

The feline blood group system was first characterised in 1962 following the identification of two major blood types (designated A and B) (Eyquem et al 1962). One blood group system with three types (A, B and AB) has since been identified (Auer & Bell 1981). Naturally occurring antibodies against foreign blood types in the cat are responsible for the premature destruction of transfused red cells, clinically severe transfusion reactions and neonatal isoerythrolysis.
Type A cats given type B blood may develop a mild transfusion reaction that is often not clinically apparent. The recipient's packed cell volume (PCV) falls to pre-transfusion levels within days of the transfusion (Giger & Bucheler 1991). Type B cats that are transfused with type A blood invariably develop rapid, potentially fatal transfusion reactions following even a single transfusion of a small volume of blood (Auer & Bell 1981).
This paper reviews current understanding of feline blood groups and alloantibodies, and their importance in transfusion medicine.
Feline blood group prevalence
In world-wide studies, type A has consistently been found to be the most common blood type. The proportions of type B cats, however, have shown considerable geographical variations (Eyquem et al 1962, Auer & Bell 1981, Ejima et al 1986, Giger et al 1989, Giger et al 1991a, 1991b, Continenza et al 1992, Giger et al 1992, Knottenbelt et al 1999a)(Table 1). The frequency of type B cats varies dramatically in different breeds (Table 2), however type AB cats are consistently rare (Auer & Bell 1981, Giger et al 1991a, 1991b, Griot-Wenk et al 1996, Knottenbelt et al 1999a). Wild felids have recently been reported to have the same blood groups as domesticated cats (Griot-Wenk & Giger 1999).
Table 1.
Frequency of blood types in domestic short and long-haired cats world-wide
| Country | n | Type A (%) | Type B (%) | Type AB (%) | References |
|---|---|---|---|---|---|
| Australia | 1895 | 73.3 | 26.3 | 0.4 | Auer & Bell (1981) |
| Austria | 101 | 97 | 3 | 0 | Giger et al (1992) |
| England | 477 | 97.1 | 2.9 | 0 | Holmes (1950) |
| Finland | 61 | 100 | 0 | 0 | Giger et al (1992) |
| France | 350 | 85.1 | 14.9 | 0 | Eyquem et al (1962) |
| Germany | 600 | 94.0 | 6.0 | 0 | Giger et al (1992) |
| Holland | 95 | 95.8 | 3.1 | 1.1 | Giger et al (1992) |
| Italy | 401 | 88.8 | 11.2 | 0 | Giger et al (1992) |
| Japan | 265 | 89.3 | 1.0 | 9.7 | Ejima et al (1986) |
| Scotland | 70 | 97.1 | 2.9 | 0 | Giger et al (1992) |
| 137 | 87.6 | 8.0 | 4.4 | Knottenbelt et al (1999a) | |
| Switzerland | 1018 | 99.6 | 0.4 | 0 | Giger et al (1992) |
| USA | 1072 | 99.7 | 0.3 | 0 | Giger et al (1989) |
| 3785 | 98.1 | 1.7 | 0.1 | Giger et al (1991b) |
Note: n=number of cats tested; %=percentage of cats.
Table 2.
Frequency of feline blood types in different breeds worldwide
| Breed | Country | n | Type A (%) | Type B (%) | Type AB (%) |
|---|---|---|---|---|---|
| Abyssinian | USA | 194 | 79.9 | 20.1 | |
| American short-hair | USA | 15 | 100.0 | ||
| Bengal | UK | 8 | 50.0 | 50.0 | |
| Birman | USA | 216 | 82.4 | 17.6 | |
| UK | 11 | 72.7 | 9.1 | 18.2 | |
| British short hair | USA | 85 | 41.2 | 58.8 | |
| UK | 105 | 41.0 | 57.1 | 1.9 | |
| Burmese | USA | 25 | 100.0 | ||
| UK | 6 | 83.3 | 16.7 | ||
| Devon Rex | USA | 100 | 57.0 | 43.0 | |
| UK | 2 | 100.0 | |||
| Norwegian Forest | USA | 20 | 100.0 | ||
| Oriental short hair | USA | 15 | 100.0 | ||
| Persian | USA | 170 | 75.9 | 24.1 | |
| UK | 16 | 87.5 | 12.5 | ||
| Italy * | 38 | 97.4 | 2.6 | ||
| Germany † | 25 | 84.0 | 16.0 | ||
| Ragdoll | UK | 2 | 100.0 | ||
| Scottish Fold | USA | 27 | 85.2 | 14.8 | |
| Siamese | USA | 99 | 100.0 | ||
| UK | 4 | 100.0 | |||
| Somali | USA | 27 | 77.8 | 22.2 | |
| UK | 9 | 77.8 | 22.2 | ||
| Tonkinese | USA | 31 | 100.0 |
Note: n=number of cats tested; %=percentage of cats.
Figures taken from Giger et al 1991a (USA) and Knottenbelt et al 1999a (UK) except:
Continenza et al 1992
Von Haarer & Grunbaum 1990.
Blood group inheritance
Feline blood types are determined by at least two alleles (a or b) at the same gene locus (Giger et al 1991a). The type a allele appears to be completely dominant over the type b allele. Therefore cats with phenotype A may have the genotype a/a or a/b, whilst only cats homozygous for the B allele express significant amounts of the type B erythrocyte antigen (Giger et al 1991a) and none of the type A antigen. With the exception of dogs and cats, inheritance of blood types within a blood group system in most other species is typically by codominant alleles (Symons & Bell 1992). Cats are therefore unusual in inheriting type B as a recessive trait (Rippee et al 1989, Giger et al 1991a).
The AB blood type was previously thought to result from matings between two phenotypic type A cats both with the genotype a/b, or between a phenotypic type A cat (genotype a/b) and a type B cat, or between two type B cats (Griot-Wenk & Giger 1991). However, more recent work suggests that type AB is independently inherited in the feline blood group system, since type AB cats are rarely the offspring of matings between type A and type B cats (Griot-Wenk et al 1996). The ab allele appears to be recessive to the a allele but dominant over the b allele (Griot-Wenk et al 1996). Type AB cats are only found in breeds in which type B cats have been identified (Griot-Wenk et al 1996).
Blood group antigens
The antigens in the feline AB blood group are not serologically related to human ABO blood group antigens (Auer & Bell 1981). By 38 days of gestation, type A and type B antigens are present on foetal erythrocytes (Auer & Bell 1981). Blood type antigens are thought to be determined by the form of neuraminic acid attached to the glycolipids of the red cell (specifically the ganglioside GD3), and possibly also a glycoprotein of the erythrocyte membrane (Andrews et al 1992). Type A blood has NeuGc-NeuGc-Galactose-Glucose-Ceramide ([NeuGc]2 GD3) (where NeuGc represents N-glycolylneuraminic acid) as the major glycolipid (Butler et al 1991a). N-acetylneuraminic acid (NeuAc) is, however, also present in variable proportions in type A blood. This variability may be due to the homozygous or heterozygous nature of cats with type A blood (Griot-Wenk et al 1993). Type B blood, in contrast to type A blood, expresses no NeuGc, the major glycolipid being NeuAc-NeuAc-Galactose-Glucose-Ceramide ([NeuAc]2GD3)(Butler et al 1991a). Type B erythrocytes are therefore characterised by [NeuAc]2 GD3, the presence of NeuAc on the 50 kD erythrocyte membrane glycoprotein and a lack of detectable NeuGc (Andrews et al 1992).
Type AB blood shows co-expression of both type A and type B antigens (Andrews et al 1992, Griot-Wenk et al 1996). The proportions of NeuAc and NeuGc, however, vary between different type AB individuals (Andrews et al 1992) and intermediate forms of glycolipids have been recovered from type AB red cells (Griot-Wenk et al 1996). Cats without either type A or B red cell antigens (analogous to the human blood group O) have not been reported.
It has been suggested that the enzyme NeuAc hydroxylase is responsible for the conversion of NeuAc to NeuGc in type A cats, and that type B cats lack this enzyme (Butler et al 1991b, Andrews et al 1992). Type AB cats may have a hereditary mutation of the enzyme or a gene that alters enzyme expression. This theory is supported by the finding that type AB cats are only produced from crosses involving an AB phenotype (Auer & Bell 1981). This theory however, fails to account for the finding of variable amounts of NeuAc in type A cats (Griot-Wenk et al 1993), unless enzyme activity was limited by an alternative mechanism in some type A cats. Measurement of NeuAc hydroxylase activity in cats has not been performed.
Alloantibodies
The proportion of type A cats with low levels of naturally occurring antibody directed against type B red cells varies geographically (Auer & Bell 1981, Ejima et al 1986, Giger et al 1989, Bucheler & Giger 1993, Knottenbelt et al 1999b). However all type B cats appear to have high levels of naturally occurring anti-A antibody (Giger et al 1989, Knottenbelt et al 1999b). Cats with type AB blood do not have alloantibodies (Griot-Wenk et al 1996). The naturally-occurring anti-B antibodies of type A cats are weak IgM agglutinins and weak haemolysins comprising equal proportions of IgM and IgG (Bucheler & Giger 1993). Anti-A alloantibodies appear to be strong anti-A haemolysins and haemagglutinins predominantly of the IgM class (Wilkerson et al 1991, Bucheler and Giger 1993), although some haemagglutinating activity has also been associated with IgG (Wilkerson et al 1991).
Naturally-occurring alloantibodies are believed to result from exposure to epitopes that are commonly found in nature (usually as structural components of a variety of organisms including plants, bacteria and protozoa), and that are similar or identical to blood group antigens (Male 1996, Tizard 1996). In man, the epitopes responsible are thought to be microbial antigens present on intestinal bacteria (Male 1996, Kuby 1997, 1997b). In cats however, the origin of these epitopes has not been established. Exposure to these epitopes results in the formation of antibodies against antigens that the individual does not naturally possess. The antibody produced will then cross-react with similar antigens (such as blood group antigens) found on foreign erythrocytes (Kuby 1997a). Exposure to epitopes that are similar to self antigens will not result in antibody formation due to self tolerance (Kuby 1997b). This mechanism explains the reported absence of alloantibodies in type AB cats (Auer & Bell 1981), since the antigens found in nature that resemble either type A or type B blood group antigens will be recognised as ‘self’. The absence of naturally-occurring alloantibodies in many domestic animal species may be due to an absence of exposure to appropriate natural epitopes. The presence of NeuAc (the type B antigen) in variable proportions on type A cells (Griot-Wenk et al 1993) may account for the variation in anti-B antibody titres recorded in type A cats, since NeuAc may be recognised as self antigen. It has not been established whether type A cats without circulating anti-B antibodies express higher proportions of NeuAc than those type A cats with higher titres of naturally-occurring alloantibodies.
Cross-matching and blood typing
Blood typing determines the nature of blood group antigens on the erythrocyte membranes, whilst cross-matching detects the presence of significant serum levels of antibody to red cell antigens. Cross-matching assesses the effect that recipient serum antibodies will have on the donor cells (major cross-match) and the effect that donor serum will have on recipient cells (minor cross-match). Since the main aim of the transfusion is to provide the recipient with red blood cells, it is most important that recipient serum antibodies do not destroy transfused donor cells and in so doing evoke an acute transfusion reaction. The minor cross-match assesses the likelihood of recipient red cell destruction by the donor serum, a much smaller risk since the volume of transfused serum will comprise only a small volume of the total recipient serum. The use of agglutination techniques for blood typing and cross-matching red cells remains the most simple, sensitive and reliable test available (Hughes-Jones 1988). Both typing and cross-matching should be performed before administration of a transfusion, as donor cells will interfere with the result (Niggermeier et al 2000).
The blood type of donor and recipient cats can often be inferred from the result of cross-matching even if neither cat's blood type is known. There are, however, a number of flaws in this approach. The presence of a positive reaction in a cross-match is not specific for mismatches involving the AB blood group antigens since a positive reaction could also be seen when an individual possesses antibodies to red cell antigens other than those of the AB system, although this has not been documented. These non-AB antibodies would be particularly important if the patient had been previously transfused. Also, the presence of a compatible cross-match on first transfusion does not rule out development of a delayed transfusion reaction, particularly if associated with potential non-AB incompatibilities.
Cross-matching with random cats will not identify type AB cats. Type AB red cells express both the type A and type B antigens, and do not possess naturally occurring alloantibodies. This means that type AB cells will act antigenically as both type A and type B, whilst serologically AB cats act as type A. The co-expression of both type A and type B antigens explains why type AB blood will show agglutination with both anti-A and anti-B sera. The agglutination of type AB cells with anti-A alloantibody is, however, not typically as marked as the agglutination seen with type A cells (Griot-Wenk et al 1996).
Traditionally, feline blood typing was performed by cross-matching erythrocytes from the cat to be tested with both anti-A and anti-B antisera obtained from cats of known blood type. However, this method posed potential problems because of the difficulty in obtaining high-titred anti-B antisera without immunising type A cats with type B blood (Butler et al 1991b). More recently, the demonstration that lectins may be used in blood typing has circumvented this problem. Triticum vulgaris lectin (a wheat germ agglutinin) specifically binds to the NeuAc terminal of the 50 kD erythrocyte membrane protein (Nagata & Burger 1974). Since the presence of the NeuAc terminal is one of the characteristics of feline type B cells (Andrews et al 1992), this lectin can be used to identify type B cells. The agglutination of type B cells by Triticum vulgaris lectin is more marked than that seen with anti-B serum antibody, and serial dilution of the lectin gives consistent results (Butler et al 1991b). Despite the presence of variable amounts of NeuAc in type A cats, the Triticum vulgaris lectin has no effect on type A erythrocytes (Butler et al 1991b, Griot-Wenk et al 1993), which may be due to the reduced number of sites available for binding. The Triticum vulgaris lectin has therefore become the standard method of detecting type B antigen on the membranes of both type B and type AB erythrocytes, eliminating the need for immunisation of type A cats. A desk-top blood typing kit (Rapid Vet-H Feline, dms laboratories, Flemington, USA) has recently been developed (Fig 1) which utilises anti-A antiserum for detecting type A antigen and the Triticum vulgaris lectin for detecting type B antigen. The kit provides an accurate method by which feline blood can be routinely typed in veterinary practice (Knottenbelt et al 1999a).
Fig 1.
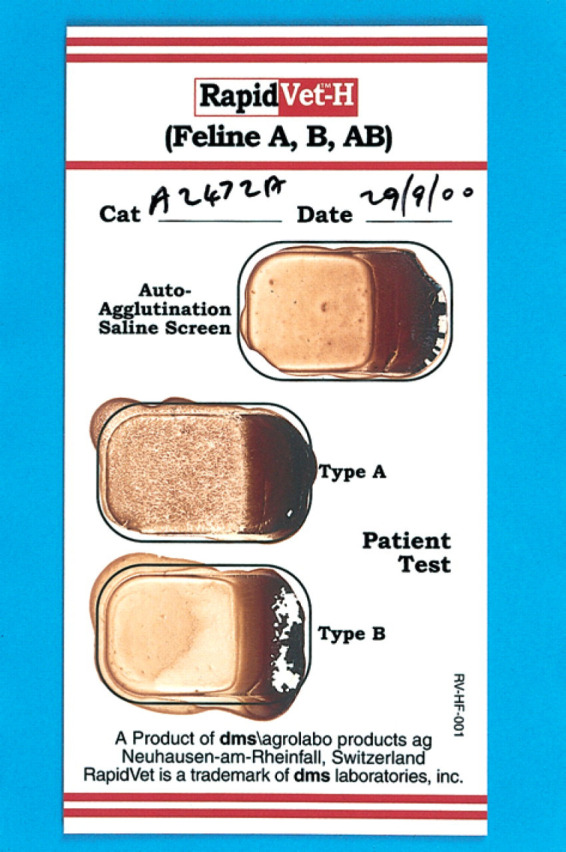
Photograph showing appearance of the Rapid Vet-H blood typing card (dms laboratories, Switzerland) following drying. Agglutination is present in the type A cell but absent from both the autoagglutination saline screen and the type B cells indicating that the cat is blood type A. Cards may be left to dry and kept as a permanent record.
Since cross-matching fails to detect blood type incompatibilities in the absence of naturally-occurring alloantibodies, blood typing remains the only method by which to ensure that AB-system-compatible blood is transfused. Donor and recipient blood typing should be performed before all first transfusions, and both cross-matching and donor blood typing performed before any subsequent transfusions. It must be noted however that a recent mismatched transfusion will interfere with recipient blood typing due to the persistence of donor cells within the recipient's blood (Niggermeier et al 2000).
Transfusion reactions
The most common aim of blood transfusion in clinical practice is to provide the recipient with sufficient erythrocytes to alleviate the more severe clinical signs of anaemia. In the majority of clinical cases, the increase in PCV achieved by transfusion allows the patient's clinical signs to be stabilised for a sufficient period to allow the underlying cause of anaemia to be diagnosed and treated. Ideally, the life span of transfused red cells would be unaffected by transfusion into another animal of the same species. However, this is often not the case in clinical transfusions. The normal feline erythrocyte life span is approximately 80 days (equivalent to a half-life of 40 days) (Marion & Smith 1983). Autologous and allogenic matched transfusions of type A and B blood are well tolerated but the half-life may be slightly shorter (29–39 days) (Giger & Bucheler 1991). In contrast, type B erythrocytes transfused into type A cats have a mean half-life of approximately 2 days (Giger & Bucheler 1991). Survival times of type A cells given to type B cats are even shorter: half the transfused cells are destroyed within minutes to hours depending on alloantibody titres. Obviously, the dramatic shortening of red cell survival time associated with such mismatched transfusions markedly diminishes the therapeutic benefits of the transfusion. Additionally, mismatched transfusions have the potential to put the patient's life in immediate danger due to the development of a transfusion reaction.
Immune-mediated destruction of transfused erythrocytes can be divided into acute and delayed haemolytic reactions. In cats without naturally-occurring alloantibodies, destruction of erythrocytes occurs after the development of an IgG immune response (usually 1–2 weeks after a first transfusion) (Marion & Smith 1983). Naturally-occurring alloantibodies are predominantly of the IgM class (anti-A alloantibodies) or equal proportions of IgM and IgG (anti-B allo antibodies) (Bucheler & Giger 1993). IgG is less effective than IgM at activating complement, complement mediated lysis is incomplete and erythrocytes are destroyed at extravascular sites (Kuby 1997b). Delayed transfusion reactions associated with IgG alloantibodies will usually have less severe clinical manifestations. Delayed reactions are seldom reported, which may be because they are uncommon, or alternatively, because they are mild in nature and therefore not observed and recorded.
Feline transfusion reactions were first reported in 1915 and are characterised by haemoglobinaemia, haemoglobinuria, jaundice, erythrophagocytosis, leucocytosis, oligura and glycosuria (Ottenburg & Thalimer 1915). Since then a number of acute and delayed transfusion reactions have been reported in cats following therapeutic transfusion of incompatible blood (Auer et al 1982, Giger & Akol 1990, Giger & Bucheler 1991, Wilkerson et al 1991b). Acute, severe transfusion reactions invariably occur on first transfusion of type A blood to type B cats (Auer & Bell 1983). The reactions are characterised by systemic anaphylaxis (hypotension, bradycardia, apnoea, urination, defaecation, vomiting and severe neurological depression) and haemolysis (haemoglobinaemia and haemoglobinuria) (Giger & Bucheler 1991).
Some type A cats that are given type B blood will show a mild transfusion reaction due to low levels of naturally-occurring anti-B antibody. Although this reaction may not be apparent clinically, the recipient's PCV will fall to pre-transfusion levels within days of receiving the transfusion (Giger & Bucheler 1991), and the transfusion will therefore have been of only transient benefit. The administration of type B blood to type A cats could result in induction of high titred anti-B antibodies which will then cause an acute severe transfusion reaction if further mismatched blood is given. The risk of a significant reaction occurring following an unmatched transfusion depends both on the blood group prevalence of the donor and recipient breeds and the alloantibody titre of the recipient. This risk will therefore vary worldwide (Table 3).
Table 3.
Summary of the risk of acute transfusion reactions occurring following unmatched transfusions
| Country | Breed of donor | Breed of recipient | Risk of acute transfusion reaction | ||
|---|---|---|---|---|---|
| Severe | Mild | ||||
| UK § | DSH/DLH | DSH/DLH | 2.6% | 7.2% | |
| British shorthair | British shorthair | 8.6% | 23.9% | ||
| DSH/DLH | British shorthair | 17.1% | 35.3% | ||
| US * | DSH/DLH | DSH/DLH | 0.3% | ||
| Persian | Persian | 18.3% | |||
| DSH/DLH | British shorthair | 58.6% | |||
| DSH/DLH | Persian | 24.0% | |||
| Australiat | DSH/DLH | DSH/DLH | 19.4% | ||
| Italy † ® | DSH/DLH | Persian | 2.3% | ||
| Persian | Persian | 2.5% | |||
Note: UK calculations based on the data from Knottenbelt et al 1999b, all other calculations based on the assumption that all type B cats have high titred anti-A antibodies.
Knottenbelt et al 1999b. The severity of transfusion reactions appears to be more closely linked to the titre of haemolytic alloantibodies (Auer & Bell 1983). Since haemolytic alloantibodies are commonly found at higher titres than haemagglutinating alloantibodies (Auer & Bell 1983), the study by Knottenbelt and others may under-estimate the proportion of cats with clinically significant naturally-occurring alloantibody titres.
Giger & Bucheler 1991.
Auer & Bell 1981.
Continenza et al 1992a.
Injection of as little as 1 ml of a 50% suspension of type A cells to type B cats can elicit severe reactions, but subsequent challenges with incompatible blood do not elicit a worsening of the transfusion reaction (Auer & Bell 1983). This is due to antigen-antibody binding causing antibody titres to fall within minutes of the incompatible red cell injection (Christian et al 1951). The severity of acute transfusion reactions appears to be more closely linked with the antibody titre of the recipient rather than the amount of antigen administered (Auer & Bell 1983).
The proportions of the immunoglobulin class involved in the reaction may influence the outcome of mismatched transfusions. IgM is more effective at triggering complement mediated lysis, whilst IgG tends to activate complement poorly (Kuby 1997b). Giger & Bucheler (1991) concluded that the rapid destruction of type A cells transfused into a type B cat occurs predominantly intravascularly and is complement and IgM-mediated. In contrast, type B cells transfused into a type A cat are haemolysed extravascularly by a process involving small amounts of IgG and IgM, without marked complement activation.
Since haemolytic anti-B antibody titres have been shown to fluctuate widely throughout the year in some type A cats (Auer & Bell 1981, 1983), the antibody status of an individual on any one occasion cannot be used to predict the severity of subsequent reactions following any given transfusion. Variations in naturally-occurring antibody titres in cats may be one reason why some type A cats show severe reactions to type B blood, while others develop minimal clinical signs except a fall in post-transfusion PCV. Despite the apparent absence of naturally-occurring antibodies in many type A cats, type B blood should not be transfused into a type A cat because subsequent sensitisation will considerably shorten erythrocyte life-span (Auer & Bell 1983).
Type AB cats pose a particular problem in transfusion medicine. The development of delayed transfusion reactions due to induction of anti-A or anti-B antibodies within the recipient should not occur in type AB recipient cats since the immune system has already been exposed to both type A and type B antigens. Induction of antibodies to antigens other than those of the AB system may, however, still occur. Since AB cats express both type A and type B erythrocyte antigens and possess no naturally-occurring antibodies (Auer & Bell 1981), AB cats should be universal recipients. Griot-Wenk & Giger (1991) substantiated this hypothesis by showing that the life-span of both type A and type B erythrocytes transfused into type AB cats was the same as that of matched transfusions to type A and B cats. No transfusion reactions were observed following these experimental transfusions.
Despite these observations and the absence of anti-A or anti-B alloantibodies in type AB cats, it has recently been suggested that type AB cats should ideally be transfused with type AB or type A blood (Griot-Wenk et al 1996). Although the administration of type B blood to a type AB cat will not result in destruction of transfused red cells, the high titres of naturally-occurring anti-A antibodies in the donor serum may result in destruction of recipient red cells. In a cat with severe anaemia, the further removal of the patient's own red cells (which are already greatly reduced in number) is unlikely to have a significant effect on the patient's post-transfusion clinical condition and PCV. In contrast, if serum containing high titres of alloantibody is given to a cat with minimal or no anaemia, there may be increased potential for red cell destruction due to the larger numbers of recipient cells available for haemolysis. Since the aim of transfusion in clinical cases is usually to maximise red cell numbers for as long a period as possible, type A or AB blood should be given to type AB cats. In situations where only a type B donor is available, the donor red cells should ideally be fractionated to remove serum before administration to a type AB recipient (Giger & Bucheler 1991).
Except for the transfusion of other type AB cats, type AB cats are poor candidates as blood donors since they express both type A and type B antigens in varying proportions. The administration of type AB blood to a type A cat without significant antibody titres will result in the induction of anti-B antibodies and the potential for either a delayed transfusion reaction or severe transfusion reactions if subsequent mismatched transfusions are given.
Summary
The presence of naturally-occurring antibodies against blood group antigens in the cat is clinically significant and may result in ineffective transfusion or severe transfusion reactions. The distribution of blood types in the local cat population may help to establish the risk of transfusion reactions occurring, whilst the magnitude of naturally-occurring antibody titres determines the severity of the reaction.
Although the apparent incidence of clinical transfusion reactions appears to be low, this probably reflects a failure to recognise the complications arising from transfusion. It is vital that screening of donor-recipient compatibility is performed prior to first transfusion of all cats. The preferred method of ensuring donor and recipient compatibility is both blood typing and cross-matching.
References
- Andrews GA, Chavey PS, Smith JE, Rich L. (1992) N-Glycolylneuraminic acid and N-Acetylneuraminic acid define feline blood group A and B antigens. Blood 79, 2485–2491. [PubMed] [Google Scholar]
- Auer L, Bell K. (1981) The AB blood group system of cats. Animal Blood Groups, Biochemistry and Genetics 12, 287–297. [DOI] [PubMed] [Google Scholar]
- Auer L, Bell K. (1983) Transfusion reactions in cats due to AB blood group incompatibility. Research in Veterinary Science 35, 145–152. [PubMed] [Google Scholar]
- Auer L, Bell K, Coates S. (1982) Blood transfusion reactions in the cat. Journal of the American Veterinary Medical Association 180, 729–730. [PubMed] [Google Scholar]
- Bucheler J, Giger U. (1993) Alloantibodies against A and B blood types in cats. Veterinary Immunology and Immunopathology 38, 283–295. [DOI] [PubMed] [Google Scholar]
- Butler M, Andrews GA, Smith JE. (1991a) Thin layer chromatography of the membrane glycolipids from type A and type B cats. Comparative Haematology International 1, 196–199. [Google Scholar]
- Butler M, Andrews GA, Smith JE, Chavey PS. (1991b) Reactivity of lectins with feline erythrocytes. Comparative Haematology International 1, 217–219. [Google Scholar]
- Christian RM, Stewart WB, Yuile D, Ervin M, Young LE. (1951) Limitation of hemolysis in experimental transfusion reactions related to depletion of complement and isoantibody in the recipient. Blood 6, 142. [PubMed] [Google Scholar]
- Continenza R, Lubas G, Gugliucci B. (1992) Indagini prelimisul sistema di gruppo sanguigno AB nel gatto allevato in Italia. Atti Societa Italiana Scienze Veterinarie XLVI, 1470–1477. [Google Scholar]
- Ejima H, Kurokawa K, Ikemoto S. (1986) Feline red blood cell groups detected by naturally occurring isoantibodies. Japanese Journal of Veterinary Science 48, 971–976. [DOI] [PubMed] [Google Scholar]
- Eyquem A, Podliachouk L, Milot P. (1962) Blood groups in chimpanzees, horses, sheep, pigs and other mammals. Annals of the New York Academy of Science 97, 320–328. [DOI] [PubMed] [Google Scholar]
- Giger U, Akol KG. (1990) Acute hemolytic transfusion reaction in an Abyssinian cat with blood type B. Journal of Veterinary Internal Medicine 4, 315–316. [DOI] [PubMed] [Google Scholar]
- Giger U, Bucheler J. (1991) Transfusion of type-A and type-B blood to cats. Journal of the American Veterinary Medical Association 198, 411–418. [PubMed] [Google Scholar]
- Giger U, Bucheler J, Patterson DF. (1991a) Frequency and inheritance of A and B blood types in feline breeds of the United States. Journal of Heredity 82, 15–20. [DOI] [PubMed] [Google Scholar]
- Giger U, Gorman NT, Hubler M, Leidinger JI, Leidinger EF, Lubas G, Niini T, Slappendel RJ. (1992) Frequencies of feline A and B blood types in Europe. Proceedings of the International Society of Animal Genetics Conference 23, Supplement I, 17–18. [Google Scholar]
- Giger U, Griot-Wenk M, Bucheler J, Smith S, Diserens D, Hale A, Patterson DF. (1991b) Geographical variation of the feline blood type frequencies in the United States. Feline Practice 19, 21–27. [Google Scholar]
- Giger U, Kilrain CG, Filippich LJ, Bell K. (1989) Frequencies of feline blood groups in the United States. Journal of the American Veterinary Medical Association 195, 1230–1232. [PubMed] [Google Scholar]
- Griot-Wenk M, Giger U. (1991) Cats with type AB blood in the United States. Journal of Veterinary Internal Medicine 5, 139. [Google Scholar]
- Griot-Wenk M, Giger U. (1999) The AB blood group system in wild felids. Animal Genetics 30, 144–147. [DOI] [PubMed] [Google Scholar]
- Griot-Wenk M, Pahlsson P, Chisholm-Chait A, Spitalnik PF, Giger U. (1993) Biochemical characterisation of the feline AB blood group system. Animal Genetics 24, 401–407. [DOI] [PubMed] [Google Scholar]
- Griot-Wenk ME, Callan MB, Casal ML, Chisholm-Chait A, Spitalnik PF, Patterson DF, Giger U. (1996) Blood type AB in the feline AB blood group system. American Journal of Veterinary Research 57, 1438–1442. [PubMed] [Google Scholar]
- Holmes R. (1950) Blood groups of cats. Journal of Physiology 111, 61. [PubMed] [Google Scholar]
- Hughes-Jones NC. (1988) Monoclonal antibodies as potential blood typing reagents. Immunology Today 9, 68–70. [DOI] [PubMed] [Google Scholar]
- Knottenbelt CM, Addie DD, Day MJ, Mackin AJ. (1999a) Determination of the prevalence of feline blood types in the UK. Journal of Small Animal Practice 40, 115–118. [DOI] [PubMed] [Google Scholar]
- Knottenbelt CM, Day MJ, Cripps PJ, Mackin AJ. (1999b) Measurement of titres of naturally-occurring alloantibodies against feline blood group antigens in the United Kingdom. Journal of Small Animal Practice 40, 365–370. [DOI] [PubMed] [Google Scholar]
- Kuby J. (1997a) Antigen-antibody interaction. In: Immunology 3rd Edition. Kuby J. (ed.) New York: W.H. Freeman and Company, pp 147–148. [Google Scholar]
- Kuby J. (1997b) Hypersensitivity reactions. In: Immunology 3rd Edition. Kuby J. (ed.) New York: W.H. Freeman and Company, pp 430–433. [Google Scholar]
- Male D. (1996) Reactions against blood cells and platelets. In: Immunology 4th Edition. Roitt I, Brostoff J, Male D. (eds). London: Mosby, pp 23.3–23.6. [Google Scholar]
- Marion RS, Smith JE. (1983) Survival of erythrocytes after autologous and allogenic transfusion in cats. Journal of the American Veterinary Medical Association 183, 1437–1439. [PubMed] [Google Scholar]
- Nagata Y, Burger MM. (1974) Wheat germ agglutinin. Molecular characteristics and specificity for sugar binding. Journal of Biological Chemistry 249, 3116–3123. [PubMed] [Google Scholar]
- Niggermeier A, Haberstroh HF, Nelson VE, Giger U. (2000) The accidental transfusion of a type A kitten with type B blood causes a transient switch from blood type A to B. Journal of Veterinary Internal Medicine 14, 214–216. [PubMed] [Google Scholar]
- Ottenburg R, Thalimer W. (1915) Studies in experimental transfusion. Journal of Medical Research 28, 213–229. [PMC free article] [PubMed] [Google Scholar]
- Rippee C, Myers J, Gindy L. (1989) Blood groups. In: Clinical practice of transfusion medicine, 2nd Edition. Peta LD, Swisher SN. (eds). New York: Churchill Livingstone, pp 59–134. [Google Scholar]
- Symons M, Bell K. (1992) Canine blood groups: Description of 20 specificities. Animal Genetics 23, 509–515. [DOI] [PubMed] [Google Scholar]
- Tizard IR. (1996) Red cell antigen and type II hypersensitivity. In: Veterinary Immunology: An Introduction. Tizard IR. (ed.) Philadelphia: W.B. Saunders, pp 359–367. [Google Scholar]
- Von Haarer M, Grunbaum EG. (1990) Zur klinischen bendentung der blutgruppen bei katzen. Die Edelkatze 1, 12–14. [Google Scholar]
- Wilkerson MJ, Meyers KM, Wardrop KJ. (1991) Anti-A isoagglutinins in two type B cats are IgG and IgM. Veterinary Clinical Pathology 20, 10–14. [DOI] [PubMed] [Google Scholar]
- Wilkerson MJ, Wardrop KJ, Meyers KM, Giger U. (1991b) Two cat colonies with A and B blood type and a clinical transfusion reaction. Feline Practice 19, 22–26. [Google Scholar]