

Abstract
In 1998 and 1999, fecal samples were collected from 669 beef cows on 39 farms located within 10 counties of Ontario. Overall prevalences of Giardia, Cryptosporidium muris, and Cryptosporidium parvum in cows were 8.7%, 10.6%, and 18.4%, respectively. Of the 39 farms sampled, Giardia was detected on 64%, Cr. muris on 72%, and Cr. parvum on 90%. Cryptosporidium parvum was detected in 28% of the cows in 1998 and in 5.2% in 1999. Differences between the 2 y were attributed to sampling during calving in 1998 and during gestation in 1999. In 1998, Giardia, Cr. muris, and Cr. parvum were detected in herds provided with municipal water. In 1998, 193 calves were sampled from 10 farms, representing 4 watersheds, in British Columbia. Thirty-six percent of the calves exhibited signs of diarrhea. Overall prevalences of Giardia and Cryptosporidium spp. in calves were 36% and 13%, respectively. There was evidence that calves with Giardia were more likely to develop scours. Restricting cattle from surface water during periods of high shedding may reduce watershed contamination.
Abstract
Résumé — Prévalence de Giardia et Cryptosporidium chez des vaches de boucherie du sud de l’Ontario et chez des veaux de boucherie du sud de la Colombie-Britannique. En 1998 et 1999, des échantillons fécaux ont été prélevés chez 669 vaches de boucherie sur 39 fermes situées dans 10 comtés de l’Ontario. La prévalence globale de Giardia, de Cryptosporidium muris et de Cryptosporidium parvum chez les vaches étaient respectivement de 8,7 %, 10,6 % et 18,4 %. Des 39 fermes participantes, Giardia a été détecté chez 64 %, Cr. muris chez 72 % et Cr. parvum chez 90 % d’entre elles. Cryptosporidium parvum a été détecté chez 28 % des vaches en 1998 et 5,2 % en 1999. Cette différence a été attribuée au moment de l’échantillonnage : au vêlage en 1998 et en cours de gestation en 1999. En 1998, Giardia, Cr. muris et Cr. parvum ont été détectés dans des fermes alimentées par un aqueduc municipal. En 1998, 193 veaux ont été échantillonnés sur 10 fermes provenant de 4 bassins versants en Colombie-Britannique. Des signes de diarrhée étaient présents chez 36 % des veaux. Les prévalences globales de Giardia et de Cryptosporidium spp. chez les veaux étaient respectivement de 36 % et de 13 %. Il y avait des preuves que les veaux avec Giardia étaient plus à risques de développer de la diarrhée profuse. L’éloignement des bovins de l’eau de surface dans les périodes de fortes excrétions fécales pourrait réduire la contamination des bassins versants.
(Traduit par Docteur André Blouin)
Introduction
Giardia and Cryptosporidium spp. have been implicated as a cause of diarrhea in dairy calves (1,2) and are common intestinal parasites of a wide range of vertebrates (3,4). Dairy calves can excrete high numbers of cysts or oocysts for weeks, and there are indications that both diseases can potentially reduce the growth performance of ruminants (5,6). Mortality from either disease is rare, but severe cryptosporidiosis is occasionally responsible for mass fatalities in calves (7,8)
There is evidence that both parasites are zoonotic, as humans have been infected with Giardia spp. from animals (9) and cryptosporidiosis in humans has been linked to infected calves (10) and household pets (11). Outbreaks of both parasites in humans have been attributed to agriculture effluent from livestock (12,13). A surface water study conducted in the vicinity of a cattle ranch in British Columbia demonstrated that numbers of Giardia cysts and Cryptosporidium oocysts in raw water from a creek passing through the ranch were higher downstream than upstream of the ranch, and that the highest concentrations of both parasites in the raw water samples were recorded coincident with peak calving time (14). Consequently, several researchers and public health officers have concluded that cattle play a major role in the contamination of surface water used for both drinking and recreational purposes (1,15,16).
In Canada, studies on the prevalence of Giardia and Cryptosporidium in cattle have focussed mainly on dairy herds (1,16). In the Fraser River valley, Giardia was identified on all 20 dairies examined, whereas Cryptosporidium parvum was found on 80% of dairy farms (1). Further examination of 505 dairy farms in Quebec indicated that 45.7% were positive for Giardia and 88.7% were positive for Cryptosporidium spp. (16). Confinement practices in dairy calf production likely increase both Giardia and Cryptosporidium infection rates, as compared with the production of beef on open range. This possibility is supported by the fact that dairy calves kept outside the barn were less likely to be infected with Giardia than those housed inside (16). Furthermore, reinfection of calves with Giardia after repeated treatment with fenbendazole illustrates that infective cysts may be readily consumed within a confined environment (17). Thus, differences in production practices may account for the one Canadian study that observed higher infection rates of Cryptosporidium in dairy calves (63.3%), as compared with beef calves (18.4%) (18).
Although Giardia and Cryptosporidium are prevalent in calves, it is likely that mature animals serve as reservoirs for the infection of younger animals. Periparturient increases in the excretion of Giardia cysts and Cryptosporidium oocysts have been reported in ewes (19) and we have observed a similar response in beef cows (20). However, the prevalence of Giardia and Cryptosporidium in mature beef cattle or calves has not been well documented within Canada. The objective of the present study was to determine the prevalence of Giardia and Cryptosporidium in beef cows in southern Ontario and southern British Columbia.
Materials and methods
Sample collection — southern Ontario
In 1998, 39 cow-calf farms were selected for participation in the study on the basis of their location within 10 counties distributed within the south, west, and central agricultural regions of southern Ontario with the highest beef cattle populations, and on the owners’ willingness to collaborate. Three to 6 farms per county participated; on each farm, 10 cows were randomly selected from the herd for sampling. During the 1998 collection (March 31 to May 22), fecal samples were collected from 382 cows on the 39 farms: 7 in the central region, 9 in the southern region, and 23 in the western region.
Due to changes in ownership, willingness to continue the collaboration, or both, repeat samples could not be collected in 1999 from 10 of the sites sampled in 1998. Thus, during the 1999 collection (December 16, 1998 to April 21, 1999), fecal samples were collected from 287 cows on 29 of the farms. Constraints with handling systems and turnover within the cow herds precluded sampling the same individual cows in both years.
The majority of the farms used wells as their source of water (34 sites), 2 sites were supplied with municipal water, and 3 farms used surface watering sites. During the spring sampling in 1998, the cows were maintained on pasture, whereas at the time of the 1999 collections, they were housed in confined lots and provided with preserved forage. The cows ranged from 2 to 14 y of age, but insufficient data from producers precluded calculation of a mean or median age.
Fecal samples (1/cow/y) were collected by a trained technician via rectal extraction at each farm. Approximate quantities (2 to 3 g) of fecal material were collected into preweighed, 15-mL screw-capped polypropylene tubes, then the tubes were reweighed to determine sample weight. After the addition of 5 mL of 10% (v/v) formalin, the tubes were capped and the contents were mixed thoroughly. Once a sufficient number of samples had been obtained, they were sent via courier to the University of Calgary for enumeration of Giardia cysts and Cryptosporidium oocysts.
Sample collection — southern British Columbia
During the 1998 calving season (March to May), 192 blood and fecal samples were collected from 10 different beef cattle ranches (13 to 20 animals per site) selected on the basis of their location within 4 different watersheds (Okanagan, Columbia, Thompson-Nicola, Kootenay) in the southeastern interior of British Columbia, and on the willingness of the producers to collaborate. Six of the sites were located in the Thompson-Nicola watershed, 2 in the Okanagan watershed, and 1 site in each of the Kootenay and Columbia watersheds.
Samples were collected by a team of veterinarians and animal technicians. All participants were provided with instructions to assure uniformity in data collection. Samples were collected from scouring calves (those exhibiting diarrhetic fecal production) and from nonscouring (producing feces of normal consistency) contemporaries at each site. An attempt was made at proportional representation of these groups to reflect the occurrence of scours within the entire herd at each site. For this study, diarrhetic feces was defined as material with a consistency loose enough to conform to the shape of the container into which it was collected. At the time of sampling, the age of the calf and its scouring status were recorded. The calves sampled ranged in age from 2 to 70 d (mean age 23.6 d, sχ̄ 1.1).
Fecal material from each calf (estimated total weight 4 to 6 g) was deposited into 2 preweighed collection tubes, which were then weighed again to determine sample weight. Five millilitres of 10% formalin solution was added to the 1st tube (2 to 3 g fecal material). The contents of the tube were then mixed and the tube shipped to the University of Calgary for enumeration of Giardia cysts and Cryptosporidium oocysts. The 2nd tube (remainder of fecal sample) was forwarded to a provincial laboratory in British Columbia and assessed for the presence of Salmonella, Campylobacter jejuni, Escherichia coli (F41, 987P, K88a, K88b or K99), rotavirus, and coronavirus.
The blood sample (10 mL) was collected from the jugular vein and serum was harvested by centrifugation at 500 × g for 5 min. Serum was frozen at −40°C until analyzed for calcium (Ca), phosphorus (P), magnesium (Mg), copper (Cu), iodine (I), selenium (Se), zinc (Zn), vitamin A, vitamin E, and IgG.
Animal care, sample collection and ethical review in Ontario and British Columbia were performed in accordance with the guidelines from the Canadian Council on Animal Care (21).
Sample analysis
Microscopic examination of Giardia and Cryptosporidium — Fecal suspensions were squeezed and rinsed through a surgical gauze sponge (Four ply, NuGauze; Johnson and Johnson, Montreal, Quebec) to yield 7 mL of filtrate. The filtrate was layered over 5 mL of 1 M sucrose (specific gravity 1.13) and centrifuged at 800 × g for 5 min to concentrate the cysts at the sucrose/water interface. The interface and upper layer of liquid were transferred by pipette to a clean tube and recentrifuged at 800 × g for 5 min. The supernatant was decanted, and the pellet was suspended in sodium phosphate buffered saline solution (PBSS) to a volume of 1 mL.
Two 150-μL samples of the concentrate were spotted onto microscope slides and allowed to air dry on a slide warmer for a minimum of 30 min. Once dry, the sample was fixed with acetone, dried, and mixed with 20 μL of either Giardia or Cryptosporidium fluorescein isothiocyanate (FITC) labelled monoclonal antibody solution (Giardi-a-glo, Crypt-o-glo; Waterborne, New Orleans, Louisiana, USA). The immunological reagents have been shown to be specific for Giardia spp. cysts and Cryptosporidium oocysts to the level of 98% (22,23) with a sensitivity of 67 cysts/oocysts per gram of feces. The sample was then incubated in a humidity chamber at 37°C for 30 min. After incubation, slides were rinsed with PBSS and the slide was allowed to air dry. Once dry, slides were mounted with glycerol (Aqua-polymount; Polysciences, Warrington, Pennsylvania, USA) and a cover slip, and the sample was examined by using an epifluorescent microscope at settings providing 200× and 400× magnification.
The number of cysts (Giardia) and oocysts (Cryptosporidium) over the spotted area were counted, which subsequently enabled the number of cysts or oocysts per gram of feces to be calculated. For samples collected from Ontario, oocysts of Cr. muris were differentiated from those of Cr. parvum based upon their size and shape (6.6 to 7.9 × 5.3 to 6.5 μm, ovoid for Cr. muris; 4.5 to 5.4 × 4.2 to 5.0 μm, spherical for Cr. parvum). This differentiation was not recorded for samples collected from calves in British Columbia. Oocysts and cysts from all samples were identified and enumerated by the same individual; previously established positive and negative control fecal samples were analyzed with each sample lot.
Bacteriology — Standard microbiologic methods were used to isolate and identify bacterial pathogens in fecal samples collected during the course of the study (24).
Isolates of E. coli were subcultured on E agar (25) containing glucose and citric acid as the only organic nutrients, and pilus antigens were typed by using a fluorescent antibody staining procedure (26). Colonies from E agar were suspended in PBSS and placed on microscope slides. A drop of heat-aggregated whole horse serum was added to the suspension on each slide and a smear of the bacterial preparation was created. Samples were air-dried for 30 min, fixed with acetone, and rinsed with PBSS. A solution of monoclonal antibodies for pilus antigens of F41, 987P, K88a, K88b, or K99 E. coli (Central Veterinary Laboratory, New Haw, Surrey, United Kingdom) was added to the samples, which were subsequently incubated in a humidity chamber at 37°C for 30 min. Slides were rinsed in PBSS; the samples were then incubated for 15 min with FITC-labeled rabbit-anti-mouse IgG, rinsed in distilled water, and air-dried. Coverslips were mounted with fluorescent mounting fluid and slides were examined at 400× magnification with an epifluorescence microscope.
Salmonella colonies were isolated by enrichment and identified by serological (Salmonella antisera; Difco Laboratories, Detroit, Michigan, USA) and standard biological techniques (27).
Campylobacter-like colonies were cultured on Blaser’s agar plates (28) at 42°C for 48 h in a microaerophilic atmosphere of 80% N2, 10% CO2, 5% O2, and 5% H2. Campylobacter-like colonies were Gram stained and examined for motility by phase-contrast microscopy. Organisms that showed corkscrew-like motility and were Gram-negative curved or spiral rods were further evaluated for their ability to hydrolyze hippurate (29).
Virology and serology — Rotavirus in fecal samples was identified by ELISA (Rotazyme II Diagnostic Kit; Abbott Laboratories, Abbott Park, Illinois, USA) and coronavirus was identified by using a standard procedure for negative staining and examination by electron microscopy (30). Radial immunodiffusion techniques (IgG SRID Kit; Veterinary Medical Research and Development, Pullman, Washington, USA) were performed to quantify serum IgG concentrations.
Serum levels of Ca, Mg, Cu, and Zn were measured by using flame atomic absorption, and P was determined by using a colorimetric kit (Diagnostic Chemicals, Charlottetown, Prince Edward Island). Selenium was determined as previously described (31). Iodine was measured by using a previously described adaptation of the Sandell-Kolthoff procedure (32) and determined by using the ferric-arsenic reduction procedure. Vitamins A and E were determined simultaneously by using a modified high-performance liquid chromatography method with dual detectors, based on the procedure of Dennison and Kirk (33).
Data analysis
Oocyst and cyst counts were log transformed and various mixed linear models (34) were used to perform an analysis of variance (ANOVA). Geometric means were calculated for positive animals within each year from each county and region, and multiple comparison of the means (least significant difference) was used to compare transformed cyst and oocyst counts. Categorical modeling of data (34) was used to control for clustering at the herd level and to compare the prevalence of parasitic infections in beef cows within regions and between the 2 sampling years. Prevalence was calculated as the number of infected individuals divided by the number of individuals sampled × 100. Spearman’s correlation was used to compare herd level summary statistics with the prevalence of parasites in calves from British Columbia.
Results
Ontario
Over the 2-year study, 669 fecal samples were collected from beef cows on farms in 10 counties in southern Ontario (Figure 1). Overall prevalences of Giardia, Cr. muris, and Cr. parvum in beef cows were 8.7%, 10.6%, and 18.4%, and these organisms were detected on 25 (64%), 28 (72%), and 35 (90%) of the 39 farms sampled, respectively. The 10 counties included in the study contained approximately 47% of Ontario’s beef cattle (35). Giardia duodenalis was identified in cows from all counties, with the exception of Victoria (Table 1). Within a county, the prevalence of Giardia was as high as 20%. Cyst counts varied among Giardia-infected cows, reaching as high as 3682 cysts/g of feces. Counts of Cr. parvum and Cr. muris oocysts in infected cows were as high as 12 323 and 175 343 per gram, respectively. In 1998, Cr. parvum was identified in all 10 counties (Table 1) and exhibited the highest prevalence of the parasites examined in all but 2 counties (Wellington and Dufferin). However, Cr. parvum was detected in only 7 of 10 counties in 1999. Prevalence across the 2 y ranged from 8.5% in Dufferin county to 38% in Bruce county. Cryptosporidium muris was also identified in all 10 counties, with prevalence over the 2 y ranging from 2% in Middlesex county to 18% in Wellington county.
Figure 1.

Prevalence of Giardia spp. and Cryptosporidium spp. infections among beef cattle in southern Ontario sampled in 1998 and 1999. Prevalences were computed as the number of samples that were positive for Giardia spp. or Cryptosporidium spp. divided by the number of samples collected within each region, multiplied by 100. Within each agricultural region, the unenclosed percentage is the prevalence of Giardia infection, the value in parentheses ( ) is the prevalence of Cr. parvum, and that in square brackets [ ] is the prevalence of Cr. muris. (n) = total number of samples collected in the region over the 2 y. Bar = 100 km.
Table 1.
Prevalence (%)a of Giardia, Cryptosporidium parvum, and Cryptosporidium muris in beef cattle in southern Ontario
| 1998 | 1999 | Both years | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Region | County | nb | Giardia | Cr. parvum | Cr. muris | n | Giardia | Cr. parvum | Cr. muris | n | Giardia | Cr. parvum | Cr. muris |
| South | Lambton | 59 | 13.6 | 20.3 | 5.1 | 29 | 0 | 6.9 | 17.2 | 88 | 9.1 | 15.9 | 9.1 |
| Middlesex | 29 | 10.3 | 41.4 | 3.5 | 20 | 0 | 5 | 0 | 49 | 6.1 | 26.5 | 2.1 | |
| Total | 88 | 12.5 | 27.3 | 4.6 | 49 | 0 | 6.1 | 10.2 | 137 | 8 | 19.7 | 6.6 | |
| West | Bruce | 30 | 10 | 56.7 | 13.3 | 20 | 0 | 10 | 5 | 50 | 6 | 38 | 10 |
| Dufferin | 56 | 12.5 | 12.5 | 16.1 | 50 | 8 | 4 | 2 | 106 | 10.4 | 8.5 | 9.4 | |
| Grey | 41 | 19.5 | 31.7 | 9.8 | 30 | 16.7 | 6.7 | 26.7 | 71 | 18.3 | 21.1 | 16.9 | |
| Huron | 30 | 3.3 | 53.5 | 30 | 30 | 3.3 | 16.7 | 3.3 | 60 | 3.3 | 35.1 | 16.7 | |
| Simcoe | 40 | 15 | 30 | 5 | 38 | 7.9 | 0 | 5.3 | 78 | 11.5 | 15.4 | 5.1 | |
| Wellington | 30 | 10 | 20 | 23.3 | 10 | 0 | 0 | 0 | 40 | 7.5 | 15 | 17.5 | |
| Total | 227 | 12.3 | 31.3 | 15.4 | 178 | 7.3 | 6.2 | 7.3 | 405 | 10.1 | 20.3 | 11.9 | |
| Central | Peterborough | 38 | 15.8 | 18.4 | 10.5 | 30 | 0 | 0 | 6.7 | 68 | 8.8 | 10.3 | 8.8 |
| Victoria | 29 | 0 | 20.7 | 13.8 | 30 | 0 | 3.3 | 10 | 59 | 0 | 11.9 | 11.9 | |
| Total | 67 | 9 | 19.4 | 11.9 | 60 | 0 | 1.7 | 8.4 | 127 | 4.7 | 11 | 10.2 | |
| Overall | 382 | 11.8 | 28.3 | 12.3 | 287 | 4.5 | 5.2 | 8 | 669 | 8.7 | 18.4 | 10.5 | |
aCalculated within each county as number of infected cows/number of cows sampled × 100%
bn = total number of cattle sampled in a given county
Overall prevalence of Giardia in cows was higher (P < 0.05) in 1998 (11.8%) than in 1999 (4.5%, Table 1). In 1998, prevalence of Giardia was fairly consistent across regions, but in 1999, it was not detected in either the southern or the central regions. Cryptosporidium parvum was detected in 28% of the samples in 1998, but in only 5.2% (P < 0.05) of the samples in 1999. In 1998, prevalence of Cr. muris was higher (P < 0.05) in the western and central regions than in the south. Its overall prevalence that year was 12.3%. As observed with the other 2 parasites, the overall prevalence of Cr. muris was lower in 1999 than in 1998. Average fecal counts of Giardia cysts and Cr. muris oocysts in positive samples were higher (P < 0.05) in 1998 (85.9 cysts/g and 230.2 oocysts/g) than in 1999 (49.7 cysts/g and 73.5 oocysts/g). Cryptosporidium parvum counts did not differ (P > 0.05) between the 2 y.
Of the samples collected, 511 were from cows provided with well water, 129 were from cows provided with surface water, and only 29 were supplied with municipal water. Giardia cysts and Cryptosporidium oocysts were detected in cattle regardless of water source (Table 2). In fact, in 1998, prevalence of Giardia cysts was numerically higher in cows with access to municipal water as compared with well or surface water. However, in 1999, Giardia was not detected in cows with access to municipal water.
Table 2.
Prevalence (%) of Giardia and Cryptosporidium spp. in beef cattle grouped by source of drinking water
| 1998 | 1999 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Water source | Number of farms | Number of cattle | Giardia | Cr. parvum | Cr. muris | Number of farms | Number of cattle | Giardia | Cr. parvum | Cr. muris |
| Municipal water | 2 | 19 | 31.6 | 31.6 | 10.5 | 1 | 10 | 0 | 10 | 0 |
| Well water | 30 | 294 | 10.2 | 26.2 | 13.6 | 22 | 217 | 5.1 | 3.2 | 9.7 |
| Surface water | 7 | 69 | 13 | 36.2 | 7.2 | 6 | 60 | 3.3 | 11.7 | 3.3 |
| Total | 39 | 382 | 11.8 | 28.3 | 12.3 | 29 | 287 | 4.5 | 5.2 | 8 |
British Columbia
A total of 192 fecal samples were collected from calves representing the 4 watersheds in southeastern British Columbia (Figure 2, Table 3). Overall prevalences of Giardia (36%) and Ca. jejuni (42%) were higher (P < 0.001) than that of Cryptosporidium (13%). Giardia and Ca. jejuni were detected on all farms, except that Ca. jejuni was not detected at Vernon site B, whereas Cryptosporidium was detected on only 50% of the sites examined. Salmonella and E. coli K99 were not detected in any of the calves that were sampled and E. coli K88a was detected in only 1 calf. Coronavirus and rotavirus were present at all sites, with the exception of Kamloops. Prevalence of G. duodenalis was as high as 70% at site G in the Thompson watershed. Although not statistically significant, this same site also exhibited the highest prevalence of scours and the greatest prevalence of both Cryptosporidium and Campylobacter. Giardia cyst counts in infected calves were as high as 113 000 cysts/g of feces, whereas Cryptosporidium counts were as high as 132 000 oocysts/g.
Figure 2.
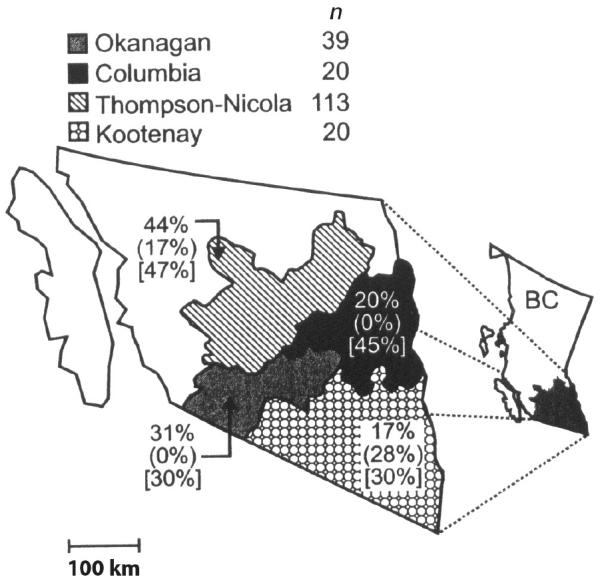
Prevalence of Giardia spp., Cryptosporidium spp., and Campylobacter jejuni infections among beef calves in southeastern British Columbia. Prevalences were computed as the number of calves infected with Giardia spp., Cryptosporidium spp. or Ca. jejuni divided by the number of calves sampled within each region, multiplied by 100. For each region, the unenclosed percentage is the prevalence of Giardia infection, the value in parentheses ( ) is the prevalence of Cryptosporidium infection, and that in square brackets [ ] is the prevalence of Campylobacter infection. (n) = total number of calves sampled in each region. Bar = 100 km.
Table 3.
Prevalence (%) of enteropathogens and scours in beef calves in southern British Columbia in 1998
| Watershed | Location | Site | na | Ageb | Giardia | Crypto | Salmonella | E. coli | Campylo | Rotavirus | Corona | Scours |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Columbia | Invermere | J | 20 | 20.6 | 20 | nd | nd | nd | 45 | 5.3 | 21.1 | 50 |
| Kootenay | Creston | F | 20 | 22.4 | 16.7 | 27.8 | nd | nd | 30 | 7.1 | 14.3 | 10 |
| Okanagan | Salmon Arm | A | 20 | 20.2 | 40 | nd | nd | nd | 60 | 30 | 60 | na |
| Vernon | B | 19 | 12.9 | 21.1 | nd | nd | nd | nd | 31.6 | 15.8 | 47.4 | |
| Average | 19.5 | 16.6 | 30.7 | nd | nd | nd | 30 | 30.8 | 38.5 | — | ||
| Thompson | Kamloops | C | 13 | 21.1 | 25 | nd | nd | nd | 38.5 | nd | nd | 30.8 |
| D | 20 | 30.3 | 42.1 | 10.5 | nd | nd | 30 | nd | 36.8 | 30 | ||
| E | 20 | 23.5 | 47.4 | nd | nd | nd | 45 | 7.7 | 53.9 | 5 | ||
| I | 20 | 10.3 | 42.1 | 26.3 | nd | nd | 25 | 8.3 | 25 | 20 | ||
| Merrit | G | 20 | 43.4 | 70 | 45 | nd | 5 | 70 | 20 | 40 | 80 | |
| H | 20 | 19.8 | 25 | 10 | nd | nd | 70 | 16.7 | 55 | 50 | ||
| Average | 18.8 | 27.3 | 43.8 | 17.1 | nd | 0.9 | 46.9 | 10.5 | 36.8 | 36.2 | ||
| Overall | 192 | 23.6 | 35.7c | 12.6d | nd | 0.5e | 41.7c | 14.9 | 33.1 | 36 |
Crypto — Cryptosporidium; Campylo — Campylobacter jejuni; E. coli — Escherichia coli K88; Corona — coronavirus; nd — not detected; na — not assessed
aNumber of calves sampled per site; not all enteropathogens were surveyed in each sample (insufficient fecal material)
bMean age (days)
cdeWithin a row, values followed by different letters differ significantly (P < 0.001)
Serum mineral, vitamin, and IgG levels were within normal range when averaged within a herd (Table 4; 36,37). Of note, however, is that although the association was not statistically significant, calves with scours exhibited markedly lower levels of serum Se and depressed levels of IgG than did those with normal fecal consistency (data not shown). This pattern was especially notable in the Merritt herd G, in which prevalence of scours was highest and levels of serum Se and IgG were lowest.
Table 4.
Serum mineral, vitamin, and immunoglobulin (Ig) G status of calves sampled for fecal enteropathogens in southern British Columbia
| Serum levels | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Watershed | Location | Site | n | Ca mmol/L | P mmol/L | Mg mmol/L | Cu μmol/L | I μmol/L | Se μmol/L | Zn μmol/L | Vit A μmol/L | Vit E μmol/L | IgG g/L |
| Columbia | Invermere | J | 20 | 2.71, s = 0.164 | 4.29, s = 1.159 | 0.87, s = 0.103 | 8.75, s = 1.613 | 0.38, s = 0.115 | 0.71, s = 0.223 | 16.10, s = 2.646 | 0.67, s = 0.204 | 3.79, s = 2.442 | 27.83, s = 5.301 |
| Kootenay | Creston | F | 20 | 2.39, s = 0.142 | 2.93, s = 0.332 | 0.66, s = 0.077 | 7.52, s = 1.165 | 1.57, s = 0.653 | 0.51, s = 0.111 | 14.20, s = 3.342 | 0.64, s = 0.192 | 1.54, s = 0.986 | 31.61, s = 7.959 |
| Okanagan | Salmon Arm | A | 20 | 2.63, s = 0.172 | 3.31, s = 0.395 | 0.73, s = 0.096 | 10.32, s = 2.156 | 0.81, s = 0.211 | 0.79, s = 0.091 | 17.32, s = 4.220 | 0.59, s = 0.136 | 4.21, s = 1.722 | 21.60, s = 3.693 |
| Vernon | B | 19 | 2.79, s = 0.243 | 3.36, s = 0.568 | 0.83, s = 0.089 | 10.40, s = 2.770 | 1.25, s = 0.540 | 0.94, s = 0.205 | 17.37, s = 3.532 | 0.61, s = 0.195 | 1.50, s = 1.250 | 25.34, s = 6.310 | |
| Average | 19.5 | 2.71, s = 0.207 | 3.34, s = 0.482 | 0.78, s = 0.093 | 10.36, s = 2.463 | 1.03, s = 0.375 | 0.86, s = 0.148 | 17.34, s = 3.876 | 0.60, s = 0.166 | 2.86, s = 1.486 | 23.45, s = 5.002 | ||
| Thompson | Kamloops | C | 13 | 2.65, s = 0.254 | 3.57, s = 0.340 | 0.83, s = 0.111 | 9.99, s = 3.714 | 1.15, s = 1.038 | 0.91, s = 0.409 | 16.47, s = 2.753 | 0.80, s = 0.456 | 3.45, s = 2.113 | 24.06, s = 4.006 |
| D | 20 | 2.49, s = 0.130 | 3.28, s = 0.736 | 0.76, s = 0.081 | 7.70, s = 2.109 | 1.06, s = 0.715 | 0.75, s = 0.101 | 16.96, s = 2.767 | 0.65, s = 0.195 | 1.66, s = 1.194 | 28.40, s = 8.192 | ||
| E | 20 | 2.54, s = 0.210 | 3.33, s = 0.361 | 0.70, s = 0.067 | 7.95, s = 2.109 | 1.31, s = 0.612 | 0.87, s = 0.480 | 16.50, s = 3.302 | 0.71, s = 0.192 | 1.79, s = 0.772 | 26.62, s = 8.155 | ||
| I | 20 | 2.40, s = 0.146 | 3.40, s = 0.390 | 0.65, s = 0.165 | 9.73, s = 3.218 | 1.28, s = 1.252 | 0.77, s = 0.572 | 15.82, s = 3.336 | 0.52, s = 0.170 | 2.38, s = 1.049 | 23.10, s = 4.512 | ||
| Merritt | G | 20 | 2.50, s = 0.103 | 3.11, s = 0.253 | 0.55, s = 0.139 | 8.84, s = 2.408 | 4.76, s = 3.792 | 0.38, s = 0.080 | 13.19, s = 4.160 | 0.62, s = 0.171 | 2.73, s = 1.311 | 19.20, s = 3.247 | |
| H | 20 | 2.72, s = 0.208 | 3.56, s = 0.755 | 0.85, s = 0.112 | 11.74, s = 3.478 | 1.02, s = 0.557 | 0.62, s = 0.139 | 15.18, s = 3.119 | 0.74, s = 0.238 | 1.88, s = 1.160 | 26.67, s = 6.244 | ||
| Average | 18.8 | 2.55, s = 0.175 | 3.37, s = 0.472 | 0.72, s = 0.113 | 9.32, s = 2.839 | 1.76, s = 1.328 | 0.72, s = 0.297 | 15.68, s = 5.437 | 0.67, s = 0.237 | 2.31, s = 1.267 | 24.68, s = 5.726 | ||
| Overall average | 2.58, s = 0.177 | 3.41, s = 0.529 | 0.74, s = 0.104 | 9.29, s = 2.474 | 1.46, s = 0.948 | 0.72, s = 0.241 | 15.91, s = 4.636 | 0.65, s = 0.215 | 2.49, s = 1.400 | 25.44, s = 5.762 | |||
| Normal levels (36,37) | 2.0 to 2.7 | 1.9 to 2.9 | 0.7 to 1.2 | 9.4 to 23.6 | 0.79 to 3.15 | 1.01 to 3.80 | 12.2 to 21.4 | 0.7 to 1.1 | 1.9 to 9.3 | 14 to 25 | |||
| Deficient levels (36,37) | < 1.5 | < 1.6 | < 0.15 | < 4.7 | < 0.4 | < 0.32 | < 6.1 | < 0.18 | < 0.2 | < 14 | |||
Prevalence of Giardia and Ca. jejuni was higher (P < 0.001) in 2- to 6-week-old calves than in 0- to 2-week-old calves (data not shown). In contrast, prevalence of Cryptosporidium was relatively constant in 0- to 6-week-old calves but notably higher (P < 0.001) in calves that were over 6 wk old.
Discussion
Few studies have examined the prevalence of Giardia and Cryptosporidium in beef cows; the majority of studies have focused on dairy cattle (1,2,17,23,24,38). Giardia was not detected when fecal samples were collected on a single occasion from 26 adult beef cows from 3 farms in Alberta (39). In another study (20), when fecal samples were collected from 20 beef cows 6 times over 25 wk, the prevalences of Giardia and Cr. andersoni among the cows ranged from 0% to 15% and 0% to 40%, respectively. Work with a single cowherd indicated that Giardia is more prevalent in calves than in cows (20). In our study, the prevalence of Giardia in beef cows on 10 different farms ranged from 0% to 20%. It has been well documented that Giardia cysts are shed intermittently in cattle (17). Consequently, the low prevalence of Giardia reported in the literature for beef cows may be associated more with the logistics of taking multiple samples from these animals under range conditions than with the actual occurrence of the infection.
As with Giardia, Cryptosporidium tends to be more prevalent in younger than older animals (40). When detection methods similar to ours are used, Cryptosporidium has generally been found to be less prevalent than Giardia in dairy calves (1,2). In the present study, Cr. parvum was more prevalent than Giardia in cows, and 90% of the farms had at least 1 cow that tested positive for Cr. parvum. Similarly, at least 1 calf was positive for a Cryptosporidium sp. on 89% of dairy farms (n = 600) examined in Quebec (16). In the present study, Cr. muris was also more prevalent than Giardia in beef cows, but, somewhat surprisingly, was not as prevalent as Cr. parvum. Cryptosporidium muris tends to reside in the abomasum (41) and it has been proposed that this species is far more common than Cr. parvum in adult cattle (42,43). The present study demonstrates that it is possible for Cr. parvum to be more prevalent than Cr. muris in beef cows. This has important implications, as clinical cryptosporidial infections in humans are caused by Cr. parvum (44). There has been only 1 reported case of Cr. muris infection in humans (44).
The decline in the prevalence of Giardia and Cryptosporidum spp. in 1999 as compared with 1998 may be the result of a number of factors. The sampling in 1999 occurred between the months of December and April, whereas the sampling in 1998 took place between March and May. Cattle were penned during the winter of 1999 but were pastured with their calves in the spring of 1998. Previous work has shown that adult cattle from herds with a high number of young calves are more likely to shed Cr. parvum oocysts than are those from herds that possess a lower number of juvenile animals (40,45). In 1998, newly born juvenile carriers in the herd may have served as a source of infection among adult animals.
It is also possible that a periparturient rise in the excretion of Giardia cysts and Crytosporidium oocysts during spring calving was responsible for the higher detection of these parasites in cows in 1998 compared with 1999. Such a scenario has been associated with the early infection of lambs (19) and calves (6) with Giardia. However, periparturient dairy cows were not identified as the source of infection for calves with Cr. parvum; rather, oocysts in the bedding and pen floor were implicated as the source of the infection (46). Others have also found Cryptosporidium to be more prevalent in beef calves in the spring than in the winter (2,18), a trend that agrees with our work in beef cows. Transmission between animals in the winter may be reduced due to the freezing of animal feces and a corresponding reduction in fecal-oral transmission of Cryptosporidium oocysts and Giardia cysts. Cryptosporidium oocysts can remain viable after freezing (47,48) and, consequently, freshly thawed feces in the spring may be a source of infection for herd members.
In the present study, provision of municipal water did not eliminate Giardia or Cryptosporidium spp. from the small number of herds that were provided with this source of water. Giardia and Cryptosporidium have been detected in dairy calves with access to both well and municipal water (1). Chlorine-based disinfectants are largely ineffective against Cryptosporidium (49) and their effectiveness against Giardia cysts declines at lower temperatures (50). Water treatment does not prevent fecal-oral contact, an event that is likely responsible for the majority of the transmission of these parasites among herd members. Consumption of water from field ponds during spring run off could also negate any benefit that water treatment has on reducing water-borne transmission of these parasites.
Most studies on the prevalence of Giardia and Cryptosporidium have been conducted on dairy calves (1,16,23,46). In the present study, the prevalence of infection with Giardia among range calves was lower than previously reported in dairy calves (1,16). Prevalence of Giardia has been shown to be greater in calves housed indoors than in those outdoors (16,23). The antihelminthic agent, fenbendazole, controls Giardia in dairy calves, but under confined conditions, these animals readily become reinfected if treatment is discontinued (51). In the natural environment, Giardia cysts are subject to desiccation and are rendered noninfective after prolonged exposure to temperatures below freezing (48). Thus, reduced confinement and the accompanying lower animal density, as well as a reduction in the survival of infective cysts in the exposed environment, may account for the reduced infection level in range calves compared with dairy calves.
Prevalence of scours was highest among calves for site G at Merritt in the Thompson watershed. There were individual calves in that herd with serum Se and IgG levels that were below normal (data not shown), a factor that contributed to this site’s low herd average of these parameters (Table 4). Prevalences of Giardia and Campylobacter were also highest in the site G herd, a finding that supports the notion that Giardia could play a role in calf scours (17). Low serum IgG concentrations at 24 h of age have been closely correlated with the prevalence of scours in calves from birth to 9 or 16 wk of age (24,52) and, in those studies, numerous enteropathogens were isolated from calf feces. The present study supports the hypothesis that passive immunity does not affect the level of infection of Cryptosporidium (52,53).
The overall prevalence of Giardia and Cryptosporidium among groups of cattle penned or pastured together is heavily influenced by the time of year at which samples are collected. Prevalence of these pathogens in cows is highest during the calving season and management strategies targeted at reducing environmental contamination are likely to be particularly critical at this point in the cow-calf cycle. Although the prevalance of Giardia and Cryptosporidium was lower in range calves than in dairy calves, there was evidence that calves exhibiting scours also were likely to harbor Giardia. Low Se levels suggests that management strategies that enhance the nutritional status of calves may also reduce the prevalence of these enteropathogens.
The public health significance of G. duodenalis and Cr. parvum isolated from cattle has been investigated with the use of polymerase chain reaction-based molecular tools (54–56). In the present study, these organisms could not be genotyped, because the formalin fixation precluded isolation of DNA from the cysts and oocysts. In North America and Australia, most human cases of cryptosporidiosis appear to be of human origin (Cryptosporidium hominis or Cr. parvum genotype 1), which are morphologically indistinguishable from the common mammalian species, Cr. parvum genotype 2 (54,57). In the United States, cattle have not been identified as the source of any waterborne outbreak of Cr. parvum in humans and in Canada, only one outbreak (in Cranbrook, British Columbia) has been attributed to the bovine Cr. parvum genotype (54). Because the specific Cr. parvum genotype could not be identified in this study, it is difficult to assess the zoonotic significance of the high prevalence of this parasite in beef cows. Giardia is also very common in calves and cows, but recent molecular analysis of cysts from a variety of animals showed that more than 95% of the samples contained cysts that were of the nonzoonotic livestock genotype (55,56).
Acknowledgments
The authors thank C. Cockwill, N. Guselle, J. Greenfield, G. Gourlay, and B. Puls for their assistance with these studies. They also thank all of the producers in Ontario and British Columbia who provided us with access to their cattle. CVJ
Footnotes
This is Lethbridge Research Centre contribution number 38703078.
References
- 1.Olson ME, Guselle NJ, O’Handley RM, et al. Giardia and Cryptosporidium in dairy calves in British Columbia. Can Vet J. 1997;38:703–706. [PMC free article] [PubMed] [Google Scholar]
- 2.Xiao LH, Herd RP, Rings DM. Concurrent infections of Giardia and Cryptosporidium on two Ohio farms with calf diarrhea. Vet Parasitol. 1993;51:41–48. doi: 10.1016/0304-4017(93)90194-R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.O’Donoghue PJ. Cryptosporidium and Cryptosporidiosis in man and animals. Int J Parasitol. 1995;25:139–155. doi: 10.1016/0020-7519(94)e0059-v. [DOI] [PubMed] [Google Scholar]
- 4.Erlandsen SL. Biotic transmission — Is giardiasis a zoonosis? In: Thompson RCA, Reynoldson JA, Lymbery AJ, eds. Giardia: From Molecules to Disease. Cambridge, UK: Cambridge Univ Pr, 1994:83–97.
- 5.Olson ME, McAllister TA, Deselliers L, et al. Effects of giardiasis on production in a domestic ruminant (lamb) model. Am J Vet Res. 1995;56:1470–1474. [PubMed] [Google Scholar]
- 6.Ralston BJ, Cockwill CL, Guselle NJ, Van Herk FH, McAllister TA, Olson ME. Prevalence of Giardia and Cryptosporidium andersoni and their effects on performance in feedlot beef cattle. Can J Anim Sci. 2003;83:153–159. [Google Scholar]
- 7.Fayer R, Speer C, Dubey J. The general biology of Cryptosporidium. In: Fayer R, ed. Cryptosporidium and Cryptosporidiosis. Boca Raton: CRC Pr, 1997:1–41.
- 8.Olson ME, Ralston BJ, O’Handley R, Guselle NJ, Applebee AJ. What is the clinical and zoonotic significance of Cryptosporidiosis in domestic animals and wildlife. In: Thompson RCA, Armson A, Ryan UM, eds. Cryptosporidium: From Molecules to Disease. Elsevier Sci, 2004 (In press).
- 9.Majewska, AC Successful experimental infections of a human volunteer and Mongolian gerbils with Giardia of animal origin. Trans R Soc Trop Med Hyg. 1993;88:360–362. doi: 10.1016/0035-9203(94)90119-8. [DOI] [PubMed] [Google Scholar]
- 10.Reif, JS, Wimmer L, Smith JA, Dargatz DA, Cheney JM. Human cryptosporidiosis associated with an epizootic in calves. Am J Public Health. 1989;79:1528–1530. doi: 10.2105/ajph.79.11.1528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Egger M, Nguyen XM, Schaad UB, Krech T. Intestinal cryptosporidiosis acquired from a cat. Infection. 1990;18:177–188. doi: 10.1007/BF01642109. [DOI] [PubMed] [Google Scholar]
- 12.Gradus MS. Water quality and waterborne protozoa. Clin Microbiol News. 1989;11:121–125. [Google Scholar]
- 13.LeChevallier MW, Norton WD, Lee RG. Occurrence of Giardia and Cryptosporidium spp. in surface water supplies. Appl Environ Microbiol. 1991;57:2610–2616. doi: 10.1128/aem.57.9.2610-2616.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ong C, Moorehead W, Ross A, Isaac-Renton J. Studies of Giardia spp. and Cryptosporidium spp. in two adjacent watersheds. Appl Environ Microbiol. 1996;62:2798–2805. doi: 10.1128/aem.62.8.2798-2805.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Garber LP, Salman MD, Hurd HS, Keefe T, Schlater JL. Potential risk factors for Cryptosporidium infection in dairy calves. J Am Vet Med Assoc. 1994;205:86–91. [PubMed] [Google Scholar]
- 16.Ruest N, Faubert GM, Couture Y. Prevalence and geographical distribution of Giardia spp. and Cryptosporidium spp. in dairy farms in Quebec. Can Vet J. 1998;39:697–700. [PMC free article] [PubMed] [Google Scholar]
- 17.O’Handley RM, Olson ME, McAllister TA, et al. Efficacy of fenbendazole for treatment of giardiasis in calves. Am J Vet Res. 1997;58:384–388. [PubMed] [Google Scholar]
- 18.Mann ED, Sekla LH, Nayer GPS, Koschik C. Infection with Cryptosporidium spp. in humans and cattle in Manitoba. Can J Vet Res. 1986;50:174–178. [PMC free article] [PubMed] [Google Scholar]
- 19.Xiao L, Herd RP, McClure KE. Periparturient rise in the excretion of Giardia sp. cysts and Cryptosporidium parvum oocysts as a source of infection for lambs. J Parasitol. 1994;80:55–59. [PubMed] [Google Scholar]
- 20.Ralston BJ, McAllister TA, Olson ME. Prevalence and infection pattern of naturally acquired giardiasis and cryptosporidiosis in range beef calves and their dams. Vet Parasitol. 2003;114:113–122. doi: 10.1016/s0304-4017(03)00134-1. [DOI] [PubMed] [Google Scholar]
- 21.Olfert EE, Cross BM, McWilliams AA, eds. Guide to the Care and Use of Experimental Animals. 2nd ed. vol. 1. Ottawa, Ontario: Can Council Anim Care, 1994.
- 22.Olson ME, Thorlakson CL, Deselliers L, Morck DW, McAllister TA. Giardia and Cryptosporidium in Canadian farm animals. Vet Parasitol. 1997;68:375–381. doi: 10.1016/s0304-4017(96)01072-2. [DOI] [PubMed] [Google Scholar]
- 23.Xiao L, Herd RP. Infection patterns of Cryptosporidium and Giardia in calves. Vet Parasitol. 1994;55:257–262. doi: 10.1016/0304-4017(93)00645-f. [DOI] [PubMed] [Google Scholar]
- 24.McDonough SP, Stull CL, Osburn BI. Enteric pathogens in intensively reared veal calves. Am J Vet Res. 1994;55:1516–1520. [PubMed] [Google Scholar]
- 25.Francis DH, Remmers GA, DeZeeuw PS. Production of K88, K99 and 987P antigens by Escherichia coli culture on synthetic and complex media. J Clin Microbiol. 1982;15:181–183. doi: 10.1128/jcm.15.1.181-183.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mullaney CD, Francis DH, Willgohs JA. Comparison of seroaggultination, ELISA and indirect fluorescent antibody staining detection of K99, K88 and 987P pilus antigens of Escherichia coli. J Vet Diagn Invest. 1991;3:115–119. doi: 10.1177/104063879100300201. [DOI] [PubMed] [Google Scholar]
- 27.Jang SS, Biberstein EL, Hirsh DC. A Diagnostic Manual of Veterinary and Clinical Bacteriology and Mycology. Davis, California: Univ California Pr, 1992:51–60.
- 28.Atlas, RM. Handbook of Microbiological Media, 2nd ed. Boca Raton: CRC Pr 1996:243–244.
- 29.Skirrow MB, Benjamin J. Campylobacters: cultural characteristics of intestinal campylobacters from man and animals. J Hyg Camb. 1980;85:427–442. doi: 10.1017/s0022172400063506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Isenberg H. Direct specimen testing: viral and chlamydial infections. In: Isenberg H, ed. Essential Procedures for Clinical Microbiology. Washington, DC: ASM Pr, 1998:533–550.
- 31.Horwitz W, ed. Official Methods of Analysis, procedure 25. 126. Washington DC: Assoc Analytical Chemists, 1980:1018 pp.
- 32.Dunn JT, Crutchfield HE, Gutekunst R, Dunn AD. Two simple methods for measuring iodine in urine. Thyroid. 1993;3:119–123. doi: 10.1089/thy.1993.3.119. [DOI] [PubMed] [Google Scholar]
- 33.Dennison DB, Kirk JR. Quantitative analysis of vitamin A in cereal products by high-performance liquid chromatography. J Food Prot. 1979;42:1376–1384. [Google Scholar]
- 34.Statistical Analysis Systems. SAS/STAT User’s Guide, Version 8. Cary, North Carolina: SAS Institute.1999.
- 35.Ontario Ministry of Agriculture and Food (homepage on the internet). Guelph, Queen’s Printer for Ontario; c2004 [last updated June 15, 2004, cited July 5, 2004]. Statistical Tables, Livestock by Country, Cattle 1998 and Cattle 1999 [about 2 screens]. Available from http://www.gov.on.ca/OMAFRA/english/stats/livestock/index.html
- 36.Puls R. Mineral Levels in Animal Health: Diagnostic Data, 2nd ed. Clearbrook British Columbia: Sherpa Int, 1994.
- 37.Puls R. Vitamin Levels in Animal Health: Diagnostic Data and Bibliographies. Clearbrook, British Columbia: Sherpa Int, 1994.
- 38.Fuente R, Luzon M, Ruiz-Santa-Quiteria JA, et al. Cryptosporidium and concurrent infections with other major enteropathogens in 1- to 30-day-old diarrheic calves in central Spain. Vet Parasitol. 1999;80:179–185. doi: 10.1016/S0304-4017(98)00218-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Buret A, denHollander N, Wallis PM, Befus D, Olson ME. Zoonotic potential of giardiasis in domestic ruminants. J Infect Dis. 1990;162:231–237. doi: 10.1093/infdis/162.1.231. [DOI] [PubMed] [Google Scholar]
- 40.Atwill ER, Johnson EM, das Gracas M, Pereira C. Association of herd composition, stocking rate and duration of calving season with fecal shedding of Cryptosporidium parvum oocysts in beef herds. J Am Vet Med Assoc. 1999;215:1833–1838. [PubMed] [Google Scholar]
- 41.Anderson BC. Abomasal cryptosporidiosis in cattle. Vet Pathol. 1987;24:235–239. doi: 10.1177/030098588702400307. [DOI] [PubMed] [Google Scholar]
- 42.Anderson BC. Prevalence of Cryptosporidium muris-like oocysts among cattle populations of the United States: preliminary report. J Protozool. 1991;38:148–152. [PubMed] [Google Scholar]
- 43.Bukhari Z, Smith HV. Detection of Cryptosporidium muris oocysts in the faeces of adult dairy cattle in Scotland. Vet Rec. 1996;138:207–208. doi: 10.1136/vr.138.9.207. [DOI] [PubMed] [Google Scholar]
- 44.Casemore DP, Wright SE, Coop RL. Cryptosporidiosis — human and animal epidemiology. In: Fayer R, ed. Cryptosporidium and Cryptosporidiosis. Boca Raton: CRC Pr 1997:65–92.
- 45.Atwill ER, Johnson EM, Klingborg DJ. Age, geographic, and temporal distribution of fecal shedding of Cryptosporidium parvum oocysts in cow-calf herds. Am J Vet Res. 1999;60:420–425. [PubMed] [Google Scholar]
- 46.Atwill ER, Harp JA, Jones T, Jardon PW, Checel S, Zylstra M. Evaluation of periparturient dairy cows and contact surfaces as a reservoir of Cryptosporidium parvum for calfhood infection. Am J Vet Res. 1998;59:1116–1121. [PubMed] [Google Scholar]
- 47.Fayer R, Nerad T. Effects of low temperature on viability of Cryptosporidium parvum oocysts. Appl Environ Microbiol. 1996;62:1431–1433. doi: 10.1128/aem.62.4.1431-1433.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Olson ME, Goh J, Phillips M, Guselle N, McAllister TA. Giardia cyst and Cryptosporidium oocyst survival in water, soil and cattle feces. J Environ Qual. 1999;28:1991–1996. [Google Scholar]
- 49.Rose JB, John JT, LeChevallier M. Waterborne Cryptosporidiosis: incidence, outbreaks and treatment strategies. In: Fayer R, ed. Cryptosporidium and Cryptosporidiosis. Boca Raton: CRC Pr 1997:93–109.
- 50.Jarroll EL, Bingham AK, Meyer EA. Giardia cyst destruction: effectiveness of six small-quantity water disinfection methods. Am J Trop Med Hyg. 1980;20:8–11. doi: 10.4269/ajtmh.1980.29.8. [DOI] [PubMed] [Google Scholar]
- 51.O’Handley RM, Cockwill C, Jelinski M, McAllister TA, Olson ME. Effects of repeat fenbendazole treatment in dairy calves with giardiosis on cyst excretion, clinical signs and production. Vet Parasitol. 1999;89:209–218. doi: 10.1016/s0304-4017(00)00200-4. [DOI] [PubMed] [Google Scholar]
- 52.Quigley JD, Martin KR, Bemis DA, et al. Effects of housing and colostrum feeding on the prevalence of selected infectious organisms in feces of Jersey calves. J Dairy Sci. 1994;7:3124–3131. doi: 10.3168/jds.S0022-0302(94)77255-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Harp JA, Woodmansee DB, Moon HW. Effects of colostral antibody on susceptibility of calves to Cryptosporidium parvum infection. Am J Vet Res. 1989;50:2117–2219. [PubMed] [Google Scholar]
- 54.Fayer R, Morgan U, Upton SJ. Epidemiology of Cryptosporidium: transmission, detection and identification. Int J Parasitol. 2000;30:1305–1322. doi: 10.1016/s0020-7519(00)00135-1. [DOI] [PubMed] [Google Scholar]
- 55.O’Handley RM, Olson ME, Fraser D, Adams P, Thompson RCA. Prevalence and genotypic characterisation of Giardia in dairy calves from Western Australia and Western Canada. Vet Parasitol. 2000;90:193–200. doi: 10.1016/s0304-4017(00)00235-1. [DOI] [PubMed] [Google Scholar]
- 56.Appelbee AJ, Frederick LM, Heitman TL, Olson ME. Prevalence and genotyping of Giardia duodenalis from beef calves in Alberta, Canada. Vet Parasitol. 2003;112:289–294. doi: 10.1016/s0304-4017(02)00422-3. [DOI] [PubMed] [Google Scholar]
- 57.Morgan-Ryan UM, Fall A, Ward LA, et al. Cryptosporidium hominis n. sp. (Apicomplexa: Cryptosporidiidae) from Homo sapiens. J Eukaryot Microbiol. 2002;49:433–440. doi: 10.1111/j.1550-7408.2002.tb00224.x. [DOI] [PubMed] [Google Scholar]