

Abstract
Background:
It is well established that unrefined sugarcane products have antioxidant activity due to phytochemicals, polyphenols, and total antioxidant capacity, which may decrease inflammation and oxidative stress. Therefore, we conducted a systematic review to evaluate the association of unrefined sugar consumption with inflammatory biomarkers.
Methods:
Google Scholar, ScienceDirect, Scopus, Cochrane Library, and ProQuest databases were searched up to December 2021 for studies that report the effect of unrefined sugar on inflammation according to inflammatory cytokines, chemokine, and adhesion molecules as outcome measures. Results: Thirty-six studies were evaluated. Across all research, five studies (two in vitro and three animal studies) reported the effect of unrefined sugar on levels of cytokines, including IL-6, TNF-α, IL-10, IL-1β, and IFN-γ. Additionally, the quality of the studies was assessed for risk of bias.
Conclusions:
it is possible to affirm that unrefined sugarcane products, including jaggery, may have a protective effect on inflammation via regulating some of the inflammatory pathways and a favorable impact on cytokines secretion according to the results of in vitro and animal model studies. However, since the findings are still insufficient, more scientific research, especially well-designed human trials, is highly recommended to conclude the outcomes confidently. Human data may encourage industries and the public to replace purified sugar with unrefined sugarcane in sugar-based food and for further health-care policy decisions.
Keywords: Brown sugar, dietary sugar, inflammation, sugarcane, systematic review, unrefined sugar
Introduction
Sweet taste, in particular from added sugars, is the most desirable flavor. Since the beginning of the 20th century, sucrose has become one of the main sweetener choices among the food industry and consumers. Sucrose, the table sugar, is produced at about 165 million tons a year, most from sugar cane (80%) and sugar beets.[1] According to Dietary Guidelines for Americans in 2020, excessive sugar intake may increase calorie intake without raising nutritional diet value. Sugar-sweetened beverages (such as soda, sports drinks, energy, and fruit drinks) and sweetened coffees and teas contribute over 40% of adult’s daily intake of added sugars.[2] Usually, foods rich in added sugar are low in fiber, minerals, vitamins, and antioxidants. Moreover, over-sugar consumption results in hypertension, cancer, adverse lipid profiles, increased inflammation and oxidative stress, cardiovascular disease, insulin resistance, obesity, glucose intolerance, and diabetes.[3,4,5,6] Thus, both the World Health Organization and the American Heart Association suggest that added sugar should not exceed 10% of total calorie intake and not more than six tablespoons full for women and nine tablespoons full for men, respectively.[3,5]
Despite the disadvantages of refined sugar consumption, white sugar is the most widely used sugar worldwide. However, these days, unrefined sugarcane alternatives, including noncentrifugal sugar (NCS), are available in the market.[7] NCS, known as whole cane sugar, has been used as a sweetener in most cane sugar-growing regions. Traditionally, the NCS uses various names depending on the country of its origin, such as brown sugar (Europe and North America), Gula Melaka (Malaysia), Jaggery (India), Kokuto (Japan), Panela (Colombia), Rapadura (Brazil), and Muscovado (Philippines) which are all produced using concentrated cane sugar juice.[8,9,10]
Many studies reported that unrefined sugarcane products have antioxidant activity due to phytochemicals and polyphenol components.[11,12,13,14,15,16] The phytochemicals and phenolics may inhibit the activity of free radicals through several mechanisms, including the destruction of free radicals, binding to metals that stimulate the production of free radicals and inhibiting the formation of free radicals as well as acting as scavengers of free radicals.[17] The antioxidants may decrease the risk of oxidative stress-related diseases such as cardiovascular, diabetes, obesity, cancer, and DNA damage.[18,19,20,21,22] In addition, phytochemicals have shown positive effects on carbohydrate metabolism during glucose transporter activity in the gut, showed protective effects on pancreatic cells from toxicity, resulted in favorable effects on insulin release, inhibition of gluconeogenesis as well as improved glucose uptake into tissues.[9,23] However, to the best of our knowledge, no comprehensive review evaluates the effectiveness of unrefined sugarcane products on inflammation. Therefore, the purpose of this systematic review is to evaluate the evidence from published studies regarding the impact of unrefined sugar on biomarkers of inflammation such as C-reactive protein (CRP), high-sensitivity C-reactive protein (hs-CRP), pro-inflammatory cytokines (e.g., interleukin-6 [IL-6], tumor necrosis factor-α [TNF-α]), chemokine, and adhesion molecules.
Materials and Methods
We conducted a systematic literature search for relevant published articles through PubMed, Google Scholar, ScienceDirect, Scopus, Cochrane Library, and ProQuest databases until December 2021. The PRISMA guideline was used in writing the present systematic review[24] and guidelines in the Cochrane Handbook for Systematic Reviews of Interventions.[25] In order to ensure that all potentially relevant publications in the English language entered the screening process, broad search terms were used in the database searches, including “unrefined sugar” OR “raw sugar” OR “NCS” OR “brown sugar” OR “jaggery” OR “Kokuto” OR “panela” AND “inflammation” OR “inflammatory cytokines” OR “pro-inflammatory cytokines” OR “CRP” OR “hs-CRP” OR “NFκB” OR “chemokine” OR “adhesion molecules” [Supplementary Table 1]. Whenever possible, Medical Subject Headings terms were used. In addition to the electronic search, the reference lists of all included articles were reviewed to find more related studies. Since the published data in this research area are limited, we included all studies (in vitro, in vivo, animal, and human intervention) that reported unrefined sugar consumption’s effect on inflammation by evaluating at least one of the inflammatory biomarkers as the outcome indicators. Since we are interested in investigating the effects of unrefined sugar on inflammation, we excluded studies that reported the effects of other sweeteners, dietary patterns, and glycemic index. Furthermore, studies that simultaneously assessed the effects of other nutrients (such as herbal extract) on sugar intake were excluded [Figure 1]. This systematic review was registered with PROSPERO on July 26, 2022 (CRD42022344140).
Supplementary Table 1.
The systematic search keywords
| Keywords | n |
|---|---|
| “unrefined sugar” | |
| “brown sugar” | |
| “non-centrifugal sugar” | |
| “kokuto” | |
| “jaggery” | |
| “raw sugar” | |
| “whole cane sugar” | |
| “panela” | |
| “muscovado” | |
| “rapadura” | |
| “gula melaka” | |
| “whole sugarcane” | |
| (1 OR 2 OR 3 OR 4 OR 5 OR 6 OR 7 OR 8 OR 9 OR 10 OR 11 OR 12) | |
| “inflammation” | |
| “inflammatory cytokines” | |
| “chemokines” | |
| “adhesion molecules” | |
| “proinflammatory cytokines” | |
| “CRP” | |
| “hs-CRP” | |
| “IL-6” | |
| “IL-1” | |
| “IL-2” | |
| “IL-8” | |
| “MCP-1” | |
| “ICAM-1” | |
| “VCAM-1” | |
| “E-selectin” | |
| “TNF-α” | |
| “adiponectin” | |
| “IL-18” | |
| “IL-1β” | |
| “CCL2” | |
| “IFN-γ” | |
| “IL-4” | |
| (14 OR 15 OR 16 OR 17 OR 18 OR 19 OR 20 OR 21 OR 22 OR 23 OR 24 OR 25 OR 26 OR 27 OR 28 OR 29 OR 30 OR 31 OR 32 OR 33 OR 34 OR 35) | |
| (13 AND 36) in Scopus | 8 |
| (13 AND 36) in PubMed | 6 |
| (13 AND 36) in Google Scholar | 13 |
| (13 AND 36) in ScienceDirect | 2 |
| (13 AND 36) in Cochrane Library | 4 |
| (13 AND 36) in ProQuest | 3 |
| Total | 36 |
Figure 1.
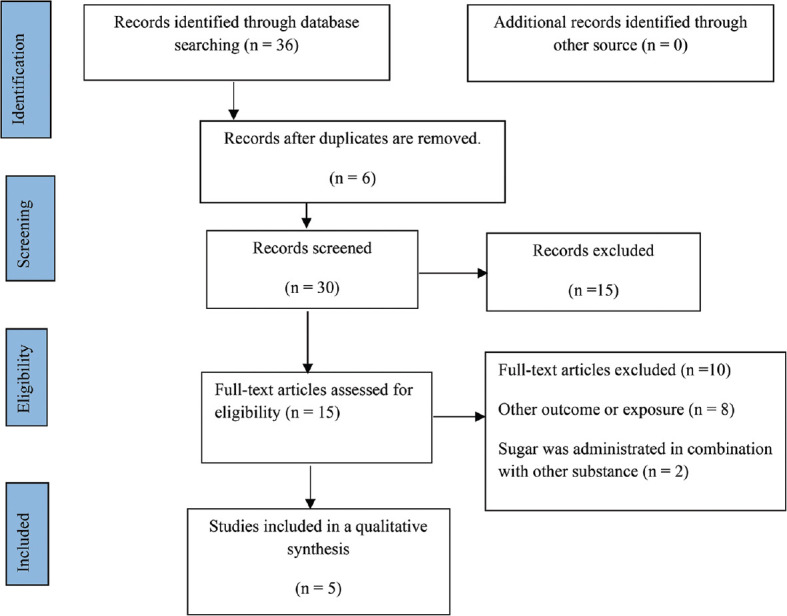
Flow chart of the study selection process
Data extraction and quality assessment of research papers
The literature search was conducted by two independent investigators (SE and AA), and they selected the related studies after reading the titles, abstract, methods, and full text. Those studies compared the outcomes before and after the intervention had been selected based on inclusion and exclusion criteria. Authors, publication year, the study’s country, duration, study design and intervention, and the primary outcomes were recorded for each study [Table 1]. Duplicated articles were excluded, and the final list was prepared for the systematic review. The quality of all studies was assessed using a checklist based on the Toxicological Data Reliability Assessment Tool (ToxRTool)[26] and Systematic Review Centre for Laboratory animal Experimentation (SYRCLE’s) Risk of Bias (Rob) tool (S2).[27] The ToxRTool has been developed to evaluate the quality of in vitro studies and consists of 18 criteria in five groups of criteria, including test substance identification, test system characterization, study-design description, study results documentation, and the plausibility of study design and results. All criteria must be answered (either by 0 or 1) while evaluating the paper. Finally, the total points assigned to each study lead to a proposal of a reliability category from 1 to 3, calculated by the tool. The SYRCLE’s Rob tool is based on the Cochrane Rob tool. It considers the following entries to assess the methodological quality of animal intervention studies: selection bias, performance bias, detection bias, attrition bias, reporting bias, and other biases.
Table 1.
Studies investigate the effect of unrefined sugar on biomarkers of inflammation- extracted data on study designs, interventions, the form of sugar, and the key conclusion
| First author, year, country | Study design | Intervention | Sugar form | Results | Key conclusions | Quality assessment score |
|---|---|---|---|---|---|---|
| Singh N, 2009,[28] India | Randomized (5 groups), 180 days, each group of 10 Swiss male albino mice with an average weight of 30±3 g | Intervention: Arsenic (0.05 ppm) Arsenic (5 ppm) Arsenic (0.05 ppm) + Jaggery (250 mg/kg) Arsenic (5 ppm) + Jaggery (250 mg/kg) Control: Neither arsenic nor jaggery | Jaggery in distilled water | 1. In the jaggery-supplemented arsenic-exposed groups, the serum levels of interleukin-1β, interleukin-6, and TNF-α were significantly decreased compared to arsenic alone treated groups 2. The levels of interleukin-1β, interleukin-6, and TNF-α were significantly higher in arsenic-treated animals than in control animals |
Jaggery effectively antagonizes many of the adverse effects of arsenic due to the presence of vitamins, minerals, carotene, and polyphenols, which may help maintain the redox equilibrium within the body to minimize the molecular and cellular oxidative damage caused by arsenic | 6/10 |
| Sánchez- Tapia M, 2019,[29] Mexico | Randomized (18 groups), 4 months, each group 6 male Wistar rats | Intervention: C+10% S C+10% F C+10% G C+2.5% SG C+10% BS C+10% H C+10% SV C+1.5% SU HF+10% S HF+10% F HF+10% G HF+2.5% SG HF+10% BS HF+10% H HF+10% SV HF+1.5% SU Control: C+W HF+W |
Brown sugar in drinking water | 1. Rats fed with sucrose, sucralose, glucose, and fructose showed the highest expression of RNA and protein abundance of TLR4 and MyD88. While BS and honey had the lowest expression of that 2. The phosphorylation of JNK was increased by SV, sucralose, sucrose, and fructose in the presence or absence of fat in the diet 3. Only groups fed with a high-fat diet stimulated the expression of NF-κB by sucralose and, to a lesser extent, by BS. There was no stimulation of NF-κB by SG and H 4. The expression of the TNF-α gene increased by sucralose, SV, Sucrose, glucose, and fructose and to a lesser extent by SG, BS, and H |
Since the intake of different sweeteners has various effects on the inflammatory state, the consumer should be aware of the type of sweetener and the fat content in the diet to prevent further metabolic complications | 6/10 |
| Elayaraja S, 2020,[30] China | Randomized (4 groups), 75 days, each group 60 healthy mono-sex tilapia | Intervention: T1: biofloc water (C: N12) T2: biofloc water (C: N15) T3: biofloc water (C: N20) Control: Dechlorinated, sand-filtered + UV satirized freshwater without JP | Jaggery powder as a carbohydrate source in biofloc water | 1. The mRNA expression of LYS, TNF-α, and IL-1β were significantly increased in the biofloc-treated fish compared to the control 2. The target genes were upregulated in proportion to the different C: N ratios, as the highest observed in C: N20 group, followed by C: N15 and C: N12 3. The increment in the expression of IL-1β and TNF-α genes reflects the effectiveness of JB-BFT in modulating the immune response of tilapia and increase its resistance to bacterial infection |
JB-BFT upregulates the immune gene expression profile, which robustly influences Nile tilapia’s innate immunity through the favorable innovation of various immune cells and enzymes. Thus, jaggery has been offered as a potential new carbon source with a unique property that satisfies all considerations of biofloc technology with high productivity and zero water exchange (eco-friendly) | 7/10 |
| Rahiman F, 2010,[31] South Africa | In vitro design using human whole blood cultures | Intervention: Stimulated (LPS/PHA) and unstimulated human blood cultures with various concentrations of sugar cane molasses Control: Stimulated (LPS/PHA) and unstimulated human blood cultures without molasses | Molasses in distilled water | 1. Molasses did not affect interleukin-6, interleukin-10, and interferon-gamma secretion in stimulated whole blood cultures 2. Molasses increased IL-6 secretion by unstimulating whole blood cultures for all concentrations compared to controls 3. Molasses samples increased the IL-10 secretion by unstimulated whole blood cultures only at 200, 400, and 800 µg/ml concentrations compared to the controls 4. Molasses samples did not affect IF-γ synthesis in both stimulated and unstimulated samples |
Molasses affects the cytokines regulating the hormonal and immune system and have inflammatory and anti-inflammatory potential. Thus, sugar cane molasses may have beneficial in promoting improved hormonal responses, but further research is suggested to conclude the results | 13/18 |
| Rahiman F, 2013,[32] South Africa | In vitro design using human whole blood cultures | Intervention: Stimulated (LPS/PHA) and unstimulated human blood cultures in the presence of various sweeteners (Artificial/natural) Control: Stimulated and unstimulated human blood cultures in the absence of sweeteners | Sweeteners in distilled water | 1. Sugar cane molasses enhanced IL-6 levels in the presence and absence of LPS, while all artificial sweeteners suppressed IL-6 levels of stimulated cultures 2. Brown sugar showed no effect on IL-6 levels compared to the distilled water control 3. None of the artificial and natural sweeteners had significant effect on levels of IL-10 in the presence and absence of PHA 4. Two artificial sweeteners significantly decreased the IL-10 levels under stimulated conditions 4. None of the artificial and natural sweeteners had significant effect on IFN-γ levels in the presence and absence of PHA |
The current article shows that artificial and natural sweeteners are not cytotoxic; however, they impact specific cellular pathways. The inflammatory potential of the sugar cane molasses may be favorable in defense against infective pathogens | 15/18 |
C=Control diet, HF=High-fat diet, W=Water, S=Sucrose, F=Fructose, G=Glucose, SG=Steviol glycosides, BS=Brown sugar, H=Honey, SV=SG + sucrose, SU=Sucralose, JP=Jaggery powder, LP=Lipopolysaccharide, LPH=Phytohemagglutinin
Results
Search results
A flow diagram visualized a detailed step in systematic search and study selection [Figure 1]. A total of 36 studies were identified during the initial search, of which 6 were removed due to duplication. Among the remaining articles (n = 30), 15 studies were omitted, considering the title and abstract. Finally, full texts of 15 articles were reviewed, and 5 studies were included in the review after omitting unrelated papers by the full-text assessment due to unrelated outcomes or exposure (n = 8), and sugar was administrated in combination with other substances (n = 2).
The main findings of the included trials in this systematic review are summarized in Table 1. The five trials addressed unrefined sugarcane products such as jaggery, brown sugar, and molasses intake as a nutritional intervention from 18 h to 180 days of exposure. Two studies were in vitro,[31,32] and the remaining (n = 3) were animal interventions.[28,29,30] Animal studies (n = 3) included male mice,[28] male rats,[29] and mono-sex tilapia fish,[30] while the in vitro studies used human whole blood cultures that were treated with raw sugar.[31,32] These studies tested different exposures (ranging from 0 μg/ml to 800 μg/ml) to unrefined sugar products in both intervention and control groups to determine the effectiveness of treatment on inflammatory biomarkers. Two studies in the animal model used diluted jaggery in water. The arsenic-exposed group supplemented with jaggery was compared to the arsenic-exposed group in the absence of jaggery treatment as a control group,[28] and the jaggery-based water system was compared to the water system with no carbon source.[30] One study compared natural (including brown sugar) and artificial sweeteners along with a high-fat diet (HF) to a control diet in the 18 randomized Wistar rats groups.[29] A study determined the in vitro effects of artificial and natural sweeteners (brown sugar and molasses) on the immune system using whole blood culture assays.[32] A previous in vitro study by the same researchers investigated the impact of the dilution range of molasses samples on endotoxin-stimulated or unstimulated diluted blood.[31]
IL-6,[28,31,32] TNF-α,[28,29,30] interleukin-1β (IL-1β),[28,30] interleukin-10 (IL-10),[31,32] and interferon-gamma (IFN-γ)[31,32] were assessed to investigate the outcomes in the intervention and control groups. Two studies evaluated the changes in the gene expression of common immune-related genes (lysozyme C, TNF-α, and IL-1β)[30] and the related inflammatory pathways [protein abundance of toll-like receptor 4 (TLR4), myeloid differentiation factor 88 (MYD88), nuclear factor kappa-light-chain enhancer of activated B cells (NF-κB), and c-Jun N-terminal kinase (JNK)].[29] Besides, two animal trials showed the protective effect of jaggery according to the study outcomes, such as IL-1β, IL-6, and TNF-α.[28,30]
Elayaraja et al. found that the jaggery-based biofloc system (JB-BFT), an economical nutrient recycling rearing system, modulated the immune response of Nile tilapia (Oreochromis niloticus) by upregulating the immune genes expression profile. In this randomized design study, the treatment groups (T1, T2, and T3) were supplied with biofloc water at C: N (carbon: nitrogen) ratios of C: N12, C: N15, and C: N20, respectively, using jaggery powder as a source of carbon, and the control group was raised in a water system without carbon substrates. The expression of different immune-related genes such as LYS, TNF-α, and IL-1β was assessed in the head kidney of the experimental fish at 75 days of experimental period. It was noticed that the JB-BFT system significantly increased the mRNA expression of the target genes in the biofloc-treated fish compared to the control, with the highest fold in C: N20.[30] The serum levels of IL-1β, IL-6, and TNF-α were significantly increased in arsenic-exposed groups [as sodium-m-arsenite at 0.05 ppm (low dose) and 5 ppm (high dose)]. In contrast, supplementation with jaggery (250 mg/mice) and arsenic (combined feeding groups) reduced their levels significantly after administrating the treatment to Swiss male albino mice for 180 days.[28]
In the study by Sánchez-Tapia and colleagues, male Wistar rats were randomized into 18 groups fed different sweeteners with (9 groups) and without high-fat diet (9 groups) at the following concentration (sucrose, fructose, glucose, brown sugar, honey, steviol glycosides plus sucrose; at 10%, steviol glycosides at 2.5%, and sucralose at 1.5%) for 4 months. The study results reported that the combination of a high-fat diet with sweeteners increased inflammation based on NF-κB gene expression, except for honey and steviol glycosides in white adipose tissue of rats by real-time PCR. Interestingly, none of the sweeteners had different induction of NF-κB protein abundance in the absence of fat in the diet. Brown sugar and honey had the lowest expression of RNA and protein abundance of TLR4. At the same time, rats fed with sucrose, sucralose, and glucose showed the highest expression of this receptor independently of the presence of high-fat diet. The expression of MYD88 followed a similar pattern to TLR4. Also, TNF-α was stimulated to a lesser extent by brown sugar, steviol glycosides, and honey compared to the other types of sweeteners, and the additional high-fat diet significantly enhanced the TNF-α gene expression.[29]
In Rahiman et al. (2013) study, human whole blood was treated with various sweeteners [including natural (white sugar, brown sugar, and sugarcane molasses) and artificial sweeteners (containing sucralose) at a concentration of 10 mg/ml] and stimulated in vitro with lipopolysaccharide (LPS) and phytohemagglutinin (PHA). The results revealed that the IL-6 levels significantly increased in the sugarcane molasses group under both stimulated and unstimulated conditions, while all artificial sweeteners suppressed the IL-6 secretion of stimulated cultures with LPS. According to the levels of the anti-inflammatory cytokines IL-10, there was no significant difference between natural and artificial sweeteners compared to the control group in the absence of PHA. However, two types of sucralose-containing sweeteners significantly decreased the IL-10 levels of stimulated cultures. Interestingly, brown sugar had no significant effect on inflammatory responses according to IL-6, IL-10, and IFN-γ parameters. Moreover, none of the sweeteners had an effect on the IFN-γ levels compared to the control group regardless of the stimulus PHA.[32] The earlier data by the same researchers determined the ability of molasses to enhance inflammatory activity by incubating the whole blood cultures with molasses (at various concentrations ranging from 12.5 to 800 mg/ml) understimulated (with LPS and PHA) and unstimulated conditions for 18 h by in vitro test system. This study showed the levels of IL-6 and IL-10 were increased in the absence of stimulus. Moreover, adding of 800 mg/ml of molasses to the unstimulated blood cultures caused significantly higher IL-6 and IL-10 values compared to control groups. On the contrary, molasses samples did not affect the IL-6, IL-10, and IFN-γ secretion in the presence of LPS and PHA.[31]
Risk of bias
The detailed results of the quality assessment are provided in Supplementary Tables 2 and 3 (S2). Based on the SYRCLE’s RoB tool, all three animal studies were judged to have low risk of bias according to the sequence generation, baseline characteristics, random housing, incomplete outcomes, and section of the reported results.[28,29,30] All studies considered as high risk of bias due to the lack of blinding of the intended interventions (performance bias) and outcome assessors (detection bias).[28,29,30] Studies were judged to have some concerns arising from random outcome assessment[28,29] and adequate allocation concealment.[28,29,30] According to the ToxRTool, numerical results lead to studies that were rated as category 1[32] and category 2.[31]
Supplementary Table 2.
Quality assessment (animal study)
| Selection bias | Performance bias | Detection bias | Attrition bias h | Reporting bias i | Other j | Total score | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|||||||||
| a | b | c | d | e | f | g | |||||
| Sánchez-Tapia et al., 2019 | yes | yes | unclear | yes | no | unclear | no | yes | yes | yes | 6 |
| Elayaraja et al., 2020 | yes | yes | unclear | yes | no | yes | no | yes | yes | yes | 7 |
| Singh et al., 2010 | yes | yes | unclear | yes | no | unclear | no | yes | yes | yes | 6 |
*a, Sequence generation; b, Baseline characteristics; c, Allocation concealment; d, Random housing; e, Blinding; f, Random outcome assessment; g, Blinding; h, Incomplete outcome data; I, Selective outcome reporting; j, Other sources of bias
Supplementary Table 3.
Quality assessment (in vitro study)
| I | II | III | IV | V | Total score | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|||||||||||||||
| a | b | c | d | e | f | g | h | i | j | k | l | m | n | o | p | q | r | ||
| Rahiman and Pool, 2013 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 15 |
| Rahiman and Pool, 2010 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 13 |
*I, test substance identification; II, test system characteristics; III, study design description; IV, study results documentation; V, plausibility of study design and data. *a, identification of test substance; b, purity of the test substance; c, source/origin of the test substance; d, nature and/or physico-chemical properties of the test substance; e, description of the test system; f, source/origin of the test system; g, test system properties; h, the method of administration; I, doses of administration; j, frequency and duration of exposure; k, negative controls; l, positive controls; m, the number of replicates; n, the study endpoint (s) and their method (s); o, description of the study results; p, the statistical method for data analysis; q, study design; r, reliability of results
Discussion
The current systematic review aimed to determine the effect of unrefined sugarcane products on inflammation by identifying five intervention studies that addressed the main objective. Unfortunately, a few studies have investigated such effects, and the overall picture needs to be more comprehensive. Furthermore, since we could not find any human interventional study on this topic, more research is needed to firmly conclude the effect of unrefined and less refined dietary sugar on inflammatory biomarkers.
We determined that administration of unrefined sugar (brown sugar, sugarcane molasses, jaggery) could reduce inflammation as indicated by various inflammatory biomarkers, including IL-6, IL-10, IL-1β, TNF-α, IF-g, and NF-κB in animal models and in vitro test systems.
Different methods, including experimental animals, tissue, and cell cultures, as well as computational simulations and clinical studies, have been used in order to find ways to treat human diseases and disorders. All methods have their own pros and cons.[33] In this review, the evidence has been provided based on in vitro and animal models. In vitro models using tissue slices or cultured cells provided valuable information on the beneficial effects of the polyphenols. In order to interpret and extrapolate the outcomes, great care is required. The concentrations in the in vitro design commonly range from mmol/l to mmol/l, while, after a regular dietary intake, the concentrations of plasma metabolites rarely exceed nmol/l. Thus, the higher doses in the in vitro models may cause forced positive results. Moreover, in animals and humans ingesting a specific polyphenol, cells are consistently exposed to this polyphenol, and prolonged exposure may have significant effects, even if the tissue concentration in tissues is low.[34] Animal studies are widely used in antioxidant research and aim to overcome differences between the in vitro models and the whole organism, as well as the restrictions of human trials.[35] Although animal models provide valuable information, differences between human and animal genomes may also lead to potential difficulties in extrapolating the data. However, most of the findings on the tissue distribution of polyphenols have been derived from earlier animal research.[34]
Previous studies showed that unrefined sugarcane products have antioxidant activity due to phytochemicals and polyphenol components.[14,15,36] The phytochemicals may also reduce the risk of DNA damage, inflammation, and various oxidative stress-related diseases.[10,19,20,21,22]
Rahiman et al. showed that the administration of molasses has both inflammatory and anti-inflammatory potential and impact the cytokines regulating humoral immune system by increasing the synthesis of IL-6 and IL-10 under unstimulated conditions.[31] These findings were confirmed by the same group later (2013), which investigated the effect of sugarcane molasses on human cell blood cultures and supported its ability to enhance inflammatory activity.[32] The work reported that IL-6 secretion of both stimulated and unstimulated cultures exposed to sugarcane molasses significantly exceeded that of the control. However, no changes were found for IL-10 and IF-g secretion. In addition, such effects were not seen for brown sugar. Based on their findings, sugarcane molasses induced inflammatory (IL-6) and anti-inflammatory (IL-10) responses. As a result, this initiated efficient immune defense mechanisms against pathogens and their toxins by increasing B cell activity, which proves the potential ability of molasses as a therapeutic agent on inflammation in a patient with a depressed immune system such as cancer.[31,32] Interleukin-6 is a cytokine with a wide range of biological activities in immune regulation, hematopoiesis, inflammation, and initiation of tumors.[37] Environmental stress factors such as infections and tissue injuries generate the expression of IL-6. The expression activates host–defense mechanisms against stress. Interleukin-6 is a mediator for notification of some emergent event generated in an infectious lesion and sends out a warning signal to the entire body.[38]
Singh et al. indicated that jaggery effectively antagonizes many of the adverse effects of arsenic by suppressing the serum levels of IL-1β, IL-6, and TNF-α in the jaggery-supplemented arsenic-exposed groups compared to the arsenic-alone-treated groups. Additionally, the serum levels of antioxidant enzymes (total antioxidant status, glutathione peroxidase, and glutathione reductase) were reduced in arsenic-exposed groups without jaggery treatment. In contrast, their levels increased in combined treatment groups (arsenic and jaggery). Such effects may be due to the presence of vitamins, minerals, and phytochemical components of jaggery, which may help minimize the molecular and cellular oxidative damage caused by arsenic.[28]
NCS (e.g., jaggery and brown sugar) includes a high amount of vitamin and total minerals (32.50–1000.79 mg/100 g) compared to refined sugar (1.95–81.20 mg/100 g), vitamin E (0.00–111.30 mg/100 g), niacin (2.14–7.00 mg/100 g), vitamin C (0.00–7.00 mg/100 g), and vitamin D2 (0.00–6.50 mg/100 g) are the most abundant vitamins in NCS, whereas no vitamin content has been found in refined sugar.[9] Since the antiradical activity of unrefined sugar significantly correlates to the total phenolic and flavonoid content, a higher amount of antioxidant capacity was determined in unrefined sugar compared to refined sugar. Unrefined sugarcane products presented a higher total phenolic content ranging from 37.00 to 1664.90 mg GAE/100 g compared to refined sugars (0.00–3.15 mg GAE/100 g).[9] The main phenolic compounds of sugarcane juice are flavonoids such as naringenin, flavones, tricin, apigenin, luteolin, and phenolic acids (caffeic, ferulic, chlorogenic, and cinnamic acids).[9,39] Azlan et al. reported that minimally refined brown sugar (MRBS) had fair amounts of antioxidants. The less refining process resulted in the retention of its phenolic acids, flavonoids, vitamins, and minerals.[40]
On the other hand, heavy sugar refinement or processing causes a considerable decrease in antioxidants (polyphenols) in sugarcane.[40] A recent randomized crossover clinical trial showed that the intake of jelly prepared using MRBS resulted in a significantly higher total antioxidant capacity (TAC) in healthy participants than in granulated white sugar jelly.[41] Thus, the health benefits of less refined sugar may encourage both industries and the public to substitute white sugar with less refined sugar in sugar-based food products and dietary sugar intake to increase antioxidant intake. The importance of antioxidant activity in NCS can be observed by the frequent usage of refined sugar consumption calculated per capita in Spain (~340 g/month). Replacement of refined sugar for an unrefined alternative could easily increase the antioxidant content of the diet from 1.4 to 9.1 mmol TE/month (depending on the type of unrefined sugar). Interestingly, the antioxidant intake from fruits with considerable TAC such as blackberries is negligible due to the infrequent per capita consumption among the Spanish population.[7,9]
Jaggery has a powerful influence on Nile tilapia’s innate immunity through good innovation of various immune cells and enzymes and upregulating the mRNA expression of LYS, TNF-α, and IL-1β. These changes showed that their influence on immune response activation of tilapia enhanced its resistance to bacterial (Aeromonas hydrophila and Streptococcus agalactiae) infection according to significant increment in the expression of IL-β and TNF-α.[30] Furthermore, the biofloc-treated groups had better growth performance, higher survival rate, and higher antioxidant activity according to superoxide dismutase, malondialdehyde, catalase, and glutathione values. This study suggests that jaggery is a new carbon source with unique properties that satisfy all considerations of biofloc technology (BFT) in an eco-friendly manner. BFT is a cost-effective system with high productivity and zero water exchange in which the nutrients can be recycled (the retention of organic nitrogenous wastes converting into a proteinaceous compound). Various aquatic species could use the technology as a food source.[42] Carbon amendment is a critical factor in BFT systems, and the C/N ratio was manipulated by the frequent addition of carbohydrates or by elevating the carbon level in fish feed. As a result, exploring new sugar with exclusive characteristics (ease of availability, reasonable price, and effective bacterial assimilation) for sustainable and profitable aquaculture is essential.[30]
In addition, Sánchez-Tapia determined that sucrose, sucralose, and glucose with and without an HF stimulated the inflammatory pathways mediated by TLR4. At the same time, the rats fed with BS had the lowest expression of RNA and protein abundance of TLR4, TNF-α, and MyD88.[29] The finding of the recent in vitro study demonstrated that NCS-based natural products have an anti-inflammatory effect on human monocytes by interfering with TLR4 signaling.[43] Among the family members of toll-like receptors, TLR4 has been demonstrated as particularly important in adipose tissue inflammation, and MyD88 is essential for the induction of proinflammatory cytokines.[44,45] It has been recognized that the interaction of these two receptors promotes the transcriptional activity of NF-κB pathway and later thereafter induces the activation of JKN, leading to inflammatory responses.[45,46] Moreover, the expression of the TNF-α gene was stimulated by sucralose, steviol glycosides with sucrose, sucrose, glucose, and fructose. This study indicated that the protein abundance of NF-κB was induced mainly by sucralose and sucrose and to a lesser extent by BS in the groups fed with HF. This study demonstrated that the intake of different sweeteners showed various effects on the inflammatory state. Therefore, the consumer should be aware of the type of sweetener and the fat content in the diet to prevent further inflammatory-related metabolic complications.[29]
TNF-α is a major proinflammatory cytokine for inducing acute and chronic inflammatory responses involved in the pathogenesis of a variety of chronic inflammatory diseases, such as type 2 diabetes, rheumatoid arthritis, and inflammatory bowel disease.[47] The NF-κB signaling pathway is a significant route activated by TNF-α and interleukin.[48,49] Activation of transcription factor translocate the nucleus, regulates of inflammatory mediators, and induces the upregulation of inflammatory gene expression by binding with the DNA.[48,50] In a nutshell, one of the primary targets for the prevention and treatment of many chronic diseases, including cancer, CVD, and diabetes, is using anti-inflammatory compounds (e.g., polyphenol) to suppress or inhibit such inflammatory and pro-inflammatory mediators.[48]
Limitations
Although this systematic review is the first that investigated the effect of unrefined sugarcane products on inflammation and the broad range of inflammatory biomarkers included, there were some limitations. First, despite our comprehensive search to include all types of sugarcane products and inflammatory biomarkers, without minimizing the type of study design, the number of trials in this field is minimal, with sustainable heterogenicity. Second, we could not find any human intervention trials to investigate the impact of unrefined dietary sugar on inflammation. Additionally, the amount of the investigated sugars, exposures, interventions, and study populations differs widely. Besides, the studies are almost similar in types of investigated sugar (jaggery, molasses, brown sugar), design (in vitro/animal), and outcome measurements (IL-6, TNF-α, and IL-1β). Since the current systematic review is based on a few published studies and there is a lack of sufficient data, especially from human interventions, more studies are suggested and needed to conclude the definite effect of NCS on inflammation.
Conclusion
The present review provides evidence that the administration of NCS products such as jaggery contributes more to induce inflammatory response than other sweeteners, including dietary sucrose and artificial sweeteners, based on in vitro and animal experiments. However, it is noteworthy that this conclusion is based on a limited number of available studies in this area of research without including any human trials due to the unavailability of published data. Thus, to provide definitive proofs of NCS’s protective role, additional evidence from further human studies with quality design is highly warranted.
Ethical Considerations
Nil.
Code of Ethics
Nil.
Author contributions
All authors contributed to the study’s conception, design, and data collection. SE and AA were involved in the initial concept of the study, investigation, prepared the manuscript, and reviewed the paper. The authors have seen and approved the final version of the manuscript.
Financial support and sponsorship
This study was funded by a Research Grant (no. 6300256-14001) to Prof Azrina Azlan, Department of Nutrition, Faculty of Medicine and Health Sciences, Universiti Putra Malaysia.
Conflicts of interest
There are no conflicts of interest.
Acknowledgment
We would like to acknowledge the library facilities of Universit Putra Malaysia that made this review possible.
References
- 1.Grembecka M. Natural sweeteners in a human diet. Rocz Państwowego Zakładu Hig. 2015;66:195–202. [PubMed] [Google Scholar]
- 2.Harper AE. U. S. Department of Agriculture and U. S. Department of Health and Human Services. Dietary guidelines for Americans, 2020-2025. [[Last accessed on 2022 Jan10]];2020 December; Available from: DietaryGuidelines.gov. [Google Scholar]
- 3.Johnson RK, Appel LJ, Brands M, Howard B V, Lefevre M, Lustig RH, et al. Dietary sugars intake and cardiovascular health a scientific statement from the american heart association. Circulation. 2009;120:1011–20. doi: 10.1161/CIRCULATIONAHA.109.192627. [DOI] [PubMed] [Google Scholar]
- 4.Jiao Y, Wang Y. The effects of sweeteners and sweetness enhancers on obesity and diabetes:A review. J Food Bioact. 2018;4:107–16. [Google Scholar]
- 5.Lemos BS, Fernandez ML, Santos MESM. Effects of consuming sweeteners on metabolic disorders. J Nutr Food Res Technol. 2018;1:34–8. [Google Scholar]
- 6.Prinz P. The role of dietary sugars in health:Molecular composition or just calories? Eur J Clin Nutr. 2019;73:1216–23. doi: 10.1038/s41430-019-0407-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Seguí L, Calabuig-Jiménez L, Betoret N, Fito P. Physicochemical and antioxidant properties of non-refined sugarcane alternatives to white sugar. Int J Food Sci Technol. 2015;50:2579–88. [Google Scholar]
- 8.Yeboah J, Santoro AM, Arrieta-Escobar JA, Caballero IM, Orjuela A, Novoa CF, et al. Heuristic-based computer-aided design of ice creams and validation by using jaggery as refined sugar substitute. Chem Eng Res Des. 2022;184:256–66. [Google Scholar]
- 9.Ebadi S, Azlan A. Nutritional composition and role of non-centrifugal sugar (NCS) in human health. Curr Nutr Food Sci. 2021;17:249–57. [Google Scholar]
- 10.Jaffé WR. Health effects of non-centrifugal sugar (NCS):A review. Sugar Tech. 2012;14:87–94. [Google Scholar]
- 11.Arif S, Batool A, Nazir W, Khan RS, Khalid N. Physiochemical Characteristics Nutritional Properties and Health Benefits of Sugarcane Juice. Elsevier Inc. 2019 Available from: https://doi.org/10.1016/B978-0-12-815270-6.00008-6. [Google Scholar]
- 12.Eggleston G, Aita G, Triplett A. Circular sustainability of sugarcane:Natural, nutritious, and functional unrefined sweeteners that meet new consumer demands. Sugar Tech. 2021;23:964–73. [Google Scholar]
- 13.Sampaio MRF, Machado MC, Lisboa MT, Vieira MA, Zimmer TBR, Otero DM, et al. Physicochemical characterization and antioxidant activity of refined and unrefined sugarcane products from Southern Brazil. Sugar Tech. 2022 Available from: https://doi.org/10.1007/s12355-022-01146-y. [Google Scholar]
- 14.Duarte-Almeida JM, Salatino A, Genovese MI, Lajolo FM. Phenolic composition and antioxidant activity of culms and sugarcane (Saccharum officinarum L.). products. Food Chem. 2011;125:660–4. [Google Scholar]
- 15.Kong F, Yu S, Zeng F, Wu X. Preparation of antioxidant and evaluation of the antioxidant activities of antioxidants extracted from sugarcane products. J Food Nutr Res. 2015;3:458–63. [Google Scholar]
- 16.Zheng R, Su S, Zhou H, Yan H, Ye J, Zhao Z, et al. Antioxidant/antihyperglycemic activity of phenolics from sugarcane (Saccharum officinarum L.). bagasse and identification by UHPLC-HR-TOFMS. Ind Crops Prod. 2017;101:104–14. [Google Scholar]
- 17.Vinayagam R, Jayachandran M, Xu B. Antidiabetic effects of simple phenolic acids:A comprehensive review. Phyther Res. 2016;30:184–99. doi: 10.1002/ptr.5528. [DOI] [PubMed] [Google Scholar]
- 18.Ji J, Flavel M, Yang X, Chen OCY, Downey L, Stough C, et al. A polyphenol rich sugarcane extract as a modulator for inflammation and neurological disorders. PharmaNutrition. 2020;12:100187. doi:10.1016/j.phanu.2020.100187. [Google Scholar]
- 19.Abbas SR, Sabir SM, Ahmad SD, Boligon AA, Athayde ML. Phenolic profile, antioxidant potential and DNA damage protecting activity of sugarcane (Saccharum officinarum) Food Chem. 2014;147:10–6. doi: 10.1016/j.foodchem.2013.09.113. [DOI] [PubMed] [Google Scholar]
- 20.Bucio-Noble D, Kautto L, Krisp C, Ball MS, Molloy MP. Polyphenol extracts from dried sugarcane inhibit inflammatory mediators in an in vitro colon cancer model. J Proteomics. 2018;177:1–10. doi: 10.1016/j.jprot.2018.02.009. [DOI] [PubMed] [Google Scholar]
- 21.Luo Y, Li S, Ho C-T. Sugarcane rind:Applications and health benefits:A review. J Food Bioact. 2018;3:1–7. [Google Scholar]
- 22.Zhang YJ, Gan RY, Li S, Zhou Y, Li AN, Xu DP, et al. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules. 2015;20:21138–56. doi: 10.3390/molecules201219753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hanhineva K, Törrönen R, Bondia-Pons I, Pekkinen J, Kolehmainen M, Mykkänen H, et al. Impact of dietary polyphenols on carbohydrate metabolism. Int J Mol Sci. 2010;11:1365–402. doi: 10.3390/ijms11041365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Moher D, Liberati A, Tetzlaff J, Altman DG. Preferred reporting items for systematic reviews and meta-analyses:The PRISMA statement. J Clin Epidemiol. 2009;62:1006–12. doi: 10.1016/j.jclinepi.2009.06.005. [DOI] [PubMed] [Google Scholar]
- 25.Higgins J, Green S. Chapter 22:Overview of reviews. John Wiley &Sons Cochrane Handbook for Systematic Reviews of Interventions. 2008 [Google Scholar]
- 26.Tran L, Tam DNH, Elshafay A, Dang T, Hirayama K, Huy NT. Quality assessment tools used in systematic reviews of in vitro studies:A systematic review. BMC Med Res Methodol. 2021;21:1–13. doi: 10.1186/s12874-021-01295-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hooijmans CR, Rovers MM, De Vries RBM, Leenaars M, Ritskes-Hoitinga M, Langendam MW. SYRCLE's risk of bias tool for animal studies. BMC Med Res Methodol. 2014;14:1–9. doi: 10.1186/1471-2288-14-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Singh N, Kumar D, Lal K, Raisuddin S, Sahu AP. Adverse health effects due to arsenic exposure :Modification by dietary supplementation of jaggery in mice. Toxicol Appl Pharmacol. 2010;242:247–55. doi: 10.1016/j.taap.2009.10.014. [DOI] [PubMed] [Google Scholar]
- 29.Sánchez-Tapia M, Martínez-Medina J, Tovar AR, Torres N. Natural and artificial sweeteners and high fat diet modify differential taste receptors, insulin, and TLR4-mediated inflammatory pathways in adipose tissues of rats. Nutrients. 2019;11:880. doi: 10.3390/nu11040880. doi:10.3390/nu11040880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Elayaraja S, Mabrok M, Algammal A, Sabitha E, Rajeswari MV, Zágoršek K, et al. Potential influence of jaggery-based biofloc technology at different C:N ratios on water quality, growth performance, innate immunity, immune-related genes expression profiles, and disease resistance against Aeromonas hydrophila in Nile tilapia (Oreochrom niloticus) Fish Shellfish Immunol. 2020;107:118–28. doi: 10.1016/j.fsi.2020.09.023. [DOI] [PubMed] [Google Scholar]
- 31.Rahiman F, Pool EJ. The effects of saccharum officinarium (SUGAR CANE) molasses on cytokine secretion by human blood cultures. J Immunoass Immunochem. 2010;31:148–59. doi: 10.1080/15321811003617453. [DOI] [PubMed] [Google Scholar]
- 32.Rahiman F, Pool EJ. The in vitro effects of artificial and natural sweeteners on the immune system using whole blood culture assays. J Immunoass Immunochem. 2013;35:26–36. doi: 10.1080/15321819.2013.784197. [DOI] [PubMed] [Google Scholar]
- 33.Saeidnia S, Manayi A, Abdollahi M. From in vitro experiments to in vivo and clinical studies;Pros and cons. Curr Drug Discov Technol. 2016;12:218–24. doi: 10.2174/1570163813666160114093140. [DOI] [PubMed] [Google Scholar]
- 34.Visioli F, de la Lastra CA, Andres-Lacueva C, Aviram M, Calhau C, Cassano A, et al. Polyphenols and human health:A prospectus. Crit Rev Food Sci Nutr. 2011;51:524–46. doi: 10.1080/10408391003698677. [DOI] [PubMed] [Google Scholar]
- 35.Vladimir-Knežević S, Blažeković B, Bival Štefan M, Babac M. Plant polyphenols as antioxidants influencing the human health, phytochemicals as nutraceuticals- Global Approaches to Their Role in Nutrition and Health. InTech. 2012. [[Last accessed on 2022 Aug 12]]. pp. 155–80. Available from: http://www.intechopen.com/books/phytochemicals-as-nutraceuticals-global-approaches-to-their-role-in- nutrition-and-health/plant-polyphenols-as-antioxidants-influencing-the-human-health .
- 36.Meerod K, Weerawatanakorn M, Pansak W. Impact of sugarcane juice clarification on physicochemical properties, some nutraceuticals and antioxidant activities of non-centrifugal sugar. Sugar Tech. 2019;21:471–80. [Google Scholar]
- 37.Kishimoto T. IL-6:From its discovery to clinical applications. Int Immunol. 2010;22:347–52. doi: 10.1093/intimm/dxq030. [DOI] [PubMed] [Google Scholar]
- 38.Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014;6:a016295. doi: 10.1101/cshperspect.a016295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zidan D, Azlan A. Non-Centrifugal Sugar (NCS) and health:A review on functional components and health benefits. Appl Sci. 2022;12:460. doi:10.3390/app12010460. [Google Scholar]
- 40.Azlan A, Khoo HE, Sajak AAB, Aizan Abdul Kadir NA, Yusof BNM, Mahmood Z, et al. Antioxidant activity, nutritional and physicochemical characteristics, and toxicity of minimally refined brown sugar and other sugars. Food Sci Nutr. 2020;8:5048–62. doi: 10.1002/fsn3.1803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Azlan A, Ebadi S, Yusof BNM, Othman NMH, Kannar D, Sultana S, et al. Satiety, glycemic profiles, total antioxidant capacity, and postprandial glycemic responses to different sugars in healthy Malaysian adults. Nutrition. 2022;97:111551. doi: 10.1016/j.nut.2021.111551. doi:10.1016/j.nut. 2021.111551. [DOI] [PubMed] [Google Scholar]
- 42.Faizullah MM, Rajagopalsamy C, Ahilan B, Dani D. Application of biofloc technology (BFT) in the aquaculture system. J Entomol Zool Stud. 2019;7:204–12. [Google Scholar]
- 43.Rueda-Gensini L, Serna JA, Bolaños N, Rodriguez J, Cruz JC, Muñoz-Camargo C. Investigating the role of thermal processing on the anti-inflammatory activity of non-centrifugal cane sugar:Implications on cytokine secretion and TLR4 signaling. Front Pharmacol. 2022;13:1–16. doi: 10.3389/fphar.2022.905347. doi:10.3389/fphar.2022.905347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.McKernan K, Varghese M, Patel R, Singer K. Role of TLR4 in the induction of inflammatory changes in adipocytes and macrophages. Adipocyte. 2020;9:212–22. doi: 10.1080/21623945.2020.1760674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Könner AC, Brüning JC. Toll-like receptors:Linking inflammation to metabolism. Trends Endocrinol Metab. 2011;22:16–23. doi: 10.1016/j.tem.2010.08.007. [DOI] [PubMed] [Google Scholar]
- 46.Zhang D, Shen X, Pang K, Yang Z, Yu A. VSIG4 alleviates intracerebral hemorrhage induced brain injury by suppressing TLR4-regulated inflammatory response. Brain Res Bull. 2021;176:67–75. doi: 10.1016/j.brainresbull.2021.08.008. [DOI] [PubMed] [Google Scholar]
- 47.Kawaguchi K, Matsumoto T, Kumazawa Y. Effects of antioxidant polyphenols on TNF-Alpha-related diseases. Curr Top Med Chem. 2011;11:1767–79. doi: 10.2174/156802611796235152. [DOI] [PubMed] [Google Scholar]
- 48.Upadhyay S, Dixit M. Role of polyphenols and other phytochemicals on molecular signaling. Oxid Med Cell Longev. 2015;2015:504253. doi: 10.1155/2015/504253. doi:10.1155/2015/504253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Park KM, Bowersa WJ. Tumor necrosis factor-alpha mediated signaling in neuronal homeostasis and dysfunction. Bone. 2010;22:977–83. doi: 10.1016/j.cellsig.2010.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Serafini M, Peluso I. Functional foods for health:The interrelated antioxidant and anti-inflammatory. Bentham Sci. 2016;6128:6701–15. doi: 10.2174/1381612823666161123094235. [DOI] [PMC free article] [PubMed] [Google Scholar]