

Abstract
Bupleurumjeholense Nakai (Apioideae, Apiaceae), originally found in the Wuling Mountain of China, was initially described as a species but later treated as a variety of B.sibiricum Vest ex Spreng. However, researchers have recently found that it is more closely related to B.chinense DC. In this study, we conducted morphological and phylogenetic analyses as well as chromosome counting to determine the taxonomic status of B.sibiricumvar.jeholense (Nakai) Chu. Our results showed that B.sibiricumvar.jeholense and B.chinense share common features (i.e., bracteoles 5 and stem solid) that distinguish both from B.sibiricumvar.sibiricum. The chromosome number of B.sibiricumvar.jeholense was found to be the same as in B.chinense (i.e., 2n = 12), whereas the chromosome number of B.sibiricumvar.sibiricum was 64. A phylogenetic tree based on complete chloroplast genome data revealed a close relationship between B.sibiricumvar.jeholense and B.chinense. Finally, B.sibiricumvar.jeholense and B.chinense were mainly found to differ in plant height, number of stems, and middle stem leaves. Based on this evidence, we propose a new combination: Bupleurumchinensevar.jeholense (Nakai) Q.R.Liu & L.H.Wang.
Key words: Apiaceae, Bupleurumchinense DC. var. jeholense (Nakai) Q.R.Liu & L.H.Wang, new combination
Introduction
Bupleurum is a monophyletic genus in the tribe Bupleureae (Apioideae, Apiaceae) (Downie et al. 2000). It contains around 180–195 species, is distributed throughout Eurasia and North Africa, with one species each in North America and South Africa, while being adventive in Australasia (Plunkett et al. 2019). China is a major diversity center of Bupleurum, containing 42 species and 16 varieties (Sheh and Watson 2005). This genus can be easily recognized based on its entire and single leaves, which usually show parallel venation and distinct bracts and bracteoles. However, the morphological characteristics used for species delimitation of this genus remain limited, thereby rendering species identification difficult. To address this problem, it is necessary to perform detailed examinations and observations in field populations.
Bupleurumjeholense Nakai was initially described on the basis of specimens collected from the Wuling Mountain (China). Nakai stated that the characters of this species are as follows: middle stem leaves oblanceolate, base tapering, apex obtuse or acute, apiculate, bracteoles 5, exceeding flowers (Nakai 1937). Later, Chu (Shan and Li 1974) treated B.jeholense as a variety of B.sibiricum Vest ex Spreng., i.e., B.sibiricumvar.jeholense (Nakai) Chu. Chu reported that B.jeholense was similar to B.sibiricum, with the main difference being in bracteole number. For example, B.jeholense commonly has five bracteoles, whereas B.sibiricum usually has 7–12 bracteoles. This difference was thought to be associated with a geographic distribution, i.e., the transition from the main distribution area in Siberia to the limited populations found in the Wuling Mountain (Liaoning Forestry Soil Research Institute 1977).
During recent fieldwork on Dongling Mountain (Beijing, China), we found that the distribution of Bupleurum species was closely linked to altitude. Normally, B.chinense is distributed at low altitudes (<1600 m). However, with the increasing altitude, we found a continuous variation in B.chinense; for example, plants became shorter; the number of branches decreased; the number of bracteoles gradually changed from 3 to 5 and varied in length from obviously shorter than the umbel to almost equal. Furthermore, as the altitude increased to 1600 m, B.chinense was gradually replaced by B.sibiricumvar.jeholense. After checking the specimens, we also found that there were misidentifications between B.chinense and B.sibiricumvar.jeholense. Subsequent phylogenetic analyses of Chinese Bupleurum spp. based on nrDNA ITS and chloroplast markers (i.e., trnH-psbA and matK) indicated that B.sibiricumvar.jeholense was more closely related to B.chinense, whereas B.sibiricumvar.sibiricum was closely related to B.smithii Wolff (Wang et al. 2011b). Hence, we doubt whether B.sibiricumvar.jeholense is a morphological variation of B.chinense that is adapted to high-altitude locations in the Yan Mountains.
We therefore collected and checked several specimens from the type locality of B.jeholense. We conducted morphological observations, statistical comparisons, cytological studies, and a phylogenetic analysis based on the complete chloroplast genome to clarify the relationship among B.sibiricumvar.jeholense, B.sibiricumvar.sibiricum, and B.chinense.
Materials and methods
Morphological observations
Using the existing scientific literature and the relevant type specimens, we collected new specimens from the type locality of B.sibiricumvar.jeholense and compared these with images of the type specimens. Specimens from field collections, CVH (https://www.cvh.ac.cn/), and online herbarium collections (MW, LE) were used to count bracteoles. In total, we obtained bracteole count data for 129 specimens of B.sibiricumvar.jeholense, 183 specimens of B.sibiricumvar.sibiricum, and 183 specimens of B.chinense. We analyzed these data using R (beanplot package) to produce boxplots (Kampstra 2008). Morphological terminology was used according to Kljuykov et al. (2004). Voucher specimens were deposited to the BNU herbarium.
Cytology
All materials used for cytological studies were obtained from the Wuling Mountain (B.sibiricumvar.jeholense), Wutai Mountain (B.chinense), and Daqing Mountain (B.sibiricumvar.sibiricum). All voucher specimens are listed in Table 1. Chromosome preparations were produced using acid digestion and hypotonic wall removal and photographed. Three technical replicates were obtained for each taxon. This procedure was adapted from Li et al. (2021).
Table 1.
Voucher information and GenBank accession numbers for newly sequenced plastome sequences.
| Taxon | Location | Voucher information | Accession |
|---|---|---|---|
| B.chinense | Dongling Mountain, Beijing, China | BNU2021HB002 (BNU) | OR387523 |
| B.smithii | Xiaowutai Mountain, Hebei, China | BNU2020DT007(BNU) | OR387522 |
| B.sibiricumvar.sibiricum | Daqing Mountain, Inner Mongolia, China | BNU2021NMG017(BNU) | OR387525 |
| B.sibiricumvar.jeholense | Wuling Mountain, Hebei, China | BNU2021HB025(BNU) | OR387524 |
Sampling and molecular analysis
Fresh plant leaves were collected from the field and quickly dried with silica gel for DNA extraction. Specimen voucher information is shown in Table 1. DNA was extracted using an HP Plant DNA hypotonic (D2485-02; Omega Bio-Tek). DNA samples were then sent to Beijing Novogene Corporation for quality testing and resequencing. An Illumina HiSeq X sequencing platform was used to generate approximately 10 GB of data for each sample. The chloroplast genome was then assembled from clean data using GetOrganelle (Jin et al. 2020). PGA (Qu et al. 2019) was used to annotate the resulting chloroplast genome. Sequences for B.yinchowense Shan et Y.Li (MT075711) and B.sikangense X.J.He & C.B.Wang (NC056803) were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/nucleotide/) to be used as references. All chloroplast genome sequences generated here were deposited in the NCBI GenBank database (accession numbers listed in Table 1). Finally, twenty-one plastid genome sequences were downloaded from NCBI (see Appendix) for phylogenetic comparisons. This included 19 species of Bupleurum and two species of Pleurospermum Hoffm., which were used as outgroups.
In total, 25 sequences were imported into PhyloSuite (Zhang et al. 2020). The mafft module (Katoh et al. 2019) was used for sequence alignment, and the ModelFinder module (Kalyaanamoorthy et al. 2017) was used to calculate the nucleotide substitution model for all aligned sequences. A maximum likelihood (ML) tree was then constructed using IQ-TREE (Minh et al. 2020), with the nucleotide substitution model set to TVM+F+R2 and a standard bootstrap value of 1000. Results were considered reliable when the bootstrap support value (BS) was ≥70% (Kress et al. 2002). A Bayesian (BI) tree was constructed using MrBayes (Huelsenbeck and Ronquist 2001), with the GTR+F+I+G4 model using the following settings: mcmcp ngen = 2,000,000, printfreq = 10,000, nchains = 4, and burninfrac = 25%. Results were considered reliable when the posterior probability (PP) was ≥0.95. The effective sample size (>200) was determined using Tracer version 1.7 (Rambaut et al. 2018).
Results
Morphological observations
The bean plot indicated that the number of bracteoles of B.sibiricumvar.jeholense was mostly 5 and occasionally 6, whereas that of B.sibiricumvar.sibiricum was (6)7–8(9). Bupleurumchinense had 5, sometimes 4 or even 3 bracteoles at lower elevations or in an understory (Figs 1, 2). The stems of B.chinense and B.sibiricumvar.jeholense were solid, lacking a cavity, whereas those of B.sibiricumvar.sibiricum were hollow at all internodes, which led to the formation of a cavity (Fig. 3). Bupleurumsibiricumvar.jeholense and B.chinense were found to mainly differ in height, number of stems, and presence of middle stem leaves. The morphological characteristics of B.sibiricumvar.jeholense plants are as follows: height below 40 cm, several stems, with 1–2 branches per stem, middle stem leaves narrower, and middle leaf length to width ratio 10–16. In contrast, B.chinense plants had single, occasionally several stem 40–90 cm high, with 2–4 branches per stem, and middle stem leaf length to width ratio 6–10. A comparison of morphological characters is shown in Table 2.
Figure 1.
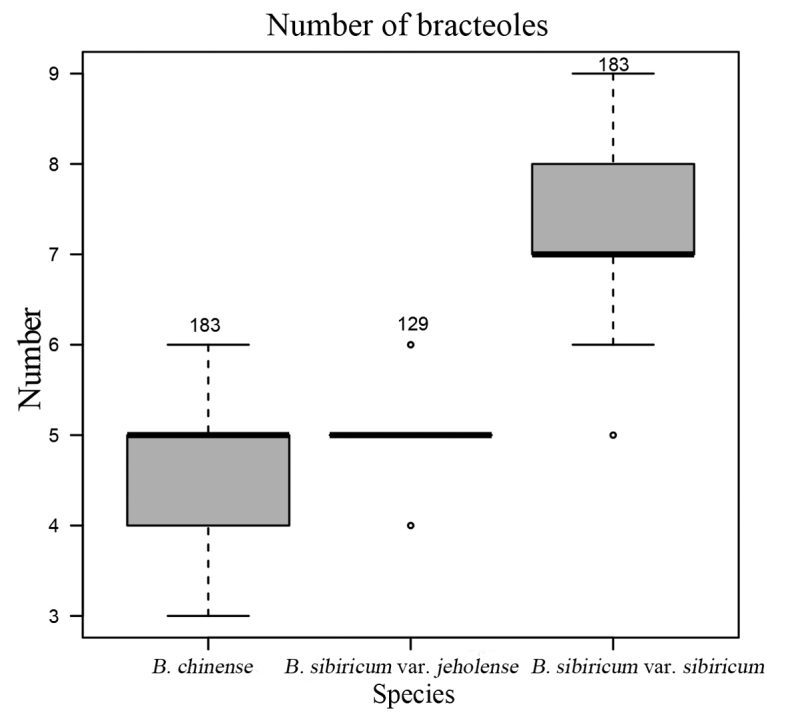
Comparison of the number of bracteoles found in Bupleurumchinense, B.sibiricumvar.jeholense and B.sibiricumvar.sibiricum.
Figure 2.

Umbel and bracteoles of the taxa under study ABupleurumchinenseBB.sibiricumvar.jeholenseCB.sibiricumvar.sibiricum.
Figure 3.

Stem transections of the taxa under study ABupleurumchinenseBB.sibiricumvar.jeholenseCB.sibiricumvar.sibiricum.
Table 2.
Comparison of the morphological characteristics and geographic distributions of three taxa under study.
| Character | B.chinense | B.sibiricumvar.jeholense | B.sibiricumvar.sibiricum |
|---|---|---|---|
| Height | 40–90 cm | 20–40 cm | 30–70 cm |
| Stem (number) | Single, occasionally several | Many, clustered | Many, clustered |
| Stem (branching) | 2–4-branched | 1–2-branched | 1–2-branched |
| Stem (presence of cavity) | Solid | Solid | Hollow in all internodes |
| Basal leaves | Withering early | Withering early | Persistent, many |
| Basal leaf size | 4–7 × 0.6–0.8 cm | 5–10 × 0.3–0.8 cm | 12–25 × 0.7–1.6 cm |
| Middle stem leaves | 4–12 × 0.6–1.8 cm | 6–12 × 0.5–1.2 cm | 6–14 × 0.5–1.6 cm |
| Middle stem leaf length to width ratio | 6–10 | 10–16 | 10–14 |
| Upper stem leaves | Not embracing | Not embracing | Rounded-cuneate, embracing |
| Bracteole relative length | Shorter than flowers | Shorter than or equal to flowers | Exceeding flowers |
| Bracteoles | (3–4) 5, lanceolate | 5 (6–7), lanceolate | (6) 7–12, elliptic-lanceolate |
| Distribution | Northeast and Central China | Yan mountains (Hebei and Beijing) | Heilongjiang, Liaoning, Inner Mongolia, Mongolia, and Russia |
Chromosome numbers
Cytological analysis revealed that the chromosome number of B.sibiricumvar.jeholense was 2n = 12 (x = 6), i.e., the same as B.chinense but different from B.sibiricumvar.sibiricum (2n = 64, x = 8) (Fig. 4). We report the chromosome number of B.sibiricumvar.jeholense for the first time, and the chromosome numbers of B.chinense and B.sibiricumvar.sibiricum determined here are consistent with previous reports (Pan et al. 1995; Qin et al. 1989).
Figure 4.

Metaphase chromosomes of the taxa under study ABupleurumchinenseBB.sibiricumvar.jeholenseCB.sibiricumvar.sibiricum.
Phylogenetic analysis
The size of the four Bupleurum chloroplast genomes ranged from 155,706 to 155,858 bp. The complete chloroplast genome had a typical circular quadripartite structure and consisted of a pair of inverted repeat regions separated by the large single copy and small single copy regions. The topologies of the ML and BI trees constructed using the cp genome sequences were consistent. These results showed that the genus Bupleurum can be divided into two clades, with all Chinese Bupleurum plants belonging to B.subg.Bupleurum. Bupleurumsibiricumvar.sibiricum was found to cluster with B.smithii and B.bicaule Helm (BS = 100% PP = 1), whereas B.chinense was found to cluster with B.sibiricumvar.jeholense (BS = 100% PP = 1) and was more distantly related to B.longiradiatum Turcz., B.falcatum L. and B.boissieuanum H. Wolff (Fig. 5).
Figure 5.

Phylogenetic tree of some Chinese Bupleurum species as inferred from chloroplast genomes using ML analyses (numbers below branches represent ML bootstrap values and BI posterior probabilities).
Discussion
Comparative morphology
Botanists have closely monitored the number of bracteoles because this character is essential in the taxonomy of Bupleurum. Moreover, bracteole number is an important indicator of the taxonomic position of B.sibiricumvar.jeholense in the genus. Five bracteoles are generally present in both B.chinense and B.sibiricumvar.jeholense. However, in environments such as the understory, B.chinense may have only three or four bracteoles. Furthermore, 6–7 bracteoles have occasionally been observed in B.sibiricumvar.jeholense. We observed that the number of bracteoles in B.sibiricum samples collected from the Daqing Mountain, which were collected at the same latitude and altitude as the samples collected from Wuling Mountain, was not 5, as would be expected for B.sibiricumvar.jeholense. Thus, we speculated that the 5-bracteoled B.sibiricumvar.jeholense may not have evolved from the 12-bracteoled B.sibiricumvar.sibiricum.
In the classification of the genus Bupleurum, stem structural characteristics have rarely been examined. After observing numerous specimens, we found that the stem of B.sibiricumvar.sibiricum was hollow and contained a substantial cavity. In contrast, the stems of B.chinense and B.sibiricumvar.jeholense lacked this cavity. The presence or absence of a stem cavity is a stable character and does not change with the period of growth. For example, B.komarovianum was once treated as a variety of B.chinense (Liaoning Forestry Soil Research Institute 1977) until Wang et al. (2011b) found that the stems of B.komarovianum were hollow. The authors combined morphological and chromosomal evidence to argue that B.komarovianum should be reinstated as a separate species. Finally, the basal leaves of B.sibiricumvar.sibiricum were found to be persistent, whereas those of B.sibiricumvar.jeholense were found to wither at the flowering and fruiting stages, as in B.chinense.
Bupleurumchinense is widely distributed throughout East Asia and is often cultivated as a medicinal plant. The morphology of this species varies with the environment. Bupleurumsibiricumvar.jeholense may be a variant of B.chinense that has adapted to the cold environments found in high-altitude mountains. In particular, the bracteole number may have increased to protect flowers at higher altitudes (Kofidis et al. 2007). Compared with B.chinense plants, B.sibiricumvar.jeholense plants are shorter, with several stems, fewer stem branches, and narrower middle stem leaves. Overall, the above evidence suggests that B.sibiricumvar.jeholense should be treated as a variety of B.chinense.
Cytological analysis
Chromosomal variation plays a vital role in species formation, and the diversity of chromosome size and number is therefore an important character that can be used to track Bupleurum species that have adapted to different habitats (Wang 2011a; Weiss-Schneeweiss and Schneeweiss 2013). Bupleurumsibiricumvar.jeholense differs from B.sibiricumvar.sibiricum in both chromosome number and basal number, but these values are identical to those of B.chinense. In this study, B.chinense and B.sibiricumvar.jeholense were both found to have a chromosome number of 12 (diploid), which is the common basic number reported for this genus (Wang 2011a). Regarding the reported chromosome number of 64 in B.sibiricum from a population collected in Inner Mongolia, Qin et al. (1989) and Jiang et al. (2002) assumed that B.sibiricum was octoploid (i.e., with a basal number of 8), marking the highest level of polyploidy found in this genus. This observation may be due to chromosomal polyploidy, a type common in colder climates (Jiang et al. 2002). Thus, in terms of both chromosome number and basal number, B.sibiricumvar.jeholense appears to be only distantly related to B.sibiricumvar.sibiricum.
Phylogenetic analysis
In this study, a phylogenetic tree was reconstructed using chloroplast genome data. Our findings were consistent with those of Wang et al. (2011b). In particular, we found that Bupleurumsibiricumvar.jeholense was embedded in B.chinense, which forms a sister clade with B.yinchowense. In addition, B.sibiricumvar.sibiricum is sister to B.smithii and B.bicaule. In contrast to B.chinense, the distributions of B.sibiricum and B.bicaule are ranging from Siberia to northeastern China.
Distribution
Bupleurumsibiricum is distributed widely throughout temperate Asia. It often co-occurs with B.scorzonerifolium in arid meadows in Inner Mongolia and Siberia at elevations of 700–2000 m. In the field, B.sibiricumvar.jeholense is often found to co-occur with B.chinense at different altitudes in the same mountain. At present, B.sibiricumvar.jeholense is found only in high-altitude areas of the Yan Mountains. Moreover, it has a narrow distribution area and is confined to altitudes of 1500–2000 m. In contrast, B.chinense is widely distributed and is found at altitudes ranging from 200 to 1600 m in Northeast China; however, it has also been found isolated at an altitude of 2100 m in Northwest China. Given these findings, we speculate that B.sibiricumvar.jeholense is a specialized morphological variant of B.chinense that has specifically adapted to high altitudes. Overall, in terms of distribution and habitat, B.sibiricumvar.jeholense differs considerably from B.sibiricum and is more similar to B.chinense.
Taxonomic treatment
. Bupleurum chinense var. jeholense
(Nakai) Q.R.Liu & L.H.Wang comb. nov.
3EBB3207-7F5B-5758-B228-F9C187CC216E
urn:lsid:ipni.org:names:77335467-1
– Bupleurumjeholense Nakai in J. Jap. Bot. 13: 482 (1937) – Bupleurumsibiricumvar.jeholense (Nakai) Chu in Shan & Li, Acta Phytotax. Sin. 12 (3): 272 (1974).
= Bupleurumjeholensevar.latifolium Nakai in J. Jap. Bot. 13: 482 (1937). Type. China. Hebei: Wuling Mountain, 1800 m, 2 Sept 1933, Nakai, Honda et Kitagawa s. n. (holotype: TI0082957!).
Type.
China. Hebei: Wuling Mountain, 1500–2000 m, 2 Sept 1933, Nakai, Honda et Kitagawa, s. n. (holotype: not barcode, fig in protologue pp 481!; isotypes: TI0082958! TI0082959! (Fig. 6) TI0082960! TI0082961!],
Figure 6.

Isotype of Bupleurumchinensevar.jeholense (Photo from TI Herbarium).
Diagnosis.
Bupleurumchinensevar.jeholense is morphologically similar to var. chinense and can be distinguished from the latter by plant size (>40 cm), the presence of multiple stems, with 1–2 branches per stem, thinner middle stem leaves, leaf length to width ratio 10–16, and the presence of 5 bracteoles.
Description.
Plants 20–40 cm, perennial. Root stout, brown, woody. Stem solitary or several, solid, petioles often purplish-red, clasping base without fibrous remnant sheaths. Basal leaves oblanceolate, 5–10 × 0.3–0.8 cm, base petioles, apex acuminate. Middle leaves sessile, oblanceolate, 4.5–12 × 0.4–1.4 cm, 7–9-nerved, base tapering, apex obtuse or acute, apiculate. Apical leaves small. Umbels 5–12, nearly equal or unequal rays 0.4–4 cm long; bracts of 1–5 unequal leaflets, often obsolete or deciduous, 3–15 × 0.6–11 mm, 4–7-nerved; bracteoles 5, lanceolate, 3–7 × 0.6–0.8 mm, exceeding flowers; umbellule 4–12 mm across, 8–14-flowered. Petals bright yellow. Stylopodium low-conic, discoid, dark yellow. Fruit oblong, brown, ca. 2.2–3.5 × 0.9–1.5 mm; ribs prominent, narrowly winged, wings pale brown; vittae 3(–4) in each furrow, 4 on commissure (Fig. 7). Fl. July–August and Fr. August–October. 2n = 12.
Figure 7.

Bupleurumchinensevar.jeholenseA plant B umbel and bract C flowers D umbel (side view) E pollen F transverse section of mericarp.
Phenology.
Flowering and fruiting from August to October.
Distribution and habit.
Hebei, Beijing. It grows in mountains at elevations of 1500–2000 m.
Additional specimens examined.
China. Beijing: Fangshan county, Jin-Wu Wang s.n. (PEY!); Mentougou county, Xiao-Liu QS-186 (BJFC!); Quan-Ru Liu 200609009 (BNU!); Xue Lin 05 05-4-114 (BJFC!); Gang-Min Zhang 201008036 (BJFC!); Xian-Yun Mu 1821 (BJFC!); Duan-Zheng Lu s.n. (BJFC!); Quan-Ru Liu DL025-2 (BNU!); Quan-Ru Liu DL026-2 (BNU!); Quan-Ru Liu DL027-1 (BNU!); Miyun county, Xian-Yun Mu 1924 (BJFC!). Hebei: Xinglong county, Li-Hua Wang BNU2021HB029 (BNU!); Li-Hua Wang BNU2021HB025 (BNU!); Jia-Yi Liu 0845 2190 (TIE!); Jin-Wu Wang s.n. (PEY!); Zhen-Fu Fang 825 (NAS!); Ze-Hui Pan 83940 (NAS!); Ze-Hui Pan 83939 (NAS!); Shen-E Liu 4833 (IFP!); Zhen-Fu Fang 826 (IFP!); Quan-Ru Liu WLS068 (BNU!); Xin-Yuan Liu 1652 (KUN!); Jia-Yi Liu 08450 (TIE!); Jia-Yi Liu 00485 (TIE!); Biaobenshi 2190 (PE!); Wu-Xiu Zhang 91 (PE!).
Supplementary Material
Acknowledgements
We gratefully acknowledge the collaboration of the curators and staff of the herbaria mentioned in the text for their assistance upon visits, as well as for access to digital specimen images. Special thanks are due to Professor Akiko of the University of Tokyo for kindly providing photographs of the type specimens. We thank Dr. Dan-Hui Liu for the analyses of molecular sequence data and Xun Sun for the field assistance.
Citation
Wang L-H, Xu X-M, He Y, Liu Q-R (2024) The identity of Bupleurum jeholense (Apiaceae). PhytoKeys 237: 231–243. https://doi.org/10.3897/phytokeys.237.111977
Additional information
Conflict of interest
The authors have declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Funding
This research was funded by the Biodiversity survey and assessment project of the Ministry of Ecological Environment (2019HJ2096001006).
Author contributions
Funding acquisition: QRL. Investigation: LHW. Software: XMX, LHW. Supervision: QRL. Writing – review and editing: YH, XMX, LHW.
Author ORCIDs
Yi He https://orcid.org/0000-0002-6925-7299
Quan-Ru Liu https://orcid.org/0000-0003-4270-4746
Data availability
All of the data that support the findings of this study are available in the main text.
References
- Downie SR, Katz-Downie DS, Watson MF. (2000) A phylogeny of the flowering plant family Apiaceae based on chloroplast DNA rpl16 and rpoC1 intron sequences: Towards a suprageneric classification of subfamily Apioideae. American Journal of Botany 87(2): 273–292. 10.2307/2656915 [DOI] [PubMed] [Google Scholar]
- Huelsenbeck JP, Ronquist F. (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics (Oxford, England) 17(8): 754–755. 10.1093/bioinformatics/17.8.754 [DOI] [PubMed] [Google Scholar]
- Jiang CM, Lu F, Peng YL. (2002) Chromosomal study on Bupleurum L. in Northeast China. Journal of Heilongjiang Nongken Normal College (4): 50–60.
- Jin JJ, Yu WB, Yang JB, Song Y, de Pamphilis CW, Yi TS, Li DZ. (2020) GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biology 21(1): 1–31. 10.1186/s13059-020-02154-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalyaanamoorthy S, Minh BQ, Wong TKF, Haeseler A, Jermiin L. (2017) ModelFinder: Fast model selection for accurate phylogenetic estimates. Nature Methods 14(6): 587–589. 10.1038/nmeth.4285 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kampstra P. (2008) Beanplot: A Boxplot Alternative for Visual Comparison of Distributions. Journal of Statistical Software 28(1): 1–9. 10.18637/jss.v028.c0127774042 [DOI] [Google Scholar]
- Katoh K, Rozewicki J, Yamada KD. (2019) MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Briefings in Bioinformatics 20(4): 1160–1166. 10.1093/bib/bbx108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kljuykov EV, Liu M, Ostroumova TA, Pimenov MG, Tilney PM, Wyk B-E. (2004) Towards a standardised terminology for taxonomically important morphological characters in the Umbelliferae. South African Journal of Botany 70(3): 488–496. 10.1016/S0254-6299(15)30233-7 [DOI] [Google Scholar]
- Kofidis G, Bosabalidis AM, Moustakas M. (2007) Combined effects of altitude and season on leaf characteristics of Clinopodiumvulgare L. (Labiatae). Environmental and Experimental Botany 60(1): 69–76. 10.1016/j.envexpbot.2006.06.007 [DOI] [Google Scholar]
- Kress WJ, Prince LM, Williams KJ. (2002) The phylogeny and a new classification of the gingers (Zingiberaceae): evidence from molecular data. American Journal of Botany 89(11): 1682–1696. 10.3732/ajb.89.10.1682 [DOI] [PubMed] [Google Scholar]
- Li HM, Zhou W, Song CF. (2021) Karyotype analysis of two species in Pternopetalum Franch. of Apiaceae. Journal of Plant Resources and Environment 30(1): 69–71. [Google Scholar]
- Liaoning Forestry Soil Research Institute (1977) Flora Plantarum Herbacearum Chinae Boreali-Orientalis vol. 6. Science Press, Beijing, 188–200.
- Minh BQ, Schmidt HA, Chernomor O, Schrempf D, Woodhams M, Haeseler A, Lanfear R. (2020) IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Molecular Biology and Evolution 37(5): 1530–1534. 10.1093/molbev/msaa015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakai T. (1937) Notulae ad plantas asiae orientale. Shokubutsu Kenkyu Zasshi 13: 471–491. [Google Scholar]
- Pan ZH, Zhuang TD, Zhou XL, Yao XM. (1995) Karyotype analysis of four taxa of Bupleurum used in Chinese drugs. Jorrnal of Plant Resources and Environment 4(4): 41–45. [Google Scholar]
- Plunkett GM, Pimenov MG, Reduron JP, Kljuykov EV, Wyk BE, Ostroumova TA, Henwood MJ, Tilney PM, Spalik K, Waston MF, Lee BY, Pu FD, Webb CJ, Hard JM, Mitchell AD, Muckensturm B. (2019) Apiaceae. In: Kadereit JW, Bittrich V (Eds) The families and genera of vascular plants, vol. 15, Flowering plants, Eudicots: Apiales, Gentianales (except Rubiaceae). Springer, Cham, 109 pp. [Google Scholar]
- Qin HZ, Pan ZH, Sheh ML, Wu ZJ. (1989) A Report on Chromosome Numbers of Chinese Umbelliferae. Zhiwu Fenlei Xuebao 27(4): 268–272. [Google Scholar]
- Qu XJ, Moore MJ, Li DZ, Yi TS. (2019) PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 15(1): 1–12. 10.1186/s13007-019-0435-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rambaut A, Drummond AJ, Xie D, Baele G, Suchard MA. (2018) Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Systematic Biology 67(5): 901–904. 10.1093/sysbio/syy032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shan RH, Li Y. (1974) On the Chinese species of Bupleurum L. Zhiwu Fenlei Xuebao 12: 261–294. [Google Scholar]
- Sheh ML, Watson MF. (2005) Bupleurum L. In: Wu ZY, Raven PH, Hong DY. (Eds) Flora of China, vol.14. Missouri Botanical Garden Press & Science Press, Beijing & St. Louis, 60–74.
- Wang CB. (2011a) Systematic and taxonomical studies on Chinese Bupleurum L. (Apiaceae) Sichuan University.
- Wang CB, Ma XG, He XJ. (2011b) A taxonomic re-assessment in the Chinese Bupleurum (Apiaceae): Insights from morphology, nuclear ribosomal internal transcribed spacer, and chloroplast (trnH-psbA, matK) sequences. Journal of Systematics and Evolution 49(6): 558–589. 10.1111/j.1759-6831.2011.00157.x [DOI] [Google Scholar]
- Weiss-Schneeweiss H, Schneeweiss GM. (2013) Karyotype diversity and evolutionary trends in angiosperms. In: Leitch IJ, Greilhuber J, Doležel J, Wendel JF (Eds) Plant Genome Diversity 2: Physical Structure, Behaviour and Evolution of Plant Genomes, Springer, Wien, 209–230. 10.1007/978-3-7091-1160-4_13 [DOI]
- Zhang D, Gao FL, Jakovlić I, Zou H, Zhang J, Li WX, Wang GT. (2020) PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Molecular Ecology Resources 20(1): 348–355. 10.1111/1755-0998.13096 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
All of the data that support the findings of this study are available in the main text.