

Abstract
The ToxRS system in Vibrio cholerae plays a central role in the modulation of virulence gene expression in response to environmental stimuli. An integration of multiple signalling inputs mediated by ToxR, -S, and -T controls virulence gene expression leading to cholera toxin (CT) production. Recently, we identified a new virulence locus, varA (virulence associated regulator), in classical V. cholerae O1 that positively controls transcription of tcpA, the major subunit of the toxin-coregulated pilus (TCP) and the production of CT, two key factors in cholera pathogenesis. The varA locus is a homolog of gacA (originally described for the soil organism Pseudomonas fluorescens), which encodes a conserved global regulator belonging to the family of two-component signal transducing molecules. GacA homologs in a number of diverse gram-negative pathogenic bacterial species have been implicated in controlling the production of diverse virulence factors. varA mutants showed reduced levels of tcpA message and TcpA protein, lacked visible signs of autoagglutination (a phenotype associated with functional TCP), produced decreased levels of CT, and were attenuated in colonizing infant mice. Transcription of varA appears to be independent of ToxR, and overexpression of the regulators tcpPH and toxT from plasmids in the varA mutant restored wild-type levels of CT production and the ability to autoagglutinate. varA represents an additional modulating factor in the coordinate expression of virulence factors in V. cholerae.
The gram-negative bacterium Vibrio cholerae is the causative agent of the diarrheal disease cholera. The pathogenic potential of V. cholerae lies in its ability to adhere to and colonize the epithelia of the small intestine of the host and secrete cholera toxin (CT). The molecular basis for the expression of virulence factors involved in colonization and toxin production depends largely on the transcriptional activator ToxR, a transmembrane protein which senses the environment and activates the expression of appropriate genes (a set of genes collectively termed the ToxR regulon) in response to these environmental signals. ToxS is a protein that stabilizes the conformation of ToxR in the periplasm. ToxR transcriptionally activates a second regulator, ToxT, which positively controls the expression of CT, encoded by the ctxAB operon, as well as the expression of the toxin-coregulated pilus (TCP), which is essential for intestinal colonization (47). While a linear signal transduction cascade consisting of ToxR, ToxS, and ToxT in V. cholerae has been previously delineated, recent studies have suggested that virulence genes may be controlled by multiple signalling inputs leading to CT production. Studies in classical V. cholerae O1 revealed that an additional pair of regulatory proteins, TcpP and TcpH, positively control the transcription of toxT and that transcription of the tcpPH operon is itself regulated by temperature and pH independently of ToxR or ToxT (7, 17). Although TcpPH can activate the toxT promoter independently of ToxR, high activation levels appear to require the synergistic interaction of ToxRS and TcpPH (17).
Many bacterial pathogens utilize signal transducing proteins of the two-component family of regulators to modulate expression of virulence genes in response to environmental signals (31, 49). For example, GacA (for global antibiotic and cyanide) control in Pseudomonas fluorescens (25) is a global regulator of virulence-related factors in Pseudomonas species, including P. syringae (42) and P. aeruginosa (38), as well as in other gram-negative bacteria, including Escherichia coli (34) and Salmonella typhimurium (22). Sequence data indicate that the GacA protein is a response regulator in the FixJ-DegU family of two-component signal transducing molecules (1, 25). In P. aeruginosa, gacA is required for full pathogenicity in both plant and animal models of infection (38), and the gacA homolog of S. typhimurium, sirA, has been shown to encode an essential transcriptional activator of invasion genes (22).
The presence of gacA homologs in diverse gram-negative bacterial species suggested that studies of this locus in additional gram-negative pathogens may contribute to our knowledge of virulence mechanisms. In this report, we describe a member of the two-component family of regulators involved in expression of the ToxR regulon in classical V. cholerae O1. Although ToxR has been shown to bind to DNA (19, 33), it does not fit into the family of two-component regulatory systems because it lacks the canonical phosphoryl-acceptor domain characteristically found in this group of regulators (33). We have identified a potential gacA homolog in V. cholerae termed varA (for virulence associated regulator) which modulates the production of two major virulence factors of this organism, the TCP and CT. VarA represents the first response regulator of the two-component family found to be involved in expression of virulence factors in V. cholerae.
MATERIALS AND METHODS
Bacterial strains and culture conditions.
Bacterial strains and plasmids used in this study are shown in Table 1. E. coli and V. cholerae strains were maintained at −70°C in Luria-Bertani (LB) broth containing 15% glycerol for long-term storage. Antibiotics added to LB medium were as follows: ampicillin (Ap), 100 μg/ml for E. coli and 25 or 50 μg/ml for V. cholerae; chloramphenicol (Cm), 25 μg/ml; kanamycin (Km), 50 μg/ml; streptomycin (Sm), 100 μg/ml; and tetracycline (Tc), 15 μg/ml for E. coli and 5 μg/ml for V. cholerae. 5-Bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-Gal) was added to LB agar at 40 μg/ml. To induce ToxR-dependent gene expression in V. cholerae, bacteria were grown in LB broth containing 85.5 mM NaCl at a starting pH of 6.5 with moderate aeration at 30°C. To repress ToxR-dependent gene expression, V. cholerae strains were grown in LB broth containing 171 mM NaCl at a starting pH of 8.5 with moderate aeration at 37°C (7). Isopropyl-β-d-thiogalactopyranoside (IPTG) was added to ToxR-inducing medium at 1 mM to induce expression of toxT from the tac promoter in pMT5K.
TABLE 1.
Bacterial strains and plasmids used in this study
| Strain or plasmid | Description | Source or reference |
|---|---|---|
| Strains | ||
| V. cholerae | ||
| O395 | Classical O1 Ogawa strain; Smr | 51 |
| SW400-12 | O395 with polar mutation in varA created by integration of the suicide plasmid pSW400-12; Smr Apr | This work |
| SW8 | SW400-12, reverted to wild type by serial passaging in the absence of Ap and selecting for loss of vector; Smr | This work |
| SW33S | O395 varA::aphA-3; Smr Kmr | This work |
| O395 toxR | O395 with mutation in toxR created by integration of the suicide plasmid pVM55; Smr Apr | This work |
| CS2-1 | O395 tcpA::TnphoA; Smr Kmr | 51 |
| CG842 | O395 ΔlacZ; Smr | 15 |
| E. coli | ||
| DH5α | F−endA1 hsdR17 supE44 thi-1 recA1 gyrA96 relA1 Δ(argF− lacZYA) U169 (φ80d lacZΔM15) | 16 |
| SM10 λ pir | thi thr leu tonA lacY supE recA::RP4-Tc::Mu λ pir R6K | 32 |
| Plasmids | ||
| pCRII | TA cloning vector; Apr | Invitrogen |
| pBR322 | Cloning vector; Apr Tcr | 5 |
| pTA400-12 | 389-bp varA PCR product from V. cholerae O395 cloned into pCRII; Apr | This work |
| pSW7H3 | 7-kb HindIII genomic fragment containing varA locus from V. cholerae O395 cloned into HindIII of pBR322; Apr Tcr | This work |
| pBluescript | Cloning vector; Apr | Stratagene |
| pSW3sac | 3-kb SacI fragment containing varA from pSW7H3 cloned into SacI of pBluescript; Apr | This work |
| pSWSA | pSW3sac digested with AatII and SalI and religated, Δpolylinker region between EcoRI and Asp718; Apr | This work |
| pPUC18K | Carries a Kmr gene cassette for creating nonpolar mutations; Apr Kmr | 30 |
| pSW36 | 800-bp SmaI fragment containing Kmr cassette from pPUC18K cloned into blunt-ended HindIII of pSWSA; Apr Kmr | This work |
| pKAS32 | Suicide vector containing rpsL gene; Apr | 48 |
| pSW37 | 3.2-kb SacI fragment from pSW36 cloned into SacI of pKAS32; Apr | This work |
| pVM55 | pJM703.1 with 630-bp EcoRI-HpaI internal fragment of toxR; Apr | 32 |
| pGP704 | Suicide vector; Apr | 32 |
| pSW400-12 | 400-bp EcoRI fragment from pTA400-12 cloned into EcoRI of pGP704; Apr | This work |
| pBBR1MCS | Cloning vector; Cmr | 23 |
| pPAC22 | PCR product of tcpPH from O395 cloned into pUC19; Apr | 7 |
| pSW1.4-1 | 1.4-kb BamHI fragment of pPAC22 containing tcpPH of V. cholerae O395 cloned into BamHI of pBBR1MCS in same orientation to the lacZ promoter; Cmr | This work |
| pSW1.4-5 | 1.4-kb BamHI fragment of pPAC22 containing tcpPH of V. cholerae O395 cloned into BamHI of pPBBR1MCS in opposite orientation to the lacZ promoter; Cmr | This work |
| pMT5 | pMMB66HE with toxT under ptac control, Apr | 13 |
| pUC71K | pBR322 and M13mp7-derived vector containing Km resistance gene from Tn903; Apr Kmr | 53 |
| pMT5K | 1.3-kb PstI insert containing Kmr cassette from pUC71K made blunted ended and cloned into PvuI (also made blunt ended) within bla of pMT5; Kmr | This work |
CT assay.
The levels of CT production after 16 to 24 h of growth in ToxR-inducing and ToxR-repressing conditions were determined by using a GM-1 enzyme-linked immunosorbent assay as previously described (20). In parallel, these same cultures were visually inspected for autoagglutination, which refers to the clumping of cells at the bottom of the culture tube and which is usually seen in ToxR-inducing conditions for classical strains of V. cholerae, such as O395 (51).
DNA and RNA analysis.
Nucleic acid manipulations were performed by standard molecular biological techniques (2). tcpA and tcpPH DNA fragments used as probes in Northern blot analyses were generated by PCR and radiolabelled by the random hexamer priming method (Stratagene, La Jolla, Calif.). Primers and PCR conditions used to generate tcpA were described previously (7). Primers used to generate the 605-bp PCR product containing tcpPH were PAC24 (5′ TTAGCAAGGTTACCGGGGATAACAA 3′) that spans positions 2264 to 2288 of the sequence with GenBank accession no. X64098 and PAC15 (5′ AAAGGATCCGTAAAGATGAAGAGATGACTT 3′) that spans positions 2870 to 2850 (7). PCR conditions were the same as those described below. Primers to generate the 644-bp PCR product containing an internal fragment of the varA open reading frame were 5′ GCGCGGTACCGGTGCAGAGCCAATAGATAAGTG 3′, which incorporates an Asp718 restriction site at the 5′ end, and 5′ GACGCGTCGACACTATAACTTCTCAGTATCATTC 3′. To confirm equal loading of RNA, one set of samples loaded in parallel was stained with ethidium bromide. Quantitation of tcpA mRNA (see Fig. 1) was done by exposing the RNA blot to phosphor screens followed by reading on a PhosphorImager (Molecular Dynamics, Sunnyvale, Calif.). DNA sequencing was performed by the DNA Sequencing Core Facility at the Massachusetts General Hospital Department of Molecular Biology. Percent amino acid identities of VarA compared to GacA homologs from P. aeruginosa, S. typhimurium, and E. coli were performed with GAP from the program manual for the Wisconsin Package (Genetics Computer Group, Madison, Wis.).
FIG. 1.
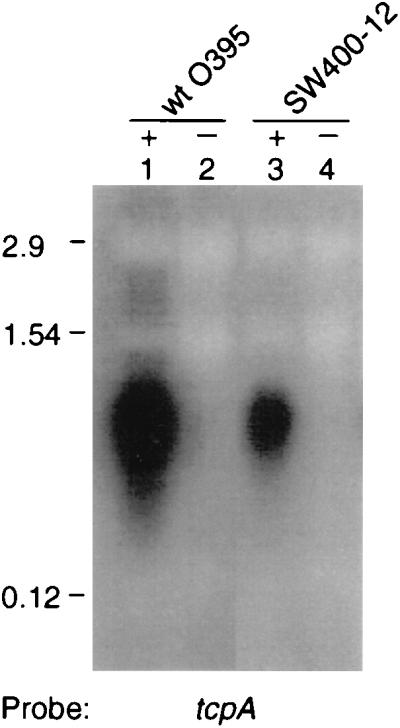
Northern blot containing 10 μg of total RNA from O395 (lanes 1 and 2) and SW400-12 (lanes 3 and 4) cultures grown under ToxR-inducing (+) and ToxR-repressing (−) conditions and probed for the tcpA message. Numbers at left are the sizes for the rRNAs, 23S (2.9 kb), 16S (1.54 kb), and 5S (0.12 kb) subunits in V. cholerae which run at the same molecular masses as the rRNAs in E. coli.
Isolation of the gacA homolog.
To identify a gacA homolog from V. cholerae classical strain O395, PCR was performed using the following degenerate oligonucleotides: 5′ ATGCCNGGNATHGGNGGNCTNGARGCG 3′, corresponding to amino acids 57 to 65 (MPGIGGLEA), and 5′ NCGRTANCGRTANGWRTTNACNGTYTTNGG 3′, corresponding to amino acids 178 to 187 (PKTVNTSYRYR) of the P. aeruginosa gacA sequence (GenBank accession no. U27988). PCR was carried out in a 100-μl reaction volume in 10 mM Tris-HCl, (pH 8.3), 15 mM MgCl2, 50 mM KCl, 200 μM deoxynucleotide triphosphates, 150 pmoles of each degenerate oligonucleotide, 1 μg of V. cholerae O395 genomic DNA, and 2.5 U of Taq DNA polymerase (Boehringer Mannheim Biochemicals, Indianapolis, Ind.). The PCR conditions were as follows: 1 cycle at 95°C for 4 min, 30 cycles at 55°C for 45 s, 72°C for 1 min, and 95°C for 30 s. A predicted PCR product of 390 bp was obtained (we also detected a similarly sized PCR product in two other V. cholerae strains, C6709 of the El Tor biotype and MO10, a V. cholerae O139 strain [not shown]). The 390-bp PCR product from O395 was gel purified (Geneclean; Bio 101 Inc., La Jolla, Calif.) and cloned into pCRII (Invitrogen, San Diego, Calif.) to generate pTA400-12. The primers 5′ CGCCAGGGTTTTCCCAGTCACGAC 3′ and 5′ AGCGGATAACAATTTCACACAGGA 3′ were used to amplify an approximately 660-bp product from pTA400-12, which includes the entire 390-bp varA insert, using the conditions described above. The amplified product was sequenced directly. Southern blots containing genomic DNA from O395 showed that the radiolabelled 390-bp PCR product hybridized to an approximately 7-kb HindIII fragment (data not shown). To identify a clone carrying a full-length gacA homolog from V. cholerae, an approximately 7-kb HindIII size-selected genomic DNA library in pBR322 was constructed and probed with the same 390-bp PCR product to obtain pSW7H3.
Strain and plasmid constructions.
To construct a polar mutation in the V. cholerae O395 varA locus, a 400-bp EcoRI fragment from pTA400-12 that contained 390 bp of varA sequence was ligated to the EcoRI site of the suicide vector pGP704 (32) to create pSW400-12. The E. coli donor strain SM10 λ pir was used to mobilize pSW400-12 into the genome of wild-type V. cholerae O395 to create SW400-12. Disruption of the varA locus was verified by DNA blotting and PCR analysis. SW8, a varA+ revertant of SW400-12, which has lost the integrated plasmid pSW400-12, was obtained by serial passage of SW400-12 for 8 days in the absence of Ap followed by screening for loss of the plasmid on LB agar plates containing Ap at 25 or 50 μg/ml. Reconstitution of the wild-type varA locus was confirmed by PCR analysis.
To generate a nonpolar varA mutant in V. cholerae O395, briefly, a 583-bp AatII-SalI restriction fragment internal to varA was deleted and replaced with a Kmr cassette from pUC18K (30) such that transcription was maintained and translation of the downstream portion of varA was reinitiated at the 3′ end of the Kmr cassette. In detail, construction of the nonpolar varA strain was as follows. (i) A 3-kb SacI fragment containing the full length varA gene derived from pSW7H3 was cloned into pBluescript KS+ (Stratagene) to generate pSW3sac. (ii) pSW3sac was digested with AatII and SalI and primer adaptors (see below) were used to reseal the AatII and SalI ends and to restore the transcription reading frame. The top strand primer was 5′ CGATATCAAGCTTGAGAAGTTATAGTG 3′ and the bottom strand primer was 5′ TCGACACTATAACTTCTCAAGCTTGATATCGACGT 3′. (The HindIII restriction site is underlined. The top and bottom primers were heated at 65°C and cooled to room temperature to facilitate self-annealing prior to ligation). (iii) The HindIII restriction site in the polylinker region of pSW3sac was removed by digesting with EcoRI and Asp718, followed by end-filling with Klenow polymerase and religation to create pSWSA. (iv) An 800-bp SmaI fragment containing the aphA-3 gene encoding Km resistance from pUC18K (30) was cloned into pSWSA at the unique HindIII site located within the primer adaptors described above, following end-filling with Klenow polymerase, to generate pSW36. (v) A 3.2-kb SacI fragment from pSW36 was cloned into the positive selection vector pKAS32 (48) at the SacI site to generate pSW37. (vi) pSW37 was mobilized from SM10 λ pir to V. cholerae O395, and transconjugants were selected on Ap and Km. (vii) Four Apr and Kmr cointegrates were streaked onto LB agar containing Sm at 1 mg/ml and Km at 50 μg/ml. Loss of the Ap-resistance-encoding gene from the vector was confirmed by the inability of Sm and Km survivors to form colonies on LB agar plates containing Ap. The mutation in one mutant, SW33S, was confirmed by PCR and sequencing of the 3′ junction between the Km resistance gene and downstream varA sequences.
SDS-polyacrylamide gel electrophoresis and Western analysis.
For Western analysis, proteins from whole-cell lysates from cultures grown overnight under ToxR-inducing and ToxR-repressing conditions were separated on a 12% polyacrylamide gel in the presence of sodium dodecyl sulfate (SDS) as previously described (24). The amount of total protein loaded (from 108 bacteria) was normalized to the optical density at 600 nm (OD600) of the overnight cultures from each strain. Proteins were transferred to Hybond-enhanced chemiluminescence nitrocellulose (Amersham, Arlington Heights, Ill.) at 4°C in electroblotting buffer (20 mM Tris–150 mM glycine [pH 8.0]) using a Mini Trans-Blot Module (Bio-Rad, Hercules, Calif.) in accordance with the manufacturer’s instructions. Immunoreactive proteins were identified as a chemiluminescent signal by using the ECL Western blotting system (Amersham Life Science) with polyclonal rabbit anti-TcpA or anti-ToxR antibodies (gifts of J. J. Mekalanos). Kaleidoscope protein molecular mass standards were from Bio-Rad.
Phage transduction.
Phage transduction assays using a derivative of CTXΦ (54) carrying the Km resistance gene were performed as previously described (9). The recipient strains, wild-type O395 and SW400-12, were grown in ToxR-inducing conditions prior to mixing with an equal volume of CTXΦ (gift of Wei Lin). The phage and bacteria mixture was incubated for 30 min at room temperature before plating at the appropriate dilutions on LB agar containing Sm and Km (control was plated on Sm alone).
Immunoelectron microscopy.
Strains used for electron microscopy were grown overnight in ToxR-inducing conditions at 30°C prior to immunogold labelling as described by Chiang et al. (10).
Animal studies.
A competition assay for intestinal colonization in infant mice was assessed as previously described (10). Briefly, 4- to 5-day-old CD1 mice were inoculated orally with 50 μl of equally mixed cultures of wild-type O395 and an isogenic Lac− strain, CG842 (15), in doses of 1.8 × 106 and 0.36 × 106 CFU or the nonpolar varA mutant SW33S and the Lac− strain in doses of 1.5 × 106 and 0.36 × 106 CFU, and mice were then placed in a 30°C incubator. Groups of two to four mice were used at each dose. After 21 to 24 h, the mice were sacrificed and the small intestines were removed, homogenized in 5 ml of LB broth, and plated on LB agar containing X-Gal at the appropriate dilutions to determine the output ratio of competing strains (number of colonies for wild-type or SW33S divided by the number of colonies for the Lac− strain).
For the in vitro competition assay, wild-type O395, SW33S, and CG842 were grown overnight at 37°C, subcultured, and grown to an approximate OD600 of 1. A volume of 50 μl containing a mixed culture (done in triplicate) of wild-type O395 and CG842 (106 CFU each) or SW33S and CG842 (106 CFU each) was inoculated into 5 ml of LB broth in ToxR-inducing conditions and grown at 30°C for 21 to 24 h. Cultures were plated at appropriate dilutions to determine the ratio of the competing strains. The output ratios for both the in vivo and in vitro competition assays were normalized to the input ratios.
Nucleotide sequence accession number.
The nucleotide sequence corresponding to the complete varA coding region has been assigned GenBank accession no. AF071105.
RESULTS
Cloning of a gacA homolog from V. cholerae O1 classical strain O395.
Since GacA homologs have been found in a variety of gram-negative bacteria and have been shown to be important virulence determinants, we examined whether V. cholerae contains a gacA homolog as well. To do this, degenerate primers corresponding to conserved amino acid stretches of GacA-related proteins from P. aeruginosa, S. typhimurium, and E. coli were used to amplify an expected 390-bp fragment by using V. cholerae DNA as template. The DNA sequence of this PCR product was determined, and analysis using the BLAST computer program revealed this sequence to be approximately 60 to 70% similar to GacA homologs from a variety of gram-negative bacteria. DNA blot analysis showed that the 390-bp PCR product hybridized to an approximately 7-kb HindIII fragment and to a 3-kb SacI fragment of V. cholerae O395 chromosomal DNA (data not shown). A size-selected genomic DNA library in pBR322 containing approximately 7-to-8-kb HindIII genomic fragments from O395 was probed with the 390-bp PCR product to obtain a clone, pSW7H3, containing the full-length gene encoding the putative GacA homolog. Partial DNA sequence analysis of pSW7H3 and DNA sequence analysis using BLAST showed that pSW7H3 encodes a putative 214-amino-acid protein with 60.46% identity to GacA from P. aeruginosa, 75.8% identity to SirA from S. typhimurium, and 74.9% identify to UvrY from E. coli (data not shown). In addition, BLAST analysis of a predicted 201-nucleotide open reading frame containing an initiation codon overlapping by a single nucleotide with the termination codon of the putative V. cholerae gacA gene revealed 68% identity to a DNA repair protein encoded by uvrC of E. coli, which is immediately downstream of the E. coli uvrY gene (data not shown) (34, 46). The conserved genomic organization of the gacA-uvrC operon structure is also observed in Pseudomonas species (25, 42) and in S. typhimurium (22).
Disruption of V. cholerae gacA homolog decreases CT production and results in lack of autoagglutination.
To determine if the gacA homolog in V. cholerae plays a role in virulence, we constructed two gacA homolog mutant strains as described in Materials and Methods and then tested the mutants for the production of CT and the ability to autoagglutinate (which requires expression of functional TCP) (52). The first mutant (strain SW400-12) contains a disruption of the gacA homolog by insertion of a pGP704 derivative which is likely to exert a polar effect on the downstream uvrC homolog. The second mutant (strain SW33S), which was constructed to eliminate polarity effects on downstream genes, contains a replacement of nearly 88% of the gacA homolog open reading frame by a Kmr cassette.
Wild-type O395 and the gacA homolog mutants were grown overnight at 30°C in LB medium with a starting pH of 6.5, conditions known to induce the expression of genes subject to ToxR regulation in classical V. cholerae, and at 37°C in LB medium with a starting pH of 8.5, conditions known to repress the expression of genes under ToxR regulation (33). As summarized in Table 2, both gacA homolog mutants failed to autoagglutinate. Moreover, strain SW400-12 (containing the polar gacA homolog) and strain SW33S (containing the nonpolar gacA homolog) exhibited approximately 60- and 10-fold reductions in the amount of CT relative to the wild type, respectively. The lower amount of CT produced by SW400-12 compared to SW33S may reflect polar effects on downstream genes in SW400-12. To verify further that the mutant phenotypes of SW400-12 and SW33S were consequences of the disruption of the gacA homolog, a revertant of SW400-12, which had lost the suicide plasmid integrated in the gene, was obtained as described in Materials and Methods. This latter strain, SW8, regained the ability to autoagglutinate and to produce near wild-type levels of CT (Table 2).
TABLE 2.
Expression of CT and autoagglutination phenotypes of O395, varA mutants, and a varA revertant following growth in ToxR-inducing and ToxR-repressing conditions
| Strain | ToxR-inducing conditionsa
|
ToxR-repressing conditionsb
|
||
|---|---|---|---|---|
| CT productionc | Agg phenotyped | CT production | Agg phenotype | |
| Expt 1 | ||||
| O395 | 11.57 ± 1.3 | + | <0.001 | − |
| SW400-12 | 0.20 ± 0.03 | − | <0.001 | − |
| SW8 | 5.59 ± 0.85 | + | <0.001 | − |
| Expt 2 | ||||
| O395 | 16.25 ± 2.0 | + | ND | − |
| SW400-12 | 0.25 ± 0.09 | − | ND | − |
| SW33S | 1.77 ± 0.41 | − | ND | − |
ToxR-inducing conditions were 30°C with a starting pH of 6.5 and 85.5 mM NaCl with moderate aeration.
ToxR-repressing conditions were 37°C with a starting pH of 8.5 and 171 mM NaCl with moderate aeration.
Expressed in micrograms per milliliter at OD600.
Autoagglutination phenotype of the cultures. +, clumping of cells; −, no clumping of cells. All strains were grown for 16 to 24 h prior to assaying for CT production and inspection for agglutination. CT values from experiments 1 and 2 are means ± standard deviations for three samples per strain. ND, not determined.
Based on the mutant phenotypes of SW400-12 and SW33S, the locus encoding the GacA homolog in V. cholerae was renamed varA, for virulence associated regulator. The results presented in this section indicate that VarA represents a previously unknown V. cholerae virulence factor.
Disruption of varA downregulates tcpA message and TcpA protein levels and decreases CT production.
To determine if lack of autoagglutination in the varA mutants, SW400-12 and SW33S, was due to a decrease in TCP expression, the mRNA and protein levels corresponding to the major pilin subunit gene, tcpA (51), were examined. Figure 1 shows a three- to fourfold reduction in the level of the 800-nucleotide tcpA transcript (6) in mutant strain SW400-12 under ToxR-inducing conditions compared to O395, whereas no detectable tcpA mRNA was detected in either O395 or the mutant under ToxR-repressing conditions. Figure 2 shows that the level of the 20.5-kDa TcpA protein was reduced in both the polar (SW400-12) and nonpolar (SW33S) varA mutants relative to wild type (O395) (lanes 1, 3, and 5). Because TcpA and the production of CT are coregulated (51), CT production was assayed in the cultures used for the immunoblot analysis. As expected, the varA mutant strains displayed reduced levels of CT relative to the wild type (Fig. 2, compare lanes 3 and 5 to lane 1). Importantly, the varA+ revertant of SW400-12 (SW8) regained wild-type levels of TcpA protein and CT under ToxR-inducing conditions (Fig. 2, lane 7), consistent with nearly full restoration of autoagglutination and CT production (Table 2). As expected, TcpA protein was undetectable in cultures grown under ToxR-repressing conditions for any of the strains tested (Fig. 2, even-numbered lanes). As an additional control, no TcpA protein was detected in strain CS2-1, which carries a TnphoA disruption in tcpA (51), under ToxR-inducing or -repressing conditions (Fig. 2, lanes 9 and 10), confirming that the 20-kDa band does in fact represent TcpA and not a cross-reactive protein. The reduction in CT production in the CS2-1 mutant under ToxR-inducing conditions (lane 9) is likely due to polar effects on toxT transcription from the insertion of TnphoA upstream (6). Taken together, these results indicate that VarA is required for full expression levels for both CT and TCP.
FIG. 2.

Western analysis of TcpA protein and CT production in O395 and varA derivatives following growth under ToxR-inducing (+) and ToxR-repressing (−) conditions. Total cell protein from each strain was adjusted for culture density prior to being run on an SDS–12% polyacrylamide gel followed by electroblotting to nitrocellulose and probing with polyclonal rabbit anti-TcpA. The 20.5-kDa TcpA protein is indicated by the arrow. O395 (lanes 1 and 2); nonpolar varA, SW33S (lanes 3 and 4); polar varA, SW400-12 (lanes 5 and 6); polar varA revertant, SW8 (lanes 7 and 8); O395 tcpA::TnphoA, CS2-1 (lanes 9 and 10). Numbers at left are Kaleidoscope protein molecular mass standards (kDa) from Bio-Rad. Numbers at bottom represent the CT amounts in the supernatant (expressed in micrograms per milliliter at OD600). nd, not determined.
Interaction of VarA with the ToxR virulence control system.
The ToxR virulence regulon is composed of a set of genes under the control of the transcriptional activator, ToxR, which activates the expression of a second regulator, ToxT, in response to environmental signals, such as pH and temperature, in classical V. cholerae (47). ToxT in turn activates transcription of tcpA (6) and ctxAB (14). Since VarA modulates expression of TcpA and CT, we examined whether transcription of varA was subject to control by ToxR as well. As shown in Fig. 3, RNA blot analysis with a varA-specific probe revealed a single relatively large hybridizing RNA species in the wild-type strain O395 that is consistent with the predicted size of a transcript containing varA as well as the downstream uvrC homolog (46). A transcript of approximately similar size and intensity was also present in the toxR mutant, indicating that transcription of varA does not require ToxR (Fig. 3, lanes 1 and 3). Indeed, varA was transcribed under environmental conditions (37°C, pH 8.5) that usually repress the expression of genes that require ToxR (Fig. 3, lanes 2 and 4). Transcription of varA appeared to be regulated in response to temperature and pH as varA mRNA was reduced but not absent in ToxR-repressing conditions (lanes 2 and 4). By comparison, the effect of environmental signals on tcpPH transcription was more dramatic, as reported previously (7) (Fig. 3, lanes 5 and 6).
FIG. 3.
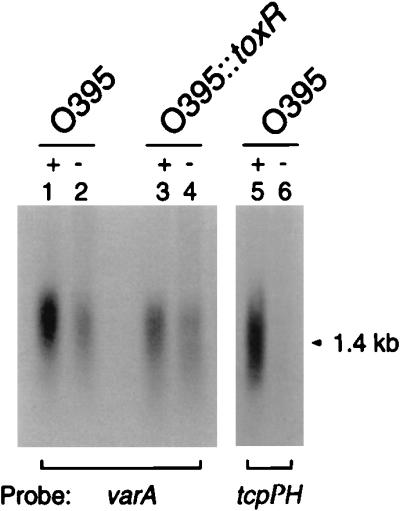
Northern blot containing 10 μg of total RNA from O395 and O395::toxR cultures grown under ToxR-inducing (+) and ToxR-repressing (−) conditions and probed for the varA or tcpPH message. O395 (lanes 1 and 2) and O395::toxR (lanes 3 and 4) are probed with varA. A parallel set of RNA samples of O395 (lanes 5 and 6) run on the same gel was probed with tcpPH (arrow indicates the 1.4-kb tcpPH transcript).
In S. typhimurium, the GacA homolog, SirA, regulates the transcription of hilA, which encodes an activator needed for invasion (22). Since HilA is reported to be a homolog of ToxR based on the similarity of the amino-terminal domain of HilA to the DNA-binding domain of ToxR (4), we examined whether VarA regulates the transcription of toxR and whether VarA affects ToxR protein levels. RNA blot analysis showed that a toxR-specific transcript was detected in both wild-type O395 and in the varA mutant SW400-12 under both ToxR-inducing and -repressing conditions (data not shown), indicating that toxR transcription does not require VarA and, as shown previously, that transcription of toxR is independent of environmental signals (12, 36). To determine if varA affects the expression of ToxR, the Western blot shown in Fig. 2 was reprobed with an anti-ToxR antibody, and no apparent changes in the level of the 32.5-kDa ToxR protein (33) in the varA mutant relative to O395 under ToxR-inducing or -repressing conditions were seen (data not shown). This result is consistent with the RNA blot data and indicated that production of the toxR message and ToxR protein does not require VarA.
Because TcpA and CT expression are reduced in a varA mutant and because, as described in the preceding paragraph, we had demonstrated that VarA does not appear to regulate toxR, it appeared likely that VarA acts downstream of ToxR. We thus carried out experiments to determine whether overexpression of toxT or tcpPH could suppress the varA mutant phenotype. ToxT is believed to activate the tcpA and ctxAB promoters (6, 14), and TcpPH is thought to activate the toxT promoter (7, 17). Consistent with this proposed circuitry, tcpPH expressed from an inducible promoter can activate a toxT::lacZ reporter construct independent of ToxR (17). As shown in Table 3, when a plasmid carrying toxT expressed from an inducible promoter was introduced into the varA mutant SW400-12, CT production and autoagglutination under ToxR-inducing conditions were restored to wild-type levels. Similarly, overexpression of tcpPH from a constitutive promoter in SW400-12 restored nearly 50 to 90% of CT production and the ability to autoagglutinate under ToxR-inducing conditions.
TABLE 3.
Expression of toxT and tcpPH from plasmids restores virulence factor expression to a varA mutant
| Strain | ToxR-inducing conditionsa
|
|
|---|---|---|
| CT production | Agg phenotype | |
| Expt 1 | ||
| O395 | 11.7 ± 2.3 | + |
| SW400-12 | 0.3 ± 0.2 | − |
| SW400-12 (pMT5K)b | 18.6 ± 7.7 | + |
| Expt 2 | ||
| O395 | 6.2 ± 1.6 | + |
| SW400-12c | 0.23 | − |
| SW400-12 (pBBR1MCS)c | 0.24 | − |
| SW400-12 (pSW1.4-1) | 5.4 ± 1.0 | + |
| SW400-12 (pSW1.4-5) | 3.0 ± 0.4 | + |
See footnotes to Table 2 for a description of these data. CT production values in experiment 1 are means ± standard deviations for five samples per strain.
Value is consistent with the approximately 1.9-fold-higher CT levels seen previously with toxT overexpressed from pMT5 relative to wild-type O395 (7). One millimolar IPTG was used to induce expression of toxT from a tac promoter in pMT5K.
Values extrapolated from ratios of CT production by SW400-12 and SW400-12 (pBBR1MCS) relative to that of wild-type O395 determined from another experiment. CT production values in experiment 2 are means ± standard deviations for triplicate samples per strain.
In summary, these results suggest that VarA modulates CT production and TcpA expression in a ToxR independent manner, most likely upstream of the toxT promoter.
In vivo colonization assay.
TcpA has been shown to be required to establish colonization in both an infant mouse colonization model (10, 51) and in human volunteer studies (18). It seemed likely that the decrease in TcpA in varA mutants would result in a decrease in colonization efficiency. To assess the role of varA in bacterial colonization in an infant mouse model, in vivo mixed infection assays with the nonpolar varA mutant SW33S were performed. As shown in Table 4, the varA mutant exhibited a competitive index of 0.17 in vivo, an approximately sixfold reduction in colonization efficiency compared to that of the wild-type strain O395. In contrast to these in vivo results, the in vitro competitive index for SW33S was 0.75, suggesting no appreciable growth defect in vitro. These results demonstrate that varA is a virulence determinant in V. cholerae that is needed for efficient colonization. Thus, VarA not only regulates the expression of TCP and the production of CT in vitro but it is likely to regulate expression of TCP in vivo as well.
TABLE 4.
Competitive index of the varA mutant straina
| Strain | Genotype | n | Competitive index
|
|
|---|---|---|---|---|
| In vivo | In vitro | |||
| O395 | Wild type | 5 | 1.3 ± 0.19 | 0.83 ± 0.12 |
| SW33S | O395 varA::kanr | 7 | 0.17 ± 0.1 | 0.75 ± 0.07 |
Competitive index (C.I.) is defined as the output ratio of mutant to wild-type bacteria divided by the input ratio of mutant to wild-type bacteria. All in vivo and in vitro competitions were done with CG842, a Lac− derivative of O395 as the competing strain. n, number of mice used in each competition study. The in vivo C.I. are means ± standard deviations for the number of mice indicated. The in vitro C.I. was the average of triplicate samples of both wild-type and mutant strains.
Quantitative assessment of pilus structural integrity.
The decrease in TCP expression in SW400-12 is consistent with the decrease in colonization efficiency in the infant mouse model. To quantitatively assess the ability of the varA mutant to assemble surface TCP, we utilized a phage, CTXΦ, which uses TCP as a receptor (54). Specifically, we performed a CTXΦ transduction assay after overnight growth in ToxR-inducing conditions. The cultures were also examined for the autoagglutination phenotype and as expected, wild-type O395 autoagglutinated, whereas the varA mutant, SW400-12, did not. Wild-type O395 showed a 100-fold higher CTXΦ transduction frequency than the varA mutant SW400-12 (the values were 0.67 × 10−1 and 0.58 × 10−3, respectively), which is consistent with the reduced amounts of TCP in the mutant.
To further evaluate surface TCP expression, we examined the varA mutant SW33S and the wild-type strain O395 by electron microscopy using primary polyclonal rabbit anti-TcpA antibodies and a gold-conjugated secondary antibody to decorated bundled pili. Wild-type O395 bacteria whose pili had been labelled with immunogold were readily located, whereas the varA mutant revealed many fewer microscopic fields that contained labelled pili (not shown). When we were able to find labelled pili in samples from the varA mutant, the pilus bundles were indistinguishable from those of wild-type O395 (not shown), indicating that varA mutants produce fewer pili but that the ones that are produced are structurally intact.
DISCUSSION
It is known that expression of virulence factors by a variety of bacterial pathogens in response to environmental signals is controlled by members of the two-component family of signal transduction proteins (31, 50). One of the components is a histidine kinase sensor which is often a transmembrane receptor whose kinase activity is modulated in response to environmental signals. The other component is the cytoplasmic response regulator which upon activation via phosphorylation at a conserved aspartate residue in the N-terminal domain by its cognate sensor kinase, controls gene expression at targeted promoters through the DNA-binding domain in the C terminus. In this paper, we describe the identification of a V. cholerae gene, varA, which is highly homologous to the two-component cytoplasmic response regulator gacA from various Pseudomonas species and show that varA is an important V. cholerae virulence factor.
V. cholerae O395 varA mutants showed decreased production of two major virulence factors in V. cholerae, TcpA, the major subunit of pili which is absolutely required for colonization, and CT, which is responsible for the severe diarrheal symptoms. We verified that the mutant phenotypes of SW400-12 were due to the disruption of the varA locus by isolating a varA+ revertant, SW8, and showing that it regained wild-type levels of CT production and TcpA protein expression and the ability to autoagglutinate. In addition, the reduced levels of CT production and lack of autoagglutination in a nonpolar varA mutant, SW33S, provided further evidence that the mutant phenotypes were a consequence of the inactivation of the varA gene. We were not able to complement the polar varA mutant, SW400-12, with varA cloned in plasmids with either low (pLAFR3) or moderate (pBBR1MCS, pBR322) copy numbers (data not shown); it is possible that the copy number of the cloning vectors used in the complementation experiments may have adversely affected the expression or function of varA, a proposed transcription factor. In P. aeruginosa, gacA carried on a plasmid also failed to complement a gacA mutation in a mouse and plant model of infection (38).
When we compared CT production in a polar versus a nonpolar varA mutant, we found that the polar mutant made about sevenfold less CT. It is possible that this is due to a polar effect on the downstream gene, a uvrC homolog. uvrC encodes an excision nuclease involved in repairing damaged DNA, and a polar mutation in gacA of P. fluorescens renders the cell highly sensitive to UV irradiation (25). Likewise, SW400-12 was highly sensitive to UV irradiation, whereas SW33S exhibited wild-type levels of resistance to UV irradiation (data not shown). Recently, an additional phenotype has been ascribed to uvrC; a uvrC mutant of the human enteropathogen, Yersinia enterocolitica, produced reduced amounts of invasin protein and was impaired in its ability to invade tissue culture cells (3).
The virulence control system in V. cholerae is a complex network of intertwining regulatory pathways. ToxR and ToxS activate the toxT promoter, and ToxT in turn activates the ctxAB and tcpA promoters. Downstream of tcpA are genes involved in pilus biogenesis as well as the toxT gene (which is encoded within the TCP cluster) (reviewed in reference 47). In addition to ToxR-dependent expression from the toxT promoter, Brown and Taylor (6) have shown that transcription of toxT also occurs as a read-through transcript from the upstream tcpA promoter, suggesting an autoregulatory loop whereby toxT regulates its own transcription through activating the tcpA promoter. Gel shift assays did not detect ToxR binding to the tcpA promoter, suggesting that ToxR activation of the TCP operon is mediated through ToxT. In addition, another pair of regulatory proteins, TcpP/H, is involved in the activation of the TCP operon, and ToxRS and TcpPH act synergistically to activate the toxT promoter (17).
We found that overexpression of toxT from an inducible promoter completely restored wild-type levels of CT production and the ability to autoagglutinate in the varA mutant SW400-12. This indicates that varA most likely affects the expression of the ToxR regulon upstream of toxT expression and is consistent with previous studies which indicate that ToxT is the last regulator within the hierarchy of the ToxRST regulatory cascade (7, 8, 13, 17). In addition, we found that overexpression of tcpPH from pSW1.4-1 restored CT production in SW400-12 to near wild-type levels (87%). A likely explanation is that overexpression of tcpPH leads to increased activation of the toxT promoter and ToxT in turn activates the expression of TcpA and CT. Preliminary results suggest that VarA does not activate transcription from the tcpPH promoter.
Because TCP has been documented to be required for colonization, it is not surprising to see that a decrease in TcpA protein in the varA mutant affected the efficiency of colonization. The nonpolar mutant showed an approximately sixfold reduction in colonization ability in the infant mouse gut compared to wild-type O395. The colonization defect of the varA mutant was not as severe as those seen for strains carrying various mutations in tcpA, which show a decrease of 2 to 4 orders of magnitude in their ability to colonize (10, 41). Rather, the colonization defect seen in the varA mutant is similar in magnitude to that in strains carrying mutations in acf (accessory colonization factors) genes (37), which are located immediately downstream of the TCP gene cluster. Transcription of acfB appears to be tcpA promoter dependent (6).
The modestly reduced colonization ability of the varA mutant is consistent with the varA mutant showing approximately 100-fold reduction in susceptibility to transduction by CTXΦ relative to O395 and producing significantly lower levels of bundled pili in immunoelectron micrographs. However, it is not clear whether the colonization deficiency of the varA mutant is due simply to a decrease in the amount of surface pili or the function of the surface pili or both. Although we cannot rule out a minor defect in pilin morphogenesis, the electron micrographs did not show any gross morphological differences.
Mutations in gacA homologs of pathogenic Pseudomonas species and S. typhimurium result in pleiotropic defects in secretion of proteases and other extracellular virulence factors (22, 28, 39, 40, 42, 43). Furthermore, sirA in S. typhimurium positively regulates the prgHIJK operon, which encodes components of a sec-independent type III secretion apparatus (22). The general secretion pathway (GSP) in V. cholerae is required for extracellular secretion of chitinase, CT, and hemagglutinin-protease (35, 45); mutations in several genes within this pathway affect secretion of these factors (44). It is possible that varA could affect expression of genes within the GSP to disrupt secretion of extracellular factors like CT. However, proteolytic activity as measured spectrophometrically in a Hide powder blue assay (52) did not show the varA mutant to be defective in general protease secretion (data not shown). This suggests that if the varA mutant has a secretion defect, the defect does not globally affect the GSP.
Although two-component regulators are common systems in a variety of bacterial pathogens, surprisingly, there have been no cited reports of these regulators in V. cholerae until recently. Lee et al. (26) reported the identification of the vieSAB operon of V. cholerae, where vieS exhibited similarity to sensor kinase components and vieA and vieB were similar to response regulators. Although it was not clear what role the vieSAB operon plays in the cholera infection process, vieB was specifically induced during colonization of the mouse intestine. Recent studies by Chiang and Mekalanos (9) using signature-tagged transposon mutagenesis to screen for random mutants affecting colonization in the infant mouse cholera model identified two genes whose products are involved in transfer of phosphate, pta (encodes phosphotransacetylase), and ptfA (encodes a phosphotransferase). Both genes are involved in modulating expression of the ToxR regulon; pta and ptfA mutants produce low amounts of CT and no TcpA. Pta controls synthesis and degradation of acetyl phosphate, which serves as a phosphodonor to some response regulators (29, 55). Since ToxR does not contain a phosphoacceptor domain, the authors postulated that perhaps Pta acts on the ToxR regulon by phosphorylating some unknown response regulator. It is tempting to speculate that VarA could be a substrate for phosphorylation by Pta. In Pseudomonas species, the cognate sensor kinase for GacA is LemA (11, 21, 27, 42). It is likely that a LemA homolog probably exists in V. cholerae as well.
ACKNOWLEDGMENTS
This work was supported by NIH grants RO1 AI34968 (S.B.C.) and T32 AI07410 and T32 AI07061 (S.M.W.) and by a grant to M.G.H. from Hoechst AG.
S.M.W. thanks Ed Ryan for providing pMT5K and members of the Mekalanos lab for providing valuable technical advice.
REFERENCES
- 1.Albright L M, Huala E, Ausubel F M. Prokaryotic signal transduction mediated by sensor and regulator protein pairs. Annu Rev Genet. 1989;23:311–336. doi: 10.1146/annurev.ge.23.120189.001523. [DOI] [PubMed] [Google Scholar]
- 2.Ausubel F M, Brent R, Kingston R E, Moore D E, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1995. [Google Scholar]
- 3.Badger J L, Miller V L. Expression of invasion and motility are coordinately regulated in Yersinia enterocolitica. J Bacteriol. 1998;180:793–800. doi: 10.1128/jb.180.4.793-800.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bajaj V, Hwang C, Lee C A. hilA is a novel ompR/toxR family member that activates the expression of Salmonella typhimurium invasion. Mol Microbiol. 1995;18:715–727. doi: 10.1111/j.1365-2958.1995.mmi_18040715.x. [DOI] [PubMed] [Google Scholar]
- 5.Bolivar F, Rodriguez R L, Greene P J, Betlach M C, Heynecker H W, Boyer H W, Crosa J W, Falkow S. Construction and characterization of new cloning vehicles. II. A multipurpose cloning system. Gene. 1977;2:95–113. [PubMed] [Google Scholar]
- 6.Brown R C, Taylor R K. Organization of tcp, acf, and toxT genes within a ToxT-dependent operon. Mol Microbiol. 1995;16:425–439. doi: 10.1111/j.1365-2958.1995.tb02408.x. [DOI] [PubMed] [Google Scholar]
- 7.Carroll P A, Tashima K T, Rogers M B, DiRita V J, Calderwood S B. Phase variation in tcpH modulates expression of the ToxR regulon in Vibrio cholerae. Mol Microbiol. 1997;25:1099–1111. doi: 10.1046/j.1365-2958.1997.5371901.x. [DOI] [PubMed] [Google Scholar]
- 8.Champion G A, Neely M N, Brennan M A, DiRita V J. A branch in the ToxR regulatory cascade of Vibrio cholerae revealed by characterization of toxT mutant strains. Mol Microbiol. 1997;23:323–331. doi: 10.1046/j.1365-2958.1997.2191585.x. [DOI] [PubMed] [Google Scholar]
- 9.Chiang S L, Mekalanos J J. Use of signature-tagged transposon mutagenesis to identify Vibrio cholerae genes critical for colonization. Mol Microbiol. 1998;27:797–806. doi: 10.1046/j.1365-2958.1998.00726.x. [DOI] [PubMed] [Google Scholar]
- 10.Chiang S L, Taylor R K, Koomey M, Mekalanos J J. Single amino acid substitutions in the N-terminus of Vibrio cholerae TcpA affect colonization, autoagglutination, and serum resistance. Mol Microbiol. 1995;17:1133–1142. doi: 10.1111/j.1365-2958.1995.mmi_17061133.x. [DOI] [PubMed] [Google Scholar]
- 11.Corbell N, Loper J E. A global regulator of secondary metabolite production in Pseudomonas fluorescens Pf-5. J Bacteriol. 1995;177:6230–6236. doi: 10.1128/jb.177.21.6230-6236.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.DiRita V J. Co-ordinate expression of virulence genes by ToxR in Vibrio cholerae. Mol Microbiol. 1992;6:451–458. doi: 10.1111/j.1365-2958.1992.tb01489.x. [DOI] [PubMed] [Google Scholar]
- 13.DiRita V J, Neely M, Taylor R K, Bruss P M. Differential expression of the ToxR regulon in classical and El Tor biotypes of Vibrio cholerae is due to biotype-specific control over toxT expression. Proc Natl Acad Sci USA. 1996;93:7991–7995. doi: 10.1073/pnas.93.15.7991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.DiRita V J, Parsot C, Jander G, Mekalanos J J. Regulatory cascade controls virulence in Vibrio cholerae. Proc Natl Acad Sci USA. 1991;88:5403–5407. doi: 10.1073/pnas.88.12.5403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gardel C, Mekalanos J J. Alterations in Vibrio cholerae motility phenotypes correlate with changes in virulence factor expression. J Bacteriol. 1996;64:2246–2255. doi: 10.1128/iai.64.6.2246-2255.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hanahan D. Studies on transformation of Escherichia coli with plasmids. J Mol Biol. 1983;166:557–580. doi: 10.1016/s0022-2836(83)80284-8. [DOI] [PubMed] [Google Scholar]
- 17.Häse C C, Mekalanos J J. TcpP protein is a positive regulator of virulence gene expression in Vibrio cholerae. Proc Natl Acad Sci USA. 1998;95:730–734. doi: 10.1073/pnas.95.2.730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Herrington D A, Hall R H, Losonsky G, Mekalanos J J, Taylor R K, Levine M M. Toxin, toxin-coregulated pili, and the toxR regulon are essential for Vibrio cholerae pathogenesis in humans. J Exp Med. 1988;168:1487–1492. doi: 10.1084/jem.168.4.1487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Higgins D E, DiRita V J. Transcriptional control of toxT, a regulatory gene in the ToxR regulon of Vibrio cholerae. Mol Microbiol. 1994;14:17–29. doi: 10.1111/j.1365-2958.1994.tb01263.x. [DOI] [PubMed] [Google Scholar]
- 20.Holmgren J. Comparison of the tissue receptors for Vibrio cholerae and Escherichia coli enterotoxins by means of gangliosides and natural cholera toxin. Infect Immun. 1973;8:851–859. doi: 10.1128/iai.8.6.851-859.1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hrabak E M, Willis D K. The lemA gene required for pathogenicity of Pseudomonas syringae pv. syringae on bean is a member of a family of two-component regulators. J Bacteriol. 1992;174:3011–3020. doi: 10.1128/jb.174.9.3011-3020.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Johnston C, Pegues D A, Hueck C J, Lee C A, Miller S I. Transcriptional activation of Salmonella typhimurium invasion genes by a member of the phosphorylated response-regulator superfamily. Mol Microbiol. 1996;22:715–727. doi: 10.1046/j.1365-2958.1996.d01-1719.x. [DOI] [PubMed] [Google Scholar]
- 23.Kovach M E, Phillips R W, Elzer P H, Roop R M N, Peterson K M. pBBR1MCS: a broad-host-range cloning vector. BioTechniques. 1994;16:800–802. [PubMed] [Google Scholar]
- 24.Laemmli U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 25.Laville J, Voisard C, Keel C, Maurhofer M, Défago G, Haas D. Global control in Pseudomonas fluorescens mediating antibiotic synthesis and suppression of black root rot of tobacco. Proc Natl Acad Sci USA. 1992;89:1562–1566. doi: 10.1073/pnas.89.5.1562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lee S H, Angelichio M J, Mekalanos J J, Camilli A. Nucleotide sequence and spatiotemporal expression of the Vibrio cholerae vieSAB genes during infection. J Bacteriol. 1998;180:2298–2305. doi: 10.1128/jb.180.9.2298-2305.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Liao C-H, McCallus D E, Fett W F. Molecular characterization of two gene loci required for production of the key pathogenicity factor pectate lyase in Pseudomonas viridiflava. Mol Plant-Microbe Interact. 1994;7:391–400. doi: 10.1094/mpmi-7-0391. [DOI] [PubMed] [Google Scholar]
- 28.Liao C H, McCallus D E, Wells J M, Tzean S S, Kang G Y. The repB gene required for production of extracellular enzymes and fluorescent siderophores in Pseudomonas viridiflava is an analog of the gacA gene of Pseudomonas syringae. Can J Microbiol. 1996;42:177–182. doi: 10.1139/m96-026. [DOI] [PubMed] [Google Scholar]
- 29.McCleary W R, Stock J B. Acetyl phosphate and the activation of two-component response regulators. J Biol Chem. 1994;269:31567–31572. [PubMed] [Google Scholar]
- 30.Menard R, Sansonetti P J, Parsot C. Nonpolar mutagenesis of the ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigella flexneri entry into epithelial cells. J Bacteriol. 1993;175:5899–5906. doi: 10.1128/jb.175.18.5899-5906.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Miller J F, Mekalanos J J, Falkow S. Coordinate regulation and sensory transduction in the control of bacterial virulence. Science. 1989;243:916–922. doi: 10.1126/science.2537530. [DOI] [PubMed] [Google Scholar]
- 32.Miller V L, Mekalanos J J. A novel suicide vector and its use in construction of insertion mutations: osmoregulation of outer membrane proteins and virulence determinants in Vibrio cholerae requires toxR. J Bacteriol. 1988;170:2575–2583. doi: 10.1128/jb.170.6.2575-2583.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Miller V L, Taylor R K, Mekalanos J J. Cholera toxin transcriptional activator ToxR is a transmembrane DNA binding protein. Cell. 1987;48:271–279. doi: 10.1016/0092-8674(87)90430-2. [DOI] [PubMed] [Google Scholar]
- 34.Moolenaar G F, van Sluis C A, Backendorf C, van de Putte P. Regulation of the Escherichia coli excision repair gene uvrC, overlap between the uvrC structural gene and the region coding for a 24 kD protein. Nucleic Acids Res. 1987;15:4273–4288. doi: 10.1093/nar/15.10.4273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Overbye L J, Sandkvist M, Bagdasarian M. Genes required for extracellular secretion of enterotoxin are clustered in Vibrio cholerae. Gene. 1993;132:101–106. doi: 10.1016/0378-1119(93)90520-d. [DOI] [PubMed] [Google Scholar]
- 36.Parsot C, Mekalanos J J. Expression of ToxR, the transcriptional activator of the virulence factors in Vibrio cholerae, is modulated by the heat shock response. Proc Natl Acad Sci USA. 1990;87:9898–9902. doi: 10.1073/pnas.87.24.9898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Peterson K M, Mekalanos J J. Characterization of the Vibrio cholerae ToxR regulon: identification of novel genes involved in intestinal colonization. Infect Immun. 1988;56:2822–2829. doi: 10.1128/iai.56.11.2822-2829.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rahme L G, Stevens E J, Wolfort S F, Shao J, Tompkins R G, Ausubel F M. Common virulence factors for bacterial pathogenicity in plants and animals. Science. 1995;268:1899–1902. doi: 10.1126/science.7604262. [DOI] [PubMed] [Google Scholar]
- 39.Rahme L G, Tan M-W, Le L, Wong S M, Tompkins R G, Calderwood S B, Ausubel F M. Use of model plant hosts to identify Pseudomonas aeruginosa virulence factors. Proc Natl Acad Sci USA. 1997;94:13245–13250. doi: 10.1073/pnas.94.24.13245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Reimmann C, Beyeler M, Latifi A, Winteler H, Foglino M, Lazdunski A, Hass D. The global activator GacA of Pseudomonas aeruginosa PAO positively controls the production of the autoinducer N-butyryl-homoserine lactone and the formation of the virulence factors pyocyanin, cyanide, and lipase. Mol Microbiol. 1997;24:309–319. doi: 10.1046/j.1365-2958.1997.3291701.x. [DOI] [PubMed] [Google Scholar]
- 41.Rhine J A, Taylor R K. TcpA pilin sequences and colonization requirements for O1 and O139 Vibrio cholerae. Mol Microbiol. 1994;13:1013–1020. doi: 10.1111/j.1365-2958.1994.tb00492.x. [DOI] [PubMed] [Google Scholar]
- 42.Rich J J, Kinscherf T G, Kitten T, Willis D K. Genetic evidence that the gacA gene encodes the cognate response regulator for the lemA sensor in Pseudomonas syringae. J Bacteriol. 1994;176:7468–7475. doi: 10.1128/jb.176.24.7468-7475.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sacherer P, Défago G, Haas D. Extracellular protease and phospholipase C are controlled by the global regulatory gene gacA in the biocontrol strain Pseudomonas fluorescens CHA0. FEMS Microbiol Lett. 1994;116:155–160. doi: 10.1111/j.1574-6968.1994.tb06694.x. [DOI] [PubMed] [Google Scholar]
- 44.Sandkvist M, Michel L O, Hough L P, Morales V M, Bagdasarian M, Koomey M, DiRita V J, Bagdasarian M. General secretion pathway (eps) genes required for toxin secretion and outer membrane biogenesis in Vibrio cholerae. J Bacteriol. 1997;179:6994–7003. doi: 10.1128/jb.179.22.6994-7003.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sandkvist M, Morales V, Bagdasarian M. A protein required for secretion of cholera toxin through the outer membrane of Vibrio cholerae. Gene. 1993;123:81–86. doi: 10.1016/0378-1119(93)90543-c. [DOI] [PubMed] [Google Scholar]
- 46.Sharma S, Stark T F, Beattie W G, Moses R E. Multiple control elements for the uvrC gene unit of Escherichia coli. Nucleic Acids Res. 1986;14:2301–2318. doi: 10.1093/nar/14.5.2301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Skorupski K, Taylor R K. Control of the ToxR virulence regulon in Vibrio cholerae by environmental stimuli. Mol Microbiol. 1997;25:1003–1009. doi: 10.1046/j.1365-2958.1997.5481909.x. [DOI] [PubMed] [Google Scholar]
- 48.Skorupski K, Taylor R K. Positive selection vectors for allelic exchange. Gene. 1996;169:47–52. doi: 10.1016/0378-1119(95)00793-8. [DOI] [PubMed] [Google Scholar]
- 49.Stock J B, Ninfa A J, Stock A M. Protein phosphorylation and regulation of adaptive responses in bacteria. Microbiol Rev. 1989;53:450–490. doi: 10.1128/mr.53.4.450-490.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Stock J B, Stock A M, Mottonen J M. Signal transduction in bacteria. Nature. 1990;344:395–400. doi: 10.1038/344395a0. [DOI] [PubMed] [Google Scholar]
- 51.Taylor R K, Miller V L, Furlong D B, Mekalanos J J. Use of phoA gene fusions to identify a pilus colonization factor coordinately regulated with cholera toxin. Proc Natl Acad Sci USA. 1987;84:2833–2837. doi: 10.1073/pnas.84.9.2833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Toder D S, Gambello M J, Iglewski B H. Pseudomonas aeruginosa LasA: a second elastase under the transcriptional control of LasR. Mol Microbiol. 1991;5:2003–2010. doi: 10.1111/j.1365-2958.1991.tb00822.x. [DOI] [PubMed] [Google Scholar]
- 53.Vieira J, Messing J. The pUC plasmids, an M13mp7-derived system for insertion mutagenesis and sequencing with synthetic universal primers. Gene. 1982;19:259–268. doi: 10.1016/0378-1119(82)90015-4. [DOI] [PubMed] [Google Scholar]
- 54.Waldor M K, Mekalanos J J. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science. 1996;272:1910–1914. doi: 10.1126/science.272.5270.1910. [DOI] [PubMed] [Google Scholar]
- 55.Wanner B L. Is cross regulation by phosphorylation of two-component response regulator proteins important in bacteria? J Bacteriol. 1992;174:2053–2058. doi: 10.1128/jb.174.7.2053-2058.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]