

Abstract
Given the key roles of the cerebellum in motor, cognitive, and affective operations and given the decline of brain functions with ageing, cerebellar circuitry is attracting the attention of the scientific community. The cerebellum plays a key role in timing aspects of both motor and cognitive operations, including for complex tasks such as spatial navigation. Anatomically, the cerebellum is connected with the basal ganglia via disynaptic loops, and it receives inputs from nearly every region in the cerebral cortex. The current leading hypothesis is that the cerebellum builds internal models and facilitates automatic behaviors through multiple interactions with the cerebral cortex, basal ganglia and spinal cord. The cerebellum undergoes structural and functional changes with ageing, being involved in mobility frailty and related cognitive impairment as observed in the physio-cognitive decline syndrome (PCDS) affecting older, functionally-preserved adults who show slowness and/or weakness. Reductions in cerebellar volume accompany ageing and are at least correlated with cognitive decline. There is a strongly negative correlation between cerebellar volume and age in cross-sectional studies, often mirrored by a reduced performance in motor tasks. Still, predictive motor timing scores remain stable over various age groups despite marked cerebellar atrophy. The cerebello-frontal network could play a significant role in processing speed and impaired cerebellar function due to ageing might be compensated by increasing frontal activity to optimize processing speed in the elderly. For cognitive operations, decreased functional connectivity of the default mode network (DMN) is correlated with lower performances. Neuroimaging studies highlight that the cerebellum might be involved in the cognitive decline occurring in Alzheimer’s disease (AD), independently of contributions of the cerebral cortex. Grey matter volume loss in AD is distinct from that seen in normal ageing, occurring initially in cerebellar posterior lobe regions, and is associated with neuronal, synaptic and beta-amyloid neuropathology. Regarding depression, structural imaging studies have identified a relationship between depressive symptoms and cerebellar gray matter volume. In particular, major depressive disorder (MDD) and higher depressive symptom burden are associated with smaller gray matter volumes in the total cerebellum as well as the posterior cerebellum, vermis, and posterior Crus I. From the genetic/epigenetic standpoint, prominent DNA methylation changes in the cerebellum with ageing are both in the form of hypo- and hypermethylation, and the presumably increased/decreased expression of certain genes might impact on motor coordination. Training influences motor skills and lifelong practice might contribute to structural maintenance of the cerebellum in old age, reducing loss of grey matter volume and therefore contributing to the maintenance of cerebellar reserve. Non-invasive cerebellar stimulation techniques are increasingly being applied to enhance cerebellar functions related to motor, cognitive and affective operations. They might enhance cerebellar reserve in the elderly. In conclusion, macroscopic and microscopic changes occur in the cerebellum during the lifespan, with changes in structural and functional connectivity with both the cerebral cortex and basal ganglia. With the ageing of the population and the impact of ageing on quality of life, the panel of experts considers that there is a huge need to clarify how the effects of ageing on the cerebellar circuitry modify specific motor, cognitive and affective operations both in normal subjects and in brain disorders such as AD or MDD, with the goal of preventing symptoms or improving the motor, cognitive and affective symptoms.
Keywords: cerebellum, ageing, motor, cognitive, affective, Alzheimer’s disease
1. Introduction (Mario Manto)
The cerebellum is a key actor for motor control, cognitive operations and affective regulation [1]. Its contributions extend to emotional control and social interactions [2]. Cerebellum contributes greatly to timing aspects of both motor and cognitive operations, including for complex tasks, in order to ensure coordination of our perception and interaction with the surrounding environment [3]. Anatomically, the cerebellum is connected with the basal ganglia and it receives inputs from nearly every region in the cerebral cortex [4]. The current leading hypothesis is that the cerebellum builds internal models and assists automatic behaviors through multiple interactions with the cerebral cortex, basal ganglia and spinal cord and thanks to numerous neuronal loops involving the cerebellar cortex and nuclei [5].
With the increasingly ageing population and the expected rise of life expectancy of the global population in the next decades, the understanding of the neurological changes occurring with ageing becomes critical for the scientific community [6]. This is relevant not only for the appraisal of physiological ageing, but also for elucidation of the pathological changes occurring in neurodegenerative disorders such as Alzheimer’s disease (AD), for which age is a risk factor [7]. Indeed, recent findings have challenged the notion that the cerebellum is unaffected by AD [8]. Older patients are particularly prone to cognitive decline, depression and to impaired motor control, with obvious consequences on quality of life. How ageing impacts the main site of concentration of neurons in the brain (the cerebellum contains more than 60% of the neurons of the brain) is thus of major importance, especially since the cerebellum is one of the structures of the brain that shows substantial loss of neurons with age [1].
This Consensus Paper aims to provide an overview on the current knowledge on the effects of ageing on cerebellar circuitry and cerebellar functions, attempting to clarify the roles of the cerebellum in motor, cognitive, and affective operations during ageing. The article gathers the viewpoints of a group of established neuroscientists specializing in various areas of cerebellar function. Structural and functional changes of the cerebellum are discussed. The complex concept emerging from this consensus paper creates a framework for further research, enriching our understanding of the effects of ageing on the cerebellum, with the ultimate goal of improving the quality of life or our patients. Although we are well aware that a final consensus cannot be reached now and that studies are ongoing, we strongly believe that the ideas presented here will help address the consequences of ageing upon motor, cognitive and affective brain functions mediated by the cerebellum.
2. Effects of ageing on behavioural performance during predictive motor timing (Pavel Filip, Martin Bareš)
Our ability to perceive time is an element essential to ensure synchrony and reliable representation of our environment, both at conscious and subconscious levels. Recent research advances in this field point towards distributed timing models, which derive temporal information from coincidental activation of various neural nodes [9]. Across this wide range of models, the representation of time is seen as an integral part of complex networks, which also encode other stimulus properties [10–11]. The neuroanatomical nodes most relevant for time perception are continually being revealed by studying the characteristic behavioural patterns or neuronal activity in both healthy individuals [12] and specific patient populations [13–14]. Complex, overlapping networks synthesizing information from multiple modalities have been proposed, involving the basal ganglia, cerebellum, posterior parietal cortex and frontal cortex, even though the precise relevance and data processing characteristics of these structures extracting temporal parameters from relevant inputs are a matter of continuous debate. Multiple research approaches have repeatedly confirmed the pivotal role of the cerebellum in predictive temporal processing, somewhat in contrast to retrospective timing tasks, for which the basal ganglia are the major driver [12].
Given this scattered, network-based nature of temporal estimation with the cerebellum in the central position, it is expected that ageing and its wide-spread brain alterations would exert substantial effects on time processing. Modern theories describing age-related performance decline mostly share the view that our brains counteract neural insults accumulated during normal ageing by continuous functional reorganization. Brain activation patterns in older adults have indeed repeatedly been described as heightened, especially in the prefrontal cortices [15–16]. A landmark study in this area has compared these age-related responses to previously described processes of a similar nature—neurocognitive scaffolding, which is generally detected during the development, learning and acquisition of new abilities in youth [17]. In its simplest form, scaffolding is envisaged as selective optimisation of the initially dispersed networks suboptimal in their performance and efficiency to the most specific and honed circuits. Ageing, accompanied by gradual alterations induced by pathological processes, is then seen as a factor hindering and diminishing the performance of these optimized pathways. However, it is presumed that our brains recruit the very same processes used in their development—re-erection of new scaffolds to compensate for lost or damaged networks.
The cerebellum holds an ambiguous position in this matter. The strongly negative correlations of cerebellar volume with age in cross-sectional studies, with accelerating rates of decline in increasingly older individuals [18], are often mirrored in reduced performance in motor tasks dependent on the cerebellum [19]. Nonetheless, this is in stark contrast with the confirmed slower genetic and epigenetic ageing of the cerebellum [20] (see also section 12). Not only does the cerebellar DNA appear to be more stable from the epigenetic point of view [21], but the cerebellum has also been shown to have fewer age-related deletions in mitochondrial DNA than the cortex [22], possibly due to lower metabolic activity in the cerebellum. Furthermore, performance decrease seems to be far from a uniform finding—predictive motor timing scores have been shown to remain stable over various age groups despite marked cerebellar atrophy exceeding the rate detected in the supratentorial structures in the very same cohort [23]. This rather counter-intuitive combination of structural changes and stable performance has been hypothesized to be the result of the previously described neural scaffolding—increased age in these subjects has been closely related to vast bilateral hyperactivation in the posterior cerebellum, a presumed correlate of the re-erection of new scaffolds [23]. All in all, the combination of surprisingly preserved behavioural performance in predictive timing, cerebellar atrophy and extensive hyperactivation of the posterior cerebellum is an evidence of a successful struggle of this versatile structure with age, escaping the general trends of functional decline seen in the supratentorial area.
The importance of these findings extends far beyond the realm of temporal estimation, given the pervasive evidence in various behavioural, psychological and imaging studies, which position the cerebellum as one of the essential nodes in a vast spectrum of cognitive and associative processes [24]. Furthermore, the cerebellum has been implicated in various diseases, often even devoid of the “traditional” cerebellar symptoms [25–26]. Ergo, the ability of the cerebellum to counteract ageing and related changes with the recruitment of new scaffolds, makes it a worthy candidate for further research into ageing processes as well as our options to maintain the performance of our ageing population across multiple domains.
3. Roles of cerebellum subserving processing speed in older adults (Clive H. Y. Wong, Tatia M. C. Lee, Chetwyn C. H. Chan)
3.1. Background
Studies on the cerebellum have revealed its essential role in processing cognitive information [27]. Earlier research reported significant neurobehavioural correlates of cerebellar volumes or sub-regional activations [28] with test performances in sustained attention [29] and executive function and working memory [30]. Processing Speed (PS), a latent factor among many cognitive functions [31], refers to how fast a person can complete a simple task with minimal involvement of working memory and executive function. Individuals’ PS would peak at around the age of 30 years, and then begins to decline in later life. In this section, we focus on the functional roles of the cerebellum, and specifically the changing in the cerebello-cerebral functional connectivities (FC) due to normal aging to address cognitive slowness, a common phenomenon among older adults.
3.2. Cerebello-cortical connectivity in the young
The latent speed factor mentioned above is associated with increased activations in neural substrates in both the cerebrum and cerebellum [32]. Topographic distributions of these neural substrates resemble those in two distinctive intrinsic cortical networks: the Dorsal Attention Network (DAN), which subserves top-down attention control, and the Salience Network (SN), which subserves goal-related mental representations. Specifically, the cerebellum’s hemisphere and vermis portions of lobule VI/VII are functionally connected to the DAN and SN [33]. Findings of other studies have also supported the associations of cerebello-cortical connectivity with younger individuals’ performances on a complex visual processing speed task (i.e., symbol-digit modality task) [34–35].
We have addressed the processing speed question by using a simple and cross-modality task. The Arrow Task has both a visual and an audial condition, which was constructed to minimize the potential biases resulting from the cognitive strategy and modality-specific brain activities [36]. We have observed that activations in the prefrontal region and cerebellum are significantly associated with the speed composites of the task conditions. Fast performers show higher prefrontal but lower cerebellar activations [36]. We have employed effective connectivity to understand the possible couplings between the frontal region and cerebellum.
Further analyses have suggested both the cerebello-cortical and cortico-cerebellar couples exert influences on the younger participants’ speed performances. Importantly, the stronger cerebello-cortical and weaker fronto-cerebellar influences are associated with higher speed. These findings indicate the likelihood that the cerebellum is the primary driver of faster speed when young individuals complete simple cognitive tasks. The results also form the basis for exploring how neurodegeneration due to normal aging would modulate the roles of the cerebellum in cognitive slowness among older adults.
3.3. Cerebello-frontal connectivity in the old
Given the cerebello-cortical connectivity revealed in younger adults, Gao et al. (2020) have employed connectome-based models to explore the differences in speed-related connectivity between younger and older individuals [37]. Brain connectivities among the pairs of 268 nodes were derived from resting-state functional brain signals correlated with the participants’ task-based reaction times. The results suggest two connective networks involving positive and negative paths, respectively. Interestingly, these two networks are significantly correlated with the task-based speed measures in older individuals but not younger individuals. In addition, a subset in the positive network shows that cerebello-frontal paths are associated with faster speed.
In contrast, a subset in the negative network has revealed that cerebello-sensorimotor paths are associated with slower speed. Our latest work on cerebello-cortical effective connectivity showed that older individuals tend to recruit more extensive neural networks than their younger counterparts in the Arrow Task [36, 38]. The other network involved is the Cognitive Cerebellar Network to the original DAN and SN for younger individuals. This additional network has been tested with a mediation model, with the inter-network effective connectivities as a mediator between age and speed [38]. Both the SN and Cerebellar Networks exert influences on the DAN. The Cerebellar-to-Dorsal Attention influences were found to mediate the age-related slowing, and the Salient-to-Dorsal Attention influences suppressed age-related slowing. We have speculated that, among older individuals, the SN would exert higher activity levels, possibly compensating for the declined cerebellar activity to optimize processing speed when performing simple tasks.
3.4. Conclusion
We assert that the cerebellum plays a mechanistic role in mediating processing speed. Extant evidence supports the existence of a cerebello-cortical network [33], an aging-related decrease in cerebello-cortical FC [39], and age-dependent speed-volume correlations in the cerebellum [40]. Here, we connect the dots and further pinpoint the cerebellar-salience-dorsal attention network interactions in cognitive slowing. Previous studies have associated the cerebellum with the automation of cognitive processes [41]. The evidence of the cerebellar and cerebello-cortical connectivity reported herein suggests age-related slowing. The cerebello-frontal network could play an even more significant role in processing speed. Taken together, declined cerebellar function may be compensated by increasing frontal activity to optimize processing speed in older adults. Future studies are to explore the reciprocal processes of the cerebellar networks and the networks’ roles in subserving other human functions.
4. Cerebellum-basal ganglia interactions with aging (Jessica A. Bernard)
The cerebellum communicates with the cerebral cortex via closed loop circuits [42]. In parallel to these cortical circuits are subcortical links between the cerebellum and basal ganglia. First discovered in non-human primates, these connections provide bidirectional connections between the cerebellum and basal ganglia [4]. Though the exact computational function for these interactions is unknown, it has been speculated that they may be foundational for broader cortical networks, and are important for learning processes [4]. In the human brain, there has been limited work using diffusion imaging to map these circuits, but the results thus far have been promising. Pelzer and colleagues [43] completed a feasibility study using tractography and demonstrated the presence of parallel circuits.
There is additional evidence for these connections from work employing resting state FC magnetic resonance imaging (fMRI). Connectivity studies investigating the cerebellar lobules have demonstrated associations between the cerebellum and basal ganglia at rest [44], and notably similar relationships were also present when examining the ventral rostral putamen [45]. Using more targeted approaches to look at cerebellar-basal ganglia connectivity focusing solely on nodes within these regions (as opposed to whole-brain approaches), robust connectivity has been demonstrated in young adults [46]. Notably, these patterns of connectivity are stronger for subregions in the cerebellum and basal ganglia that are associated with motor and cognitive cortical circuits [46], respectively, as defined based on lobular connectivity, regional striatal connectivity, and work in non-human primates on the closed-loop cerebello-thalamo-cortical networks [45, 47]. These patterns of robust connectivity in young adults have also subsequently replicated in a large sample [48].
Understanding this circuit in aging is of great interest given the broad functional contributions of both the cerebellum and basal ganglia, and the growing literature implicating these regions and their circuits in aging processes [39, 49–50]. Initial suggestions that there may be age differences in connectivity between these regions came from a whole-brain investigation of lobular cerebellar connectivity in older adults [44]. In this investigation, when comparing connectivity of each hemispheric and vermal lobule in young and older adults, one of the most consistent patterns was a lower connectivity between the cerebellum and basal ganglia in older adults relative to young adults [44]. Furthermore, connectivity between the cerebellum and caudate nucleus was associated with motor performance in older adults such that better performance was related to higher connectivity [44]. In a large sample of adults (n = 590) between the ages of 18 and 88 years, there were negative correlations with age between the cerebellar dentate nucleus and putamen [50], providing further support for the impacts of age on this circuit.
In a targeted follow-up focusing on the cerebellum and basal ganglia, connectivity in older adults was not just lower than that of young adults, it also differed in direction. In young adults, there were positive correlations between seeds in the cerebellum and basal ganglia, while in older adults the predominant pattern was that of anti-correlations [46]. Furthermore, connectivity between Crus I and the superior ventral striatum was positively associated with self-report measures of balance. That is, older individuals with higher correlations between these regions also reported better balance when completing everyday activities, highlighting the functional relevance of these networks [46]. Finally, while in young adults there was a clear dissociation in terms of connectivity strength for connections within motor and cognitive circuits, this was no longer present in older adults [46], broadly consistent with ideas of dedifferentiation in the aging brain [51].
Mechanistic explanations for these differences in cerebellar-basal ganglia connectivity remain largely speculative. However, we have previously suggested that these differences are related, at least in part, to age differences in dopaminergic function [44; 46]. In individuals with Parkinson’s disease, there are difference in cerebellar connectivity when on and off l-dopa medication, demonstrating the broad sensitivity of these circuits to dopamine [52]. Notably however, the administration of l-dopa to healthy young adults modulates connectivity between the basal ganglia and cerebellum [53]. Given that there is a degree of normative decline in dopamine in advanced age [54], this may, at least in part, be driving connectivity differences in advanced age. More generally, this work points to the relative importance of considering sub-cortical circuits in our understanding of the processes of aging.
5. Cognitive Cerebellum and Aging (Colby L. Samstag, Erik S. Carlson)
The cerebellum is classically recognized for its role in sensory error prediction signaling in motor learning and postural control, facilitating the detection and correction of differences between intended and performed motions. Recent research has begun to elucidate the extent to which the cerebellum also participates in diverse cognitive domains including reward signaling, spatial navigation memory, and nonmotor learning in emotional domains like fear conditioning, all of which contribute to higher level executive function [1]. Due to the uniform repeated architecture of the cerebellar cortex, it is thought that cerebellar circuitry functions as an error prediction in these cognitive tasks. Furthermore, since the cerebellum receives inputs from nearly every region in the cerebral cortex, as well as ascending inputs from the spinal cord [55] and cranial nerves, it serves as a nexus between cortical and subcortical communication. In this way, it is thought that the cerebellum coordinates the functions of other brain regions at the circuit level, contributes to the building of an internal model, and facilitates automatic behaviors [1]. Some evidence even suggests that total cerebellar volume can independently predict scores on intelligence tests [56]. Despite the increasing attention on the role of the cerebellum in cognitive processing, it remains poorly understood the extent to which the cellular pathological changes that occur in the cerebellum during ageing affect cognitive processing.
Much of our knowledge of the cerebellum’s role in cognitive processing comes from the study of human patients exhibiting gross cerebellar lesions as the consequence of strokes, prenatal or developmental lesions [57]. Cerebellar damage causes defects in classical cerebellar functions, resulting in ataxia and deficiencies in motor learning. However, it frequently also causes a constellation of altered cognitive and social dysfunction phenotypes collectively referred to as cerebellar cognitive affective disorder, affecting virtually every cognitive domain including attention, perception, declarative memory, language, and working memory [57]. In particular, patients with cerebellar lesions show difficulty with executive function, including planning, abstract reasoning, and multitasking, tasks which are also impaired in dementia [57]. Classical cerebellar functions including eyeblink classical conditioning and postural control decline with age [55], but few studies link changes in the cerebellum with age-related cognitive decline.
The cerebellum undergoes structural and functional changes with age, and conditions of ageing present with phenotypes reminiscent of acute cerebellar injury [57]. Longitudinal neuroimaging of healthy subjects demonstrates that regions of the cerebellum undergo reductions in volume [58]. Furthermore, overall cerebellar volume is closely associated with gait speed, itself a strong predictor of cognitive decline [59]. Neuroimaging of patients who progress from mild cognitive impairment to dementia show reduced volume of Crus I and Crus II [60], and patients with Alzheimer’s disease also show reductions in cerebellar volume [61]. Age-related reductions in cerebellar volume are not uniformly distributed, and parts of the cerebellum implicated in cognition appear particularly vulnerable. Convergent evidence from humans, monkeys, and rodents have highlighted the importance of the lateral cerebellum and the dentate nucleus as centers of cognitive cerebellar processing, regions that also show degeneration with age (reviewed in [62]). The dentate nucleus is a region that has expanded greatly in primate lineages, and experiments in rodents show that neurons in the dentate nucleus participate in a host of cognitive functions including working memory and spatial navigation [63]. The density of these regions follow an “inverted U” curve throughout the human lifespan, peaking at about age 30 and declining with age [64–65]. Neurons of the dentate nucleus acquire other pathological hallmarks with age, including an accumulation of both iron [66] and lipofuscin [67] in normal ageing, and increased DNA damage and cell cycle events in Alzheimer’s disease [68]. Taken together, these data suggest that decreases in cerebellar volume accompany ageing and are at least correlated with cognitive decline, though a causal relationship has not been clearly established.
Several models have emerged to explain how age-related cerebellar damage may impact cognition. In addition to its role as an error prediction circuit, the cerebellum connects “low level processing” subcortical and “high level processing” cortical regions, and it buffers the effects of increased attentional burdens from these other brain regions (reviewed in [69]). Collectively, the cerebellum functions as a cognitive reserve to free up resources for prefrontal cortical regions to perform other tasks (see also section 14). This model posits that age-related cerebellar defects lead to attentional burden on other systems and results in more disorganized and slower function. As ageing leads to cerebellar volumetric loss and alterations of FC, there is a decrease in the capacity for cortical regions to offload automatic tasks to the cerebellum. Regions of the cerebellum that undergo age-related changes may therefore become an important target for treatments aiming to slow or reverse cognitive decline. In support of this hypothesis, recent work shows that bilateral transcranial magnetic stimulation of Crus II increases cognitive performance in patients with Alzheimer’s disease by strengthening FC networks with nodes of the prefrontal cortex [70]. In summary, compelling evidence shows that the cerebellum participates in cognitive tasks, but also undergoes age-related decline in function, yet a definitive mechanistic link between cerebellar function and cognitive decline remains elusive. Future work should clarify the role of cerebellum in cognitive decline with age, and whether interventions targeting the cerebellum can reduce attentional burden on cortical regions in cognitive decline.
6. Cerebellum, healthy ageing, and spatial navigation (Stephen Ramanoël, Christophe Habas, Angelo Arleo)
Spatial navigation constitutes a complex behaviour that encompasses perceptual, cognitive, and motor processes underlying efficient goal-oriented actions in most daily activities. This manifold ability requires the integration of different sensory modalities, the acquisition and maintenance of spatial knowledge in memory during the locomotion phase, and the flexible use of adaptive spatial strategies [71]. Spatial navigational abilities are impacted by healthy ageing and by changes occurring in multiple brain regions, including the frontal, temporal, and cerebellar cortices [72–73]. Notably, older adults display prominent impairments in their capacity to integrate spatial information from the environment, to reorient or navigate in unfamiliar environments, and to flexibly switch among spatial strategies when necessary [74]. While an extensive body of literature has highlighted the importance of the age-related atrophy and dysfunction of the hippocampus in the decline of navigational abilities, the cerebellum has only recently gained interest in the field of ageing and spatial navigation.
Accumulating evidence indicates that cerebellar-hippocampal interactions support the implementation and the efficient use of spatial representations during goal-directed behaviour [75–77]. These findings extend the role of the cerebellum beyond its traditional implication in motor control. Igloi and colleagues (2015) have shown that the use of a sequence-based navigational strategy, which relies on the formation of associations between spatial cues and a directional behaviour, is underpinned by co-activation of the right cerebellum (Crus I), left hippocampus, and frontal regions [75]. The authors also reported that the implementation of a place-based strategy, defined as the formation of a flexible mental map-like representation of the absolute position of the goal, was subtended by different interactions between left Crus I, right hippocampus, and medial parietal cortex. In the same vein, Hauser and colleagues (2020) found that modulation of cerebello-hippocampal activity could differentiate spatial reference frames (word-centred vs. self-centred) during passive perception of visuo-spatial changes [76].
In the context of healthy ageing, few studies have indicated age-related structural or functional differences in the cerebellum during navigational tasks. One cross-sectional study and one longitudinal study have reported age-related changes in turning preference [78] and distance-dependent navigation performance [79] in a virtual Morris Water Maze task that were associated with global cerebellar atrophy. These results are consistent with a non-motor role of the cerebellum in navigation, in relation to the multisensory integration of spatial information to guide behaviour [77]. Moreover, results from functional studies have allowed the cognitive role of the cerebellum to be precisely outlined by exploring the activity of cerebellar subregions recruited during navigation. fMRI studies conducted on young and healthy older adult populations have reported the involvement of the Crus I and the lobule VI regions of the cerebellum during distinct spatial memory tasks in virtual environments [74, 80–81]. Ramanoël and colleagues (2020) have found that older participants exhibit stronger lateralized cerebellar activity in the right Crus I than younger subjects when orienting using visual landmarks [74]. In addition, in a recent voxel-based-morphometric study conducted on the same sample of participants, the authors have shown an age-related reduction of grey matter volume (GMV) only for the Crus I region in the left cerebellum [82]. These lateralization effects were interpreted as a neural correlate of the age-related shift from the use of a place-based strategy, involving left cerebellar and right hippocampal coactivation, to the use of a sequence-based strategy underpinned by right Crus I, left hippocampal, and frontal regions [75]. This pattern of results could reflect older adults’ reduced capability to form map-like representations of the environment. They could also reveal a functional adaptation to the left Crus I atrophy that favours the use of the relatively preserved right Crus I in terms of grey matter atrophy and that supports sequence-based strategy. An alternative, not exclusive interpretation could be related to age-dependent impairments in executive functions that may impact older adults’ capacity to flexibly switch between navigational strategies. Indeed, Crus I region has been reported to be connected to a large associative network supporting executive control in various aspects of cognition [33].
Taken together, these studies highlight the role of the cerebellum in complex cognitive abilities such as spatial navigation. However, it is important to note that none of the aforementioned studies on healthy ageing and navigation had the primary aim of investigating cerebellar functioning. In addition, only one voxel-based-morphometric study used a specific pipeline optimized for cerebellum analysis [82]. These elements stress the need for future studies to use specific a priori hypotheses on human cerebellar dynamic in order to better characterize the impact of healthy ageing on navigation. These findings on the cognitive cerebellum provide the basis for new fundamental and applied research to maintain autonomy of healthy older adults and to characterize age-related pathologies in terms of spatial navigation deficits [83–84].
7. Cerebellum and the physio-cognitive decline syndrome (Chih-Ping Chung, Liang-Kung Chen)
7.1. Physio-cognitive decline syndrome, concurrent physical and cognitive impairments, as a phenotype to identify the elderly at risk of dementia or/and disability
Physical and cognitive abilities are critical elements for maintaining functional independence in the elderly [85]. The accumulated evidence suggests that physical and cognitive declines have synergistic, harmful interactions on healthy aging [86–88]. In addition, the risk for dementia is higher among older people with concurrent physical frailty and cognitive impairment compared to those with only cognitive impairment [88]. Many cohort studies have found that gait slowness and handgrip weakness (mobility subtype of physical frailty) are the two physical functions most-strongly associated with cognitive impairment in the elderly.
Physio-cognitive decline syndrome (PCDS) is defined based on these observations and serial research studies, affecting older, functionally-preserved adults who have slowness and/or weakness (by cut-offs from the 2019 consensus update of the Asian Working Group for Sarcopenia) [89] plus cognitive performance ≥ 1.5 SD below the mean for age-, sex-, and education-matched controls in all cognitive domains, as assessed by comprehensive objective neuropsychological tests. In large cohort studies, PCDS delineated a subpopulation of 10–15% of community-dwelling older persons with neither dementia nor disability [90]. These studies also showed that PCDS had high predictive value for incident dementia (HR 3.4, 95% CI 2.4– 5.0) among 4570 ≥ 65-year-olds followed for 36 months [91] and for incident disability (HR 3.9, 95% CI 3.0–5.1) among 9936 ≥ 65-year-olds followed for 24 months [92]. Since the operational definition of PCDS successfully identifies an appreciable population of at-risk older people, PCDS has been proposed as a phenotype and potential treatment target of unhealthy aging at an early stage, and also a framework for studying the pathophysiological mechanisms of mobility frailty-associated cognitive impairment [90].
7.2. Cerebellar structure and neurocircuit abnormalities as the neural signature of PCDS
Neuroimaging studies of community-dwelling older people in the I-Lan Longitudinal Aging Study (ILAS) and National Institute for Longevity Sciences-Longitudinal Study of Aging (NILS-LSA) cohorts revealed a specific pattern of gray matter volume (GMV) changes in physical frailty and PCDS [93–95]. Using voxel-based analysis of the whole brain magnetic resonance T1-weighted imaging, researchers first identified regions of GMV reduction in the elderly with physical frailty. These regions included the cerebellum, hippocampi, middle frontal gyri, right inferior parietal lobule, precentral gyrus, and left insula, anterior cingulate, and middle occipital gyrus. The study also revealed that each physical frailty subtype had distinct GMV changes [93]. The cerebellum was the region with the greatest GMV reduction related to mobility subtype frailty (gait slowness or/and handgrip weakness) compared with the other components of physical frailty (exhaustion and involuntary body-weight loss) [93]. The neuroanatomic correlate of PCDS, concurrent mobility frailty and cognitive impairment, was also evaluated. Voxel-based whole-brain analysis showed that participants with PCDS had lower GMV than non-PCDS ones in the right and left amygdala, right and left thalamus, right hippocampus, right temporal occipital fusiform cortex, right occipital pole, and left cerebellum VI and V (family-wise error cluster corrected P-value < 0.05). The regions with GMV reduction in PCDS subjects were similar between the middle-aged and older adults; whereas larger clusters with more extensive GMV-reduction regions were observed in ≥ 65-year-olds with PCDS than in non-PCDS contemporaries (Table 1, family-wise cluster corrected p-value < 0.05) [94].
Table 1.
Reduced gray-matter volume in physio-cognitive decline syndrome subjects of two aged groups
| Anatomical region | MNI-space coordinates | Maximum intracluster t value | ||||
|---|---|---|---|---|---|---|
|
| ||||||
| Cluster size | Side | Structure | X | Y | Z | |
| Reduced GMV in PCDS < 65 years old | ||||||
|
| ||||||
| 598 | Left | Occipital pole | −13.5 | −100.5 | 0 | 3.99 |
| 405 | Left | Precentral gyrus | −58.5 | 7.5 | 15.0 | 4.32 |
| 298 | Right | Amygdala | 25.5 | 0 | −27.0 | 3.69 |
| 255 | Left | Cerebellum V | −18.0 | −45.0 | −16.5 | 3.40 |
| 138 | Left | Cerebellum VIIIa | −24.8 | −62.3 | −47.3 | 4.38 |
|
| ||||||
| Reduced GMV in PCDS ≥ 65 years old | ||||||
|
| ||||||
| 30986 | Right | Hippocampus | 32.3 | −15.8 | −10.5 | 6.97 |
| Right | Thalamus | 1.5 | −12.0 | −10.5 | 6.51 | |
| Left | Amygdala | −27.0 | −12.0 | −15.0 | 6.19 | |
| 2580 | Right | Cerebellum Crus I | 30.8 | −66.0 | −33.0 | 4.29 |
| 2013 | Left | Cerebellum Crus I | −31.5 | −69.0 | −31.5 | 4.37 |
| 1274 | Right | Occipital pole | 13.5 | −91.5 | 30.0 | 4.68 |
| 869 | Cerebellum vermis VIIIa | 4.5 | −67.5 | −39.0 | 3.99 | |
| 598 | Right | 58.0 | −25.5 | 16.0 | 4.14 | |
| 584 | Right | Postcentral gyrus | 45.0 | −19.5 | 63.0 | 3.90 |
| 562 | Right | Lateral occipital cortex | 52.5 | −63.0 | 13.5 | 4.32 |
| 545 | Right | Cingulate gyrus | 7.5 | 1.5 | 40.5 | 4.23 |
Analyses adjusted for age, sex, Centre for Epidemiologic Studies Depression Scale and cardiovascular risk factors (hypertension, diabetes, hypderlipidemia, smoking and obesity). MNI = Montreal Neurological Institute. GMV = gray-matter volume.
These studies consistently found cerebellum involvement in mobility frailty and related cognitive impairment (e.g., PCDS). Among other identified brain regions with GMV reduction in PCDS, hippocampus/amygdala GMV has long been recognized as an early marker of cognitive impairment in the elderly [96]. Therefore, a neurocircuit connecting the left cerebellum and hippocampus/amygdala that might be involved in the pathophysiology of PCDS has been postulated [94]. Researchers used diffusion-weighted tractography to establish a population-based probability map of hypothetical neuroanatomical connections. Among 1,006 included participants, delineated tracts successfully connected cerebellar regions of interest to the left and right hippocampus/amygdala area in 72.5%, but to the right amygdala/hippocampus area in only 20.8%. The tract probability map (likelihood of participant’s cerebellar-hippocampus/amygdala connection passing through the voxel) described a highly coherent connection path across participants, with maximal probability of 60.8%/72.5% on the left side and 15.0%/20.8%) on the right (Figure 1) [94]. Tract-specific analyses showed that within bilateral cerebellum to amygdala/hippocampus connections, PCDS subjects had significantly higher values than non-PCDS ones in three different diffusivity indices (mean, radial and axial) [94].
Figure 1. Group-wise probabilistic connections between cerebellar and bilateral amygdala/hippocampus areas.
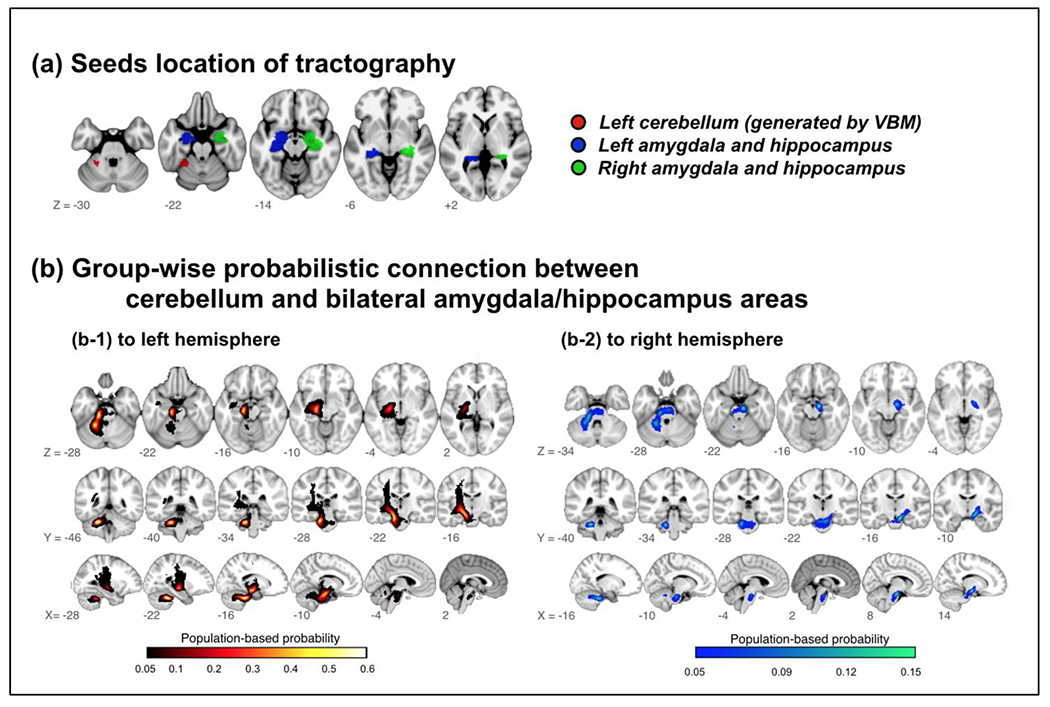
VBM = voxel-based morphometry.
The neuroanatomic characteristics revealed by these studies have provided evidence for cerebellar structure and neurocircuitry involved in concomitant physio-cognitive declines in older people. These findings may provide clues to the early neuroanatomic pathology of age-related concomitant physical and cognitive decline. Cerebellum GMV and hippocampus-amygdala-cerebellum neurocircuits might constitute imaging biomarkers in future preventive interventions for health aging.
8. Cerebello-cerebral functional connectivity with ageing (Giusy Olivito, Maria Leggio)
Since the cerebellar role in cognitive and behavioural functions has been well established [24, 97], the cerebellum has attracted an increased interest and importance in the context of ageing [44, 46]. Healthy ageing is typically characterized by decline in both motor and cognitive functions [17]. Consistently, MRI studies have shown age differences in brain volume [98] as well as in resting state brain networks [99–100]. In particular, age differences in FC have been related to both motor and cognitive performance in older adults [44, 100], thus supporting the functional relevance of these differences.
While the investigation of age-related differences in neural substrate has been primarily focused on the cerebral cortex [98, 101], the study of cortico-cerebellar FC has provided crucial insight into understanding both motor and cognitive declines associated with healthy aging [44]. Furthermore, differences in cerebello-cerebral FC across life span have been shown [44, 46], demonstrating in particular lower cerebellar connectivity as individuals age [44, 46–47].
Functional segregation of the cerebellum has been evidenced by several connectivity studies [102–103] showing that it plays a crucial role in motor functions as well as in cognition through functional interactions with cerebral regions. In particular, based on FC results, the human cerebellum has been divided into a primary sensorimotor zone, having connections with motor and somatosensory cortices, and a supramodal zone, mainly encompassing the cerebellar Crus II, having connections with prefrontal and posterior parietal cortices [103] and strongly related to cognition [102].
It has been shown that in older adult, within-cerebellum connectivity greatly overlaps with the pattern observed in young adults, with cerebellar regions implicated in motor networks (i.e., Lobules I–IV, V, VI and VIIIa) strongly correlated between themselves, similar to how the posterior lobules are correlated between themselves [44]. Conversely, in older adults the patterns of cerebello-cortical connectivity do not overlap with findings in the young adults. Consistent with classical functional segregation of the cerebellum [24, 103], in young adults cerebello-cerebral connectivity has been shown between lobules I–IV, V, and VI and pre-motor and primary motor cortical regions [103] while in older adults, a more widespread pattern of correlations has been found between regions including the hippocampus, middle frontal gyrus, parahippocampal gyrus, anterior cingulate cortex, and dorsal pre-motor cortex [44]. In contrast, the pattern of correlation between posterior cerebellar lobules (i.e., Crus I and Crus II) and the cerebral cortex in older adults was similar to that found in young adults involving the prefrontal and temporal cortex, as well as subcortical structures, such as the caudate and thalamus. However, unlike the findings in young adults [103], no functional correlations were found between lobules of the vermis and the primary and pre-motor cortical regions [44].
Beyond the qualitative comparison of research findings in young and older adults described above, the direct comparison between the two groups has provided interesting insights into age-related cerebellar FC differences, showing in particular a global FC decrease in cerebello-cerebral networks of older adults compared to young adults with no differential patterns by cerebellar functional regions thus involving both motor and cognitive cerebello-cerebral modules [44, 103]. This finding suggests that cerebello-cerebral networks are equally impacted by aging, regardless of functional specialization [44], and is further supported by evidence that both dorsal (motor) and ventral (cognitive) regions of the dentate nucleus show age-related negative correlations with motor and association cortices, respectively [50].
Of particular interest is the pattern of decreased FC involving the cerebellum and the medial temporal lobe, thus suggesting a relation between disrupted cerebellar-hippocampal connectivity and memory deficits typically reported in healthy aging [104]. Importantly, the opposite pattern of FC has been also described in older adults with greater connectivity than that observed in young brains mainly involving the prefrontal cortex and cerebellar regions predominantly implicated in intrinsic motor network [102–103]. This is likely to be related to compensation mechanisms in response to the overall cerebello-cerebral functional decrease [44].
Among the intrinsic FC networks of the brain, the DMN has been especially well studied in older brains [99–100]. Studies show that decreased FC in the DMN is related to performance on cognitive tasks and contributes to cognitive decline across multiple domains [100]. Indeed, the DMN has a crucial role in the context of internally directed mental states, such as remembering, planning, and related cognitive functions [100] and has been shown to have strong FC with posterior cerebellar Crus II in healthy subjects [102], suggesting that cerebellar contribution to the DMN may be important for understanding cognitive variability in older adults.
Age differences in connectivity patterns between the cerebellum and basal ganglia have been also been found in a recent study [46] showing that altered FC is related to behavioural performances in older adults. Indeed, both structures strongly contribute to the performance of motor and cognitive tasks [24] and are reciprocally connected, forming an integrated network with the cortex [4].
Overall, all these observations raise important implications for understanding pathological mechanisms underlying motor and cognitive decline in advanced age as well as improving the understanding of age-related neurodegenerative conditions, such as Alzheimer’s disease (AD) (an overview of the cerebellum in Alzheimer disease can be found in the following sections).
9. Role of the Cerebellum in Cognitive Decline in Alzheimer’s Disease (Muriel M. K. Bruchhage)
9.1. Cerebellum and Alzheimer’s disease progression
AD is the most prevalent form of dementia [105] and is defined by progressively degrading cognitive, motor and socio-emotional function. AD cognitive deficits precede symptoms by on average 20 years and are associated with structural and functional changes in the brain (for example, [106]). While traditionally focus has been placed on hippocampal and cerebral tissue loss, accumulating findings have implicated posterior cerebellar atrophy in AD disease progression (for a review, [8]). A recent meta-analysis (84.Gellersen et al., 2021) confirmed that associations between posterior GMV loss and reduced cognitive performance persisted for healthy ageing and AD even when controlling for total cerebral and hippocampal volume, indicating an independent contribution of the posterior cerebellum to ageing processes and AD progression. Our group applied voxel-based prediction maps of cerebellar GM and white matter volume fraction myelin (VFM) comparing whole brain, cerebral and cerebellar contributions to different stages of cognitive impairment in AD and healthy ageing [107]. While GM cerebellar prediction accuracy was higher than for the whole brain and cerebrum across all stages of cognitive decline, cerebellar VFM prediction accuracy was highest for mild/moderate stages of AD but performed lowest for mild cognitive impairment across modalities. This prevailed even when dividing the cerebellum into anterior and posterior regions, with the highest AD prediction accuracy for posterior cerebellar lobe in advanced cognitive decline and the anterior cerebellum in the first stage of cognitive decline. While these results confirm the importance of the cerebellum to cognitive decline in AD progression independent of cerebral contributions, they also potentially indicate differences in timing of cerebellar tissue and regional contributions to stages of AD. While early stages of cognitive decline seem to be driven by anterior regions of the cerebellum, the posterior cerebellum contributes to established cognitive decline in AD, increasing with symptom severity. This parallels cerebellar functionality and AD symptom disease progression, as motor phenomena classically associated with anterior cerebellar dysfunction are often present in early AD symptomatology, preceding cognitive symptoms associated with posterior cerebellar dysfunction.
9.2. The cerebellum is part of a dynamic affective network
The cerebellum is highly interconnected structurally and functionally, enabling cerebellar networks to impact both cerebellar and cerebral functioning and connectivity through short- and long-term plasticity [108]. These cerebellar plastic changes are not restricted to motor skills but expand to cognitive and emotional functions as early as childhood (for example [107]). Neuroanatomic connections and clinical observations stemming from cerebellar injury have led to functional divisions of the cerebellum into the mainly anterior ‘sensorimotor cerebellum’, and the posterior ‘cognitive cerebellum’ [24]. This applies further to lateralization effects. Damage to the right cerebellar hemisphere results in language difficulties and right vermal damage leads to social and affective processing deficits, while damage to the left hemisphere results in spatial difficulties (for a review [109]). The same meta-analysis [84] confirming posterior GM loss associations with reduced cognitive performance in AD further identified a right lateralization of the posterior lobe unique to AD. Together with the cerebellum’s widespread connections, this could indicate more specialized atrophy processes at play, exacerbating along the right posterior cerebellum to the cerebrum affecting cognitive and language functions typical to AD symptomatology.
9.3. Cerebellar contributions to AD symptom development
The human cerebellum is one of the earliest regions of the brain to develop [110], but continues to grow into adulthood, with posterior regions maturing late [111]. This long duration makes it especially vulnerable for disorder and disease, including the role of early-life adverse events in AD genetic markers increasing chances of AD development and progression. In other words, environmental perturbations at a very early period determine the susceptibility to pathological conditions later in life (for a review, [112]). AD neurodegeneration follows specific topographic patterns of grey and white matter loss that emerge during its early stages, with white matter loss speculated to precede GM atrophy [113]. Coupled with differences in timing of cognitive decline and accumulating findings of right posterior cerebellar atrophy in AD progression, it could indicate that early abnormalities in cerebro-cerebellar networks together with genetic vulnerability affect early stages of AD symptom development. Thus, differential subregional vulnerability to AD pathology and normal ageing may be due to early downstream influences resulting from cerebro-cerebellar synergism (for example, [114]), and structural decline may also have selective effects on distant cerebral regions enabled by networks established already at very early development [115].
9.4. Conclusion
Increasing evidence supports an active role of cerebellar abnormalities in early AD development and associated cognitive decline. However, AD diagnosis is often supported by positron emission tomography scans, where overall volume is normed against cerebellar volume to assess differences due to ageing processes. As outlined, recent evidence suggests a potentially more dominant role of the cerebellum in AD cognitive decline. Thus, clinical AD diagnosis and treatment should account for, rather than norm for, potential cerebellar atrophy at different stages of AD progression and view AD as a disease affecting dynamic networks rather than isolated cerebral and hippocampal atrophies. The increasing role of the cerebellum in AD development might further stimulate new research directions needed after recent controversy around the amyloid hypothesis [116].
10. Cerebellar grey matter loss in healthy ageing and Alzheimer’s disease (Heidi I.L. Jacobs, Xavier Guell, Jeremy D. Schmahmann)
The discovery of the cerebellar cognitive affective syndrome and its localization to the cognitive cerebellum in the posterior lobe [117–118] directed attention to the previously unrecognized possibility of cerebellar contribution to the cognitive and neuropsychiatric symptoms typical of AD [8]. Cerebral hemispheric volumetric changes in AD overlap only partially with healthy ageing, a concept that is now expanded to include the cerebellum [84].
10.1. Ageing and Mild Cognitive Impairment (MCI)
Cerebral cortical thinning is observed in most brain regions with advancing age and increases linearly over time [119]. Widespread cerebellar volumetric decline mirrors these changes in the cerebral hemispheres, and is thought to reflect principally the loss of white matter in cerebellar folia and corpus medullare in the vermis and both hemispheres [58, 120].
In older individuals with mild cognitive deficits but without dementia, lower GMV was seen in association with worse overall cognitive performance, while posterior lobe GMV loss was associated with lower executive function scores [121]. Volume loss may reflect neuronal drop-out, as patients with MCI or AD have fewer cerebellar Purkinje cells than cognitively normal older individuals [122]. Synaptic alterations are observed in the mossy fibers (MFs), granule cell dendrites, parallel fibers and Purkinje cell dendrites with substantial loss of dendritic spines, and a decrease in the number of granule and Golgi cells in the granule cell layer [123–124]. These changes are most frequent in Crus I, consistent with observations from structural neuroimaging studies demonstrating lower grey matter volume in the vermis and posterior lobe in the early stages of the disease, whereas the anterior lobe is more affected in the later stages [125]. In patients with MCI, functional neuroimaging studies report similar topographic findings of lower task-related activity predominantly in cerebellar posterior regions initially, and present in anterior cerebellar regions in later stages [8]. This evolution of atrophy corresponds to the clinical phenotypes of early cognitive and emotional disturbances and later motor deficits. Lower cerebellar volume is associated with higher odds of being in the prodromal stages of the disease, even when controlling for hippocampal volume and APOE-status [107, 126]. In healthy older individuals the cerebellar DMN and cerebral DMN are negatively correlated during rest, whereas in amnestic MCI there is degradation of the anti-correlation between the cerebellar DMN and the medial frontal cortex [127]. This relationship correlates with worse memory performance and suggests that reduced anti-correlation impacts the modulatory role of the cerebellum on cognitive functioning, particularly the executive control of memory [127].
10.2. Ageing and AD
The question of differential grey matter volume loss in healthy ageing versus mild to moderate AD was evaluated in a meta-analysis of 18 studies including data from 2,441 participants [84], leading to three conclusions. First, areas of spatial overlap between regions of decreased grey matter volume were small between the two groups, including only one cluster of overlap in right cerebellar lobules VI-Crus I. Second, despite a lack of prominent spatial overlap, functional networks linked to areas of volumetric changes were similar across both groups, including cerebellar regions connected to default-mode, frontoparietal, and ventral attention networks. This conclusion was further supported by functional gradient analyses that showed a similar localization of areas of volumetric changes in both groups along the two principal gradients of cerebellar functional organization [128–130]. And third, despite the relative degree of functional similarity between the two groups, functional differences were detected when using an alternative functional gradient that relates to lateralization of non-motor function between left and right cerebellar hemispheres. Areas of volumetric changes in AD showed a right-hemispheric preference along this gradient, compared to volumetric changes in healthy ageing that were distributed similarly in both hemispheres.
10.3. AD
The view that the cerebellum was largely spared the neuropathologic hallmarks of AD arose because cerebellar deposition of beta-amyloid plaques and neurofibrillary tangles were detected only in late-stage disease [131–132]. In autosomal dominant AD, however, cerebellar beta-amyloid plaques can be detected in unimpaired carriers 10 years before clinical onset [133–135]. Detailed immunocytochemistry assessments in both autosomal dominant and sporadic AD reveal that the cerebellum accumulates the earlier and possibly more toxic soluble, diffuse forms of beta-amyloid, and occasionally, sparse neuritic plaques [123]. Neurofibrillary tangles are only rarely observed in the cerebellum, as in individuals with early and advanced familial AD with the Osaka mutation in the amyloid precursor protein (APP) gene [136]. These earlier diffuse beta-amyloid accumulations in AD are present in the molecular layer of the cerebellum and in the vicinity of Purkinje cells [124]. Plaque formation can be detected in the same location, but less commonly, and is not observed in the dentate nucleus. These observations are significant because misfolding of tau and beta-amyloid commences two to three decades prior to the first clinical symptoms, highlighting the importance of careful phenotyping, including in older individuals who appear cognitively normal. Severity of cerebellar beta-amyloid deposits is correlated with the number of plaques and tangles observed in the locus coeruleus; the locus coeruleus commits afferent fibers to the cerebellum, is one of the first regions vulnerable to AD pathology [133, 137], and may serve as a route of transneuronal spread of pathology. Indeed, FC studies have reported lower connectivity between cerebellum and locus coeruleus [138], and between cortical resting-state networks and specific cerebellar regions when comparing controls to patients with prodromal or clinically established AD [125, 139–141]. When matching voxel-based morphometry patterns to findings from FC in AD patients, focal atrophy was noted bilaterally in regions of cerebellar Crus I and II that were associated with the DMN [125].
10.4. Mouse models
In older humans lower cerebellar volumes are linked to degraded performance on the classical eyeblink conditioned response [142]. In the ageing mouse model, the decline in this conditional associative learning paradigm occurs together with Purkinje cell loss and diminished synaptic plasticity [143]. In transgenic AD mice, full-length APP and C-terminal fragments are detected early in the cerebellum, indicating processing of the beta-amyloid precursor protein [122]. Increasing intracellular beta-amyloid-related processes and smaller beta-amyloid plaques were observed in the mouse cerebellum as well as in the cerebrum, starting at age 2 months. Cerebellar dysfunction as well as abnormal Purkinje cell activity preceded the detection of these smaller beta-amyloid plaques. Even though non-fibrillar forms of beta-amyloid are not always detected with PET-imaging, this suggests that their presence can disrupt the structure, function and modulatory capacity of the cerebellum.
11. Depression and cerebellar volumes in ageing (Vonetta M. Dotson, Hannah R. Bogoian, Maria Misiura)
The cerebellum’s role in affect and emotion processing was first identified through studies of cerebellar cognitive affective syndrome, a syndrome often marked by emotional lability (i.e., uncontrollable laughter or tearfulness) and changes in affect due to cerebellar lesions [144]. Vermal and paravermal lesions in particular have been observed to induce disruption to emotion and behavior [144], and have led researchers to speculate about the cerebellum’s role in the emotional as well as cognitive aspects of depression [145].
A number of structural imaging studies have since identified a relationship between depressive symptoms and cerebellar GMV. Major depressive disorder (MDD) and higher depressive symptom burden are associated with smaller GMV in the total cerebellum as well as cerebellar subregions, including the posterior cerebellum, vermis, and posterior Crus I [146–149]. Depression-related increases in cerebellar GMV have also been observed. Individuals experiencing an acute episode of MDD were shown to have increased volume in left cerebellar area IX compared to participants with remitted MDD, whereas remitted participants had bilaterally increased volumes in this area [150]. Another study of participants with severe depression found that individuals with MDD had increased right cerebellar area VIIIa volume and increased left cerebellar area VIIb volume compared to healthy controls at baseline [151]. In this study, structural change in the left cerebellum area VIIA Crus I was associated with depressive symptom improvement following a trial of electroconvulsive therapy [151].
To date, investigations into the cerebellum’s role in depression have been conducted primarily with middle-aged adults. However, a recent study examined symptom dimensions of depression and subregions of the cerebellum in a sample of community dwelling late-middle-aged and older adults [152]. Results indicated that greater subthreshold depressive symptoms, as indicated by self-report of depressive symptoms on the Center of Epidemiologic Studies Depression Scale (CES-D), were associated with larger vermis VI volumes. When symptom dimensions of depression were examined through CES-D subscales, positive relationships were identified between the somatic symptom subscale score and vermis VI volumes, and the lack of positive affect subscale score and vermis VIII volume. In contrast, an imaging study of older adults with mild cognitive impairment found that cerebellar GMV was not associated with psychological symptoms of depression or apathy [121].
Aside from grey matter volumes, other imaging modalities have detected cerebellar changes in depressed older adults. FC changes, or correlated brain activity, within the cerebellum or between the cerebellum and other brain regions, have been detected in late-life depression [153–154]. One study found reduced connectivity of the cerebellum in depressed older adults compared to controls [153], while another found increased connectivity in posterior cerebellar regions in treatment-naive older adults with first-episode depression [154]. The discrepancy in findings may relate to the chronicity and treatment status of the two samples, but more research on cerebellar functional connectivity (FC) in depressed older adults is needed to clarify patterns and moderators of depression-related FC alterations in late life.
Cerebellar white matter volumes have been correlated with apathy and depression severity in one study [155]. However, there is a gap in knowledge about white matter integrity of the cerebellum and depression in older adults, despite the well-known link between late-life depression and white matter integrity [156]. Indeed, the presence of white matter lesions, areas of abnormal white matter visualized as white matter hyperintensities (WMH) on MRI scans, characterizes a subtype of late-life depression called vascular depression. There is clearly a need to investigate whether WMH in the cerebellum are related to depressive symptoms, particularly because white matter lesions in the cerebellum typically cause motor symptoms that may be more recognizable to ]caregivers and patients than the slower, more gradual changes in mood seen in LLD [157]. Alternatively, if the cerebellum is relatively spared from WMH in individuals with vascular disease, understanding the protective and compensatory cellular mechanisms inform interventions for depression.
12. DNA methylation and the ageing cerebellum (Patrick Bryant)
Changes in DNA methylation at different genomic sites are related to development and used for regulating gene expression. These changes have also been found to be related to ageing [158], connecting epigenetic changes to lifespan [159]. “Epigenetic clocks” [21] assess this proxy for biological ageing in terms of DNA methylation to predict deviations between chronological and biological ageing. However, the relationship between DNA methylation and age is tissue-specific, suggesting that the rate of the epigenetic clock and DNA methylation changes are not universal, which is also why tissue-specific epigenetic clocks have been developed [160]. This tissue specificity is especially prominent in the cerebellum, where a systematic underestimation of ageing has been observed in older subjects [161].
The underestimation of epigenetic clocks in older subjects has been observed across other tissues as well and one explanation is that there are fewer older samples present in the fitting of these models [21, 162]. Another issue is the difference in sample availability. Due to difficulty in obtaining brain tissue samples and the relative ease with which blood samples can be obtained, blood samples are overrepresented. This is reflected in the higher correlation between predicted and chronological ages in blood compared to, for example, the cerebellum [161. The prominent deviation in the cerebellum is thereby not necessarily functional, but may instead be related to limitations in model fitting and data preparation.
Previously, mainly linear models have been used to relate methylation markers to age. The construction of these models assumes constant change rates and the models are outperformed by ones which do not [160, 163]. A recent study agnostic to the linearity of the methylation-age relationship showed that the methylation markers that change most during ageing tend to have both linear and non-linear relationships with ageing [164]. The rate of methylation in cerebellum markers was also found to be saturated with age, which provides an explanation for the systematic underestimation of models that assume constant change rates throughout life.
By selecting the DNA methylation markers with the biggest age-related changes in 407 cerebellum samples, two distinct types are found: one that is increasingly methylated (hypermethylation) with age and one that is decreasingly so (hypomethylation) [164]. A study relating the functional impact of gene expression on methylation found that hypomethylation is more common with age in monocytes and T-cells [165]. How the hyper- and hypomethylated markers in the cerebellum impact gene expression on a functional basis is unknown. Neither is it known if the variation in methylation is in fact functional for the ageing cerebellum. It is possible that the variation in methylation with ageing is only spurious and perhaps only the hypomethylation is functional.
What is known about the epigenetic age of the cerebellum is that it seems to be functional. This is exemplified by the study of genetic variants in certain gene clusters affecting neurological disorders such as AD and Parkinson’s disease. Genetic variants that impact DNA methylation resulting in acceleration of predicted age according to epigenetic clocks are also related to these disorders [166]. It is suggested that ageing and disease are related to the misregulation of gene expression through altered methylation patterns [167]. The exact mechanisms are likely local, however, as seen through the tissue-specific DNA methylation changes throughout ageing [168].
Since the most prominent DNA methylation changes in the cerebellum are both in the form of hypo- and hypermethylation [164], one can hypothesize that the expression of certain genes is both increased and decreased with age [169]. This instability in gene expression may be related to lower mobility and/or coordination in older age groups [170]. This remains to be investigated, as it is currently not known whether the DNA methylation changes found in the cerebellum relate to changes in gene expression.
Several questions related to the ageing process of the cerebellum and DNA methylation remain unanswered. An important study would be to select the genes related to the most prominent DNA methylation changes (both hyper- and hypomethylated) and investigate their impact on gene expression. Similar to the findings observed in AD and Parkinson’s disease, single-nucleotide polymorphisms (SNPs) in these genes could be related to methylation signatures and phenotypes related to the ageing cerebellum such as coordination. Likely, such data is readily available through open-source transcriptomics datasets with age annotations.
Studying the functional relationship between DNA methylation and ageing will inform many important aspects of phenotype control and lead to improved age predictors. The most interesting aspects of age predictors are not to accurately assess age, but to assess phenotypes through deviating methylation signatures [171]. A model that connects changes in DNA methylation at different ages to gene expression would do exactly this by providing a proxy for differential gene expression and outcomes. Several key issues remain as DNA methylation is tissue-specific and a biopsy of the cerebellum is too invasive. Future models using DNA methylation data from blood could be able to assess health states in other tissues and overall health risks. The relationships of methylation statuses between different tissues have not been studied so far, leaving these questions unanswered.
13. What potential does lifelong musical instrument training possess to avoid age-related cerebellar atrophy? (Masatoshi Yamashita, Kaoru Sekiyama)
MRI studies have revealed key structural characteristics of the aging brain. For example, GMV of the cerebellum and hippocampus decreases with age [172–173] (see also section 10). Atrophy of these brain regions is associated with age-related decline in various cognitive and motor functions [174] (see also sections 5 and 15). Considering the rise in aging-associated issues in the current society, identifying lifestyle habits that effectively mitigate age-related cognitive decline and brain atrophy is essential. Playing musical instruments is a candidate for such lifestyle because it is reportedly associated with a reduced risk of dementia [175]. Nevertheless, it remains unclear whether lifelong involvement in playing a musical instrument is effective in counteracting age-related brain atrophy. This section will introduce the structural and functional advantages of the cerebellum in older musicians who have participated in lifelong musical instrument training.
Musicians are an excellent model to understand learning-related brain plasticity due to their long-term practice. Structurally, young musicians demonstrate increased GMV in the auditory and motor-related regions compared to age-matched non-musicians [176]. Among the various brain regions where young musicians demonstrate gray matter enlargement, the cerebellum may be particularly relevant to behavioural characteristics. For example, Hutchinson et al. (2003) have reported that larger cerebellar volume was associated with daily practice intensity [177]. Furthermore, a recent study on young musicians reported that larger GMV in Crus I of the cerebellum was associated with better temporal discrimination of musical tones [178]. These findings indicate that the cerebellum is associated with instrumental music training and musically relevant cognitive skills. Although previous research has highlighted the need for multimodal investigations on older musicians to assess positive effects of musical activity during aging, neuroimaging studies are lacking. Our study discovered that older musicians (mean age, 70.8 years; musical experience, 52.7 years; age at the commencement of musical instrument training, 8.6 years) had larger bilateral GMVs in Crus I of the cerebellum than age-matched non-musicians (Figure 2a). Moreover, we demonstrated that cerebellar (Crus I) GMVs in the regions of interest of non-musicians sharply decreased with age, whereas volume and age were not significantly correlated in musicians (Figure 2b) [179]. These findings indicate that lifelong practice of a musical instrument is associated with structural maintenance of the cerebellum in old age. Interestingly, a recent study has reported that reduction in GMV of Crus I is associated with executive dysfunction [180]. This suggests that structural changes in Crus I play an important role in the maintenance of motor planning and monitoring [181], which are needed for skillful execution of finger movement during music-making.
Figure 2.
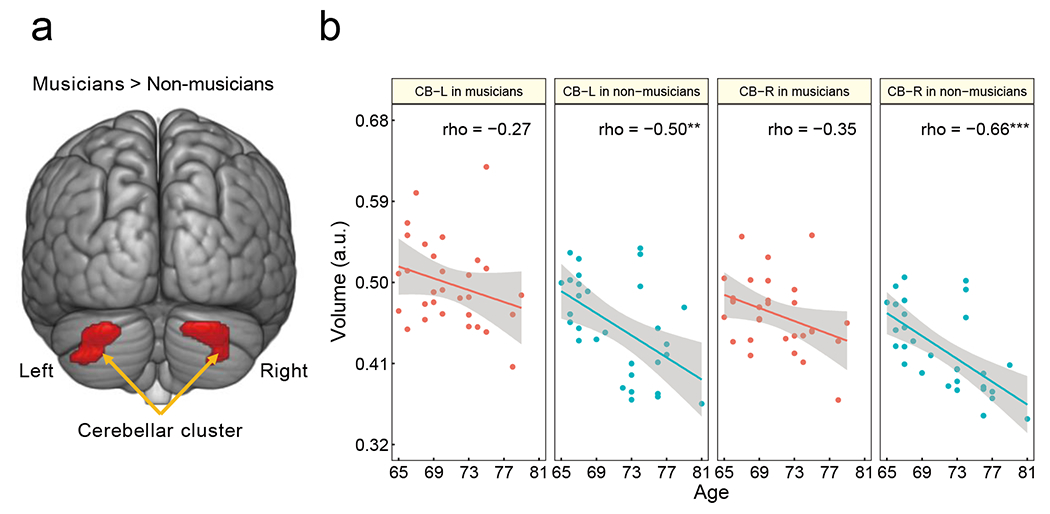
Cerebellar changes related to musical instrument experience [partial modification from [179]. (a) Older musicians had higher bilateral GMVs in the cerebellum than non-musicians. (b) For the cerebellar ROIs, the non-musicians demonstrated a negative correlation between their GMV and age, whereas such correlation was not significant. ** P < 0.01, *** P < 0.001. CB-L, left cerebellum; CB-R, right cerebellum; GMV, gray matter volume; ROI, region of interest; a.u., arbitrary units
fMRI studies provide additional support for the specificity of music-related cerebellum activation in musicians. Musician-specific activation of the cerebellum, as well as auditory-related areas, was observed during passive listening to piano melodies [181], suggesting a degree of audio-motor transformation when listening to music. Moreover, some fMRI studies have reported that activation of the cerebellum was observed in imagery tasks involving specific timing and sequential finger coordination, such as imagery of playing the piano and a finger-tapping task [182–183], suggesting a role of the cerebellum in the imagery of music-related finger movements. Furthermore, our task-related FC analyses revealed that older musicians possessed greater cerebellar–hippocampal FC during melodic working memory tasks, which was correlated with tapping speed [179]. Thus, musicians may encode melodies to sequences of finger movements for working memory maintenance, considering that the right hippocampus has been implicated in melodic retrieval [184] and memory based-music imagery [185]. The implication of these findings is that such a music imagery-related network would be strengthened in musicians through musical instrument training.
In conclusion, the characteristics of musicians may be underscored by their vigorous musical training with complex physical and mental operations, such as the motor imagery of accurate sound, high speed and skillful execution of finger movement to realize melodies and musical impressions, and memorization of long musical phrases. Such extensive and time-pressured effort may be associated with cortical reorganization including motor-related regions in musicians. In particular, lifelong active engagement in musical instrument training is related to structural and functional advantages in the neural system involving the cerebellum.
14. Ageing and cerebellar reserve (Hiroshi Mitoma, Shinji Kakei, Mario Manto)
Cerebellar reserve is defined as the capacity of the cerebellum for compensation and restoration following pathological changes affecting this part of the brain [186–189]. In other words, cerebellar reserve constitutes remarkable inherent forces of resilience to cerebellar pathologies and ability for recovery. This description dates back to the classic paper by Sir G. Holmes [190]. Two types of cerebellar reserve have been described based on the nature of the pathoetiology of cerebellar injury. First, when the etiology elicits immediate structural damage in a limited area of the cerebellum (e.g., stroke or traumatic injury), the ensuing cerebellar functional deficit can be restored through compensation by other cerebellar areas unaffected by the structural damage (structural cerebellar reserve) [187]. On the other hand, when the etiology weakens cerebellar neurons (as well as glial cells) in diverse regions (e.g., degenerative cerebellar ataxias (CAs), immune-mediated CAs, and metabolic/toxic CAs), it gradually leads to cell death, and the affected lesion itself can replenish deteriorating cerebellar functions (functional cerebellar reserve) [187].
There is little or no information on the effects of age on cerebellar reserve. The aim of this section is to describe possible mechanisms underlying the resistance to ageing and ageing-related pathologies and the decline in ageing within the frame of the cerebellar reserve.
14.1. Physiology underlying cerebellar reserve
The concept of reserve has been proposed first in other degenerative diseases, such as cognitive reserve in AD [191–192] and motor reserve in Parkinson’s disease [193]. Compared with these types of reserve, cerebellar reserve is probably unique by showing outstanding capacities for compensation and restoration following cerebellar injury. Such capacity depends on cerebellar-specific structural and functional features that are lacking in other parts of the central nervous system (CNS).
About 60-80% of the whole brain’s 85-100 billion neurons are located in the cerebellum, although the cerebellum forms only about 10% of the brain mass [194–195]. The vast cerebellar neuronal networks are functionally organized to act as internal forward models. The motor domain integrates various inputs to predict the state of the future apparatus for a command [196–199]. The cerebellum is organized to operate as an internal model machine supported by two distinctive inherent mechanisms in the cerebellar neural circuits: “redundant afferents to the microzone” and “multiple forms of synaptic plasticity.”
Indeed, the cerebellum integrates two lines of inputs necessary for an internal forward model, conveyed through MFs: 1) an efference copy (copy of a motor command) from the controller; and 2) a sensory feedback signal that describes the slightly past (~100 ms) state of the motor apparatus [198]. Notably, the branching patterns of individual MFs are intensively divergent, especially along the mediolateral axis [200], leading to the convergence of vast numbers of combinations of multiple MF inputs into the microzone, the rostro-caudal strip of the cerebellar cortex [201]. On the other hand, the divergent forms of plasticity in the cerebellar cortex cooperate synergistically to create the optimal output for any behavior [202]. A separate role for each form of plasticity in motor learning has also been proposed [203].
In essence, cerebellar circuitry is enriched in plasticity mechanisms and multimodal cerebral and peripheral inputs are integrated through multiple plastic modifications at widely distributed synapses, which allows the organization of the internal forward model. While these unique characteristics underlie cerebellar learning, they also provide the cerebellum at the same time with the ability to reorganize any damage to the internal forward model.
14.2. Cerebellar reserve-related resilience to ageing and neurodegenerative diseases
The cerebellum is connected with nearly all regions of cerebral association and paralimbic areas [204–206]. Each cerebellar region shows highly correlated patterns of activation to the corresponding cortical region [24]. In this context, cerebellar structural and functional disorders are not only associated with motor deficits, but are also associated with degradation of thought and affect, clinically known as the cerebellar cognitive affective syndrome (Schmahmann syndrome) [117, 208].
Evidence suggests that the cerebellum ages more slowly than other brain regions [62]. In addition, earlier research showed that amyloid deposition in the cerebellum occurred late in AD with the rare presence of neurofibrillary tangles [131, 209]. Another study showed milder synapse loss in the cerebellum compared with the hippocampus and frontal cortex in patients with AD [210]. Thus, the cerebellum seems to be resistant to developing specific pathology in the early stages of AD. Given that the cerebellum is relatively unaffected in early AD, it might may play a role in compensation for AD-related deficits [62]. Consistently, accumulating evidence points to differences between various brain regions (mainly, the cerebral cortex) and the cerebellum with regard to changes in their structures and functions in diseases that impact cognition and affect, such as AD [8, 125], frontotemporal dementia [125], Parkinson’s disease [211], autism spectrum disorder [212], schizophrenia [213], and MDD [145].
In a Consensus paper, Guell and Schmahmann pointed out the existence of cerebellar cognitive resilience and recovery in cerebellar-linked disorders [187]. They termed these mechanisms cerebellar cognitive reserve [187]. The cerebellum can compensate for ageing-related and cortically-based cognitive declines and deficits by reorganizing the internal forward model through redundant afferents and multiple forms of cerebellar plasticity, as in motor domains.
14.3. Ageing-related decline in cerebellar reserve
Cerebellar reserve is also affected by ageing. Whereas total cerebellar cortical volume decreases by 10.8% over the life span, total cerebellar white matter decreases by 25.9%, and the latter is attributed to the loss of Purkinje and granule cells [214]. Disruption in the connectivity of the cortico-cerebellar resting state network was also reported in older adults [44]. Consistently, age-related changes are found in cerebellar functions, timing, and postural controls [55]. Notably, adaptation speed becomes slower in prism adaptation of ball slowing [215–216] and prism adaptation of hand-reaching [217].
Since motor learning abilities and cerebellar reserve are two sides of the same coin, the above studies point to an age-related decline in cerebellar reserve. The latter also means the disappearance of the time-lag between cerebral and cerebellar disorders, in which the cerebellum escapes the damage in normal ageing and early AD.
14.4. Therapeutic strategies: use of time-lag
One possible therapeutic strategy is to maximize the time-lag between cerebral and cerebellar disorders and potentiate cerebellar reserve. For example, non-invasive cerebellar stimulation has been applied for potentiation of cerebellar reserve (see also next section) [189]. Some beneficial results with this technique have been reported [218], where the plasticity of cerebellar cortical synapses is assumed to be the neural basis for the long-lasting modulation [219]. Interestingly, larger volumes of cerebellar lobules IV, VI and VIIIB are positively correlated with a positive outcome following a stroke, independent of the severity of initial impairment, age and lesion volume, whereas the total volume of the cerebellum is not associated with the outcome following the vascular injury [220]. These observations highlight a heterogeneity within the cerebellum in terms of resilience. Further molecular and transplantation techniques are also needed to manipulate the re-organization of redundant cerebellar afferents using synaptic plasticity. Globally, the cerebellum is now considered a critical source of scaffolding or reserve for cortical function in ageing, but studies are needed to clarify the secrets of resilience [69]. Techniques of optimization of resilience and recruitment of key lobules within the cerebellum need to be identified in the elderly in order to improve the management of the numerous cerebellar disorders encountered in the clinic.
15. Cerebellar transcranial direct current stimulation (ctDCS) and the aging cerebellum (Zeynab Rezaee)
The cerebellum and cerebral cortex have extensive reciprocal connectivity [221] and different cerebellar regions are involved in various cognitive and motor networks. The volume loss and morphological changes in the cerebellum as a result of aging negatively affect cognitive functions such as working memory, processing speed, spatial processing, and long-term memory [39, 222–223]. Moreover, aging in the cerebellum is associated with a decline in motor performance, including gait and balance [224]. The aging brain compensates for decreased performance by recruiting additional neural resources, enabling older adults to maintain higher performance levels (see also previous section). However, a considerable decline in both motor and cognitive performance is still observable. Therefore, further investigations of therapeutic approaches to improve brain function in older adults are needed. One way to help the brain use more neural connectivity and modulate neural excitability is using electric stimulation of the brain via transcranial direct current stimulation (tDCS), a non-invasive brain stimulation technique that modulates neural activity via electrical stimulation. Depending on the direction of the current, the stimulation cause excitation or inhibition in neural activity [225]. Cerebellar tDCS (ctDCS) has been found to affect the learning and performance of cognitive and motor tasks [226–227]. It has also been proposed that ctDCS can be beneficial in the aging brain by modulating the cerebro-cerebellar network and improving the cerebellum’s function as an adjuvant treatment to address age-related cerebellar motor and cognitive decline effectively [39].
In the motor domain, anodal ctDCS has been shown to positively affect motor learning and adaptation in older adults. Anodal ctDCS decreased the reaction time of only older adults in a study of manual dexterity and motor learning [228]. Another study [229] noted that simultaneous postural training and bilateral anodal ctDCS had therapeutic effects in older adults with high fall risk. Investigators did not observe any significant positive results in postural training or bilateral anodal ctDCS alone. In a healthy aging sample, Hardwrick and Celnik (2014) showed that age-related declines in motor adaptation can be diminished by cerebellar tDCS stimulation [230]. Motor adaptation improvement as a result of applying anodal ctDCS was also reported by Weightman et al. (2020) [231]. However, the positive effect was only shown in the motor adaptation of arm movement but not hand movement [231]. However, results have not been consistent. Hulst et al. (2017) found no effect of anodal ctDCS on force field reach adaptation in both controls and patients with cerebellar degeneration [232], and other studies have reported either no impact or adverse effects on motor learning in older healthy adults [233]. While research in the cognitive domain is less extensive than motor investigations, the Cerebellum plays an essential and supportive role in cognitive learning and performance via connections between the posterior cerebellum and prefrontal cortex. The effectiveness of ctDCS in the cognitive domain has been demonstrated in studies using different paradigms, i.e., semantic processing, language, visuospatial attention, verbal working memory, and implicit categorization [234]. However, the work on the cognitive domain is notably less than on motor investigations, and most of the ctDCS effect on cognitive learning has been reported on the younger adult population. Given the outcome variability, the effectiveness of ctDCS in the aging brain in improving cognitive and motor domains needs further investigation.
Some factors influencing outcome variability are specific to the methodology, i.e., study design, tasks, divergent sample size, and stimulation parameters (intensity, duration, electrode size, electrode placement (see below)), and others are specific to aging. Different regions of the cerebellum shrink differently during aging, impacting task performance and electrical properties. It has been shown that the posterior and anterior vermis [235–236] and the superior posterior cerebellum (i.e., Lobules Crus I, Crus II, VI, and VIIb) [237–239] display pronounced age-related volume change. Based on neuroimaging studies [129], different groups of cerebellar lobules are involved in various tasks. For example, lobules IV and V of the anterior cerebellum contribute to force-field perturbations. In contrast, lobule VI is important in visuomotor adaptation [240], executive functioning tasks depend on Crus II and lobule VIIb, and spatial adaptation is related to pontocerebellum stimulation (for more, see [39], [222]). Therefore, individual age-related morphological change of the cerebellar lobules as cerebellar functional units must be considered in defining task-based ctDCS protocol for the aging population.
As GMV declines with age, the volume of CSF in the subarachnoid space increases. This increase in scalp-cortex distance influence current dispersion. Parazzini and colleagues found that individual anatomical variability impacted electrical field distributions [241]. Other aging morphological changes affecting current dispersion include skull thickness and head size. Computational modeling can aid in predicting the effect of these factors on tDCS response. In a computational modeling study [242] analyzing 20 healthy subjects, the electric field strength and focality showed the magnitude of the electric field in the targeted cerebellar area decreased with increasing distance between the targeted gray matter and the scalp. The same study reported an improvement in the field focality when scalp–cortex distance decreased by positioning the electrodes below the occipital cortex. Hence, using methods to evaluate scalp-cortex distance, i.e., structural MRI scans, may reduce variability and allow future studies to adjust the current intensity.
Another source of inter- and intra-individual variability in ctDCS results is electrode montage. To modulate neural activity, in the majority of experimental studies, two electrodes have been used to apply a weak direct current to the cerebellum. In the three common montages for ctDCS, the active electrode can be placed over one cerebellar hemispheres, 1-2 cm below and 3-4 cm lateral to the inion or in the midline to target whole cerebellum [242–244]. The location of the return electrode can be over 1) the buccinator muscle, 2) ipsilateral shoulder, or 3) the contralateral supraorbital muscle [242, 245–246]. In our computational modeling study [243], we showed that using the same electrode montage across different individuals for ctDCS in various aging groups led to differences in the maximum electric field, electric field distribution, and stimulation patterns. Therefore, subject-specific montages can partially address inter-individual variability. Computational modeling conducted before stimulation is an effective method for individualizing ctDCS protocols, providing effective dose control of the intensity of the applied stimulation. In our computational studies, one comparing different montages [247] and another assessing the impact of age on a single montage [243], we showed that various electrode montages affect local and network activity differently. In the aging cerebellum, one must also consider the additional neural connectivity from the recruitment of other cerebellar areas and networks to maintain performance when selecting a target and to propose a stimulation protocol. For example, increasing excitability by ctDCS in one area can lead to inhibition in other brain areas that are part of the same functional network [248]. Therefore, optimizing targeting by minimizing electric field distribution outside the desired location is important in ctDCS investigations and may reduce outcome variability.
Inter-individual variability effects can emerge due to task dependency. An individual’s baseline task performance is effective in the outcome effects of the ctDCS [228]. It has been postulated that a more significant ctDCS-induced impact might be expected in older adults as the task is likely to be more challenging and motivating to the elderly [228]. Thus, further research to compare the effects of ctDCS on high-functioning and lower-functioning older adults seems necessary.
Although numerous studies reported well-founded results on the normal brain’s electric stimulation, a prediction of this approach on older adults’ brains still needs further investigation [225]. The possible harmful effects of tDCS on older adults’ brains is a crucial consideration [249]. Despite existing evidence of the benefits of ctDCS in the rehabilitation of motor function and cognitive performance, evidence from the investigation of electric field dosing and distribution in different computational studies [250–252] indicate that ctDCS parameters need to be modified for the older adults brain to establish a ctDCS protocol for individuals with brain atrophy. Computational modeling studies can facilitate the investigation and prediction of electric field and current density distribution across cerebellar regions and lobules [251, 253].
The subject and task-specific optimization in defining stimulation parameters for ctDCS is an excellent consideration in its clinical potential. The individual differences in the cerebellar cortex’s anatomical and physiological characteristics (e.g., folding structure and orientation of neuronal population), cerebellar lobules’ contribution to different tasks, and the reciprocal connection of different cerebellar regions to multiple cortical areas can lead to a variety of ctDCS protocols is based on the target population needs. The possible harmful effects of ctDCS on older adults’ brains because of certain age-specific variables (e.g., gray matter shrinkage) are a crucial consideration. Despite existing evidence of the benefits of ctDCS in improving motor function and cognitive performance, the dosing parameters need to be modified for the elderly population to apply an unharmful current intensity. The computational analysis and optimization of electric field focality and current intensity before clinical study help configure age-related requirements for the ctDCS protocol. Future studies may focus on maintaining comparable lobule-specific dosing across subjects. Multimodal studies which combine modeling prior to clinical application, behavioural measurement of ctDCS effects, and neuroimaging approach to assessing functional changes help to understand better the age-related changes in current flow at the targeted region based on tissue resistivity and head anatomy. Additionally, lobule-specific electric field and current flow information for the task-specific ctDCS are necessary to estimate the spillover to non-targeted cerebellar lobules in older adults and reduce undesired inter-individual variability in the ctDCS effects.
16. Discussion
The cerebellum mediates numerous functions that are significantly affected by ageing [1]. Hence, there is a need for a unified characterization of how cerebellar adaptation shapes motor, perceptual, and cognitive processes, to maintain efficient behaviour in older adults. With the quickly rising population of older adults, a better understanding of the effects of ageing on the brain becomes more and more important, considering that ageing impacts on motor, cognitive and affective operations. Throughout this Consensus Paper, the panel of experts discuss the current knowledge on the effects of ageing on cerebellar circuitry and cerebellar functions, attempting to clarify up the contributions of the cerebellum in motor/cognitive/affective operations during ageing and in prevalent disorders such as AD or MDD.
In the field of dementia, methodological developments to characterize individuals more comprehensively, and to examine whether these effects are modified by the presence of AD pathology in the cerebral cortex and possibly cerebellum are necessary. Increases in neural activation can be mediated by beta-amyloid, starting from the earliest stages of the disease, but two important methodological challenges need to be resolved to better understand the relationship between beta-amyloid and compensatory activation and to investigate the possibility of translating this new knowledge to therapeutic interventions. First, current imaging methods that visualize the spatial deposition of AD neuropathology detect fibrillar forms, but not the diffuse plaques which are present in the cerebellum early in the disease. In vivo methods will need to be developed that measure human nonfibrillar forms of beta-amyloid to investigate whether the magnitude of diffuse amyloid in the cerebellum correlates with cognitive and cerebral changes. Second, increased neuronal activity in cerebral cortex is associated with both protective and detrimental effects, and it is likely that similar observations may be made in the cerebellum. In the cognitive field, it has been observed that the cerebellar volume is reduced in AD [61]. GMV loss in AD appears distinct from that observed in normal ageing: it starts initially in cerebellar posterior lobe regions and is associated with neuronal, synaptic and beta-amyloid neuropathology. There is growing evidence for a contribution of the right posterior cerebellar atrophy in AD progression. The lateralization of atrophy in AD differs from the bilateral atrophy in healthy ageing [84]. The panel of experts considers that the cerebellum is thus not viewed anymore as an innocent bystander in AD [125].
Another point of Consensus is that the cerebellum is a site of structural and functional changes with ageing, and older patients (including older adults without disease) present clinical symptoms reminiscent of acute cerebellar injury [57]. In terms of motor control, there is an agreement that the cerebellum is a key-player for timing operations and that motor control is impaired in the elderly. Neuroimaging studies demonstrate that the cerebellar regions show a reduction in volume and that cerebellar circuitry is involved in the PCDS observed in older subjects [58] with a correlation between global cerebellar volume and gait speed [59]. Studies on the cerebellar-hippocampal interactions support the hypothesis that cerebellum contributes to the implementation and the efficient use of spatial representations [77]. Age-dependent differences have been also demonstrated in cerebello-striatal FC [46]. In fact, reduced FC has been observed in elderly adults compared to young adults, equally impacting both motor and cognitive cerebello-cerebral modules [44]. This connectivity has also been linked to behavioural performance. Thus, cerebellum GMV, cerebello-cerebral FC, and hippocampus-amygdala-cerebellum neurocircuit might constitute neuroimaging biomarkers for healthy aging. We underline the importance of functional MRI, also targeting the cerebellum, in tracking normal aging and the stage and intensity of the neurodegenerative process in different age-related conditions. Data from resting state fMRI studies presented here evidence that FC can be used as a biomarker to disentangle normal aging from age-related pathological conditions. Since functional brain changes are thought to precede structural brain change [254], the functional damage in certain brain areas may result from a “pre-symptomatic condition”, i.e., the functional damage (reduced FC) may precede the clinical manifestation.
17. Conclusion and perspectives
In summary, compelling evidence shows that the cerebellum participates in cognitive tasks, and also undergoes age-related decline in function, yet a definitive mechanistic link between cerebellar function and cognitive decline remains elusive. Future work should elucidate the role of cerebellum in cognitive decline with age, and whether interventions targeting the cerebellum can reduce the attentional burden on cortical regions in cognitive decline.
There is currently insufficient information regarding the prevalence of cerebellar related dysfunction in ageing and dementia. This may be addressed by the development of measures sensitive to cerebellar function and dysfunction, including assessments of context dependent speed of information processing, sequencing, temporal order, associative learning, and automaticity. This could include computerized remote tools and machine learning methods that identify patterns reflecting cerebellar dysfunction in pre-symptomatic individuals.
Neuroimaging and neuropathological data reveal that grey matter volume loss in AD appears to be distinct from that seen in normal ageing, occurs initially in cerebellar posterior lobe regions that support the cognitively relevant cerebro-cerebellar intrinsic connectivity networks, and is associated with neuronal, synaptic, and beta-amyloid neuropathology, providing support for the contention that the cerebellum is no innocent bystander in AD [125]. Given that AD neuropathologic changes can be present in cognitively normal older individuals, and that not every brain of older individuals with memory impairments harbors elevated beta-amyloid deposits, it will be important for future studies to carefully characterize the etiologic processes of both the unimpaired and impaired populations. This will provide a spatiotemporal model of changes within the cerebellum during disease progression, and advance our understanding of a potential cerebellar contribution to the heterogeneity of clinical phenotypes of AD. Furthermore, collaborative studies capitalizing on physiological and optogenetic methods to study the relationship between neuronal codes or firing patterns (e.g., changes in tonic versus burst firing) and the direction of change of motor and cognitive performance in animal models will be key to developing physiologic interventions in translational research studies in humans that aim to delay AD-related cognitive decline.
Spatial navigation offers an integrative framework to uncover relationships between distinct cerebellar functions and ageing. In particular, the coupling of cerebellar neuroimaging and behavioural assessment of spatial navigation under naturalistic conditions can provide a promising perspective, by leveraging the recent technological advances in mobile brain imaging [255–256] and cerebellar functional near-infrared spectroscopy (fNIRS) recordings [257]. This approach would allow neuro-behavioural biomarkers to be identified, in order to differentiate age-related motor and cognitive deficits and to develop personalized rehabilitation programs.
It remains to be shown whether cerebellar structural and functional changes in healthy ageing, mild cognitive impairment, and dementia have a primarily positive or negative impact on behavior. Correlational studies described in this Consensus paper highlight the relevance of cerebellar structure and function in young vs older subjects and in demented vs cognitively intact individuals, but they do not determine whether these changes are pathological/detrimental or compensatory/beneficial in nature. The underlying underpinnings require also a clarification. Interventional studies that modify cerebellar physiology, such as noninvasive cerebellar stimulation studies, may help clarify this question. The importance of assessing the effectiveness of new interventions and treatments specifically targeting the cerebellum and the development of rehabilitation protocols combining MRI approach with non-invasive cerebellar stimulation should be underlined. According to the rehabilitation potentiality of the Non Invasive Brain stimulation (NIBS) techniques [258–259], further research in the future could be crucial to evaluate neuromodulation approaches to improve functions mediated by the cerebellum [234]. In particular, previous research findings have already demonstrated that non-invasive neurostimulation modulates a brain network rather than just the local stimulation site [259] and that ctDCS can induce neurophysiological changes in the cerebello-brain interaction [243]. This emphasizes the therapeutic and clinical implications of this approach potentially extensible to different pathological conditions. Targeting the regions of cerebellum affected age-related changes may therefore prove an important target for treatments to slow or reverse cognitive decline. In support of this hypothesis, recent work shows that bilateral transcranial magnetic stimulation of Crus II increases cognitive performance in patients with Alzheimer’s disease by strengthening FC networks with nodes of the prefrontal cortex [70]. Cerebellar resources might be important for cerebral cortical processing and it is possible that degraded cerebellar output disrupts cortical activation and FC [260]. NIBS might be applied to counter-act these remote detrimental effects.
Techniques aiming to improve cerebellar reserve are particularly relevant not only to reduce the impacts of ageing on the functioning of a healthy ageing cerebellum but also to promote recovery after injury. The interactions between cerebellum, cerebral cortex and subcortical nodes such as basal ganglia are dynamic across lifespan as a result of variable computational demands between newborns, young children and the elderly [261]. This should be taken into account in future studies addressing the effects of ageing on cerebellar functions. Models of motor/cognitive/affective/social ageing should incorporate the cerebellar circuitry [69].
The panel of experts considers that areas of future research/next steps should include:
the validation of the role of cerebellum and its related neurocircuit in age-related functional declines in larger, multi-ethnic cohorts
methods to unravel the cerebellum biological age for personalized, precision analysis and evaluate whether this measurement could serve as an unhealthy biomarker as well as a treatment target
more direct tests of new models of cerebellum in cognitive aging, such as those outlined by Bernard [69]
timeline of change and senescence in cerebellum – a closer look at mid-life and incorporation of impacts of sex differences/menopause (research indicating sex steroid hormones impact cerebellar network dynamics during menstrual cycle – may also play a role in menopause)
investigations and incorporation of cerebello-thalamo-cortical white matter tracts to have a more integrated network approach
further studies on AD – this is an emerging literature though consensus here highlights the need for additional detailed and targeted work in this domain
the question of the roles of WMH in depression remains unanswered and deserves further studies
in terms of epigenetics, the question of the impact of hyper- and hypomethylation of the DNA within the cerebellum and gene expression needs to be clarified.
Fundings
KS and MY are supported by the Japan Society for the Promotion of Science through a Grant-in-Aid for Scientific Research (21H04422).
VMD is supported by the National Institute on Aging (U19AG073172, U19AG065169, R21AG077307), the National Science Foundation (2112455), and the Alzheimer’s Association (AARG-NTF-21-852145).
JAB is supported by the National Institute on Aging (R01AG064010).
Abbreviations:
- AD
Alzheimer’s disease
- APP
amyloid precursor protein
- CES-D
Center of epidemiology studies depression scale
- ctDCS
Cerebellar transcranial direct current stimulation
- DAN
Dorsal attention network
- DMN
Default mode network
- FC
Functional connectivity
- fNIRS
Functional near-infrared spectroscopy
- GMV
Grey matter volume
- ILAS
I-LAN longitudinal ageing study
- MCI
Mild cognitive impairment
- MF
Mossy fiber
- NIBS
Non invasive brain stimulation
- PCDS
Physio-cognitive decline syndrome
- PS
Processing speed
- SN
Salience Network
- SNPs
Single-nucleotide polymorphisms
- VFM
Volume fraction myelin
- WMH
White matter hyperintensities
Footnotes
Conflicts of interest
The authors declare no conflict of interest.
Data availability
The data shown in the Figures are available on request.
References
- 1.Manto M, Gruol DL, Schmahmann JD, Koibuchi N, Sillitoe RV. Handbook of Cerebellum and Cerebellar Disorders. Springer, 2022 [Google Scholar]
- 2.Van Overwalle F, Manto M, Cattaneo Z, Clausi S, Ferrari C, Gabrieli JDE, Guell X, Heleven E, Lupo M, Ma Q, Michelutti M, Olivito G, Pu M, Rice LC, Schmahmann JD, Siciliano L, Sokolov AA, Stoodley CJ, van Dun K, Vandervert L, Leggio M. Consensus Paper: cerebellum and social cognition. Cerebellum 2020;19(6):833–868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bareš M, Apps R, Avanzino L, Breska A, D’Angelo E, Filip P, Gerwig M, Ivry RB, Lawrenson CL, Louis ED, Lusk NA, Manto M, Meck WH, Mitoma H, Petter EA. Consensus paper: decoding the contributions of the cerebellum as a time machine. from neurons to clinical applications. Cerebellum 2019;18(2):266–286 [DOI] [PubMed] [Google Scholar]
- 4.Bostan AC, Strick PL. The basal ganglia and the cerebellum: nodes in an integrated network. Nat Rev Neurosci 2018;19(6):338–350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.D’Angelo E. Physiology of the cerebellum. Handb Clin Neurol 2018;154:85–108 [DOI] [PubMed] [Google Scholar]
- 6.GBD 2017 US Neurological Disorders Collaborators, Feigin VL, Vos T, Alahdab F, Amit AML, Bärnighausen TW, Beghi E, Beheshti M, Chavan PP, Criqui MH, Desai R, Dhamminda Dharmaratne S, Dorsey ER, Wilder Eagan A, Elgendy IY, Filip I, Giampaoli S, Giussani G, Hafezi-Nejad N, Hole MK, Ikeda T, Owens Johnson C, Kalani R, Khatab K, Khubchandani J, Kim D, Koroshetz WJ, Krishnamoorthy V, Krishnamurthi RV, Liu X, Lo WD, Logroscino G, Mensah GA, Miller TR, Mohammed S, Mokdad AH, Moradi-Lakeh M, Morrison SD, Shivamurthy VKN, Naghavi M, Nichols E, Norrving B, Odell CM, Pupillo E, Radfar A, Roth GA, Shafieesabet A, Sheikh A, Sheikhbahaei S, Shin JI, Singh JA, Steiner TJ, Stovner LJ, Wallin MT, Weiss J, Wu C, Zunt JR, Adelson JD, Murray CJL. Burden of neurological disorders across the US from 1990-2017: a global burden of disease study. JAMA Neurol 2021;78(2):165–176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hou Y, Dan X, Babbar M, Wei Y, Hasselbalch SG, Croteau DL, Bohr VA. Ageing as a risk factor for neurodegenerative disease. Nat Rev Neurol 2019;15(10):565–581 [DOI] [PubMed] [Google Scholar]
- 8.Jacobs HIL, Hopkins DA, Mayrhofer HC, Bruner E, van Leeuwen FW, Raaijmakers W, Schmahmann JD. The cerebellum in Alzheimer’s disease: evaluating its role in cognitive decline. Brain 2018;141(1):37–47 [DOI] [PubMed] [Google Scholar]
- 9.Buhusi CV, Meck WH. What makes us tick? Functional and neural mechanisms of interval timing. Nat Rev Neurosci 2005;6(10):755–65 [DOI] [PubMed] [Google Scholar]
- 10.Wencil EB, Coslett HB, Aguirre GK, Chatterjee A. Carving the clock at its component joints: neural bases for interval timing. J Neurophysiol 2010;104(1):160–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Buonomano DV, Maass W. State-dependent computations: spatiotemporal processing in cortical networks. Nat Rev Neurosci 2009;10(2):113–25 [DOI] [PubMed] [Google Scholar]
- 12.Filip P, Lošák J, Kašpárek T, Vaníček J, Bareš M. Neural network of predictive motor timing in the context of gender differences. Neural Plasticity 2016;2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Filip P, Gallea C, Lehéricy S, Bertasi E, Popa T, Mareček R, et al. Disruption in cerebellar and basal ganglia networks during a visuospatial task in cervical dystonia. Mov Disord 2017;32(5):757–68 [DOI] [PubMed] [Google Scholar]
- 14.Bares M, Husarova I, Lungu OV. Essential tremor, the cerebellum, and motor timing: towards integrating them into one complex entity. Tremor Other Hyperkinetic Mov 2012;2:1–9 [PMC free article] [PubMed] [Google Scholar]
- 15.Naccarato M, Calautti C, Jones PS, Day DJ, Carpenter TA, Baron JC. Does healthy aging effect the hemispheric activation balance during paced index-to-thumb opposition task? An fMRI study. Neuroimage 2006;32(3):1250–6 [DOI] [PubMed] [Google Scholar]
- 16.Mattay VS, Fera F, Tessitore A, Hariri AR, Das S, Callicott JH, et al. Neurophysiological correlates of age-related changes in human motor function. Neurology 2002;58(4):630. [DOI] [PubMed] [Google Scholar]
- 17.Park DC, Reuter-Lorenz P. The adaptive brain: aging and neurocognitive scaffolding. Ann Rev Psychol 2009;60:173–96 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jernigan TL, Archibald SL, Fennema-Notestine C, Gamst AC, Stout JC, Bonner J, et al. Effects of age on tissues and regions of the cerebrum and cerebellum. Neurobiol Aging 2001;22(4):581–94 [DOI] [PubMed] [Google Scholar]
- 19.Bernard J, Nguyen AD, Hausman HK, Maldonado T, Ballard HK, Eakin SM, et al. Shaky scaffolding: age differences in cerebellar activation revealed through activation likelihood estimation meta-analysis. bioRxiv 2019;716530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Horvath S, Mah V, Lu AT, Woo JS, Choi OW, Jasinska AJ, et al. The cerebellum ages slowly according to the epigenetic clock. Aging (Albany NY) 2015;7(5):294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Horvath S. DNA methylation age of human tissues and cell types. Genome Biol 2013;14(10):3156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mecocci P, MacGarvey U, Kaufman AE, Koontz D, Shoffner JM, Wallace DC, et al. Oxidative damage to mitochondrial DNA shows marked age-dependent increases in human brain. Ann Neurol 1993;34(4):609–16 [DOI] [PubMed] [Google Scholar]
- 23.Filip P, Gallea C, Lehéricy S, Lungu O, Bareš M. Neural scaffolding as the foundation for stable performance of aging cerebellum. Cerebellum 2019;18(3):500–10 [DOI] [PubMed] [Google Scholar]
- 24.Stoodley CJ, Schmahmann JD. Functional topography in the human cerebellum: a meta-analysis of neuroimaging studies. Neuroimage 2009;44(2):489–501 [DOI] [PubMed] [Google Scholar]
- 25.Filip P, Lungu OV, Bareš M. Dystonia and the cerebellum: A new field of interest in movement disorders? Clin Neurophysiol 2013;124(7):1269–76 [DOI] [PubMed] [Google Scholar]
- 26.Marcián V, Filip P, Bareš M, Brázdil M. Cerebellar dysfunction and ataxia in patients with epilepsy: coincidence, consequence, or cause? Tremor Other Hyperkinetic Mov 2016; 6:376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Koziol LF, Budding D, Andreasen N, D’Arrigo S, Bulgheroni S, Imamizu H, Yamazaki T. Consensus Paper: the cerebellum’s role in movement and cognition. Cerebellum 2014; 13:151–77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kansal K, Yang Z, Fishman AM, Sair HI, Ying SH, Jedynak BM, Onyike CU. Structural cerebellar correlates of cognitive and motor dysfunctions in cerebellar degeneration. Brain 2016;140. doi: 10.1093/brain/aww327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Langner R, Eickhoff SB. Sustaining attention to simple tasks: a meta-analytic review of the neural mechanisms of vigilant attention. Psychol Bull. 2013;139(4):870–900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.E KH, Chen SH, Ho MH, Desmond JE. A meta-analysis of cerebellar contributions to higher cognition from PET and fMRI studies. Hum Brain Mapp. 2014;35(2):593–615 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Roberts RD, Stankov L. Individual differences in speed of mental processing and human cognitive abilities: troward a taxonomic model. Learning Individ Diff 1999; 11. doi: 10.1016/S1041-6080(00)80007-2 [DOI] [Google Scholar]
- 32.Habeck C, Gazes Y, Razlighi Q, Steffener J, Brickman A, Barulli D, Stern Y. The reference ability neural network study: life-time stability of reference-ability neural networks derived from task maps of young adults. Neuroimage 2016; 125. doi: 10.1016/j.neuroimage.2015.10.077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Habas C. Functional connectivity of the cognitive cerebellum. Frontiers System Neurosci 2021; 15. doi: 10.3389/fnsys.2021.642225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Forn C, Ripollés P, Cruz-Gómez AJ, Belenguer A, González-Torre JA, Ávila C. Task-load manipulation in the symbol digit modalities test: an alternative measure of information processing speed. Brain Cognition 2013;82. doi: 10.1016/j.bandc.2013.04.003 [DOI] [PubMed] [Google Scholar]
- 35.Silva PHR, Spedo CT, Baldassarini CR, Benini CD, Ferreira DA, Barreira AA, Leoni RF. Brain functional and effective connectivity underlying the information processing speed assessed by the Symbol Digit Modalities Test. Neuroimage 2019; 184. doi: 10.1016/j.neuroimage.2018.09.080 [DOI] [PubMed] [Google Scholar]
- 36.Wong CHY, Liu J, Lee TMC, Tao J, Wong AWK, Chau BKH, Chan CCH. Fronto-cerebellar connectivity mediating cognitive processing speed. Neuroimage 2021;226. doi: 10.1016/j.neuroimage.2020.117556 [DOI] [PubMed] [Google Scholar]
- 37.Gao M, Wong CHY, Huang H, Shao R, Huang R, Chan CCH, Lee TMC. Connectome-based models can predict processing speed in older adults. Neuroimage 2020; 223. doi: 10.1016/j.neuroimage.2020.117290 [DOI] [PubMed] [Google Scholar]
- 38.Wong CHY, Liu J, Tao J, Chen LD, Yuan HL, Wong MNK, Xu YW, Lee TMC, Chan CCH. Causal influences of salience/cerebellar networks on dorsal attention network subserved age-related cognitive slowing. GeroScience 2022; 10.1007/s11357-022-00686-1. Advance online publication. 10.1007/s11357-022-00686-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bernard JA, Seidler RD. Moving forward: Age effects on the cerebellum underlie cognitive and motor declines. Neurosci Biobehav Rev 2014; 42. doi: 10.1016/j.neubiorev.2014.02.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Eckert MA. Slowing down: age-related neurobiological predictors of processing speed. Frontiers Neurosci 2011;5. doi: 10.3389/fnins.2011.00025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Chapter Ramnani N. 10 Automatic and controlled processing in the corticocerebellar system. Progr Brain Res 2014;210. doi: 10.1016/B978-0-444-63356-9.00010-8 [DOI] [PubMed] [Google Scholar]
- 42.Strick PL, Dum RP, Fiez JA. Cerebellum and nonmotor function. Annu Rev Neurosci. 2009;32:413–434. [DOI] [PubMed] [Google Scholar]
- 43.Pelzer EA, Hintzen A, Goldau M, von Cramon DY, Timmermann L, Tittgemeyer M. Cerebellar networks with basal ganglia: Feasibility for tracking cerebello-pallidal and subthalamo-cerebellar projections in the human brain. Eur J Neurosci 2013;38:3106–3114. [DOI] [PubMed] [Google Scholar]
- 44.Bernard JA, Peltier SJ, Wiggins JL, Jaeggi SM, Buschkuehl M, Fling BW, Kwak Y, Jonides J, Monk CS, Seidler RD. Disrupted cortico-cerebellar connectivity in older adults. Neuroimage 2013; 83: 103–119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Di Martino A, Scheres A, Margulies DS, Kelly AMC, Uddin LQ, Shehzad Z, Biswal B, Walters JR, Castellanos FX, Milham MP. Functional connectivity of human striatum: a resting state fMRI study. Cereb Cortex 2008;18:2735–2747 [DOI] [PubMed] [Google Scholar]
- 46.Hausman HK, Jackson TB, Goen JRM, Bernard JA. From synchrony to asynchrony: cerebellar-basal ganglia functional circuits in young and older adults. Cereb Cortex 2020;30:718–729 [DOI] [PubMed] [Google Scholar]
- 47.Bernard JA, Seidler RD, Hassevoort KM, Benson BL, Welsh RC, Wiggins JL, Jaeggi SM, Buschkuehl M, Monk CS, Jonides J, Peltier SJ. Resting state cortico-cerebellar functional connectivity networks: a comparison of anatomical and self-organizing map approaches. Front Neuroanat. 2012. Aug 10;6:31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jackson TB, Bernard JA. Cerebello-basal ganglia networks and cortical network global efficiency. Cerebellum 2022. Jun 3. doi: 10.1007/s12311-022-01418-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Klostermann EC, Braskie MN, Landau SM, O’Neil JP, Jagust WJ. Dopamine and frontostriatal networks in cognitive aging. Neurobiol Aging 2012; 33:623.e15–623.e24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bernard JA, Ballard HK, Jackson TB. Cerebellar dentate connectivity across adulthood: a large-scale resting state functional connectivity investigation. Cereb Cortex Commun 2021;2 :tgab050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Carp J, Park J, Polk TA, Park DC. Age differences in neural distinctiveness revealed by multi-voxel pattern analysis. Neuroimage 2011;56:736–743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Festini SB, Bernard JA, Kwak Y, Peltier S, Bohnen NI, Müller MLTM, Dayalu P, Seidler RD. Altered cerebellar connectivity in parkinson’s patients ON and OFF L-DOPA medication. Front Hum Neurosci 2015;9:214. doi: 10.3389/fnhum.2015.00214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kelly C, De Zubicaray G, Di Martino A, Copland DA, Reiss PT, Klein DF, Castellanos FX, Milham MP, McMahon K. L-dopa modulates functional connectivity in striatal cognitive and motor networks: A double-blind placebo-controlled study. J Neurosci 2009;29:7364–7378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fearnley JM, Lees AJ. Ageing and Parkinson’s disease : substantia nigra regional selectivity. Brain 1991;114:2283–2301. [DOI] [PubMed] [Google Scholar]
- 55.Hogan MJ. The cerebellum in thought and action: a fronto-cerebellar aging hypothesis. New Ideas Psychol 2004;22, 97–125, doi: 10.1016/j.newideapsych.2004.09.002 [DOI] [Google Scholar]
- 56.Hogan MJ, Staff RT, Bunting BD et al. Cerebellar brain volume accounts for variance in cognitive performance in older adults. Cortex 2011;47, 441–450, doi: 10.1016/j.cortex.2010.01.001 [DOI] [PubMed] [Google Scholar]
- 57.Schmahmann JD. Disorders of the cerebellum: ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J Neuropsychiatry Clin Neurosci 2004;16, 367–378, doi: 10.1176/jnp.16.3.367 [DOI] [PubMed] [Google Scholar]
- 58.Han S, An Y, Carass A, Prince JL, Resnick SM. Longitudinal analysis of regional cerebellum volumes during normal aging. Neuroimage 2020;220, 117062, doi: 10.1016/j.neuroimage.2020.117062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Nadkarni NK, Nunley KA, Aizenstein H, et al. Association between cerebellar gray matter volumes, gait speed, and information-processing ability in older adults enrolled in the Health ABC study. J Gerontol A Biol Sci Med Sci 2014;69, 996–1003, doi: 10.1093/gerona/glt151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kim HJ, Cheong EN, Jo S et al. The cerebellum could serve as a potential imaging biomarker of dementia conversion in patients with amyloid-negative amnestic mild cognitive impairment. Eur J Neurol 2021;28, 1520–1527, doi: 10.1111/ene.14770 [DOI] [PubMed] [Google Scholar]
- 61.Andersen K, Andersen BB, Pakkenberg B. Stereological quantification of the cerebellum in patients with Alzheimer’s disease. Neurobiol Aging 2012;33, 197 e111–120, doi: 10.1016/j.neurobiolaging.2010.06.013 [DOI] [PubMed] [Google Scholar]
- 62.Liang KJ, Carlson ES. Resistance, vulnerability and resilience: A review of the cognitive cerebellum in aging and neurodegenerative diseases. Neurobiol Learn Mem 2020;170, 106981, doi: 10.1016/j.nlm.2019.01.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Locke TM, Soden ME, Miller SM, et al. Dopamine D1 receptor-positive neurons in the lateral nucleus of the cerebellum contribute to cognitive behavior. Biol Psychiatry 2018;84, 401–412, doi: 10.1016/j.biopsych.2018.01.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bernard JA, Leopold DR, Calhoun VD, Mittal VA. Regional cerebellar volume and cognitive function from adolescence to late middle age. Hum Brain Mapp 2015. ;36, 1102–1120, doi: 10.1002/hbm.22690 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Romero JE, Coupe P, Lanuza E, et al. Toward a unified analysis of cerebellum maturation and aging across the entire lifespan: A MRI analysis. Hum Brain Mapp 2021;42, 1287–1303, doi: 10.1002/hbm.25293 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Diedrichsen J, Maderwald S, Küper M, et al. Imaging the deep cerebellar nuclei: a probabilistic atlas and normalization procedure. Neuroimage 2011;54, 1786–1794, doi: 10.1016/j.neuroimage.2010.10.035 [DOI] [PubMed] [Google Scholar]
- 67.Gilissen EP, Leroy K, Yilmaz Z, et al. A neuronal aging pattern unique to humans and common chimpanzees. Brain Struct Funct 2016;221, 647–664, doi: 10.1007/s00429-014-0931-5 [DOI] [PubMed] [Google Scholar]
- 68.Chen J, Cohen ML, Lerner AJ, Yang Y, Herrup K. DNA damage and cell cycle events implicate cerebellar dentate nucleus neurons as targets of Alzheimer’s disease. Mol Neurodegener 2010;5, 60, doi: 10.1186/1750-1326-5-60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bernard JA. Don’t forget the little brain: A framework for incorporating the cerebellum into the understanding of cognitive aging. Neurosci Biobehav Rev 2022;137, 104639, doi: 10.1016/j.neubiorev.2022.104639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Yao Q, Tang F, Wang Y, et al. Effect of cerebellum stimulation on cognitive recovery in patients with Alzheimer disease: A randomized clinical trial. Brain Stimul 2022;15, 910–920 doi: 10.1016/j.brs.2022.06.004 [DOI] [PubMed] [Google Scholar]
- 71.Arleo A, Rondi-Reig L. Multimodal sensory integration and concurrent navigation strategies for spatial cognition in real and artificial organisms. J Integr Neurosci 2007;6:327–66 10.1142/s0219635207001593 [DOI] [PubMed] [Google Scholar]
- 72.Colombo D, Serino S, Tuena C, Pedroli E, Dakanalis A, Cipresso P, et al. Egocentric and allocentric spatial reference frames in aging: A systematic review. Neurosci Biobehav Rev 2017;80:605–21. 10.1016/j.neubiorev.2017.07.012 [DOI] [PubMed] [Google Scholar]
- 73.Lester AW, Moffat SD, Wiener JM, Barnes CA, Wolbers T. The aging navigational system. Neuron 2017;95:1019–35. 10.1016/j.neuron.2017.06.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ramanoël S, Durteste M, Bécu M, Habas C, Arleo A. Differential brain activity in regions linked to visuospatial processing during landmark-based navigation in young and healthy older adults. Front Hum Neurosci 2020;14:552111. 10.3389/fnhum.2020.552111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Igloi K, Doeller CF, Paradis A-L, Benchenane K, Berthoz A, Burgess N, et al. Interaction between hippocampus and cerebellum Crus I in sequence-based but not place-based navigation. Cereb Cortex 2015;25:4146–54. 10.1093/cercor/bhu132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hauser MFA, Heba S, Schmidt-Wilcke T, Tegenthoff M, Manahan-Vaughan D. Cerebellar-hippocampal processing in passive perception of visuospatial change: An ego- and allocentric axis? Hum Brain Mapp 2020;41:1153–66. 10.1002/hbm.24865 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Rondi-Reig L, Paradis A-L, Fallahnezhad M. A liaison brought to light: cerebellum-hippocampus, partners for spatial cognition. Cerebellum 2022;21:826–37. 10.1007/s12311-022-01422-3 [DOI] [PubMed] [Google Scholar]
- 78.Yuan P, Daugherty AM, Raz N. Turning bias in virtual spatial navigation: Age-related differences and neuroanatomical correlates. Biol Psychol 2014;96:8–19. 10.1016/j.biopsycho.2013.10.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Daugherty AM, Raz N. A virtual water maze revisited: two-year changes in navigation performance and their neural correlates in healthy adults. Neuroimage. 2017;146:492–506. 10.1016/j.neuroimage.2016.09.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Moffat SD, Elkins W, Resnick SM. Age differences in the neural systems supporting human allocentric spatial navigation. Neurobiol Aging 2006;27:965–72. 10.1016/j.neurobiolaging.2005.05.011 [DOI] [PubMed] [Google Scholar]
- 81.Antonova E, Parslow D, Brammer M, Dawson GR, Jackson SHD, Morris RG. Age-related neural activity during allocentric spatial memory. Memory 2009;17(2):125–43. 10.1080/09658210802077348 [DOI] [PubMed] [Google Scholar]
- 82.Ramanoël S, Durteste M, Perot V, Habas C, Arleo A. An appraisal of the role of the neocerebellum for spatial navigation in healthy aging. Cerebellum 2023;22(2):235–239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mirino P, Pecchinenda A, Boccia M, Capirchio A, D’Antonio F, Guariglia C. Cerebellum-cortical interaction in spatial navigation and its alteration in dementias. Brain Sci 2022;12. https://www.mdpi.com/2076-3425/12/5/523 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Gellersen HM, Guell X, Sami S. Differential vulnerability of the cerebellum in healthy ageing and Alzheimer’s disease. Neuroimage Clin 2021;30:102605. 10.1016/j.nicl.2021.102605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.World Health Organization. Decade of healthy ageing: baseline report. World Health Organization, Geneva, Switzerland; 2021. https://cdn.who.int/media/docs/default-source/mca-documents/decade-of-healthy-ageing-baseline-report_06012021.pdf. Accessed 25 June 2021. [Google Scholar]
- 86.Panza F, Lozupone M, Solfrizzi V, Sardone R, Dibello V, Di Lena L, D’Urso F, Stallone R, Petruzzi M, Giannelli G, Quaranta N, Bellomo A, Greco A, Daniele A, Seripa D, Logroscino G. Different cognitive frailty models and health- and cognitive-related outcomes in older age: from epidemiology to prevention. J Alzheimers Dis 2018;62:993–1012. doi: 10.3233/JAD-170963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Robertson DA, Savva GM, Kenny RA. Frailty and cognitive impairment—a review of the evidence and causal mechanisms. Ageing Res Rev 2013;12:840–851. doi: 10.1016/j.arr.2013.06.004 [DOI] [PubMed] [Google Scholar]
- 88.Zheng L, Li G, Gao D, Wang S, Meng X, Wang C, Yuan H, Chen L. Cognitive frailty as a predictor of dementia among older adults: A systematic review and meta-analysis. Arch Gerontol Geriatr 2020;87:103997. doi: 10.1016/j.archger.2019.103997 [DOI] [PubMed] [Google Scholar]
- 89.Chen LK, Woo J, Assantachai P, Auyeung TW, Chou MY, Iijima K, Jang HC, Kang L, Kim M, Kim S, Kojima T, Kuzuya M, Lee JSW, Lee SY, Lee WJ, Lee Y, Liang CK, Lim JY, Lim WS, Peng LN, Sugimoto K, Tanaka T, Won CW, Yamada M, Zhang T, Akishita M, Arai H. Asian Working Group for Sarcopenia: 2019 Consensus Update on Sarcopenia Diagnosis and Treatment. J Am Med Dir Assoc 2020;21:300–307.e2. doi: 10.1016/j.jamda.2019.12.012 [DOI] [PubMed] [Google Scholar]
- 90.Chung CP, Lee WJ, Peng LN, Shimada H, Tsai TF, Lin CP, Arai H, Chen LK. Physio-cognitive decline syndrome as the phenotype and treatment target of unhealthy aging. J Nutr Health Aging. 2021;25(10):1179–1189. doi: 10.1007/s12603-021-1693-4. [DOI] [PubMed] [Google Scholar]
- 91.Shimada H, Doi T, Lee S, Makizako H, Chen LK, Arai H. Cognitive Frailty Predicts Incident Dementia among Community-Dwelling Older People. J Clin Med 2018;7:250. doi: 10.3390/jcm7090250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Tsutsumimoto K, Doi T, Nakakubo S, Kim M, Kurita S, Ishii H, Shimada H. Cognitive Frailty as a Risk Factor for Incident Disability During Late Life: A 24-Month Follow-Up Longitudinal Study. J Nutr Health Aging 2020;24:494–499. doi: 10.1007/s12603-020-1365-9 [DOI] [PubMed] [Google Scholar]
- 93.Chen WT, Chou KH, Liu LK, Lee PL, Lee WJ, Chen LK, Wang PN, Lin CP. Reduced cerebellar gray matter is a neural signature of physical frailty. Hum Brain Mapp 2015;36:3666–3676. doi: 10.1002/hbm.22870 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Liu LK, Chou KH, Hsu CCH, Peng LN, Lee WJ, Chen WT, Lin CP, Chung CP, Wang PN, Chen LK. Cerebellar-limbic neurocircuit is the novel biosignature of physio-cognitive decline syndrome. Aging (Albany NY) 2020;12:25319–25336. doi: 10.18632/aging.104135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Nishita Y, Nakamura A, Kato T, Otsuka R, Iwata K, Tange C, Ando F, Ito K, Shimokata H, Arai H. Links Between Physical Frailty and Regional Gray Matter Volumes in Older Adults: A Voxel-Based Morphometry Study. J Am Med Dir Assoc 2019;20:1587–1592.e7. doi: 10.1016/j.jamda.2019.09.001 [DOI] [PubMed] [Google Scholar]
- 96.Bettio LEB, Rajendran L, Gil-Mohapel J. The effects of aging in the hippocampus and cognitive decline. Neurosci Biobehav Rev. 2017. Aug;79:66–86. doi: 10.1016/j.neubiorev.2017.04.030. [DOI] [PubMed] [Google Scholar]
- 97.Tedesco AM, Chiricozzi FR, Clausi S , Lupo M, Molinari M, Leggio MG. The cerebellar cognitive profile. Brain 2011;134: 3672–3686 [DOI] [PubMed] [Google Scholar]
- 98.Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, Williamson A, Dahle C, Gerstorf D, Acker JD. Regional brain changes in aging healthy adults: general trends, individual differences and modifiers. Cereb Cortex 2005;15:1676–1689 [DOI] [PubMed] [Google Scholar]
- 99.Damoiseaux JS, Beckman CF, Sanz Arigita EJ, Barkhof F, Scheltens Ph, Stam CJ, Rombouts SARB. Reduced resting-state brain activity in the “default network” in normal aging. Cerebral Cortex 2008;18:1856–1864. [DOI] [PubMed] [Google Scholar]
- 100.Andrews-Hanna JR, Snyder AZ, Vincent JL, Lustig C, Head D, Raichle ME, Buckner RL. Disruption of large-scale brain systems in advanced aging. Neuron 2007;56:924–935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Wu T, Zang Y, Wang L, Long X, Hallett M, Chen Y, Chan P. Aging influence on functional connectivity of the motor network in the resting state. Neurosci Lett 2007;42:164–168. [DOI] [PubMed] [Google Scholar]
- 102.Habas C, Kamdar N, Nguyen D, Prater K, Beckmann CF, Menon V, Greicius MD. Distinct cerebellar contributions to intrinsic connectivity networks. J Neurosci 2009;29:8586–8594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.O’Reilly JX, Beckmann CF, Tomassini V, Ramnani N, Johansen-Berg H. Distinct and overlapping functional zones in the cerebellum defined by resting state functional connectivity. Cerebral Cortex 2010;20:953–965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Maguire EA, Frith CD. Aging affects the engagement of the hippocampus during autobiographical memory retrieval. Brain 2003;126:1511–1523 [DOI] [PubMed] [Google Scholar]
- 105.Reitz C, Brayne C, Mayeux R. Epidemiology of Alzheimer’s disease. Nat Rev Neurol 2011; 7: 137–152. doi: 10.1038/nrneurol.2011.2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Camicioli RM. Mild motor impairment: motor change preceding mild cognitive impairment and dementia. Alzheimers Dementia 2010;6, S92. doi: 10.1016/j.jalz.2010.05.282 [DOI] [Google Scholar]
- 107.Bruchhage MMK, Correia S, Malloy P, Salloway S, Deoni S. Machine learning classification identifies cerebellar contributions to early and moderate cognitive decline in Alzheimer’s disease. Frontiers Aging Neurosci 2020;12, 524024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Ito M. The Cerebellum: Brain for an implicit self. Upper Saddle River: New Jersey, 2012 [Google Scholar]
- 109.Stoodley CJ, Desmond JE, Schmahmann JD. Functional topography of the human cerebellum revealed by functional neuroimaging studies. In: Handbook of the cerebellum and cerebellar disorders. Dordrecht: Springer Netherlands, 2013, pp. 1717–1751 [Google Scholar]
- 110.Leto K, Arancillo M, Becker EBE, Buffo A, Chiang C, et al. Consensus paper: cerebellar development. Cerebellum 2016;15(6), 789–828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Tiemeier H, Lenroot RK, Greenstein DK, Tran L, Pierson R, Giedd JN. Cerebellar development during childhood and adolescence: a longitudinal morphometric MRI study. NeuroImage 2010; 49(1), 67–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Gauvrit T, Benderradji H, Buee L, Blum D, Vieau D. Early-Life Environment Influence on Late-Onset Alzheimer’s Disease. Frontiers Cell Dev Biol 2022;10, 834661–834661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Serra L, Cercignani M, Lenzi D, Perri R, Fadda L, Caltagirone C, Macaluso E, Bozzali M. Grey and white matter changes at different stages of Alzheimer’s disease. J Alzheimers Dis 2010;19(1):147–59 [DOI] [PubMed] [Google Scholar]
- 114.Sullivan EV, Pfefferbaum A. Diffusion tensor imaging and aging. Neurosci Biobehav Rev 2006;30(6), 749–761 [DOI] [PubMed] [Google Scholar]
- 115.Limperopolous C, Chilingaryan G, Sullivan N, Guizard N, Robertson RL, du Plessis AJ. Injury to the premature cerebellum: outcome is related to remote cortical development. Cereb Cortex 2014; 24(3), 728–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Piller S. Blots on a field? Science 2022. ; 377 (6604). doi: 10.1126/science.ade0209. [DOI] [PubMed] [Google Scholar]
- 117.Schmahmann JD, Sherman JC. The cerebellar cognitive affective syndrome The cerebellar cognitive affective syndrome. Brain 1998; 141(May 1998):561–79 [DOI] [PubMed] [Google Scholar]
- 118.Schmahmann JD, Guell X, Stoodley CJ, Halko MA. The Theory and Neuroscience of Cerebellar Cognition. Annu Rev Neurosci 2019; 42(1): 337–364 [DOI] [PubMed] [Google Scholar]
- 119.Frangou S, Modabbernia A, Williams SCR, Papachristou E, Doucet GE, et al. Cortical thickness across the lifespan: Data from 17,075 healthy individuals aged 3–90 years. Hum. Brain Mapp 2022; 43(1):431–451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Hoogendam YY, van der Geest JN, van der Lijn F, van der Lugt A, Niessen WJ, et al. Determinants of cerebellar and cerebral volume in the general elderly population. Neurobiol Aging 2012;33(12):2774–2781 [DOI] [PubMed] [Google Scholar]
- 121.Buhrmann A, Brands AMA, van der Grond J, Schilder C, van der Mast RC, et al. Cerebellar Grey Matter Volume in Older Persons Is Associated with Worse Cognitive Functioning. Cerebellum 2021;20:9–20 [DOI] [PubMed] [Google Scholar]
- 122.Chaudhari K, Wang L, Kruse J, Winters A, Sumien N, et al. Early loss of cerebellar Purkinje cells in human and a transgenic mouse model of Alzheimer’s disease. Neurol Res 2021. ;43(7):570–581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Mavroudis I, Petridis F, Kazis D, Njau S, Costa V, Baloyannis S. Purkinje cells pathology in Alzheimer’s disease. Am J Alzheimers Dis Other Demen 2019;34(7):439–49 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Mavroudis IA, Fotiou DF, Adipepe LF, Manani MG, Njau SD, et al. Morphological changes of the human purkinje cells and deposition of neuritic plaques and neurofibrillary tangles on the cerebellar cortex of Alzheimer’s disease. Am J Alzheimers Dis Other Demen 2010;25(7):585–591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Guo CC, Tan R, Hodges JR, Hu X, Sami S, Hornberger M. Network-selective vulnerability of the human cerebellum to Alzheimer’s disease and frontotemporal dementia. Brain 2016;139(5):1527–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Lin CY, Chen CH, Tom SE, Kuo SH. Cerebellar Volume Is Associated with Cognitive Decline in Mild Cognitive Impairment: Results from ADNI. Cerebellum 2020;19(2):217–225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Pagen LHG, Van De Ven VG, Gronenschild EHBM, Priovoulos N, Verhey FRJ, Jacobs HIL. 2020. Contributions of cerebro-cerebellar default mode connectivity patterns to memory performance in mild cognitive impairment. J Alzheimer Dis 2020;75(2):633–647 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Guell X. Functional Gradients of the Cerebellum: a Review of Practical Applications. Cerebellum 2022; 21(6):1061–72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Guell X, Schmahmann J. Cerebellar Functional Anatomy: a Didactic Summary Based on Human fMRI Evidence. Cerebellum 2020;19:1–5 [DOI] [PubMed] [Google Scholar]
- 130.Guell X, Schmahmann J, Gabrieli J, Ghosh S. Functional gradients of the cerebellum. Elife 2018; 7:e36652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Braak H, Braak E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol 1991;82(4), 239–259 [DOI] [PubMed] [Google Scholar]
- 132.Nelson PT, Alafuzoff I, Bigio EH, Bouras C, Braak H, et al. Correlation of Alzheimer disease neuropathologic changes with cognitive status: A review of the literature. J Neuropathol Exp Neurol 2012;71:362–381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Cole G, Neal JW, Singhrao SK, Jasani B, Newman GR. The distribution of amyloid plaques in the cerebellum and brain stem in Down’s syndrome and Alzheimer’s disease: a light microscopical analysis. Acta Neuropathol 1993;85(5):542–552 [DOI] [PubMed] [Google Scholar]
- 134.Ghisays V, Lopera F, Goradia DD, Protas HD, Malek-Ahmadi MH, et al. PET evidence of preclinical cerebellar amyloid plaque deposition in autosomal dominant Alzheimer’s disease-causing Presenilin-1 E280A mutation carriers. NeuroImage Clin 2021; 31:102749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Wang H, D’Andrea M, Nagele R. Cerebellar diffuse amyloid plaques are derived from dendritic Abeta42 accumulations in Purkinje cells. Neurobiol Aging 2002; 23(2):213–23 [DOI] [PubMed] [Google Scholar]
- 136.Shimada H, Minatani S, Takeuchi J, Takeda A, Kawabe J, et al. Heavy tau burden with subtle amyloid beta accumulation in the cerebral cortex and cerebellum in a case of familial alzheimer’s disease with app osaka mutation. Int J Mol Sci. 2020; 21(12):4443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Jacobs HIL, Becker JA, Kwong K, Engels-Domínguez N, Prokopiou PC, et al. In vivo and neuropathology data support locus coeruleus integrity as indicator of Alzheimer’s disease pathology and cognitive decline. Sci Transl Med 2021;13(612):eabj2511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Del Cerro I, Villarreal MF, Abulafia C, Duarte-Abritta B, Sánchez SM, et al. Disrupted functional connectivity of the locus coeruleus in healthy adults with parental history of Alzheimer’s disease. J Psychiatr Res 2020;123:81–88 [DOI] [PubMed] [Google Scholar]
- 139.Olivito G, Serra L, Marra C, Di Domenico C, Caltagirone C, et al. Cerebellar dentate nucleus functional connectivity with cerebral cortex in Alzheimer’s disease and memory: a seed-based approach. Neurobiol Aging 2020;89:32–40 [DOI] [PubMed] [Google Scholar]
- 140.Tang F, Zhu D, Ma W, Yao Q, Li Q, Shi J. Differences changes in cerebellar functional connectivity between mild cognitive impairment and Alzheimer’s disease: a seed-based approach. Front Neurol 2021;12:645171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Zhou Z, Zhu R, Shao W, Zhang SJ, Wang L, et al. Changes in Resting-State Functional Connectivity of Cerebellum in Amnestic Mild Cognitive Impairment and Alzheimer’s Disease: A Case-Control Study. Front Syst Neurosci 2021;15:596221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Woodruff-Pak DS. Eyeblink Classical Conditioning Differentiates Normal Aging from Alzheimer’s Disease. Integr. Physiol Behav Sci 2001;36(2):87–108 [DOI] [PubMed] [Google Scholar]
- 143.Woodruff-Pak DS, Foy MR, Akopian GG, Lee KH, Zach J, et al. Differential effects and rates of normal aging in cerebellum and hippocampus. Proc Natl Acad Sci (USA) 2010;107(4): 1624–1629 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Schmahmann JD, Weilburg JB and Sherman JC. The neuropsychiatry of the cerebellum - insights from the clinic. Cerebellum 2007: 6:254–67. doi 10.1080/14734220701490995 [DOI] [PubMed] [Google Scholar]
- 145.Depping MS, Schmitgen MM, Kubera KM and Wolf RC. Cerebellar Contributions to Major Depression. Front Psychiatry 2018: 9:634. doi 10.3389/fpsyt.2018.00634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Grieve SM, Korgaonkar MS, Koslow SH, Gordon E and Williams LM. Widespread reductions in gray matter volume in depression. NeuroImage Clin 2013: 3:332–9. doi 10.1016/j.nicl.2013.08.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Lai CH and Wu YT. The gray matter alterations in major depressive disorder and panic disorder: Putative differences in the pathogenesis. J Affective Disord 2015: 186:1–6. doi 10.1016/j.jad.2015.07.022 [DOI] [PubMed] [Google Scholar]
- 148.Xu LY, Xu FC, Liu C, Ji YF, Wu JM, Wang Y, Wang HB and Yu YQ. Relationship between cerebellar structure and emotional memory in depression. Brain Behavior 2017: 7:e00738. doi 10.1002/brb3.738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Zhang YN, Li H, Shen ZW, Xu C, Huang YJ and Wu RH. Healthy individuals vs patients with bipolar or unipolar depression in gray matter volume. World J Clin Cases 2021: 9:1304–17. doi 10.12998/wjcc.v9.i6.1304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Depping MS, Wolf ND, Vasic N, Sambataro F, Hirjak D, Thomann PA and Wolf RC. Abnormal cerebellar volume in acute and remitted major depression. Progr Neuro-psychopharmacol Biol Psych 2016: 71:97–102. doi 10.1016/j.pnpbp.2016.06.005 [DOI] [PubMed] [Google Scholar]
- 151.Depping MS, Nolte HM, Hirjak D, Palm E, Hofer S, Stieltjes B, Maier-Hein K, Sambataro F, Wolf RC and Thomann PA. Cerebellar volume change in response to electroconvulsive therapy in patients with major depression. Progr Neuro-psychopharmacol Biol Psych 2017: 73:31–5. doi 10.1016/j.pnpbp.2016.09.007 [DOI] [PubMed] [Google Scholar]
- 152.Bogoian HR, King TZ, Turner JA, Semmel ES and Dotson VM. Linking depressive symptom dimensions to cerebellar subregion volumes in later life. Translat Psychiatry 2020: 10:201. doi 10.1038/s41398-020-00883-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Li J, Gong H, Xu H, Ding Q, He N, Huang Y, Jin Y, Zhang C, Voon V, Sun B, Yan F and Zhan S. Abnormal voxel-wise degree centrality in patients with late-life depression: a resting-state functional magnetic resonance imaging study. Front Psychiatry 2019: 10:1024. doi 10.3389/fpsyt.2019.01024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Liu F, Hu M, Wang S, Guo W, Zhao J, Li J, Xun G, Long Z, Zhang J, Wang Y, Zeng L, Gao Q, Wooderson SC, Chen J and Chen H. Abnormal regional spontaneous neural activity in first-episode, treatment-naive patients with late-life depression: a resting-state fMRI study. Progr Neuro-psychopharmacol Biol Psych 2012: 39:326–31. doi 10.1016/j.pnpbp.2012.07.004 [DOI] [PubMed] [Google Scholar]
- 155.Avedisova AS, Samotaeva IS, Luzin RV, Semenovyh NS, Sergunova KA, Akzhigitov RG and Zakharova RV. [Apathy in depression: a morphometric analysis]. Zh Nevrol Psikhiatr Im S S Korsakova 2019: 119:141–7. doi 10.17116/jnevro2019119051141 [DOI] [PubMed] [Google Scholar]
- 156.Salo KI, Scharfen J, Wilden ID, Schubotz RI and Holling H. Confining the Concept of Vascular Depression to late-onset depression: a meta-analysis of MRI-defined hyperintensity burden in major depressive disorder and bipolar disorder. Front Psychol 2019: 10:1241. doi 10.3389/fpsyg.2019.01241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Ataullah AHM, Naqvi IA. Cerebellar dysfunction. StatPearls. Treasure Island (FL); 2022, Aug 22. [Google Scholar]
- 158.Sen P, Shah PP, Nativio R, Berger SL. Epigenetic mechanisms of longevity and aging. Cell 2016;166:822–839. doi: 10.1016/j.cell.2016.07.050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Flurkey K, Papaconstantinou J, Miller RA, Harrison DE. Lifespan extension and delayed immune and collagen aging in mutant mice with defects in growth hormone production. Proc Natl Acad Sci (USA). 2001;98: 6736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Choi H, Joe S, Nam H. Development of tissue-specific age predictors using DNA methylation data. Genes 2019;10. doi: 10.3390/genes10110888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.El Khoury LY, Gorrie-Stone T, Smart M, Hughes A, Bao Y, Andrayas A, et al. Systematic underestimation of the epigenetic clock and age acceleration in older subjects. Genome Biol 2019;20: 1–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, Klotzle B, Bibikova M, Fan JB, Gao Y, Deconde R, Chen M, Rajapakse I, Friend S, Ideker T, Zhang K. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell 2013;49: 359–367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Li X, Li W, Xu Y. Human age prediction based on DNA methylation using a gradient boosting regressor. Genes . 2018;9. doi: 10.3390/genes9090424 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Bryant P, Elofsson A. The relationship between ageing and changes in the human blood and brain methylomes. NAR Genom Bioinform 2022;4. doi: 10.1093/nargab/lqac001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Reynolds LM, Taylor JR, Ding J, Lohman K, Johnson C, Siscovick D, et al. Age-related variations in the methylome associated with gene expression in human monocytes and T cells. Nat Commun 2014;5. doi: 10.1038/ncomms6366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Lu AT, Hannon E, Levine ME, Hao K, Crimmins EM, Lunnon K, et al. Genetic variants near MLST8 and DHX57 affect the epigenetic age of the cerebellum. Nat Commun 2016;7. doi: 10.1038/ncomms10561 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Jin Z, Liu Y. DNA methylation in human diseases. Genes Diseases 2018;5:1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Slieker RC, Relton CL, Gaunt TR, Slagboom PE, Heijmans BT. Age-related DNA methylation changes are tissue-specific with ELOVL2 promoter methylation as exception. Epigenetics Chromatin 2018;11: 1–11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Moore LD, Le T, Fan G. DNA Methylation and Its Basic Function. Neuropsychopharmacol 2012;38: 23–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Seidler RD, Bernard JA, Burutolu TB, Fling BW, Gordon MT, Gwin JT, et al. Motor control and aging: links to age-related brain structural, functional, and biochemical effects. Neurosci Biobehav Rev 2010;34: 721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Lu AT, Quach A, Wilson JG, Reiner AP, Aviv A, Raj K, et al. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging 2019;11. doi: 10.18632/aging.101684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Raz N, Ghisletta P, Rodrigue KM, Kennedy KM, Lindenberger U. Trajectories of brain aging in middle-aged and older adults: regional and individual differences. Neuroimage 2010;51(2):501–11. 10.1016/j.neuroimage.2010.03.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Ramanoël S, Hoyau E, Kauffmann L, Renard F, Pichat C, Boudiaf N, et al. Gray matter volume and cognitive performance during normal aging. A voxel-based morphometry study. Front Aging Neurosci 2018;10:235. 10.3389/fnagi.2018.00235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Nyberg L, Lövdén M, Riklund K, Lindenberger U, Bäckman L. Memory aging and brain maintenance. Trends Cogn Sci 2012;16(5):292–305. 10.1016/j.tics.2012.04.005 [DOI] [PubMed] [Google Scholar]
- 175.Verghese J, Lipton RB, Katz MJ, Hall CB, Derby CA, Kuslansky G, et al. Leisure activities and the risk of dementia in the elderly. N Engl J Med 2003;348(25):2508–16. 10.1056/NEJMoa022252 [DOI] [PubMed] [Google Scholar]
- 176.Zatorre RJ, Chen JL, Penhune VB. When the brain plays music: auditory-motor interactions in music perception and production. Nat Rev Neurosci 2007;8(7):547–58. 10.1038/nrn2152 [DOI] [PubMed] [Google Scholar]
- 177.Hutchinson S, Lee LH, Gaab N, Schlaug G. Cerebellar volume of musicians. Cereb Cortex 2003;13(9):943–49. 10.1093/cercor/13.9.943 [DOI] [PubMed] [Google Scholar]
- 178.Paquette S, Fujii S, Li HC, Schlaug G. The cerebellum’s contribution to beat interval discrimination. Neuroimage 2017;163:177–82. 10.1016/j.neuroimage.2017.09.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Yamashita M, Ohsawa C, Suzuki M, Guo X, Sadakata M, Otsuka Y, et al. Neural advantages of older musicians involve the cerebellum: implications for healthy aging through lifelong musical instrument training. Front Hum Neurosci 2022;15:784026. 10.3389/fnhum.2021.784026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Olivito G, Lupo M, Iacobacci C, Clausi S, Romano S, Masciullo M, et al. Structural cerebellar correlates of cognitive functions in spinocerebellar ataxia type 2. J Neurol 2018;265(3):597–606. 10.1007/s00415-018-8738-6 [DOI] [PubMed] [Google Scholar]
- 181.Baumann S, Koeneke S, Schmidt CF, Meyer M, Lutz K, Jancke L. A network for audio-motor coordination in skilled pianists and non-musicians. Brain Res 2007;1161:65–78. 10.1016/j.brainres.2007.05.045 [DOI] [PubMed] [Google Scholar]
- 182.Hanakawa T, Immisch I, Toma K, Dimyan MA, Van Gelderen P, Hallett M. Functional properties of brain areas associated with motor execution and imagery. J Neurophysiol 2003;89(2):989–1002. 10.1152/jn.00132.2002 [DOI] [PubMed] [Google Scholar]
- 183.Meister IG, Krings T, Foltys H, Boroojerdi B, Müller M, Töpper R, et al. Playing piano in the mind—an fMRI study on music imagery and performance in pianists. Brain Res Cogn Brain Res 2004;19(3):219–28. 10.1016/j.cogbrainres.2003.12.005 [DOI] [PubMed] [Google Scholar]
- 184.Watanabe T, Yagishita S, Kikyo H. Memory of music: roles of right hippocampus and left inferior frontal gyrus. Neuroimage 2008;39(1):483–91. 10.1016/j.neuroimage.2007.08.024 [DOI] [PubMed] [Google Scholar]
- 185.Herholz SC, Coffey EBJ, Pantev C, Zatorre RJ. Dissociation of neural networks for predisposition and for training-related plasticity in auditory-motor learning. Cereb Cortex 2016;26(7):3125–34. 10.1093/cercor/bhv138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Mitoma H, Manto M, Hampe CS. Time is cerebellum. Cerebellum 2018;17(4):387–391 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Mitoma H, Buffo A, Gelfo F, Guell X, Fucà E, Kakei S, et al. Consensus paper. Cerebellar reserve: From cerebellar physiology to cerebellar disorders. Cerebellum 2020;19(1):131–153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Mitoma H, Kakei S, Yamaguchi K, Manto M. Physiology of cerebellar reserve: redundancy and plasticity of a modular machine. Int J Mol Sci 2021;22(9):4777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Manto M, Kakei S, Mitoma H. The critical need to develop tools assessing cerebellar reserve for the delivery and assessment of non-invasive cerebellar stimulation. Cerebellum Ataxias 2021;8(1):2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Holmes G. The symptoms of acute cerebellar injuries due to gunshot injuries. Brain 1917;40:461–535 [Google Scholar]
- 191.Stern Y. Cognitive reserve in ageing and Alzheimer’s disease. Lancet Neurol 2012;11(11):1006–1012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Stern Y. An approach to studying the neural correlates of reserve. Brain Imaging Behav 2017;11(2):410–416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Palmer SJ, Ng B, Abugharbieh R, Eigenraam L, McKeown MJ. Motor reserve and novel area recruitment: Amplitude and spatial characteristics of compensation in Parkinson’s disease. Eur J Neurosci 2009;29(11):2187–2196 [DOI] [PubMed] [Google Scholar]
- 194.Colin F, Ris L, Godaux E. Neuroanatomy of the cerebellum. In The cerebellum and Its Disorders; Manto M, Pandolfo M, Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 6–29 [Google Scholar]
- 195.Walloe S, Pakkenberg B, Fabricius K. Stereological estimation of total cell numbers in the human cerebral and cerebellar cortex. Front Hum Neurosci 2014; 8: 508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Wolpert DM, Ghahramani Z, Jordan MI. An internal model for sensorimotor integration. Science 1995;269(5232):1880–1882 [DOI] [PubMed] [Google Scholar]
- 197.Popa LS, Hewitt AL, Ebner TJ. Purkinje cell simple spike discharge encodes error signals consistent with a forward internal model. Cerebellum 2013;12(3):331–333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Tanaka H, Ishikawa T, Kakei S. Neural evidence of the cerebellum as a state predictor. Cerebellum 2019;18(3):349–371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Tanaka H, Ishikawa T, Lee J, Kakei S. The cerebro-cerebellum as a locus of forward model; A review. Front Syst Neurosci 2020;14:19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Wu HS, Sugihara I, Shinoda Y. Projection patterns of single mossy fibers originating from the lateral reticular nucleus in the rat cerebellar cortex and nuclei. J Comp Neurol 1999;411:97–118 [DOI] [PubMed] [Google Scholar]
- 201.Apps R, Hawkes R, Aoki S, Bengtsson F, Brown AM, Chen G, et al. Cerebellar modules and their role as operational cerebellar processing units: a consensus paper [corrected]. Cerebellum 2018;17(5):654–682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.De Zeeuw CI, Lisberger SG, Raymond JL. Diversity and dynamics in the cerebellum. Nat Neurosci 2021, 24, 160–167 [DOI] [PubMed] [Google Scholar]
- 203.Hirano T. Around LTD hypothesis in motor learning. Cerebellum 2014;13(5):645–50 [DOI] [PubMed] [Google Scholar]
- 204.Schmahmann JD. From movement to thought: Anatomic substrates of the cerebellar contribution to cognitive processing. Hum Brain Mapp 1996;4(3):174–98 [DOI] [PubMed] [Google Scholar]
- 205.Schmahmann JD, Pandya DN. The cerebrocerebellar system. Int Rev Neurobiol 1997;41:31–60 [DOI] [PubMed] [Google Scholar]
- 206.Kelly RM, Strick PL. Cerebellar loops with motor cortex and prefrontal cortex of a nonhuman primate. J Neurosci 2003;23(23):8432–44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Stoodley CJ, Schmahmann JD. Evidence for topographic organization in the cerebellum of motor control versus cognitive and affective processing. Cortex 2010;46(7): 831–844 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Guell X, Hoche F, Schmahmann JD. Metalinguistic deficits in patients with cerebellar dysfunction: empirical support for the dysmetria of thought theory. Cerebellum 2015. Feb;14(1):50–8 [DOI] [PubMed] [Google Scholar]
- 209.Thal DR, Rüb U, Orantes M, Braak H. Phases of A beta-deposition in the human brain and its relevance for the development of AD. Neurology 2002;58(12):1791–800 [DOI] [PubMed] [Google Scholar]
- 210.Blennow K, Bogdanovic N, Alafuzoff I, Ekman R, Davidsson P. Synaptic pathology in Alzheimer’s disease: Relation to severity of dementia, but not to senile plaques, neurofibrillary tangles, or the ApoE4 allele. J Neural Transm 1996;103(5):603–618 [DOI] [PubMed] [Google Scholar]
- 211.Wu T, Hallett M. The cerebellum in Parkinson’s disease. Brain 2013;136(3):696–709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Arnold Anteraper S, Guell X, D’Mello A, Joshi N, Whitfield- Gabrieli S, Joshi G. Disrupted cerebro-cerebellar intrinsic functional connectivity in young adults with high-functioning autism spectrum disorder: A data-driven, whole-brain, high temporal resolution fMRI study. Brain Connect 2019;9(1):48–59 [DOI] [PubMed] [Google Scholar]
- 213.Moberget T, Doan NT, Alnæs D, Kaufmann T, Córdova-Palomera A, Lagerberg TV, et al. Cerebellar volume and cerebellocerebral structural covariance in schizophrenia: a multisite mega-analysis of 983 patients and 1349 healthy controls. Mol Psychiatry 2018;23(6):1512–20 [DOI] [PubMed] [Google Scholar]
- 214.Andersen BB, Gundersen HJ, Pakkenberg B. Aging of the human cerebellum: A stereological study. J Comp Neurol 2003;466(3): 356–365 [DOI] [PubMed] [Google Scholar]
- 215.Fernández-Ruiz J, Hall C, Vergara P, Díaz R. Prism adaptation in normal aging: slower adaptation rate and larger aftereffect. Brain Res Cogn Brain Res 2009;9(3):223–226 [DOI] [PubMed] [Google Scholar]
- 216.King BR, Fogel SM, Albouy G, Doyon J. Neural correlates of the age-related changes in motor sequence learning and motor adaptation in older adults. Front Hum Neurosci 2013;7: 142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Hashimoto Y, Honda T, Matsumura K, Nakao M, Soga K, Katano K, et al. Quantitative evaluation of human cerebellum-dependent motor learning through prism adaptation of hand-reaching movement. PLoS ONE 2015;10(3):e0119376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Van Dun K, Mitoma H, Manto M. Cerebellar cortex as a therapeutic target for neurostimulation. Cerebellum 2018;17(6);777–787 [DOI] [PubMed] [Google Scholar]
- 219.Nuzzo C, Ruggiero F, Cortese F, Cova I, Priori A, Ferrucci R. Non-invasive cerebellar stimulation in cerebellar disorders. CNS Neurol Disord Drug Targets 2018;17(3):193–198 [DOI] [PubMed] [Google Scholar]
- 220.Sadeghihassanabadi F, Frey BM, Backhaus W, Choe CU, Zittel S, Schön G, Bönstrup M, Cheng B, Thomalla G, Gerloff C, Schulz R. Structural cerebellar reserve positively influences outcome after severe stroke. Brain Commun 2022. Aug 4;4(6):fcac203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Pini L, Manenti R, Cotelli M, Pizzini FB, Frisoni GB, Pievani M. Non-invasive brain stimulation in dementia: a complex network story. NDD [Internet]. 2018. [cited 2019 May 28];18:281–301. Available from: https://www.karger.com/Article/FullText/495945 [DOI] [PubMed] [Google Scholar]
- 222.Koppelmans V, Hoogendam YY, Hirsiger S, Mérillat S, Jäncke L, Seidler RD. Regional cerebellar volumetric correlates of manual motor and cognitive function. Brain Struct Function 2017;222:1929–44. 10.1007/s00429-016-1317-7 [DOI] [PubMed] [Google Scholar]
- 223.Reuter-Lorenz PA, Park DC. Human neuroscience and the aging mind: a new look at old problems. J Gerontol B Psychol Sci Soc Sci 2010;65B:405–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Holviala J, Kraemer WJ, Sillanpää E, Karppinen H, Avela J, Kauhanen A, et al. Effects of strength, endurance and combined training on muscle strength, walking speed and dynamic balance in aging men. Eur J Appl Physiol 2012;112:1335–47. 10.1007/s00421-011-2089-7 [DOI] [PubMed] [Google Scholar]
- 225.Woods AJ, Antal A, Bikson M, Boggio PS, Brunoni AR, Celnik P, et al. A technical guide to tDCS, and related non-invasive brain stimulation tools. Clinical Neurophysiol 2016;127:1031–48 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Nitsche MA, Paulus W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J Physiol (Lond) 2000;527:633–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Fregni F, Boggio PS, Nitsche M, Bermpohl F, Antal A, Feredoes E, et al. Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory. Exp Brain Res 2005;166:23–30 [DOI] [PubMed] [Google Scholar]
- 228.Lindberg PG, Verneau M, Boterff QL, Cuenca-Maia M, Baron J-C, Maier MA. Age- and task-dependent effects of cerebellar tDCS on manual dexterity and motor learning–A preliminary study. Neurophysiologie Clin 2022;52(5):354–365 [DOI] [PubMed] [Google Scholar]
- 229.Yosephi MH, Ehsani F, Zoghi M, Jaberzadeh S. Multi-session anodal tDCS enhances the effects of postural training on balance and postural stability in older adults with high fall risk: Primary motor cortex versus cerebellar stimulation. Brain Stim 2018;11:1239–50 [DOI] [PubMed] [Google Scholar]
- 230.Hardwick RM, Celnik PA. Cerebellar direct current stimulation enhances motor learning in older adults. Neurobiol Aging 2014;35:2217–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Weightman M, Brittain J-S, Punt D, Miall RC, Jenkinson N. Targeted tDCS selectively improves motor adaptation with the proximal and distal upper limb. Brain Stim 2020;13:707–16 [DOI] [PubMed] [Google Scholar]
- 232.Hulst T, John L, Küper M, van der Geest JN, Göricke SL, Donchin O, et al. Cerebellar patients do not benefit from cerebellar or M1 transcranial direct current stimulation during force-field reaching adaptation. J Neurophysiol 2017;118:732–48 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Rauscher M, Yavari F, Batsikadze G, Ludolph N, Ilg W, Nitsche MA, et al. Lack of cerebellar tDCS effects on learning of a complex whole body dynamic balance task in middle-aged (50–65 years) adults. Neurol Res Pract 2020;2:38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Oldrati V, Schutter DJLG. Targeting the Human Cerebellum with transcranial direct current stimulation to modulate behavior: a meta-analysis. Cerebellum 2018;17:228–36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Raz N, Torres IJ, Spencer WD, White K, Acker JD. Age-related regional differences in cerebellar vermis observed in vivo. Arch Neurol 1992;49:412–6 [DOI] [PubMed] [Google Scholar]
- 236.Shah SA, Doraiswamy PM, Husain MM, Figiel GS, Boyko OB, McDonald WM, et al. Assessment of posterior fossa structures with midsagittal MRI: The effects of age. Neurobiol Aging 1991;12:371–4 [DOI] [PubMed] [Google Scholar]
- 237.Abe O, Yamasue H, Aoki S, Suga M, Yamada H, Kasai K, et al. Aging in the CNS: Comparison of gray/white matter volume and diffusion tensor data. Neurobiol Aging 2008;29:102–16 [DOI] [PubMed] [Google Scholar]
- 238.Lee JY, Lyoo IK, Kim SU, Jang HS, Lee DW, Jeon HJ, et al. Intellect declines in healthy elderly subjects and cerebellum. Psychiatry Clin Neurosci 2005;59:45–51 [DOI] [PubMed] [Google Scholar]
- 239.Paul R, Grieve SM, Chaudary B, Gordon N, Lawrence J, Cooper N, et al. Relative contributions of the cerebellar vermis and prefrontal lobe volumes on cognitive function across the adult lifespan. Neurobiol Aging 2009;30:457–65 [DOI] [PubMed] [Google Scholar]
- 240.Donchin O, Rabe K, Diedrichsen J, Lally N, Schoch B, Gizewski ER, et al. Cerebellar regions involved in adaptation to force field and visuomotor perturbation. J Neurophysiol 2012;107:134–47 [DOI] [PubMed] [Google Scholar]
- 241.Parazzini M, Rossi E, Ferrucci R, Liorni I, Priori A, Ravazzani P. Modelling the electric field and the current density generated by cerebellar transcranial DC stimulation in humans. Clin Neurophysiol 2014;125:577–84 [DOI] [PubMed] [Google Scholar]
- 242.Ferrucci R, Cortese F, Priori A. Cerebellar tDCS: How to Do It. Cerebellum 2015;14:27–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Galea JM, Jayaram G, Ajagbe L, Celnik P. Modulation of cerebellar excitability by polarity-specific non-invasive direct current stimulation. J Neurosci 2009;29:9115–22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Galea JM, Vazquez A, Pasricha N, Orban de Xivry J-J, Celnik P. Dissociating the roles of the cerebellum and motor cortex during adaptive learning: the motor cortex retains what the cerebellum learns. Cerebral Cortex 2011;21:1761–70 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Rampersad SM, Janssen AM, Lucka F, Aydin Ü, Lanfer B, Lew S, et al. Simulating Transcranial Direct Current Stimulation With a Detailed Anisotropic Human Head Model. IEEE Trans Neural Syst Rehab Eng 2014;22:441–52 [DOI] [PubMed] [Google Scholar]
- 246.Grimaldi G, Argyropoulos GP, Bastian A, Cortes M, Davis NJ, Edwards DJ, et al. Cerebellar transcranial direct current stimulation (ctDCS): a novel approach to understanding cerebellar function in health and disease. Neuroscientist 2016;22:83–97 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Rezaee Z, Dutta A. Cerebellar Lobules Optimal Stimulation (CLOS): a computational pipeline to optimize cerebellar lobule-specific electric field distribution. Frontiers Neurosci 2019;13 :266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Summers JJ, Kang N, Cauraugh JH. Does transcranial direct current stimulation enhance cognitive and motor functions in the ageing brain? A systematic review and meta- analysis. Ageing Res Rev 2016;25:42–54 [DOI] [PubMed] [Google Scholar]
- 249.Habich A, Fehér KD, Antonenko D, Boraxbekk C-J, Flöel A, Nissen C, et al. Stimulating aged brains with transcranial direct current stimulation: opportunities and challenges. Psychiatry Res: Neuroimaging. 2020;306:111179. [DOI] [PubMed] [Google Scholar]
- 250.Klaus J, Schutter DJLG. Electrode montage-dependent intracranial variability in electric fields induced by cerebellar transcranial direct current stimulation. Sci Rep 2021;11:22183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Rezaee Z, Dutta A. Lobule-specific dosage considerations for cerebellar transcranial direct current stimulation during healthy aging: a computational modeling study using age-specific magnetic resonance imaging templates. Neuromodulation 2020;23:341–65 [DOI] [PubMed] [Google Scholar]
- 252.Mahdavi S, Towhidkhah F. Computational human head models of tDCS: Influence of brain atrophy on current density distribution. Brain Stim 2018;11:104–7 [DOI] [PubMed] [Google Scholar]
- 254.Damoiseaux S Jessica. Resting-state fMRI as a biomarker for Alzheimer’s disease? Alzheimers Res Ther 2012; 4(2): 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.De Sanctis P, Solis-Escalante T, Seeber M, Wagner J, Ferris DP, Gramann K. Time to move: Brain dynamics underlying natural action and cognition. Eur J Neurosci 2021; 54(12), 8075–8080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Delaux A, de Saint Aubert JB, Ramanoël S, Bécu M, Gehrke L, Klug M, Chavarriaga R, Sahel JA, Gramann K, Arleo A. Mobile brain/body imaging of landmark-based navigation with high-density EEG. Eur J Neurosci 2021;54(12), 8256–8282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Rocco G, Delaire E, Ramanoël S, Meste O, Magnié-Mauro MN, Grova C, Lebrun J. Densifying optodes montage to enhance cerebellar fNRIS. The Society for functional Near Infrared Spectroscopy (fNIRS), Boston, USA, 2022. [Google Scholar]
- 258.Ferrucci R, Giannicola G, Rosa M, Fumagalli M, Boggio PS, Hallett M, Zago S, Priori A. Cerebellum and processing of negative facial emotions: cerebellar transcranial DC stimulation specifically enhances the emotional recognition of facial anger and sadness. Cogn Emot 2012;26:786–799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.To WT, De Ridder D, Hart J Jr, Vanneste S. Changing brain networks through non-invasive neuromodulation. Front Hum Neurosci 2018;12:128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260.Maldonado T, Jackson TB, Bernard JA. Anodal cerebellar stimulation increases cortical activation: Evidence for cerebellar scaffolding of cortical processing. Hum Brain Mapp 2023;44(4):1666–1682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Beuriat PA, Cristofori I, Gordon B, Grafman J. The shifting role of the cerebellum in executive, emotional and social processing across the lifespan. Behav Brain Funct 2022;18(1):6. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data shown in the Figures are available on request.