

Abstract
Background
Therapeutic strategies based on scavenging reactive oxygen species (ROS) and suppressing inflammatory cascades are effective in improving functional recovery after spinal cord injury (SCI). However, the lack of targeting nanoparticles (NPs) with powerful antioxidant and anti-inflammatory properties hampers the clinical translation of these strategies. Here, CD44-targeting hyaluronic acid-selenium (HA-Se) NPs were designed and prepared for scavenging ROS and suppressing inflammatory responses in the injured spinal cord, enhancing functional recovery.
Results
The HA-Se NPs were easily prepared through direct reduction of seleninic acid in the presence of HA. The obtained HA-Se NPs exhibited a remarkable capacity to eliminate free radicals and CD44 receptor-facilitated internalization by astrocytes. Moreover, the HA-Se NPs effectively mitigated the secretion of proinflammatory cytokines (such as IL-1β, TNF-α, and IL-6) by microglia cells (BV2) upon lipopolysaccharide-induced inflammation. In vivo experiments confirmed that HA-Se NPs could effectively accumulate within the lesion site through CD44 targeting. As a result, HA-Se NPs demonstrated superior protection of axons and neurons within the injury site, leading to enhanced functional recovery in a rat model of SCI.
Conclusions
These results highlight the potential of CD44-targeting HA-Se NPs for SCI treatment.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12951-024-02302-0.
Keywords: CD44 targeting, Inflammation, Reactive oxygen species, Selenium nanoparticles, Spinal cord injury
Introduction
Spinal cord injury (SCI) is a catastrophic condition that results in severe disability, including paraplegia or quadriplegia, and gives rise to a range of serious multi-system complications, such as recurrent bladder and kidney infections, intestinal problems, as well as cardiac and respiratory dysfunction, placing a heavy burden on individuals, families, and society [1, 2]. In clinical practice, the conventional approaches for treating SCI encompass administration of high-dose methylprednisolone, implementation of surgical interventions aimed at stabilizing and decompressing the spinal cord, and provision of rehabilitative therapy [3, 4]. However, currently available therapies are mainly palliative and do not offer substantial functional recovery [5–8]. Hence, there is an impetus to explore alternative therapeutic strategies that consider the pathophysiology of SCI. The deleterious pathophysiological events that occur following SCI are the consequence of a biphasic process that encompasses both primary and secondary impairments [9–12]. The primary damage stems from the mechanical force and the initial impairment that subsequently initiates a rapidly escalating sequence of degenerative occurrences, commonly referred to as the secondary injury [13–15]. The secondary injury comprises various pathological events, such as the generation of reactive oxygen species (ROS) and the occurrence of inflammation [16–18]. In particular, overproduction of ROS at the injury site leads to the depletion of endogenous antioxidants and the disruption of redox balance, causing extensive damage to intracellular biomacromolecules, such as DNA and proteins, followed by consequent neuronal cell death [17, 19]. Moreover, excess ROS provoke an aggravated inflammatory response, recruiting and activating inflammatory cells, such as microglia and astrocytes, which further exacerbates neuronal cell death [14, 17]. Hence, it can be argued that ROS significantly contribute to the occurrence and progression of secondary injury in SCI.
It is well-documented that timely therapeutic intervention through ROS scavenging and inflammation suppression at the initial phase of SCI can significantly improve neurological and functional recuperation [14, 18, 20–23]. Recently, the utilization of nano-biomaterials as a therapeutic approach for SCI has garnered considerable interest [5, 15, 24–26]. Among them, selenium (Se)-based nanoparticles (NPs) have emerged as promising candidates due to their capacity for scavenging ROS and mitigating inflammation [27–29]. It has been proven that Se supplementation can forestall ROS accumulation, suppress lipid peroxidation, and prevent cellular damage [30–33]. Moreover, Se has been shown to mitigate functional deficits in the central nervous system due to its antioxidation properties [15, 34, 35]. Notwithstanding, conventional Se NPs have drawbacks, such as limited targeting ability, substandard stability, and limited solubility, which impede their application in basic research and the clinic [36]. It is therefore imperative to formulate innovative approaches to ensure the accumulation of Se NPs at the lesion site, favorable biological effects, and safety for therapeutic applications.
Hyaluronic acid (HA), a polysaccharide originating from the natural extracellular matrix, serves as a ligand for cluster of differentiation 44 (CD44) receptor [37]. Furthermore, it has been demonstrated that HA is beneficial for repairing spinal cord tissue by preventing the formation of glial scars [38]. In the current work, we focused the engineering and fabrication of HA-stabilized selenium nanoparticles (HA-Se NPs) specifically targeting the overexpressed CD44 receptor within the injured spinal cord, with the aim of mitigating secondary injury (Scheme 1). The HA-Se NPs were prepared via reduction of seleninic acid in the presence of sodium ascorbate. The obtained HA-Se NPs exhibited an exceptional ROS scavenging ability. Moreover, in vivo assessments confirmed that the HA-Se NPs tended to accumulate at the injury site through active binding to CD44, which is upregulated in activated astrocytes. By virtue of these targeting and ROS-scavenging abilities, the HA-Se NPs achieved superior neuroprotection and enhanced functional recovery in a rat model of SCI.
Scheme 1.

Schematic illustration of the preparation of CD44-targeting HA-Se NPs for promoting functional recovery after SCI
Materials and methods
High-molecular-weight HA (with a molecular weight of 2 × 105 g mol−1) and seleninic acid were procured from Huaxia Chemical Reagent Co., Ltd. (Chengdu, China). Sodium ascorbate was obtained from Aladdin Bio-Chem Technology Co., Ltd. (Shanghai, China). Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum were purchased from Gibco (Thermo Fisher Scientific, Waltham, MA, USA). 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA; D6883), and 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Hydrogen peroxide solution (30 wt% in water) was obtained from Sinopharm Chemical Reagent Co., Ltd. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) was purchased from Shanghai Macklin Biochemical Co., Ltd. Anti-GFAP, anti-NF200, and anti-NeuN primary antibodies were obtained from Abcam (Cambridge, UK). Anti-CD44 and anti-caspase-3 antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). The dialysis membrane was obtained from Greenbird Technology Co., Ltd (Shanghai, China). The remaining compounds were obtained from commercial providers and used without any further manipulation.
Preparation and characterization of HA-Se NPs
Briefly, 45 mL of aqueous HA solution (1.5 mg/mL) was mixed with 600 µL of seleninic acid (0.1 M) in a round-bottom flask. Subsequently, 3 mL of ascorbic acid aqueous solution (0.1 M) was gradually introduced into the mixture with constant stirring at ambient temperature (~ 22 °C). The gradual addition of sodium ascorbate solution resulted in a noticeable alteration of the solution’s color from a clear hue to a red tint. Upon completion of the reaction, the mixture was subjected to dialysis for 48 h using ultrapure water and a dialysis bag with a molecular weight cutoff (MWCO) of 300 kDa. Thereafter, the solution was centrifuged at 4,000 rpm for 20 min. Finally, the HA-Se NPs were obtained via lyophilization as red powder (yield: 82.5%).
The size and zeta potential of HA-Se nanoparticles were evaluated via dynamic light scattering on a Malvern Zetasizer Nano ZS (ZEN3600, Malvern Instruments, Worcestershire, UK). The morphology of HA-Se NPs was observed through transmission electron microscopy (TEM) (JOEL-1011, Tokyo, Japan) at an accelerating voltage of 200 kV and emission-scanning electron microscopy (SEM) (Zeiss, Oberkochen, Germany). Using the KBr pellet method, Fourier transform infrared spectra (FT IR) were recorded with a Win-IR device (Bio-Rad Laboratories, Hercules, CA, USA). The amount of selenium in HA-Se NPs was determined using an inductively coupled plasma mass spectrometer (ICP-MS, Xseries II, Thermo Scientific, USA). The antioxidant capacity of HA-Se NPs was explored by analyzing their influence on DPPH free radicals. In the control group, a combination of water (2 mL) and DPPH anhydrous ethanol solution (0.4 mM, 2 mL) was utilized. The experimental group, on the other hand, included HA-Se NPs (250/1, 000 µg/mL, 2 mL) and DPPH anhydrous ethanol solution (0.4 mM, 2 mL), while the blank group included HA-Se NPs (250/1,000 µg/mL, 2 mL) and absolute ethanol (2 mL). The background was modified through the utilization of a mixture of 2 mL of water and 2 mL of anhydrous ethanol. The aforementioned groups were exposed to darkness for a period of 30 min, after which the absorbance at 517 nm was recorded on a Bio-Rad 680 microplate reader (TECAN Trading AG, Switzerland). The free radical-scavenging rate (%) was determined as per the following formula: (1−(Aexperimental − Ablank)/Acontrol) ×100, where Aexperimental, Ablank, and Acontrol represent the absorbance values of the experimental, blank, and control group, respectively.
Cytotoxicity and neuroprotective activity assay in vitro
Astrocytes, pheochromocytoma 12 (PC12) cells, and the BV2 microglia cell line were procured from the Cell Bank of the Chinese Academy of Science (Shanghai, China). The MTT method was employed to evaluate the cytotoxicity of HA-Se NPs in astrocytes and PC12 cells [22]. In brief, astrocytes or PC12 cells were seeded onto 96-well plates at a concentration of 8,000 cells per well and incubated for 12 h in DMEM medium. Subsequently, different concentrations of HA-Se NPs were added to the wells, ranging from 3.125 to 100 µg/mL. Control group cells were subjected to treatment with phosphate-buffered saline (PBS). Three parallel tests were carried out for every concentration. After a further incubation for either 24 or 48 h, solution containing 5 mg/mL MTT was added to each well at a volume of 20 µL and allowed to incubate for a further 4 h. Thereafter, the medium was discarded and exchanged with 150 µL of dimethyl sulfoxide. The absorbance of each well at 492 nm was determined using an absorbance microplate reader (Infinite M200, Tecan, Switzerland). In an effort to assess the potential protective properties of HA-Se NPs against H2O2-induced oxidative stress [22], astrocytes were grown in 96-well plates, with each well containing 8,000 cells. Prior to H2O2 exposure, the cells underwent 30-min pre-treatment with either PBS or HA-Se NPs at 50 and 100 µg/mL. Thereafter, the cells were exposed to 100 µM H2O2 for 24 h, and their viability was assessed via MTT assay. To further evaluate cell viability, we employed a commercial live-dead cell staining kit from Sigma-Aldrich. After staining, cells were observed through a confocal laser-scanning microscope (CLSM) (LSM 780, Zeiss). Dead cells were then quantified using the ImageJ software (1.51k, NIH, Bethesda, MD, USA). DCFH-DA fluorescence intensity was measured to quantitatively determine ROS levels in both astrocytes and PC12 cells.
Assessment of proinflammatory cytokine levels in vitro and in vivo
In order to examine the capacity of HA-Se NPs to inhibit inflammation, BV2 cells were first seeded onto 6-well plates at a density of 2 × 105 cells/well and permitted to settle for 12 h. Cells were then subjected to pre-treatment with either PBS or HA-Se NPs (20 µg/mL) for 2 h. Thereafter, the cells were exposed to 1 µg/mL lipopolysaccharide (LPS) for 24 h. To determine the levels of interleukin (IL)-1β, tumor necrosis factor α (TNF-α), and IL-6, we employed commercially available enzyme-linked immunosorbent assay kits (Anoric Bio-technology Co., Ltd) and an absorbance microplate reader (Infinite M200, Tecan, Switzerland). The assays were performed in triplicate. To confirm the in vivo suppressive effects of HA-Se NPs on inflammatory cells, which typically exhibit a peak at the lesion site within 3–7 days post-injury, immunofluorescence analysis of spinal cord tissue samples was carried out using anti-CD68 and anti-Iba-1 antibodies.
Animal model of SCI
Approval for animal procedures was granted by the Animal Ethics Committee of Jilin University (approval No. SY202103013). Female Sprague-Dawley rats, aged 6–8 weeks, were procured from Liaoning Changsheng Biotechnology Ltd. and provided with ad libitum access to food and water. Sprague-Dawley rats were anesthetized via intraperitoneal administration of pentobarbital sodium intraperitoneally at a dose of 5 mg per kilogram of body weight. After disinfecting with iodine volts, a T10 laminectomy procedure was performed to reveal the spinal cord, and the injury model was established using a weight-drop device (C4P01-001, Shenzhen, China). A 40-g rod was allowed to fall from a height of 50 mm, making contact with the spinal cord’s uncovered dorsal surface, and causing an initial depth of penetration of 2.5 mm. Following the surgical intervention, the fascia, muscle, and skin were sequentially closed in layers. The Sprague-Dawley rats were housed in a temperature-regulated setting at 25 ± 2 °C and were provided with ad libitum access to both water and food. Moreover, the Sprague-Dawley rats were treated with cefazolin, given twice daily for a duration of 5 days at a dose of 25 mg/kg. The urinary bladders of Sprague-Dawley rats were manually emptied twice daily until they regained the ability to urinate autonomously.
The SCI rats were randomly divided into four groups (n = 9). The rats were treated with saline, HA-Se NPs (1 mg kg−1), HA-Se NPs (5 mg kg−1), or HA-Se NPs (10 mg kg−1). All rats received intravenous injections. The animals underwent anesthesia and were then perfused with 4% paraformaldehyde and PBS at the 12-week interval post-injury. After being harvested from the center of injury, the spinal cords, which had a length of roughly 2 cm, were treated with a 4% paraformaldehyde solution. Thereafter, the specimens were embedded in paraffin for further analysis. These sections were employed in subsequent experiments.
Analysis of CD44 expression
CD44 expression in the spinal cord was evaluated seven days after injury. The tissue slices underwent dual labeling using a primary anti-CD44 antibody in conjunction with primary anti-glial fibrillary acidic protein (GFAP), anti-NeuN, anti-CD68, or anti-Iba-1 antibodies. For immunofluorescence staining, the antibody utilized is documented in Additional file 1: Table S1 . CLSM was utilized to obtain images. Immunofluorescence experiments were performed three times. To examine the potential for inflammation to stimulate CD44 expression in astrocytes, the cells were grown on 6-well plates and subjected to varying concentrations of LPS (10, 20, or 40 µg/mL) or glutamate (100, 200, or 400 µM) for a duration of 24 h. CD44 expression was evaluated via western blot and immunofluorescence analyses. Following LPS activation for 24 h, astrocytes were rinsed with PBS and were then fixed with 4% PFA. Nonspecific binding sites were obstructed using blocking buffer for a period of 1.5 h. Thereafter, the cells were incubated with an anti-CD44 primary antibody and incubated overnight at a temperature of 4 °C. After being washed with PBS, the cells were exposed to secondary antibodies and incubated for a duration of 2 h at room temperature. Another PBS wash was then carried out, and the cells were subsequently exposed to 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (Dil), a fluorescent probe for cell membranes, for a duration of 15 min. Ultimately, the astrocytes were examined and visualized via CLSM. This experiment was also repeated three times.
To ascertain the potential association between the CD44 receptor and the absorption of HA-Se NPs, a competitive cellular uptake experiment was performed. Following a 24-h incubation with LPS, astrocytes were subjected to a 4-h pretreatment with either HA (1 mg/mL) or PBS. Samples were then subjected to incubation with 0.1 mg/mL HA-Se NPs for 4 and 8 h, followed by five washes with PBS and fixation with 4% PFA for a period of 20 min. Cell nuclei were stained with DAPI staining prior to observation via confocal microscopy (LSM 780, Zeiss, Germany). Mean intracellular fluorescence intensity was determined using ImageJ (1.51k, NIH, Bethesda, MD, USA).
Biodistribution of HA-Se NPs
NH2-Cy5-labeled HA-Se NPs were synthesized as previously described [39]. To visualize the in vivo biodistribution of HA-Se NPs, rats were administered Cy5-labeled HA-Se NPs at a dose of 10 mg/kg subsequent to the injury. At specific intervals (4, 12, and 24 h), the rats were humanely sacrificed, and their primary organs (heart, lung, liver, spleen, brain, kidney, and spinal cord) were obtained. Control samples were obtained by administering Cy5-labeled HA-Se NPs to sham rats, which were then euthanized 12 h post-injection. The obtained organs were imaged and analyzed using a Maestro In Vivo Imaging System (IVIS Lumina LT Series III, PerkinElmer, USA).
Behavioral analysis of SCI rats
The Basso, Beattie, Bresnahan (BBB) locomotor rating scale was employed to evaluate the motor functions of rats. Based on the 21-point BBB open-field grading scale, rats were individually assessed in an open field for 5 min. The BBB score assesses the voluntary movements of rat limbs through a scale of 0 to 21. A score of 0 represents paralysis, whereas 21 indicates normal movements. Evaluation was performed by two separate evaluators, and the marks were ascertained through mutual accord.
Histological and immunofluorescence analyses
Fixed spinal cord samples were embedded in paraffin, followed by sectioning into 4 μm thick slices in the coronal plane. Through H&E staining, pathological changes, which included enlargement of cavity area and inflammatory cell infiltration, were investigated. The extent of demyelination was determined by staining spinal cord slices from each group with 0.1% Luxol fast blue (LFB) at the 12-week mark following injury.
To evaluate the myelin sheath ultrastructure, we prepared tissue for transmission electron microscopy (TEM) as per our previously established method [22]. The spinal cord segments were fixed overnight at a temperature of 4 °C using 2.5% glutaraldehyde. Thereafter, the segments were sliced into 1 mm3 pieces, osmicated for 90 min, and then dehydrated for a period of 135 min. Subsequently, the ultrathin sections were subjected to staining with uranyl acetate and lead citrate, followed by a thorough examination using TEM. To assess the expression of neurofilament 200 (NF200) and NeuN, the slices were first subjected to permeation with a PBS solution that contained 0.1% Triton X-100 for a period of 15 min. Thereafter, a 2-h incubation period with 5% bovine serum albumin (BSA) was utilized for blocking, followed by thorough washing with PBS containing 0.2% Tween 20. Subsequently, the portions were subjected to an overnight incubation at a temperature of 4 °C, utilizing primary antibodies against NF200 and NeuN. The axons and neurons were distinguished using anti-NF200 and anti-NeuN, respectively. Additional file 1: Table S1 provides details on antibody dilution for immunofluorescence. Images were acquired through CLSM. The potential toxicity of HA-Se NPs was assessed 12 weeks post-treatment through H&E staining of vital organs.
Western blotting
Spinal cord tissue was lysed using a buffer solution that contained proteinase inhibitors. The lysate was then centrifuged at 13,300 rpm for 15 min. Protein concentration was determined using a BCA protein kit (Beyotime, Shanghai, China). The procedure involved separation of 10 µg protein from each sample using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and subsequent transfer of the separated protein onto polyvinylidenedifluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA). For blocking, the membranes were treated with 5% bovine serum albumin for 1.5 h. Thereafter, membranes were incubated with either anti-CD44 or anti-caspase-3 antibodies at 4 °C overnight. After washing, the membranes were incubated with a secondary antibody conjugated to horseradish peroxidase for 2 h at room temperature. Target protein expression was normalized based on that of GAPDH or β-actin. Western blotting antibody data is available in Additional file 1: Table S2. An enhanced chemiluminescence Western blot detection system (AI600 Imager; GE Healthcare, Chicago, IL, USA) was used to capture protein bands. Densitometric analysis of the protein bands was carried out using Multi Gauge software (Fuji, Tokyo, Japan). Western blotting experiments were repeated three times.
Statistical analysis
To determine the statistical significance of differences, a one-way analysis of variance or t-tests were performed utilizing Prism 8.0.2 (GraphPad Software, San Diego, CA, USA). *P < 0.05 was deemed statistically significant. **P < 0.01 and ***P < 0.001 were deemed highly significant.
Results
Preparation and characterization of HA-Se NPs
The synthesis of HA-Se NPs was accomplished through a straightforward reduction reaction, utilizing water-soluble HA as a stabilizing and capping agent [40]. The HA acts as a capping agent to facilitate NP formation and also provides a protective shell to prevent NP aggregation. The HA-Se NPs exhibited a size of 101.2 ± 1.4 nm and a polydispersity index of 0.17 ± 0.02 (Fig. 1A). The particle size of HA-Se NPs did not change significantly over a period of 3 days (Additional file 1: Figure S1), which indicates a high degree of stability. Through TEM, we observed that HA-Se NPs possessed a core-shell structure, with a diameter averaging roughly 95 nm (Fig. 1B and Additional file 1: Figure S2). Moreover, SEM analysis revealed that HA-Se NPs exhibited a uniform spherical morphology, with an approximate size of 94 nm (Fig. 1C). The zeta potential of HA-Se NPs was − 26.2 ± 0.47. Their composition was investigated in greater detail using X-ray photoelectron spectroscopy, which confirmed that the nanoparticles were predominantly composed of carbon, nitrogen, oxygen, and selenium, as indicated by the distinctive peaks detected in the C 1s, N 1s, O 1s, and Se 3d spectra. (Additional file1: Figure S3). The selenium content of HA-Se NPs was 10.8%, as determined via ICP-MS. As oxidative stress can cause significant and continuous harm to the central area and adjacent spinal cord [6, 24], the capacity of HA-Se NPs to eliminate ROS was subsequently examined. HA-Se NPs exhibited remarkable ability in eliminating DPPH free radicals that was concentration- and exposure duration-dependent, as shown in Fig. 1D.
Fig. 1.

Characterization of hyaluronic acid-selenium nanoparticles (HA-Se NPs). A Dynamic light scattering measurement of HA-Se NPs. B Transmission and C scanning electron microscopy imaging of HA-Se NPs. D The efficacy of HA-Se NPs in scavenging free radicals was evaluated at varying concentrations
Biocompatibility of HA-Se NPs, protection from H2O2-induced oxidative stress, and inflammation suppression in vitro
The biocompatibility of the synthesized HA-Se NPs was investigated through MTT tests. The impact of HA-Se NPs on astrocytes and PC12 cells was evaluated through observation and analysis after 24 or 48 h of exposure at concentrations ranging from 0 to 100 mg/L. Notwithstanding the highest concentration tested, the HA-Se NPs induced no apparent cytotoxicity, as shown in Additional file 1: Figure S4. Oxidative stress occurs when excessive ROS are generated after SCI [41–43]. HA-Se NPs successfully protected astrocytes from oxidative stress induced by H2O2, preventing cell death (Figure S5). Live/dead cell staining further confirmed the protective effect of HA-Se NPs (Fig. 2A, B and Additional file 1: Figure S6A, B). Consistently, the presence of HA-Se NPs effectively reduced the amount of ROS during in vitro culture of astrocytes and PC12 cells, as indicated by the reduced fluorescence intensity of a ROS probe (Fig. 2C, D and Additional file 1: Figure S6C, D). The anti-inflammatory efficacy of HA-Se NPs was determined in BV2 cells exposed to LPS (1 µg/mL). Incubation with HA-Se NPs completely reversed the LPS-induced increase in primary cytokines (TNF-α, IL-1β, and IL-6) (Fig. 2E–G).
Fig. 2.

The efficacy of HA-Se NPs in preventing H2O2-induced oxidative stress and inflammation was evaluated. A Live/dead staining of astrocytes. Scale bar = 20 μm. The concentration of H2O2 was 100 µM. B Quantitative analysis of astrocyte cell death. *P < 0.05. C Levels of reactive oxygen species within astrocytes were determined via DCFH-DA staining. D Quantitative analysis of DCF fluorescence intensity in astrocytes. **P < 0.01. Quantification of E interleukin (IL)-1β, F IL-6, and G tumor necrosis factor (TNF)-α levels in BV2 cell culture medium in the presence or absence of HA-Se NPs. *P < 0.05
Targeting efficiency of HA-Se NPs in vitro and in vivo
Insufficient information exists regarding CD44 expression within the injured spinal cord [44]. Thus, we evaluated CD44 expression in the injured spinal cord of rats. Increases in CD44 levels were detected at 1-, 3-, 5-, and 7-days post-injury (Fig. 3A–C). To further characterize the cells which exhibited CD44 upregulation, we performed immunofluorescence staining. As indicated in Fig. 3D and E, GFAP and CD44 signals completely co-localized, while Iba-1 and CD44 exhibited partial co-localization. In addition, CD68 and CD44 fluorescence signals as well as those of NueN and CD44 exhibited incomplete colocalization (Fig. 3F and G). Taken together, these findings suggest that CD44 was expressed in astrocytes and a subset of microglial cells.
Fig. 3.
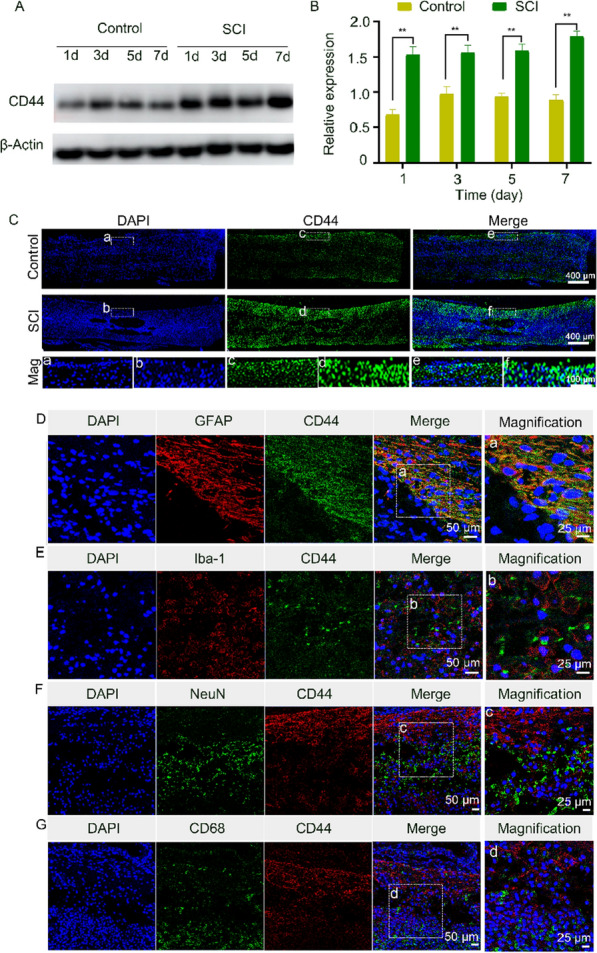
CD44 expression in the injured spinal cord. A Representative western blot images of CD44 expression in the control and SCI groups. B Densitometric analysis of CD44 levels based on data in (A). **P < 0.01 in comparison to the control group. C CD44 immunofluorescence staining of injured spinal cord samples. Mag: Magnification. D Immunofluorescence staining of injured spinal cord samples for GFAP (red) and CD44 (green). E Immunofluorescence staining of injured spinal cord samples for CD44 (green) and Iba-1 (red). F Immunofluorescence staining of injured spinal cord samples for NeuN (green) and CD44 (red). G Immunofluorescence staining of injured spinal cord samples for CD68 (green) and CD44 (red)
It has been previously established that an inflammatory microenvironment, such as that induced by LPS, promotes the secretion of proinflammatory cytokines by microglial cells and astrocytes [45–48]. Further, research has shown that is CD44 upregulated during the inflammatory response [49–51]. Thus, we exposed astrocytes to LPS for 24 h, observing notably enhanced CD44 expression in the LPS group as compared to controls, indicative of a substantial elevation in CD44 following LPS activation (Fig. 4A and Additional file 1: Figure S7). Western blotting confirmed the upregulation of CD44 in LPS-stimulated astrocytes (Additional file 1: Figure S8A, B). Conversely, the presence of glutamate did not alter CD44 expression in astrocytes, as indicated by the data presented in Additional file 1: Figure S8C and D.
Fig. 4.

CD44 immunofluorescence staining in astrocytes and internalization of Cy5-HA-Se NPs. A Astrocytes stained for CD44 and Dil. Scale bar = 50 μm. B Cy5-HA-Se NPs internalization by astrocytes after 4 and 8 h of incubation. Scale bar = 50 μm. C Targeting efficiency of HA-Se NPs in vivo. Fluorescence images of the major organs after intravenous injection of rats with Cy5-labeled HA-Se NPs. The organs were collected at 24 h post-injection from non-treated rats and at 6, 12, and 24 h post-injection from SCI rats
Next, we explored the targeting efficiency of HA-Se NPs in vitro, using LPS to induce CD44 expression in astrocytes. The time-dependent internalization of HA-Se NPs by astrocytes was confirmed based on a gradual increase in red fluorescence, which we attributed to CD44 expression. To corroborate this observation, we blocked CD44 with HA [37, 52, 53]. In LPS-activated astrocytes, time-dependent internalization was observed in both the HA pretreatment and non-pretreatment groups, being stronger in the latter group during the initial 4 and 8 h (Fig. 4B and Additional file 1: Figure S9). These results validated that enhanced HA-Se NPs internalization was achieved via CD44. To assess NP targeting efficacy in vivo, rats were administered Cy5-labeled HA-Se NPs after SCI. Following injection, the injured spinal cord exhibited a high fluorescent signal within a mere 6-h window, with the fluorescent signal remaining strong at 12 h and subsequently waning at 24 h, possibly due to deterioration of the HA-Se NP shell structure. Cy5-labeled HA-Se NPs accumulated in the heart, liver, spleen, lungs, and kidneys, particularly in the liver and kidneys, which may suggest that the NPs are metabolized via these two organs (Fig. 4C). The fluorescence signal of the liver was gradually increased and reached to maximum at 12 h post-injection.
Neuroprotective activity of HA-Se NPs in vivo
To investigate the neuroprotective activity of HA-Se NPs in vivo, BBB scores were utilized to evaluate the restoration of locomotor function over a 12-week post-injury duration. The recovery of locomotor function was notably accelerated in all treatment groups as opposed to the saline group within a 12-week period following SCI. The group receiving 10 mg kg−1 exhibited a notably elevated BBB score compared to the other groups (Fig. 5A). This observation is also supported by visual inspection of SCI rat spinal cord tissues (Fig. 5B). Assessment of the pathological alterations that transpired at the site of the lesion revealed that the condition of spinal cords was notably inferior in the saline group, with greater numbers of inflammatory cells detected. In contrast, the animals receiving HA-Se NPs exhibited varying degrees of improvement in spinal cord integrity. Figure 5C illustrates that the animals receiving 10 mg/kg HA-Se NPs displayed the least extensive lesion cavity, with a sparse presence of inflammatory cells (Additional file 1: Figure S10). Based on the aforementioned observations, it can be inferred that the application of HA-Se NPs was effective in halting the progression of secondary injury cascades.
Fig. 5.

Neuroprotective effects of HA-Se NPs. A Basso Beattie Bresnahan (BBB) scores of SCI rats. B Gross images of the spinal cord in different groups at 12 weeks post-injury. The lesion site is denoted by the red circle. C Images of hematoxylin and eosin (H&E) right, three magnified images of the areas marked by the blue squares. Scale bars = 500 and 50 μm, respectively. Dotted line and arrows indicate the lesion cavity and inflammatory cells, respectively. D Images of an injured spinal cord stained with LFB. Scale bars = 500 and 50 μm, respectively. Dotted line indicates the lesion cavity. E Representative TEM images of different experimental groups after treatment with saline or HA-Se NPs at the indicated concentrations. Scale bar = 2 μm. Arrows indicate myelin sheaths. F Western blot analysis of cleaved caspase-3 in injured spinal cord tissues 24 h after injury. GAPDH was used as the internal control. G Densitometric analysis of cleaved caspase-3 levels based on (D). **P < 0.01 in relation to the saline group
Axonal demyelination is a common result of SCI [6, 54, 55]. In light of this, we utilized LFB staining to assess the degree of myelination in axons. As indicated by Fig. 5D, there was a reduction in the number of intact myelin sheaths in the group treated with saline. HA-Se NPs treatment alleviated the severity of demyelination, indicating that HA-Se NPs preserved myelin sheaths following injury. To further verify changes in myelination, we performed TEM analysis of the myelin sheaths. In the saline-treated group, TEM images indicated that the myelin sheaths were disorganized and not compactly arranged (Fig. 5E). Meanwhile, the group treated with HA-Se NPs demonstrated greater amount, size, and thickness of myelin sheaths, with the best outcomes observed in rats treated with 10 mg/kg of HA-Se NPs.
It is worth mentioning that the application of HA-Se NPs brought about a significant reduction in the levels of cleaved caspase-3, which were significantly elevated following SCI (Fig. 5F, G). Western blot confirmed the significant reduction in cleaved caspase-3 levels within the spinal cords of animals treated with HA-Se NPs in comparison to those treated with saline. Subsequently, we assessed the remedial impacts of HA-Se NPs on both neurons and axons. Saline-treated controls exhibited a substantial loss of axons and neurons, as observed via confocal imaging. Conversely, animals administered different concentrations of HA-Se NPs displayed greater myelin sheath and axon retention, which highlighted the protective potential of HA-Se NPs in the affected area (Fig. 6). This outcome is consistent with the observed improvement in locomotive function (Fig. 5A). In addition, a considerable number of CD68- and Iba-1-positive cells (macrophage/microglia) were observed at the injury site in the saline group. In comparison, the HA-Se NPs group exhibited a notable decrease in CD68- and Iba-1-positive cells (Additional file 1: Figure S11). Taken together, these observations supported the therapeutic value of HA-Se NPs in enhancing locomotor function recovery through the preservation of neurons and axons. It should also be noted that the major organs obtained from rats administered HA-Se NPs exhibited no visible alterations (Additional file 1: Figure S12). It can therefore be inferred that HA-Se NPs are safe and effective for the management of SCI.
Fig. 6.

Immunohistochemistry of scar tissue labeled with NeuN (green) and NF200 (red). The group that received a dosage of 10 mg/kg exhibited greater preservation of both neurons and axons within the affected spinal cord tissue, even at 12 weeks post-SCI. Scale bar = 50 μm
Discussion
In the present study, we employed a facile redox approach to prepare the HA-Se NPs that specifically target CD44 receptor. HA-Se NPs scavenged ROS and restricted inflammatory responses. Further, HA-Se NPs efficiently accumulated at lesions by targeting CD44. Consequently, HA-Se NPs protected axons and neurons within the injury site, improving functional recuperation in a rat model of SCI. Therefore, we believe that CD44-targeting HA-Se NPs represent a viable therapeutic option for the treatment of SCI.
Therapeutic approaches that involve scavenging ROS and inhibiting inflammatory cascades within the damaged spinal cord have demonstrated efficacy in enhancing functional recovery following SCI [56, 57]. In the preliminary in vitro experiments, HA-Se NPs effectively scavenged DPPH free radicals, in a manner dependent on concentration and exposure time. To explore the capacity of HA-Se NPs for reducing ROS at the cellular level, H2O2 was used to mimic oxidative stress. HA-Se NPs significantly reduced ROS levels during in vitro culture of astrocytes and PC12 cells, in addition to reducing cell death. The damaged and surrounding spinal cord tissues are subject to infiltration by microglia/macrophages, which subsequently secrete proinflammatory cytokines, shaping a hostile microenvironment that hinders SCI repair [58–60]. Our results showed that HA-Se NPs dramatically suppress LPS-induced inflammatory cytokine (TNF-α, IL-1β, and IL-6) production. The anti-inflammatory capacity of HA-Se NPs was further demonstrated in our rat SCI model, based on a reduction in CD68- and Iba-1-positive cell infiltration. Taken together, these observations highlight the protective effect of HA-Se NPs, which shield spinal cord cells from oxidative stress-induced injury and aberrant inflammation.
Excessive ROS generation and the consequent aggressive inflammatory response are known to impede SCI repair [18]. Therefore, a prompt therapeutic intervention that involves the elimination of ROS and suppression of inflammation can yield substantial benefits in terms of sustained neurological and functional recovery [14]. In addition, targeting ability is an essential aspect in the enhanced curative effect and lesser side effects associated with NPs [61]. Our results showed that the CD44 receptor was highly expressed in astrocytes and microglia following SCI, in accordance with previous research [44, 62]. HA is a well-known ligand of the CD44 receptor [63]. Through series in vitro studies, we found that the HA-Se NPs were effectively internalized by astrocytes. In addition, the HA-Se NPs effectively accumulated within damaged spinal cord tissue, where they persist for a duration of 24 h. The positive impact of HA-Se NPs may be attributed to their exceptional targeting capability, which enables prolonged accumulation within spinal cord tissue.
Conclusions
Herein, we developed CD44-targeting HA-Se NPs for enhanced SCI recovery through ROS elimination and the suppression of inflammatory responses. Upon SCI, astrocytes and certain microglial cells exhibit upregulated CD44 expression, which facilitate the accumulation HA-Se NPs and consequent protection of axons and neurons within the injury site. The aforementioned observations underscore the auspicious therapeutic potential of HA-Se NPs in oxidative stress-induced disease or their viability as potential nanocarriers for the treatment of SCI.
Supplementary Information
Additional file 1: Figure S1. The stability of HA-Se NPs in pH 7.4 PBS buffer. Figure S2. Transmission electron microscopy (TEM) image of HA-Se NPs revealing a spherical shape, with a mean diameter of approximately 95 nm. Figure S3. X-ray photoelectron spectrum of hyaluronic acid-selenium nanoparticles (HA-Se NPs). Figure S4. In vitro biocompatibility of HA-Se NPs. Viability of PC12 cells and astrocytes incubated with different concentrations of HA-Se NPs for (A) 24 and (B) 48 h. Data are presented as mean ± SD (n = 3 for each group). Figure S5. The HA-Se nanoparticles scavenge reactive oxygen species (ROS) to protect astrocytes from oxidative damage. [H2O2] =100 μM. Figure S6. HA-Se NPs scavenge ROS to protect PC12 cells in vitro. (A) Live/dead staining of PC12 cells. Scale bar = 20 μm. (B) Quantitative analysis of dead cells. (C) Intracellular ROS levels in PC12 cells were measured using DCFH-DA staining. Scale bar = 20 μm. (D) Quantitative analysis of DCF fluorescence intensity in the cells. **P<0.01. Figure S7. Quantitative analysis of the mean fluorescence intensity of CD44 staining in Fig. 4A. **P<0.01 compared to the control group. Figure S8. Astrocytes overexpress CD44 upon LPS exposure. Western blot analysis of CD44 in astrocytes upon (A) LPS and (C) glutamate activation. (B, D) Densitometric analysis of CD44 levels based on the data in (A) and (C), respectively. *P<0.05, **P<0.01. Figure S9. Quantitative analysis of the mean fluorescence intensity of Cy5-HA-Se NPs in Fig. 4B. **P<0.01 in comparison to the LPS/8 h group. Figure S10. Quantitative analysis of inflammatory cells in Figure 5C,* P < 0.05, in the saline group compared with the 10 mg/kg group. Figure S11. Immunohistochemistry staining of scar tissue labeled with Iba-1 (green) and CD68 (red) 12 weeks after SCI. Scale bar = 50 μm. Figure S12. Hematoxylin & eosin (H&E) staining of the major organs in the experimental group. Table S1. Information on the antibodies used for immunofluorescence (IF) staining. Table S2. Antibodies used for western blotting (WB).
Acknowledgements
Not applicable.
Abbreviations
- BBB
Basso Beattie Bresnahan
- BSA
Bovine serum albumin
- CLSM
Confocal laser-scanning microscope
- DMEM
Dulbecco’s modified Eagle’s medium
- LPS
Lipopolysaccharide
- GFAP
Glial fibrillary acidic protein
- H&E
Hematoxylin and eosin
- HA
Hyaluronic acid
- PBS
Phosphate-buffered saline
- ROS
Reactive oxygen species
- SCI
Spinal cord injury
- SEM
Scanning electron microscopy
- TEM
Transmission electron microscopy
- NPs
Nanoparticles
Author contributions
WL: Conceptualization, Formal analysis, Writing-original draft and editing. YL: Conceptualization, Writing-review and editing. JZ: Formal analysis, Writing-review and editing. RN: Data curation, Writing-review and editing. CX: Writing-review and editing. MZ: Data curation. CX: Writing-review and editing, Funding acquisition. WL: Conceptualization, Writing-review and editing, Supervision, Funding acquisition. RG: Conceptualization, Funding acquisition, Writing review and editing, Supervision.
Funding
This work was supported by grants from the National Natural Science Foundation of China (grant number: 82271411, 51803072), Jilin Provincial Finance Program (Grant number: 2022SCZ10, 2022SCZ25, 2023SCZ41, and 2021SCZ07), Jilin Provincial Science and Technology Program (Grant number: YDZJ202201ZYTS038, 20200201341JC), Youth Support Programmed Project of China Japan Union Hospital of Jilin University(2022qnpy11).
Availability of data and materials
No datasets were generated or analysed during the current study.
Declarations
Ethics approval and consent to participate
All animal procedures were performed under the guidelines of the institutional review board and the ethics committee of Jilin University (approval No. SY202103013).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Wenqi Luo and Yueying Li contributed equally to this work.
Contributor Information
Wanguo Liu, Email: liuwanguo6016@jlu.edu.cn.
Rui Gu, Email: gurui@jlu.edu.cn.
References
- 1.Ong W, Pinese C, Chew SY. Scaffold-mediated sequential drug/gene delivery to promote nerve regeneration and remyelination following traumatic nerve injuries. Adv Drug Deliv Rev. 2019;149–150:19–48. doi: 10.1016/j.addr.2019.03.004. [DOI] [PubMed] [Google Scholar]
- 2.Hutson TH, Di Giovanni S. The translational landscape in spinal cord injury: focus on neuroplasticity and regeneration. Nat Rev Neurol. 2019;15:732–745. doi: 10.1038/s41582-019-0280-3. [DOI] [PubMed] [Google Scholar]
- 3.Huang H, Young W, Skaper S, Chen L, Moviglia G, Saberi H, et al. Clinical neurorestorative therapeutic guidelines for spinal cord Injury (IANR/CANR version 2019) J Orthop Translat. 2020;20:14–24. doi: 10.1016/j.jot.2019.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Silva NA, Sousa N, Reis RL, Salgado AJ. From basics to clinical: a comprehensive review on spinal cord injury. Prog Neurobiol. 2014;114:25–57. doi: 10.1016/j.pneurobio.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 5.Li L, Xiao B, Mu J, Zhang Y, Zhang C, Cao H, et al. A MnO2 nanoparticle-dotted hydrogel promotes spinal cord repair via regulating reactive oxygen species microenvironment and synergizing with mesenchymal stem cells. ACS Nano. 2019;13:14283–14293. doi: 10.1021/acsnano.9b07598. [DOI] [PubMed] [Google Scholar]
- 6.Ahuja CS, Wilson JR, Nori S, Kotter MRN, Druschel C, Curt A, et al. Traumatic spinal cord injury. Nat Rev Dis Primers. 2017;3:17018. doi: 10.1038/nrdp.2017.18. [DOI] [PubMed] [Google Scholar]
- 7.Führmann T, Anandakumaran PN, Shoichet MS. Combinatorial therapies after spinal cord injury: how can biomaterials help? Adv Healthc Mater. 2017;6:1601130. doi: 10.1002/adhm.201601130. [DOI] [PubMed] [Google Scholar]
- 8.Wu W, Lee SY, Wu X, Tyler JY, Wang H, Ouyang Z, et al. Neuroprotective ferulic acid (FA)-glycol chitosan (GC) nanoparticles for functional restoration of traumatically injured spinal cord. Biomaterials. 2014;35:2355–2364. doi: 10.1016/j.biomaterials.2013.11.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cerqueira SR, Oliveira JM, Silva NA, Leite-Almeida H, Ribeiro-Samy S, Almeida A, et al. Microglia response and in vivo therapeutic potential of methylprednisolone-loaded dendrimer nanoparticles in spinal cord injury. Small. 2013;9:738–749. doi: 10.1002/smll.201201888. [DOI] [PubMed] [Google Scholar]
- 10.Wang XJ, Shu GF, Xu XL, Peng CH, Lu CY, Cheng XY, et al. Combinational protective therapy for spinal cord injury medicated by sialic acid-driven and polyethylene glycol based micelles. Biomaterials. 2019;217:119326. doi: 10.1016/j.biomaterials.2019.119326. [DOI] [PubMed] [Google Scholar]
- 11.Wang XJ, Peng CH, Zhang S, Xu XL, Shu GF, Qi J, et al. Polysialic-acid-based micelles promote neural regeneration in spinal cord injury therapy. Nano Lett. 2019;19:829–838. doi: 10.1021/acs.nanolett.8b04020. [DOI] [PubMed] [Google Scholar]
- 12.Shi Y, Kim S, Huff TB, Borgens RB, Park K, Shi R, et al. Effective repair of traumatically injured spinal cord by nanoscale block copolymer micelles. Nat Nanotechnol. 2010;5:80–87. doi: 10.1038/nnano.2009.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Song YH, Agrawal NK, Griffin JM, Schmidt CE. Recent advances in nanotherapeutic strategies for spinal cord injury repair. Adv Drug Deliv Rev. 2019;148:38–59. doi: 10.1016/j.addr.2018.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Andrabi SS, Yang J, Gao Y, Kuang Y, Labhasetwar V. Nanoparticles with antioxidant enzymes protect injured spinal cord from neuronal cell apoptosis by attenuating mitochondrial dysfunction. J Control Release. 2020;317:300–311. doi: 10.1016/j.jconrel.2019.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rao S, Lin Y, Du Y, He L, Huang G, Chen B, et al. Designing multifunctionalized selenium nanoparticles to reverse oxidative stress-induced spinal cord injury by attenuating ROS overproduction and mitochondria dysfunction. J Mater Chem B. 2019;7:2648–2656. doi: 10.1039/c8tb02520g. [DOI] [PubMed] [Google Scholar]
- 16.Wang H, Lin F, Wu Y, Guo W, Chen X, Xiao C, et al. Carrier-free nanodrug based on co-assembly of methylprednisolone dimer and Rutin for combined treatment of spinal cord injury. ACS Nano. 2023;17:12176–12187. doi: 10.1021/acsnano.3c00360. [DOI] [PubMed] [Google Scholar]
- 17.Nukolova NV, Aleksashkin AD, Abakumova TO, Morozova AY, Gubskiy IL, Kirzhanova ЕА, et al. Multilayer polyion complex nanoformulations of superoxide dismutase 1 for acute spinal cord injury. J Control Release. 2018;270:226–236. doi: 10.1016/j.jconrel.2017.11.044. [DOI] [PubMed] [Google Scholar]
- 18.Gao Y, Vijayaraghavalu S, Stees M, Kwon BK, Labhasetwar V. Evaluating accessibility of intravenously administered nanoparticles at the lesion site in rat and pig contusion models of spinal cord injury. J Control Release. 2019;302:160–168. doi: 10.1016/j.jconrel.2019.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mu X, He H, Wang J, Long W, Li Q, Liu H, et al. Carbogenic nanozyme with ultrahigh reactive nitrogen species selectivity for traumatic brain injury. Nano Lett. 2019;19:4527–4534. doi: 10.1021/acs.nanolett.9b01333. [DOI] [PubMed] [Google Scholar]
- 20.Zhang T, Lin F, Liu W, Liu Y, Guo Z, Xiao C, et al. Reactive oxide species-scavenging lipid-polymer nanoparticles for neuroprotection after spinal cord injury. Appl Mater Today. 2021;24:101109. doi: 10.1016/j.apmt.2021.101109. [DOI] [Google Scholar]
- 21.Thuret S, Moon LDF, Gage FH. Therapeutic interventions after spinal cord injury. Nat Rev Neurosci. 2006;7:628–643. doi: 10.1038/nrn1955. [DOI] [PubMed] [Google Scholar]
- 22.Luo W, Wang Y, Lin F, Liu Y, Gu R, Liu W, et al. Selenium-doped carbon quantum dots efficiently ameliorate secondary spinal cord injury via scavenging reactive oxygen species. Int J Nanomedicine. 2020;15:10113–10125. doi: 10.2147/IJN.S282985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jiang X, Liu X, Yu Q, Shen W, Mei X, Tian H, et al. Functional resveratrol-biodegradable manganese doped silica nanoparticles for the spinal cord injury treatment. Mater Today Bio. 2022;13:100177. doi: 10.1016/j.mtbio.2021.100177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kim JW, Mahapatra C, Hong JY, Kim MS, Leong KW, Kim HW, et al. Functional recovery of contused spinal cord in rat with the injection of optimal-dosed cerium oxide nanoparticles. Adv Sci (Weinh) 2017;4:1700034. doi: 10.1002/advs.201700034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gong W, Zhang T, Che M, Wang Y, He C, Liu L, et al. Recent advances in nanomaterials for the treatment of spinal cord injury. Mater Today Bio. 2023;18:100524. doi: 10.1016/j.mtbio.2022.100524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liu Y, Lin F, Zhang T, Wu C, Liu W, Wang H, et al. Toxic aldehyde-scavenging polypeptides mitigate secondary injury after spinal cord injury. Sci China Mater. 2023;66:2925–2937. doi: 10.1007/s40843-022-2409-4. [DOI] [Google Scholar]
- 27.Zeng L, Chen J, Ji S, Chan L, Zheng W, Chen T. Construction of a cancer-targeted nanosystem as a payload of iron complexes to reverse cancer multidrug resistance. J Mater Chem B. 2015;3:4345–4354. doi: 10.1039/c4tb02010c. [DOI] [PubMed] [Google Scholar]
- 28.Nagy G, Pinczes G, Pinter G, Pocsi I, Prokisch J, Banfalvi G. In situ electron microscopy of Lactomicroselenium particles in probiotic bacteria. Int J Mol Sci. 2016;17:1047. doi: 10.3390/ijms17071047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Li Y, Li X, Wong YS, Chen T, Zhang H, Liu C, et al. The reversal of cisplatin-induced nephrotoxicity by selenium nanoparticles functionalized with 11-mercapto-1-undecanol by inhibition of ROS-mediated apoptosis. Biomaterials. 2011;32:9068–9076. doi: 10.1016/j.biomaterials.2011.08.001. [DOI] [PubMed] [Google Scholar]
- 30.Xu C, Qiao L, Guo Y, Ma L, Cheng Y. Preparation, characteristics and antioxidant activity of polysaccharides and proteins-capped selenium nanoparticles synthesized by Lactobacillus casei ATCC 393. Carbohydr Polym. 2018;195:576–585. doi: 10.1016/j.carbpol.2018.04.110. [DOI] [PubMed] [Google Scholar]
- 31.Zhai X, Zhang C, Zhao G, Stoll S, Ren F, Leng X. Antioxidant capacities of the selenium nanoparticles stabilized by chitosan. J Nanobiotechnol. 2017;15:4. doi: 10.1186/s12951-016-0243-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ferreira CA, Ni D, Rosenkrans ZT, Cai W. Scavenging of reactive oxygen and nitrogen species with nanomaterials. Nano Res. 2018;11:4955–4984. doi: 10.1007/s12274-018-2092-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Guo L, Xiao J, Liu H, Liu H. Selenium nanoparticles alleviate hyperlipidemia and vascular injury in ApoE-deficient mice by regulating cholesterol metabolism and reducing oxidative stress. Metallomics. 2020;12:204–217. doi: 10.1039/c9mt00215d. [DOI] [PubMed] [Google Scholar]
- 34.Cardoso R, Hare B, Lind DJ, McLean M, Volitakis CA, Laws I. The APOE ε4 allele is Associated with lower selenium levels in the brain: implications for Alzheimer’s Disease. ACS Chem Neurosci. 2017;8:1459–1464. doi: 10.1021/acschemneuro.7b00014. [DOI] [PubMed] [Google Scholar]
- 35.Ji Z, Zheng J, Ma Y, Lei H, Lin W, Huang J, et al. Emergency treatment and photoacoustic assessment of spinal cord injury using reversible dual-signal transform‐based selenium antioxidant. Small. 2023;19:e2207888. doi: 10.1002/smll.202207888. [DOI] [PubMed] [Google Scholar]
- 36.Cong W, Bai R, Li YF, Wang L, Chen C. Selenium nanoparticles as an efficient nanomedicine for the therapy of Huntington’s disease. ACS Appl Mater Interfaces. 2019;11:34725–34735. doi: 10.1021/acsami.9b12319. [DOI] [PubMed] [Google Scholar]
- 37.Choi KY, Han HS, Lee ES, Shin JM, Almquist BD, Lee DS, et al. Hyaluronic acid-based activatable nanomaterials for stimuli-responsive imaging and therapeutics: beyond CD44-mediated drug delivery. Adv Mater. 2019;31:e1803549. doi: 10.1002/adma.201803549. [DOI] [PubMed] [Google Scholar]
- 38.Khaing ZZ, Milman BD, Vanscoy JE, Seidlits SK, Grill RJ, Schmidt CE. High molecular weight hyaluronic acid limits astrocyte activation and scar formation after spinal cord injury. J Neural Eng. 2011;8:046033. doi: 10.1088/1741-2560/8/4/046033. [DOI] [PubMed] [Google Scholar]
- 39.Lin F, Liu Y, Luo W, Liu S, Wang Y, Gu R, et al. Minocycline-loaded poly(alpha-lipoic acid)-methylprednisolone prodrug nanoparticles for the combined anti-inflammatory treatment of spinal cord injury. Int J Nanomedicine. 2022;17:91–104. doi: 10.2147/IJN.S344491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhang Y, Wang J, Zhang L. Creation of highly stable selenium nanoparticles capped with hyperbranched polysaccharide in water. Langmuir. 2010;26:17617–17623. doi: 10.1021/la1033959. [DOI] [PubMed] [Google Scholar]
- 41.Yang L, Conley BM, Cerqueira SR, Pongkulapa T, Wang S, Lee JK, et al. Effective modulation of CNS inhibitory microenvironment using bioinspired hybrid-nanoscaffold-based therapeutic interventions. Adv Mater. 2020;32:e2002578. doi: 10.1002/adma.202002578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xiao L, Wei F, Zhou Y, Anderson GJ, Frazer DM, Lim YC, et al. Dihydrolipoic acid-gold nanoclusters regulate microglial polarization and have the potential to alter neurogenesis. Nano Lett. 2020;20:478–495. doi: 10.1021/acs.nanolett.9b04216. [DOI] [PubMed] [Google Scholar]
- 43.Wang J, Li D, Liang C, Wang C, Zhou X, Ying L, et al. Scar tissue-targeting polymer micelle for spinal cord injury treatment. Small. 2020;16:e1906415. doi: 10.1002/smll.201906415. [DOI] [PubMed] [Google Scholar]
- 44.Moon C, Heo S, Sim KB, Shin T. Upregulation of CD44 expression in the spinal cords of rats with clip compression injury. Neurosci Lett. 2004;367:133–136. doi: 10.1016/j.neulet.2004.05.101. [DOI] [PubMed] [Google Scholar]
- 45.Greenhalgh AD, David S, Bennett FC. Immune cell regulation of glia during CNS injury and disease. Nat Rev Neurosci. 2020;21:139–152. doi: 10.1038/s41583-020-0263-9. [DOI] [PubMed] [Google Scholar]
- 46.Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541:481–487. doi: 10.1038/nature21029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liddelow SA, Barres BA. Reactive astrocytes: production, function, and therapeutic potential. Immunity. 2017;46:957–967. doi: 10.1016/j.immuni.2017.06.006. [DOI] [PubMed] [Google Scholar]
- 48.Sofroniew MV. Astrocyte barriers to neurotoxic inflammation. Nat Rev Neurosci. 2015;16:249–263. doi: 10.1038/nrn3898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Solier S, Müller S, Cañeque T, Versini A, Mansart A, Sindikubwabo F, et al. A druggable copper-signaling pathway that drives inflammation. Nature. 2023;617:386–394. doi: 10.1038/s41586-023-06017-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ponta H, Sherman L, Herrlich PA. CD44: from adhesion molecules to signaling regulators. Nat Rev Mol Cell Biol. 2003;4:33–45. doi: 10.1038/nrm1004. [DOI] [PubMed] [Google Scholar]
- 51.Puré E, Cuff CA. A crucial role for CD44 in inflammation. Trends Mol Med. 2001;7:213–221. doi: 10.1016/s1471-4914(01)01963-3. [DOI] [PubMed] [Google Scholar]
- 52.Takasugi M, Firsanov D, Tombline G, Ning H, Ablaeva J, Seluanov A, et al. Naked mole-rat very-high-molecular-mass hyaluronan exhibits superior cytoprotective properties. Nat Commun. 2020;11:2376. doi: 10.1038/s41467-020-16050-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Skandalis SS, Karalis T, Heldin P. Intracellular hyaluronan: importance for cellular functions. Semin Cancer Biol. 2020;62:20–30. doi: 10.1016/j.semcancer.2019.07.002. [DOI] [PubMed] [Google Scholar]
- 54.Floriddia EM, Lourenço T, Zhang S, van Bruggen D, Hilscher MM, Kukanja P, et al. Distinct oligodendrocyte populations have spatial preference and different responses to spinal cord injury. Nat Commun. 2020;11:5860. doi: 10.1038/s41467-020-19453-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cunha MI, Su M, Cantuti-Castelvetri L, Müller SA, Schifferer M, Djannatian M, et al. Pro-inflammatory activation following demyelination is required for myelin clearance and oligodendrogenesis. J Exp Med. 2020;217:e20191390. doi: 10.1084/jem.20191390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rao S, Lin Y, Lin R, Liu J, Wang H, Hu W, et al. Traditional Chinese medicine active ingredients-based selenium nanoparticles regulate antioxidant selenoproteins for spinal cord injury treatment. J Nanobiotechnol. 2022;20:278. doi: 10.1186/s12951-022-01490-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lin S, Li D, Zhou Z, Xu C, Mei X, Tian H. Therapy of spinal cord injury by zinc modified gold nanoclusters via immune-suppressing strategies. J Nanobiotechnol. 2021;19:281. doi: 10.1186/s12951-021-01035-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Xi K, Gu Y, Tang J, Chen H, Xu Y, Wu L, et al. Microenvironment-responsive immunoregulatory electrospun fibers for promoting nerve function recovery. Nat Commun. 2020;11:4504. doi: 10.1038/s41467-020-18265-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ma D, Zhao Y, Huang L, Xiao Z, Chen B, Shi Y, et al. A novel hydrogel-based treatment for complete transection spinal cord injury repair is driven by microglia/macrophages repopulation. Biomaterials. 2020;237:119830. doi: 10.1016/j.biomaterials.2020.119830. [DOI] [PubMed] [Google Scholar]
- 60.Liu W, Xu B, Xue W, Yang B, Fan Y, Chen B, et al. A functional scaffold to promote the migration and neuronal differentiation of neural stem/progenitor cells for spinal cord injury repair. Biomaterials. 2020;243:119941. doi: 10.1016/j.biomaterials.2020.119941. [DOI] [PubMed] [Google Scholar]
- 61.Stater EP, Sonay AY, Hart C, Grimm J. The ancillary effects of nanoparticles and their implications for nanomedicine. Nat Nanotechnol. 2021;16:1180–1194. doi: 10.1038/s41565-021-01017-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Matsumoto T, Imagama S, Hirano K, Ohgomori T, Natori T, Kobayashi K, et al. CD44 expression in astrocytes and microglia is associated with ALS progression in a mouse model. Neurosci Lett. 2012;520:115–120. doi: 10.1016/j.neulet.2012.05.048. [DOI] [PubMed] [Google Scholar]
- 63.Salathia S, Gigliobianco MR, Casadidio C, Di Martino P, Censi R. Hyaluronic acid-based nanosystems for CD44 mediated anti-inflammatory and antinociceptive activity. Int J Mol Sci. 2023;24:7286. doi: 10.3390/ijms24087286. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Figure S1. The stability of HA-Se NPs in pH 7.4 PBS buffer. Figure S2. Transmission electron microscopy (TEM) image of HA-Se NPs revealing a spherical shape, with a mean diameter of approximately 95 nm. Figure S3. X-ray photoelectron spectrum of hyaluronic acid-selenium nanoparticles (HA-Se NPs). Figure S4. In vitro biocompatibility of HA-Se NPs. Viability of PC12 cells and astrocytes incubated with different concentrations of HA-Se NPs for (A) 24 and (B) 48 h. Data are presented as mean ± SD (n = 3 for each group). Figure S5. The HA-Se nanoparticles scavenge reactive oxygen species (ROS) to protect astrocytes from oxidative damage. [H2O2] =100 μM. Figure S6. HA-Se NPs scavenge ROS to protect PC12 cells in vitro. (A) Live/dead staining of PC12 cells. Scale bar = 20 μm. (B) Quantitative analysis of dead cells. (C) Intracellular ROS levels in PC12 cells were measured using DCFH-DA staining. Scale bar = 20 μm. (D) Quantitative analysis of DCF fluorescence intensity in the cells. **P<0.01. Figure S7. Quantitative analysis of the mean fluorescence intensity of CD44 staining in Fig. 4A. **P<0.01 compared to the control group. Figure S8. Astrocytes overexpress CD44 upon LPS exposure. Western blot analysis of CD44 in astrocytes upon (A) LPS and (C) glutamate activation. (B, D) Densitometric analysis of CD44 levels based on the data in (A) and (C), respectively. *P<0.05, **P<0.01. Figure S9. Quantitative analysis of the mean fluorescence intensity of Cy5-HA-Se NPs in Fig. 4B. **P<0.01 in comparison to the LPS/8 h group. Figure S10. Quantitative analysis of inflammatory cells in Figure 5C,* P < 0.05, in the saline group compared with the 10 mg/kg group. Figure S11. Immunohistochemistry staining of scar tissue labeled with Iba-1 (green) and CD68 (red) 12 weeks after SCI. Scale bar = 50 μm. Figure S12. Hematoxylin & eosin (H&E) staining of the major organs in the experimental group. Table S1. Information on the antibodies used for immunofluorescence (IF) staining. Table S2. Antibodies used for western blotting (WB).
Data Availability Statement
No datasets were generated or analysed during the current study.