbia University; (1938). [Google Scholar]
- 27.Magnussen S., Spillmann L., Stürzel F. & Werner J. S. Unveiling the foveal blue scotoma through an afterimage. Vision Research 44, 377–383 (2004). 10.1016/j.visres.2003.09.023 [DOI] [PubMed] [Google Scholar]
- 28.Zaidi Q., Ennis R., Cao D. & Lee B. Neural locus of color afterimages. Curr Biol 22, 220–224 (2012). 10.1016/j.cub.2011.12.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Marriott F. H. Thresholds for negative after-images. J Physiol 180, 888–892 (1965). 10.1113/jphysiol.1965.sp007738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Long G. M. & Kling S. C. Positive and negative afterimages from brief target gratings. Vision Res 23, 959–963 (1983). 10.1016/0042-6989(83)90005-6 [DOI] [PubMed] [Google Scholar]
- 31.Shimojo S., Kamitani Y. & Nishida S. Afterimage of perceptually filled-in surface. Science 293, 1677–1680 (2001). 10.1126/science.1060161 [DOI] [PubMed] [Google Scholar]
- 32.Hamburger K., Geremek A. & Spillmann L. Perceptual filling-in of negative coloured afterimages. Perception 41, 50–56 (2012). 10.1068/p7066 [DOI] [PubMed] [Google Scholar]
- 33.van Lier R., Vergeer M. & Anstis S. Filling-in afterimage colors between the lines. Curr Biol 19, R323–324 (2009). 10.1016/j.cub.2009.03.010 [DOI] [PubMed] [Google Scholar]
- 34.Downey J. E. An experiment on getting an after-image from a mental image. Psychological Review 8, 42–55 (1901). [Google Scholar]
- 35.Weiskrantz L. An unusual case of after-imagery following fixation of an “imaginary” visual pattern. Q J Exp Psychol 2, 170–175 (1950). [Google Scholar]
- 36.Barber T. X. The afterimages of “hallucinated” and “imagined” colors. J Abnorm Psychol 59, 136–139 (1959). 10.1037/h0047334 [DOI] [PubMed] [Google Scholar]
- 37.Davies P. Conditioned after-images. I. Br J Psychol 65, 191–204 (1974). 10.1111/j.2044-8295.1974.tb01394.x [DOI] [PubMed] [Google Scholar]
- 38.Day R. H. On interocular transfer and the central origin of visual after-effects. Am J Psychol 71, 784–790 (1958). 10.2307/1420346 [DOI] [PubMed] [Google Scholar]
- 39.Gerling J. & Spillmann L. Duration of visual afterimages on modulated backgrounds: postreceptoral processes. Vision Res 27, 521–527 (1987). 10.1016/0042-6989(87)90038-1 [DOI] [PubMed] [Google Scholar]
- 40.Wright J. M. v. A note on interocular transfer and the colour of visual after-images. Scandinavian Journal of Psychology 4 (1963). 10.1111/j.1467-9450.1963.tb01329.x [DOI] [Google Scholar]
- 41.Gilroy L. A. & Blake R. The interaction between binocular rivalry and negative afterimages. Curr Biol 15, 1740–1744 (2005). 10.1016/j.cub.2005.08.045 [DOI] [PubMed] [Google Scholar]
- 42.Shevell S. K., St Clair R. & Hong S. W. Misbinding of color to form in afterimages. Vis Neurosci 25, 355–360 (2008). 10.1017/S0952523808080085 [DOI] [PubMed] [Google Scholar]
- 43.Davies P. The role of central processes in the perception of visual after-images. Br J Psychol 64, 325–338 (1973). 10.1111/j.2044-8295.1973.tb01358.x [DOI] [PubMed] [Google Scholar]
- 44.Dong B., Holm L. & Bao M. Cortical mechanisms for afterimage formation: evidence from interocular grouping. Sci Rep 7, 41101 (2017). 10.1038/srep41101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cerf-Beare A. Pattern and orientation effects on afterimage duration. Perception 13, 443–453 (1984). 10.1068/p130443 [DOI] [PubMed] [Google Scholar]
- 46.van Boxtel J. J. A., Tsuchiya N. & Koch C. Opposing effects of attention and consciousness on afterimages. Proc Natl Acad Sci 107, 8883–8888 (2010). 10.1073/pnas.0913292107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Brascamp J. W., van Boxtel J. J. A., Knapen T. H. J. & Blake R. A dissociation of attention and awareness in phase-sensitive but not phase-insensitive visual channels. J Cogn Neurosci 22, 2326–2344 (2010). 10.1162/jocn.2009.21397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Shimizu M. & Kimura E. Afterimage duration depends on how deeply invisible stimuli were suppressed. J Vis 23, 1 (2023). 10.1167/jov.23.8.1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Baijal S. & Srinivasan N. Types of attention matter for awareness: a study with color afterimages. Conscious Cogn 18, 1039–1048 (2009). 10.1016/j.concog.2009.09.002 [DOI] [PubMed] [Google Scholar]
- 50.Suzuki S. & Grabowecky M. Attention during adaptation weakens negative afterimages. J Exp Psychol Hum Percept Perform 29, 793–807 (2003). 10.1037/0096-1523.29.4.793 [DOI] [PubMed] [Google Scholar]
- 51.Utz S. & Carbon C. C. Afterimages are biased by top-down information. Perception 44, 1263–1274 (2015). 10.1177/0301006615596900 [DOI] [PubMed] [Google Scholar]
- 52.Lupyan G. Object knowledge changes visual appearance: Semantic effects on color afterimages. Acta Psychol (Amst) 161, 117–130 (2015). 10.1016/j.actpsy.2015.08.006 [DOI] [PubMed] [Google Scholar]
- 53.Hayhoe M. M. & Williams D. R. Disappearance of afterimages at ‘impossible’ locations in space. Perception 13, 455–459 (1984). 10.1068/p130455 [DOI] [PubMed] [Google Scholar]
- 54.Daw N. W. Why after-images are not seen in normal circumstances. Nature 196, 1143–1145 (1962). 10.1038/1961143a0 [DOI] [PubMed] [Google Scholar]
- 55.On Z. X. & van Boxtel J. J. A. The role of transparency cues in afterimage color perception. Sci Rep 7, 9183 (2017). 10.1038/s41598-017-09186-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sperandio I., Lak A. & Goodale M. A. Afterimage size is modulated by size-contrast illusions. J Vis 12 (2012). 10.1167/12.2.18 [DOI] [PubMed] [Google Scholar]
- 57.Qian J., Liu S. & Lei Q. Illusory Distance Modulates Perceived Size of Afterimage despite the Disappearance of Depth Cues. PLoS One 11, e0159228 (2016). 10.1371/journal.pone.0159228 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Dwyer J., Ashton R. & Broerse J. Emmert’s law in the Ames room. Perception 19, 35–41 (1990). 10.1068/p190035 [DOI] [PubMed] [Google Scholar]
- 59.Emmert E. Grössenverhältnisse der Nachbilder. Klinische Monatsblätter für Augenheilkunde 19, 443–450 (1881). [Google Scholar]
- 60.Bonnet C. & Pouthas V. Apparent size and duration of a movement after-effect. Q J Exp Psychol 24, 275–281 (1972). 10.1080/14640747208400281 [DOI] [PubMed] [Google Scholar]
- 61.Sperandio I., Chouinard P. A. & Goodale M. A. Retinotopic activity in V1 reflects the perceived and not the retinal size of an afterimage. Nature Neuroscience 15, 540–542 (2012). 10.1038/nn.3069 [DOI] [PubMed] [Google Scholar]
- 62.O’Halloran W. J. & Weintraub D. J. Delboeuf “illusions” of circle size induced via photoflashgenerated afterimages. Perception & Psychophysics 22, 171–176 (1977). 10.3758/BF03198751 [DOI] [Google Scholar]
- 63.Broerse J., Ashton R. & Shaw C. The apparent shape of afterimages in the Ames room. Perception 21, 261–268 (1992). 10.1068/p210261 [DOI] [PubMed] [Google Scholar]
- 64.Schwarzkopf D. S., Song C. & Rees G. The surface area of human V1 predicts the subjective experience of object size. Nat Neurosci 14, 28–30 (2011). 10.1038/nn.2706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ernst M. O. & Bülthoff H. H. Merging the senses into a robust percept. Trends Cogn Sci 8, 162–169 (2004). 10.1016/j.tics.2004.02.002 [DOI] [PubMed] [Google Scholar]
- 66.Anderson R. H. & Deffenbacher K. A. Effect of sound stimulation on visual afterimages. Percept Mot Skills 32, 343–346 (1971). 10.2466/pms.1971.32.2.343 [DOI] [PubMed] [Google Scholar]
- 67.Gregory R. L., Wallace J. G. & Campbell F. W. Changes in size and shape of visual afterimages observed in complete darkness during changes of position in space. Q J Exp Psychol 11, 1–64 (1959). 10.1080/17470215908416288 [DOI] [Google Scholar]
- 68.Davies P. Effects of movements upon the appearance and duration of a prolonged visual afterimage: 1. Changes arising from the movement of a portion of the body incorporated in the afterimaged scene. Perception 2, 147–153 (1973). 10.1068/p020147 [DOI] [Google Scholar]
- 69.Hogendoorn H., Kammers M. P. M., Carlson T. A. & Verstraten F. A. J. Being in the dark about your hand: Resolution of visuo-proprioceptive conflict by disowning visible limbs. Neuropsychologia 47, 2698–2703 (2009). 10.1016/j.neuropsychologia.2009.05.014 [DOI] [PubMed] [Google Scholar]
- 70.Stone B. W. & Tinker J. Multisensory Tracking of Objects in Darkness: Capture of Positive Afterimages by the Tactile and Proprioceptive Senses. PLoS One 11, e0150714 (2016). 10.1371/journal.pone.0150714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ingle D. Central visual persistences: I. Visual and kinesthetic interactions. Perception 34, 1135–1151 (2005). 10.1068/p5408 [DOI] [PubMed] [Google Scholar]
- 72.Gersztenkorn D. & Lee A. G. Palinopsia revamped: A systematic review of the literature. Surv Ophthalmol 60, 1–35 (2015). 10.1016/j.survophthal.2014.06.003 [DOI] [PubMed] [Google Scholar]
- 73.Schimansky S., Bennetto L. & Harrison R. Palinopsia. Pract Neurol 22, 392–395 (2022). 10.1136/practneurol-2022-003347 [DOI] [PubMed] [Google Scholar]
- 74.Critchley M. Types of visual perseveration: “Paliopsia” and “illusory visual spread”. Brain 74, 267–299 (1951). 10.1093/brain/74.3.267 [DOI] [PubMed] [Google Scholar]
- 75.Kawasaki A. & Purvin V. Persistent palinopsia following ingestion of lysergic acid diethylamide (LSD). Arch Ophthalmol 114, 47–50 (1996). 10.1001/archopht.1996.01100130045007 [DOI] [PubMed] [Google Scholar]
- 76.Crutch S. J. et al. Abnormal visual phenomena in posterior cortical atrophy. Neurocase 17, 160–177 (2011). 10.1080/13554794.2010.504729 [DOI] [PubMed] [Google Scholar]
- 77.Chan D., Crutch S. J. & Warrington E. K. A disorder of colour perception associated with abnormal colour after-images: a defect of the primary visual cortex. J Neurol Neurosurg Psychiatry 71, 515–517 (2001). 10.1136/jnnp.71.4.515 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hoke E. S., Ross B. & Hoke M. Auditory afterimage: Tonotopic representation in the auditory cortex. Neuroreport 9, 3065–3068 (1998). 10.1097/00001756-199809140-00027 [DOI] [PubMed] [Google Scholar]
- 79.Engelen T., Rademaker R. L. & Sack A. T. Reduced Fading of Visual Afterimages after Transcranial Magnetic Stimulation over Early Visual Cortex. J Cogn Neurosci 31, 1368–1379 (2019). 10.1162/jocn_a_01415 [DOI] [PubMed] [Google Scholar]
- 80.Virsu V. & Laurinen P. Long-lasting afterimages caused by neural adaptation. Vision Res 17, 853–860 (1977). 10.1016/0042-6989(77)90129-8 [DOI] [PubMed] [Google Scholar]
- 81.James W. The Principles of Psychology. Vol. II (Henry Holt and Company, 1890). [Google Scholar]
- 82.Dance C. J., Ipser A. & Simner J. The prevalence of aphantasia (imagery weakness) in the general population. Conscious Cogn 97, 103243 (2022). 10.1016/j.concog.2021.103243 [DOI] [PubMed] [Google Scholar]
- 83.Dance C. J., Ward J. & Simmer J. What is the link between mental imagery and sensory sensitivity? Insights from aphantasia. Perception 50, 757–782 (2021). 10.1177/03010066211042186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sparing R. et al. Visual cortex excitability increases during visual mental imagery—a TMS study in healthy human subjects. Brain Res 938, 92–97 (2002). 10.1016/s0006-8993(02)02478-2 [DOI] [PubMed] [Google Scholar]
- 85.Hartman A. M. The apparent size of after-images in delusional and non-delusional schizophrenics. Am J Psychol 75, 587–595 (1962). 10.2307/1420281 [DOI] [PubMed] [Google Scholar]
- 86.Thakkar K. N., Silverstein S. M. & Brascamp J. W. A review of visual aftereffects in schizophrenia. Neurosci Biobehav Rev 101, 68–77 (2019). 10.1016/j.neubiorev.2019.03.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Khadjevand F., Shahzadi S. & Abbassian A. Reduction of negative afterimage duration in Parkinson’s disease patients: A possible role for dopaminergic deficiency in the retinal Interplexiform cell layer. Vision Res 50, 279–283 (2010). 10.1016/j.visres.2009.09.026 [DOI] [PubMed] [Google Scholar]
- 88.Kay L., Keogh R., Andrillon T. & Pearson J. The pupillary light response as a physiological index of aphantasia, sensory and phenomenological imagery strength. Elife 11 (2022). 10.7554/eLife.72484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Marks D. F. Visual imagery differences in the recall of pictures. Br J Psychol 64, 17–24 (1973). 10.1111/j.2044-8295.1973.tb01322.x [DOI] [PubMed] [Google Scholar]
- 90.Peirce J. W. PsychoPy—Psychophysics software in Python. J Neurosci Methods 162, 8–13 (2007). 10.1016/j.jneumeth.2006.11.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All data and scripts will be made available prior to publication.