

Abstract
The GhNAC2 transcription factor identified from G. herbaceum improves root growth and drought tolerance through transcriptional reprogramming of phytohormone signaling. The promoter of such a versatile gene could serve as an important genetic engineering tool for biotechnological application. In this study, we identified and characterized the promoter of GhNAC2 to understand its regulatory mechanism. GhNAC2 transcription factor increased in root tissues in response to GA, ethylene, auxin, ABA, mannitol, and NaCl. In silico analysis revealed an overrepresentation of cis-regulatory elements associated with hormone signaling, stress responses and root-, pollen-, and seed-specific promoter activity. To validate their role in GhNAC2 function/regulation, an 870-bp upstream regulatory sequence was fused with the GUS reporter gene (uidA) and expressed in Arabidopsis and cotton hairy roots for in planta characterization. Histochemical GUS staining indicated localized expression in root tips, root elongation zone, root primordia, and reproductive tissues under optimal growth conditions. Mannitol, NaCl, auxin, GA, and ABA, induced the promoter-driven GUS expression in all tissues while ethylene suppressed the promoter activity. The results show that the 870 nt fragment of the GhNAC2 promoter drives root-preferential expression and responds to phytohormonal and stress signals. In corroboration with promoter regulation, GA and ethylene pathways differentially regulated root growth in GhNAC2-expressing Arabidopsis. The findings suggest that differential promoter activity governs the expression of GhNAC2 in root growth and stress-related functions independently through specific promoter elements. This multifarious promoter can be utilized to develop yield and climate resilience in cotton by expanding the options to control gene regulation.
Supplementary Information
The online version contains supplementary material available at 10.1007/s12298-024-01411-2.
Keywords: Cotton, Gene regulation, GhNAC2, Hormone signaling, Inducible promoter, Root
Introduction
Cotton (Gossypium hirsutum) is one of the essential fiber crops used by the textile industry worldwide. Cotton cultivation is exposed to several environmental stresses, such as drought, heat, and flooding (Dabbert and Gore 2014; Li et al. 2019a; Zahid et al. 2021). The root system is a vital interface through which plants respond to various environmental factors. They are the lifeline of the plant to water and nutrient supply (Meshram et al. 2021). A well-developed root system is critical for anchoring cotton growth, stress adaptation, and producing quality fiber (Niu et al. 2018). Phytohormones are the key regulators of root growth, development, and environmental interactions. Multiple hormones, including auxin, ethylene, and gibberellins (GA), govern root cell proliferation and elongation in the root apical meristem and elongation zone, respectively, through complex crosstalk (Li et al. 2015; Pacifici et al. 2015; Qin and Huang 2018). The primary hormonal signaling is perceived through specific transcription factor (TF) families, such as AP2/ERF (APETALA2/ethylene responsive factor), ARF (Auxin-responsive Factor), AREB/ABF (ABRE-binding protein/ABRE-binding factor (AREB/ABF), GRAS-domain proteins, etc., which in turn, signal the central transcription factors (Fujita et al. 2013; Li et al. 2016; Xie et al. 2019).
NAC (No Apical Meristem, ATAF1/2, and CUC2) TFs are the central regulators of growth and stress signaling in plants (Nuruzzaman et al. 2013; Srivastava et al. 2022b). NAC proteins control transcriptional reprogramming associated with root development (Yarra and Wei 2021; Takasaki et al. 2010; Xu et al. 2022), seed germination (Kim et al. 2008; Yu et al. 2021; Park et al. 2011), floral development (Yang et al. 2018; Sun et al. 2020), seed development (Sánchez-Montesino et al. 2019), stress responses (Guo et al. 2017; Srivastava et al. 2022a), etc. Previous studies showed that NAC TFs regulate the downstream metabolism and signaling of phytohormones. For instance, ATAF1 directs ABA biosynthesis to activate senescence, and JUB1 regulates GA catabolism to suppress flowering in Arabidopsis, respectively (Jensen et al. 2013; Shahnejat-Bushehri et al. 2016). In addition, the phytohormonal cascades act as upstream signals tuning the expression of NAC genes in response to stress and developmental cues (He et al. 2005). For instance, auxin and an interplay between ABA and ethylene direct NAC-mediated xylem development and fruit ripening in poplar and tomato, respectively (Johnsson et al. 2019; Kou et al. 2021). Thus, NAC proteins form the core of hormonal pathways.
The promoter of NAC genes carries crucial regulatory elements and TF binding sites bound by the hormone- and stress-responsive TFs to execute specific spatiotemporal gene expression patterns (Nakashima et al. 2012). GUS-reporter-based study of promoter activity of GhNAC79, a drought-responsive NAC gene in cotton, indicated stress-induced and predominant expression in cotyledons of Arabidopsis (Guo et al. 2017). Similarly, the GhNAC4 promoter is induced by multiple hormones and shows high expression in the vascular bundle and guard cells in tobacco (Trishla et al. 2020). Indeed, promoter analysis provides significant insight into the mechanism of hormone signaling and plant-environment interaction. Regardless, limited information is available about NAC promoters, especially in cotton.
Gossypium hirsutum, the widely cultivated cotton species for natural fiber production, is susceptible to drought stress (Gao et al. 2020). Several cultivars of G. herbaceum, species with poor fiber quality, are inherently drought tolerant and could serve as a useful genetic resource to explore target genes to develop stress adaptation (Kulkarni et al. 2009). Our previous study showed that GhNAC2, a NAC transcription factor isolated from G. herbaceum, promoted root growth not only during optimum growth conditions but also during drought stress in cotton. Differential gene expression study in Arabidopsis pointed towards its role in transcriptional reprogramming of phytohormonal metabolism and stress-responsive genes (Gunapati et al. 2016). To understand the role of promoter in determining the functional versatility of GhNAC2, we cloned the upstream region of GhNAC2 and characterized the tissue-specific, hormone- and stress-responsive activities in cotton hairy roots and Arabidopsis, using a GUS reporter-based approach. Further, the phytohormonal regulation of root growth was examined in GhNAC2-expressing Arabidopsis lines.
Materials and methods
Plant materials and growth conditions
For gene expression analysis, mature cotton seeds (Gossypium herbaceum cv. Vagad) were delinted using concentrated H2SO4 for one minute, neutralized with a saturated aqueous solution of NaHCO3, followed by washing with deionized water (4–5 times) and dried at 28 °C for 2–3 days. Healthy seeds were surface-sterilized using 0.1% (w/v) HgCl2 for 5 min and washed with sterile deionized water (4–5 times). The seeds were soaked in absolute ethanol for one minute, and flame scorched. Sterilized seeds were germinated on Murashige–Skoog Agar medium [MSA: 0.5X MS + 0.8% (w/v) agar + 2% (w/v) glucose, pH 5.8] and incubated at 28 °C for 48 h under dark. The emerging radicals were transferred to the culture tubes containing 1X Hoagland solution (Hoagland and Arnon 1950) and grown for three weeks at 28 °C under 16 h:8 h (light:dark) photoperiodic conditions. For promoter isolation, genomic DNA was isolated from the leaf tissue of three-week-old cotton seedlings using the CTAB method (Ali et al. 2019). To generate cotton hairy roots expressing the GUS-fused GhNAC2 promoter construct and control vectors, two-week-old seedlings (Gossypium hirsutum cv. Coker 310) germinated in MSA media containing tube were used.
To study phytohormonal regulation of root growth in Arabidopsis, healthy seeds of wild type (WT) and four independent homozygous T3 lines (L1, L4, L8, and L15) expressing the GhNAC2 gene (Gunapati et al. 2016) were surface-sterilized with 70% (v/v) alcohol (2 min) and 2% (v/v) sodium hypochlorite (NaOCl) + 0.02% (v/v) Triton X-100 (5 min), followed by washing with sterile deionized water (4–5 times). The seeds were placed on MSA medium [0.5X MS + 0.8% (w/v) agar + 2% (w/v) sucrose, pH 5.8] and kept for stratification at 4 °C for two days under dark conditions. The plates were then transferred to the growth chamber maintained at 22 °C and 16 h:8 h (light:dark) photoperiodic conditions. Root lengths were measured manually after the indicated time interval. To generate Arabidopsis lines expressing the GUS-fused GhNAC2 promoter and control vectors, WT (Columbia-0) plants were grown in soilrite pots supplied with nutrient media for one month at 22 °C (Misra et al. 2017). For GUS histochemical staining at the seedling stage, surface-sterilized seeds of control and three independent homozygous T3 Arabidopsis lines (L1, L2, and L3) expressing the GhNAC2 promoter were grown on MSA plates for the indicated time as mentioned earlier in the study. To study GUS expression in mature vegetative tissues (one-month-old), reproductive organs (flower, stigma, anther, and pollen), siliques, and seeds, the control and promoter-expressing Arabidopsis lines were grown in soilrite as mentioned earlier in the study.
Phytohormone and abiotic stress treatment
For gene expression analysis in cotton, WT seedlings were transferred to Hoagland media supplemented with 1.0 µM GA3 (Gibberellin A3), 1.0 µM ACC (1-aminocyclopropane-1-carboxylic acid, an ethylene precursor), 100 µM ABA (Abscisic acid), 1.0 µM IAA (Indole-3-acetic acid), and deionized water (mock). After two hours, the root tissues were harvested, flash-frozen, and stored at − 80 °C. For root growth analysis in Arabidopsis, the seeds were germinated on MSA plates supplemented with phytohormones: 0.5 µM GA3, 1.0 µM ACC, 1.0 µM ABA, 0.1 µM IAA, and their respective inhibitors: 1.0 µM PAC (Paclobutrazol), 0.1 µM AgNO3 (Silver nitrate), 1.0 µM Fluridon, and 1.0 µM TIBA (Triiodobenzoic acid) and grown for the indicated time. For the GUS expression study, control and promoter-expressing cotton hairy roots were transferred to MSA plates containing 1.0 µM GA3, 1.0 µM ACC, 1.0 µM ABA, 0.1 nM IAA, 1.0 µM PAC, 0.1 µM AgNO3, 1.0 µM Fluridon, 1.0 µM TIBA, 200 mM Mannitol, and 150 mM NaCl for 24 h. The statistical analysis was performed using Duncan’s test at 5% probability level by subjecting the data to one-way ANOVA using SPSS software 20.0.
Real-time qPCR analysis
Total RNA was isolated from the frozen control and treated root tissues using the CTAB-LiCl RNA extraction method (Asif et al. 2000). First strand cDNA synthesis was carried out using GeneSure H-Minus cDNA Synthesis Kit (Puregene, Genetix Biotech). To estimate the transcript abundance of GhNAC2 (EU706339), real-time qRT-PCR was run on StepOnePlus Real-Time PCR System (Applied Biosystems, USA) using the GeneSure SYBR Green qPCR 2X Master Mix (Puregene, Genetix Biotech). GhUBI1 (EU604080) and GhActin7 (XM_016840071) were used as internal controls (references) to normalize the expression values. The expression analysis was performed with three biological and technical replicates, and the relative gene expression was calculated by the 2−ΔΔCT method (Livak and Schmittgen 2001). Student’s t- test with a threshold p-value < 0.05 was used to compare the treated samples with the mock controls. The primers used in the study are listed in Supplementary Table S1.
Isolation and cloning of GhNAC2 promoter by genome walking
2.5 μg of genomic DNA was digested with four different blunt-end restriction enzymes viz. DraI, EcoRV, RsaI, and HincII (ThermoFisher Scientific, USA) separately and purified by phenol:chloroform extraction. To construct the genome walking libraries, the purified digested products were ligated with the BD Genome Walker adapters using the standard protocol of BD Genome Walker Universal Kit (BD Biosciences, Clontech, USA). For isolation of the GhNAC2 promoter sequence, the mixture of genomic libraries was used as the template for the nested touch-down PCR. The standard reaction and thermocycling parameters mentioned in the user manual were followed to obtain the amplicon. The 870 bp DNA fragment was purified and cloned in the pTZ57R/T vector, followed by Sanger sequencing (3730xl DNA Analyzer, Applied Biosystems). The promoter sequence was submitted to National Center for Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/) with the accession number: OQ190160. For in silico analysis, the sequence was analyzed using the databases of plant cis-acting regulatory elements, such as PlantCare (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) (Rombauts et al. 1999) and PLACE (https://www.dna.affrc.go.jp/PLACE/?action=newplace) (Higo et al. 1999). The transcription factor binding sites (TFBS) were screened using PlantRegMap (http://plantregmap.gao-lab.org/binding_site_prediction.php) (Tian et al. 2020). The 870 bp promoter fragment was cloned in the BamHI site in the binary expression vector pBI101 to generate its in-frame fusion with GUS (ProGhNAC2::GUS). The primers used in the study are listed in Supplementary Table S1.
Generation of transgenic Arabidopsis expressing GhNAC2 promoter
The ProGhNAC2::GUS construct was introduced into Agrobacterium tumefaciens (strain GV3101) and Arabidopsis transformed using the floral dip method (Clough and Bent 1998). Transformed seeds were collected, surface-sterilized, and grown on the MSA medium supplemented with kanamycin (50 mg/L). The kanamycin-resistant T1 seedlings were allowed to self-fertilize. T-DNA insertion was confirmed by antibiotic selection (50 mg/L Kanamycin) and genomic PCR. The homozygous T3 transgenic lines were used for detailed promoter analysis.
Generation of cotton hairy roots expressing GhNAC2 promoter
The cotyledonary and newly emerged leaves were excised into one cm2 sections and used as explants for the hairy root transformation (Verma et al. 2007; Gu et al. 2014). The explants were randomly pricked at 3–4 positions with the help of a sterile needle and immersed into Agrobacterium rhizogenes (strain A4) culture carrying the ProGhNAC2::GUS construct and kept for gentle shaking for 30 min at 25 °C. The infected explants were soaked on sterile filter paper and transferred to MSP media (1X MS + 0.22% (w/v) phytagel + 3% (w/v) glucose, pH 5.8) and co-cultivated at 25 °C under dark conditions for 48 h. The explants were washed with Cefotaxime + Augmentin (125 mg/L each) for 5 min, followed by washing with sterile deionized water (4–5 times) to remove the excess Agrobacterium. For hairy root generation, the explants were transferred to MSP media plates supplemented with Kanamycin (50 mg/L), Cefotaxime (125 mg/L), and Augmentin (125 mg/L) and incubated at 25 °C under 16 h:8 h (light:dark) photoperiodic conditions. The hairy roots emerged at the wounded sites after three weeks of selection, which were sub-cultured further to increase the root biomass. T-DNA insertion was confirmed by genomic PCR, and the transgenic hairy roots were used for detailed promoter analysis.
Histochemical GUS staining and fluorometric assay
The transgenic Arabidopsis lines expressing the ProGhNAC2::GUS construct were analyzed by histochemical GUS staining. Plants expressing pBI121 (CaMV35S::GUS) and pBI101 (empty vector) served as the positive and negative control, respectively. The histochemical GUS staining was performed as described previously (Jefferson et al. 1987). The tissues were immersed in the staining buffer: 50 mM phosphate buffer (pH 7.2), 2.0 mM potassium ferricyanide [K3Fe(CN)6], 2.0 mM potassium ferrocyanide [K4Fe(CN)6], and 0.1% (v/v) Triton X-100, supplied with 1.0 mg/mL of X-gluc substrate and kept at 37 °C for two hours. The stained samples were washed with 70% (v/v) ethanol to remove the chlorophyll pigment and examined under the light microscope (Leica DM1000LED, Germany) and stereo-microscope (Leica S8APO, Germany). To examine promoter activity in the cotton hairy roots, the phytohormone- and stress-treated tissues were subjected to GUS staining at 37 °C for eight hours (Jefferson et al. 1987). Hairy roots expressing pBI121 and pBI101 served as the positive and negative control, respectively. The images were captured by stereo-microscope (Leica S8APO, Germany). In addition, the stained roots samples were fixed in Optimal Cutting Temperature (OCT) Embedding Medium (Scigen, USA) at − 80 °C for 30 min followed by transverse sectioning using Microtome (Leica DM1950) at 4 °C. The sections were examined under Light Microscopy (Leica DM2500, Germany).
GUS activity was quantified by a fluorometric assay using 4-trifluoromethylumbelliferyl-β-D-glucuronic acid (4-MUG salt) as a substrate. 1.0 mM MUG was dissolved in GUS extraction buffer [50 mM sodium phosphate buffer (pH 7.0), 10 mM EDTA (pH 8.0), 10 mM β-mercaptoethanol, 0.1% (w/v) sarcosine, 0.1% (v/v) Triton X-100] to prepare GUS assay buffer and pre-heated at 37 °C for 15 min. 50 µl of crude protein (0.5 mg/ml), extracted from hairy root tips using Bradford’s assay (Bradford 1976), was mixed with 900 µl of GUS assay buffer and incubated at 37 °C to produce 4-Methylumbelliferone (4-MU). 100 µl aliquots of the mixture were removed successively at 15 min intervals and added to 900 µl of 0.2 M Na2CO3 to terminate the reaction. The fluorescence intensity (FI) was measured by Spectrophotometer Tecan Infinite M200 Pro (excitation 365 nm, emission 455 nm). The GUS activity of the extract was calculated in terms of pmol of 4-MU per minute per mg protein according to the equation below.
Results
GhNAC2 gene expression is induced by phytohormones and abiotic stress
NAC transcription factors (TFs) are the central receptors of hormonal and stress signals (Jensen et al. 2010). GhNAC2 is a transcriptional repressor belonging to the NAC family of cotton that governs root growth and drought response (Gunapati et al. 2016). To examine the phytohormonal regulation of GhNAC2 expression, WT cotton seedlings were treated with growth- and stress-signaling hormones for two hours. The qRT-PCR indicated an increase in the abundance of GhNAC2 transcript in response to GA3 (~ threefold), ACC (~ 4.7-fold), ABA (~ 27-fold), IAA (~ tenfold), mannitol (~ 35-fold), and NaCl (~ threefold) in the root tissues (Fig. 1a).
Fig. 1.

Expression and in-silico promoter analysis of GhNAC2 transcription factor. a GhNAC2 was significantly induced by the exogenous treatment of GA3, ACC, ABA, IAA, mannitol, and NaCl. All data are means ± SE (n = 9). Student’s t- test with a threshold p value < 0.05 was used to compare the treated samples with the mock controls (a = p value < 0.001, b = p value < 0.01, and c = p value < 0.05). b Schematic diagram representing the core and distal promoter elements, 5’-UTR, exon/intron arrangement, and 3’-UTR. c Histogram of the over-represented cis-acting elements in the distal promoter region involved in the stress, hormonal, light, and tissue-specific gene regulation. d Histogram representing the TF binding sites occurring in the promoter. (UTR untranslated region, TSS transcription start site)
GhNAC2 promoter carries hormone and TF regulatory signals
A DNA fragment of ~ 870 bp was amplified from the upstream of the GhNAC2 gene by genome walking PCR (Supplementary Fig. S1) and sequenced. The retrieved sequence was analyzed using in silico databases to identify the 5’-untranslated region (UTR), transcription start site (TSS), and the cis-regulatory elements (CRE) present in the core, proximal and distal region, governing the initiation, intensity, and specificity of the transcriptional activity of GhNAC2 gene (Biłas et al. 2016).The GhNAC2 gene carried a 5′-UTR of 120 bp upstream of the TSS [+ 1], marking the site of transcriptional initiation. The core promoter region carried a canonical TATA box at − 24 bp required for the binding of RNA polymerase. The proximal promoter region was detected with CAAT [− 140 bp] box and GC box [− 78 bp], which is required for the primary regulation (Supplementary Table S2). The distal promoter region [− 150 bp to − 750 bp] was rich in the CREs required for hormone-and stress-specific regulation (Supplementary Table S3). Three motifs, PYRIMIDINEBOXOSRAMY1A (1 site), GADOWNAT (1 site), and WRKY71OS (9 sites), involved in the GA signaling were found. ABRERATCAL, an ABA-responsive element (ABRE), was detected at three sites. Three CREs involved in auxin response: ARFAT (1 site), ASF1MOTIFCAMV (2 sites), and SGBFGMGMAUX28 (1 site), were predicted (Higo et al. 1999).
Dehydration-induced cis-acting elements such as MYCCONSENSUSAT (5 sites), ACGTATERD1 (3 sites), and CBFHV (3 sites), as well as salt-responsive element GT1GMSCAM4 (3 sites), were also overrepresented (Fig. 1b and Supplementary Table S3). Several promoter motifs controlling tissue-specific expression were found in the GhNAC2 promoter. For instance, ROOTMOTIFTAPOX1 (4 sites) and ACGTROOT1 (2 sites) are involved in the root-specific expression, POLLEN1LELAT52 (6 sites) and GTGANTG10 (12 sites) regulate pollen-specific expression (Filichkin et al. 2004), and DOFCOREZM (8 sites), -300ELEMENT (3 sites), and RYREPEATBNNAPA (1 site) are known for seed-specific promoter activity (Ezcurra et al. 1999). In addition, the binding sites for several TFs, such as bHLH, bZIP, ERF, WRKY, including NAC, known to govern stress and developmental responses were also predicted (Das et al. 2019; Tian et al. 2020) (Fig. 1c). The study indicated a complex regulation of GhNAC2 promoter by hormone, stress, tissue-specific, and transcription factors signals.
GhNAC2 promoter was predominantly expressed in the roots, reproductive tissues, and immature seeds
To gain insight into the tissue-specific GhNAC2 promoter activity, GUS expression was examined in different tissues of homozygous progenies of three independent transgenic Arabidopsis lines (L1, L2, and L3) expressing the GhNAC2 gene under GhNAC2 promoter (Supplementary Fig. S2). Strong promoter-driven GUS expression was detected in the emerging radical tip of 36 h-old germinating seedlings, unlike the control (Fig. 2a and b). After 48 h and 72 h of germination, GUS expression was observed in the root tips (Fig. 2c and d) and hypocotyl elongation zone (Fig. 2e). During the post-germinative growth (15-day-old), preferential GUS expression was observed in the root tissue compared to the leaves (Fig. 2f and g). When examined closely, strong GUS expression was localized to the elongation zone and tip of the primary root (Fig. 2h), and lateral root primordia and lateral root tip (Fig. 2i and j). However, no GUS activity was observed in the differentiation zone of the roots (Fig. 2d and i). At the rosette stage (one month old), the GUS expression was limited to the leaf veins, leaf trichomes, root hairs, and lateral roots (Fig. 21–o). Later during the reproductive stage, GhNAC2 promoter-driven GUS expression was localized to the anther, stigma, pollen grains, and apical bud (Fig. 2q–t). Besides, strong GUS expression was detected in the immature silique valves and seeds (Fig. 2v and w) and mature silique valves (Fig. 2x). Interestingly, no GUS expression was detected in the sepal and petals of the flower and mature seeds (Fig. 2q and y).
Fig. 2.

Tissue-specific GUS expression driven by GhNAC2 promoter in transgenic Arabidopsis. GUS histo-chemical staining was performed in different tissues of transgenic Arabidopsis lines expressing the ProGhNAC2::GUS construct. Tissues expressing the empty pBI101 vector served as control (a: 72 h-old root, f: 15-day-old seedling, k: one-month-old leaf, p: flower, and u: silique). Prominent GUS expression was observed in the root tips and elongation zones of b 36 h-old, c 48 h-old, and d, e 72 h-old seedlings. In the 15-day-old seedlings, g preferential GUS expression was localized in the root tissues: h, i elongation zone and primary root tip, j lateral root tip and lateral root primordia. In the one-month-old soil-grown plants, GUS expression was visible in l leaf veins, m trichomes, n root hairs, along with o the lateral roots. In the two-month-old plants, prominent GUS expression was observed in the q, r stigma and anthers, s pollen grains, t apical buds, v, w immature silique valves and seeds, and x, y mature silique valves. The images represent the results obtained for three independent T3 generation lines. (RT root tip, EZ elongation zone, DZ differentiation zone, LRP lateral root primordia, LRT lateral root tip, RH root hair, AB apical bud, IV immature silique valve, IS immature seed, MV mature silique valve, and MS mature seed)
Further, to understand GhNAC2 promoter regulation in the homologous cotton system, transgenic hairy roots expressing the ProGhNAC2::GUS construct were developed. Unlike control (Fig. 3a and b), faint GUS expression was detected in the elongation zone (Fig. 3c) along with an intense GUS expression in the root tips (Fig. 3d) and root primordia (Fig. 3e) of the transgenic hairy roots. However, no GUS activity was observed in the differentiation zone (Fig. 3f). Further, the TS sections of the respective GUS-expressing zones were sliced for detailed visualization. In the elongation zone, the GUS expression was limited to the epidermal layer (Fig. 3c) whereas, in the root tips, strong GUS expression was found in the root stele (pericycle, vascular tissues, and pith) and cortex region, besides the epidermal layer (Fig. 3d).
Fig. 3.

Tissue-specific GUS expression driven by GhNAC2 promoter in cotton hairy roots. GUS localization was visualized in the intact (top) and transverse sections (bottom). a Empty pBI101 and b pBI121 vectors served as the negative and positive control, respectively. GhNAC2 promoter-driven GUS expression was observed in the c elongation zone, d root tip, and e root primordia, except f differentiation zone of cotton hairy roots. The images represent the results obtained for 10–12 independent hairy roots expressing each construct. (EZ elongation zone, DZ differentiation zone, RT root tip, RP root primordia, EC epidermal cell, CC cortical cell, PC pericycle cell)
GhNAC2 promoter is activated by hormones and abiotic stress
GhNAC2 transcript was induced in response to hormone treatment and abiotic stress (Fig. 1a). To validate the hormone regulation at the promoter level, cotton hairy roots expressing the ProGhNAC2::GUS construct (Supplementary Fig. S2) were treated with GA, ethylene, ABA, auxin and their inhibitors followed by GUS staining (Fig. 4 and Supplementary Fig. S3). Strong GUS expression was observed in the whole tissue upon treatment with GA (1.0 µM), ABA (1.0 µM), and low concentration auxin (0.1 nM IAA), unlike the mock treatment showing a localized GUS staining in the tip, primordia, and elongation zone of the hairy root (Fig. 4a). GUS expression was lost upon treatment with ethylene (1.0 µM ACC: Fig. 4c). When the transgenic hairy roots were treated with the respective inhibitors (1.0 µM PAC, 1.0 µM Fluridon, and 1.0 µM TIBA), no GUS expression was observed, except in the case of 0.1 µM AgNO3 (ethylene signal inhibitor) inducing strong GUS expression (Fig. 4c). To examine stress-induced promoter activity, GUS expression was analyzed after osmotic stress (200 mM mannitol) and salt (150 mM NaCl) stress treatment. Similar to the hormonal response, stress-induced promoter-driven GUS expression was detected in the whole root tissue, including the differentiation zone (Fig. 5d and 5e) like the positive control (Fig. 5b). Further, the GUS enzymatic activity was quantified using the fluorimetric assay (Gallagher 1992). The GUS activity was higher in the GA (1.4-fold), ABA (1.4-fold), and auxin (2.2-fold) treated hairy roots, but highly suppressed in the case of ethylene treatment down to 30% (Fig. 6a). The GUS activity was 5.2-fold and 4.5-fold higher in osmotic and salt stresses, respectively (Fig. 6b).
Fig. 4.

Phytohormonal regulation of GhNAC2 promoter in cotton hairy roots. Promoter-driven GUS expression was examined in response to a, f, k Mock treatment, b GA3, c ACC (an ethylene precursor), d ABA, e IAA, their respective inhibitors: g PAC (GA biosynthesis inhibitor), h AgNO3 (ethylene signal inhibitor), i TIBA (auxin transport inhibitor), and j Fluridon (ABA biosynthesis inhibitor), and their combinations: l IAA and PAC, m (IAA and ACC, n IAA and AgNO3, and o ACC and TIBA. Strong GUS expression is indicated by arrows. The images represent the results obtained for 10–12 independent hairy roots for each treatment
Fig. 5.

Abiotic stress-induced regulation of GhNAC2 promoter in Arabidopsis (top) and cotton hairy roots (bottom). GUS histochemical staining was performed to study the promoter activity upon treatment with osmotic and salt stress. a Negative control (empty pBI101), b Positive control (empty pBI121), c Mock (no stress), d osmotic stress (mannitol), and e salt stress (NaCl) treatment. The images represent the results obtained for three independent T3 generation Arabidopsis lines and 10–12 independent cotton hairy roots for each treatment
Fig. 6.

Quantitative measurement of the GUS enzyme activity. GUS fluorometric assay was performed in the transgenic cotton hairy root tips to estimate the promoter-driven GUS expression upon a phytohormone and b stress treatment. The GUS activity was determined in terms of the pmol of 4-MU (4-methylumbelliferone) produced per mg of crude protein per min. All data are means ± SE (n = 9)
GhNAC2-promoted root growth is regulated by phytohormones in transgenic Arabidopsis
Our previous study revealed that GhNAC2 induces root growth when expressed in Arabidopsis (Gunapati et al. 2016). When the WT and transgenic lines (L1, L4, L8, and L15) expressing GhNAC2 were grown in 0.5X MSA, a significant increase of 21–40% was observed in the primary root length of transgenic lines (5.02 ± 0.09 cm, 4.35 ± 0.37 cm, 4.78 ± 0.53 cm, and 5.05 ± 0.02 cm) after 15 days compared to the WT (3.61 ± 0.10 cm). To understand the role of phytohormonal signaling in the GhNAC2-governed growth phenotype, the differential change in the primary root lengths of WT and transgenic Arabidopsis seedlings was studied in the presence and absence of exogenous GA, ethylene, ABA, auxin, and their respective biosynthesis/signal transduction inhibitors: PAC, AgNO3, Fluridon, and TIBA (Fig. 7 and Supplementary Fig. S4). When treated with 0.5 µM GA3, the root lengths in the transgenic lines were 23–28% longer than in the respective untreated control (6.26 ± 0.0.92 cm, 5.35 ± 0.88 cm, 6.03 ± 0.45 cm, and 6.47 ± 0.82 cm), whereas the root length in WT (4.97 ± 0.69 cm) was ~ 38% longer. The observations indicate a differential increase in the root growth of WT and transgenic lines due to GA treatment. In contrast, treatment of 1.0 µM PAC (an inhibitor GA biosynthesis) suppressed the root growth of both WT (2.08 ± 0.31 cm) and the transgenic lines (2.02 ± 0.0.22 cm, 2.01 ± 0.34 cm, 2.03 ± 0.18 cm, and 2.09 ± 0.36 cm), representing a differential decrease in root length of WT (42%) and transgenic lines (54–58%; Fig. 7a). This result indicated a GA-dependent root growth promotion mediated by GhNAC2.
Fig. 7.

GhNAC2-promoted root growth is controlled by phytohormone signaling. a GA3 differentially increased the root growth in WT and transgenic Arabidopsis lines (L1, L4, L8, and L15) expressing the GhNAC2 gene. Whereas in the presence of PAC, the root growth was decreased. b ACC and AgNO3 differentially suppressed and promoted root growth in WT and transgenic lines, respectively. c The transgenic lines showed less ABA sensitivity as compared to WT, whereas fluridon decreased the root growth of both WT and transgenic lines. d In the presence of IAA, no significant root stimulation was observed in either WT or transgenic lines. In addition, TIBA suppressed the root growth in both WT and transgenic lines. All data are means ± SE (n = 9). Different letters indicate significant differences between treatments and genotypes based on the Duncan’s multiple range test (a = p value < 0.001, b = p value < 0.01, and c = p value < 0.05)
Unlike GA, ethylene is an inhibitor of root growth (Shani et al. 2013; Street et al. 2015). The ethylene precursor, ACC (1.0 µM), differentially inhibited the root growth of WT (1.01 ± 0.09 cm, 40% decrease) and transgenic lines (1.40 ± 0.10 cm, 1.21 ± 0.11 cm, 1.20 ± 0.08 cm, and 1.49 ± 0.07 cm, 22–26% decrease). However, 0.1 µM AgNO3 (an ethylene signal inhibitor) induced the root length by 3% (WT) and 21–58% (transgenic lines) resulting in longer primary roots in the transgenic lines (2.30 ± 0.09 cm, 2.42 ± 0.30 cm, 2.63 ± 0.23 cm, and 2.45 ± 0.45 cm) than WT (1.72 ± 0.29 cm). The most contrasting changes in the root phenotype, in response to ethylene and its inhibitor, were observed after a 7-day treatment (Fig. 7b). This indicated that GhNAC2 regulates root growth by suppressing ethylene signaling. When treated with 1.0 µM ABA, a differential decrease in the root length was observed in WT (20%) and transgenic lines (4–12%), indicating less ABA sensitivity in the transgenic roots, while in the presence of 1.0 µM Fluridon (an ABA biosynthesis inhibitor), both WT (62%) and transgenic lines (52–62%) showed severe root growth inhibition. The most contrasting phenotype was observed after a 20-day treatment (Fig. 7c). Auxin is a master regulator of root growth and development, promoting root growth in a dose-dependent manner (Saini et al. 2013). In the presence of low-concentration auxin (0.1 µM IAA), root growth was induced in both WT (9%) and transgenic lines (3–13%). Similarly, treatment with 1.0 µM TIBA (an auxin transport inhibitor) resulted in decreased root length in both WT (32%) and transgenic lines (25–34%; Fig. 7d).
Discussion
Our previous study revealed promotion of root growth and drought tolerance by GhNAC2 in Arabidopsis and cotton. The spatio-temporal expression study in root, leaf, fibre, stem, and flower, showed that under normal growth conditions, GhNAC2 is predominantly expressed in root tissues. While microarray analysis had revealed differential regulation of the GA and ABA pathway and down-regulation of ethylene biosynthesis/signaling genes suggesting GhNAC2-mediated transcriptional modulation of hormonal metabolism and signaling (Gunapati et al. 2016). Here, we examined the induced expression of GhNAC2 in response to GA, ethylene, ABA, and auxin cotton roots (Fig. 1a) and tried to correlate these with cis-regulatory elements and TF binding sites identified in its promoter (Fig. 1c and d). Tissue/cell-specific study of the GhNAC2 promoter revealed preferential GUS expression in root tissues of Arabidopsis (Fig. 2g) that were further confirmed in cotton hairy roots for hormone-and stress-responsive promoter activities (Figs. 3, 4, 5 and 6). In addition, we also studied how exogenous supply of hormones and their inhibitors alter GhNAC2-mediated root growth in Arabidopsis (Fig. 8).
Fig. 8.
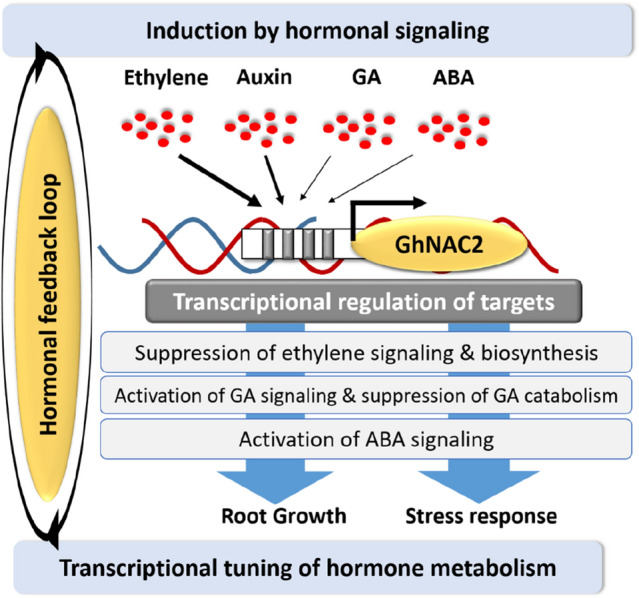
Phytohormonal signaling of GhNAC2 transcription factor. Ethylene, ABA, GA, and auxin are the upstream hormonal regulators determining GhNAC2 promoter activity. Further, the GhNAC2 protein regulates the downstream genes involved in the biosynthesis and/or signaling of ethylene, ABA, and GA. This suggested that the actions of GhNAC2 are governed by the ethylene/ABA/GA pathway through a feedback loop
In plants, the tissue-specific patterns of gene expression and interaction with environmental signals at various developmental stages are largely mediated by transcriptional regulation, which is fundamentally driven by the binding of transcription factors (TFs) to the gene promoter (Priest et al. 2009). The TFs (singly or in combination with other TFs and regulators) interact with specific cis-regulatory elements (CREs) in the distal promoter region to influence spatio-temporal gene expression. Thus, analyzing the CREs and TF-binding sites (TFBSs) in the promoter is a crucial part of the functional analysis of a gene.
The gene expression study showed regulation of GhNAC2 by GA, ethylene, ABA, auxin, mannitol, and NaCl in cotton roots (Fig. 1a). The highest fold change in expression was found in response to ABA and mannitol. This increase, that may drive its drought responsive function, may be explained partly by an over-representation of ABA responsive CREs such as ABRERATCAL (MACGYGB), an element recognized by ABRE-binding factors (ABFs) to induce Ca2+-dependent ABA response (Oh et al. 2005; Kaplan et al. 2006) (Supplementary Table S3). Similarly, CREs regulating GA signalling (both, positively and negatively) and also under the influence of ABA were found in the GhNAC2 promoter. Of these, PYRIMIDINEBOXOSRAMY1A (CCTTTT) might positively regulate GA-dependent activation (Mena et al. 2002) while the presence of GADOWNAT (ACGTGTC), generally present in genes that are down regulated by GA signaling during Arabidopsis seed germination (Ogawa et al. 2003) and the WRKY71OS (TGAC) motif, recognized by WRKY71 and responsible for blocking GA-induced activation in aleurone cells in rice (Zhang et al. 2004), may down regulate GhNAC2. Such counteracting GA-related elements might govern the antagonistic regulation of GA and ABA pathways to control seed germination, maturity, and dormancy (Liu and Hou 2018). The presence of the auxin-regulatory elements ARFAT (TGTCTC) and ASF1MOTIFCAMV (TGACG) that are targeted by ARF and TGA TFs, respectively (Hagen and Guilfoyle 2002; Li et al. 2019b) may control auxin-dependent growth responses as in Arabidopsis and rice while the over-representation of TF binding sites recognized by ERF (ethylene-responsive factors) may control auxin- and ethylene-responsive expression of GhNAC2 (Fig. 1a).
In Arabidopsis, AtMYC2 and MYB2 proteins bind to MYCCONSENSUSAT (CANNTG) and MYB2CONSENSUSAT (YAACKG) motifs to activate the expression of drought-responsive genes such as CBF2/DREB1C, CBF3, rd22, etc. (Abe et al. 2003) while CBFHV (RYCGAC) is recognized by HvCBF1 and HvCBF2 to activate cold- and drought-response in barley (Abedini et al. 2017). ACGTATERD1 (ACGT) motif regulates the early dehydration-responsive induction of rd22 while GT1GMSCAM4 (GAAAAA), present in SCaM4, interacts with GT1-like TFs to induce pathogen and NaCl response in soybean (Park et al. 2004). The abundance of these stress-regulatory motifs in GhNAC2 promoter correlates with the mannitol- and salt-induced expression of GhNAC2 (Fig. 1a).
In view of the expression of GhNAC2 in roots and its regulation by and response to several hormones, the promoter-GUS studies were performed for detailed in planta characterization. The promoter shows strong and predominant GUS expression in the roots of transgenic Arabidopsis (Fig. 2g and o), mainly in the root tips and elongation zones (Fig. 2h), root primordia (Fig. 2i), and root hairs (Fig. 2n), indicating root-preferred expression of GhNAC2 at seedling and rosette stage. In case of the shoot, localized GUS activity was observed in the elongation zone near the hypocotyl of germinating seedlings (Fig. 2b and c), leaf veins, and trichomes of rosette leaves consisting of vascular tissues and epidermal cells, respectively (Fig. 2l and m). The root-preferential expression was validated in the homologous system through studies in cotton hairy roots and showed cell-specific changes with localization of GUS expression to the epidermal layer of the elongation zone (Fig. 3c) while the root tips showed strong GUS expression in the cortex and stele as well (Fig. 3d). The presence of the ROOTMOTIFTAPOX1 (ATATT) and ACGTROOT1 (GCCACGTGGC) motifs in the GhNAC2 promoter, that are known for root-related expression in tobacco (Elmayan and Tepfer 1995; Welchen et al. 2009) may explain the root related expression in cotton.
The root tip and elongation zones of the root and shoot consist of meristematic tissue, unlike the differentiation zone. The presence of meristem-specific CREs, such as SITEIIATCYTC and LEAFYATAG (Kamiya et al. 2003; Welchen and Gonzalez 2005) might explain the higher GhNAC2 expression in the actively dividing tissues of the meristematic cells suggesting differential transcriptional reprogramming in these tissues by GhNAC2. A localized GUS expression was also noted in the stigma, anther, pollens, and immature seeds (Fig. 2q and w) and are correlated with the detection of pollen- and seed-specific CREs in the promoter (Ezcurra et al. 1999; Filichkin et al. 2004) (Supplementary Table S3) suggesting the involvement of GhNAC2 in seed and reproductive organ development. In support of this, several NAC genes such as OsNTL5, ZmNAC84, NST1/2, etc., have been reported to play significant roles in seed development and reproduction by regulating inflorescence fertility (Guo et al. 2018), pollen development (Yang et al. 2018), and anther dehiscence (Mitsuda et al. 2005).
When cotton hairy roots expressing the GUS reporter gene under a GhNAC2 promoter were treated with low concentrations of auxin, an intense GUS expression was detected (Fig. 4d and Supplementary Fig. S3d). A strong expression across the root was also observed upon treatment with GA and ABA (Fig. 4b, e and Supplementary Fig. S3c and S3e). However, ethylene significantly suppressed the promoter-driven GUS activity (Fig. 4c and Supplementary Fig. S3c). Expectedly, treatment with inhibitors of their biosynthesis such as paclobutrazol (GA biosynthesis), fluridone (ABA biosynthesis), TIBA (auxin transport) reduced GUS expression in most tissues with the exception of the root tip while AgNO3 (ethylene perception inhibitor) promoted GUS expression (Fig. 4g–j). Combination of treatments however provided interesting results hinting at more complex regulation. Combined treatment with PAC and auxin resulted in a loss in activity, indicating that auxin was unable to activate the GhNAC2 promoter if GA levels were limiting (Fig. 4l). In tune with this, when PAC and IAA were administered simultaneously in growth media, no increment in root length was observed in the transgenic lines compared to WT (Supplementary Fig. 4a) confirming that auxin-mediated promotion of root growth requires GA (Fu and Harberd 2003). High ethylene levels inhibited the low auxin-induced promoter activity (Fig. 4m). Ethylene is known to direct the efflux of auxin and may thereby suppress auxin effects (Muday et al. 2012). Interestingly, AgNO3 also suppressed the auxin response suggesting that an intact ethylene signaling pathway was also necessary for auxin effects (Strader et al. 2009). Surprisingly, simultaneous treatment with ethylene and TIBA caused activation of promoter-driven GUS expression (Fig. 4o). Since both individually suppress GhNAC2 expression, the activation by a combination of both probably leads to latent activation of GhNAC2 by other hormone pathways that will require further studies. The GUS-reporter-aided analysis of promoter activities suggests that auxin, ABA, ethylene and GA are major upstream hormonal regulators of GhNAC2. Mannitol and NaCl treatment also resulted in stronger promoter activity, validating the GhNAC2 induction in response to osmotic- and salt-stress (Fig. 5d, e, 6b, and Fig. 1a) and may probably involve the ABA/ethylene pathways. The phytohormone and stress-induced GUS expression was not localized, but detected in whole tissue including the differentiation zone, unlike the basal expression (Figs. 4 and 5). This indicates that the function of GhNAC2 is tissue- and organ-specific under normal conditions. Indeed, hormone and stress-directed expression of GhNAC2 might govern the developmental responses and plant-environment interactions.
Root growth regulation by NAC proteins is directed by multiple phytohormonal signals in a dose-dependent manner to exert synergistic, antagonistic, or compensative actions (Jensen et al. 2010). In the GhNAC2-expressing transgenic Arabidopsis, root growth promotion was observed under every treatment except when GA biosynthesis was inhibited through PAC treatment suggesting that endogenous GA was necessary for GhNAC2 action (Fig. 7a). In support of this, GA2OX2, a GA catabolic gene was down regulated in GhNAC2-expressing transgenic lines (Gunapati et al. 2016; Mach 2017) while GhNAC2 is significantly induced by exogenous GA (Fig. 1a). Thus the GA pathway seems to play a direct role in the regulation of GhNAC2-mediated root elongation through a feedback loop. Indeed, GA treatment could promote root growth in the GhNAC2-expressing Arabidopsis lines despite TIBA-inhibited auxin transport (Supplementary Fig. S4b) suggesting that GA signaling is crucial for auxin-dependent promoter activity (Fig. 4l) and GA-dependent root growth. GA is known to promote root growth by stimulating root meristem elongation when accumulated in dividing endodermal cells or supplied exogenously at low concentrations (Tanimoto 2012; Shani et al. 2013).
In contrast, root growth inhibition through negative regulation of meristem cell proliferation and epidermal elongation is one of the characteristics of ethylene signaling (Street et al. 2015; Vaseva et al. 2018). The suppression of ethylene signaling by AgNO3 stimulated the root growth in the GhNAC2-expressing Arabidopsis lines (Fig. 7b). Moreover, AgNO3 treatment could mitigate the inhibitory effects of high auxin concentration on the root growth in the transgenic lines (Supplementary Fig. S4c). This phenotype is supported by the microarray study (Gunapati et al. 2016) suggesting the down-regulation of ethylene biosynthesis and signaling genes in the transgenic Arabidopsis lines. Thus, GhNAC2 is a negative regulator of ethylene-induced root inhibition. ABA also inhibits root growth via ethylene biosynthesis (Luo et al. 2014). Less ABA-induced root growth inhibition in the transgenic roots suggested increased ethylene suppression through GhNAC2 transcriptional regulation (Fig. 7c). Nevertheless, auxin treatment, as well as inhibition of auxin signaling, caused no differential effect on the root growth of transgenic lines and WT seedlings (Fig. 7d). This observation suggests that activation of GA pathway and suppression of ethylene signaling determines the GhNAC2-mediated root growth stimulation, in an auxin independent manner.
Concluding remarks
The study suggests that the GhNAC2 transcription factor governs crucial growth and stress-related functions through differential promoter regulation during optimum and non-favorable environmental conditions though a combination of hormonal signals, such as GA, ABA, ethylene and auxin. Activation of GA and suppression of ethylene signaling play a prominent role in stimulating the promoter activity resulting in enhanced root growth due to decreased GA catabolism and ethylene biosynthesis conferred by GhNAC2. The tissue-preferential and localized promoter activity in growing tissues such as the root tip and elongation zone indicates the specific role of GhNAC2 in signaling growth responses. Such a balanced mechanism that governs growth and stress tolerance with fewer unwanted trade-offs is useful for agricultural applications where stress adaptation is desired along with simultaneous gain in biomass and yield (Srivastava and Sahoo 2021). Thus, the native promoter could be utilized to further improve GhNAC2-controlled phenotypes in cotton to develop improved root architecture and yield of quality fiber.
Supplementary Information
Below is the link to the electronic supplementary material.
Acknowledgements
We thank Dr. Praveen C. Verma (Principal Scientist), Division of Molecular Biology and Biotechnology, CSIR-NBRI, for providing the Agrobacterium rhizogenes A4 strain. We are grateful to Dr. P. A. Shirke (Chief Scientist) and Dr. Baleshwar (Principal Scientist), CSIR-NBRI, for availing of the microscopy facility. Dr. Manoj Kumar (Senior Scientist), CSIR-NBRI, provided the micro-sectioning facility. Mr. Ram Awadh is acknowledged for the care and maintenance of transgenic plants.
Author contributions
VAS and APS conceptualized and designed the study. RN, SG, and RS carried out the experiments. Data analysis and interpretation were performed by RN, RS, VAS, and APS. RS and RN wrote the manuscript. APS and VAS reviewed and edited the manuscript. All authors read and approved the final version of the manuscript.
Funding
VAS acknowledges project fund GAP3427 from DST-Science and Engineering Research Board (SERB) and research fund MLP0035 from Council of Scientific & Industrial Research (CSIR); RN acknowledges Department of Biotechnology (DBT), India, for JRF fellowship for this study.
Declarations
Conflict of interest
All authors declare not having competing or conflicting interests.
Data availability statement
The promoter sequence (Accession number: OQ190160) is available in the Genbank repository (https://www.ncbi.nlm.nih.gov/genbank/). Other data generated or analyzed during this study are included in this manuscript and supplementary information files.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell. 2003;15(1):63–78. doi: 10.1105/tpc.006130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abedini R, GhaneGolmohammadi F, PishkamRad R, Pourabed E, Jafarnezhad A, Shobbar Z-S, Shahbazi M. Plant dehydrins: shedding light on structure and expression patterns of dehydrin gene family in barley. J Plant Res. 2017;130(4):747–763. doi: 10.1007/s10265-017-0941-5. [DOI] [PubMed] [Google Scholar]
- Ali Q, Salisu IB, Raza A, Shahid AA, Rao AQ, Husnain T. A modified protocol for rapid DNA isolation from cotton (Gossypium spp.) MethodsX. 2019;6:259–264. doi: 10.1016/j.mex.2019.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asif MH, Dhawan P, Nath P. A simple procedure for the isolation of high quality RNA from ripening banana fruit. Plant Mol Biol Rep. 2000;18(2):109–115. doi: 10.1007/BF02824018. [DOI] [Google Scholar]
- Biłas R, Szafran K, Hnatuszko-Konka K, Kononowicz AK. Cis-regulatory elements used to control gene expression in plants. Plant Cell Tissue Organ Cult. 2016;127(2):269–287. doi: 10.1007/s11240-016-1057-7. [DOI] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72(1–2):248–254. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
- Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16(6):735–743. doi: 10.1046/j.1365-313x.1998.00343. [DOI] [PubMed] [Google Scholar]
- Dabbert T, Gore MA. Challenges and perspectives on improving heat and drought stress resilience in cotton. J Cotton Sci. 2014;18(3):393–409. [Google Scholar]
- Das A, Pramanik K, Sharma R, Gantait S, Banerjee J. In-silico study of biotic and abiotic stress-related transcription factor binding sites in the promoter regions of rice germin-like protein genes. PLoS ONE. 2019;14(2):e0211887. doi: 10.1371/journal.pone.0211887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elmayan T, Tepfer M. Evaluation in tobacco of the organ specificity and strength of therolD promoter, domain A of the 35S promoter and the 35S2 promoter. Transg Res. 1995;4(6):388–396. doi: 10.1007/BF01973757. [DOI] [PubMed] [Google Scholar]
- Ezcurra I, Ellerström M, Wycliffe P, Stålberg K, Rask L. Interaction between composite elements in the napA promoter: both the B-box ABA-responsive complex and the RY/G complex are necessary for seed-specific expression. Plant Mol Biol. 1999;40(4):699–709. doi: 10.1023/a:1006206124512. [DOI] [PubMed] [Google Scholar]
- Filichkin SA, Leonard JM, Monteros A, Liu P-P, Nonogaki H. A novel endo-β-mannanase gene in tomato LeMAN5 is associated with anther and pollen development. Plant Physiol. 2004;134(3):1080–1087. doi: 10.1104/pp.103.035998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu X, Harberd NP. Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature. 2003;421(6924):740–743. doi: 10.1038/nature01387. [DOI] [PubMed] [Google Scholar]
- Fujita Y, Yoshida T, Yamaguchi-Shinozaki K. Pivotal role of the AREB/ABF-SnRK2 pathway in ABRE-mediated transcription in response to osmotic stress in plants. Physiol Plant. 2013;147(1):15–27. doi: 10.1111/j.1399-3054.2012.01635.x. [DOI] [PubMed] [Google Scholar]
- Gallagher SR (1992) Quantitation of GUS activity by fluorometry. In: GUS protocols: using the GUS gene as a reporter of gene expression, pp 47–59
- Gao M, Snider JL, Bai H, Hu W, Wang R, Meng Y, Wang Y, Chen B, Zhou Z. Drought effects on cotton (Gossypium hirsutum L.) fibre quality and fibre sucrose metabolism during the flowering and boll-formation period. J Agron Crop Sci. 2020;206(3):309–321. doi: 10.1111/jac.12389. [DOI] [Google Scholar]
- Gu Z, Huang C, Li F, Zhou X. A versatile system for functional analysis of genes and micro RNA s in cotton. Plant Biotechnol J. 2014;12(5):638–649. doi: 10.1111/pbi.12169. [DOI] [PubMed] [Google Scholar]
- Gunapati S, Naresh R, Ranjan S, Nigam D, Hans A, Verma PC, Gadre R, Pathre UV, Sane AP, Sane VA. Expression of GhNAC2 from G herbaceum, improves root growth and imparts tolerance to drought in transgenic cotton and Arabidopsis. Sci Rep. 2016;6(1):1–14. doi: 10.1038/srep24978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo Y, Pang C, Jia X, Ma Q, Dou L, Zhao F, Gu L, Wei H, Wang H, Fan S. An NAM domain gene, GhNAC79, improves resistance to drought stress in upland cotton. Front Plant Sci. 2017;8:1657. doi: 10.3389/fpls.2017.01657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo S, Dai S, Singh PK, Wang H, Wang Y, Tan JL, Wee W, Ito T. A membrane-bound NAC-like transcription factor OsNTL5 represses the flowering in Oryza sativa. Front Plant Sci. 2018;9:555. doi: 10.3389/fpls.2018.00555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagen G, Guilfoyle T. Auxin-responsive gene expression: genes, promoters and regulatory factors. Plant Mol Biol. 2002;49(3):373–385. doi: 10.1023/A:1015207114117. [DOI] [PubMed] [Google Scholar]
- He XJ, Mu RL, Cao WH, Zhang ZG, Zhang JS, Chen SY. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005;44(6):903–916. doi: 10.1111/j.1365-313X.2005.02575.x. [DOI] [PubMed] [Google Scholar]
- Higo K, Ugawa Y, Iwamoto M, Korenaga T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucl Acids Res. 1999;27(1):297–300. doi: 10.1093/nar/27.1.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoagland DR, Arnon DI. The water-culture method for growing plants without soil. Calif Agric Exp Stn Circular. 1950;347:32. [Google Scholar]
- Jefferson RA, Kavanagh TA, Bevan MW. GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987;6(13):3901–3907. doi: 10.1002/j.1460-2075.1987.tb02730.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen MK, Kjaersgaard T, Petersen K, Skriver K, behavior, NAC genes: time-specific regulators of hormonal signaling in Arabidopsis. Plant Signal Behav. 2010;5(7):907–910. doi: 10.4161/psb.5.7.12099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen MK, Lindemose S, De Masi F, Reimer JJ, Nielsen M, Perera V, Workman CT, Turck F, Grant MR, Mundy J. ATAF1 transcription factor directly regulates abscisic acid biosynthetic gene NCED3 in Arabidopsis thaliana. FEBS Open Bio. 2013;3:321–327. doi: 10.1016/j.fob.2013.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnsson C, Jin X, Xue W, Dubreuil C, Lezhneva L, Fischer U. The plant hormone auxin directs timing of xylem development by inhibition of secondary cell wall deposition through repression of secondary wall NAC-domain transcription factors. Physiol Plant. 2019;165(4):673–689. doi: 10.1111/ppl.12766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamiya N, Nagasaki H, Morikami A, Sato Y, Matsuoka M. Isolation and characterization of a rice WUSCHEL-type homeobox gene that is specifically expressed in the central cells of a quiescent center in the root apical meristem. Plant J. 2003;35(4):429–441. doi: 10.1046/j.1365-313x.2003.01816.x. [DOI] [PubMed] [Google Scholar]
- Kaplan B, Davydov O, Knight H, Galon Y, Knight MR, Fluhr R, Fromm H. Rapid transcriptome changes induced by cytosolic Ca2+ transients reveal ABRE-related sequences as Ca2+-responsive cis elements in Arabidopsis. Plant Cell. 2006;18(10):2733–2748. doi: 10.1105/tpc.106.042713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim SG, Lee AK, Yoon HK, Park CM. A membrane-bound NAC transcription factor NTL8 regulates gibberellic acid-mediated salt signaling in Arabidopsis seed germination. Plant J. 2008;55(1):77–88. doi: 10.1111/j.1365-313X.2008.03493.x. [DOI] [PubMed] [Google Scholar]
- Kou X, Zhou J, Wu CE, Yang S, Liu Y, Chai L, Xue Z. The interplay between ABA/ethylene and NAC TFs in tomato fruit ripening: a review. Plant Mol Biol. 2021;106(3):223–238. doi: 10.1007/s11103-021-01128-w. [DOI] [PubMed] [Google Scholar]
- Kulkarni VN, Khadi BM, Maralappanavar MS, Deshapande LA, Narayanan S (2009) The worldwide gene pools of Gossypium arboreum L. and G. herbaceum L., and their improvement. In: Genetics and genomics of cotton. Springer, pp 69–97. 10.1007/978-0-387-70810-2_4
- Li J, Zhao Y, Chu H, Wang L, Fu Y, Liu P, Upadhyaya N, Chen C, Mou T, Feng Y. SHOEBOX modulates root meristem size in rice through dose-dependent effects of gibberellins on cell elongation and proliferation. PLoS Genet. 2015;11(8):e1005464. doi: 10.1371/journal.pgen.1005464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li H-M, Liu S-D, Ge C-W, Zhang X-M, Zhang S-P, Chen J, Shen Q, Ju F-Y, Yang Y-F, Li Y. Association analysis of drought tolerance and associated traits in upland cotton at the seedling. Int J Mol Sci. 2019;20(16):3888. doi: 10.3390/ijms20163888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S-B, Xie Z-Z, Hu C-G, Zhang J-Z. A review of auxin response factors (ARFs) in plants. Front Plant Sci. 2016;7:47. doi: 10.3389/fpls.2016.00047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li N, Muthreich M, Huang LJ, Thurow C, Sun T, Zhang Y, Gatz C. TGACG-BINDING FACTORs (TGAs) and TGA-interacting CC-type glutaredoxins modulate hyponastic growth in Arabidopsis thaliana. New Phytol. 2019;221(4):1906–1918. doi: 10.1111/nph.15496. [DOI] [PubMed] [Google Scholar]
- Liu X, Hou X. Antagonistic regulation of ABA and GA in metabolism and signaling pathways. Front Plant Sci. 2018;9:251. doi: 10.3389/fpls.2018.00251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods. 2001;25(4):402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- Luo X, Chen Z, Gao J, Gong Z. Abscisic acid inhibits root growth in Arabidopsis through ethylene biosynthesis. Plant J. 2014;79(1):44–55. doi: 10.1111/tpj.12534. [DOI] [PubMed] [Google Scholar]
- Mach J. A time to divide and a time to expand: histone deacetylases flip a gibberellin oxidase-mediated switch in root meristem cells. Plant Cell. 2017;29(9):2082–2083. doi: 10.1105/tpc.17.00732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mena M, Cejudo FJ, Isabel-Lamoneda I, Carbonero P. A role for the DOF transcription factor BPBF in the regulation of gibberellin-responsive genes in barley aleurone. Plant Physiol. 2002;130(1):111–119. doi: 10.1104/pp.005561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meshram JH, Mahajan SS, Nagrale D, Gokte-Narkhedkar N, Kumbhalkar H. Understanding root biology for enhancing cotton production. Plant Roots. 2021 doi: 10.5772/intechopen.95547. [DOI] [Google Scholar]
- Misra A, Khan K, Niranjan A, Kumar V, Sane VA. Heterologous expression of two GPATs from Jatropha curcas alters seed oil levels in transgenic Arabidopsis thaliana. Plant Sci. 2017;263:79–88. doi: 10.1016/j.plantsci.2017.07.003. [DOI] [PubMed] [Google Scholar]
- Mitsuda N, Seki M, Shinozaki K, Ohme-Takagi M. The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell. 2005;17(11):2993–3006. doi: 10.1105/tpc.105.036004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muday GK, Rahman A, Binder BM. Auxin and ethylene: collaborators or competitors? Trends Plant Sci. 2012;17(4):181–195. doi: 10.1016/j.tplants.2012.02.001. [DOI] [PubMed] [Google Scholar]
- Nakashima K, Takasaki H, Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K. NAC transcription factors in plant abiotic stress responses. Biochim Biophys Acta. 2012;1819(2):97–103. doi: 10.1016/j.bbagrm.2011.10.005. [DOI] [PubMed] [Google Scholar]
- Niu J, Zhang S, Liu S, Ma H, Chen J, Shen Q, Ge C, Zhang X, Pang C, Zhao X. The compensation effects of physiology and yield in cotton after drought stress. J Plant Physiol. 2018;224:30–48. doi: 10.1016/j.jplph.2018.03.001. [DOI] [PubMed] [Google Scholar]
- Nuruzzaman M, Sharoni AM, Kikuchi S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front Microbiol. 2013;4:248. doi: 10.3389/fmicb.2013.00248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ogawa M, Hanada A, Yamauchi Y, Kuwahara A, Kamiya Y, Yamaguchi S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell. 2003;15(7):1591–1604. doi: 10.1105/tpc.011650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh S-J, Song SI, Kim YS, Jang H-J, Kim SY, Kim M, Kim Y-K, Nahm BH, Kim J-K. Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol. 2005;138(1):341–351. doi: 10.1104/pp.104.059147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacifici E, Polverari L, Sabatini S. Plant hormone cross-talk: the pivot of root growth. J Exp Bot. 2015;66(4):1113–1121. doi: 10.1093/jxb/eru534. [DOI] [PubMed] [Google Scholar]
- Park HC, Kim ML, Kang YH, Jeon JM, Yoo JH, Kim MC, Park CY, Jeong JC, Moon BC, Lee JH. Pathogen-and NaCl-induced expression of the SCaM-4 promoter is mediated in part by a GT-1 box that interacts with a GT-1-like transcription factor. Plant Physiol. 2004;135(4):2150–2161. doi: 10.1104/pp.104.041442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park J, Kim Y-S, Kim S-G, Jung J-H, Woo J-C, Park C-M. Integration of auxin and salt signals by the NAC transcription factor NTM2 during seed germination in Arabidopsis. Plant Physiol. 2011;156(2):537–549. doi: 10.1104/pp.111.177071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Priest HD, Filichkin SA, Mockler TC. Cis-regulatory elements in plant cell signaling. Curr Opin Plant Biol. 2009;12(5):643–649. doi: 10.1016/j.pbi.2009.07.016. [DOI] [PubMed] [Google Scholar]
- Qin H, Huang R. Auxin controlled by ethylene steers root development. Int J Mol Sci. 2018;19(11):3656. doi: 10.3390/ijms19113656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rombauts S, Déhais P, Van Montagu M, Rouzé P. PlantCARE, a plant cis-acting regulatory element database. Nucl Acids Res. 1999;27(1):295–296. doi: 10.1093/nar/27.1.295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saini S, Sharma I, Kaur N, Pati PK. Auxin: a master regulator in plant root development. Plant Cell Rep. 2013;32(6):741–757. doi: 10.1007/s00299-013-1430-5. [DOI] [PubMed] [Google Scholar]
- Sánchez-Montesino R, Bouza-Morcillo L, Marquez J, Ghita M, Duran-Nebreda S, Gómez L, Holdsworth MJ, Bassel G, Oñate-Sánchez L. A regulatory module controlling GA-mediated endosperm cell expansion is critical for seed germination in Arabidopsis. Mol Plant. 2019;12(1):71–85. doi: 10.1016/j.molp.2018.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shahnejat-Bushehri S, Tarkowska D, Sakuraba Y, Balazadeh S. Arabidopsis NAC transcription factor JUB1 regulates GA/BR metabolism and signalling. Nat Plants. 2016;2(3):1–9. doi: 10.1038/nplants.2016.13. [DOI] [PubMed] [Google Scholar]
- Shani E, Weinstain R, Zhang Y, Castillejo C, Kaiserli E, Chory J, Tsien RY, Estelle M. Gibberellins accumulate in the elongating endodermal cells of Arabidopsis root. Proc Natl Acad Sci. 2013;110(12):4834–4839. doi: 10.1073/pnas.1300436110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava R, Sahoo L. Balancing yield trade-off in legumes during multiple stress tolerance via strategic crosstalk by native NAC transcription factors. J Plant Biochem Biotechnol. 2021;30:708–729. doi: 10.1007/s13562-021-00749-y. [DOI] [Google Scholar]
- Srivastava R, Kobayashi Y, Koyama H, Sahoo L. Cowpea NAC1/NAC2 transcription factors improved growth and tolerance to drought and heat in transgenic cowpea through combined activation of photosynthetic and antioxidant mechanisms. J Integr Plant Biol. 2022 doi: 10.1111/jipb.13365. [DOI] [PubMed] [Google Scholar]
- Srivastava R, Kobayashi Y, Koyama H, Sahoo L. Overexpression of cowpea NAC transcription factors promoted growth and stress tolerance by boosting photosynthetic activity in Arabidopsis. Plant Sci. 2022;319:111251. doi: 10.1016/j.plantsci.2022.111251. [DOI] [PubMed] [Google Scholar]
- Strader LC, Beisner ER, Bartel B. Silver ions increase auxin efflux independently of effects on ethylene response. Plant Cell. 2009;21(11):3585–3590. doi: 10.1105/tpc.108.065185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Street IH, Aman S, Zubo Y, Ramzan A, Wang X, Shakeel SN, Kieber JJ, Schaller GE. Ethylene inhibits cell proliferation of the Arabidopsis root meristem. Plant Physiol. 2015;169(1):338–350. doi: 10.1104/pp.15.00415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Q, Huang J, Guo Y, Yang M, Guo Y, Li J, Zhang J, Xu W. A cotton NAC domain transcription factor, GhFSN5, negatively regulates secondary cell wall biosynthesis and anther development in transgenic Arabidopsis. Plant Physiol Biochem. 2020;146:303–314. doi: 10.1016/j.plaphy.2019.11.030. [DOI] [PubMed] [Google Scholar]
- Takasaki H, Maruyama K, Kidokoro S, Ito Y, Fujita Y, Shinozaki K, Yamaguchi-Shinozaki K, Nakashima K. The abiotic stress-responsive NAC-type transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in rice. Mol Genet Genomic. 2010;284(3):173–183. doi: 10.1007/s00438-010-0557-0. [DOI] [PubMed] [Google Scholar]
- Tanimoto E. Tall or short? Slender or thick? A plant strategy for regulating elongation growth of roots by low concentrations of gibberellin. Ann Bot. 2012;110(2):373–381. doi: 10.1093/aob/mcs049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tian F, Yang D-C, Meng Y-Q, Jin J, Gao G. PlantRegMap: charting functional regulatory maps in plants. Nucl Acids Res. 2020;48(D1):D1104–D1113. doi: 10.1093/nar/gkz1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trishla VS, Marriboina S, Boyidi P, Kirti PB. GUS-reporter based analysis of the promoter activity of Gossypium hirsutum NAC transcription factor, GhNAC4 that is induced by phytohormones and environmental stresses. Plant Cell Tissue Organ Cult. 2020;141(3):643–654. doi: 10.1007/s11240-020-01825-2. [DOI] [Google Scholar]
- Vaseva II, Qudeimat E, Potuschak T, Du Y, Genschik P, Vandenbussche F, Van Der Straeten D. The plant hormone ethylene restricts Arabidopsis growth via the epidermis. Proc Natl Acad Sci. 2018;115(17):E4130–E4139. doi: 10.1073/pnas.1717649115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verma PC, Negi AS, Jain DC, Khanuja S, Banerjee S. Agrobacterium rhizogenes-mediated transformation of Picrorhiza kurroa Royle ex Benth.: establishment and selection of superior hairy root clone. Plant Biotechnol Rep. 2007;1(3):169–174. doi: 10.1007/s11816-007-0029-0. [DOI] [Google Scholar]
- Welchen E, Gonzalez DH. Differential expression of the Arabidopsis cytochrome c genes Cytc-1 and Cytc-2. Evidence for the involvement of TCP-domain protein-binding elements in anther-and meristem-specific expression of the Cytc-1 gene. Plant Physiol. 2005;139(1):88–100. doi: 10.1104/pp.105.065920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welchen E, Viola IL, Kim HJ, Prendes LP, Comelli RN, Hong JC, Gonzalez DH. A segment containing a G-box and an ACGT motif confers differential expression characteristics and responses to the Arabidopsis Cytc-2 gene, encoding an isoform of cytochrome c. J Exp Bot. 2009;60(3):829–845. doi: 10.1093/jxb/ern331. [DOI] [PubMed] [Google Scholar]
- Xie Z, Nolan TM, Jiang H, Yin Y. AP2/ERF transcription factor regulatory networks in hormone and abiotic stress responses in Arabidopsis. Front Plant Sci. 2019;10:228. doi: 10.3389/fpls.2019.00228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu P, Ma W, Hu J, Cai W. The nitrate-inducible NAC transcription factor NAC056 controls nitrate assimilation and promotes lateral root growth in Arabidopsis thaliana. PLoS Genet. 2022;18(3):e1010090. doi: 10.1371/journal.pgen.1010090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Q, Zhang H, Liu C, Huang L, Zhao L, Zhang A. A NAC transcription factor ZmNAC84 affects pollen development through the repression of ZmRbohH expression in maize. J Plant Biol. 2018;61(6):366–373. doi: 10.1007/s12374-018-0227-z. [DOI] [Google Scholar]
- Yarra R, Wei W. The NAC-type transcription factor GmNAC20 improves cold, salinity tolerance, and lateral root formation in transgenic rice plants. Funct Integr Genomics. 2021;21(3):473–487. doi: 10.1007/s10142-021-00790-z. [DOI] [PubMed] [Google Scholar]
- Yu J, Mao C, Zhong Q, Yao X, Li P, Liu C, Ming F. OsNAC2 is involved in multiple hormonal pathways to mediate germination of rice seeds and establishment of seedling. Front Plant Sci. 2021 doi: 10.3389/fpls.2021.699303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zahid Z, Khan MKR, Hameed A, Akhtar M, Ditta A, Hassan HM, Farid G. Dissection of drought tolerance in upland cotton through morpho-physiological and biochemical traits at seedling stage. Front Plant Sci. 2021;12:627107. doi: 10.3389/fpls.2021.627107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z-L, Xie Z, Zou X, Casaretto J, T-hD Ho, Shen QJ. A rice WRKY gene encodes a transcriptional repressor of the gibberellin signaling pathway in aleurone cells. Plant Physiol. 2004;134(4):1500–1513. doi: 10.1104/pp.103.034967. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The promoter sequence (Accession number: OQ190160) is available in the Genbank repository (https://www.ncbi.nlm.nih.gov/genbank/). Other data generated or analyzed during this study are included in this manuscript and supplementary information files.