

Abstract
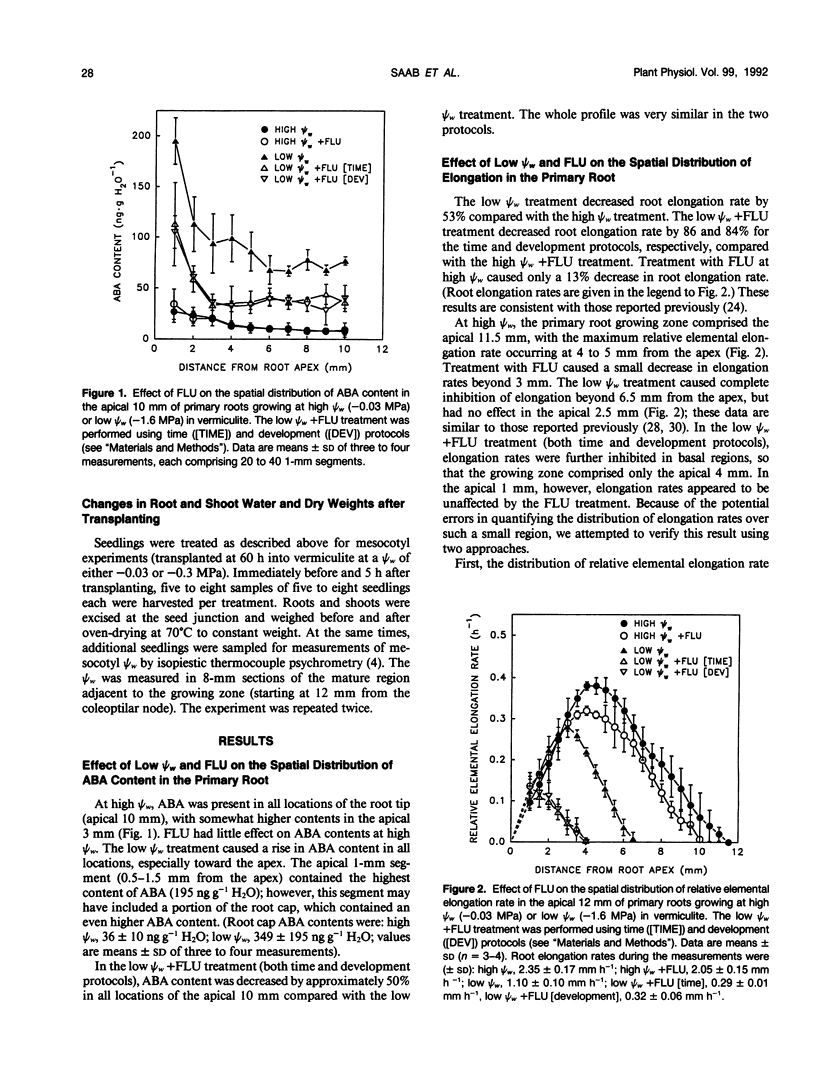
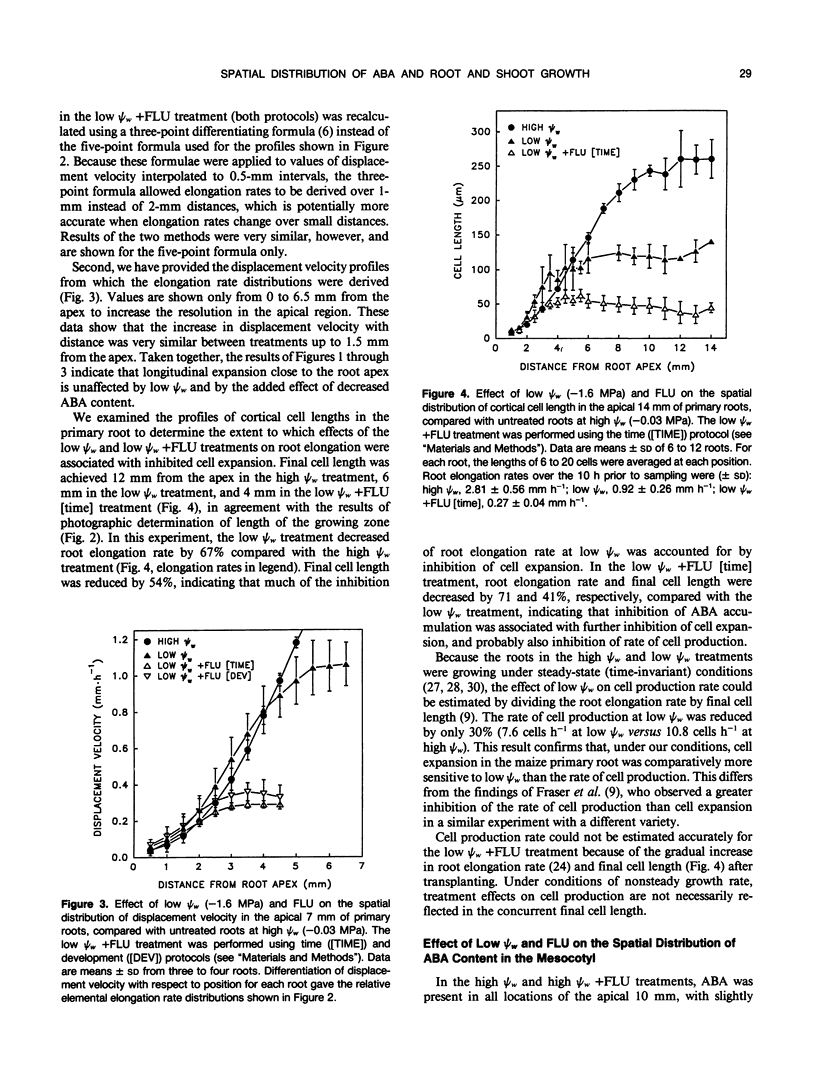
Previous work showed that accumulation of endogenous abscisic acid (ABA) acts both to maintain primary root growth and inhibit shoot growth in maize seedlings at low water potentials (ψw) (IN Saab, RE Sharp, J Pritchard, GS Voetberg [1990] Plant Physiol 93: 1329-1336). In this study, we have characterized the growth responses of the primary root and mesocotyl of maize (Zea mays L. cv FR27 × FRMo 17) to manipulation of ABA levels at low ψw with a high degree of spatial resolution to provide the basis for studies of the mechanism(s) of ABA action. In seedlings growing at low ψw and treated with fluridone to inhibit carotenoid (and ABA) biosynthesis, ABA levels were decreased in all locations of the root and mesocotyl growing zones compared with untreated seedlings growing at the same ψw. In the root, low ψw (−1.6 megapascals) caused a shortening of the growing zone, as reported previously. The fluridone treatment was associated with severe inhibition of root elongation rate, which resulted from further shortening of the growing zone. In the mesocotyl, low ψw (−0.3 megapascal) also resulted in a shortened growing zone. In contrast with the primary root, however, fluridone treatment prevented most of the inhibition of elongation and the shortening of the growing zone. Final cell length measurements indicated that the responses of both root and mesocotyl elongation to ABA manipulation at low ψw involve large effects on cell expansion. Measurements of the relative changes in root and shoot water contents and dry weights after transplanting to a ψw of −0.3 megapascal showed that the maintenance of shoot elongation in fluridone-treated seedlings was not attributable to increased water or seed-reserve availability resulting from inhibition of root growth. The results suggest a developmental gradient in tissue responsiveness to endogenous ABA in both the root and mesocotyl growing zones. In the root, the capacity for ABA to protect cell expansion at low ψw appears to decrease with increasing distance from the apex. In the mesocotyl, in contrast, the accumulation of ABA at low ψw appears to become increasingly inhibitory to expansion as cells are displaced away from the meristematic region.
Full text
PDF




Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- Cosgrove D. Biophysical control of plant cell growth. Annu Rev Plant Physiol. 1986;37:377–405. doi: 10.1146/annurev.pp.37.060186.002113. [DOI] [PubMed] [Google Scholar]
- Fraser T. E., Silk W. K., Rost T. L. Effects of low water potential on cortical cell length in growing regions of maize roots. Plant Physiol. 1990 Jun;93(2):648–651. doi: 10.1104/pp.93.2.648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hydén H., Lange P. W. Correlation of the S100 brain protein with behavior. Exp Cell Res. 1970 Sep;62(1):125–132. doi: 10.1016/0014-4827(79)90514-7. [DOI] [PubMed] [Google Scholar]
- Saab I. N., Sharp R. E., Pritchard J., Voetberg G. S. Increased endogenous abscisic Acid maintains primary root growth and inhibits shoot growth of maize seedlings at low water potentials. Plant Physiol. 1990 Aug;93(4):1329–1336. doi: 10.1104/pp.93.4.1329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaer J. A., Mandoli D. F., Briggs W. R. Phytochrome-mediated cellular photomorphogenesis. Plant Physiol. 1983 Jul;72(3):706–712. doi: 10.1104/pp.72.3.706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp R. E., Hsiao T. C., Silk W. K. Growth of the Maize Primary Root at Low Water Potentials : II. Role of Growth and Deposition of Hexose and Potassium in Osmotic Adjustment. Plant Physiol. 1990 Aug;93(4):1337–1346. doi: 10.1104/pp.93.4.1337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp R. E., Silk W. K., Hsiao T. C. Growth of the maize primary root at low water potentials : I. Spatial distribution of expansive growth. Plant Physiol. 1988 May;87(1):50–57. doi: 10.1104/pp.87.1.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silk W. K., Walker R. C., Labavitch J. Uronide Deposition Rates in the Primary Root of Zea mays. Plant Physiol. 1984 Mar;74(3):721–726. doi: 10.1104/pp.74.3.721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spollen W. G., Sharp R. E. Spatial distribution of turgor and root growth at low water potentials. Plant Physiol. 1991 Jun;96(2):438–443. doi: 10.1104/pp.96.2.438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twente J. W., Twente J. A. Regulation of hibernating periods by temperature. Proc Natl Acad Sci U S A. 1965 Oct;54(4):1044–1051. [PMC free article] [PubMed] [Google Scholar]