

Abstract
Yams (Dioscorea species) are an important food resource in Madagascar, where both cultivated winged yam (D. alata) and wild edible yams are consumed. However, there is limited knowledge on the nutrient composition of wild edible yams in Madagascar, and on how they compare with the cultivated winged yam. Therefore, in this study, nine wild edible yam species, one with two subspecies from Madagascar (D. bako, D. buckleyana, D. irodensis, D. maciba, D. orangeana, D. pteropoda, D. sambiranensis subsp. bardotiae and subsp. sambiranensis, D. seriflora, and Dioscorea species Ovy valiha), were analyzed for their nutrient composition, compared with cultivated D. alata. They include 6/6 of the most favored wild edible yam species in Madagascar. New nutrient composition data (protein, carbohydrate/starch, energy, lipid, β‐carotene, and minerals) are presented for these nine wild edible yam species. The results show that they contain comparable levels of lipids and starch to D. alata, but none are better sources of protein than D. alata. The results show that D. irodensis contains a significantly higher β‐carotene content when compared to all other edible yams analyzed, and that D. buckleyana, D. irodensis, and D. sambiranensis subsp. bardotiae have a higher calcium content than cultivated D. alata, while all nine wild edible yam species analyzed contain a higher iron content, compared to cultivated D. alata. The nutrient composition data presented could provide new incentives to conserve wild edible yams and inform on strategies to select Dioscorea species for sustainable cultivation and use, providing opportunities to enhance future food security in Madagascar.
Keywords: carotenoids, Dioscorea, macronutrients, micronutrients, minerals, protein, yams
New information on the nutrient composition of nine edible yams (Dioscorea species) endemic to Madagascar is presented. None of the wild edible yams are a better source of protein compared to cultivated D. alata, but D. irodensis contains a significantly higher β‐carotene content compared to all other edible yams analyzed. Compared to cultivated D. alata, all nine wild edible yam species contain higher iron contents, while D. buckleyana, D. irodensis, and D. sambiranensis subsp. bardotiae have a higher calcium content.
1. INTRODUCTION
Yam tubers, which are the underground organs of species in the plant genus Dioscorea (Dioscoreaceae), are important food resources in Madagascar, an island in the Indian Ocean located off the eastern coast of Africa. Thirty‐eight Dioscorea species have been identified in Madagascar, representing 6% of the world's diversity of this genus; 30 of these species are endemic and occur in the wild, and they are virtually all edible (Wilkin et al., 2022). Wild edible Dioscorea species are found in all parts of the island, both in humid areas such as in the east of the country and in the more arid regions in the west and south.
While cultivated varieties of nonendemic D. alata L. (which is of southeast Asian origin) are available in Madagascar, their supply is limited; thus, rural communities harvest or buy locally sourced wild edible Dioscorea species as a supplementary food or as a substitute for rice during periods of hardship and famine. During these “hungry gaps”, particularly in the late dry season and early rainy season when other foods such as rice may be used up, the demand for wild edible yams for food and nutrition escalates, significantly increasing their price in local markets (Wilkin et al., 2022). Indeed, wild edible yams in Madagascar are an important seasonal food in certain regions such as Diana and Menable and are widely used in much of the island nation in times of hardship.
Although yam tubers are an important food resource in Madagascar, there remains limited published data on the nutrient composition of wild edible yams endemic to Madagascar (Jeannoda et al., 2007). Of the Dioscorea species known to occur in Madagascar, those that have been evaluated have almost all been sourced from other countries, so their nutrient composition may not reflect that of tubers from species harvested in Madagascar. Dioscorea alata tubers from south Asia contain a high proportion of starch (65%–84%) and approximately 7% protein (dry weight), though different cultivars may vary in their content of these macronutrients (Rinaldo, 2020; Wanasundera & Ravindran, 1994).
Wild edible yams in Madagascar are threatened by habitat degradation and by over or unsustainable use, thus risking food security and biodiversity loss, with some species (D. bako, D. buckleyana, D. irodensis, D. orangeana) already categorized as endangered according to the International Union for the Conservation of Nature (IUCN) Red List (IUCN, 2022). Dioscorea sambiranensis subsp. bardotiae is also categorized as endangered, while D. sambiranensis subsp. sambiranensis is categorized as vulnerable (IUCN, 2022). Dioscorea sambiranensis is divided into two subspecies based on the vegetative and infructescence traits (Wilkin et al., 2009). These challenges to biodiversity conservation and to the role of wild edible yams in food security are exacerbated by the impact of climate change, which may result in increased temperatures, altered precipitation patterns leading to floods and drought and extreme weather events, which are already concerns in Madagascar, impacting on agriculture and the economy and on food security (WHO & UN, 2022). The impact of climate change on food security is an additional concern that may intensify the triple burden of malnutrition (undernutrition, micronutrient deficiencies, and obesity) and metabolic and lifestyle factors for diet‐related noncommunicable diseases; indeed, in 2021, 43.1% of the adult population in Madagascar were considered undernourished, while 4.5% of the adult population were considered obese, and 3.9% had diabetes (WHO & UN, 2022). Furthermore, almost 8% of children under age 5 in Madagascar are affected by wasting, which is higher than the average (6%) of the Africa region (GNR, 2022).
New knowledge of the nutritional value of wild edible yams in Madagascar may facilitate conservation actions by providing a scientific evidence base, and therefore incentives, for their sustainable cultivation and use, in addition to providing new insights into their nutritional value that could enable informed dietary choices. In this context, the aims of this study are to assess the nutrient composition of tubers from nine wild edible yam species, one with two subspecies, from Madagascar (D. bako, D. buckleyana, D. irodensis, D. maciba, D. orangeana, D. pteropoda, D. sambiranensis subsp. bardotiae and subsp. sambiranensis, D. seriflora, and Dioscorea species Ovy valiha, an undescribed species), and to compare their nutrient content with that of the cultivated nonendemic winged yam, D. alata. The wild edible yams analyzed include 6/6 (D. bako, D. buckleyana, D. maciba, D. orangeana, D. sambiranensis, and D. seriflora) of the most favored wild edible yam species in Madagascar (Wilkin et al., 2022).
2. MATERIALS AND METHODS
2.1. Samples
The tubers of nine wild edible yam (Dioscorea) species, in addition to cultivated D. alata, were harvested in Madagascar. The Dioscorea species collected, their geographical collection locations, common names, and conservation status are shown in Table 1 (additional details, including GPS coordinates for collection locations and month and year of harvest, are shown in Table S1). Harvested tubers were dried (details are indicated in Table S1) and sliced, then stored in boxes at room temperature in the dark (protected from light) prior to their analysis for nutritional composition. For nutrient analyses, samples were ground to a fine powder using a coffee grinder.
TABLE 1.
Wild edible yam tubers collected: species, common names, and collection locations. a
| Dioscorea species | IUCN conservation status b | Common name | Collection location | District (collection location) |
|---|---|---|---|---|
| Dioscorea species | Unknown | Ovy valiha | Andranomadiro | Antsiranana |
| D. sambiranensis subspecies bardotiae Wilkin | Endangered | Ovy jaune | Betahitra | |
| D. pteropoda Boivin ex H. Perrier | Vulnerable | Ovindambo | Ambilo | |
| D. buckleyana Wilkin | Endangered | Taravy | Betahitra | |
| D. orangeana Wilkin | Endangered | Ovy jia | Ivovona | |
| D. irodensis Wilkin, Rajaonah & Randriamb. | Endangered | Ovy laglasy | Ivovona | |
| D. maciba Jum. & H.Perrier | Least Concern | Ovy cable | Mahagaga | |
| D. sambiranensis subspecies sambiranensis | Vulnerable | Angona | Ankariha | Ambanja |
| D. seriflora Jum. & H.Perrier | Least Concern | Ovy sofy | Anaboranosalama | |
| D. bako Wilkin | Endangered | Bako | Tsaraotana | Morondava |
2.2. Protein analysis
Protein content was determined from the total nitrogen content by the Kjeldahl method (Bradstreet, 1965). In brief, 0.25 g of powdered tuber sample was placed in a flask containing a solution of a concentrated sulfuric acid and Kjeldahl catalyst. After approximately 3 h, when the solution was translucent, 50 mL of sodium hydroxide (40%) was added to neutralize the pH. After the distillation phase, the distillate was collected in aqueous boric acid solution (4%) prior to titration with sulfuric acid (0.1 N). Protein content was estimated from the total nitrogen content of food using 6.25 as the conversion factor. Protein content was also determined using the colorimetric Bradford assay, as described previously (Maehre et al., 2018). Protein was extracted from each sample (500 mg yam powder) using an extraction buffer (0.1 M NaOH in 3.5% NaCl). The solution was vortexed and incubated for 30 min at 60°C, followed by centrifugation for 10 min at 5,000 rpm. The supernatant was collected and protein content was measured with a commercially available assay kit (Bio‐Rad, Inc.) using bovine serum albumin as a standard. Data are calculated as mg/g dry weight and presented as % values.
2.3. Carbohydrate and starch analysis
Total carbohydrate content was determined by subtracting the protein, lipid, ash, and water contents in the total weight of the tuber samples (FAO, 1998). Total starch content was measured using a Total Starch HK Assay Kit (K‐TSHK; Megazyme, Bray, Ireland). The assay followed the AOAC Official Method 996.11 for total starch determination (Englyst & Cummings, 1988; McCleary et al., 1994, 1997). Samples were assayed in triplicate. Total starch content was calculated in accordance with the manufacturer's (Megazyme, Bray, Ireland) instructions and data are presented as g/100 g dry weight.
2.4. Energy analysis
The energy value of tuber samples was determined using the Atwater method, based on the heat combustion of nutrients (1 g of protein provides 4 kcal/17 kJ; 1 g of fat provides 9 kcal/37 kJ; 1 g of carbohydrate provides 4 kcal/17 kJ; Atwater & Woods, 1896). The energy was also evaluated using the ballistic bomb calorimeter method to measure the gross energy content of each yam sample (Miller & Payne, 1959). Analysis was carried out using a Gallenkamp Ballistic Bomb Calorimeter (IKA©C1 Compact Calorimeter) and gross energy content is presented as kcal/100 g dry weight.
2.5. Lipid analysis
Lipid content was determined using the Folch method (Folch et al., 1957). In brief, 1.0 g tuber sample was added to a solution of chloroform/methanol (2/1, v/v) prior to homogenization and vacuum filtration. The lower phase of the filtrate was recovered prior to evaporation at 65°C, followed by steaming at 85°C.
2.6. Ash analysis
Ash content was determined by placing 5.0 g tuber material in a porcelain crucible prior to incineration in a muffle furnace at 550°C until white/gray ash consisting of inorganic material was obtained. After cooling in a desiccator, the mass of the ash was determined (AOAC International, 2000).
2.7. β‐Carotene analysis
All reactions were carried out in a darkened fume hood with all sample tubes wrapped in foil to protect samples from light. Carotenoids were extracted from samples of yam (500 mg) using a ternary solvent system added, which consisted of 10 mL hexane, 5 mL methanol, and 5 mL HPLC‐grade water (Bóna‐Lovász et al., 2013). The sample was first dissolved in hexane and methanol, followed by a 2‐min vortex. Tubes were wrapped with foil in order to protect the carotenoids from daylight and incubated overnight. Subsequently, HPLC‐grade water was added to each tube to separate the methanol from the solvent. The solution was then fully vortexed for 1 min and centrifuged (3000 rpm, 30 min) to further separate the hexane layer (containing the carotenoids). Following centrifugation, the upper hexane layer was collected and filtered. Absorbance was measured at 490 nm, and carotenoid content was measured using a standard curve with β‐carotene as the reference compound. The data are expressed as μg β‐carotene equivalents/100 g dry weight.
2.8. Mineral analysis
Mineral analysis to quantify zinc, iron, and calcium was performed using inductively coupled plasma–optical emission spectrometry (ICP‐OES, Thermo Fisher Scientific). Yam samples (50 mg) were mixed with 10 mL nitric acid (40%) and subjected to microwave digestion (CEM Corporation, UK). The total volume of each tube was adjusted to 14 mL with HPLC‐grade water, and yttrium (1 μg/mL final concentration) was added to each tube as an internal standard. Tubes were mixed and centrifuged for 5 min at 2000 rpm before analysis. Mineral content was calculated using standard curves generated using ICP‐grade standards for iron, zinc, and calcium, and data are presented as mg/100 g dry weight.
2.9. Statistical analysis
Data are presented as the mean ± SEM from at least three independent experiments. Statistical differences were determined using one‐way ANOVA followed by either Tukey's (for multiple comparisons) or Dunnett's (for comparison of wild yam species with D. alata) post hoc tests where appropriate. Differences where p < .05 were considered statistically significant. Statistical analysis was performed using SigmaPlot (version 14.5).
3. RESULTS
3.1. Protein content
Nine wild edible Dioscorea species and cultivated D. alata from Madagascar were analyzed over 12 harvest seasons from 2018 to 2020 (Table 2). Differences in protein content occurred between different harvest seasons, but protein content was not consistently high or low for any harvest season, thus indicating that harvest season is not a determinant for protein content in the Dioscorea species analyzed. The mean protein content of cultivated D. alata (6.4 g/100 g dry weight [DW] 2018–2019) did not differ significantly from the mean protein content of any of the wild edible species analyzed. In tubers harvested in 2019–2020, the mean protein content of D. alata (8.5 g/100 g DW) was significantly higher compared to all wild edible species analyzed (D. buckleyana p < .03; all other species p < .01), except for D. irodensis; D. sambiranensis subsp. bardotiae, and D. bako.
TABLE 2.
Protein, energy, starch, total carbohydrate, lipid, and ash contents of wild edible yams (Dioscorea species) and cultivated winged yam (D. alata) collected over different harvest seasons in Madagascar, calculated as dry weight.
| Dioscorea speciesa | Protein (%) rangeb Aug 2018–Mar 2019 (mean %) | Protein (%) rangec Aug 2019–Jun 2020 (mean %) | Energy ranged (kcal/100 g) Aug 2018–Mar 2019 (mean kcal/100 g) | Energy rangee (kcal/100 g) Aug 2019–Jun 2020 (mean kcal/100 g) | Starch content rangef (g/100 g) Aug 2019–Jun 2020 (mean g/100 g) | Total carbohydrate content rangeg (g/100 g) Aug 2019–Jun 2020 (mean g/100 g) | Total lipid content rangeh (g/100 g) Aug 2019–Jun 2020 (mean g/100 g) | Total ash content rangei (g/100 g) Aug 2019–Jun 2020 (mean g/100 g) |
|---|---|---|---|---|---|---|---|---|
| D. alata |
5.7–7.1 (6.4 ± 0.5) |
5.8–11.1 (8.5 ± 2.4) |
363.8–366.0 (364.9 ± 1.1) |
376.4–384.7 (380.0 ± 3.8) |
57.2–58.3 (57.8 ± 1.6) |
81.6–89.2 (84.9 ± 3.4) |
0.3–1.0 (0.7 ± 0.2) |
4.2–7.0 (5.9 ± 1.1) |
| D. bako |
2.7–6.1 (4.6 ± 0.3) |
4.6–10.0 (6.7 ± 2.5) |
361.3–372.3 (366.9 ± 3.0) |
384.8–397.1 (388.7 ± 4.8) |
41.7–64.3 (53.9 ± 2.6) |
79.9–92.6 (87.1 ± 4.7) |
0.9–2.8 (1.5 ± 0.8) |
1.9–7.3 (4.7 ± 2.0) |
| D. buckleyana |
3.7–10.8 (7.6 ± 0.6) |
4.1–7.0 (5.5 ± 1.1) |
304.0–372.4 (340.4 ± 18.1) |
373.9–388.6 (382.7 ± 5.4) |
34.7–54.0 (46.2 ± 2.4) |
86.5–89.0 (87.6 ± 0.9) |
0.8–1.8 (1.2 ± 0.4) |
3.9–7.6 (5.8 ± 1.2) |
| D. irodensis |
5.3–7.1 (6.5 ± 0.3) |
4.1–11.3 (6.3 ± 2.6) |
346.5–360.8 (360.1 ± 0.7) |
361.7–396.4 (382.9 ± 11.6) |
34.2–47.3 (42.3 ± 2.2) |
77.0–91.3 (86.3 ± 4.9) |
0.8–2.7 (1.4 ± 0.8) |
3.9–10.7 (6.0 ± 2.6) |
| D. maciba |
4.1–7.0 (5.4 ± 0.2) |
3.3–9.7 (5.2 ± 2.4) |
340.7–359.6 (355.2 ± 2.1) |
378.2–401.5 (387.9 ± 8.7) |
47.6–61.8 (54.9 ± 2.1) |
83.2–92.9 (89.5 ± 3.5) |
0.7–1.5 (1.0 ± 0.4) |
1.5–6.3 (4.3 ± 1.8) |
| D. orangeana |
2.4–5.1 (4.4 ± 0.2) |
2.7–6.5 (4.6 ± 1.2) |
320.8–377.2 (366,1 ± 5.9) |
377.2–390.8 (386.5 ± 4.8) |
44.0–58.5 (51.3 ± 1.8) |
88.2–91.2 (89.7 ± 1.3) |
0.7–1.5 (1.0 ± 0.4) |
3.3–6.7 (4.7 ± 1.2) |
| D. pteropoda |
5.0–7.8 (6.1 ± 0.4) |
2.4–5.2 (3.7 ± 1.0) |
344.3–373.7 (340.5 ± 12.3) |
390.2–400.7 (393.4 ± 4.1) |
26.0–54.3 (45.1 ± 3.5) |
90.4–92.8 (91.5 ± 1.0) |
0.9–2.0 (1.4 ± 0.4) |
2.3–4.2 (3.4 ± 0.7) |
| D. sambiranensis subsp. bardotiae |
2.6–6.1 (4.2 ± 0.2) |
3.5–8.6 (5.7 ± 1.0) |
353.8–366.3 (360.1 ± 3.4) |
380.2–393.2 (385.7 ± 4.6) |
39.9–58.3 (49.3 ± 2.2) |
84.2–91.6 (88.9 ± 2.9) |
0.7–1.0 (0.8 ± 0.1) |
2.6–6.2 (4.6 ± 1.2) |
| D. sambiranensis subsp. sambiranensis |
1.9–4.4 (3.1 ± 0.3) |
2.3–3.7 (3.0 ± 0.6) |
348.4–365.8 (356.7 ± 4.7) |
389.6–403.2 (393.1 ± 5.1) |
54.8–59.1 (57.8 ± 0.8) |
91.7–94.2 (92.9 ± 1.0) |
0.8–1.8 (1.0 ± 0.7) |
2.2–3.6 (3.0 ± 0.5) |
| D. seriflora |
4.0–7.3 (5.0 ± 0.3) |
2.2–5.0 (2.9 ± 1.0) |
352.3–376.2 (363.7 ± 6.5) |
386.1–398.1 (393.1 ± 4.1) |
51.6–61.8 (57.2 ± 1.5) |
90.1–93.9 (92.6 ± 1.4) |
0.7–1.5 (1.2 ± 0.4) |
2.3–4.3 (3.3 ± 0.7) |
| Dioscorea species (Ovy valiha) |
5.8–9.4 (6.9 ± 0.3) |
3.6–6.5 (4.6 ± 1.1) |
350.5–362.7 (357.0 ± 2.5) |
380.3–394.7 (386.1 ± 5.6) |
39.6–57.9 (51.8 ± 6.1) |
86.7–92.1 (89.7 ± 2.0) |
0.7–1.5 (1.0 ± 0.4) |
3.2–6.8 (4.7 ± 1.4) |
Note: aDried tubers consisting of parenchyma and periderm tissue. Protein content determined using the bBradford and cKjeldahl methods (n = 10–16); energy content determined using the dballistic bomb calorimeter and eAtwater methods (n = 2–12); starch content determined using the fAOAC Official Method 996.11 for total starch determination (n = 6–12); gtotal carbohydrate content determined by subtracting the protein, lipid, ash, and water contents (n = 10–12); lipid content determined by hFolch method (n = 10–12); and ash content determined by imass after incineration (n = 10–12). Data presented for the mean are shown as ± SEM.
3.2. Starch, carbohydrate, energy, and lipid contents
The nine wild edible Dioscorea species and cultivated D. alata from Madagascar were analyzed over four harvest seasons for starch content, which was not consistently high or low for any one harvest season. The mean starch content of cultivated D. alata (57.8 g/100 g DW) across all four harvest seasons did not differ significantly from the mean starch content of the other edible species analyzed (Table 2). The nine wild edible Dioscorea species and cultivated D. alata were analyzed over six harvest seasons for total carbohydrate content, which ranged from 84.9 g/100 g (mean, DW) in cultivated D. alata to 92.9 g/100 g (mean, DW) in D. sambiranensis subsp. sambiranensis (Table 2). Total carbohydrate content in D. alata was significantly lower than that observed for Dioscorea sp. (Ovy valiha; p < .04), D. pteropoda, D. sambiranensis subsp. sambiranensis, D. seriflora (all p < .01), D. orangeana (p < .04), and D. maciba (p < .04).
The nine wild edible yam species and cultivated D. alata from Madagascar were analyzed for energy (kcal/100 g; Table 2). The mean energy provided by the edible yam species across all harvest seasons ranged from 340.4 kcal/100 g (D. buckleyana) to 393.4 kcal/100 g (D. pteropoda). The energy calculated for D. alata was significantly lower (p < .01) than that observed for D. pteropoda, D. sambiranensis subsp. sambiranensis, and D. seriflora. The total lipid content was calculated as 0.7–1.5 g/100 g (mean, DW) across all of the edible yam species analyzed (Table 2), with the lipid content of all species analyzed not significantly different to that observed for D. alata.
3.3. β‐Carotene content
The edible yam species (wild and cultivated D. alata) were analyzed for total carotenoid content over eight harvest seasons between August 2018 and October 2019, with results expressed as β‐carotene equivalents (DW; Figure 1). Differences in the β‐carotene content of these species occurred between different harvest seasons, but the content was not consistently high or low for any one harvest season. This indicates that harvest season is not an informative indicator of β‐carotene content in the edible yam species analyzed.
FIGURE 1.
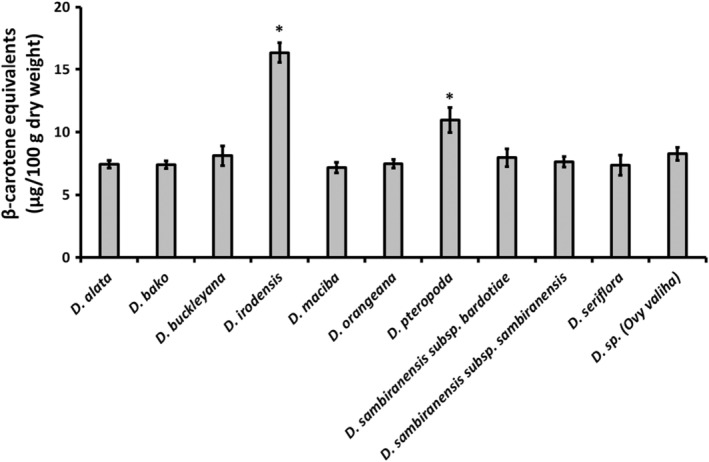
β‐Carotene content of nine wild edible yam species and cultivated Dioscorea alata across eight harvest seasons, expressed as β‐carotene equivalents (μg/100 g dry weight) for mean values (n = 6–8) + SEM. *Significantly different from D. alata (p < .01).
The β‐carotene content of D. irodensis was consistently higher than the β‐carotene content of all other species analyzed in all harvest seasons, except for the October 2019 harvest, when the β‐carotene content of D. pteropoda (10.6 μg/100 g) and D. sambiranensis subsp. bardotiae (10.0 μg/100 g) was higher than that of D. irodensis (9.6 μg/100 g), but not significantly. The average (mean) β‐carotene content of D. irodensis (16.4 μg/100 g) across all harvest seasons was significantly higher than the mean β‐carotene content of all other species analyzed, including D. alata (7.4 μg/100 g; p < .001; one‐way ANOVA and Tukey's post hoc test). The analysis also showed that the average β‐carotene content of D. pteropoda was significantly higher than D. alata (p < .02).
To assess the distribution of β‐carotene in yam tuber parenchyma and periderm, the two species with the highest mean β‐carotene content (D. irodensis: 16.4 μg/100 g and D. pteropoda: 11.0 μg/100 g) were subjected to additional analysis for β‐carotene content, compared with D. alata. The tuber parenchyma and periderm tissue of these three species across four harvest seasons (December 2019–June 2020) were separated and each tissue was analyzed separately to determine β‐carotene content. β‐Carotene was not significantly different between parenchyma and periderm tissue for each species analyzed (Figure 2), indicating that peeling the tubers of these species to remove the periderm would not adversely affect the β‐carotene content of the tubers (w/w). Consistent with the whole tuber β‐carotene content (Figure 1), the mean β‐carotene content of the separate parenchyma and periderm from D. irodensis (17.7 ± 0.6 and 18.0 ± 0.9 μg/100 g, respectively; mean ± SEM, n = 12) was significantly higher than the β‐carotene content of D. pteropoda (14.4 ± 0.8 and 13.2 ± 0.5 μg/100 g, respectively; mean ± SEM, n = 12) and D. alata (12.2 ± 0.4 and 11.9 ± 0.5 μg/100 g, respectively; mean ± SEM, n = 12; parenchyma, p < .001; periderm, p < .001; one‐way ANOVA and Tukey's post hoc test). The β‐carotene content of D. irodensis was also significantly higher than D. alata for both parenchyma and periderm in each harvest season (Figure 2).
FIGURE 2.

β‐Carotene content of separated parenchyma and periderm tissue of two wild edible yam species and cultivated Dioscorea alata across four harvest seasons, expressed as β‐carotene equivalents (μg/100 g dry weight) for mean values (n = 3) + SEM. *Significantly different from D. alata in each harvest season (p < .01).
3.4. Mineral and ash content
The total ash content, representing the inorganic (crude mineral) content determined for the edible yam species analyzed in this study was in the range of 3.0–6.0 g/100 g (mean, DW). The total ash content of D. alata was significantly higher than that observed in D. pteropoda (p < .03), D. sambiranensis subsp. sambiranensis (p < .01), and D. seriflora (p < .02). The wild edible yam species and cultivated D. alata were also analyzed for calcium, iron, and zinc content over four harvest seasons between August 2018 and March 2019, with DW content determined by ICP‐OES. Differences in the content of these minerals occurred between different harvest seasons, but the content was not consistently high or low for any one harvest season. This indicates that harvest season is not an informative indicator of calcium, iron, or zinc content in the edible yam species analyzed.
Although there were no statistical differences in mean mineral content across these four harvest seasons, the mean calcium content of D. buckleyana (2456 mg/kg), D. irodensis (2354 mg/kg), and D. sambiranensis subsp. bardotiae (2346 mg/kg) was 2.8–3 times greater than D. alata (833 mg/kg; Figure 3a). For iron, the content in D. alata (58 mg/kg) was lower than the mean iron content of all other species analyzed; the highest levels were found in D. irodensis (311 mg/kg; Figure 3b). The mean zinc content of D. alata (35 mg/kg) across the same period was higher than the mean zinc content of all other species analyzed, though not significantly (Figure 3c).
FIGURE 3.
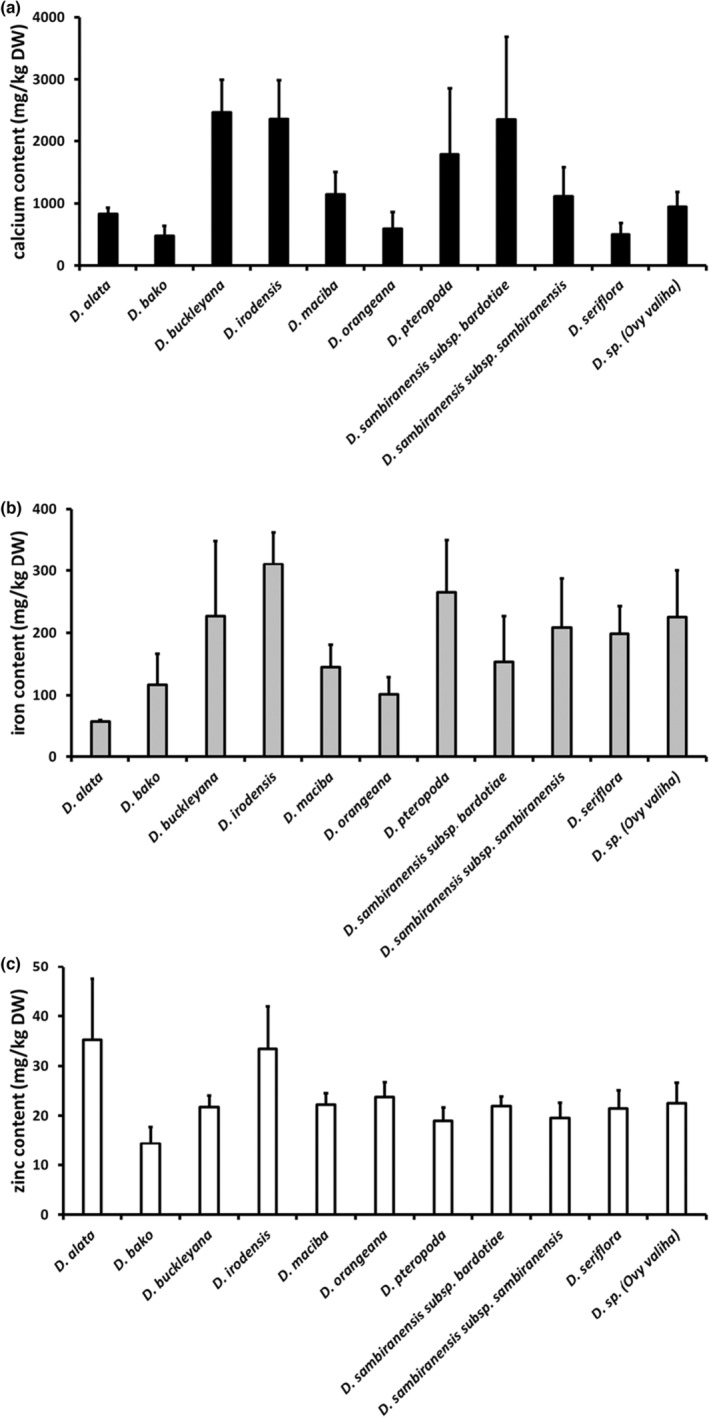
Mineral (a: calcium, b: iron, c: zinc) content of nine wild edible yam species and cultivated Dioscorea alata across four harvest seasons. (n = 3–4) + SEM.
4. DISCUSSION
In Madagascar, wild edible yams are consumed in addition to the nonendemic cultivated winged yam, Dioscorea alata. However, the nutrient composition of wild edible yams (an important Malagasy food resource, particularly in times of hardship and famine) has been understudied. Consequently, there is a need to further understand how wild edible yams may contribute to dietary nutrition in Madagascar and how their nutrient profiles compare with the cultivated D. alata. This study addresses this knowledge gap by presenting new nutrient composition data for nine wild edible Dioscorea species in comparison with cultivated D. alata, also sourced from Madagascar.
Dietary protein is required for optimal growth, development, and health in humans. Although protein from animal sources is considered to provide higher quantities and more balanced proportions of amino acids compared to protein from plant sources, plant‐based foods contribute 35% of protein globally to human diets, while consumption of <65% of total protein from animal‐sourced foods can lead to protein undernutrition (Wu, 2016). In this context, plant‐based protein can contribute to dietary protein requirements, particularly in circumstances or periods where access to other food sources of protein may be limited. Indeed, although yams have been primarily considered to be a starchy crop, they may also contribute to dietary protein, as has been documented for edible yams in West Africa (Asiedu & Sartie, 2010). The protein content of cultivated D. alata was analyzed and compared with the protein content of the nine edible wild yam species in this study. The mean protein content of cultivated D. alata did not differ significantly from the mean protein content of any of the wild edible species analyzed (tubers harvested 2018–2019). In tubers harvested in 2019–2020, the protein content of D. alata was significantly higher than all wild edible species analyzed, except for D. irodensis, D. sambiranensis subsp. bardotiae, and D. bako. These results indicate that none of the wild edible yams analyzed in this study are a better source of protein in comparison with cultivated D. alata, but certain species may provide comparable protein levels to those provided by D. alata. However, it is also recognized that the amino acid profiles of the protein occurring in D. alata and the wild edible yams require analysis to further evaluate their contribution as plant‐based protein sources for dietary needs.
The observed protein content of D. alata and all edible wild yam species analyzed in this study (range for mean values: 2.9%–8.5% DW determined using two different protein analysis methods; Table 2) compares favorably with 2.4%–7.8% protein (DW) reported for different cultivars of sweet potato (Kourouma et al., 2020; though other studies report a lower range: 0.6%–2.5% DW; Neela & Fanta, 2019), and is higher than protein reported for cassava (1%–3% DW; Ferraro et al., 2016; Shewry, 2003). Cultivated D. alata and wild edible yams in Madagascar may therefore be a richer protein source compared to some other tuber crops such as cassava.
Starch forms the bulk of energy provided by edible Dioscorea species. For example, D. esculenta (Lour.) Burkill tubers have been shown to contain similar levels of starch (78% DW) compared to D. alata (65%–84% starch, tubers not sourced from Madagascar; Wilkin et al., 2022). In this study, the mean starch content of cultivated D. alata across four harvest seasons did not differ significantly from the mean starch content of the wild edible Dioscorea species analyzed. The edible Dioscorea species were also analyzed over six harvest seasons for total carbohydrate content, which ranged from 84.9 g/100 g (mean, DW) in cultivated D. alata to 92.9 g/100 g (mean, DW) in D. sambiranensis subsp. sambiranensis. Total carbohydrate in D. alata was significantly lower than in Dioscorea sp. (Ovy valiha), D. pteropoda, D. sambiranensis subsp. sambiranensis, D. seriflora, D. orangeana, and D. maciba. These results indicate that D. alata has comparable starch levels to the wild edible Dioscorea species analyzed, while certain wild edible species may provide a higher total carbohydrate content (which represents starch and individual sugars) compared to D. alata. These results compare with 80–90 g/100 g DW carbohydrate, of which 80 g/100 g is starch, for cassava root (Ferraro et al., 2016); and 51.9–69.2 g/100 g DW starch content which has been reported for different cultivars of sweet potato (Kourouma et al., 2020), while the total carbohydrate for different varieties of sweet potato is reported as 81.3–87.3 g/100 g DW (Cartier et al., 2017). Thus, D. alata and the wild edible yams analyzed in this study have a comparable total carbohydrate and starch content to the tuber crops sweet potato and cassava.
Yams are a major source of calories for millions of people in tropical and subtropical regions (Asiedu & Sartie, 2010). In this study, the calculated energy for D. alata was significantly lower than that calculated for D. pteropoda, D. sambiranensis subsp. sambiranensis, and D. seriflora. However, the energy levels observed for D. alata and the wild edible Dioscorea species analyzed are comparable to energy values for orange‐fleshed sweet potato (345 kcal/100 g DW; Neela & Fanta, 2019). Furthermore, the total lipid content across all of the edible yam species analyzed (which did not differ significantly between species) was comparable with orange‐fleshed sweet potato (0.03–0.9 g/100 g DW; Neela & Fanta, 2019) and with other cultivars of sweet potato (0.24–1.11 g/100 g DW; Kourouma et al., 2020). However, there are some reports of lipid content of sweet potato being as high as 2.2 g/100 g DW (Cartier et al., 2017).
Vitamin A is a fat‐soluble vitamin that occurs in the diet in various forms (e.g., as retinol isomers in animal products), depending on the source. It is essential for growth, development, and maintenance of epithelial tissue and for vision, while a deficiency can result in ocular complications (e.g., xeropthalmia which may progress to blindness; Martindale, 2022). Certain edible plants, including various fruits and vegetables, are a source of provitamin A carotenoids, which have vitamin A activity when consumed, with β‐carotene being the most important (McCance & Widdowson, 2015). To understand the contribution of the nine wild edible yams and the cultivated D. alata from Madagascar on dietary vitamin A intake, they were assessed for their carotenoid content over eight harvest seasons, with results expressed as β‐carotene equivalents (DW). The β‐carotene content of D. irodensis was consistently higher than the β‐carotene content of all other species analyzed in all harvest seasons and the average (mean) β‐carotene content of D. irodensis across all harvest seasons was significantly higher than the mean β‐carotene content of all other species analyzed, including the cultivated D. alata.
To assess the distribution of β‐carotene in yam tuber parenchyma (which is consumed by people) and periderm (which is not consumed by people but may be used as livestock feed), the two species with the highest mean β‐carotene content (D. irodensis and D. pteropoda) and D. alata were subjected to additional analysis for β‐carotene content in the parenchyma and periderm tissue. β‐Carotene content was not significantly different between parenchyma and periderm tissue for each species analyzed (Figure 2), indicating that peeling the tubers of these species to remove the periderm would not adversely affect the β‐carotene content of the tubers (w/w), and that the inclusion of periderm tissue in livestock feed could provide a source of β‐carotene (albeit at low levels). Consistent with the whole tuber β‐carotene content (Figure 1), the mean β‐carotene content of the separate parenchyma and periderm from D. irodensis was also significantly higher than that in D. alata.
Food tables indicate that yam contains only trace amounts of carotenoids (McCance & Widdowson, 2015). Our data showing β‐carotene equivalents in the low μg range are consistent with food table values and contrast with 416 μg/100 g to 20,811 μg/100 g DW β‐carotene reported for different cultivars of sweet potato (Kourouma et al., 2020). Although the β‐carotene content of D. irodensis was higher than all other edible yam species analyzed in this study, this value is still at least 25‐fold less than that observed for certain sweet potato varieties (Kourouma et al., 2020). Therefore, considering the low β‐carotene content compared to certain other edible plants such as sweet potatoes, consumption of cultivated D. alata, or the wild edible yam species investigated in this study is unlikely to provide a meaningful contribution to dietary vitamin A intake or to prevent vitamin A deficiency. It should also be considered, however, that carotenoids may have functions beyond their role in nutrition as vitamin A precursors. Dietary intake of carotenoids has been associated with a reduced risk of certain chronic diseases and carotenoids show mechanistic effects (e.g., antioxidant) relevant to human health (Bohn et al., 2021; Monjotin et al., 2022). Therefore, although the β‐carotene contents of the wild and cultivated yams from Madagascar are unlikely to significantly contribute to dietary vitamin A requirements, the other functions of carotenoids, and those of other phytochemical constituents, merit further consideration for other potential health benefits.
The total ash content, representing the inorganic (crude mineral) content determined for the edible yam species analyzed in this study ranged from 3.0 to 6.0 g/100 g (mean, DW), which compares similarly with that observed for different sweet potato cultivars (2.0–4.5 g/100 g DW; Kourouma et al., 2020; Cartier et al., 2017). The wild edible yam species and cultivated D. alata were also analyzed for calcium, iron, and zinc contents over four harvest seasons. Although there were no statistical differences in mean mineral content across these four harvest seasons, the mean calcium contents of D. buckleyana, D. irodensis, and D. sambiranensis subsp. bardotiae were 2.8–3 times greater than D. alata (Figure 3a). For iron, the content in D. irodensis was 5 times higher than in D. alata (Figure 3b). Analysis showed that mean zinc content was similar across all yam species (Figure 3c).
All mineral values were substantially higher than those reported for raw yam tubers in food tables (McCance & Widdowson, 2015), which may indicate that a significant proportion of the mineral content lies in the periderm. The difference in mineral content observed for different collections of the same species could be explained by different environmental conditions, that is, soil fertility and climate of the region, which may be associated with the origin or time of harvest (Huang et al., 2007). Many studies on wild edible plant species show that they contain a considerable percentage of minerals, ranging from 2.48% to 6.36% (Afiukwa et al., 2013; Oko & Famurewa, 2015; Shajeela et al., 2011), which is comparable to the observations in our study. These findings, therefore, further support other research that shows reduced access to wild foods can negatively affect food security and nutrition, specifically micronutrient consumption (Moore et al., 2022).
Different varieties of orange‐fleshed sweet potato contain calcium in the range 219.8–273.5 mg/kg DW, iron in the range 9.1–14.0 mg/kg DW, and zinc in the range 28.5–42.4 mg/kg DW (Alam et al., 2020), while other varieties improved for disease tolerance are reported to contain calcium, iron, and zinc in the ranges 207.0–250.9, 50.9–101.8, and 21.8–31.8 mg/kg DW, respectively (Mitiku & Teka, 2017). The mineral content of the tubers of different cassava (yellow‐fleshed) genotypes is reported as 1149.7–1444.7 mg/kg DW for calcium, 8.5–9.5 mg/kg DW for iron, and 8.7–9.7 mg/kg DW for zinc (Alamu et al., 2020). Other cassava genotypes are reported to contain 1360–3690 mg/kg DW calcium, 290–400 mg/kg DW iron, and 130–190 mg/kg DW zinc (Charles et al., 2005). Thus, the calcium contents of wild yams and cultivated D. alata observed in this study are higher than calcium levels reported for certain varieties of sweet potato, and within a similar range to those reported for certain genotypes of cassava. The iron and zinc contents observed for wild yams and cultivated D. alata were within the content range of these minerals that are reported to occur in sweet potato and cassava tubers, depending on the variety or genotype of these tuber crops. An important caveat is that the reported mineral values represent the content of the raw plant tissue. To date there have been no reports of the bioavailability of these minerals (i.e., the fraction that can be absorbed in the intestine and utilized in the body) from either raw material or from meals containing cooked yam.
Knowledge of the nutrient composition of wild edible yams in Madagascar may have different implications for food security and conservation of edible species. First, the knowledge that certain wild edible yams contain comparable nutrient contents (e.g., protein, starch) to the cultivated D. alata could potentially enhance the demand for wild edible yams, particularly when cultivated crops (D. alata and others) may be in limited supply. Second, although wild edible yams are harvested in certain regions of Madagascar to supplement crops such as cassava and maize, especially during hunger periods (Andriamparany et al., 2014), the knowledge that wild edible yams may contain a higher protein content than other crops such as cassava (1%–3% DW; Ferraro et al., 2016; Shewry, 2003) may provide an additional incentive to harvest wild edible yams during other periods, exacerbating existing threats to their survival. Overharvesting of edible plants from the wild may have consequences for their conservation and for food security. Indeed, certain Dioscorea species are threatened and overharvesting of tubers from the wild is a contributing factor, with some species already categorized as endangered according to the IUCN Red List (Table 1). However, although wild edible species may be exploited for their useful properties, knowledge of how they may benefit people, for example, as food or medicines, may also motivate the sustainable management of natural resources (Howes et al., 2020). From this perspective, knowledge of the nutrient content of wild edible food species could provide incentives for their sustainable cultivation and use, to align with strategies for their conservation and for food security, in addition to supporting livelihoods. It is recognized, however, that other factors are to be considered when selecting edible yams to harvest from the wild, or indeed to cultivate. For example, local use of wild edible yams may depend on taste or personal preferences, local needs, availability, market prices, location, and quantities harvested (Andriamparany et al., 2014) among other complexities, as has been discussed previously (Moore et al., 2022). In this study, the wild edible yams analyzed include 6/6 (D. bako, D. buckleyana, D. maciba, D. orangeana, D. sambiranensis, and D. seriflora) of the most favored wild edible yam species in Madagascar (Wilkin et al., 2022). Here, we therefore present new nutrient composition data for the six most favored wild edible yams in Madagascar, to complement other food research studies on factors that influence food choice (e.g., taste, texture), while providing new nutrient data for popular edible yams in Madagascar, relevant to help support their sustainable use and conservation.
As a small island, Madagascar is vulnerable to the impacts of climate change on food and nutrition security due to factors such as fragile natural environments and lack of arable land, impacting on agriculture and contributing to risks including water insecurity (WHO & UN, 2022). Furthermore, climate change is likely to exacerbate the triple burden of metabolic and lifestyle risk factors for noncommunicable diseases and malnutrition (WHO & UN, 2022), in a country that already has the highest rate of chronic malnutrition in the world, with 42% of children under age 5 suffering from stunting (Moore et al., 2022). Potential strategies to ameliorate the effects of climate change on food security and dietary diversity include creating more diverse and climate‐resilient agricultural production systems in addition to improved knowledge and cultivation of naturally stress‐resistant plants, aligned with methods to maintain the genetic diversity of crops (Herrera et al., 2021; Ulian et al., 2020). Another approach to address these challenges is to promote the cultivation of nutrient‐dense indigenous vegetables, rather than more recently introduced vegetables that may not be adapted to local growing conditions or be culturally suitable; for example, nonprofit organizations in Madagascar have aided the cultivation of indigenous yams (Moore et al., 2022). Furthermore, community involvement in conservation projects has greater potential to achieve positive outcomes for both biodiversity conservation and food security (Moore et al., 2022). In this context, dissemination of information on the nutrient content of wild edible plants, such as for wild edible yams in Madagascar as presented in this study, could facilitate motivation and opportunities for the sustainable cultivation and use of endemic yams to benefit people and the environment. Finally, it is recognized that though new nutrient composition data for wild edible yams from Madagascar is a first step to understanding how these food resources may contribute to dietary nutrient requirements, this knowledge can also underpin future studies to further evaluate the role of wild edible yams in the diet, including research to assess the effects of preparation and cooking on nutrient content, and the bioavailability and bioaccessibility of nutrients.
5. CONCLUSION
This study presents new information on the nutrient composition of nine edible yams (Dioscorea species) endemic to Madagascar, presenting new data for their protein, carbohydrate/starch, mineral, β‐carotene, and lipid contents, as well as the energy they provide. The results show that though none of the wild edible yams analyzed in this study are a better source of protein compared to cultivated D. alata, all of the Dioscorea species analyzed contain higher levels of protein compared to those published for cassava. The wild edible D. irodensis contains a significantly higher β‐carotene content compared to all other edible yams analyzed, but levels are still considerably lower (at least 25‐fold) than published data for certain sweet potato varieties. This study has also revealed that of the wild edible Dioscorea species analyzed, D. buckleyana, D. irodensis, and D. sambiranensis subsp. bardotiae have a higher calcium content compared to the cultivated D. alata, while all nine wild edible yam species contain higher iron contents compared to cultivated D. alata. In conclusion, these results provide new insights into the nutrient composition of wild edible yams endemic to Madagascar which could underpin the direction of future research to further understanding of how they may contribute to dietary nutritional requirements and dietary diversity in Madagascar. Dissemination of new knowledge on the nutritional value of wild edible yams could also facilitate opportunities to incentivize their conservation, sustainable cultivation, and use to complement strategies to enhance food security in Madagascar.
AUTHOR CONTRIBUTIONS
Mirana K. Ratsimbazafy: Conceptualization (equal); data curation (equal); formal analysis (equal); investigation (equal); methodology (equal); writing – review and editing (equal). Paul A. Sharp: Formal analysis (equal); investigation (supporting); methodology (equal); resources (supporting); supervision (supporting); writing – review and editing (equal). Louisette Razanamparany: Conceptualization (equal); investigation (equal); methodology (equal); resources (equal); supervision (equal); writing – review and editing (supporting). Mamy Tiana Rajaonah: Data curation (equal); investigation (supporting); writing – review and editing (supporting). Feno Rakotoarison: Data curation (supporting); writing – review and editing (supporting). Kholoud K. Khoja: Investigation (equal); methodology (supporting); writing – review and editing (supporting). Paul Wilkin: Conceptualization (equal); funding acquisition (lead); project administration (lead); supervision (equal); writing – review and editing (equal). Melanie‐Jayne R. Howes: Conceptualization (supporting); data curation (equal); project administration (supporting); supervision (equal); writing – original draft (lead); writing – review and editing (lead).
FUNDING INFORMATION
The Darwin Initiative funded the following projects to support this research: “Sustainable yam markets for conservation and food security in Madagascar” (EIDPO049) and “Conserving Madagascar's yams through cultivation for livelihoods and food security” (22‐005).
CONFLICT OF INTEREST STATEMENT
The authors declare no conflict of interest.
Supporting information
Table S1
ACKNOWLEDGMENTS
The authors thank the Darwin Initiative for funding the following projects to support this research: “Sustainable yam markets for conservation and food security in Madagascar” (EIDPO049) and “Conserving Madagascar's yams through cultivation for livelihoods and food security” (22‐005). The authors acknowledge A. & M. Hladik and the Board of Trustees, RBG Kew, for the photo of Dioscorea bako shown in the graphical abstract.
Ratsimbazafy, M. K. , Sharp, P. A. , Razanamparany, L. , Rajaonah, M. T. , Rakotoarison, F. , Khoja, K. K. , Wilkin, P. , & Howes, M.‐J. (2024). Wild edible yams from Madagascar: New insights into nutritional composition support their use for food security and conservation. Food Science & Nutrition, 12, 280–291. 10.1002/fsn3.3757
DATA AVAILABILITY STATEMENT
Data relating to the yam tubers collected and analyzed in this study are shared in the supplementary material of this article. Other research data are not shared.
REFERENCES
- Afiukwa, C. A. , Ogah, O. , Ugwu Okechukwu, P. C. , Oguguo, J. O. , Ali, F. U. , & Ossai, E. C. (2013). Nutritional and antinutritional characterization of two wild yam species from Abakaliki, Southeast Nigeria. Research Journal of Pharmaceutical, Biological and Chemical Sciences, 4, 840–848. [Google Scholar]
- Alam, M. K. , Sams, S. , Rana, Z. H. , Akhtaruzzaman, M. , & Islam, S. N. (2020). Minerals, vitamin C, and effect of thermal processing on carotenoids composition in nine varieties orange‐fleshed sweet potato (Ipomoea batatas L.). Journal of Food Composition and Analysis, 92, 103582. 10.1016/j.jfca.2020.103582 [DOI] [Google Scholar]
- Alamu, E. O. , Maziya‐Dixon, B. , Sibeso, C. , Parkes, E. , & Dixon, A. G. (2020). Variations of macro‐ and microelements in yellow‐fleshed cassava (Manihot esculenta Crantz) genotypes as a function of storage root portion, harvesting time, and sampling method. Applied Sciences, 10, 5396. 10.3390/app10165396 [DOI] [Google Scholar]
- Andriamparany, J. N. , Brinkmann, K. , Jeannoda, V. , & Buerkert, A. (2014). Effects of socio‐economic household characteristics on traditional knowledge and usage of wild yams and medicinal plants in the Mahafaly region of South‐Western Madagascar. Journal of Ethnobiology and Ethnomedicine, 10, 82. 10.1186/1746-4269-10-82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- AOAC . (2000). Official methods of analysis of AOAC International (17th ed.). USA Association of Analytical Communities. [Google Scholar]
- Asiedu, R. , & Sartie, A. (2010). Crops that feed the world 1. Yams. Food Security, 2, 305–315. 10.1007/s12571-010-0085-0 [DOI] [Google Scholar]
- Atwater, W. O. , & Woods, C. D. (1896). The chemical composition of American food materials. US Official Experiment Stations, Experiment Station Bulletin No. 28. [Google Scholar]
- Bohn, T. , Bonet, M. L. , Borel, P. , Keijer, J. , Landrier, J.‐F. , Milisav, I. , Ribot, J. , Riso, P. , Winklhofer‐Roob, B. , Sharoni, Y. , Corte‐Real, J. , Van Helden, Y. , Loizzo, M. R. , Poljšak, B. , Porrini, M. , Roob, J. , Trebše, P. , Tundis, R. , Wawrzyniak, A. , … Duliñska‐Litewka, J. (2021). Mechanistic aspects of carotenoid health benefits – where are we now? Nutrition Research Reviews, 34, 276–302. 10.1017/S0954422421000147 [DOI] [PubMed] [Google Scholar]
- Bóna‐Lovász, J. , Bóna, A. , Ederer, M. , Sawodny, O. , & Ghosh, R. (2013). A rapid method for the extraction and analysis of carotenoids and other hydrophobic substances suitable for systems biology studies with photosynthetic bacteria. Metabolites, 3, 912–930. 10.3390/metabo3040912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradstreet, R. S. (1965). The Kjeldahl method for organic nitrogen. Academic Press. [Google Scholar]
- Cartier, A. , Woods, J. , Sismour, E. , Allen, J. , Ford, E. , Githinji, L. , & Xu, Y. (2017). Physiochemical, nutritional and antioxidant properties of fourteen Virginia‐grown sweet potato varieties. Journal of Food Measurement and Characterization, 11, 1333–1341. 10.1007/s11694-017-9511-8 [DOI] [Google Scholar]
- Charles, A. L. , Sriroth, K. , & Huang, T.‐C. (2005). Proximate composition, mineral contents, hydrogen cyanide and phytic acid of 5 cassava genotypes. Food Chemistry, 92, 615–620. 10.1016/j.foodchem.2004.08.024 [DOI] [Google Scholar]
- Englyst, H. N. , & Cummings, J. H. (1988). Improved method for measurement of dietary fiber as non‐starch polysaccharides in plant foods. Journal – Association of Official Analytical Chemists, 71, 808–814. 10.1093/jaoac/71.4.808 [DOI] [PubMed] [Google Scholar]
- FAO . (1998). Carbohydrates in human nutrition. Report of a Joint FAO/WHO Expert Consultation. FAO Food and Nutrition Paper No. 66. [PubMed] [Google Scholar]
- Ferraro, V. , Piccirillo, C. , Tomlins, K. , & Pintado, M. E. (2016). Cassava (Manihot esculenta Crantz) and yam (Dioscorea spp.) crops and their derived foodstuffs: Safety, security and nutritional value. Critical Reviews in Food Science and Nutrition, 56, 2714–2727. 10.1080/10408398.2014.922045 [DOI] [PubMed] [Google Scholar]
- Folch, J. , Lees, M. , & Sloane, G. H. (1957). A simple method for the isolation and purification of total lipides from animal tissues. Journal of Biological Chemistry, 226, 497–509. 10.1016/S0021-9258(18)64849-5 [DOI] [PubMed] [Google Scholar]
- GNR: Global Nutrition Report . (2022). https://globalnutritionreport.org/resources/nutrition‐profiles/africa/eastern‐africa/madagascar/
- Herrera, J. P. , Rabezara, J. Y. , Ravelomanantsoa, N. A. F. , Metz, M. , France, C. , Owens, A. , Pender, M. , Nunn, C. L. , & Kramer, R. A. (2021). Food insecurity related to agricultural practices and household characteristics in rural communities of Northeast Madagascar. Food Security, 13, 1393–1405. 10.1007/s12571-021-01179-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howes, M.‐J. R. , Quave, C. L. , Collemare, J. , Tatsis, E. C. , Twilley, D. , Lulekal, E. , Farlow, A. , Li, L. , Cazar, M.‐E. , Leaman, D. J. , Prescott, T. A. K. , Milliken, W. , Martin, C. , De Canha, M. N. , Lall, N. , Qin, H. , Walker, B. E. , Vásquez‐Londoño, C. , Allkin, B. , … Nic Lughadha, E. (2020). Molecules from nature: Reconciling biodiversity conservation and global healthcare imperatives for sustainable use of medicinal plants and fungi. Plants, People, Planet, 2, 463–481. 10.1002/ppp3.10138 [DOI] [Google Scholar]
- Huang, C.‐C. , Chiang, P.‐Y. , Chen, Y.‐Y. , & Wang, C.‐C. R. (2007). Chemical compositions and enzyme activity changes occurring in yam (Dioscorea alata L.) tubers during growth. LWT – Food Science & Technology, 40, 1498–1506. 10.1016/j.lwt.2006.12.006 [DOI] [Google Scholar]
- IUCN: International Union for Conservation of Nature . (2022, October 31). Red List of Threatened Species. Version 2022‐1. https://www.iucnredlist.org
- Jeannoda, V. H. , Razanamparany, J. , Rajaonah, M. T. , Monneuse, M. O. , Hladik, A. , & Hladik, C. M. (2007). Les ignames (Dioscorea spp.) endémiques et cultivées de Madagascar: diversité, perception, valeur nutritionnelle et systèmes de gestion. Revue d'Ecologie, Terre et Vie, 62, 191–207. https://hal.archives‐ouvertes.fr/hal‐00546221 [Google Scholar]
- Kourouma, V. , Mu, T.‐H. , Zhang, M. , & Sun, H.‐N. (2020). Comparative study on chemical composition, polyphenols, flavonoids, carotenoids and antioxidant activities of various cultivars of sweet potato. International Journal of Food Science and Technology, 55, 369–378. 10.1111/ijfs.14336 [DOI] [Google Scholar]
- Mæhre, H. K. , Dalheim, L. , Edvinsen, G. K. , Elvevoll, E. O. , & Jensen, I.‐J. (2018). Protein determination – Method matters. Food, 7, 5. 10.3390/foods7010005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martindale: The Complete Drug Reference . (2022, May 20 & November 24). Medicines Complete. Pharmaceutical Press. www.medicinescomplete.com [Google Scholar]
- McCance, R. A. , & Widdowson, E. M. (2015). McCance & Widdowson's The Composition of Foods (7th Summary Edition). Royal Society of Chemistry. [Google Scholar]
- McCleary, B. V. , Gibson, T. S. , & Mugford, D. C. (1997). Measurement of total starch in cereal products by amyloglucosidase‐α‐amylase method: Collaborative study. Journal of AOAC International, 80, 571–579. 10.1093/jaoac/80.3.571 [DOI] [Google Scholar]
- McCleary, B. V. , Gibson, T. S. , Solah, V. , & Mugford, D. C. (1994). Total starch measurement in cereal products: Interlaboratory evaluation of a rapid enzymic test procedure. Cereal Chemistry, 71, 501–505. [Google Scholar]
- Miller, D. S. , & Payne, P. R. (1959). A ballistic bomb calorimeter. British Journal of Nutrition, 13, 501–508. 10.1079/BJN19590064 [DOI] [PubMed] [Google Scholar]
- Mitiku, D. H. , & Teka, T. A. (2017). Nutrient and antinutrient composition of improved sweet potato [Ipomea batatas (L) Lam] varieties grown in eastern Ethiopia. Nutrition and Food Science, 47, 369–380. 10.1108/NFS-07-2016-0098 [DOI] [Google Scholar]
- Monjotin, N. , Amiot, M. J. , Fleurentin, J. , Morel, J. M. , & Raynal, S. (2022). Clinical evidence of the benefits of phytonutrients in human healthcare. Nutrients, 14, 1712. 10.3390/nu14091712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore, M. , Alpaugh, M. , Razafindrina, K. , Trubek, A. B. , & Niles, M. T. (2022). Finding food in the hunger season: A mixed methods approach to understanding wild plant foods in relation to food security and dietary diversity in southeastern Madagascar. Frontiers in Sustainable Food Systems, 6, 929308. 10.3389/fsufs.2022.1929308 [DOI] [Google Scholar]
- Neela, S. , & Fanta, S. W. (2019). Review on nutritional composition of orange‐fleshed sweet potato and its role in management of vitamin a deficiency. Food Science and Nutrition, 7, 1920–1945. 10.1002/fsn3.1063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oko, A. , & Famurewa, A. (2015). Estimation of nutritional and starch characteristics of Dioscorea alata (water yam) varieties commonly cultivated in the south‐eastern Nigeria. British Journal of Applied Science & Technology, 6, 145–152. 10.9734/BJAST/2015/14096 [DOI] [Google Scholar]
- Rinaldo, D. (2020). Carbohydrate and bioactive compounds composition of starchy tropical fruits and tubers, in relation to pre and postharvest conditions: A review. Journal of Food Science., 85, 249–259. 10.1111/1750-3841.15002 [DOI] [PubMed] [Google Scholar]
- Shajeela, P. S. , Mohan, V. R. , Jesudas, L. L. , & Soris, P. T. (2011). Nutritional and antinutritional evaluation of wild yam (Dioscorea spp.). Tropical and subtropical agroecosystems, 14, 723–730. [Google Scholar]
- Shewry, P. R. (2003). Tuber storage proteins. Annals of Botany, 91, 755–769. 10.1093/aob/mcg084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulian, T. , Diazgranados, M. , Pironon, S. , Padulosi, S. , Liu, U. , Davies, L. , Howes, M.‐J. R. , Borrell, J. S. , Ondo, I. , Pérez‐Escobar, O. A. , Sharrock, S. , Ryan, P. , Hunter, D. , Lee, M. A. , Barstow, C. , Łuczaj, Ł. , Pieroni, A. , Cámara‐Leret, R. , Noorani, A. , … Mattana, E. (2020). Unlocking plant resources to support food security and promote sustainable agriculture. Plants, People, Planet, 2, 421–445. 10.1002/ppp3.10145 [DOI] [Google Scholar]
- Wanasundera, J. P. D. , & Ravindran, G. (1994). Nutritional assessment of yam (Dioscorea alata) tubers. Plant Foods for Human Nutrition, 46, 33–39. 10.1007/BF01088459 [DOI] [PubMed] [Google Scholar]
- WHO (World Health Organization) and UN (United Nations Framework Convention on Climate Change) . (2022). Madagascar Health & Climate Change Country Profile 2021. https://www.who.int/publications/i/item/WHO‐HEP‐ECH‐CCH‐21.01.08
- Wilkin, P. , Bennett, B. , Cameron, S. , Howes, M.‐J. R. , Jeannoda, V. , Rajaonah, M. T. , Rakotoarison, F. , Razanamparany, L. , & Viruel, J. (2022). Dioscoreaceae. In Goodman S. M. (Ed.), The new natural history of Madagascar (pp. 545–550). Princeton University Press. [Google Scholar]
- Wilkin, P. , Hladik, A. , Jeannoda, V. , & Weber, O. (2009). The threatened edible yams of the Dioscorea sambiranensis R. Knuth species complex (Dioscoreaceae): A new species and subspecies. Adansonia, 31, 249–266. 10.5252/a2009n2a2 [DOI] [Google Scholar]
- Wu, G. (2016). Dietary protein intake and human health. Food & Function., 7, 1251–1265. 10.1039/C5FO01530H [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1
Data Availability Statement
Data relating to the yam tubers collected and analyzed in this study are shared in the supplementary material of this article. Other research data are not shared.