

Abstract
Rhoptry-associated protein 1 (RAP1) of Plasmodium falciparum is a potential component of a malaria vaccine. We have expressed in Escherichia coli eight recombinant RAP1 proteins representing almost the entire sequence of the mature protein and assessed the antigenicity of the proteins by immunization of mice. Antisera to six of the recombinant proteins reacted specifically with parasite-derived RAP1 (PfRAP1), as determined by indirect immunofluorescence and by immunoblotting. These proteins were then used in enzyme-linked immunosorbent assays to evaluate human antibody responses to RAP1 during naturally transmitted infections in The Gambia. Immunoglobulin G (IgG) antibodies specifically reactive with the recombinant RAP1 proteins are directed mostly towards fragments containing the N-terminal sequences of mature PfRAP1. The most N-terminal segment (residues 23 to 175) contains only minor epitopes, while major epitopes are outside this region. Antibodies from malaria patients do not compete for a linear epitope recognized by an inhibitory anti-RAP1 monoclonal antibody. Analysis of IgG subclass distribution shows that human anti-RAP1 antibodies are predominantly IgG1.
Rhoptries are club-shaped organelles located at the apical end of the merozoite, the extracellular form of malaria parasites responsible for invasion of erythrocytes. Antigens associated with these organelles are among targets for antimalaria vaccines (19). One such antigen of Plasmodium falciparum is a complex of two noncovalently linked proteins, rhoptry-associated protein 1 (RAP1) and RAP2 (4, 5, 7, 10, 18, 28, 30, 31).
Unlike many other malaria antigens, RAP1 shows minimal genetic polymorphism, and its amino acid sequence is highly conserved between isolates. Full or partial amino acid sequences of the protein deduced from gene sequences obtained from seven distinct isolates of different geographical origins were more than 99% identical, with only nine amino acid substitutions identified (14–16, 28). This suggests that interstrain antigenic diversity may not be a problem for a RAP1-based vaccine.
Experimental immunizations of Saimiri monkeys with affinity-purified RAP1-RAP2 complex conferred partial protection against P. falciparum infection (27). The epitopes responsible for this immunity were not determined. Monoclonal antibodies (MAbs) to conserved linear epitopes of RAP1 inhibit the development of P. falciparum in vitro (13, 31), suggesting that antibodies to this antigen may reduce the replication of the parasite. Since RAP1 is a component of an endotoxin-like exoantigen that stimulates in vitro production of tumor necrosis factor by human mononuclear cells (22), it was proposed that antibodies against RAP1 might protect against the disease by removing the toxin-like exoantigen from circulation.
The knowledge of human immune recognition of RAP1 is inadequate. To date, only four studies of human immune responses to RAP1 during natural malaria infection have been reported. Jakobsen and colleagues (20) showed that lymphocytes from most of 21 Ghanaian donors proliferated in vitro in response to a recombinant protein representing the N-terminal third of RAP1 (amino acids [aa] 23 to 294), suggesting the presence of T-cell epitopes in this region. Sera from these donors also contained antibodies to the recombinant RAP1 (rRAP1). A larger study using the same rRAP1 and sera of 425 Tanzanians living in an area where malaria is holoendemic showed that the proportion of responders increased with age and, in addition, indicated an association between high levels of anti-RAP1 immunoglobulin G (IgG) antibodies and protection against high P. falciparum densities in children (21). A more recent study of 100 Papua New Guineans confirmed that the recognition of RAP1 correlated with age (35). Only one study compared the relative immunogenicities of different regions of RAP1 (15). Testing sera of 26 individuals by immunoblotting for antibodies to rRAP1 antigens and visual scoring of results, the study indicated that most antibodies detectable by this method were against epitopes within an N-terminal region (aa 1 to 122) (15).
The work presented here describes the production and immunological characterization of a new set of rRAP1 proteins and their use in an enzyme-linked immunosorbent assay (ELISA) for evaluation of antibody responses in Gambian malaria patients. We show that although patients have IgG antibodies to an rRAP1 containing the N-terminal sequence from aa 23 to 175, more antibodies are targeted to major epitopes outside this region. The antibodies are mainly of the IgG1 subclass.
MATERIALS AND METHODS
Production of rRAP1 antigens.
To express in Escherichia coli sufficient amounts of rRAP1 proteins, the RAP1 gene of the K1 strain of P. falciparum was modified, without altering the primary amino acid sequence of the protein, as follows: (i) P. falciparum codons rarely used in E. coli were replaced by abundant codons (25), (ii) potential transcriptional terminators were destroyed, and (iii) putative internal ribosomal binding sites were eliminated (reference 37 and unpublished data).
GST fusion proteins.
Two rRAP1 proteins were produced as fusions to the C terminus of glutathione S-transferase (GST) by using pGEX expression vectors (33). Forward primer 5′ TGGATCCATCAATGTAAACGGTGA 3′ and reverse primer 5′ TGAATTCCAGTAGTACTTTCTTCAG 3′ were used to amplify by PCR a DNA fragment (nucleotides 67 to 525) encoding a protein termed C1 in this paper and corresponding to aa 23 to 175 of RAP1 (Fig. 1). BamHI and EcoRI endonuclease restriction sites (underlined) were incorporated into the forward and reverse primers, respectively, to facilitate cloning of the PCR fragment into the pGEX-2T vector. A second set of primers, 5′ CCCCGGGTCTTCTGAAGAAAGTACTA 3′ (forward) and 5′ TGAATTCCTAATGATTCTATAGTATC 3′ (reverse), amplified a DNA fragment (nucleotides 505 to 1098) encoding a protein designated C2 including aa 169 to 366. SmaI and EcoRI restriction sites in these forward and reverse primers, respectively, were designed for cloning of the amplified DNA into the pGEX-3X vector.
FIG. 1.



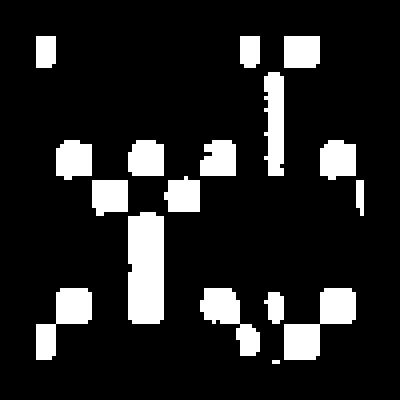



Schematic representation of P. falciparum RAP1 and rRAP1 proteins. C1 and C2 and P2 to P7 are codes for GST and His6 recombinant proteins, respectively. The first and last amino acid residues of RAP1 included in each recombinant protein are indicated. Numbering of amino acid residues is as in reference 28. , region of repeats related to the KSSSPS motif (between aa 123 and 164); , proteolytic cleavage site between residues 190 and 191 yielding p65 product from a mature p80 (26); , inhibitory MAb epitope (LTPLEELYP) (13); , cysteine-rich region (between aa 353 and 616); ▪, His6 tag; , DHFR; , GST.
RAP1 fragments cloned in these expression vectors were then used to transform E. coli TG1. Recombinant clones expressing GST-RAP1 fusion proteins were selected by the small-scale expression method (32).
The GST protein alone was purified from cultures transformed with pGEX-2T vector (without a RAP1 insert) and used as a control antigen in ELISAs and immunoblots.
His6 fusion proteins.
Six rRAP1 proteins (P2 to P7 [Fig. 1]) with a C-terminal His6 tag were produced. DNA fragments encoding the His6 proteins were amplified by PCR from the modified RAP1 gene, except for DNA encoding the P2 protein which was amplified from parasite genomic DNA. DNA fragments encoding proteins P2 to P6 were integrated between the start codon for translation and the BglII site of the pDS56/RBSII-(His)6 expression vector (37). A DNA fragment encoding P7 was ligated into the BglII site of the plasmid pDHFRS-(0/0)-His6 (37) and expressed as a fusion with mouse dihydrofolate reductase (DHFR) at the N terminus. The ligation products were used to transform E. coli W3100, initially transformed with pREP4 repressor plasmid. Recombinant clones expressing rRAP1-His6 proteins were detected by the presence of a novel protein in isopropyl-β-d-thiogalactopyranoside (IPTG)-induced samples that was absent in the corresponding uninduced samples.
DHFR-His6 was purified from E. coli cells transformed with plasmid pDHFRS-(0/0)-His6 and served as a control antigen for the P7 rRAP1.
Sequences of rRAP1 proteins.
DNA sequences of all RAP1 inserts were determined by the dideoxy-mediated chain terminator method (29), and the deduced amino acid sequences of the recombinant proteins were compared to that of RAP1 of the reference K1 strain (28). Recombinant proteins C1, P2, P4, P5, and P6 have amino acid sequences identical to the known RAP1 sequence. Sequences encoding proteins P3, P7, and C2 each contain a single nucleotide change which has led to a different amino acid change in each protein. P3 has a cysteine (codon TGT) in position 406 changed to arginine (CGT). In protein P7, a tyrosine (TAC) in position 580 is changed to phenylalanine (TTC). C2 has an asparagine (AAT) instead of lysine (AAA) in position 260.
Purification of rRAP1.
GST fusion proteins were soluble after sonication of E. coli cells in phosphate-buffered saline (pH 7.4) (PBS) containing 2% Triton X-100 and were affinity purified under nondenaturing conditions by adsorption on glutathione agarose beads (Sigma, Poole, Dorset, United Kingdom) (33).
Attempts to purify the His6 proteins under nondenaturing conditions were not successful. These proteins could not be extracted from E. coli cells lysed in the presence of mild detergents such as 2% Nonidet P-40 or 2% Triton X-100. The His6 proteins could only be first extracted in 6 M guanidine hydrochloride and then purified in the presence of 8 M urea on nickel nitrilotriacetic acid (Ni2+-NTA) agarose (Qiagen) (37). The His6 proteins were eluted in 8 M urea buffer (pH 6.3) containing 300 mM imidazole.
The purity of the recombinant proteins was assessed on Coomassie blue-stained sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels (not shown) and by immunoblotting (Fig. 2). Single protein bands were observed for P6 and P7 (not shown). The other six proteins each contained a major protein and multiple smaller fragments. The proteins and the fragments were revealed by immunoblotting with RAP1-specific MAb 2.29 (7, 13) for those proteins containing the MAb’s target sequence (P2, P3, P4, P5, and C2 [Fig. 2]) and also with mouse antisera raised to the different rRAP1 antigens (not shown). Thus, preparations of P2, P3, P4, P5, C1, and C2 included degradation products of these recombinant proteins.
FIG. 2.

Immunoblot of purified recombinant proteins with RAP1-specific MAb 2.29. The rRAP1 proteins were resolved on an SDS-PAGE gel and electroblotted onto a nitrocellulose membrane, and the membrane was probed with MAb 2.29 and developed with peroxidase-conjugated rabbit anti-mouse immunoglobulin. P2, P3, P4, P5, and C2 reacted with MAb 2.29, specific for the sequence LTPLEELYP contained in these proteins. Note antibody reactivity also with degradation fragments of these proteins. P6, P7, and C1, without the epitope, showed no reactivity with MAb 2.29. The marker lane contained the following prestained molecular weight markers (Bio-Rad): phosphorylase b (107,000), bovine serum albumin (76,000), ovalbumin (52,000), carbonic anhydrase (36,800), soybean trypsin inhibitor (27,200), and lysozyme (19,200).
To resolve whether the degradation of the GST fusions occurred during sonication of E. coli cells and the gentle purification of proteins C1 and C2, protease inhibitors (0.1 mM tosyl-l-lysine chloromethyl ketone, 1 mM phenylmethylsulfonyl fluoride, 0.5% [wt/vol] aprotinin, 50 μg of chymostatin per ml, 50 μg of pepstatin per ml, and 50 μg of leupeptin per ml) were included in the extraction buffer. However, the degradation was still observed, indicating that it had occurred in E. coli before the purification process. This approach was not tested for the His6 proteins, since it was unlikely that any protease activity would be retained under the strong denaturing conditions of their extraction and purification.
The yields of the purified recombinant proteins were determined by use of the Bradford protein assay kit (Bio-Rad, Hertfordshire, United Kingdom) before storage of the proteins at −70°C until required.
Immunogenicity of recombinant proteins.
Mice were immunized with all eight rRAP1 antigens (Fig. 1). The His6 proteins were dialyzed against PBS to remove urea prior to immunization. The dialysis led to precipitation of these proteins. Groups of five 6- to 8-week-old female outbred mice (MF1 strain) were immunized intraperitoneally with 50 μg of a protein in complete Freund’s adjuvant. Primary immunization sera were collected 28 days later. The mice were given boosters of 50 μg of the same proteins in incomplete Freund’s adjuvant on day 35, and secondary immunization sera were collected 13 and 21 days later.
IFA.
P. falciparum K1 (38) was grown by the continuous culture method (39) for use in the indirect immunofluorescence assay (IFA) and immunoblots.
Sera of individual mice obtained after primary and secondary immunizations were titrated by doubling dilutions between 1:50 and 1:12,150. Samples (25 μl) of these diluted sera were tested on acetone-fixed P. falciparum-infected erythrocytes by IFA as described previously (8). RAP1-specific MAb 2.29 (7) served as a positive control.
Immunoblotting.
Schizonts were enriched from in vitro culture by the percoll gradient method (17). A 50-μl sample of packed schizonts was extracted in 200 μl of Laemmli sample buffer (with or without 5% 2-mercaptoethanol) and boiled for 3 min, and the supernatant (15 μl/well) containing parasite proteins was resolved in an SDS–10% PAGE gel. Western blotting of total parasite proteins was carried out as described previously (7), and the blots were probed with mouse antisera to rRAP1 proteins. RAP1-specific MAb 2.29 was used as a positive control to detect RAP1 among the parasite’s proteins.
Immunoblots of 2 μg of purified recombinant proteins were analyzed with the same mouse antisera or MAb 2.29.
Human sera and ELISA.
Sera from 44 patients (1 to 38 years old) presenting with uncomplicated P. falciparum malaria to the Outpatient Department of the Medical Research Council at Fajara, The Gambia, West Africa, were collected during the malaria transmission season from October to December 1983. A 0.3-ml serum sample was obtained from each donor as part of a venous blood sample for other studies (9) after consent, under the approval of the Scientific and Ethical Committees of the Medical Research Council and of the Gambian Government. A positive standard was a pool of sera from adult Gambians immune to malaria; titrations of this standard were performed in each ELISA to control the quality of experiments and interexperimental variation. Sera of 31 healthy adult donors to the Scottish Blood Transfusion Service who had not been exposed to malaria were used as negative controls to determine the specificity of the assay. All sera were stored at −70°C.
rRAP1 or control proteins were diluted to 0.5 μg/ml in coating buffer (0.015 M Na2CO3, 0.035 M NaHCO3; pH 9.6). Wells of a 96-well microtiter plate (Immunolon 4; Dynatech, Billingshurst, United Kingdom) were coated with 100 μl (0.05 μg) of an antigen at 4°C overnight. Wells coated with the GST protein alone served as controls for GST-RAP1 fusions. Wells treated with coating buffer or DHFR-His6 were controls for the P2- to P6- or the P7-His6 rRAP1 proteins, respectively. The ELISA procedure was performed as described elsewhere (6). Human sera were tested at a dilution of 1:500 in duplicate wells, horseradish peroxidase-conjugated rabbit anti-human IgG (Dako Ltd., High Wycombe, United Kingdom) was used diluted 1:5,000, and reactions were developed with ortho-phenylenediamine dihydrochloride (OPD) (Sigma) substrate.
Optical density (OD) values specific for antibody reactivity with rRAP1 proteins were obtained by subtracting averaged OD values of appropriate antigen controls (coating buffer, DHFR-His6, or GST) from averaged OD values of duplicate tests with His6- or GST-fused RAP1 antigens.
Mean OD values plus 2 standard deviations for sera of 31 control individuals who had not been exposed to malaria were taken as negative-to-positive cutoff values. The cutoff values for individual rRAP1 proteins were 0.205 for C1, 0.159 for C2, 0.148 for P2, 0.219 for P3, 0.191 for P4, 0.253 for P5, 0.163 for P6, and 0.199 for P7.
IgG subclass-specific ELISA.
Anti-RAP1 IgG subclasses were determined for sera that pretested positive for IgG antibodies by using polyclonal peroxidase-conjugated sheep immunoglobulins specific for each human IgG subclass (supplied by The Binding Site Ltd., Birmingham, United Kingdom). Specificities and working dilutions of the anti-human IgG1 (AP006), IgG2 (AP007), IgG3 (AP008), and IgG4 (AP009) were determined by titrations of these sera (1:500 to 1:64,000) on purified myeloma proteins [IgG1(κ), IgG2(κ), IgG3(κ), IgG4(κ), and IgM(κ); The Binding Site]. All conjugates were entirely specific for the appropriate IgG subclass and were used at a working dilution of 1:500. Reactivities of thus-diluted subclass-specific sera with purified human myeloma proteins over the concentration range of 1 to 0.032 μg/well were 2.33 to 0.18 for anti-IgG1, 1.30 to 0.11 for anti-IgG2, 2.5 to 0.37 for anti-IgG3, and 2.65 to 0.25 for anti-IgG4. The ELISA procedure was as for total IgG (6) except that human sera were tested for subclasses diluted 1:100.
Competition ELISA.
A competition ELISA was carried out to compare various rRAP1 antigens for epitope similarity. Increasing concentrations (from 0.1 to 40 μg/ml) of antigens used as competitors were preincubated for 5 h with aliquots of human sera diluted 1:500. Subsequently, 100-μl aliquots of the preincubated sera were added to duplicate wells coated with the same rRAP1 or a different rRAP1; the rest of the procedure was done as described above with anti-human IgG conjugate (Dako Ltd.).
RESULTS
Specificity for parasite RAP1 of mouse antisera to recombinant proteins.
Mice were immunized with the purified rRAP1s to show that all eight proteins were immunogenic and to establish the proteins’ similarity to parasite-derived RAP1 (PfRAP1). Antisera to each immunogen recognized the respective rRAP1 in Western blots, and the results of immunoblotting with the homologous proteins were positive (data not shown). Antibodies raised against the rRAP1 antigens were tested for recognition of PfRAP1 by IFA and immunoblotting. IFA was used to assess stage specificity and subcellular distribution of positive reactions. All IFA-positive sera gave reactions typical of PfRAP1, a double-dot rhoptry staining pattern with segmented schizonts and free merozoites and more-diffuse reactions with young ring parasites. The strength of responses induced by different recombinant proteins varied (Table 1). All mice immunized with P3, C1, or C2 responded to primary and secondary immunization by production of antibodies recognizing the parasites in IFA. The responses of mice immunized with P2, P4, or P5 were more variable, with only some animals producing antibodies reactive with the parasites. All sera from mice immunized with P6 or P7 failed to react with parasites.
TABLE 1.
Immunogenicity of recombinant RAP1 proteins in outbred MF1 mice
| Immunizing protein | Mouse no. | Immunofluorescence with schizontsa
|
||
|---|---|---|---|---|
| Primary response, day 28 | Secondary response
|
|||
| Day 13 | Day 21 | |||
| P2 | 1 | − | − | − |
| 2 | − | − | − | |
| 3 | − | − | ++ | |
| 4 | ++ | ++ | ++ | |
| 5 | ++ | +++ | +++ | |
| P3 | 6 | +++ | +++ | +++ |
| 7 | +++ | +++ | +++ | |
| 8 | +++ | +++ | +++ | |
| 9 | +++ | +++ | +++ | |
| 10 | +++ | +++ | +++ | |
| P4 | 11 | − | − | − |
| 12 | + | + | + | |
| 13 | + | + | + | |
| 14 | + | +++ | +++ | |
| 15 | + | +++ | +++ | |
| P5 | 16 | − | − | − |
| 17 | − | − | − | |
| 18 | + | ++ | ++ | |
| 19 | + | ++ | ++ | |
| 20 | +++ | +++ | +++ | |
| P6 | 21 | − | − | − |
| 22 | − | − | − | |
| 23 | − | − | − | |
| 24 | − | − | − | |
| 25 | − | − | − | |
| P7 | 26 | − | − | − |
| 27 | − | − | − | |
| 28 | − | − | − | |
| 29 | − | − | − | |
| 30 | − | − | − | |
| C1 | 31 | + | ++ | ++ |
| 32 | + | +++ | +++ | |
| 33 | + | +++ | +++ | |
| 34 | + | +++ | +++ | |
| 35 | + | +++ | +++ | |
| C2 | 36 | + | ++ | ++ |
| 37 | + | +++ | +++ | |
| 38 | + | +++ | +++ | |
| 39 | + | +++ | +++ | |
| 40 | + | +++ | +++ | |
Primary and secondary immune response antisera were scored as follows by IFA for the characteristic punctate fluorescence pattern: −, negative at 1:50; +, weak (IFA titer up to 1:50); ++, medium (IFA titer up to 1:150); +++, strong (IFA titer up to 1:500). No positive reactions were observed for dilutions above 1:500. Immunoblots of recombinant proteins probed with pooled secondary-response mouse antisera were positive.
After secondary immunization, antisera from each group of mice were pooled and used to probe blots of reduced or nonreduced (with or without 2-mercaptoethanol) total parasite proteins for specific recognition of the PfRAP1 band pattern. In mature schizonts, RAP1 exists as two major protein bands, the mature protein, p80, and its proteolytically cleaved product, p65 (7, 13).
In immunoblots, antisera against the P2, P3, P4, P5, and C2 proteins recognized specifically both RAP1 bands (p80 and p65) and no other protein in the total parasite extract (Fig. 3, lanes 1 to 4 and 8). Antiserum against the C1 protein recognized only the mature RAP1 (p80) but not its processed form (p65) (Fig. 3, lane 7). This specificity correlates well with the fact that the proteolytic cleavage that yields the p65 fragment occurs between aa 190 and 191 (Fig. 1). The C1 protein contains only RAP1 sequences upstream of this cleavage site; hence, antibodies to C1 specifically react with the p80 but not with the p65 fragment, with which the C1 antigen shares no sequence. The reduced (Fig. 3) and nonreduced (not shown) blots were identical except for a typical difference in relative mobility of the two RAP1 bands, which run higher under nonreducing than reducing conditions (28).
FIG. 3.

Immunoblot reactivity of mouse antisera raised to recombinant RAP1 proteins against parasite RAP1 in extract of total schizont proteins. Enriched schizonts were boiled in Laemmli buffer, and proteins were resolved on an SDS-PAGE gel and electroblotted onto a nitrocellulose membrane. Membrane strips were probed with pooled secondary immunization antisera to the indicated rRAP1 proteins and developed with peroxidase-conjugated rabbit anti-mouse immunoglobulin. The positions of parasite RAP1 p80 and its natural fragment p65 are indicated on the right. Lanes 1 to 4 and 8, specific reactivity of antisera to P2, P3, P4, P5, and C2, respectively, with both p80 and p65; lane 7, reactivity of anti-C1 serum to p80 only. Antisera to P6 and P7 reacted with no parasite protein (lanes 5 and 6). The marker lane contained the same prestained molecular weight markers as in Fig. 2.
The P6 and P7 proteins both induced antibodies which recognized these recombinant immunogens but failed to recognize RAP1 or any other protein of the parasite (Fig. 3, lanes 5 and 6).
Human antibody reactivity with rRAP1 proteins.
Once the antigenic similarity of the rRAP1 proteins to PfRAP1 was established, the proteins were used in an ELISA to detect specific antibodies in sera of malaria patients. Titrations of a positive standard serum included in each assay were highly reproducible and allowed interexperimental comparison of results. Sera of 31 non-malaria-exposed donors served to determine negative-to-positive cutoff values that were used to calculate the percentages of antibody-positive patients.
The reactivities of the proteins with serum IgG antibodies from Gambian malaria patients are illustrated in Fig. 4. The proportions of patients who had antibodies reactive with the individual proteins were variable. Of 44 patients tested, 39, 48, 57, 64, 66, and 82% had IgG reactive with the C1, C2, P2, P3, P4, and P5 proteins, respectively. In contrast, only 23 and 11% of patients had antibodies reactive with the P6 and P7 proteins, respectively. Thus, detectable antibodies appear to be directed to epitopes present on proteins that contain N-terminal sequences of mature RAP1.
FIG. 4.

Dot plot showing IgG antibodies detectable with recombinant RAP1 proteins in sera from Gambian malaria patients. Each circle represents the reactivity of 1 serum sample from each of 44 individuals. Horizontal bars indicate cutoffs (mean + 2 standard deviations of 31 European sera).
Epitope similarities among rRAP1 proteins.
Competition ELISAs were used for more-accurate epitope comparisons between selected pairs of rRAP1 proteins. To assess epitopes in the N-terminal region, we compared C1 and P5, the shortest and the longest rRAP1 antigens which share N-terminal aa 23 to 175 (an example is shown in Fig. 5a). Antibodies in a serum preincubated with either C1 or P5 as a competitor protein were completely blocked from binding to C1. In contrast, only partial inhibition of antibodies reactive with the P5 antigen was achieved with C1 as the competitor antigen. Thus, although this serum contains some antibodies to determinants within the N-terminal part that is common to the two proteins (aa 23 to 175), most antibodies are against additional epitopes present only on the P5 antigen. Identical results were obtained with four other tested human sera.
FIG. 5.

(a) Competition ELISA between C1 and P5 antigens showing cross-reactivity of human IgG antibodies with two different rRAP1 proteins. C1 competitor partially inhibits antibody binding to P5 antigen (□) and completely inhibits antibody binding to C1 antigen (▪) on wells. Preincubation of the serum with the P5 competitor completely blocks binding of antibodies to either C1 (○) or P5 (•). (b) Competition ELISA between P4 and P5 antigens showing the presence of minor epitopes in the C-terminal region of rRAP1 P5. Substantial partial inhibition of antibodies binding to P5 antigen on wells was observed with the P4 competitor (□); P4 competitor completely inhibits antibodies binding to P4 on wells (×). Preincubation of the serum with P5 as a competitor completely blocks binding of antibodies to either P4 (○) or P5 (•).
Competition ELISAs between the P4 and P5 antigens were performed to assess whether there were additional epitopes on P5, the longer of the two proteins (aa 23 to 769), compared to P4 (aa 23 to 608). The results suggest that P5 has only minor additional epitopes compared to P4 protein (Fig. 5b). Antibodies preincubated with P5 as the competitor were completely inhibited from binding to either P4 or P5. Preincubation of the antibodies with P4 as the competitor almost completely blocked binding to P5. Of nine sera used to test competition between this antigen pair, reactions of only two other sera revealed the presence of minor epitopes at the C-terminal region of P5.
Human antibodies do not block binding of inhibitory MAb 2.29.
We tested by competition ELISA whether human antibodies recognize an epitope that is a target for an inhibitory MAb. Ten sera containing antibodies to the P4 protein were tested for their ability to block binding of inhibitory MAb and vice versa. The human antibodies did not block binding of MAb 2.29 to P4 (Fig. 6a). In reciprocal experiments, increasing concentrations of MAb 2.29 (Fig. 6b) did not block the binding of human antibodies to P4. Identical results were obtained with the C2 protein (not shown). These results suggest that antibodies from malaria patients bind mostly to epitopes different from that recognized by the inhibitory MAb.
FIG. 6.

(a) Competition ELISA showing that anti-RAP1 human antibodies do not block binding of MAb 2.29. Increasing concentrations of serum M5 or 2D2, containing antibodies to rRAP1 P4, or a control normal serum were reacted with wells coated with P4 antigen. A dilution of RAP1 MAb 2.29 (1:100,000) was then reacted with the wells and developed with peroxidase rabbit anti-mouse IgG. (b) Competition ELISA showing that anti-RAP1 MAb 2.29 does not block binding of human antibodies in serum M5 or serum 2D2. Increasing concentrations of MAb 2.29 and a control MAb, 13.4 were reacted with P4-coated wells. Human sera diluted 1:500 were reacted with the wells and developed with mouse Ig-adsorbed peroxidase-conjugated goat anti-human IgG (Caltag, Burlingame, Calif.).
RAP1 IgG subclass.
The rRAP1 proteins C1, C2, P4, and P5 were used to examine IgG subclasses in sera of 22 individuals who tested positive for IgG antibodies with at least one of these antigens. Results obtained with the four antigens were similar. IgG1 was the predominant antibody subclass (Fig. 7). A few sera contained traces of IgG2, IgG3, or IgG4 at levels close to the negative cutoffs except for two individuals with IgG3 reactive to the C2 antigen.
FIG. 7.

Dot plot of human anti-RAP1 IgG antibody subclasses. Sera were tested for IgG antibody subclasses reactive with rRAP1 antigens C1 (a), C2 (b), P4 (c), and P5 (d). Reactivities of 22 individual endemic sera (circles) and of 9 European control sera (squares) and cutoff levels (mean + 2 standard deviations of the European sera) (horizontal bars) are indicated.
DISCUSSION
The aim of this study was to develop a panel of recombinant proteins antigenically similar to PfRAP1 for the evaluation of human antibody responses to RAP1 during malaria infection. Although RAP1 is a potential malaria vaccine candidate, it has received less attention than other malaria molecules in seroepidemiological studies of human immunity (15, 20, 21, 35). In addition, the studies of human anti-RAP1 responses reported so far used recombinant proteins, most of which were not characterized for antigenic similarities with PfRAP1. This is probably because the production of suitable rRAP1 antigens has been difficult.
Here, two sets of rRAP1 proteins were successfully purified in a soluble form required for antibody ELISAs. Recombinant proteins fused to GST were readily soluble in PBS in the presence of a mild detergent. His6 recombinant proteins, however, had to be extracted in 6 M guanidine hydrochloride and could be maintained in solution only in the presence of 8 M urea. Stowers et al. (34) reported similar difficulties in the purification of a His6 rRAP1 (the same as P4 reported here) in the absence of strong detergents. Our results indicate that the solubility of rRAP1 is improved to some extent by the GST fusion partner. However, the parasite RAP1 is lipophilic and requires the use of detergents for its extraction and purification (28). It is thus uncertain whether any recombinant expression system could ameliorate the inherent poor solubility of the RAP1 molecule.
Degradation was a feature of rRAP1 proteins reported in this study, a problem also encountered by Stowers et al. (34). Since protease inhibitors added during purification did not prevent this degradation, it probably occurs before purification of the recombinant proteins, possibly within E. coli cells following expression. Alternatively, autocatalysis could be an inherent property of the RAP1 protein, an idea originally suggested by Ridley et al. (28). Whatever the reason, the degradation would be a problem for production of a defined rRAP1 vaccine. Though this problem remains unresolved, it can be circumvented by isotachophoresis, which has been used to purify a defined rRAP1 protein from its multiple breakdown products (36).
Immunization of mice showed that all our rRAP1 proteins were immunogenic, and antibodies to six proteins (C1, C2, P2, P3, P4, and P5) recognized PfRAP1, indicating that at least these proteins had epitopes of the native protein. Hence, we were confident in using these proteins for serological assays.
We have used the rRAP1 proteins in an ELISA to show that Gambian malaria patients had IgG antibodies reactive with the proteins. Together with reports of others (15, 20, 21, 35), this indicates that native RAP1 is antigenic during naturally transmitted infections. Previous studies established that human antibodies are directed to epitopes on recombinant proteins containing N-terminal sequences, aa 1 to 292 (15) and aa 23 to 294 (20, 21). In addition, Howard et al. (15) mapped major epitopes to the N-terminal region from aa 1 to 122. Although our results are in general agreement with these reports, the results of competition between the C1 and P5 antigens (Fig. 5a) indicate the presence of stronger or more numerous human B-cell epitopes outside the N-terminal region from aa 23 to 175. Also, when the percentages of patients who have antibodies reactive with the C1 and C2 proteins (39 and 48%, respectively) are compared, the larger proportion had antibodies to C2 antigen (aa 169 to 366). Thus, although there are B-cell epitopes at a segment of the N-terminal region (aa 23 to 175), additional and stronger epitopes exist C terminal to this region.
We have investigated whether human antibodies can compete for an epitope (LTPLEELYP) targeted by an inhibitory murine MAb. Although we have obtained no evidence for antibodies to this epitope in malaria patients, the possibility that the sequence could be recognized by immune individuals has not been excluded.
Competition ELISA between the P4 and P5 antigens indicated that only a few individuals have antibodies to the C terminus and thus that this region of RAP1 may be poorly immunogenic. Previous studies also suggested that the C-terminal parts of RAP1 may be poorly immunogenic in humans. Howard et al. (15) reported that 8 of 17 sera tested reacted weakly with SDS-denatured rRAP1, representing the C-terminal part of the protein. A pool of human immune sera used to probe affinity-purified denatured PfRAP1 strongly recognized the whole protein (p80) but only weakly recognized its natural C-terminal fragment p65 (27). It has been speculated that the C-terminal region contains amphiphilic helices which may account for its apparent poor immunogenicity (28). Alternative explanations, not considered in the interpretation of the earlier results, are that the antigenic structure of the C-terminal part of RAP1 was destroyed by denaturation and/or not properly formed in recombinant proteins.
In PfRAP1, cysteine residues form intramolecular disulfide bonds (28), though neither the precise pattern of disulfide bond formation nor the effect of the bonds on the epitope structure of RAP1 is known. We find that antibodies elicited in mice by P6 and P7 do not recognize PfRAP1 at all, and these proteins are also poorly recognized by antibodies from malaria patients. The P6 and P7 proteins span the cysteine-rich region of PfRAP1, but they lack one and six cysteine residues, respectively, and thus they may not be properly folded. Poor recognition of the P6 and P7 antigens by only a few human sera suggests that if natural antibodies to the cysteine-rich region of RAP1 exist, they may be directed to conformation-dependent epitopes lacking in these proteins. Indeed, the incomplete cysteine-rich region in P6 and P7 may be folded so inappropriately that the proteins bear no antigenic resemblance to the parasite-produced RAP1.
Protein P5, however, contains 98% of the sequence of the mature RAP1 and all its cysteine residues. Thus, in principle, P5 could be appropriately folded to express putative conformational epitopes that may react with antibodies. Though some conformation-dependent MAbs inhibit parasite growth in vitro (31), immunization of Saimiri monkeys with denatured RAP1-RAP2 complex afforded protection against P. falciparum infection (27). This suggests that conformation-dependent epitopes may not be critical for anti-RAP1 immunity.
We have evaluated for the first time the distribution of anti-RAP1 IgG subclasses and have found IgG1 to be the predominant subclass. The subclass of IgG antibodies is likely to be an important factor modulating cellular immunity to malaria. Cytophilic IgG1 and IgG3 antibodies that bind to receptors on monocytes, macrophages, and neutrophils enhance antiparasite mechanisms, including phagocytosis (11, 23), antibody-dependent cellular inhibition (3), and respiratory burst in vitro (24). In vivo, the presence of IgG1 and IgG3 antibodies has been correlated with individuals’ ability to maintain low parasite densities (1, 2, 12). Whether antibodies to RAP1 are protective is not known yet, but the predominantly IgG1 response could be of significance since the intracellular location of RAP1 may require antibody-dependent cellular inhibition. Alternatively, a possible mechanism of anti-RAP1 immunity could be the removal from circulation of RAP1-containing endotoxin-like antigen complex, which may have a role in malaria pathology (22).
In summary, we have developed a panel of recombinant antigens which retain epitopes similar to parasite-derived RAP1. The proteins were used to show that the most N-terminal segment (residues 23 to 175) contains only minor epitopes, whereas major epitopes are outside this region. Subclass analyses of human anti-RAP1 antibodies show predominantly IgG1 responses. Longitudinal studies are in progress to provide a better understanding of the kinetics of human immune responses to RAP1.
ACKNOWLEDGMENTS
We are grateful to Kay Samuel for immunization of mice and Sabine Demmak for technical assistance. We thank Brian Greenwood for access to Gambian sera and Eleanor Riley for supplying a pool of malaria-immune serum.
This investigation received financial support from UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases (project 940030) and from the Wellcome Trust (013163/Z/96/B).
REFERENCES
- 1.Aribot G, Rogier C, Sarthou J L, Trape J F, Balde A T, Druilhe P, Roussilhon C. Pattern of immunoglobulin isotype response to Plasmodium falciparum blood-stage antigens in individuals living in a holoendemic area of Senegal (Dielmo, West Africa) Am J Trop Med Hyg. 1996;54:449–457. doi: 10.4269/ajtmh.1996.54.449. [DOI] [PubMed] [Google Scholar]
- 2.Bouharoun-Tayoun H, Druilhe P. Plasmodium falciparum malaria: evidence for an isotype imbalance which may be responsible for delayed acquisition of protective immunity. Infect Immun. 1992;60:1473–1481. doi: 10.1128/iai.60.4.1473-1481.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bouharoun-Tayoun H, Oeuvrey C, Lunel F, Druilhe P. Mechanism underlying the monocyte-mediated antibody-dependent killing of Plasmodium falciparum asexual blood stages. J Exp Med. 1995;182:409–418. doi: 10.1084/jem.182.2.409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bushell G R, Ingram L T, Fardoulys C A, Cooper J A. An antigenic complex in the rhoptries of Plasmodium falciparum. Mol Biochem Parasitol. 1988;28:105–112. doi: 10.1016/0166-6851(88)90057-6. [DOI] [PubMed] [Google Scholar]
- 5.Campbell G H, Miller L H, Hudson D, Franco E L, Andrysiak P M. Monoclonal antibody characterisation of Plasmodium falciparum antigens. Am J Trop Med Hyg. 1984;33:1051–1054. doi: 10.4269/ajtmh.1984.33.1051. [DOI] [PubMed] [Google Scholar]
- 6.Cavanagh D R, McBride J S. Antigenicity of recombinant protein derived from Plasmodium falciparum merozoite surface protein 1. Mol Biochem Parasitol. 1997;85:197–211. doi: 10.1016/s0166-6851(96)02826-5. [DOI] [PubMed] [Google Scholar]
- 7.Clark J T, Anand R, Akoglu T, McBride J S. Identification and characterisation of proteins associated with the rhoptry organelles of Plasmodium falciparum merozoites. Parasitol Res. 1987;73:425–434. doi: 10.1007/BF00538200. [DOI] [PubMed] [Google Scholar]
- 8.Conway D J, Greenwood B M, McBride J S. The epidemiology of multiple-clone Plasmodium falciparum infections in Gambian patients. Parasitology. 1991;103:1–6. doi: 10.1017/s0031182000059217. [DOI] [PubMed] [Google Scholar]
- 9.Conway D J, Greenwood B M, McBride J S. Longitudinal study of Plasmodium falciparum polymorphic antigens in a malaria-endemic population. Infect Immun. 1992;60:1122–1127. doi: 10.1128/iai.60.3.1122-1127.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Crewther P E, Culvenor J G, Silva A, Cooper J A, Anders R F. Plasmodium falciparum: two antigens of similar size are located in different compartments of the rhoptry. Exp Parasitol. 1990;70:193–206. doi: 10.1016/0014-4894(90)90100-q. [DOI] [PubMed] [Google Scholar]
- 11.Druilhe P, Khusmith S. Epidemiological correlation between levels of antibodies promoting merozoite phagocytosis of Plasmodium falciparum and malaria immune status. Infect Immun. 1987;55:888–891. doi: 10.1128/iai.55.4.888-891.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ferreira M U, Kimura E A S, Souza J M, Katzin A M. The isotype composition and avidity of naturally acquired anti-Plasmodium falciparum antibodies: differential patterns in clinically immune Africans and Amazonian patients. Am J Trop Med Hyg. 1996;55:315–323. doi: 10.4269/ajtmh.1996.55.315. [DOI] [PubMed] [Google Scholar]
- 13.Harnyuttanakorn P, McBride J S, Donachie S, Heidrich H G, Ridley R G. Inhibitory monoclonal antibodies recognise epitopes adjacent to a proteolytic cleavage site on the RAP 1 protein of Plasmodium falciparum. Mol Biochem Parasitol. 1992;55:177–186. doi: 10.1016/0166-6851(92)90138-a. [DOI] [PubMed] [Google Scholar]
- 14.Howard R F. The sequence of the p82 rhoptry protein is highly conserved between two Plasmodium falciparum isolates. Mol Biochem Parasitol. 1992;51:327–330. doi: 10.1016/0166-6851(92)90083-v. [DOI] [PubMed] [Google Scholar]
- 15.Howard R F, Jensen J B, Franklin H L. Reactivity profile of human anti-82-kilodalton rhoptry protein antibodies generated during natural infection with Plasmodium falciparum. Infect Immun. 1993;61:2960–2965. doi: 10.1128/iai.61.7.2960-2965.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Howard R F, Peterson C. Limited RAP1 sequence diversity in field isolates of Plasmodium falciparum. Mol Biochem Parasitol. 1996;77:95–98. doi: 10.1016/0166-6851(96)02576-5. [DOI] [PubMed] [Google Scholar]
- 17.Howard R F, Reese R T. Synthesis of merozoite proteins and glycoproteins during the schizogony of Plasmodium falciparum. Mol Biochem Parasitol. 1984;10:319–334. doi: 10.1016/0166-6851(84)90030-6. [DOI] [PubMed] [Google Scholar]
- 18.Howard R F, Stanley H A, Campbell G H, Reese R T. Proteins responsible for a punctate fluorescence pattern in Plasmodium falciparum merozoites. Am J Trop Med Hyg. 1984;33:1055–1059. doi: 10.4269/ajtmh.1984.33.1055. [DOI] [PubMed] [Google Scholar]
- 19.Howard R J, Pasloske B L. Target antigens for asexual malaria vaccine development. Parasitol Today. 1993;9:369–372. doi: 10.1016/0169-4758(93)90085-t. [DOI] [PubMed] [Google Scholar]
- 20.Jakobsen P H, Hviid L, Theander T G, Afare E A, Ridley R G, Heegaard P M H, Stüber D, Dalsgaard K, Nkrumah F K. Specific T-cell recognition of the merozoite proteins rhoptry-associated protein 1 and erythrocyte-binding antigen 1 of Plasmodium falciparum. Infect Immun. 1993;61:268–273. doi: 10.1128/iai.61.1.268-273.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jakobsen P H, Lemnge M M, Abu-Zeid Y A, Msangeni H A, Salum F M, Mhina J I K, Akida J A, Ruta A S, Ronn A M, Heegaard P M H, Ridley R G, Bygbjerg I C. Immunoglobulin G reactivities to rhoptry-associated protein-1 associated with decreased levels of Plasmodium falciparum parasitaemia in Tanzanian children. Am J Trop Med Hyg. 1996;55:642–646. doi: 10.4269/ajtmh.1996.55.642. [DOI] [PubMed] [Google Scholar]
- 22.Jakobsen P H, Moon R, Ridley R G, Bate C A W, Taverne J, Hansen M B, Takacs B, Playfair J H L, McBride J S. Tumour necrosis factor and interleukin-6 production induced by components associated with merozoite proteins of Plasmodium falciparum. Parasite Immunol. 1993;15:229–237. doi: 10.1111/j.1365-3024.1993.tb00605.x. [DOI] [PubMed] [Google Scholar]
- 23.Khusmith S, Druilhe P. Specific arming of monocytes by cytophilic IgG promotes Plasmodium falciparum merozoite ingestion. Trans R Soc Trop Med Hyg. 1982;76:423–424. doi: 10.1016/0035-9203(82)90208-5. [DOI] [PubMed] [Google Scholar]
- 24.Kumaratilake L M, Ferrante A, Jaeger T, Rzepczyk C M. Effects of cytokines, complement, and antibody on the neutrophil respiratory burst and phagocytic response to Plasmodium falciparum merozoites. Infect Immun. 1992;60:3731–3738. doi: 10.1128/iai.60.9.3731-3738.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Prapunwattana P, Sirawaraporn W, Yuthavong Y, Santi D V. Chemical synthesis of the Plasmodium falciparum dihydrofolate reductase-thymidylate synthase gene. Mol Biochem Parasitol. 1996;83:93–106. doi: 10.1016/s0166-6851(96)02756-9. [DOI] [PubMed] [Google Scholar]
- 26.Ridley R G, Lahm H W, Takacs B, Scaife J G. Genetic and structural relationships between components of a protective rhoptry antigen complex from Plasmodium falciparum. Mol Biochem Parasitol. 1991;47:245–246. doi: 10.1016/0166-6851(91)90184-8. [DOI] [PubMed] [Google Scholar]
- 27.Ridley R G, Takacs B, Etlinger H, Scaife J G. A rhoptry antigen of Plasmodium falciparum is protective in Saimiri monkeys. Parasitology. 1990;101:187–192. doi: 10.1017/s0031182000063228. [DOI] [PubMed] [Google Scholar]
- 28.Ridley R G, Takacs B, Lahm H W, Delves C J, Goman M, Certa U, Matile H, Woollett G R, Scaife J G. Characterisation and sequence of a protective rhoptry antigen from Plasmodium falciparum. Mol Biochem Parasitol. 1990;41:125–134. doi: 10.1016/0166-6851(90)90103-s. [DOI] [PubMed] [Google Scholar]
- 29.Sanger F, Nicklen S, Coulson A R. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA. 1977;74:5463–5467. doi: 10.1073/pnas.74.12.5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Saul A, Cooper J, Hauquitz D, Irving D, Cheng Q, Stowers A, Limpaiboon T. The 42-kilodalton rhoptry-associated protein of Plasmodium falciparum. Mol Biochem Parasitol. 1992;50:139–150. doi: 10.1016/0166-6851(92)90251-e. [DOI] [PubMed] [Google Scholar]
- 31.Schofield L, Bushell G R, Cooper J A, Saul A J, Upcroft J A, Kidson C. A rhoptry antigen of Plasmodium falciparum contains conserved and variable epitopes recognised by inhibitory monoclonal antibodies. Mol Biochem Parasitol. 1986;18:183–195. doi: 10.1016/0166-6851(86)90037-x. [DOI] [PubMed] [Google Scholar]
- 32.Smith D B, Corcoran L M. Expression and purification of glutathione-S-transferase fusion protein. In: Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: Greene Publishing Associates and Wiley Interscience; 1995. pp. 16.7.1–16.7.7. [DOI] [PubMed] [Google Scholar]
- 33.Smith D B, Johnson K S. Single-step purification of polypeptides expressed in Escherichia coli as fusions with glutathione S-transferase. Gene. 1988;67:31–40. doi: 10.1016/0378-1119(88)90005-4. [DOI] [PubMed] [Google Scholar]
- 34.Stowers A, Prescott N, Cooper J, Takacs B, Stüber D, Kennedy P, Saul A. Immunogenicity of recombinant Plasmodium falciparum rhoptry associated proteins 1 and 2. Parasite Immunol. 1995;17:631–642. doi: 10.1111/j.1365-3024.1995.tb01009.x. [DOI] [PubMed] [Google Scholar]
- 35.Stowers A, Taylor D, Prescott N, Cheng Q, Cooper J, Saul A. Assessment of the humoral immune response against Plasmodium falciparum rhoptry-associated proteins 1 and 2. Infect Immun. 1997;65:2329–2338. doi: 10.1128/iai.65.6.2329-2338.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stowers A W, Spring K J, Saul A. Preparative scale purification of recombinant proteins to clinical grade by isotachophoresis. Bio/Technology. 1995;13:1498–1503. doi: 10.1038/nbt1295-1498. [DOI] [PubMed] [Google Scholar]
- 37.Stüber D, Matile H, Garotta G. System for high level production in Escherichia coli and rapid purification of recombinant proteins: application to epitope mapping, preparation of antibodies and structure-function analysis. In: Lefkovits J, Pernis B, editors. Immunological methods. IV. Orlando, Fla: Academic Press; 1990. pp. 121–152. [Google Scholar]
- 38.Thaithong S, Beale G H. Resistance of ten Thai isolates of Plasmodium falciparum to chloroquine and pyrimethamine by in vitro tests. Trans R Soc Trop Med Hyg. 1981;75:271–273. doi: 10.1016/0035-9203(81)90333-3. [DOI] [PubMed] [Google Scholar]
- 39.Trager W, Jensen J B. Human malaria parasites in continuous culture. Science. 1976;193:673–675. doi: 10.1126/science.781840. [DOI] [PubMed] [Google Scholar]