

Abstract
Primordial germ cells (PGCs) are the founder cells of the germline. The ability to generate PGC-like cells (PGCLCs) from pluripotent stem cells has advanced our knowledge of gametogenesis and holds promise for developing infertility treatments. However, generating an ample supply of PGCLCs for demanding applications such as large-scale genetic screens has been a limitation. Here, we demonstrated that simultaneous overexpression of 4 transcriptional factors - Nanog and three PGC master regulators Prdm1, Prdm14 and Tfap2c - in suspended mouse epiblast like cells (EpiLCs) and formative embryonic stem cells (ESCs) results in efficient and cost-effective production of PGCLCs. Nanog overexpression enhanced the PGC regulatory network, suppressed differentiation of somatic lineages, and maintained PGC fate. PGCLCs generated in this manner have transcriptomes similar to in vivo PGCs and are more advanced than cytokine-induced PGCLCs. These differentiated PGCLCs could be sustained over prolonged periods of culture and could differentiate into spermatogonia-like cells in vitro. We exploited this platform to conduct a bulk epigenome-wide CRISPRi screen to identify genes that decreased the efficiency of PGCLC formation when downregulated. One of the identified candidates was Ncor2, a transcriptional repressor that acts via recruitment of Class I and Class IIa histone deacetylases (HDACs). Consistently with this finding, the HDAC inhibitors valproic acid (VPA; an anti-convulsant) and sodium butyrate (SB; a widely-used dietary supplement) suppressed PGCLC differentiation. Furthermore, exposure of developing mouse embryos to SB or VPA caused hypospermatogenesis. This work demonstrates the feasibility of this platform to efficiently conduct large-scale functional screens of genes, chemicals, or other agents that may impact germline development.
Introduction
In mammals, gametogenesis is a complex and long process that is initiated by the specification of primordial germ cells (PGCs), the precursors of oocytes and sperm. Unraveling the mechanisms of PGC development is crucial for understanding key processes such as sexually dimorphic epigenetic inheritance and the bases of infertility, a relatively common health condition impacting ~15% of all couples (1). In mammals, PGCs specified in the embryonic epiblast respond to extrinsic and intrinsic signaling pathways including bone morphogenetic protein (BMP) and WNT signaling. In mice, BMP4 activates WNT3, and subsequently a master mesodermal transcription factor (TF) brachyury (also known as T). T activates critical early germline markers Prdm1 (also known as Blimp1), Prdm14, and Tfap2c (also known as AP2γ) (2). Prdm1 is expressed in the precursors of PGCs, which then stimulates Prdm14 and Tfap2c transcription to activate the germline pathway. The core TF network containing Prdm1, Prdm14, and Tfap2c is critical for mouse PGC specification; it represses somatic differentiation while driving reacquisition of a pluripotency network and epigenetic reprogramming (2). After migration to and colonization of the genital ridges, male germ cells differentiate into gonocytes. Thereafter, they become spermatogonial stem cells that subsequently seed recurrent rounds of mitotic expansion yielding spermatogonia that enter meiosis, and eventually, haploid spermatozoa.
Characterization of these gene regulatory pathways in PGCs formed the basis for creating mouse primordial germ cell-like cells (PGCLCs) from pluripotent stem cells in vitro (2). Pluripotency has at least three distinct states that mirror early embryonic development: naïve or ground state ESCs resembling the preimplantation epiblast; formative ESCs resembling early post-implantation gastrulating epiblast; and primed epiblast stem cells (EpiSCs) resembling late post-implantation gastrulating epiblast (3–5). Naïve ESCs typically are cultured with inhibitors of fibroblast growth factor and mitogen-activated protein kinase (FGF/MEK) as well as glycogen synthase kinase 3β (GSK3β) in the presence of leukemia inhibitory factor (LIF; “2i+LIF” medium). EpiSCs self-renew when exposed to Activin A and FGF2. Naïve ESCs do not readily respond to germ cell inductive stimuli, while primed EpiSCs display limited germline competence. By modulating activin and WNT signaling, a formative state has been stably captured in vitro (4–7). In this intermediate pluripotency state, formative ESCs possess dual competency for mouse chimera formation (when introduced into host blastocysts) and direct responsiveness to PGC specification.
The ability to model germ cell development in culture holds great potential for applications such as dissecting the genomic regulation of gametogenesis, and identifying genetic causes of infertility that would be important for assisted reproductive technologies (ART) involving gene corrections and optimization of in vitro gametogenesis (IVG). However, these applications remain challenging due to limited scalability of PGCLC generation. Conventional methods involve a transient conversion from naïve ESCs to germline-competent epiblast-like cells (EpiLCs), which represent a formative state. Aggregated EpiLC clusters are then exposed to high concentrations of BMPs and supporting cytokines, leading to their differentiation into PGCLCs (8). Alternatively, PGCLC specification can be induced by overexpressing three TFs (Prdm1, Prdm14, and Tfac2c), and to a lesser extent, two TFs (Prdm1 + Tfap2c or Prdm14 + Tfap2c) or even a single TF (Prdm14), in EpiLC aggregates (9). Nanog, a core pluripotency factor expressed in PGCs, is not essential for emergence of PGCs or germline transmission, but its deletion results in a significant reduction in PGC numbers both in vitro and in vivo (10). Nanog is also expressed in naïve ESCs, and its expression undergoes a transient downregulation as cells transition into the EpiLC state (8,11). The overexpression of Nanog can promote efficient PGCLC differentiation by binding endogenous enhancers of Prdm1 and Prdm14, even without BMP4 and associated cytokines (12). Furthermore, Nanog overexpression induces cytokine-free PGCLC specification by repressing Otx2, encoding a TF essential for anterior neuroectoderm specification (13,14).
Promoting cell proliferation is another strategy for producing a large number of PGCs/ PGCLCs. However, extended culture of mouse PGCs promotes dedifferentiation into pluripotent embryonic germ cells (EGCs) (15,16). Saitou and colleagues found that Rolipram and Forskolin, which stimulate cAMP signaling, enables expansion of PGCLCs up to ~50-fold in the presence of SCF (stem cell factor) on m220 feeders (stromal cells expressing a membrane-bound form of human SCF), and the expanded PGCLCs can contribute to spermatogenesis after transplantation to neonatal testes (17–19). Nevertheless, PGCLCs cultured under these conditions cannot be expanded for more than 7 days. Although these methods have been instrumental in understanding and improving PGCLC formation, they are associated with either low efficiency or high costs (for reagents such as cytokines).
While several genetic causes of infertility have been identified (20,21), pinpointing specific epigenetic factors that perturb gene expression is more challenging. Sperm counts have been reported to have declined worldwide since the mid 1900s - possibly due to environmental factors introduced after the industrial revolution - but there are major concerns regarding data analysis and interpretations underlying these claims (22). Exposure to certain chemicals during fetal development, such as endocrine disruptors, can cause transgenerational alterations in epigenetic landscapes and hinder reproductive function and fertility in both males and females (23–26). During PGC development, there is a global loss of repressive histone marks such as H3K9me2 and an increase of active histone marks such as H3K4me3 and H3K27ac (27,28). These dynamic epigenetic landscapes are crucial, and exposure to specific environmental toxins or chemicals that interfere with the epigenetic profile may impact PGC development (29,30). Histone acetylation and deacetylation regulate gene expression and repression, respectively. Nuclear receptor co-repressor 2 (NCOR2) is a well-studied nuclear receptor co-repressor that facilitates histone deacetylation (31,32). There are 18 histone deacetylases (HDACs) that are evolutionally conserved across all eukaryotes and can be divided into four classes (33). Both Class I (HDAC1, 2, and 3) and Class IIa (HDAC4, 5, 7, and 9) are known to interact with NCOR2 complexes to help downregulate gene expression (31,32). Trichostatin A, an inhibitor of Class I and II HDACs (HDACi), promotes PGC dedifferentiation into EGCs, suggesting that histone deacetylation is important for maintaining the PGC state (34).
Here, we report a simple, scalable and economical approach to induce the differentiation of PGCLCs from formative ESCs and EpiLCs at scale by overexpressing four TFs (Prdm1, Prdm14, Tfap2c and Nanog). The resulting PGCLCs exhibit transcriptional and epigenetic profiles resembling those of late (E11.5) PGCs, and could be further differentiated into spermatogonia-like cells. Using this platform, we performed a CRISPR inhibition (CRISPRi) screen and identified several epigenetic factors that impacted the efficiency of PGCLC specification and proliferation, including Ncor2. Furthermore, we found that the exposure to Class I and Class IIa HDAC inhibitors attenuated PGCLC formation in vitro and germ cell formation in mice exposed during development. Our in vitro differentiation system holds promise for facilitating high-throughput functional genomic studies of germ cell development, characterization of potential infertility-causing genetic variants, and improving IVG.
Results
Establishing a cytokine-free platform for efficient production of PGCLCs from EpiLCs.
To establish an efficient, cytokine-free system for generating PGCLCs, we began with a mouse ESC line containing enhanced GFP (eGFP) under the regulatory control of Stella (also called Dppa3/Pgc7), a PGC marker (35), and validated the ability of this line to form germline-transmitting chimeric mice after blastocyst microinjection (Fig. S1A–D), demonstrating its pluripotency. The Stella-eGFP reporter enables real-time visualization of PGCLC development from ESCs in vitro. Next, based on the finding that Nanog promotes transition of ESCs to PGCLCs by binding the enhancer regions of Prdm1 and Prdm14 and inducing their expression (12), we introduced a stably-integrated doxycycline (Dox)-inducible Nanog. To test the effectiveness of this transgene, we utilized a high efficiency enhancer activity detection system (36) to derive ESC lines containing targeted integrations of a lacZ reporter under the control of Prdm1 or Prdm14 enhancers into the safe harbor H11 locus. Dox treatment of embryoid bodies (EBs; ESCs>EpiLCs>EBs) derived from these lines triggered β-galactosidase production, indicating the effectiveness of NANOG-stimulated transcription (Fig. S1E–H). Besides promoting expression of Prdm1 and Prdm14, NANOG represses Otx2, a TF that promotes differentiation of pluripotent cells towards a neuroectodermal fate (12,14). Mutant cells lacking OTX2 enter the germline from an EpiLC state with high efficiency, even in the absence of the essential PGCLC-promoting cytokine signals (37). In summary, NANOG plays a crucial role in PGCLC differentiation by promoting the expression of PGC core regulatory TF-encoding genes Prdm1 and Prdm14, while simultaneously inhibiting the somatic fate inducer Otx2 (Fig. S1I).
Overexpression of Prdm1, Prdm14 and Tfap2c (hereafter called “3TF”) can induce PGCLCs without cytokines, however, the efficiency of this differentiation system is relatively low with no increase in the fraction of cells becoming PGCLCs after day 4 post-induction (9,38). Because Nanog can induce PGCLCs independently of BMP signaling and performs better when combined with cytokines (12), we hypothesized that simultaneous overexpression of NANOG and downstream targets (i.e. Prdm1, Prdm14, Tfap2c and Nanog, hereafter called “4TF”) in EpiLCs might render PGCLCs robustly. We isolated several Dox-inducible clonal cell lines harboring combinations of these TFs in Stella-eGFP ESCs that also contained rtTAM2 (reverse tetracycline-controlled transactivator mutant; Fig. 1A and B). The resulting ESCs, cultured in 2i+LIF medium, were differentiated to EpiLCs by 2 days of Activin A and bFGF exposure. To induce PGCLC differentiation, the EpiLCs aggregates were treated with Dox instead of cytokines. We first tested whether ectopic expression of one or more TFs drove EpiLC-to-PGCLC differentiation in U-bottom 96-well plates. Overexpression of Nanog alone led to ~35% Stella-eGFP+ cells in day 6 EBs. This percentage increased when Nanog was combined with one of the core TF transgenes (i.e., Nanog+Prdm1, Nanog+Prdm14 and Nanog+Tfap2c), even though these genes are upregulated by NANOG during PGCLC differentiation (12). Remarkably, simultaneous overexpression of all 4TFs caused ~80% of cells to become eGFP+ (Figs. 1A–E).
Figure 1. Construction of the 4TF-induced PGCLC differentiation platform.

A. Percentages of day 6 PGCLCs (Stella-eGFP+ cells) differentiated from Dox-inducible ESCs harboring distinct combinations of TFs. Each individual clonal line is represented by a dot. B. The process for constructing 4TF-inducible ESCs. The 4TFs and rtTA are in bold. C. Schema illustrating the 4TF-induced PGCLC differentiation from ESCs. D. Representative EBs with eGFP signal in the presence or absence of Dox during the 6-day period. Scale bar in D represents 100 μm. E. FACS patterns of Stella-eGFP+ cells (indicated by dashed lines) during differentiation. F. Relative fluorescence density of H3K9me2 and K3K27me3 in GFP−, GFP dim and GFP bright cells from Dox-induced day 6 EBs. G. Relative mRNA expression of Dnmt3a and Dnmt3b in ESCs, EpiLCs and day 6 EBs. H. Bisulphite sequence analysis of methylated cytosine in the DMRs of the imprinted genes in Stella-eGFP+ and Stella-eGFP− cells in day 6 EBs. White and black circles represent unmethylated and methylated cytosines, respectively. Data in G are represented as the mean ± SEM. Data in F and G were analyzed using one-way ANOVA with Tukey’s post hoc test.
In vivo, PGCs undergo substantial epigenetic reprogramming, characterized by the loss of H3K9me2 and the acquisition of H3K27me3, as well as global erasure of DNA methylation (2). To test the fidelity of epigenetic changes in PGCLCs produced in our 4TF system, we examined histone modifications, CpG methylation status of imprinted genes, and capacity for dedifferentiation into pluripotent EGCs. Differentiated cells with higher eGFP fluorescence displayed lower H3K9me2 and higher H3K27me3 labeling (Fig. 1F). The mRNA levels of epigenetic modifiers Dnmt3a and Dnmt3b were increased in EpiLCs and subsequent downregulated in EBs (Fig. 1G), which aligns with previous findings (8). Bisulfite sequencing of PGCLCs (eGFP+ cells) revealed reduced methylation in differentially methylated regions for both paternally (H19) and maternally (Snrpn) imprinted loci, while eGFP− cells (presumably somatic-like) exhibited a more methylated state (Fig. 1H). In addition, PGCLCs could de-differentiated to EGCs in vitro when transferred to 2i+LIF medium (Fig. S2). Collectively, our data indicate that simultaneously overexpression of 4TFs effectively induces the differentiation of EpiLCs into PGCLCs.
Transcriptomic properties of 4TF-induced PGCLCs
To examine the fidelity of our 4TF-induced PGCLCs, i.e., Stella-eGFP+ cells (Fig. S3), RNA sequencing (RNA-seq) was performed and the transcriptomes were compared to those of in vivo germ cells (encompassing PGCs at E9.5, E10.5, and E11.5 and male/female germ cells at E12.5 and E13.5) and cytokine (Ck)-induced PGCLCs (those at days 2, 4 and 6 after induction, referred to as Ck_D2/4/6PGCLC) (17,39,40). Principal component analysis (PCA) showed that 4TF PGCLCs at days 2, 4 and 6 (4TF_D2/4/6PGCLC) clustered tightly as a group separate from naïve ESCs, EpiLCs and Ck_D2PGCLCs, representing a transient state towards the acquisition of a PGC-like state from EpiLC/ epiblast states (9) (Fig. 2A). Notably, 4TF-induced PGCLCs grouped closely with PGCs at E10.5 and E11.5, while Ck_D4/6PGCLCs grouped closely with E9.5 and E10.5 PGCs (Fig. 2A). These data suggest that 4TF-induced PGCLCs are more akin to late PGCs than Ck_D2/4/6PGCLCs. We next applied two-sided rank–rank hypergeometric overlap (RRHO) analysis to compare the gene expression signatures between samples in a threshold-free manner (41). We observed robust overlap in both up- and down-regulated transcripts amongst differentiating cells in both Ck- and 4TF-induced groups relative to either ESCs or EpiLCs (Fig. 2B). In the Ck-induced group, ~60% of genes exhibited concordant differential expression (53% when compared to ESCs and 67% when compared to EpiLCs) between D2 and D6 PGCLCs. The concordance was higher for the D2 and D6 4TF-induced group (74% and 78%, respectively). In both induction systems, D4 and D6 PGCLC shared around 84% concordantly regulated genes (Fig. 2C). These data demonstrated that 4TF-induced PGCLCs at early stages exhibited greater consistency in their transcriptomic changes compared those in Ck-induced PGCLCs. Given that only microarray (not RNA-seq) datasets were available for 3TF- and Nanog-induced PGCLCs, we performed a Spearman correlation coefficient analysis to compare their similarities with Ck-induced PGCLCs. Microarray datasets of 3TF- and Nanog-induced PGCLCs (referred to as 3TF_D2/4PGCLC and Nanog_D4PGCLC; datasets for 3TF_D6PGCLC and Nanog_D2/6PGCLC are not available) were compared to the corresponding microarray datasets of Ck-PGCLCs generated using the same platforms. The 3TF_D2/4PGCLCs showed high correlation with Ck_D4/6PGCLCs but not Ck_D2PGCLCs; Nanog_D4PGCLC showed a similar correlation to Ck_D4PGCLCs (Fig. 2D). Consistent with the PCA, 4TF_D2/4/6PGCLCs are highly correlated with Ck_D4/6PGCLCs but not Ck_D2PGCLCs (Fig. 2D).
Figure 2. 4TF-induced PGCLCs share similar transcriptomes with cytokine- and 3TF-induced PGCLCs.

A. PCA of indicated transcriptomes from 4TF- and cytokine (Ck)-induced PGCLCs, in vivo germ cells at indicated times of gestation, EpiLCs and ESCs. “D” = days of induction. PGCLCs were isolated by flow sorted for diagnostic markers. B. Comparison of differential expression profiles between D6PGCLC (x-axis) and D2/4PGCLC (y-axis) with RRHO2 maps. The extent of overlap transcriptome was represented by each heatmap colored based on -log10(p-value) from a hypergeometric test. Each map shows the extent of overlapped upregulated genes, versus ESCs or EpiLCs, in the bottom left corner, whereas shared downregulated genes are displayed in the top right corners. C. Percentage of concordant genes encompassing both up and down regulated genes compared with D6PGCLCs. D. Spearman correlation coefficients between PGCLCs. 4TF_D2/4/6PGCLCs were compared with RNA-seq datasets of Ck_D2/4/6PGCLCs. 3TF_D2/4PGCLCs were compared with microarray datasets of Ck_D2/4/6PGCLCs, generated using the Affymetrix Mouse Genome 430 2.0 Array. Nanog_D4PGCLC was compared with microarray dataset of Ck_D4PGCLC, generated using the Illumina MouseWG-6 v2.0 expression beadchip. N.A., not available. E. GSVA score of Wnt signaling pathways (10 out of the 11 pathways in total were selected, with one specific to the planar cell polarity being excluded) in D2PGCLC between Ck-, 3TF- and 4TF-induced conditions (Ck_D2, 3TF_D2 and 4TF_D2). ESC, EpiLC and PGCLC in each group were used to calculate GSVA score and only the scores of D2PGCLC were displayed. F. Heatmap of selected genes associated with WNT signaling and targets. G. GSVA score of neurodevelopment-related pathways (see Fig. S4C) in D2PGCLC between Ck-, 3TF- and 4TF-induced conditions (Ck_D2, 3TF_D2 and 4TF_D2). H. Heatmap of selected genes associated with neurodevelopment.
In vivo, PGC specification follows WNT-dependent induction, which activates the master mesodermal TF, T. This triggers the core TF network for PGCs (i.e., Prdm1, Prdm14, and Tfap2c). Whereas the Ck induction system relies on WNT signaling, the 3TF and Nanog induction systems do not. To investigate WNT signaling activity during 4TF-induced PGCLC differentiation, we compared the transcriptomes of Ck- and TF-induced PGCLCs. Gene set variation analysis (GSVA) revealed strong enrichment of Wnt signaling pathways in Ck_D2PGCLCs (including downstream target genes Wnt3, Wnt5a, Wnt5b, Axin2, Mixl1, Cdx2, and T), as previously reported (9), but not in 3TF_D2PGCLCs or in our 4TF_D2PGCLCs (Fig. 2E and F). Overall, unlike Ck induction, the 4TF, 3TF and Nanog induce PGCLCs from EpiLCs independently of the BMP-WNT-T pathway.
Unlike our 4TF system, the 3TF system exhibits low differentiation efficiency that did not increase after day 4 after induction (9,38). To explore the impact of Nanog overexpression in driving efficient differentiation into PGCLCs, we performed Gene set enrichment analysis (GSEA). This revealed that pathways predominantly related to neurodevelopment were suppressed in D2PGCLCs compared to EpiLCs (Figs. S4A and B), and Otx2 was involved in the majority of these pathways (Fig. S4C). Otx2 plays a crucial role in brain development and exhibits reciprocal antagonism with Nanog. Induction of transgenic Nanog in EpiLCs shifts the balance between these mutual antagonists, favoring Nanog (42). When Otx2 levels decrease sufficiently, this enables NANOG to influence the regulatory elements controlling genes associated with PGC-specific TFs (42). GSVA revealed that neurodevelopment-related pathways and genes were highly repressed in 4TF_D2PGCLCs, while 3TF_D2PGCLCs showed a modest level of suppression. Notably, these pathways and genes were enriched in Ck_D2PGCLCs (Fig. 2G and H, and S4D).
To better characterize the transcriptome landscapes during PGCLC differentiation, we applied k-Means analysis of top 500 differentially expressed genes (DEGs) and defined one cluster (C4) representing genes expressed in 4TF_D2/4/6PGCLCs (Fig. 3A). Gene Ontology (GO) analysis revealed that 11 of the top 20 pathways in this cluster were associated with reproductive processes (Fig. 3B). GSVA confirmed enrichment in germ cell development terms such as gamete generation, oogenesis, piRNA metabolic process, meiosis and reproduction were enriched in 4TF-induced but not Ck- and 3TF- induced PGCLCs compared to ESCs and EpiLCs (Fig. 3C and S5). GSEA revealed upregulation of genes related to gamete generation, meiotic cell cycle and reproduction in 4TF-induced PGCLCs. Processes related to piRNA processing and meiotic cell cycle also enriched in 3TF-induced D4PGCLCs. However, upregulation of these gene sets were barely observed in Ck- and Nanog-induced PGCLCs (Fig. 3D and Fig. S6). We next compared the DEGs among Ck-, 3TF- and 4TF-induced PGCLCs. Across all three groups, the naïve pluripotency gene Klf4 was highly expressed in ESCs, while genes related to the formative state (Otx2, Pou3f1, Fgf5, Dnmt3a and Dnmt3b) were highly expressed in EpiLCs (Fig. 3E). Early PGC-related genes (e.g., Prdm1, Prdm14, Nanog, Dnd1, Tfap2c, Dppa3 and Itgb3) were notably expressed in Ck_D4PGCLC, 3TF_D2PGCLC and 4TF_D2PGCLC, indicating that both 3TF and 4TF induction systems cause molecular differentiation more rapidly than cytokine induction. Some PGC genes, including Rhox9, Rhox6, Rhox5 and Gm9, were highly expressed in Ck_D6PGCLC, 3TF_D4PGCLC and 4TF_D4PGCLC. Interestingly, late PGC, meiosis and piRNA-related genes were consistently expressed in 4TF-induced PGCLCs but not in Ck-induced PGCLCs (Fig. 3E and S7). Network analysis identified a regulatory axis facilitates the transition from early to late PGCLCs (Fig. S8; see Discussion).
Figure 3. Gene expression profiling of 4TFinduced PGCLCs.

A. Heatmap of k-Means clustering of variably expressed genes in ESCs, EpiLCs, and 4TF-induced D2/4/6PGCLCs (n = 500, k = 4). Genes were grouped into four clusters (“C1–4”) on the basis of expression similarity. B. Biological process GO term analysis of Cluster 4 (C4). Terms of germ cell development (highlighted by red) was highly enriched for terms in C4. C. Heatmap showing the average GSVA enrichment score of selected germ cell development pathways. D. Representative GSEA enrichment plots depicting significant enrichment of germ cell signatures in D6PGCLCs compared with ESCs. E. Heatmap of representative gene expression of pluripotency and germ cells among ESCs, EpiLCs and PGCLCs.
Further differentiation of 4TF-induced PGCLCs in vitro
We next examined the potential of PGCLCs to differentiate into advanced stages in vitro. D6PGCLCs (Stella-eGFP+ cells) were aggregated with dissociated neonatal testicular cells from germ cell deficient KitW/Wv mice in U bottom 96-well plates. Then, the cell aggregates were transferred to a trans-well plate and cultured in medium containing GNDF and bFGF that are essential for promoting spermatogonial growth (43) (Fig. 4A). After a two-week induction period, we conducted immunofluorescence (IF) staining to assess whether the aggregates express late PGC or spermatogonia proteins. We detected cell clusters that expressed NANOS3, DAZL and DDX4 within the aggregates (Figs. 4B). Over 70% of DDX4-positive cells also expressed GCNA (Germ cell nuclear antigen) (Fig. 4C), a robust marker of germ cells from E11 onwards (44,45). Interestingly, certain DDX4-positive cells also contained the spermatogonial marker PLZF (formally ZBTB16; Zinc finger and BTB domain-containing protein 16) (46) but not the Sertoli cell marker SOX9 (Fig. 4C). To test whether the germ cells can developed into advanced stages, the aggregates were cultured using a gas-liquid interphase method for an additional week in the medium containing KSR (knockout serum replacement) at 34°C (Fig. 4A) (47). Interestingly, cells expressing both GCNA and STRA8 were identified in the aggregates (Fig. 4D). These data suggest that the 4TF-induced PGCLCs have the capacity of further development in vitro.
Figure 4. Development of cells past PGCLC stage.
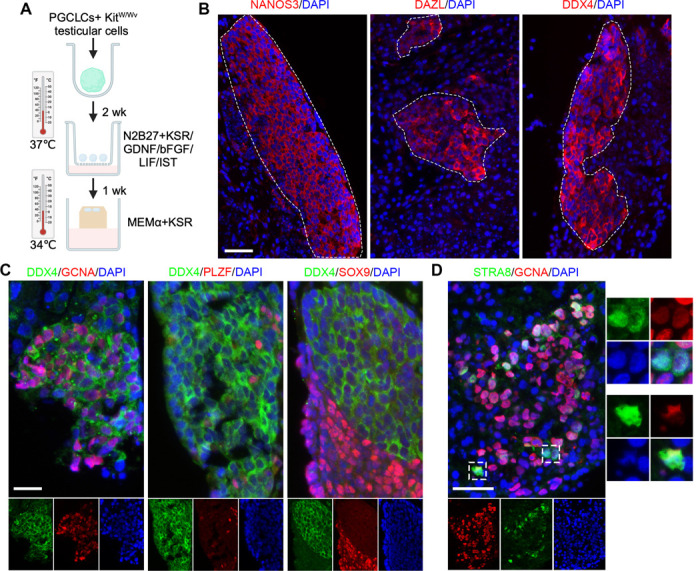
A. Further development of 4TF_D6PGCLCs by aggregation with testicular somatic cells isolated from Kitw/wv mice. The cultured aggregates were subjected to IF. B. Clusters of germ cells labeling by NANOS3, DAZL and DDX4 were identified from aggregates cultured after 2 weeks. Dashed lines denote regions with germ cell clusters. C. Dual IF staining of DDX4 with GCNA, PLZF and SOX9 in the aggregates after 2-week culture. D. Dual IF staining of GCNA and STRA8 in the aggregates cultured for an additional week. The boxed region are magnifications of representive dual positive cells. Scale bars in B, C and D represent 50 μm.
Scale-up differentiation and long-term culture of PGCLCs
To validate the Stella-eGFP transgene as an accurate marker of PGCLCs, we analyzed differentiating cultures with two additional PGC surface markers, ITGB3 (also known as CD61) and SSEA1 (8). Doubly positive cells were shown to differentiate into functional gametes (48). As expected, day 6 EBs were significantly enriched for ITGB3+/SSEA1+ and ITGB3+/Stella-eGFP+ cells compared to ESCs and EpiLCs (Fig. 5A). Next, we sought to determine the scalability of 4TF-induction system. We seeded 2×103 cells/ well to U bottom 96-well plates, 8×104 cells/well to untreated 12-well plates (suitable for suspension culture) and 1×106 cells to a single 100mm bacteriological Petri dish, then assessed differentiation in day 6 EBs. The percentages of both ITGB3+/SSEA1+ and ITGB3+/Stella-eGFP+ populations were highest in 12-well plate and lowest in 96-well plates (Fig. 5A and B). Suspecting that EB size influences differentiation efficiency, we measured the diameters of day 6 EBs. In the U bottom 96-well plates, EB sizes were homogeneous and by far the largest (~2.5× on average) amongst the culture formats (Fig. 5C). The results indicate an inverse relationship between efficiency of PGCLC differentiation and EB size, which is consistent with observation in the human PGCLC differentiation system (49). The scalability of the Nanog-only induction system was also observed by seeding varying numbers of EpiLCs into different plates/ dish. However, the percentages of both ITGB3+/SSEA1+ and ITGB3+/Stella-eGFP+ populations were 30% and 60% lower, respectively, than those in the 4TF induction system (Fig. S9A; 12 well plate cultures). EpiLCs represent a transient, germline-competent formative state, with day 2 EpiLCs demonstrating the highest efficiency in response to cytokine induction compared to day 1 or 3 EpiLCs (8). We found that 4TFs can also induce PGC fate in day 1 EpiLCs, albeit with lower efficiency than in day 2 EpiLCs (Fig. 5D and E). Nanog overexpression alone was insufficient to differentiate day 1 EpiLCs into PGCLCs (Fig. S9B and C).
Figure 5. Scale-up and long-term differentiation of 4TF-induced PGCLCs.

A. Representative FACS pattern of Itgb3+/Ssea1+ and Itgb3+/eGFP+ cells in ESCs, EpiLCs and day 6 EBs from U bottom 96-well plates, 12-well nontreated plates and 100mm bacteriological petri dishes. eGFP, Stella-eGFP. B. Quantification of PGCLC populations in day 6 EBs at different plates/dish. C. Boxplots showing the distribution of the diameter of day 6 EBs in different plates/dish. D. Induction of 4TF-induced PGCLCs from day 1 EpiLCs. E. Quantification of 4TF-induced PGCLC populations in day 6 EBs in the presence or absence of Dox. Day 1 EpiLCs were used for EB formation in non-treated 12-well plates. F. Schema depicting the process of EB passage. The dissociated EB cells were passaged every 4 days into each well of non-treated 12-well plates. G. Growth curves of long-term cultured EBs induced by 4TFs, Nanog or not. H. Doubling time of passaged EBs in 4TF- and Nanog-induced PGCLC differentiation systems. I. FACS pattern of Itgb3+/Ssea1+ and Itgb3+/eGFP+ cells induced by 4TFs in day 20 and 43 EBs cultured in 12-well non-treated plates. Data in B, C, E, G and H are represented as the mean ± SEM. Data in B and C were analyzed using one-way ANOVA with Tukey’s post hoc test. Data in E were analyzed using a two-tailed paired t test. Data in H were analyzed using a two-tailed unpaired t test. Scale bar in C represents 200 μm.
Next, we investigated the potential for maintaining PGCLCs over long periods. Day 6 EBs from both 4TF- and Nanog-induction systems were dispersed, and 8×104 cells/ well were seeded into a 12-well plate. The cells were passaged every 4 days in suspension (Fig. 5F). We successfully maintained the EBs for 10 continuous passages with Dox-induced transgene expression. However, cell numbers declined without Dox in both systems, indicating that TFs, particularly Nanog, are essential for sustaining EB cell proliferation (Fig. 5G). While the doubling times between 4TF- and Nanog-induced cells were comparable (Fig. 5H), only the 4TF system consistently maintained the PGC identity (>70% of cells), whereas nearly all cells in the Nanog-only system lost PGC marker expression (Fig. 5I; S9D and E). Lastly, we investigated the impact of culture plate type on PGCLC differentiation by seeding 8×104 cells/well onto treated 12-well plates designed for adherent culture. After differentiation, we observed the presence of attached cells, along with both floating and attached EBs (Fig. S10A). While PGCLCs were robustly induced, the differentiation efficiency was lower compared to those cultured in untreated plates (Fig. S10B and C), indicating that suspension culture is more effective in promoting PGCLC differentiation compared to adherent culture.
High throughput CRISPR perturbation screen identifies epigenetic regulators impacting PGC development.
A major motivation for developing an inexpensive, simple and scalable germ cell culture system was to conduct genome-scale functional genomic screens for genes involved in various aspects of germ cell biology and development. Epigenetic reprogramming is an especially important aspect of normal PGC development. To identify epigenetic regulators impacting PGC development, we designed a CRISPRi screening strategy to target all the genes with known or putative functions in epigenetic regulation (Fig. 6A). We built a custom lentiviral library encoding 7,360 sgRNAs targeting 701 “epigenetic” genes, plus 350 non-targeting sgRNAs as controls (Tables S1 and S2), and introduced these into a 4TF-inducible ESC line containing an integrated dCas9-KRAB (Krüppel-associated box) transgene (Fig. S11A–D). The library-infected ESCs contained 89% of all the sgRNAs in the library, with at least 5 sgRNAs present for 694 genes (Fig. S11E and Table S3). PGCLC differentiation was initiated in 100mm bacteriological Petri dishes (Fig. 6A), and both Stella-eGFP+ and Stella-eGFP− cells were isolated from dispersed day 6 EBs (Fig. S11F). In each resulting population and ESCs, sgRNAs were PCR amplified and then sequenced. Quality control measures indicated a high degree of uniformity and coverage of the sgRNA library in each cell population (Fig. S12). The MAGeCKFlute algorithm was utilized to calculate a beta score for every potential gene by comparing each population to the ESCs (50). Genes with a negative beta score suggests negative selection of the epigenetic factor within the cell population, whereas a positive beta score signifies positive selection. Using one standard deviation from the mean as a threshold, we identified 53 and 61 under- and over-represented genes, respectively, from the 701 queried genes (Fig. S13A and B; Table S4).
Figure 6. Application of 4TF-inducible system for high throughput screening epigenetic regulators.

A. Schema of CRISPRi library generation and validation is shown on the top. The uniformity of sgRNA library representation was tested by sequencing of batch-cloned plasmids and lentivirus-infected ESCs (colored cell clusters). On the bottom is a schema of the PGCLC differentiation protocol and CRISPRi screening process. Day 6 EBs were sorted into Stella-eGFP+ and Stella-eGFP− cells to generate count representatives for downstream analysis. Candidate gene Ncor2 and HDAC inhibitors SB and VPA were selected for validation. OE, overexpression. B. Experimental schema showing SB and VPA treatment on pregnant mice and analysis their pups. i.p., intraperitoneal. C. Testis weight of 2-month-old pups. n.s., no significant difference. D. Sperm counts. E. Histological analyses of 2-month-old testes and cauda epididymis. The boxed region are magnifications of atrophic seminiferous tubules. Data in C and D are represented as the mean ± SEM and were analyzed using one-way ANOVA with Tukey’s post hoc test. Scale bars in E represent 50 μm.
Transduction of sgRNAs targeting Ncor2, which encodes a nuclear receptor co-repressor that facilitates histone deacetylation (31,32), had the most negative impact on 4TF-induced PGCLC in our screen (Fig. S13 and Table S4). Treatment with Ncor2-targeting siRNA caused significant reductions in ITGB3+/Stella-eGFP+ cells (Fig. S14A–D), consistent with a previous observation from a Ck-induction system (51). NCOR2 contains three repression domains (RD1, RD2, and RD3) capable of interacting with various classes of HDACs (52) (Fig. S14E). Because one motivation of this project was to identify environmental agents (including drugs and dietary supplements) that might cause deleterious epigenetic germline perturbations, we tested the effects of two HDACi on PGCLC development: sodium butyrate (SB) and valproic acid (VPA). Both are short-chain fatty acids that inhibit HDACs by binding to the zinc-containing domains of these enzymes (Fig. S14E) (53). Treatment with VPA or SB resulted in a reduction in PGCLC and increased H3K27ac levels (Fig. S14F–I). To investigate why HDACi treatment dramatically decreased the efficiency of PGCLC differentiation, we conducted RNA-seq to analyze the transcriptomes of Stella-GFP+ and Stella-GFP− cells on day 6 of differentiation from control and HDACi-treated groups. Our findings revealed that HDACi treatment result in a loss of germline identity because the genes associated with somatic lineages were upregulated (Fig. S15 and S16; Text S1).
To assess the impact of histone deacetylation on in vivo PGC development, HDACi was administered to pregnant mice and their offspring were examined for potential reproductive defects. From E5.5 to E11.5, pregnant females received daily injections of either 600mg/kg or 800mg/kg of SB per body weight or 300mg/kg of VPA per body weight (Fig. 6B). All treated dams were viable and produced a comparable number of pups to the water-injected control group. Quantification of follicle numbers in 1-month-old ovaries revealed no significant differences compared to the controls (Fig. S17). In 2-month-old males, testis weights were unaffected by HDACi treatment during pregnancy (Fig. 6C). However, these animals exhibited a 30% reduction in sperm numbers in the SB groups and an 18% decrease in the VPA group (Fig. 6D). Histological analysis identified atrophic seminiferous tubules (1–3 per section of a whole testis), despite normal morphologies in the cauda epididymides (Fig. 6E). TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labeling) staining revealed no apoptotic cells within atrophic tubules (Fig. S18). These findings suggest that fetal exposure to SB and VPA leads to PGC loss, or possibly spermatogonial stem cell loss, contributing to hypospermatogenesis in adulthood.
Direct differentiation of PGCLCs from formative 4TF-inducible ESCs.
To determine if 4TFs can efficiently induce PGCLCs from formative ESCs, a pluripotent stem cell that directly responds to PGC specification, the 4TF-inducible ESCs were transformed to a formative state through exposure to bFGF, Activin A and GSK3β inhibitor (CHIR99021) (Fig. 7A) as described previously (5). Naïve ESCs typically grow as tightly packed colonies with a dome shape. Upon EpiLC induction, the cells quickly underwent a morphological conversion that includes flattening, diminished cell-cell interactions, and the formation of cellular protrusions. However, formative ESCs displayed distinctive morphologies (Fig. 7B). Immunolabeling of ZO1 (zonula occludens-1, also known as tight junction protein 1) revealed that formative ESCs and EpiLCs were more similar to one another than naïve ESCs (Fig. 7B). The regulation of Oct4 expression in naïve and primed pluripotent cells is differentially controlled by distal (DE) and proximal enhancers, respectively (54). The DE also exhibits activity in the formative state, albeit at a lower level compared to that in the naïve state (5). To monitor the exit from the naïve state during naïve-to-formative conversion, we compared the GFP signal intensity of Oct4-DE-GFP (“OG2”) iPSCs during this transition. Consistent with previous observations (5), the majority of formative iPSCs and EpiLCs were GFP+ but the average signal intensity was lower than that in naïve iPSCs (Fig. 7B). Collectively, these results confirm the establishment of formative ESCs/iPSCs from naïve cells.
Figure 7. Direct differentiation of PGCLCs from formative 4TF-inducible ESCs.

A. Schema for inducing PGCLCs from formative ESCs. B. Characterization of formative ESC derived from naïve ESCs. Top panel: representative morphologies of naïve ESCs, formative ESCs and EpiLCs. Middle panel: IF staining of tight junction protein ZO1 on naïve ESCs, formative ESCs and EpiLCs. Bottom panel: GFP expression in naïve and formative OG2 (Oct4-DE-GFP) iPSCs, and EpiLCs differentiated from naïve OG2 iPSCs. C. Representative FACS pattern of Itgb3+/Ssea1+ and Itgb3+/Stella-eGFP+ cells in formative ESCs and day 6 EBs with or without Dox treatment in 12-well non-treated plates. D. Quantification of PGCLC subpopulations after 4TF induction. E. IF staining of germ cell markers in cultured D6PGCLCs (Stella-eGFP+) differentiated from formative ESCs aggregated with testicular cells from Kitw/wv mice. F. RRHO analysis of transcriptomes from 4TF formative ESCs and EpiLCs compared to those from a published study (55). Sampled time points during differentiation are indicated (6h, 12h, 24h, and 48h). Data in D are represented as the mean ± SEM and analyzed using a two-tailed paired t test. Scale bars in top and bottom panels of B represent 100μm, and in middle panel of B and E represent 50 μm.
We next tested if formative 4TF-inducible ESCs respond directly to PGC specification. After removing the feeder layer, PGCLC differentiation was conducted by seeding 8×104 ESCs/well to untreated 12-well plates (Fig. 7A). FACS analysis of day 6 EBs revealed distinct ITGB3+/SSEA1+ and ITGB3+/Stella-eGFP+ populations with induction by Dox; such cells were not observed in undifferentiated formative ESCs and without induction by Dox (Fig. 7C and D). However, the efficiency was lower compared to that observed when EpiLC were induced to PGCLCs (Fig. 5A and B). Treatment with an FGF receptor (FGFR) inhibitor PD173074 was reported to significantly increase Ck-induced PGCLC formation from formative ESC via unknown mechanisms (5). However, this enhancement was not observed in 4TF-induced PGCLCs (Fig. S19). Interestingly, overexpression of Nanog alone is insufficient to direct formative ESC toward a PGC fate (Fig. S20). We next aggregated the D6PGCLC (Stella-eGFP+ cells) with somatic cells from KitW/Wv testes to assess further differentiation potential. Similar to EpiLC derived PGCLC (Fig. 4A), the formative ESC-derived PGCLCs demonstrated the capacity for further development (Fig. 7E). Collectively, these findings demonstrated that overexpressing 4TFs can directly and effectively induce differentiation of formative ESCs into PGCLCs.
To elucidate the differences in efficiency in generating PGCLCs from formative 4TF ESCs vs. day 2 EpiLCs, and to explore why Nanog alone is insufficient to drive differentiation of formative ESCs to PGCLCs, we compared the transcriptomes of our derived EpiLCs to those in a published dataset of formative ESCs cultured and differentiated under identical conditions (55). RRHO analysis indicated substantial similarities in gene expression changes during differentiation and a significant correlation between 48h EpiLC and naïve ESC transcriptomes, indicating a shared cell state (Fig. 7F). Interestingly, while formative ESCs overlapped with EpiLCs at various stages in co-upregulated genes, they did not show overlap in co-downregulated genes (Fig. 7F). This suggests that formative ESCs represent distinct sub-stages, leading to differences in PGCLC induction efficiencies.
Discussion
PGC development entails important regulatory and epigenetic events that lay the groundwork for subsequent sex-specific gametogenesis programs. Although in vitro PGCLC induction systems have facilitated molecular studies of the specification and developmental events of the germ cell lineage (9,12,37,56–60), current methodologies have shortcomings in terms of efficiency, scalability and long-term culture. Our 4TF-induction system addresses these limitations, having several advantages compared to Ck-, 3TF-, and Nanog-only inducible systems (Fig. S21). We demonstrated that simultaneous overexpression of Nanog, Prdm1, Prdm14 and Tfap2c in mouse EpiLCs or formative ESCs can render PGCLCs in a highly efficient and cost-effectively manner (Fig. S21). The induced PGCLCs exhibit late PGC hallmarks, and they can develop into spermatogonium-like cells in vitro. Overexpression of Nanog had the important effect of enhancing the PGC regulatory network, suppressing the formation of non-germline lineages and maintaining PGC fate, significantly improving PGCLC production.
Specification of mouse PGCs requires coordinated actions of Prdm1, Prdm14, and Tfap2c. Here, we found that forced overexpression of Nanog combined with one of these core TFs can drive PGCLC specification more efficiently that Nanog alone, and 4TFs yielded the highest efficiency, indicating additive or synergistic effects. Given that NANOG can bind and activate enhancers of Prdm1 and Prdm14 in EpiLCs in vitro (12) as we confirmed here, we conclude that Nanog overexpression strengthens the PGC core regulatory network in the 4TF system. NANOG also plays a role in suppressing somatic lineage induction development. For example, OTX2, a crucial TF for anterior neuroectoderm specification, shows reciprocal antagonism with NANOG and thus restricts mouse germline entry (37). 4TF-induced PGCLCs exhibited Otx2 downregulation, Nanog upregulation, and significant downregulation of neurodevelopment pathways in D2PGCLCs vs EpiLCs. However, these pathways were activated in Ck-induced PGCLCs or not fully repressed in 3TF-induced PGCLCs. Using Nanog-eGFP transgene iPSCs, Hayashi et al. demonstrated that Nanog was upregulated as early as day 2 in Ck-induced PGCLCs (8). The distinct kinetics of NANOG expression in the 4TF and Ck induction systems correlated with the repression and activation of neurodevelopment pathways, respectively. Lastly, NANOG is essential for maintaining PGC fate. Depletion of Nanog in PGCLCs showed decreased proliferation and increased apoptosis (10). Those findings indicate multifunctional roles of NANOG in driving differentiation into PGCLCs.
The transcriptional profiles of Ck- and 4TF-induced PGCLCs resembled PGCs at different stages. This disparity partially arises from the essential role of WNT signaling and the mesodermal program in the Ck-induction system, whereas the 4TF-induction system circumvents these requirements. Combining insights from previous studies (9,12), all PGCLC systems using TF overexpression (Nanog, 3TF and 4TF) drive differentiation independently of the BMP4-WNT-T pathway. The gene regulatory axis of early-to-late PGC development established in 4TF-induced PGCLCs propels their differentiation beyond what Ck achieved within the same timeframe. Consequently, in the 4TF but not Ck-induced PGCLCs, the late PGC regulatory network was triggered as indicated by expression of genes involved in piRNA processes (such as Piwil2/Mili, Tdrd5, Mov10l1, Tdrd7 and Tex19.2).
It was surprising that several meiosis genes were expressed in the 4TF-induced PGCLCs, including Stra8, Sycp1/2/3, Meiob and Smc1b. One possible explanation is that although the engineered ESCs were male, some fraction of cells may have initiated a meiotic transcription program as would normally occur in females absent sex determination signals that emanate from somatic cells in vivo. We also observed expression of Mael, involved in pachytene piRNA processes (namely LINE-element suppression) a gene expressed strongly in mouse spermatocytes and which is essential for normal spermatogenesis in mice (61). However, MAEL protein is present at substantial levels in fetal oocytes and primordial follicles in both humans and mice (62,63) where it is also involved in suppressing LINE-1 activity that contributes to fetal oocyte attrition. Regarding the mechanisms by which these meiosis genes are transcribed before clear evidence of meiocytes in the D6PGCLC cultures, network analysis gives a clue. During germ cell development, Tfap2c is activated by Prdm1 and Prdm14 (2), directly targeting Dmrt1 in PGCLCs (38,64). Human DMRT1 upregulates late PGC markers (such as DAZL and PIWIL2), while DAZL enhances DMRT1’s impact by repressing pluripotency and promoting expression of late PGC (65) and meiosis genes. This beginning of the meiosis transcriptional program in these cultures may precede developmental or cellular hallmarks of meiosis.
At present, gonadal and testicular somatic cells are needed for further differentiation of male PGCLCs (18,66,67). Seminiferous tubule-like structures can be constructed by aggregating Ck-induced PGCLCs with gonadal somatic cells under a gas-liquid interphase culture condition (18,67). Given that 4TF-induced PGCLCs exhibit more advanced development compared to Ck-induced PGCLCs, we opted for the readily accessible neonatal testicular somatic cells to facilitate PGCLC differentiation, along with key cytokines (i.e., GDNF and bFGF) crucial for spermatogonial proliferation. While no seminiferous tubule-like structures were observed, clusters of germ cells were formed and surrounded by somatic cells. Similar to previous observations (18,67), a subset of germ cells underwent further differentiation into cells with spermatogonium-like characteristics. Some germ cells displayed the early meiosis marker STRA8 after culture at 34°C, but further work is necessary to explore whether these are truly meiotic cells that can complete meiosis and progress to later stages of spermatogenesis in vivo or in vitro.
Unraveling the genetic and epigenetic factors governing gametogenesis is crucial for understanding causes of developmental or reproductive abnormalities. Though genomic analyses have characterized gene expression patterns and epigenetic modifications during germ cell development (68–70), the field has been hampered by the lack of high-throughput platforms for genomewide functional studies. Massively parallel CRISPR-based screens require large number of cells to accommodate libraries containing tens of thousands of sgRNAs. Previously, a screen of 199 epigenetic modifiers was performed using individual RNAi knockdowns under standard conditions, i.e., exposure of individual EBs to cytokines in 96-well plates (51). By contrast, the 4TF-inducible system enabled en masse gene knockdowns using Petri dishes and sequencing-based screening of 701 “epigenetic” genes for key roles in PGC development. Our study not only identified epigenetic events influencing the generation and development of PGCs, but also to draw links between epigenetic modifications and environmental factors (such as medications and dietary supplements) that might influence gametogenesis and possibly fertility.
The 4TF-induction system bypasses the need for BMP4 and WNT3, enabling direct programming of EpiLCs into PGCLCs. Consequently, our CRISPRi screen likely did not query epigenetic regulators involved during the period of BMP signaling and the activation of germline-specific TFs. As mentioned earlier, an RNAi-based epigenetic screen was described that utilized a Blimp1 reporter for PGCLC identification. Blimp1 expression is equivalent to ~E6.25, and they likely preferentially identified epigenetic genes that regulate the transition of epiblast cells to the PGC fate, as well as the repression of somatic networks. In contrast, our platform preferentially queried epigenetic factors that regulate PGC differentiation downstream of germline TF expression. Disruption of these factors holds the potential to impede or override the germline transcription network.
The dietary supplement SB is beneficial for intestinal homeostasis and energy metabolism by enhancing intestinal barrier function and mucosal immunity with its anti-inflammatory properties (71). However, the potential effects of maternal SB supplementation upon fetal development are poorly explored. VPA is a widely prescribed drug used to treat epilepsy, bipolar disorder, and migraines. There is an elevated risk of birth defects (such as cleft lip, heart and neural tube defects) in children born to women taking VPA during pregnancy (72), but possible impacts on germ cell development remain unknown. Our in vitro and in vivo results raise the possibility that fetal exposure to HDAC inhibitors can deplete PGCs, manifesting as atrophic seminiferous tubules and hypospermatogenesis in adulthood. However, the ovarian reserve was unaffected even though treatment occurred before sex determination. It is possible that compromised cells were preferentially lost in the process of fetal oocyte attrition, which normally eliminates up to 80% of oocytes (73,74). Alternatively, the molecular impacts of treatments may only manifest in prospermatogonia or spermatogonial stem cells. Although the recommended clinical doses of SB and VPA are lower than those utilized in our study, we cannot say if human fetuses are more or less sensitive to these drugs than mouse embryos with respect to germline development. Our findings contribute to an expanding body of evidence indicating that epigenetic factors and dietary choices can impact male fertility (75,76).
In mice, competence for germline specification distinguishes the formative state of pluripotency from the naïve and primed states (77). Studies using Ck-,3TF- and Nanog-induction systems have demonstrated that a formative state is essential for robust induction of a PGC-like state (8,9,12). Induction of 3TF in naïve ESC resulted in a unique phenotype characterized by intense activation of the Stella reporter without forming a PGC-like state. The transcriptomes of these Stella+ cells were close to those of ESCs, but not PGCLCs/PGCs (9). Since Nanog abundantly expressed in naïve ESC, the above result also indicates that induction of 4TFs in naïve ESCs fails to form PGCLCs. Given that both formative ESCs and EpiLCs respond directly to PGC specification, it was unclear whether exposure of formative ESCs to 4TF overexpression was compatible with efficient differentiation into PGCLCs. Recent advancements have identified four distinct sub-stages of formative pluripotent stem cells, exhibiting functional characteristics and WNT/ β-catenin signaling modulation (4–7). In our study, we cultivated formative ESCs using one of these protocols (5), and revealed that 4TFs can drive both formative ESCs and EpiLCs into PGCLCs. PGCLCs (Stella-eGFP+ cells) differentiated from formative ESCs exhibit similar further developmental potential with those from EpiLCs. Importantly, enables simplification of the PGCLC differentiation process, as formative ESCs can be stably maintained and would not require preliminary induction into the transient EpiLC-like state from naïve ESCs.
This scalable and efficient 4TF-induction system offers a valuable tool to conduct unbiased genetic screens impacting PGCLC formation. Besides CRISPRi as described here, this platform enables other CRISPR-based perturbation screens including knockouts, upregulation via CRISPR activation (CRISPRa), combinatorial disruptions, and single-cell transcriptomics as a phenotypic readout (78,79). Furthermore, a useful aspect of the 4TF system is that the derived PGCLCs persisted for prolonged periods in culture, enabling various studies involving, for example, large amounts of material for biochemical analyses, genetic manipulations initiated after cells reach this advanced stage, responses to drug or biologic exposures, or as a starting point for exploring further differentiation. Finally, considering that genetic causes underlie ~50% of human infertility (1), parallelized screening of genetic variants, particularly rare or minor alleles that are classified as variants of uncertain significance (VUS), is another potential application. Functionally interpreting the VUS within genes that essential for gametogenesis, along with achieving a comprehensive understanding of epigenetic networks, are important for genetic counseling, clinical applications, and potentially gene correction therapies for individuals to produce viable gametes in vitro or in vivo (1,80,81).
Materials and Methods
Mice and cell lines
All animal usage was approved by Cornell University’s Institutional Animal Care and Use Committee, under protocol 2004–0038 to J.C.S. Four- to eight-day-old KitW/Wv male mice generated by crossing C57BL/6J-KitWv/J and WB/ReJ KitW/J were used in this study. C57BL/6J-KitWv/J (#000049), WB/ReJ KitW/J (#000692), B6(Cg)-Tyrc−2J/J (#000058) and C57BL/6J (# 000664) mice were purchased from The Jackson Laboratory. The Stella-eGFP transgenic ESC line (Stella-eGFP ESCs) with Stella flanking sequences coupled to eGFP as the reporter was kindly provided by Prof. M. Azim Surani (35).
Vectors
To construct pPBhCMV1-Prdm1-pA and pPBhCMV1-Prdm14-pA, mouse Prdm1 and Prdm14 coding sequences were cloned into pPBhCMV1-Nanog-pA (a gift from M. Azim Surani) by PCR flanked with XhoI and NotI restriction sites. To construct targeting vectors containing enhancer-reporter transgenes, PCR-amplified sequences of mouse Prdm1 and Prdm14 enhancers (12) bearing terminal NotI restriction sites were cloned into PCR4-Shh::lacZ-H11 (Addgene, #139098). Plasmid clones were validated by Sanger sequencing. pPyCAG-Pbase and pPB-CAG-rtTA-IN (Addgene, #60612) were kindly provided by M. Azim Surani. pSpCas9(BB)-2A-PuroR (px459) V2.0 (Addgene, #62988), psPAX2 (Addgene, #12260), pMD2.G (Addgene, #12259), pLV-tetO-Tfap2c (Addgene, #70269), pLX_311-KRAB-dCas9 (Addgene, #96918) and pXPR_050 (Addgene, # 96925) were obtained from Addgene. Primer sequences are listed in Table S5.
Microinjection for chimera production
The ESCs were microinjected to B6(Cg)-Tyrc−2J/J blastocysts by standard methods in Cornell’s Transgenic Core Facility (82).
Derivation of genetically modified ESC clones
A piggyBac transposon expression system was used to stably integrate single copies of various vectors that were doxycycline (Tet) inducible. To establish the Tet-on expression system, pPBhCMV1-Prdm1-pA, pPBhCMV1-Prdm14-pA and pPBhCMV1-Nanog-pA vectors were transfected using TransIT®-LT1 Transfection Reagent (Mirus Bio™, MIR2304) into ESCs together with pPBCAG-rtTA and pPyCAG-PBase vectors. After 1 week of Geneticin™ (80 μg/ml; Life Technologies, 10131035) selection (all vectors except pPyCAG-PBase contain neomycin resistance genes), clones containing desired inserts were identified. Cells infected with pLV-tetO-Tfap2c lentivirus (described below) were selected by Zeocin® (InvivoGen, ant-zn-05) to identify transformants.
To construct ESC lines containing enhancer-reporters, the targeting vector PCR4-Shh::lacZ-H11 and H11 gRNA-expression plasmid px459 were transfected into cells bearing Tet-inducible Nanog along. After puromycin selection for 48 hours, single colonies were expanded and correctly targeted clones were identified using primers listed in Table S5.
ESC, iPSC culture and PGCLC differentiation
Mouse ESCs maintained on γ-irradiated feeder MEFs (mouse embryonic fibroblast cells) in DMEM containing 15% FBS, penicillin-streptomycin, NEAA, β-mercaptoethanol, 1,000U/ml LIF (Peprotech, 300–05). Naïve ESCs were cultured in 2i+ LIF medium containing 1×N2B27, 1 μM PD0325901 (Reprocell, 04–0006-02), 3 μM CHIR99021 (Reprocell, 04–0004-02) and 1,000 U/ml LIF on wells coated with 0.01% poly-L-ornithine (Sigma-Aldrich, P3655) and 10 ng/ml laminin (Corning, 354232).
For the formative state conversion, naïve ESCs and iPSCs were dissociated into single cells and plated on MEFs plate in 2i +LIF medium. Follow the protocol describe previously (5), the next day, medium was changed to formative medium (N2B27 medium containing 10 ng/ml bFGF, 10 ng/ml Activin A and 3 μM CHIR99021) for further culture. It usually takes about 2–4 passages for fully conversion. Formative ESCs and iPSCs were maintained in formative medium.
For PGCLC differentiation, EpiLCs were induced by naïve ESCs as described previously (48). Briefly, 1×105 ESCs were placed per well of a 12-well plate coated with 16.7 μg/mL bovine plasma fibronectin (Sigma-Aldrich, F1141) in N2B27 medium containing 20 ng/mL Activin A (Peprotech, 120–14E), 12 ng/mL bFGF (Gibco, 13256–029), and 1% knockout serum replacement (KSR, ThermoFisher, 10828010). The medium was changed the following day. A total of 48 hours after plating, 2×103 EpiLCs per well were seeded in a U-bottom 96-well plate (ThermoFisher Scientific, 268200) in GK15 medium (GMEM with 15% KSR, 0.1mM NEAA, 1mM sodium pyruvate, 0.1mM β-mercaptoethanol, penicillin-streptomycin, and 2mM L-glutamine) with Dox (Sigma, D9891). To induce PGCLCs from formative ESCs, after removing the feeder cells, 2×103 cells per well were seeded in the U-bottom 96-well plate. Alternatively, 8×104 cells per well were seeded in an untreated 12-well plate (ThermoFisher Scientific, 150200), or 1×106 cells were seeded into a 100mm Petri dish in GK15 medium. To sustain the PGCLCs, day 6 EBs in a 12-well plate were dissociated into single cells, and 8×104 cells per well were seeded in an untreated 12-well plate. The cells were passaged every 4 days. Transgenes were induced by addition of 2 μg/ml Dox at day 0 of PGCLC induction. Cells were cultured at 37 °C in a 5% CO2-95% air atmosphere.
Flow cytometry
To flow sort PGCLCs, single-cell suspensions were stained with 1 μg/ml propidium iodide (PI, Invitrogen, P3566) and filtered through a 70 μm cell strainer. eGFP+ cells were sorted using a BD FACSMelody 4-Way Sorter. To analyze the PGCLCs, suspended cells were washed with PBS and fixed in 70% ethanol. For staining, cells were washed in MACS buffer (1×PBS, 0.5%BSA and 2mM EDTA) and blocked with 5% goat serum (Sigma, NS02L) in MACS buffer at 4°C for 1 hour. Cells were then stained with PE (Phycoerythrin) mouse/rat anti-CD61 and Alexa Fluor® 647 mouse/human anti-SSEA1 antibodies (Table S6) at 4°C for 1 hour. Cells were then washed in MACS buffer twice and filtered through a 35 μm cell strainer (Falcon, 352235). The stained cells were analyzed using a BD FACSymphony A3 cytometer. FACS data were analyzed using FlowJo™ v10.4 or flowCore package in R.
Cell aggregate cultures for further differentiation
Sorted D6PGCLCs were mixed with dissociated neonatal testicular cells from KitW/Wv mice with 10:1 ratio and co-cultured for 2 days in a U bottom 96-well using GK15 medium. The aggregates were subsequently transferred to a trans-well plate in N2B27 medium containing 20 ng/ml bFGF, 40 ng/ml GDNF (PeproTech, 450–44), 1,000 U/ml LIF, 1×Insulin-Transferrin-Selenium (ThermoFisher Scientific, 41400045) and 15% KSR for additional 2 weeks. The aggregates were cultured at 37 °C in a 5% CO2-95% air atmosphere. Two weeks later, the aggregates were cultured on agarose gel stand in MEMα (Minimum essential medium α; ThermoFisher Scientific, 12571063) containing 15% KSR for an additional week, as described previously (47). The aggregates in this stage were cultured at 34 °C in a 5% CO2-95% air atmosphere.
Cell staining
ALP staining was performed using the Vector Red Alkaline Phosphatase Substrate Kit (Vector Laboratories, Burlingame, CA, USA) according to the manufacturer’s directions. LacZ staining was performed the Senescence β-Galactosidase Staining Kit (Cell Signaling Technology, #9860) according to the manufacturer’s directions. For TUNEL staining, sections were deparaffinized and performed TUNEL staining using the DeadEnd™ Fluorometric TUNEL System (Promega; G3250) following manufacturer’s instructions.
For cell IF staining, the cells growing on cover slides were washed with PBS, fixed in 4% paraformaldehyde (PFA) for 10 min at RT, washed twice with PBS. For section IF staining, cell aggregates were fixed in 4% PFA and embedded in paraffin. The 6 μm sections were deparaffinized and performed antigen retrieval using sodium citrate buffer. The slides were incubated for 10 min at 37 °C in blocking buffer (PBS containing 10% normal goat serum). Then, they were incubated overnight in a humidified chamber at 4 °C with the antibodies listed in Table S6. After washing twice with PBS, the slides were incubated at 37 °C for 30 min with a 1:500 dilution of secondary antibodies (Table S6), then incubated at 37 °C for 5 min with 500 ng/mL of DAPI and mounted using Vectashield antifade mounting medium (Vector, H-1000).
Bisulfite sequencing
Methylation patterns in the differentially methylated domains (DMDs) of the paternally imprinted gene H19 and maternally imprinted gene Snrpn were determined using bisulfite genomic sequencing as previously described (83). Genomic DNA was isolated and bisulfite treatment was conducted with an EZ DNA methylation-lightning kits (ZYMO Research, D5030) according to the manufacturer’s protocol. Bisulfate-converted DNA was subjected to PCR amplification of the H19 and Snrpn DMDs. The primer sequences and PCR conditions for amplification are previously described and listed in Table S5 (83). PCR products were sub-cloned into the pGEM-T Easy vector (Promega, A1360) and sequenced. Sequences were determined and analyzed using BiQ Analyzer software (84).
Reverse transcription and qRT-PCR
Total RNA extraction from cells was performed by E.Z.N.A.® Total RNA Kit I (Omega BIO-TEK) according to the manufacturer’s instructions or extracted with TRIzol® reagent (Thermo Fisher Scientific, 15596018). Reverse transcription was performed using a qScript™ cDNA SuperMix kit (Quantabio, 95048). qRT-PCR was performed with a C1000 Touch™ Thermal Cycler (Bio-Rad) amplification system using RT² SYBR® Green qPCR Mastermixes (Qiagen) and primer sets specific for each gene (Table S5). The relative levels of transcripts were calculated using the 2−ΔΔCT method and gene expression levels were normalized to Gapdh.
CRISPRi library design and gRNA cloning
Seven hundred and one epigenetic factors were selected from Epifactors, and 10 unique sgRNAs were designed for each gene using the GPP sgRNA Design tool. The CRISPRi library sgRNA oligos with Bsmb1 overhangs were synthesized by GenScript (Piscataway, NJ) and cloned into the lentiviral vector pXPR_050 by Golden Gate cloning following published protocols (85). Briefly, oligo pools were PCR amplified using primers listed in Table S5. pXPR_050 plasmids were digested using the Bsmb1 restriction enzyme (NEB, R0580). The synthesized double-strand oligo pools and digested pXPR_050 plasmids were ligated using T7 ligase (NEB, M0318S). The pXPR_050 plasmids were then transformed into Stbl4 competent cells (Invitrogen, 11635018). The transformed bacteria were then plated on LB plates with ampicillin resistance and grown for 24 hours at 30°C. To ensure high coverage of the CRISPRi library, bacterial colonies over 100× of the CRISPRi oligo library (~800,000) were collected and grown in SOC medium (Sigma, S1797). Plasmid DNA was isolated using a midi-prep kit (Invitrogen, K210005). The integrity of the CRISPRi library was validated by Illumina Miseq through Cornell Genomics Facility (detailed library preparation steps are presented below in “Library validation and CRISPRi dropout screening”).
For CRISPR-assisted insertion of PCR4-Shh::lacZ-H11 vector into the H11 locus, we used a gRNA that was previously designed for this purpose (36). Synthesized oligonucleotides were annealed and cloned into px459 vector. The sequences of all gRNAs are listed in Table S5.
Lentiviral packaging and transduction
Lentiviral particles were produced by transfecting HEK293T cells with the following plasmids and amounts: 2.5μg pMD2.G, 7.5μg psPAX2, and 10μg vectors (from pLX_311 lentiviral expression of dCas9-KRAB, the entire constructed pXPR_050 CRISPRi sgRNA expression library and pLV-tetO-Tfap2c) using TransIT-LT1 (Mirus, MIR2305). HEK293T culture media was collected at both 48 and 72 hours post transfection and concentrated using Amicon Ultra-15 columns (Millipore, UFC903024). ESCs were first transduced with lentiviral packaged pLX_311 and selected with Blasticidin (Gibco, A1113903) to generate stable dCas9-KRAB expressing cell lines, which were validated by PCR (Table S5) and Western blotting as described below. Approximately 1.6×107 ESCs with stable dCas9-KRAB expression were then transduced with the pXPR_050 CRISPRi sgRNA expression library at a multiplicity of infection of 0.3 to ensure at least 500× coverage of the library. The cells were selected in puromycin (Gibco, A1113803) (1.2mg/ml) for 2 days. All cells were infected with lentivirus for 48 hours.
Library validation and CRISPRi dropout screening.
Genomic DNAs from cells carrying CRISPRi sgRNA library were extracted using a Genomic DNA Purification Kit (Thermo Scientific, K0722). To recover sgRNA sequences, both CRISPRi plasmid pools and genomic DNAs were PCR amplified with Miseq_Adaptor primers (Table S5) using Q5 High-Fidelity DNA Polymerase (NEB, M0491L). PCR samples were purified from 2% agarose gels using a Gel Extraction Kit (Qiagen, 28704), then sequenced on an Illumina Miseq (Cornell Genomics Facility) using the seqRead_Miseq primer (Table S5). To validate the integrity of the CRISPRi library, the raw counts were mapped to the original CRISPRi library using ScreenProcessing Tools (86). To identify CRISPRi dropout candidates, sgRNA representations from ESCs, Stella-eGFP+, and Stella-eGFP− cells were analyzed using MAGeCK, MAGeCK-VISPR, and MAGeCKFlute Tools (50).
Knockdown and drug treatment during PGCLC differentiation.
Twenty-four hours before PGCLC differentiation, EpiLCs were transfected with control siNT (ON-TARGETplus Non-targeting Control siRNA, Dharmacon, D-001810–1005) and siNcor2 (ON-TARGETplus siRNA SMARTPool, Dharmacon, L-045364–00-0005) short dsRNA molecules using jetPRIME transfection reagent (Polyplus, 101000046) at a final concentration of 50nM. EpiLCs were then differentiated into PGCLCs over 6 days. Knockdown efficiencies were measured at day 2. Sodium butyrate (Sigma; B5887) and valproic acid (Sigma, P4543) were dissolved in water at stocks concentrations of 100mg/ml and 1M, respectively. To treat cells, dilutions were added to the PGCLC culture media at the start of PGCLC differentiation. Cells were collected 2, 4, or 6 days later. To prepare the cells for RNA-seq, the EBs were treated with 0.05mg/ml SB or 0.5mM VPA.
Western blotting
Cells were lysed and homogenized using RIPA buffer (Thermo Fisher Scientific, 89900) with protease inhibitors (Sigma, 04693159001) and phosphatase inhibitors (Sigma, 4906845001). The samples were centrifuged at 13,000rpm (~15,000G), and supernatants were used for western blot analysis. The protein concentrations were measured using a BCA Protein Assay Kit (Pierce, 23250). Samples were run through a gradient polyacrylamide gel (Bio-Rad, 4561083EDU) and transferred to PVDF membranes (Millipore, IPVH00010). Membranes were blocked by 5% nonfat milk for 1 hour. The antibodies used for western blotting were listed in Table S6. The HRR substrate (Millipore, WBLUC0500) and ChemiDoc Imaging System (Bio-Rad) were used for protein visualization.
Mouse treatment and reproductive phenotyping
For administration of drugs during pregnancy, E0.5 is the morning in which a vaginal plug was detected. Pregnant females received daily injections of water, SB (600mg/kg and 800mg/kg), or VPA (300mg/kg) from E5.5 to E11.5. Histology analysis, sperm and follicle counts were performed as described previously (80,87). To prepare the paraffin blocks, tissues were fixed overnight in Bouin’s solution or 4% PFA at room temperature and were embedded by Histology Lab at Cornell University. Paraffin sections at 6μm thick were deparaffinized and stained with hematoxylin and eosin. One cauda epididymis per male was used for sperm counting. To quantified follicle number, the ovary block was serially sectioned and stained by H&E. Every fifth section was scored for follicles at various stages.
RNA-seq and data analysis
Total RNAs were purified from cells using TRIzol® reagent (Invitrogen, 15596018). Purified RNAs were used for Illumina RNA-seq library preparation with NEBNext Ultra II Directional RNA Library Prep Kit (NEB, E7765), and a minimum of 20 million raw reads were obtained. The raw counts were then aligned to mouse mm10 genome with ENSEMBL gene annotations. Differential analysis of RNA-seq data was done using the R package DESeq2. RRHO2 analysis was performed also using an R package (41). K-Means clustering analysis was performed using iDEP 1.12 (88). GSEA was performed to explore enriched pathways and interpret RNA-seq data using predefined gene sets from the Molecular Signatures Database (v.7.1) in R packages (89). GSVA was carried out using the GSVA package in R (90). Gene sets enriched in the GSEA were organized into a graphical network produced using EnrichmentMap (91) plugins in Cytoscape (v.3.10.1) (92), as described previously (93). The online STRING database (version 12.0) was used for generation of protein interaction (94). The upregulated DEGs (log2FC>2, p<0.05 and count>250) from PGCLCs on days 2, 4 and 6 compared to ESC were integrated for the network analysis.
Accession numbers
The RNA-seq data generated in this study have been deposited at GEO (Gene Expression Omnibus) (GSE256104, reviewer token: mpgbukuqdnmrxgv; GSE259290, reviewer token: ivwzgkugrbijlyz). The RNA-seq data of ESCs/EpiLCs/Ck_D2/4/6PGCLCs (GSE67259) and in vivo germ cells (GSE74094 and GSE87644) were downloaded from the GEO database. The ESCs/EpiLCs/3TF_D2/4PGCLCs microarray dataset is accession # GSE43775. CRISPRi (GSE259289; reviewer token: gdadeiwgvzafxmb). The GEO number for RNA-seq data of formative ESCs is GSE135989. The GEO number for microarray dataset of ESCs/Nanog_D4PGCLC/Ck_D4PGCLC is GSE71933. The GEO number for RNA-seq datasets of 6h, 12h, 24h, and 48h EpiLCs is GSE256381.
Image process and data statistics
Fluorescent images were captured by an Olympus XM10 camera. Bright-field images were captured by the Olympus SC30 camera. Cropping, color, and contrast adjustments were made with Adobe Photoshop CC 2024, using identical background adjustments for all images. Fluorescence intensity was measured by ImageJ. All data were expressed as mean ± SEM. Statistical tests were carried out using a paired student’s t-test or one-way analysis of variance (ANOVA) followed by the Tukey’s post hoc test with R. Graph generation was performed using R. The schematic diagrams were created using BioRender.
Software and websites
Epifactors: https://epifactors.autosome.org/
GPP sgRNA Design tool: https://portals.broadinstitute.org/gpp/public/analysis-tools/sgrna-design
ScreenProcessing Tools: https://github.com/mhorlbeck/ScreenProcessing
MAGeCK, MAGeCK-VISPR, and MAGeCKFlute Tools: https://github.com/WubingZhang/MAGeCKFlute
DESeq2: https://bioconductor.org/packages/release/bioc/html/DESeq2.html
SARTools: https://github.com/PF2-pasteur-fr/SARTools
RRHO2: https://github.com/RRHO2/RRHO2
GSVA: https://alexslemonade.github.io/refinebio-examples/
GSEA: https://www.gsea-msigdb.org/gsea/index.jsp
iDEP 1.1: http://149.165.154.220/idep11/
g:Profiler: https://biit.cs.ut.ee/gprofiler/gost
ShinyGo 0.80: http://bioinformatics.sdstate.edu/go80/
ggplot2: https://ggplot2.tidyverse.org
STRING 12.0: https://string-db.org
FlowJo: https://www.flowjo.com
BioRender: www.biorender.com
Cytoscape: https://cytoscape.org
EnrichmentMap: https://apps.cytoscape.org/apps/enrichmentmap
Supplementary Material
Acknowledgements
The authors would like to thank R. Munroe and C. Abratte of Cornell’s transgenic facility for generating the mice, A. Surani for Stella-eGFP ESCs and several plasmids, and J.C. Bloom for OG2 iPSCs. This work is supported by a grant from the National Institutes of Health (R01 HD082568 to JCS), a Center grant (1P50HD096723) from the National Institute of Child Health and Human Development (K. Orwig: PI; JCS: Lead Project 2), and contract CO29155 from the NY State Stem Cell Program (NYSTEM). X.D. was supported by a postdoctoral fellowship from the Empire State Stem Cell Fund through New York State Department of Health contract no. C30293GG.
Footnotes
Competing interests
The authors declare no competing or financial interests.
References
- 1.Ding X, Schimenti JC. Strategies to identify genetic variants causing infertility. Trends Mol Med. 2021. Aug;27(8):792–806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Saitou M, Hayashi K. Mammalian in vitro gametogenesis. Science. 2021. Oct 1;374(6563):eaaz6830. [DOI] [PubMed] [Google Scholar]
- 3.Nichols J, Smith A. Naive and primed pluripotent states. Cell Stem Cell. 2009. Jun 5;4(6):487–92. [DOI] [PubMed] [Google Scholar]
- 4.Kinoshita M, Barber M, Mansfield W, Cui Y, Spindlow D, Stirparo GG, et al. Capture of Mouse and Human Stem Cells with Features of Formative Pluripotency. Cell Stem Cell. 2021. Mar 4;28(3):453–471.e8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Yu L, Wei Y, Sun H-X, Mahdi AK, Pinzon Arteaga CA, Sakurai M, et al. Derivation of intermediate pluripotent stem cells amenable to primordial germ cell specification. Cell Stem Cell. 2021. Mar 4;28(3):550–567.e12. [DOI] [PubMed] [Google Scholar]
- 6.Luo Q, Pui H-P, Chen J, Yu L, Jannig PR, Pei Y, et al. Epiblast-like stem cells established by Wnt/β-catenin signaling manifest distinct features of formative pluripotency and germline competence. Cell Rep. 2023. Jan 31;42(1):112021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang X, Xiang Y, Yu Y, Wang R, Zhang Y, Xu Q, et al. Formative pluripotent stem cells show features of epiblast cells poised for gastrulation. Cell Res. 2021. May;31(5):526–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hayashi K, Ohta H, Kurimoto K, Aramaki S, Saitou M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell. 2011. Aug 19;146(4):519–32. [DOI] [PubMed] [Google Scholar]
- 9.Nakaki F, Hayashi K, Ohta H, Kurimoto K, Yabuta Y, Saitou M. Induction of mouse germ-cell fate by transcription factors in vitro. Nature. 2013. Sep 12;501(7466):222–6. [DOI] [PubMed] [Google Scholar]
- 10.Zhang M, Leitch HG, Tang WWC, Festuccia N, Hall-Ponsele E, Nichols J, et al. Esrrb Complementation Rescues Development of Nanog-Null Germ Cells. Cell Rep. 2018. Jan 9;22(2):332–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Acampora D, Di Giovannantonio LG, Garofalo A, Nigro V, Omodei D, Lombardi A, et al. Functional Antagonism between OTX2 and NANOG Specifies a Spectrum of Heterogeneous Identities in Embryonic Stem Cells. Stem Cell Reports. 2017. Nov 14;9(5):1642–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Murakami K, Günesdogan U, Zylicz JJ, Tang WWC, Sengupta R, Kobayashi T, et al. NANOG alone induces germ cells in primed epiblast in vitro by activation of enhancers. Nature. 2016. Jan 21;529(7586):403–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vojtek M, Zhang J, Sun J, Zhang M, Chambers I. Differential repression of Otx2 underlies the capacity of NANOG and ESRRB to induce germline entry. BioRxiv. 2021. Jun 14; [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Acampora D, Mazan S, Lallemand Y, Avantaggiato V, Maury M, Simeone A, et al. Forebrain and midbrain regions are deleted in Otx2−/− mutants due to a defective anterior neuroectoderm specification during gastrulation. Development. 1995. Oct;121(10):3279–90. [DOI] [PubMed] [Google Scholar]
- 15.Shamblott MJ, Axelman J, Wang S, Bugg EM, Littlefield JW, Donovan PJ, et al. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc Natl Acad Sci USA. 1998. Nov 10;95(23):13726–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Matsui Y, Zsebo K, Hogan BL. Derivation of pluripotential embryonic stem cells from murine primordial germ cells in culture. Cell. 1992. Sep 4;70(5):841–7. [DOI] [PubMed] [Google Scholar]
- 17.Ohta H, Kurimoto K, Okamoto I, Nakamura T, Yabuta Y, Miyauchi H, et al. In vitro expansion of mouse primordial germ cell-like cells recapitulates an epigenetic blank slate. EMBO J. 2017. Jul 3;36(13):1888–907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ishikura Y, Ohta H, Sato T, Murase Y, Yabuta Y, Kojima Y, et al. In vitro reconstitution of the whole male germ-cell development from mouse pluripotent stem cells. Cell Stem Cell. 2021. Dec 2;28(12):2167–2179.e9. [DOI] [PubMed] [Google Scholar]
- 19.Murase Y, Yabuta Y, Ohta H, Yamashiro C, Nakamura T, Yamamoto T, et al. Long-term expansion with germline potential of human primordial germ cell-like cells in vitro. EMBO J. 2020. Nov 2;39(21):e104929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gajbhiye R, Fung JN, Montgomery GW. Complex genetics of female fertility. NPJ Genom Med. 2018. Oct 12;3:29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Krausz C, Riera-Escamilla A. Genetics of male infertility. Nat Rev Urol. 2018. Jun;15(6):369–84. [DOI] [PubMed] [Google Scholar]
- 22.Jørgensen N, Lamb DJ, Levine H, Pastuszak AW, Sigalos JT, Swan SH, et al. Are worldwide sperm counts declining? Fertil Steril. 2021. Dec;116(6):1457–63. [DOI] [PubMed] [Google Scholar]
- 23.Brieño-Enríquez MA, Larriba E, Del Mazo J. Endocrine disrupters, microRNAs, and primordial germ cells: a dangerous cocktail. Fertil Steril. 2016. Sep 15;106(4):871–9. [DOI] [PubMed] [Google Scholar]
- 24.Rattan S, Zhou C, Chiang C, Mahalingam S, Brehm E, Flaws JA. Exposure to endocrine disruptors during adulthood: consequences for female fertility. J Endocrinol. 2017. Jun;233(3):R109–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pivonello C, Muscogiuri G, Nardone A, Garifalos F, Provvisiero DP, Verde N, et al. Bisphenol A: an emerging threat to female fertility. Reprod Biol Endocrinol. 2020. Mar 14;18(1):22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Skinner MK. Endocrine disruptors and epigenetic transgenerational disease etiology. Pediatr Res. 2007. May;61(5 Pt 2):48R–50R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kurimoto K, Yabuta Y, Hayashi K, Ohta H, Kiyonari H, Mitani T, et al. Quantitative Dynamics of Chromatin Remodeling during Germ Cell Specification from Mouse Embryonic Stem Cells. Cell Stem Cell. 2015. May 7;16(5):517–32. [DOI] [PubMed] [Google Scholar]
- 28.Tang WWC, Dietmann S, Irie N, Leitch HG, Floros VI, Bradshaw CR, et al. A unique gene regulatory network resets the human germline epigenome for development. Cell. 2015. Jun 4;161(6):1453–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lombó M, Getino-Álvarez L, Depincé A, Labbé C, Herráez MP. Embryonic Exposure to Bisphenol A Impairs Primordial Germ Cell Migration without Jeopardizing Male Breeding Capacity. Biomolecules. 2019. Jul 25;9(8). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li Z, Fang F, Zhao Q, Li H, Xiong C. Supplementation of vitamin C promotes early germ cell specification from human embryonic stem cells. Stem Cell Res Ther. 2019. Nov 15;10(1):324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nagy L, Kao HY, Chakravarti D, Lin RJ, Hassig CA, Ayer DE, et al. Nuclear receptor repression mediated by a complex containing SMRT, mSin3A, and histone deacetylase. Cell. 1997. May 2;89(3):373–80. [DOI] [PubMed] [Google Scholar]
- 32.Huang EY, Zhang J, Miska EA, Guenther MG, Kouzarides T, Lazar MA. Nuclear receptor corepressors partner with class II histone deacetylases in a Sin3-independent repression pathway. Genes Dev. 2000. Jan 1;14(1):45–54. [PMC free article] [PubMed] [Google Scholar]
- 33.Park S-Y, Kim J-S. A short guide to histone deacetylases including recent progress on class II enzymes. Exp Mol Med. 2020. Feb 19;52(2):204–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Durcova-Hills G, Tang F, Doody G, Tooze R, Surani MA. Reprogramming primordial germ cells into pluripotent stem cells. PLoS ONE. 2008. Oct 27;3(10):e3531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Payer B, Chuva de Sousa Lopes SM, Barton SC, Lee C, Saitou M, Surani MA. Generation of stella-GFP transgenic mice: a novel tool to study germ cell development. Genesis. 2006. Feb;44(2):75–83. [DOI] [PubMed] [Google Scholar]
- 36.Kvon EZ, Zhu Y, Kelman G, Novak CS, Plajzer-Frick I, Kato M, et al. Comprehensive in vivo interrogation reveals phenotypic impact of human enhancer variants. Cell. 2020. Mar 19;180(6):1262–1271.e15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zhang J, Zhang M, Acampora D, Vojtek M, Yuan D, Simeone A, et al. OTX2 restricts entry to the mouse germline. Nature. 2018. Oct 3;562(7728):595–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Magnúsdóttir E, Dietmann S, Murakami K, Günesdogan U, Tang F, Bao S, et al. A tripartite transcription factor network regulates primordial germ cell specification in mice. Nat Cell Biol. 2013. Aug;15(8):905–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yamashiro C, Hirota T, Kurimoto K, Nakamura T, Yabuta Y, Nagaoka SI, et al. Persistent requirement and alteration of the key targets of PRDM1 during primordial germ cell development in mice. Biol Reprod. 2016. Jan;94(1):7. [DOI] [PubMed] [Google Scholar]
- 40.Sasaki K, Yokobayashi S, Nakamura T, Okamoto I, Yabuta Y, Kurimoto K, et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell. 2015. Aug 6;17(2):178–94. [DOI] [PubMed] [Google Scholar]
- 41.Cahill KM, Huo Z, Tseng GC, Logan RW, Seney ML. Improved identification of concordant and discordant gene expression signatures using an updated rank-rank hypergeometric overlap approach. Sci Rep. 2018. Jun 25;8(1):9588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Vojtek M, Zhang J, Sun J, Zhang M, Chambers I. Differential repression of Otx2 underlies the capacity of NANOG and ESRRB to induce germline entry. Stem Cell Reports. 2022. Jan 11;17(1):35–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kubota H, Avarbock MR, Brinster RL. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc Natl Acad Sci USA. 2004. Nov 23;101(47):16489–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Enders GC, May JJ. Developmentally regulated expression of a mouse germ cell nuclear antigen examined from embryonic day 11 to adult in male and female mice. Dev Biol. 1994. Jun;163(2):331–40. [DOI] [PubMed] [Google Scholar]
- 45.Carmell MA, Dokshin GA, Skaletsky H, Hu Y-C, van Wolfswinkel JC, Igarashi KJ, et al. A widely employed germ cell marker is an ancient disordered protein with reproductive functions in diverse eukaryotes. eLife. 2016. Oct 8;5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Buaas FW, Kirsh AL, Sharma M, McLean DJ, Morris JL, Griswold MD, et al. Plzf is required in adult male germ cells for stem cell self-renewal. Nat Genet. 2004. Jun;36(6):647–52. [DOI] [PubMed] [Google Scholar]
- 47.Sato T, Katagiri K, Gohbara A, Inoue K, Ogonuki N, Ogura A, et al. In vitro production of functional sperm in cultured neonatal mouse testes. Nature. 2011. Mar 24;471(7339):504–7. [DOI] [PubMed] [Google Scholar]
- 48.Hayashi K, Saitou M. Generation of eggs from mouse embryonic stem cells and induced pluripotent stem cells. Nat Protoc. 2013. Aug;8(8):1513–24. [DOI] [PubMed] [Google Scholar]
- 49.Jo K, Teague S, Chen B, Khan HA, Freeburne E, Li H, et al. Efficient differentiation of human primordial germ cells through geometric control reveals a key role for Nodal signaling. eLife. 2022. Apr 8;11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang B, Wang M, Zhang W, Xiao T, Chen C-H, Wu A, et al. Integrative analysis of pooled CRISPR genetic screens using MAGeCKFlute. Nat Protoc. 2019. Mar;14(3):756–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Mochizuki K, Hayashi Y, Sekinaka T, Otsuka K, Ito-Matsuoka Y, Kobayashi H, et al. Repression of somatic genes by selective recruitment of HDAC3 by BLIMP1 is essential for mouse primordial germ cell fate determination. Cell Rep. 2018. Sep 4;24(10):2682–2693.e6. [DOI] [PubMed] [Google Scholar]
- 52.Guenther MG, Lane WS, Fischle W, Verdin E, Lazar MA, Shiekhattar R. A core SMRT corepressor complex containing HDAC3 and TBL1, a WD40-repeat protein linked to deafness. Genes Dev. 2000. May 1;14(9):1048–57. [PMC free article] [PubMed] [Google Scholar]
- 53.Kim H-J, Bae S-C. Histone deacetylase inhibitors: molecular mechanisms of action and clinical trials as anti-cancer drugs. Am J Transl Res. 2011. Feb;3(2):166–79. [PMC free article] [PubMed] [Google Scholar]
- 54.Choi HW, Joo JY, Hong YJ, Kim JS, Song H, Lee JW, et al. Distinct enhancer activity of oct4 in naive and primed mouse pluripotency. Stem Cell Reports. 2016. Nov 8;7(5):911–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lyu X, Cui Y, Kong Y, Yang M, Shen H, Liao S, et al. A transient transcriptional activation governs unpolarized-to-polarized morphogenesis during embryo implantation. Mol Cell. 2024. Jul 25;84(14):2665–2681.e13. [DOI] [PubMed] [Google Scholar]
- 56.Aramaki S, Hayashi K, Kurimoto K, Ohta H, Yabuta Y, Iwanari H, et al. A mesodermal factor, T, specifies mouse germ cell fate by directly activating germline determinants. Dev Cell. 2013. Dec 9;27(5):516–29. [DOI] [PubMed] [Google Scholar]
- 57.Tischler J, Gruhn WH, Reid J, Allgeyer E, Buettner F, Marr C, et al. Metabolic regulation of pluripotency and germ cell fate through α-ketoglutarate. EMBO J. 2019. Jan 3;38(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hackett JA, Huang Y, Günesdogan U, Gretarsson KA, Kobayashi T, Surani MA. Tracing the transitions from pluripotency to germ cell fate with CRISPR screening. Nat Commun. 2018. Oct 16;9(1):4292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Naitou Y, Nagamatsu G, Hamazaki N, Shirane K, Hayashi M, Hayashi M, et al. Dual role of Ovol2 on the germ cell lineage segregation during gastrulation in mouse embryogenesis. Development. 2022. Feb 15;149(4). [DOI] [PubMed] [Google Scholar]
- 60.Yin Y, Cao S, Fu H, Fan X, Xiong J, Huang Q, et al. A noncanonical role of NOD-like receptor NLRP14 in PGCLC differentiation and spermatogenesis. Proc Natl Acad Sci USA. 2020. Sep 8;117(36):22237–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Soper SFC, van der Heijden GW, Hardiman TC, Goodheart M, Martin SL, de Boer P, et al. Mouse maelstrom, a component of nuage, is essential for spermatogenesis and transposon repression in meiosis. Dev Cell. 2008. Aug;15(2):285–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kim M-S, Pinto SM, Getnet D, Nirujogi RS, Manda SS, Chaerkady R, et al. A draft map of the human proteome. Nature. 2014. May 29;509(7502):575–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Malki S, van der Heijden GW, O’Donnell KA, Martin SL, Bortvin A. A role for retrotransposon LINE-1 in fetal oocyte attrition in mice. Dev Cell. 2014. Jun 9;29(5):521–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Schemmer J, Araúzo-Bravo MJ, Haas N, Schäfer S, Weber SN, Becker A, et al. Transcription factor TFAP2C regulates major programs required for murine fetal germ cell maintenance and haploinsufficiency predisposes to teratomas in male mice. PLoS ONE. 2013. Aug 13;8(8):e71113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Irie N, Lee S-M, Lorenzi V, Xu H, Chen J, Inoue M, et al. DMRT1 regulates human germline commitment. Nat Cell Biol. 2023. Oct;25(10):1439–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Eguizabal C, Montserrat N, Vassena R, Barragan M, Garreta E, Garcia-Quevedo L, et al. Complete meiosis from human induced pluripotent stem cells. Stem Cells. 2011. Aug;29(8):1186–95. [DOI] [PubMed] [Google Scholar]
- 67.Ishikura Y, Yabuta Y, Ohta H, Hayashi K, Nakamura T, Okamoto I, et al. In Vitro Derivation and Propagation of Spermatogonial Stem Cell Activity from Mouse Pluripotent Stem Cells. Cell Rep. 2016. Dec 6;17(10):2789–804. [DOI] [PubMed] [Google Scholar]
- 68.Bleckwehl T, Crispatzu G, Schaaf K, Respuela P, Bartusel M, Benson L, et al. Enhancer-associated H3K4 methylation safeguards in vitro germline competence. Nat Commun. 2021. Oct 1;12(1):5771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li L, Li L, Li Q, Liu X, Ma X, Yong J, et al. Dissecting the epigenomic dynamics of human fetal germ cell development at single-cell resolution. Cell Res. 2021. Apr;31(4):463–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Alexander AK, Rice EJ, Lujic J, Simon LE, Tanis S, Barshad G, et al. A-MYB and BRDT-dependent RNA Polymerase II pause release orchestrates transcriptional regulation in mammalian meiosis. Nat Commun. 2023. Mar 29;14(1):1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Liu H, Wang J, He T, Becker S, Zhang G, Li D, et al. Butyrate: A Double-Edged Sword for Health? Adv Nutr. 2018. Jan 1;9(1):21–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Jentink J, Loane MA, Dolk H, Barisic I, Garne E, Morris JK, et al. Valproic acid monotherapy in pregnancy and major congenital malformations. N Engl J Med. 2010. Jun 10;362(23):2185–93. [DOI] [PubMed] [Google Scholar]
- 73.Kurilo LF. Oogenesis in antenatal development in man. Hum Genet. 1981;57(1):86–92. [DOI] [PubMed] [Google Scholar]
- 74.Baker TG. A quantitative and cytological study of germ cells in human ovaries. Proc R Soc Lond, B, Biol Sci. 1963. Oct 22;158:417–33. [DOI] [PubMed] [Google Scholar]
- 75.Babakhanzadeh E, Nazari M, Ghasemifar S, Khodadadian A. Some of the factors involved in male infertility: A prospective review. Int J Gen Med. 2020. Feb 5;13:29–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mann U, Shiff B, Patel P. Reasons for worldwide decline in male fertility. Curr Opin Urol. 2020. May;30(3):296–301. [DOI] [PubMed] [Google Scholar]
- 77.Smith A. Formative pluripotency: the executive phase in a developmental continuum. Development. 2017. Feb 1;144(3):365–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Chavez A, Scheiman J, Vora S, Pruitt BW, Tuttle M, P R Iyer E, et al. Highly efficient Cas9-mediated transcriptional programming. Nat Methods. 2015. Apr;12(4):326–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Dixit A, Parnas O, Li B, Chen J, Fulco CP, Jerby-Arnon L, et al. Perturb-Seq: Dissecting Molecular Circuits with Scalable Single-Cell RNA Profiling of Pooled Genetic Screens. Cell. 2016. Dec 15;167(7):1853–1866.e17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ding X, Singh P, Schimenti K, Tran TN, Fragoza R, Hardy J, et al. In vivo versus in silico assessment of potentially pathogenic missense variants in human reproductive genes. Proc Natl Acad Sci USA. 2023. Jul 25;120(30):e2219925120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Ding X, Schimenti JC. Female infertility from oocyte maturation arrest: assembling the genetic puzzle. EMBO Mol Med. 2023. Jun 7;15(6):e17729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Luo Y, Hartford SA, Zeng R, Southard TL, Shima N, Schimenti JC. Hypersensitivity of primordial germ cells to compromised replication-associated DNA repair involves ATM-p53-p21 signaling. PLoS Genet. 2014. Jul 10;10(7):e1004471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhou Q, Wang M, Yuan Y, Wang X, Fu R, Wan H, et al. Complete Meiosis from Embryonic Stem Cell-Derived Germ Cells In Vitro. Cell Stem Cell. 2016. Mar 3;18(3):330–40. [DOI] [PubMed] [Google Scholar]
- 84.Bock C, Reither S, Mikeska T, Paulsen M, Walter J, Lengauer T. BiQ Analyzer: visualization and quality control for DNA methylation data from bisulfite sequencing. Bioinformatics. 2005. Nov 1;21(21):4067–8. [DOI] [PubMed] [Google Scholar]
- 85.Read A, Gao S, Batchelor E, Luo J. Flexible CRISPR library construction using parallel oligonucleotide retrieval. Nucleic Acids Res. 2017. Jun 20;45(11):e101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Liu SJ, Horlbeck MA, Weissman JS, Lim DA. Genome-Scale Perturbation of Long Noncoding RNA Expression Using CRISPR Interference. Methods Mol Biol. 2021;2254:323–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ding X, Fragoza R, Singh P, Zhang S, Yu H, Schimenti JC. Variants in RABL2A causing male infertility and ciliopathy. Hum Mol Genet. 2020. Dec 18;29(20):3402–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ge SX, Son EW, Yao R. iDEP: an integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinformatics. 2018. Dec 19;19(1):534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci USA. 2005. Oct 25;102(43):15545–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Hänzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinformatics. 2013. Jan 16;14:7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Merico D, Isserlin R, Stueker O, Emili A, Bader GD. Enrichment map: a network-based method for gene-set enrichment visualization and interpretation. PLoS ONE. 2010. Nov 15;5(11):e13984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003. Nov;13(11):2498–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Reimand J, Isserlin R, Voisin V, Kucera M, Tannus-Lopes C, Rostamianfar A, et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat Protoc. 2019. Feb;14(2):482–517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Szklarczyk D, Kirsch R, Koutrouli M, Nastou K, Mehryary F, Hachilif R, et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023. Jan 6;51(D1):D638–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.