

Abstract
Interaction of the human immunodeficiency virus type 1 (HIV-1) gp120 envelope glycoprotein with the primary receptor, CD4, promotes binding to a chemokine receptor, either CCR5 or CXCR4. The chemokine receptor-binding site on gp120 elicits CD4-induced (CD4i) antibodies in some HIV-1-infected individuals. Like CCR5 itself, the CD4i antibody 412d exhibits a preference for CCR5-using HIV-1 strains and utilizes sulfated tyrosines to achieve binding to gp120. Here, we show that 412d binding requires the gp120 β19 strand and the base of the V3 loop, elements that are important for the binding of the CCR5 N terminus. Two gp120 residues in the V3 loop base determined 412d preference for CCR5-using HIV-1 strains. A chimeric molecule in which the 412d heavy-chain third complementarity-determining loop sequence replaces the CCR5 N terminus functioned as an efficient second receptor, selectively supporting the entry of CCR5-using HIV-1 strains. Sulfation of N-terminal tyrosines contributed to the function of this chimeric receptor. These results emphasize the close mimicry of the CCR5 N terminus by the gp120-interactive region of a naturally elicited CD4i antibody.
Human immunodeficiency virus type 1 (HIV-1) entry into target cells is mediated by sequential binding to the primary receptor, CD4, and either of two coreceptors, CCR5 or CXCR4 (1, 3, 7, 13, 14, 20, 29). CD4 binding to the HIV-1 gp120 exterior envelope glycoprotein results in a change in gp120 conformation that is favorable for CCR5 or CXCR4 binding (42, 43). Receptor binding is thought to trigger further conformational changes in the HIV-1 envelope glycoproteins, ultimately leading to fusion of the viral and cell membranes.
The binding sites for CD4 and the CCR5 or CXCR4 chemokine receptors on the HIV-1 gp120 glycoprotein are potential targets for intervention. The HIV-1 gp120 glycoprotein is composed of regions conserved among virus strains (C1 to C5) and regions that exhibit significant variation (V1 to V5). The binding site for CD4 has been visualized by x-ray crystallography and includes a highly conserved pocket on the gp120 surface (25, 26). The gp120 structures involved in chemokine receptor binding include the well-conserved β19 strand and the third variable (V3) loop, which governs chemokine receptor choice (2, 23, 30, 35, 39). Most of the HIV-1 strains that are transmitted horizontally and that predominate in the first few years of infection utilize CCR5 as a coreceptor (28, 31, 33). Thus, understanding gp120-CCR5 interaction may facilitate the development of effective therapies and vaccines.
Like all G protein-coupled receptors, CCR5 and CXCR4 are thought to span the membrane seven times. The CCR5 N terminus and second extracellular loop have been shown to be important for the ability of the receptor to support HIV-1 entry (15, 16). The CCR5 N terminus is electronegative; in addition to being rich in acidic residues, several of the tyrosines in this segment are sulfated posttranslationally (18). The negatively charged tyrosine sulfates contribute to the efficiency of gp120 binding and HIV-1 entry (8). Sulfated peptides corresponding in sequence to the CCR5 N terminus bind gp120 glycoproteins from CCR5-using (R5) HIV-1 strains after incubation with soluble CD4 (sCD4) (11, 17, 19). Studies of gp120 mutants suggest that the binding of the CCR5 N terminus requires sequences in the β19 strand and the base of the V3 loop (12, 17). Other gp120 sequences near the tip of the V3 loop are thought to contribute to the ability of gp120 to interact with the body of the chemokine receptor (22, 34, 35). Both sets of interaction are required for functional, high-affinity gp120-CCR5 binding leading to virus entry.
The binding sites for CD4 and chemokine receptor on the HIV-1 gp120 envelope glycoprotein serve as targets for neutralizing antibodies generated during natural infection. Some potent neutralizing antibodies, such as immunoglobulin G1b12, bind near the CD4-binding site of gp120 (38, 48). Other monoclonal antibodies derived from HIV-1-infected individuals recognize a conserved gp120 structure that is closely related to the chemokine receptor-binding site. The binding of these antibodies to gp120 is induced by CD4 binding; hence, they are designated CD4-induced (CD4i) antibodies (41). CD4i antibodies block the binding of gp120-sCD4 complexes to the chemokine receptors (42, 43). CD4i antibodies exhibit various degrees of potency in neutralizing HIV-1. The neutralizing efficacy of CD4i antibodies is limited by steric constraints on antibody binding after the HIV-1 envelope glycoproteins have engaged the CD4 glycoprotein on the target cell surface (27). Some CD4i antibodies have long complementarity-determining region 3 (CDR3) loops on their heavy chains (8, 21). These long CDR3 loops may allow CD4i antibodies to access conserved gp120 structures in the β19 strand and V3 base, bypassing the variable gp120 loops that protect these conserved elements. In some cases, sulfated tyrosine residues on the heavy-chain CDR3 loops contribute to the interaction of the CD4i antibody with HIV-1 gp120 (8). Thus, several of the CD4i antibodies appear to mimic the CCR5 receptor.
One CD4i antibody, 412d, preferentially recognizes gp120 glycoproteins from CCR5-using HIV-1 strains (8). When a number of gp120 glycoproteins from different HIV-1 variants were examined, a strong correlation was observed between 412d binding and the ability of the gp120 glycoprotein to use CCR5 as a second receptor. Thus, the 412d antibody appears to mimic CCR5 quite closely. Here, we characterize the gp120 epitope for the 412d antibody. To examine whether the structural similarity of the 412d antibody and CCR5 translates into functional similarity, we replaced the CCR5 N terminus by sequences of the 412d heavy-chain CDR3 loop. The ability of these chimeric receptors to support the entry of a variety of HIV-1 strains was tested.
MATERIALS AND METHODS
Cell lines and antibodies.
The 293T cells were used for expressing HIV-1 gp120 proteins and also for producing recombinant viruses. Cf2Th cells were used for expressing CD4 and CCR5 or CXCR4 to study the entry of viruses and antibody neutralization.
The human monoclonal antibody 412d was isolated from an Epstein-Barr virus-transformed B-cell line, which was established from an HIV-1-infected individual (AC-01) undergoing early structured treatment interruption (37). Epstein-Barr virus transformation of B cells was performed as previously described (36). The antibody was purified on protein A-Sepharose and eluted with 3 M MgCl2 at room temperature.
Construction of HIV-1 envelope glycoprotein and CCR5 mutants.
The panel of HIV-1 YU2 gp120 mutants used for 412d epitope mapping was made by site-directed mutagenesis as previously described (35). The starting plasmid, YU2wtΔ, encodes a gp120 derivative from the YU2 HIV-1 strain containing deletions of gp120 residues 31 to 81 and 128 to 194, which represent the N terminus and V1/V2 variable loops, respectively.
The overlap-PCR approach was employed to generate mutations and deletions for the V3 loop-deletion mutants of the envelope glycoproteins of the HXBc2 and ADA HIV-1 strains. The latter envelope glycoproteins were expressed by as ADA ΔV1/2gp160 glycoproteins. The V3 loop exchange mutants in the HXBc2, ADA, YU2, and MN envelope glycoproteins; the human CCR5 N terminus deletion mutants; and the chimeric CCR5 mutant containing the 412d CDRH3 loop were all made by the overlap-PCR approach. All introduced changes were confirmed by DNA sequencing.
Numbering of the gp120 amino acid residues is based on the sequence of the prototypic HXBc2 strain of HIV-1, according to current conventions (24).
Expression and analysis of CCR5 variants.
The wild-type CCR5 and CCR5 variants were expressed from plasmids containing a codon-optimized CCR5 gene and have a C-terminal C9 epitope tag. For analysis of sulfation, Cf2Th cells in a 100-mm dish were transfected using Effectene (QIAGEN). The cells were washed three times in PBS and then used for cysteine/methionine labeling or sulfate labeling. For cysteine/methionine labeling, cells were incubated overnight in cysteine/methionine-free Dulbecco's modified Eagle medium (DMEM) with 200 μCi (each) of 35S-labeled cysteine and 35S-labeled methionine (Protein Labeling Mix; Perkin-Elmer). For sulfate labeling, cells were incubated in sulfate-free medium (M8028; Sigma) with 500 μCi of 35S-sulfate (Na235SO4; Perkin-Elmer) for 1 day at 37°C. Cells were lysed in solubilization buffer, consisting of 1% CHAPSO {3-[(-cholamidopropyl) dimethylammonio]-2-hydroxyl-1-propanesulfonic acid}, 10% glycerol, 20 mM Tris, pH 7.5, 100 mM (NH4)2SO4, and protease inhibitors, and incubated at 4°C for 45 min with rocking. The cell lysates were cleared by centrifugation at 14,000 × g for 15 min at 4°C. The cleared lysates were incubated with the anti-C9 antibody 1D4 and protein A-Sepharose beads. The precipitated CCR5 proteins were visualized by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and autoradiography.
To produce target cells for HIV-1 infection, Cf2Th-CD4 cells were transfected with plasmids expressing wild-type or variant CCR5 proteins. An aliquot of the transfected cells was stained with phycoerythrin-conjugated anti-human CCR5 antibody, 2D7, and analyzed by fluorescence-activated cell sorter. The remaining cells were used for infection by recombinant HIV-1 expressing luciferase, as described below.
Transient expression of HIV-1 envelope glycoproteins.
293T cells grown to 70% confluence in 100-mm dishes were transfected with 4 μg of an envelope protein-expressing plasmid DNA and 1 μg of an HIV-1 Tat-expressing plasmid DNA with the Effectene transfection reagent (QIAGEN). One day later, the medium was removed, the cells were washed once with 10 ml of phosphate-buffered saline (PBS), and labeling medium (4.5 ml of DMEM; 0.5 ml of heat-inactivated, dialyzed FBS; 50 μg of penicillin-streptomycin solution) and 20 μl (∼230 μCi) of 35S-methionine protein labeling mixture were added. The cells were incubated at 37°C for another day. The medium was harvested, centrifuged to remove cells debris, and then stored at 4°C.
Immunoprecipitation of envelope glycoproteins.
For precipitation of radiolabeled HIV-1 envelope glycoproteins, 100 μl of medium (for soluble gp120 expression) or 400 μl of medium (for full-length gp160) containing the labeled proteins was incubated overnight with 100 μl of 10% protein A-Sepharose (Pharmacia), 50 μl of 4% bovine serum albumin, and 1 μg of monoclonal antibody with or without sCD4 (8 to 10 μg/ml) or, instead of the monoclonal antibody, 2 μl of a mixture of sera from HIV-1-infected individuals. PBS was added to bring the total volume to 1 ml. The Sepharose beads were then washed twice with 1 ml of 0.5 M NaCl in PBS and once with 1 ml of PBS. The beads were mixed with 2× gel loading buffer and boiled for 3 min. Following the removal of the beads by centrifugation, the supernatants were loaded on a 10% SDS-PAGE gel. The gel was enhanced with Autoflour (National Diagnostic) for 45 min before being dried at 80°C for 2 h and exposed to film. The gel was also used for PhosphorImager (Molecular Dynamics) analysis.
Virus preparation.
Recombinant HIV-1 containing the firefly luciferase gene was produced by transfection of 293T cells with the pCMV-Gag-Pol packaging plasmid, the pHIV-luc vector, and the pSVIIIenv plasmid expressing the wild-type or mutant envelope glycoproteins. Three days after transfection, the cell supernatants were harvested, assayed for virus by reverse transcriptase measurement, and frozen in aliquots.
Virus infection assay.
The Cf2Th cells expressing CD4 and either wild-type or mutant CCR5 were seeded at a density of 6,000 cells/well in a 96-well plate and cultured overnight at 37°C. About 2,000 reverse transcriptase units of virus were incubated with the cells overnight at 37°C in a 50-μl volume. Then, 200 μl of fresh medium was added to the cells, and the cells were cultured for 2 days. The cells were washed once with PBS and lysed with 30 μl of luciferase lysis buffer. The plates were stored in a −80°C freezer until the assays were performed. Luciferase assays were performed with the EG & G Berthold Microplate Luminometer LB 96V (Promega).
Neutralization assay.
The viral entry and neutralization assays were performed in 24-well plates for the wild-type HXBc2 and the HXBc2 NM/DI asparagine/methionine (NM) mutant, in which the asparagine/methionine (NM) pair was changed to aspartic acid/isoleucine (DI). Cf2Th-CD4/CCR5 cells were used to examine the use of CCR5 for entry by the HXBc2 NM/DI mutant. Cf2Th-CD4/CXCR4 cells were used to test the sensitivity of the viruses to neutralization by the 412d antibody. Viruses were incubated with 10 μg/ml 412d in 0.5 ml of DMEM for 60 min at 37°C. The virus-antibody mixtures were then transferred to wells containing the target Cf2Th-CD4/CCR5 cells. After overnight incubation, 1.5 ml of fresh medium was added to each well, and the cells were cultured for 2 more days. The cells were then lysed and assayed for luciferase activity (Tuner 20; Promega).
RESULTS
Mapping of the 412d epitope.
The CD4i monoclonal antibody 412d is derived from an HIV-1-infected individual, AC-01 (8, 37). As has been found for other CD4i antibodies (42-46), the 412d antibody blocked the ability of HIV-1 gp120-sCD4 complexes to bind the CCR5 receptor (Fig. 1).
FIG. 1.
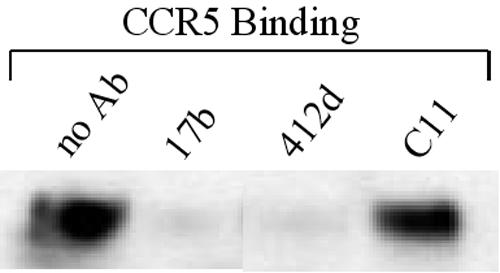
Inhibition of gp120 binding to CCR5 by the 412d antibody. Radiolabeled YU2 HIV-1 gp120 glycoprotein was incubated in the presence of sCD4 (10 μg/ml) with Cf2Th-CCR5 cells. The incubation was performed in the absence (no Ab) or presence of the 17b, 412d, or C11 antibody. The bound gp120 glycoprotein was detected by lysis of the Cf2Th-CCR5 cells and precipitation with sera pooled from HIV-1-infected individuals.
To characterize the gp120 epitope of 412d, a panel of 30 YU2 gp120 mutants was tested for 412d recognition, as previously described (44, 46). The amount of each mutant gp120 protein in the reaction was normalized by precipitating the samples in parallel with a mixture of polyclonal sera from HIV-1-infected individuals (data not shown). In the absence of sCD4, compared with recognition of wild-type gp120, recognition by the 412d antibody was significantly reduced for several gp120 mutants tested (e.g., 117K/D, 121K/D [C1 region], 207K/D, 257T/D [C2 region], 383F/S [C3 region], and 435Y/S [C4 region]). However, 412d binding to these mutants could be largely restored by adding sCD4. By contrast, even in the presence of sCD4, 412d did not efficiently precipitate the 420I/R mutant, which alters the β19 strand of gp120, or the 298R/G mutant, which alters the V3 loop (Fig. 2). Deletion of the entire V3 loop also abolished 412d binding, even in the presence of sCD4 (Fig. 2). The 381E/R mutant is disrupted for binding several conformation-dependent ligands (35, 44, 46), explaining its inefficient precipitation by the 412d antibody. In comparison with previously described epitope maps for CD4i antibodies such E51, 17b, and 23e, the gp120 residues important for 412d binding are located in an overlapping region of gp120. However, unlike most CD4i antibodies (44-46), the 412d antibody requires elements of the gp120 V3 loop for recognition. Thus, the 412d antibody epitope includes the β19 strand and V3 loop of HIV-1 gp120.
FIG. 2.

Precipitation of HIV-1 gp120 mutants by the 412 antibody. Radiolabeled wtΔ or mutant derivatives were incubated with the 412d antibody in the absence (−) or presence (+) of sCD4 (8 μg/ml). The precipitated proteins are shown. The indicated conserved region (C1-C4) or the V3 region represents the location of the altered amino acid residue on the primary HIV-1 gp120 sequence.
V3 loop sequences critical for 412d recognition.
The 412d antibody is unusual among CD4i antibodies in demonstrating a marked preference for the gp120 glycoproteins of CCR5-using HIV-1 isolates (8). As CCR5 use is dictated by sequences in the V3 loop (5, 22) and 412d recognition demonstrates a strong and unusual dependence on the integrity of the V3 loop, we wished to define better the V3 elements that contribute to 412d binding. The effect of changes in arginine 298 on 412d binding suggested the importance of the base of the V3 loop to 412d recognition. Therefore, a set of deletions extending from the V3 tip was created and analyzed (Fig. 3a). Figure 3b shows that the ΔV3-16 and ΔV3-18 glycoproteins were precipitated by the 412d antibody in both the absence and presence of sCD4. By contrast, the ΔV3 and ΔV3-15 glycoproteins were not efficiently recognized by the 412d antibody. We conclude that the 17 residues at the base of the V3 loop are important for 412d recognition of gp120.
FIG. 3.


Effect of gp120 V3 loop deletions on 412d recognition. (a) The sequences of the gp120 V3 loop of the X4 HIV-1 strain HXBc2, the R5X4 strain 89.6, and the R5 strains YU2 and ADA are aligned. The conserved sequences are boxed. The asterisks indicate the particular amino acid residues altered in this study. The extent of the deletions made in the HXBc2 and ADA gp120 glycoproteins is indicated by the horizontal arrows; the linkers substituted for the deleted residues are indicated in parentheses next to the name of the mutant. The ΔV3-12 and ΔV3-20 constructs were made for the HXBc2 gp120 glycoprotein only. The V3 deletion mutants in the ADA envelope glycoproteins were constructed in the context of the ΔV1/2 gp120 glycoprotein. The numbering scheme is based on that recommended by Korber et al. (24). (b) Precipitation of the ADA ΔV1/2 envelope glycoprotein and mutant derivatives by the 412d antibody in the absence and presence of 10 μg/ml sCD4 is shown. Precipitation of the proteins by pooled sera (P.S.) from HIV-1-infected individuals is also shown. The two bands represent glycoforms of the ΔV1/2 gp120 glycoprotein (data not shown).
The 412d antibody specifically precipitates the gp120 glycoproteins of the CCR5-using HIV-1 strains, YU2 and ADA, and the dualtropic (R5X4) strain 89.6, but not the gp120 from the CXCR4-using HXBc2 or MN strains (Fig. 4). A V3 loop-directed antibody, F425b, recognized the MN, ADA, YU2, and 89.6 gp120 glycoproteins but did not precipitate the HXBc2 gp120 glycoprotein (Fig. 4). To examine the gp120 determinants of this differential recognition, we utilized chimeric gp120 glycoproteins that contained the V3 loop from a heterologous strain. Some chimerae contained a heterologous V3 loop and an amino acid (residue 440) from the fourth conserved region of gp120. These changes in gp120 have been shown to alter chemokine receptor preference (4, 8). The HXBc2(YU2V3) glycoprotein, which contains the V3 loop of the YU2 R5 HIV-1 strain in the background of HXBc2 gp120, was efficiently recognized by the 412d and F425b antibodies. This result indicates that the V3 loop is the major determinant of the poor recognition of the HXBc2 gp120 glycoprotein by these antibodies. The V3 loop was also the determinant of the differential recognition of the MN and ADA gp120 glycoproteins by the 412d antibody (Fig. 4). Chimeric gp120 glycoproteins derived from the MN strain but containing the ADA V3 loop [MN(ADAV3) and MN(ADAV3 plus E440R)] were efficiently precipitated by the 412d antibody. By contrast, chimeric gp120 glycoproteins derived from the ADA strain but containing MN V3 loop [ADA(MNV3) and ADA(MNV3 plus R440E)] were not precipitated by the 412d antibody. The alteration of residue 440, which can influence the efficiency of CXCR4 usage (4, 19, 47), had no significant effect on 412d recognition. The MN-ADA chimeric gp120 glycoproteins were all precipitated by the F425b antibody, as expected from the efficient recognition of the MN and ADA parent glycoproteins. These results demonstrate that variation in V3 loop sequences accounts for the different recognition of gp120 from distinct HIV-1 strains by the 412d and F425b CD4i antibodies.
FIG. 4.

Effect of V3 loop exchange on gp120 recognition by the F425b and 412d antibodies. The gp120 glycoproteins of the X4 HXBc2 and MN HIV-1 strains, the R5X4 89.6 HIV-1 strain, and the R5 YU2 and ADA HIV-1 strains were precipitated by the indicated monoclonal antibody or by pooled sera (P.S.) from HIV-1-infected individuals. In addition, chimeric gp120 glycoproteins in which the gp120 V3 loop or V3 loop and residue 440 derived from a heterologous strain were tested. For example, the HXBc2(YU2V3) glycoprotein contains the V3 loop from the YU2 gp120 in the background of the HXBc2 gp120 glycoprotein.
We wished to further define the differences between the HXBc2 and YU2 V3 loop that accounted for the differences in 412d recognition. Because the base of the V3 loop is similar in these two gp120 glycoproteins, we hypothesized that sequences in the tip of the HXBc2 V3 loop might play an inhibitory role in 412d binding. To test this, progressive deletions of the tip of the V3 loop were made in the HXBc2 gp120 envelope glycoprotein. The ΔV3-20 and ΔV3-18 glycoproteins, which are missing the V3 tip (Fig. 3a), were recognized by the 412d antibody slightly better than the wild-type HXBc2 gp120 glycoprotein (Fig. 5). Additional deletion of V3 residues resulted in loss of 412d recognition (ΔV3-16 and ΔV3-12) (Fig. 5). Thus, sequences in the HXBc2 V3 tip may contribute to the poor recognition of the HXBc2 gp120 glycoprotein by the 412d antibody. However, even the ΔV3-18 mutant is recognized inefficiently by 412d compared with the HXBc2 (YU2V3) glycoprotein (Fig. 5). Therefore, other sequence differences between the HXBc2 and YU2 V3 loops apparently account for the differential recognition by the 412d antibody.
FIG. 5.

Precipitation of HXBc2 V3 loop deletion mutants by the 412d antibody. The indicated HXBc2 wild-type (WT) or mutant gp120 envelope glycoproteins were precipitated either by the 412d antibody in the presence of 10 μg/ml sCD4 or by pooled sera (P.S.) from HIV-1-infected individuals.
Comparison of the sequences in the base of the V3 loop of the HXBc2 and YU2 gp120 glycoproteins reveals differences in amino acid residues 325 and 326 (Fig. 3a). These residues are asparagine and methionine in the HXBc2 gp120 but are aspartic acid and isoleucine in the gp120 glycoproteins of viruses that can utilize CCR5 as a coreceptor. Alteration of the asparagine-methionine pair in the HXBc2 ΔV3-16 glycoprotein to aspartic acid-isoleucine resulted in efficient recognition by the 412d antibody (Fig. 6a). These changes were not sufficient to allow this envelope glycoprotein to bind two other antibodies, the CD4i F425b (6) antibody or the V3 loop-directed 39F antibody (Fig. 6a). The altered HXBc2 ΔV3-16 glycoprotein was not able to bind CCR5-expressing cells (Fig. 6a), consistent with the documented role of the V3 tip in chemokine receptor binding (2, 10, 35).
FIG. 6.

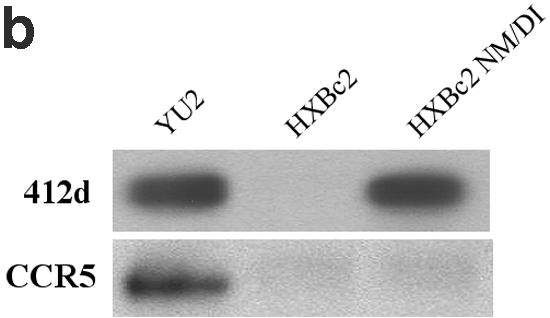


Effect of changes in residues 325 and 326 on 412d recognition. (a) The HXBc2 ΔV3-16 glycoproteins containing either asparagine/methionine (NM) or aspartic acid/isoleucine (DI) at V3 loop positions 325 and 326 were precipitated by the indicated monoclonal antibodies or by pooled sera (P.S.) from HIV-1-infected humans. All envelope glycoproteins were incubated with 10 μg/ml sCD4 prior to immunoprecipitation. The F425b and 39F antibodies are directed against the V3 loop. The ability of the envelope glycoproteins to bind CCR5-expressing cells is shown in the right panel (CCR5). (b) The precipitation of the indicated full-length gp120 glycoproteins by the 412d antibody in the presence of 10 μg/ml sCD4 is shown in the top panel. The ability of the envelope glycoproteins to bind Cf2Th-CCR5 cells after incubation with 10 μg/ml sCD4 is shown in the bottom panel. (c) Luciferase activity in Cf2Th-CD4/CCR5 cells incubated with recombinant luciferase-expressing HIV-1 viruses containing the indicated HIV-1 envelope glycoproteins is shown. (d) Recombinant luciferase-expressing viruses with the wild-type HXBc2 or HXBc2 NM/DI envelope glycoproteins were used to infect Cf2Th-CD4/CXCR4 cells. In some cases, viruses were first incubated with the 412d antibody. Luciferase activity in the target cells is shown. Data in panels c and d represent the means and standard deviation derived from two independent experiments.
We also examined the effect of changes in residues 325 and 326 in the context of the complete HXBc2 gp120 glycoprotein (Fig. 6b). The aspartic acid-isoleucine substitutions at these positions in the HXBc2 gp120 allowed 412d recognition but not CCR5 binding (HXBc2 NM/DI) (Fig. 6b). We conclude that the major determinants of the differential recognition by the 412d antibody of gp120 glycoproteins of the HXBc2 HIV-1 strain and CCR5-using strains are the residues at positions 325 and 326. CCR5 binding apparently has other requirements besides the changes in these residues. Consistent with this interpretation, when recombinant HIV-1 luciferase vectors were pseudotyped with the different envelope glycoproteins, neither the HXBc2 nor the HXBc2 NM/DI envelope glycoproteins was able to support entry into cells expressing CD4 and CCR5 (Fig. 6c). Both of these envelope glycoproteins mediated efficient entry into cells expressing CD4 and CXCR4 (Fig. 6d). Moreover, infection mediated by the HXBc2 NM/DI envelope glycoproteins, but not that mediated by the HXBc2 NM/DI envelope glycoproteins, was neutralized by the 412d antibody. These results demonstrate that the requirements for 412d antibody binding and CCR5 recognition can be separated in the context of a functional HIV-1 envelope glycoprotein.
Functional mimicry of the CCR5 N terminus by the 412 antibody.
Several lines of evidence indicate that the heavy-chain CDR3 loops of the CD4i antibodies play a major role in contacting gp120 (21, 25, 26). The similarities, including the presence of sulfated tyrosines, between the heavy-chain CDR3 of some CD4i antibodies (such as 412d) and the CCR5 N terminus prompted us to ask whether these structures might be functionally equivalent. The N terminus of CCR5 was replaced by sequences derived from the heavy-chain CDR3 loop of the 412d antibody (N412dR5) (Fig. 7a). To examine the contribution of tyrosine sulfation to the functional activity exhibited by this chimeric protein, the N412dR5-FF mutant was created (Fig. 7a). In this mutant, two of the tyrosine residues in the 412d heavy-chain CDR3 insert were converted to phenylalanine, preventing the addition of sulfate at these sites. We also examined the function of a CCR5 protein lacking the N terminus (ΔNR5) (Fig. 7a). The ΔNR5 protein was previously shown to be efficiently expressed on the cell surface but is significantly decreased in its ability to support the entry of R5 HIV-1 (17). Plasmids expressing the wild-type and mutant CCR5 proteins were transfected into Cf2Th canine thymocytes, which were radiolabeled with 35S-cysteine/methionine or 35S-sulfate. All of the CCR5 variants were expressed and, with the exception of the ΔNR5 mutant, incorporated labeled sulfate (Fig. 7b). The N412dR5-FF mutant was labeled with 35S-sulfate, suggesting that the two tyrosines remaining in the N terminus of this protein may be modified by sulfation.
FIG. 7.




Functional replacement of the CCR5 N terminus with the 412d heavy-chain CDR3 sequence. (a) The sequences of the CCR5 N terminus, the 412d heavy-chain CDR3, and the chimeric receptor constructs are shown. The ΔNR5 mutant has a deletion affecting the N terminus of CCR5. (b) Cf2Th cells expressing the indicated CCR5 variants were radiolabeled with 35S-cysteine/methionine (CYS/MET) or 35S-sulfate (SO4). The lysates were precipitated with the 1D4 antibody directed against the C9 C-terminal epitope tag on the CCR5 variants. Precipitates were analyzed by SDS-PAGE and autoradiography. (c) Cf2Th-CD4 cells expressing the indicated CCR5 variants were incubated with phycoerythrin-conjugated 2D7 antibody and analyzed by fluorescence-activated cell sorter. (d) The graph shows the luciferase activity in target Cf2Th-CD4 cells expressing the indicated CCR5 variants, following incubation with recombinant luciferase-expressing HIV-1 containing the indicated HIV-1 envelope glycoproteins. The values shown represent the means and standard deviation of four replicate experiments. Because of the high level of infectivity of the virus with the HXBc2(YUV3) glycoproteins, the values shown represent 1/10 of the actual luciferase activity observed.
The plasmids expressing the wild-type and mutant CCR5 proteins were transfected into Cf2Th cells expressing CD4, and the cell surface expression of the CCR5 variants and the susceptibility of these cells to recombinant luciferase-expressing viruses with various HIV-1 envelope glycoproteins were examined. All four CCR5 variants were expressed equivalently on the cell surface (Fig. 7c). The N412dR5 protein functioned as a receptor for viruses with the YU2, ADA, and JR-FL HIV-1 envelope glycoproteins as efficiently as wild-type CCR5 (Fig. 7d and data not shown). The N412dR5 protein was less efficient than wild-type CCR5 in supporting infection by dualtropic viruses from clade B (89.6), clade C (SA32, MCGP1.3, and 9279), and clade E (CF402) (Fig. 7d and data not shown). The integrity of the tyrosine residues in the substituted 412d CDR3 sequence contributes to the efficiency of the receptor activity (see N412dR5-FF in Fig. 7d). Neither wild-type CCR5 nor the N412dR5 protein supported infection of X4 viruses (HXBc2 and MN) (Fig. 7d). Utilization of wild-type CCR5 and N412dR5 was determined by V3 loop sequences [compare HXBc2 and HXBc2(YUV3) or MN and MN(ADAV3) in Fig. 7d]. We conclude that the 412d heavy-chain CDR3 loop sequence can functionally replace the CCR5 N terminus with respect to its activity and specificity as an HIV-1 receptor.
DISCUSSION
Binding of the HIV-1 gp120 glycoprotein to the CCR5 chemokine receptor is thought to involve two sets of contacts: (i) the interaction of the β19 strand and V3 loop of gp120 with the CCR5 N terminus and (ii) the binding of the tip of the V3 loop to the body of the chemokine receptor. These CCR5-interactive elements of gp120 are more conserved than the sequences along the flanks of the V3 loop, variations which can determine chemokine receptor preference (4, 22, 32, 40). Additional variation in V3 sequences is probably driven by pressure from neutralizing antibodies directed against both variable and more conserved gp120 structures. The CD4i antibodies presumably contribute to the selection of virus variants with minimal spontaneous exposure of the chemokine receptor-binding site on the envelope glycoprotein complex.
All CD4i antibodies bind gp120 better after CD4 binding and block the binding of gp120-sCD4 complexes to the chemokine receptor (41-43, 46). They do so by interacting with the first of the two chemokine receptor-binding surfaces on gp120 described above, i.e., the contacts for the N terminus of the chemokine receptor. The gp120 β19 strand and V3 loop base represent the most positively charged structures on the gp120 surface (25, 26) and thus complement the acidic N termini of the chemokine receptors. Contributing to this acidity are sulfated tyrosines in the N termini of both CCR5 and CXCR4. These sulfotyrosine residues are particularly important for HIV-1 entry via CCR5 (8, 11, 12, 15, 18). Some CD4i antibodies, like 412d, have evolved sulfated tyrosines in their long heavy-chain CDR3 loops that capitalize on the increased opportunities for molecular contacts with gp120 afforded by this posttranslational modification (8, 21). The 412d antibody is unusual among CD4i antibodies in exhibiting a marked preference for the gp120 glycoproteins of CCR5-using HIV-1 strains (8). Our results indicate that the amino acid residues at positions 325 and 326 in the base of the V3 loop are the major determinants of this 412d preference. Examination of the V3 loop sequences of numerous HIV-1 strains reveals that the aspartic acid-isoleucine pair, which is preferred by the 412d antibody, is much more common than the asparagine-methionine pair found in the HXBc2 gp120. As the 412d antibody was derived from an HIV-1-infected individual (8, 37), the viruses that elicited this antibody most likely had V3 sequences resembling the aspartic acid-isoleucine pair. Changes in residues 325 and 326 have been shown to decrease CCR5 binding (10), so the aspartic acid-isoleucine pair may also contribute to the interaction with the CCR5 N terminus. Our results, however, clearly indicate that CCR5 binding and utilization require specific V3 loop structures other than those specified by residues 325 and 326. Thus, the V3 loop requirements for CCR5 binding are more stringent than those for 412d recognition. For CCR5 N-terminal peptides and for 412d, deletions affecting only the tip of the V3 loop have little effect on binding; on the other hand, the V3 tip is required for binding the complete CCR5 protein (10, 12, 35). This is consistent with a model in which the 412d heavy-chain CDR3 loop mimics only the CCR5 N terminus and not the remainder of the CCR5 molecule, which contributes necessary contacts with the V3 loop tip.
The 412d heavy-chain CDR3 sequences effectively substitute for the natural CCR5 N terminus in supporting HIV-1 infection. This suggests that the two sets of gp120-CCR5 contacts described above are not significantly misaligned by this substitution. Dualtropic HIV-1 strains utilized the chimeric receptor somewhat less efficiently than wild-type CCR5, possibly related to their generally lower affinity for CCR5 than that exhibited by pure R5 viruses (9). The chimeric receptor exhibited the same preference for different HIV-1 strains as CCR5 itself. These results underscore the high degree to which the 412d antibody mimics the CCR5 N terminus. Although not fully understood, the structural requirements in the CCR5 N terminus for gp120 binding extend beyond the mere presence of sulfotyrosine and acidic residues. For example, the tyrosine-sulfated CXCR4 N terminus inefficiently substitutes for the CCR5 N terminus in supporting infection by R5 HIV-1 isolates (32). Studies in which individual CCR5 N-terminal residues were altered indicate that some specificity for the tyrosines at particular locations exists (15, 16).
Further attempts to define the precise interaction of the 412d antibody with gp120 are warranted. Our results suggest that such information may provide important insights into the interaction of gp120 and CCR5.
Acknowledgments
We thank Yvette McLaughlin and Sheri Farnum for manuscript preparation.
This work was supported by NIH grants (AI24755, AI31783, AI39420, and AI40895 to J.S. and AI24030 to J.R.), by a Center for AIDS Research grant (AI42848), by an unrestricted research grant from the Bristol-Myers Squibb Foundation, by a gift from the late William F. McCarty-Cooper, and by funds from the International AIDS Vaccine Initiative.
REFERENCES
- 1.Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger. 1996. CC CKR5: a RANTES, MIP-1α, MIP-1β receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272:1955-1958. [DOI] [PubMed] [Google Scholar]
- 2.Basmaciogullari, S., G. J. Babcock, D. Van Ryk, W. Wojtowicz, and J. Sodroski. 2002. Identification of conserved and variable structures in the human immunodeficiency virus gp120 glycoprotein of importance for CXCR4 binding. J. Virol. 76:10791-10800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bleul, C. C., M. Farzan, H. Choe, C. Parolin, I. Clark-Lewis, J. Sodroski, and T. A. Springer. 1996. The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry. Nature 382:829-833. [DOI] [PubMed] [Google Scholar]
- 4.Carrillo, A., and L. Ratner. 1996. Human immunodeficiency virus type 1 tropism for T-lymphoid cell lines: role of the V3 loop and C4 envelope determinants. J. Virol. 70:1301-1309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Carrillo, A., D. B. Trowbridge, P. Westervelt, and L. Ratner. 1993. Identification of HIV-1 determinants for T lymphoid cell line infection. Virology 197:817-824. [DOI] [PubMed] [Google Scholar]
- 6.Cavacini, L., M. Duval, L. Song, R. Sangster, S. H. Xiang, J. Sodroski, and M. Posner. 2003. Conformational changes in env oligomer induced by an antibody dependent on the V3 loop base. AIDS 17:685-689. [DOI] [PubMed] [Google Scholar]
- 7.Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, C. Gerard, and J. Sodroski. 1996. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell 85:1135-1148. [DOI] [PubMed] [Google Scholar]
- 8.Choe, H., W. Li, P. L. Wright, N. Vasilieva, M. Venturi, C. C. Huang, C. Grundner, T. Dorfman, M. B. Zwick, L. Wang, E. S. Rosenberg, P. D. Kwong, D. R. Burton, J. E. Robinson, J. G. Sodroski, and M. Farzan. 2003. Tyrosine sulfation of human antibodies contributes to recognition of the CCR5 binding region of HIV-1 gp120. Cell 114:161-170. [DOI] [PubMed] [Google Scholar]
- 9.Clapham, P. R., J. D. Reeves, G. Simmons, N. Dejucq, S. Hibbitts, and A. McKnight. 1999. HIV coreceptors, cell tropism and inhibition by chemokine receptor ligands. Mol. Membr. Biol. 16:49-55. [DOI] [PubMed] [Google Scholar]
- 10.Cormier, E. G., and T. Dragic. 2002. The crown and stem of the V3 loop play distinct roles in human immunodeficiency virus type 1 envelope glycoprotein interactions with the CCR5 coreceptor. J. Virol. 76:8953-8957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Cormier, E. G., M. Persuh, D. A. Thompson, S. W. Lin, T. P. Sakmar, W. C. Olson, and T. Dragic. 2000. Specific interaction of CCR5 amino-terminal domain peptides containing sulfotyrosines with HIV-1 envelope glycoprotein gp120. Proc. Natl. Acad. Sci. USA 97:5762-5767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cormier, E. G., D. N. Tran, L. Yukhayeva, W. C. Olson, and T. Dragic. 2001. Mapping the determinants of the CCR5 amino-terminal sulfopeptide interaction with soluble human immunodeficiency virus type 1 gp120-CD4 complexes. J. Virol. 75:5541-5549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. Di Marzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J. Schall, D. R. Littman, and N. R. Landau. 1996. Identification of a major co-receptor for primary isolates of HIV-1. Nature 381:661-666. [DOI] [PubMed] [Google Scholar]
- 14.Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Nagashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton. 1996. HIV-1 entry into CD4+ cells is mediated by the chemokine receptor CC- CKR-5. Nature 381:667-673. [DOI] [PubMed] [Google Scholar]
- 15.Dragic, T., A. Trkola, S. W. Lin, K. A. Nagashima, F. Kajumo, L. Zhao, W. C. Olson, L. Wu, C. R. Mackay, G. P. Allaway, T. P. Sakmar, J. P. Moore, and P. J. Maddon. 1998. Amino-terminal substitutions in the CCR5 coreceptor impair gp120 binding and human immunodeficiency virus type 1 entry. J. Virology. 72:279-285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Farzan, M., H. Choe, L. Vaca, K. Martin, Y. Sun, E. Desjardins, N. Ruffing, L. Wu, R. Wyatt, N. Gerard, C. Gerard, and J. Sodroski. 1998. A tyrosine-rich region in the N terminus of CCR5 is important for human immunodeficiency virus type 1 entry and mediates an association between gp120 and CCR5. J. Virol. 72:1160-1164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Farzan, M., S. Chung, W. Li, N. Vasilieva, P. L. Wright, C. E. Schnitzler, R. J. Marchione, C. Gerard, N. P. Gerard, J. Sodroski, and H. Choe. 2002. Tyrosine-sulfated peptides functionally reconstitute a CCR5 variant lacking a critical amino-terminal region. J. Biol. Chem. 277:40397-40402. [DOI] [PubMed] [Google Scholar]
- 18.Farzan, M., T. Mirzabekov, P. Kolchinsky, R. Wyatt, M. Cayabyab, N. P. Gerard, C. Gerard, J. Sodroski, and H. Choe. 1999. Tyrosine sulfation of the amino terminus of CCR5 facilitates HIV-1 entry. Cell 96:667-676. [DOI] [PubMed] [Google Scholar]
- 19.Farzan, M., N. Vasilieva, C. E. Schnitzler, S. Chung, J. Robinson, N. P. Gerard, C. Gerard, H. Choe, and J. Sodroski. 2000. A tyrosine-sulfated peptide based on the N terminus of CCR5 interacts with a CD4-enhanced epitope of the HIV-1 gp120 envelope glycoprotein and inhibits HIV-1 entry. J. Biol. Chem. 275:33516-33521. [DOI] [PubMed] [Google Scholar]
- 20.Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger. 1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science 272:872-877. [DOI] [PubMed] [Google Scholar]
- 21.Huang, C.-C., M. Venturi, S. Majeed, M. J. Moore, S. Phogat, M.-Y. Zhang, D. S. Dimitrov, W. A. Hendrickson, J. Robinson, J. Sodroski, R. Wyatt, H. Choe, M. Farzan, and P. D. Kwong. 2004. Structural basis of tyrosine sulfation and VH-gene usage in antibodies that recognize the HIV type 1 coreceptor-binding site on gp120. Proc. Natl. Acad. Sci. USA 101:2706-2711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hung, C. S., N. Vander Heyden, and L. Ratner. 1999. Analysis of the critical domain in the V3 loop of human immunodeficiency virus type 1 gp120 involved in CCR5 utilization. J. Virol. 73:8216-8226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hwang, S. S., T. J. Boyle, H. K. Lyerly, and B. R. Cullen. 1991. Identification of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science 253:71-74. [DOI] [PubMed] [Google Scholar]
- 24.Korber, B., F. Foley, C. Kuiken, S. Pillai, and J. Sodroski. 1998. Numbering positions in HIV relative to HXBc2, p. III-102-III-103. In B. Korber, C. Kuiken, B. Foley, B. Hahn, F. McCutchan, J. Mellors, and J. Sodroski (ed.), Human retroviruses and AIDS. Los Alamos National Laboratory, Los Alamos, N.Mex.
- 25.Kwong, P. D., R. Wyatt, S. Majeed, J. Robinson, R. W. Sweet, J. Sodroski, and W. A. Hendrickson. 2000. Structures of HIV-1 gp120 envelope glycoproteins from laboratory-adapted and primary isolates. Structure Fold Des. 8:1329-1339. [DOI] [PubMed] [Google Scholar]
- 26.Kwong, P. D., R. Wyatt, J. Robinson, R. W. Sweet, J. Sodroski, and W. A. Hendrickson. 1998. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 393:648-659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Labrijn, A. F., P. Poignard, A. Raja, M. B. Zwick, K. Delgado, M. Franti, J. Binley, V. Vivona, C. Grundner, C. C. Huang, M. Venturi, C. J. Petropoulos, T. Wrin, D. S. Dimitrov, J. Robinson, P. D. Kwong, R. T. Wyatt, J. Sodroski, and D. R. Burton. 2003. Access of antibody molecules to the conserved coreceptor binding site on glycoprotein gp120 is sterically restricted on primary human immunodeficiency virus type 1. J. Virol. 77:10557-10565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Meng, G., X. Wei, X. Wu, M. T. Sellers, J. M. Decker, Z. Moldoveanu, J. M. Orenstein, M. F. Graham, J. C. Kappes, J. Mestecky, G. M. Shaw, and P. D. Smith. 2002. Primary intestinal epithelial cells selectively transfer R5 HIV-1 to CCR5+ cells. Nat. Med. 8:150-156. [DOI] [PubMed] [Google Scholar]
- 29.Oberlin, E., A. Amara, F. Bachelerie, C. Bessia, J. L. Virelizier, F. Arenzana-Seisdedos, O. Schwartz, J. M. Heard, I. Clark-Lewis, D. F. Legler, M. Loetscher, M. Baggiolini, and B. Moser. 1996. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and prevents infection by T-cell-line-adapted HIV-1. Nature 382:833-835. [DOI] [PubMed] [Google Scholar]
- 30.O'Brien, W. A., Y. Koyanagi, A. Namazie, J. Q. Zhao, A. Diagne, K. Idler, J. A. Zack, and I. S. Chen. 1990. HIV-1 tropism for mononuclear phagocytes can be determined by regions of gp120 outside the CD4-binding domain. Nature 348:69-73. [DOI] [PubMed] [Google Scholar]
- 31.Philpott, S. M. 2003. HIV-1 coreceptor usage, transmission, and disease progression. Curr. HIV Res. 1:217-227. [DOI] [PubMed] [Google Scholar]
- 32.Polzer, S., M. T. Dittmar, H. Schmitz, and M. Schreiber. 2002. The N-linked glycan g15 within the V3 loop of the HIV-1 external glycoprotein gp120 affects coreceptor usage, cellular tropism, and neutralization. Virology 304:70-80. [DOI] [PubMed] [Google Scholar]
- 32a.Pontow, S., and L. Ratner. 2001. Evidence for common structural determinants of human immunodeficiency virus type 1 coreceptor activity provided through functional analysis of CCR5/CXCR4 chimeric coreceptors. J. Virol. 75:11503-11514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Pope, M., and A. T. Haase. 2003. Transmission, acute HIV-1 infection and the quest for strategies to prevent infection. Nat. Med. 9:847-852. [DOI] [PubMed] [Google Scholar]
- 34.Rizzuto, C., and J. Sodroski. 2000. Fine definition of a conserved CCR5-binding region on the human immunodeficiency virus type 1 glycoprotein 120. AIDS Res. Hum. Retrovir. 16:741-749. [DOI] [PubMed] [Google Scholar]
- 35.Rizzuto, C. D., R. Wyatt, N. Hernandez-Ramos, Y. Sun, P. D. Kwong, W. A. Hendrickson, and J. Sodroski. 1998. A conserved HIV gp120 glycoprotein structure involved in chemokine receptor binding. Science 280:1949-1953. [DOI] [PubMed] [Google Scholar]
- 36.Robinson, J. E., D. Holton, S. Pacheco-Morell, J. Liu, and H. McMurdo. 1990. Identification of conserved and variant epitopes of human immunodeficiency virus type 1 (HIV-1) gp120 by human monoclonal antibodies produced by EBV-transformed cell lines. AIDS Res. Hum. Retrovir. 6:567-579. [DOI] [PubMed] [Google Scholar]
- 37.Rosenberg, E. S., M. Altfeld, S. H. Poon, M. N. Phillips, B. M. Wilkes, R. L. Eldridge, G. K. Robbins, R. T. D'Aquila, P. J. Goulder, and B. D. Walker. 2000. Immune control of HIV-1 after early treatment of acute infection. Nature 407:523-526. [DOI] [PubMed] [Google Scholar]
- 38.Saphire, E. O., P. W. Parren, C. F. Barbas III, D. R. Burton, and I. A. Wilson. 2001. Crystallization and preliminary structure determination of an intact human immunoglobulin, b12: an antibody that broadly neutralizes primary isolates of HIV-1. Acta Crystallogr. D Biol. Crystallogr. 57:168-171. [DOI] [PubMed] [Google Scholar]
- 39.Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1991. Macrophage and T cell-line tropisms of HIV-1 are determined by specific regions of the envelope gp120 gene. Nature 349:167-169. [DOI] [PubMed] [Google Scholar]
- 40.Stamatatos, L., and C. Cheng-Mayer. 1995. Structural modulations of the envelope gp120 glycoprotein of human immunodeficiency virus type 1 upon oligomerization and differential V3 loop epitope exposure of isolates displaying distinct tropism upon virion-soluble receptor binding. J. Virol. 69:6191-6198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Thali, M., J. P. Moore, C. Furman, M. Charles, D. D. Ho, J. Robinson, and J. Sodroski. 1993. Characterization of conserved human immunodeficiency virus type 1 gp120 neutralization epitopes exposed upon gp120-CD4 binding. J. Virol. 67:3978-3988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Trkola, A., T. Dragic, J. Arthos, J. M. Binley, W. C. Olson, G. P. Allaway, C. Cheng-Mayer, J. Robinson, P. J. Maddon, and J. P. Moore. 1996. CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5. Nature 384:184-187. [DOI] [PubMed] [Google Scholar]
- 43.Wu, L., N. P. Gerard, R. Wyatt, H. Choe, C. Parolin, N. Ruffing, A. Borsetti, A. A. Cardoso, E. Desjardin, W. Newman, C. Gerard, and J. Sodroski. 1996. CD4-induced interaction of primary HIV-1 gp120 glycoproteins with the chemokine receptor CCR-5. Nature 384:179-183. [DOI] [PubMed] [Google Scholar]
- 44.Xiang, S. H., N. Doka, R. K. Choudhary, J. Sodroski, and J. E. Robinson. 2002. Characterization of CD4-induced epitopes on the HIV type 1 gp120 envelope glycoprotein recognized by neutralizing human monoclonal antibodies. AIDS Res. Hum. Retrovir. 18:1207-1217. [DOI] [PubMed] [Google Scholar]
- 45.Xiang, S. H., P. D. Kwong, R. Gupta, C. D. Rizzuto, D. J. Casper, R. Wyatt, L. Wang, W. A. Hendrickson, M. L. Doyle, and J. Sodroski. 2002. Mutagenic stabilization and/or disruption of a CD4-bound state reveals distinct conformations of the human immunodeficiency virus type 1 gp120 envelope glycoprotein. J. Virol. 76:9888-9899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Xiang, S. H., L. Wang, M. Abreu, C. C. Huang, P. D. Kwong, E. Rosenberg, J. E. Robinson, and J. Sodroski. 2003. Epitope mapping and characterization of a novel CD4-induced human monoclonal antibody capable of neutralizing primary HIV-1 strains. Virology 315:124-134. [DOI] [PubMed] [Google Scholar]
- 47.Yuan, W., S. Craig, Z. Si, M. Farzan, and J. Sodroski. 2004. CD4-induced T-20 binding to human immunodeficiency virus type 1 gp120 blocks interaction with the CXCR4 coreceptor. J. Virol. 78:5448-5457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zwick, M. B., P. W. Parren, E. O. Saphire, S. Church, M. Wang, J. K. Scott, P. E. Dawson, I. A. Wilson, and D. R. Burton. 2003. Molecular features of the broadly neutralizing immunoglobulin G1 b12 required for recognition of human immunodeficiency virus type 1 gp120. J. Virol. 77:5863-5876. [DOI] [PMC free article] [PubMed] [Google Scholar]