

Abstract
Aminergic signaling is known to play a critical role in regulating female reproductive processes in both mammals and insects. In Drosophila, the ortholog of noradrenaline, octopamine, is required for ovulation as well as several other female reproductive processes. Two octopamine receptors have already been shown to be expressed in the Drosophila reproductive tract and to be required for egg-laying: OAMB and Octβ2R. The Drosophila genome contains 4 additional octopamine receptors—Octα2R, Octβ1R, Octβ3R, and Oct-TyrR—but their cellular patterns of expression in the reproductive tract and potential contribution(s) to egg-laying are not known. In addition, the mechanisms by which OAMB and Octβ2R regulate reproduction are incompletely understood. Using a panel of MiMIC Gal4 lines, we show that Octα2R, Octβ1R, Octβ3R, and Oct-TyrR receptors are not detectable in either epithelium or muscle but are clearly expressed in neurons within the female fly reproductive tract. Optogenetic activation of neurons that express at least 3 types of octopamine receptors stimulates contractions in the lateral oviduct. We also find that octopamine stimulates calcium transients in the sperm storage organs and that its effects in spermathecal, secretory cells, can be blocked by knock-down of OAMB. These data extend our understanding of the pathways by which octopamine regulates egg-laying in Drosophila and raise the possibility that multiple octopamine receptor subtypes could play a role in this process.
Keywords: octopamine, octopamine receptor, egg-laying, spermatheca, oviposition
Introduction
The regulation of some reproductive processes is surprisingly conserved, allowing the use of relatively simple systems to explore the underlying mechanisms: these include the aminergic regulation of oocyte development and ovulation mediated by noradrenalin in mammals and its structural ortholog octopamine (OA) in Drosophila melanogaster (White et al. 2021b) (Kim et al. 2021) (Deady and Sun 2015) (Andreatta et al. 2018) (Meiselman et al. 2018) (Yoshinari et al. 2020). D. melanogaster expresses 6 OA receptors (El-Kholy et al. 2015) (Qi et al. 2017) (Evans and Maqueira 2005) (Balfanz et al. 2005) (Maqueira et al. 2005), and 2 of them, OAMB and Octβ2R, have been proposed to regulate egg-laying and female fertility (Lee et al. 2003) (Lee et al. 2009) (Lim et al. 2014) (Li et al. 2015). OAMB is expressed in a sheath of follicle cells that surround the egg and is required for OA-dependent rupture of the sheath (Deady and Sun 2015). Both Octβ2R and OAMB are expressed in epithelial cells that line the oviducts, and expression of OAMB at this site is required for fertility (Lee et al. 2003) (Lee et al. 2009) (Lim et al. 2014) (Deshpande et al. 2022). OAMB also regulates the function of the seminal receptacle and spermathecae (Avila et al. 2012). Both OAMB and Octβ2R are expressed in central and peripheral neurons that innervate the reproductive tract, and Octβ2R-expressing neurons in the reproductive tract may regulate oviduct contractions (Deshpande et al. 2022). Surprisingly, despite the proposed roles of both OAMB and Octβ2R in muscle contractility, neither is detectably expressed in muscle using the MiMIC GAL4 lines that we have used (Deshpande et al. 2022). In addition, while some of the signaling pathways responsible for the effects of OAMB and Octβ2R are known, others, including the mechanism by which OAMB regulates sperm storage, remain incompletely understood.
Drosophila express 4 other OA receptors in addition to OAMB and Octβ2R: Octα2R, Octβ1R, Octβ3R, and Oct-TyrR (El-Kholy et al. 2015) (Qi et al. 2017) (Evans and Maqueira 2005) (Balfanz et al. 2005) (Maqueira et al. 2005). At the cellular level, their patterns of expression in the reproductive tract are not known (El-Kholy et al. 2015) and it is not known whether they may play any role in reproduction. We show here that Octα2R, Octβ1R, Octβ3R, and Oct-TyrR are expressed in neurons and neuronal processes in the reproductive tract, but unlike OAMB and Octβ2R, they are not expressed in epithelial cells. To further probe the potential function of OA in the reproductive tract, we have tested the effects of optogenetically stimulating OA receptor–expressing cells and the effects of OA on the acute regulation of calcium transients in the sperm storage organs.
Materials and methods
Experimental model and subject details
D. melanogaster were raised in mixed sex vials on cornmeal/sucrose/yeast/sucrose/dextrose/agar medium at 25°C and 50–70% humidity under a 12:12 light:dark cycle unless otherwise noted. All experiments used mated or virgin female flies 4–6 days post eclosion. All fly lines used in this study are listed in Table 1.
Table 1.
Fly lines and antibodies used in this study.
| Reagent type | Symbol | Identifier | Source | Full genotype |
|---|---|---|---|---|
| Drosophila line | OAMB-T2A-Gal4 | #84675 | BDSC | y[1], w[*]; Mi{Trojan-GAL4.1}Oamb[MI12417-TG4.1] |
| Octα2R-T2A-Gal4 | #84610 | BDSC | y[1] w[*]; Mi{Trojan-GAL4.0}Octalpha2R[MI10227-TG4.0]/TM3, Sb[1] Ser[1] | |
| Octβ1R-T2A-Gal4 | #86139 | BDSC | TI{RFP[3xP3.cUa]=2A-GAL4}Octbeta1R[2A-C.GAL4.KI]/TM3, Sb[1] | |
| Octβ2R-T2A-Gal4 | #84678 | BDSC | w[*]; TI{2A-GAL4}Octbeta2R[2A-ACDE.GAL4] | |
| Octβ3R-T2A-Gal4 | #84680 | BDSC | w[*]; TI{2A-GAL4}Octbeta3R[2A-FG.GAL4]/TM3, Sb[1] | |
| Oct-TyrR-T2A-GAL4 | #86138 | BDSC | TI{RFP[3xP3.cUa]=2A-GAL4}Oct-TyrR[2A-GAL4.KI] | |
| UAS-mCD8::GFP | #32194 | BDSC | w[*]; P{y[+t7.7] w[+mC]=20XUAS-IVS-mCD8::GFP}attP2 | |
| Ppk1.0-LexA | N/A | Bing Ye | w ; Sp / CyO ; Ppk1.0-LexA / TM6 | |
| UAS-mCD8::GFP, LexAop-CD2::RFP | #67093 | BDSC | y[1] w[*]; P{y[+t7.7] w[+mC]=lexAop-rCD2::RFP-p10.UAS-mCD8::GFP-p10}su(Hw)attP5/CyO; TM3, Sb[1]/TM6B, Tb[1] | |
| UAS-ChR2-XXM | N/A | Robert Kittel | w[1118]; UAS-CHR2(XXM)::tdTomato / CyO | |
| 24B-Gal4 | #1767 | BDSC | w[*]; P{w[+mW.hs]=GawB}how[24B] | |
| 40B09-Gal4 | #41235 | BDSC Jianjun Sun |
w[1118]; P{y[+t7.7] w[+mC]=GMR40B09-GAL4}attP2 | |
| UAS-RCaMP1b | #63793 | BDSC | w[*]; PBac{y[+mDint2] w[+mC]=20XUAS-IVS-NES-jRCaMP1b-p10}VK00005 | |
| UAS-OAMB-RNAi | #31233 | BDSC | y[1] v[1]; P{y[+t7.7] v[+t1.8]=TRiP.JF01732}attP2 | |
| UAS-dicer2 | #24651 | BDSC | w[1118]; P{w[+mC]=UAS-Dcr-2.D}10 | |
| Antibodies/labeling reagents | Mouse anti-GFP | Cat# A11120 | Invitrogen | |
| AF488-conjugated goat-anti-mouse | Cat# A28175 | Invitrogen | ||
| AF555-conjugated phalloidin | Cat# A34055 | Invitrogen | ||
| Rabbit anti-dsRed | Cat# 632496 | Takara Bio | ||
| AF568-conjugated goat-anti-rabbit | Cat# A11036 | Invitrogen |
Fly husbandry and stocks
Publicly available fly lines with noted identifiers were identified using Flybase (Gramates et al. 2022) obtained from the Bloomingtom Drosophila Stock Center (BDSC) (listed in Table 1). We would like to thank Jianjun Sun (University of Connecticut) for information on the spermatheca expression of the 40B09-Gal4 allele. We also thank the following people for generously supplying the following additional lines: Bing Ye (University of Michigan) for ppk1.0-LexA (Gou et al. 2014), Soo Hong Min (Harvard), and Robert Kittel (University of Würzberg) for UAS-ChR2-XXM (Dawydow et al. 2014) (Scholz et al. 2017).
Immunofluorescent labeling
To visualize the expression patterns of the OA receptor genes, the reproductive systems from flies harboring UAS-mCD8-GFP and OA receptor MiMIC-T2A-Gal4s were dissected in phosphate buffered saline (PBS) at 25°C with the ventral nerve cord (VNC) and medial abdominal nerve (MAN) connection left intact. The tissue was fixed in 4% paraformaldehyde (PFA) for 15 min and washed in PBS containing 0.3% (vol/vol) Triton X-100 (PBT) for 30 min. Following fixation, preparations were washed for 1 h in blocking buffer containing 5% (vol/vol) normal goat serum (NGS) (Cat# G9023, Sigma-Aldrich) in PBT then incubated for 24–48 h at 4°C in the primary antibody mouse anti-GFP (1:500 in blocking buffer, Cat# A11120, Invitrogen). The sample was washed in PBT for 3 h at 25°C then incubated in the secondary antibody AF488-conjugated goat-anti-mouse (1:500 in blocking buffer, Cat# A28175, Invitrogen) at 4°C for 24 h. After washing in PBT for 1 h at 25°C, the preparations were optically cleared in 25% glycerol for 1 h at 25°C and then mounted on Superfrost slides (Cat# 12-550-143, Fisherbrand) with bridged glass cover slips #0 (Cat# 72198-10, Electron Microscopy Sciences) in Fluoromount-G mounting media (Cat# 0100-01, SouthernBiotech). For colabeling of muscle cells, preparations were dissected and processed as described above, except AF555-conjugated phalloidin (1:500, Ref# A34055, Invitrogen) was included in the secondary antibody solution. For colabeling of ppk1.0-LexA, preparations were dissected and processed as described above, except rabbit anti-dsRed (1:500, Cat# 632496, Takara Bio) was added to the primary antibody solution and AF568-conjugated goat-anti-rabbit (1:500, Cat# A11036, Invitrogen) was added to the secondary antibody solution.
Optogenetic stimulation of lateral oviduct contractions
For optogenetic experiments, mated female flies were raised in standard food containing 80 μM all-trans retinal from 1 day post eclosion until tested (5–7 days). To minimally disrupt nerves in and around the reproductive tract, optogenetic stimulations of OA receptor-expressing cells were performed on “abdominal fillet” preparations as previously described (Deshpande et al. 2022). In brief, the abdomen was separated from the rest of the fly body using microscissors and pinned to a Sylgard dish, and the sternal plates removed to expose the reproductive organs. Stimulation was performed using a Lambda DG-4 light source (Sutter), Chroma filter set 41001, and the light path of an Axio Examiner Z1 microscope to illuminate the entire field of view at 1 mW/mm2 power. The preparation was also illuminated from the side using an external LED to facilitate the visualization of contractions. Contractions of the LO were defined by a decrease in the distance between ovaries and a characteristic contraction of the oviduct tissue as described (Deshpande et al. 2022) and counted in video recordings acquired with a charge-coupled device (CCD) camera (Andor iXon 897, Oxford Instruments, Oxfordshire, England) at a capture rate of 12 frames/s and using Andor IQ2 software. Flies harboring 1 copy of Octβ1R, Octβ3R, or Oct-TyrR wMiMIC-T2A-Gal4 and 1 copy of UAS-ChR2-XXM::tdTomato were compared with controls that expressed UAS-ChR2-XXM::tdTomato alone.
Live imaging of sperm storage organs
To specifically determine OA's effects on the accessory gland cellular activity, without simultaneously activating peripheral circuits associated with the reproductive tract, we performed live imaging using a previously described “isolated preparation” (Deshpande et al. 2022). The reproductive tract was dissected out of the abdomen, and the abdominal cuticle, the gut, and fat bodies were removed. The MAN was cut and the CNS was also removed. The anterior tip of the ovaries and the distal end of the uterus were pinned to a Sylgard substrate with insect pins. Live imaging of RCaMP1b expressed in the seminal receptacle and spermathecae was performed using a 555 nm LED light source (Thorlabs), the standard Chroma filter set 41007a, and a Zeiss Achroplan water immersion 10× objective on a Zeiss Axio Examiner Z1 microscope with a CCD camera (Andor iXon 897, Oxford Instruments, Oxfordshire, England) at a capture rate of 12 frames/s using Andor IQ2 software. Images were analyzed using Fiji/ImageJ software (Schindelin et al. 2012). For all regions of interest (ROIs), an off-target area of equal size was selected as background. In all experiments, 1 min of baseline activity was recorded before OA was bath applied to the preparation at the indicated concentration. Changes in fluorescence are reported as the background-subtracted difference in the change in fluorescence divided by baseline {ΔF/F = [(F peak − F baseline)/F baseline], where F baseline = average RCaMP signal during the 1 min before OA bath application}. Recording was conducted for 4 min following OA addition. All flies tested were 4 days post eclosion and carried either 1 copy of 24b-Gal4 or 40B09-Gal4 and 1 copy of UAS-RCaMP1b. RNAi experiments used flies that additionally included 1 copy each of UAS-dicer2 and UAS-OAMB-RNAi. Mated flies were cohoused with CS males after sorting 1 day post eclosion. Virgin flies were sorted and housed without males.
Statistical analysis
The optogenetic data shown in Fig. 8 were analyzed using Dunnett's test in the base R package. The data in Fig. 8g–h were analyzed using 2-way ANOVA in GraphPad Prism.
Fig. 8.

OA induces calcium transients in sperm storage organs. UAS-RCaMP1b was expressed in either muscle cells in the seminal receptable with 24B-Gal4 (a–c) or in spermathecal secretory cells with 40B09-Gal4 (d–i) followed by application of OA or vehicle. The red ovals represent examples of ROIs used for quantitation. Sample images (a) of seminal receptacle muscle cells pre- and postaddition (maximal response) of 1 μM OA at 60 s and a trace showing the time course of the response (b). A dose response plot shows seminal receptacle muscle cells increase intracellular calcium in response to OA doses ranging from 0.01 to 1000 μM (c). Sample images (d) of spermathecal secretory cells pre- and postaddition of 1 μM OA at 60 s and the time course of the response (e). Spermathecal secretory cells increase intracellular calcium in response to OA doses ranging from 0.5 to 1000 μM (f). Knocking down OAMB in the spermathecal secretory cells with UAS-OAMB-RNAi (indicated as “RNAi: +”) reduces the maximal RCaMP response to OA in the spermathecal secretory cells of mated flies (g), the time to maximal ΔF/F response in both virgin and mated flies (h), and the baseline RCaMP signal in virgin flies (i, arbitrary units × 103) relative to matched controls (controls express 40B09-Gal4, UAS- RCaMP1b, and UAS-dicer2 but not UAS-OAMB-RNAi. Mean+/− SEM shown; 1-way ANOVA with multiple comparison test across all conditions within each panel, *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001, ****P ≤ 0.0001. Scale bars = 50 μm.
Results
All OA receptors are expressed in the reproductive tract
Using a panel of MiMIC-T2A-Gal4 lines, each 1 of the 6 Drosophila OA receptors has previously been shown to be expressed in the fly central nervous system (McKinney et al. 2020). We previously used these lines to investigate the expression and function of Octβ2R and OAMB in the reproductive tract, focusing on the oviducts (Deshpande et al. 2022). We have now extended the analysis of Octβ2R and OAMB to other regions of the reproductive tract and used additional MiMIC lines to determine the expression patterns of Octα2R, Octβ1R, Octβ3R, and Oct-TyrR (Fig. 1a–i). We observe fine processes at the base of the ovaries that express Octβ2R (Fig. 1d) but not OAMB (Fig. 1e). Octβ3R- and Oct/TyrR-expressing processes also extend into the ovary (Fig. 1f and g), but we do not detect expression of OctβR- or Octα2R-expressing processes in this region (Fig. 1h and i).
Fig. 1.

OA receptor expression in the ovaries. a) A schematized dorsal view of the female Drosophila reproductive tract showing a mature egg (white arrowhead), the ovaries (Ov), lateral oviducts (LO), common oviduct (CO), and the uterus (Ut). Two sperm storage organs are attached to the uterus: the seminal receptacle (SR, indicated in gray because it is on the ventral side of the Ut) and the 2 spermathecae (Sp). An additional pair of parovarian/accessory glands (Pa) is attached to the uterus posterior to the spermatheca. b) A dissected preparation viewed from the dorsal side showing the relative sizes and positions of the Ov, Sp, and Ut. c) A schematize sagittal view of the reproductive tract as it appears in vivo. d–i) MiMIC Gal4 lines for the indicated receptors were used to express membrane-attached GFP (green) and colabeled for phalloidin (magenta) as a marker for muscle cells. The boxed areas highlight the presence or absence of fine processes that ascend into the ovaries. Faint labeling of follicle cells within the boxed area can be seen with OAMB (e). Labeling of the oviduct epithelium is also visible for Octβ2 and OAMB (black arrowheads). Scale bars: 100 μm.
For Octβ2, some labeled processes in the reproductive tract represent autoreceptors expressed in Tdc2(+) neurons as seen in colabeling experiments for Tdc2 and Octβ2R (Deshpande et al. 2022). In contrast to Octβ2R, we do not detect coexpression of Octβ3R or Oct-TyrR in Tdc2(+) neurons (data not shown). Therefore, the Octβ3R and Oct-TyrR processes that we detect in the ovaries and elsewhere in the reproductive tract represent other types of neurons that remain to be identified.
Consistent with previous reports (Lee et al. 2003) (Deady and Sun 2015), we detect OAMB in follicle cells that surround the mature oocyte (Fig. 1e, boxed area). In addition, we detect a swath of labeling at the base of the ovaries; this may represent the insect analog of the corpus luteum that consists of follicle cells that have been shed from the activated oocyte (data not shown) (Deady and Sun 2015). We do not detect expression of any other OA receptors in the follicle cells (Fig. 1 and data not shown).
Both OAMB and Octβ2 are expressed in the epithelial cells that line the lumen of the oviducts (Lee et al. 2003) (Deshpande et al. 2022), and genetic rescue studies have indicated that both may function in the epithelium to promote egg-laying (Lim et al. 2014). In contrast to OAMB and Octβ2, we do not detect expression of Octα2R, Octβ1R, Octβ3R, or Oct-TyrR in the epithelial cells that line the oviducts (Fig. 2). For comparison, an example of epithelial cells in the LO expressing OAMB is shown in Fig. 2e. Similar to both Octβ2R and OAMB, all other OA receptors including Octα2R, Octβ1R, Octβ3R, and Oct-TyrR can be detected in fine processes within both the LO and CO (Fig. 2a–d). Processes that express all 6 OA receptors are also present in the uterus, while the SR is innervated only by OAMB-, Octα2R-, and Oct/TyrR-expressing cells (Fig. 3). A summary of these and all other expression data are shown in Table 2.
Fig. 2.
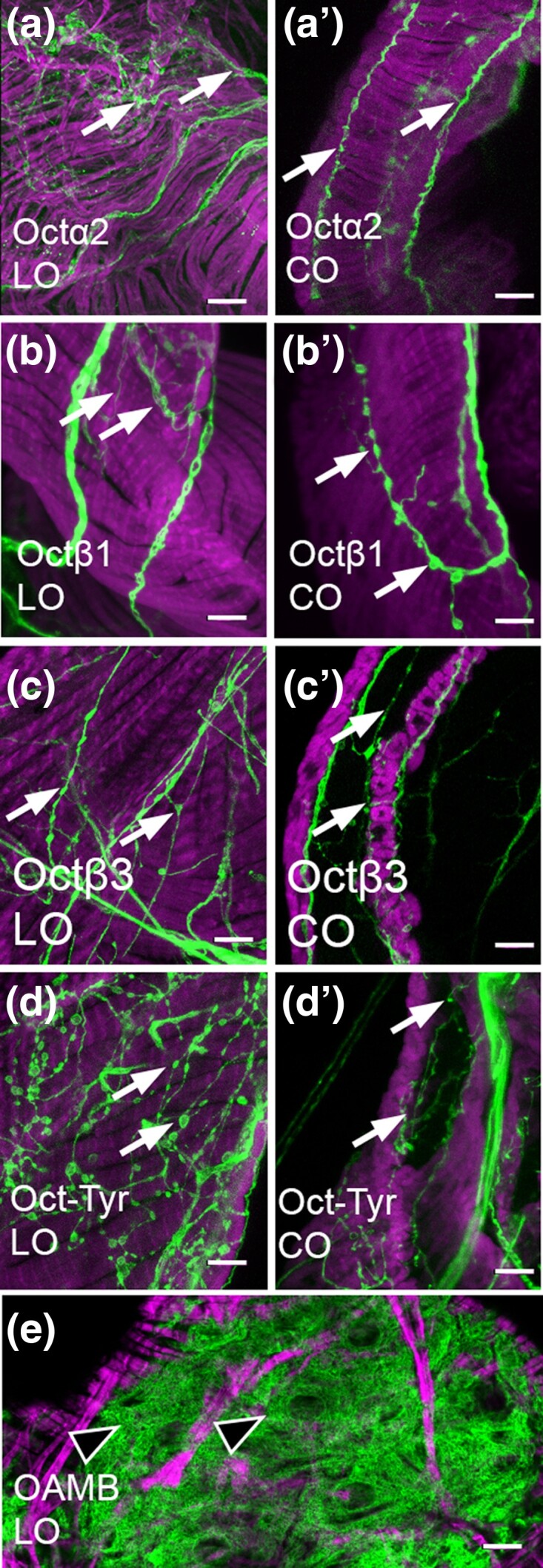
Processes in the oviducts. a–e). MiMIC Gal4 lines for the indicated receptors were used to express membrane-attached GFP (green) and colabeled for phalloidin (magenta) as in Fig. 1. Fine processes (white arrows) are present for Octα2R, Octβ1R, Octβ3R, and Oct-TyrR as previously reported for Octβ2R and OAMB. Labeling in muscle or epithelium was not detected. An example of epithelial cells expressing OAMB is shown (e, black arrowheads). Scale bars: 10 μm.
Fig. 3.

Processes in the uterus and seminal receptacle. a–f) MiMIC Gal4 lines for the indicated receptors were used to express membrane-attached GFP (green) and colabeled for phalloidin (magenta) as in Figs. 1 and 2. Processes in the uterus (a–f) are visible for all receptors. Processes in the seminal receptable (a′–f′) are visible for OAMB, Octα2R, and Oct-TyrR but not Octβ1R, Octβ2R, or Octβ3R. Scale bars: 10 μm.
Table 2.
Data summary.
| OAMB | Octα2R | Octβ1R | Octβ2R | Octβ3R | Oct-TyrR | ||
|---|---|---|---|---|---|---|---|
| CNS neurons | + | + | + | + | + | + | |
| Ovarian follicle cells | + | − | − | − | − | − | |
| Epithelium | LO | + | − | − | + | − | − |
| CO | + | − | − | − | − | − | |
| Presumptive secretory cells | Spermatheca | + | − | − | − | − | − |
| Parovarian gland | + | − | − | − | − | − | |
| Innervation | Ovaries | − | − | − | + | + | + |
| LO | + | + | + | + | + | + | |
| CO | + | + | + | + | + | + | |
| Ut | + | + | + | + | + | + | |
| SR | + | + | − | − | − | + | |
| Peripheral neurons | Anterior Ut | + | + | − | + | + | − |
| Posterior Ut | + | − | + | − | − | − | |
| “Free-hanging” | − | + | + | − | + | + | |
| ChR2-induced oviduct contractions | − | ND | + | + | +/− | + | |
The expression of the receptor GAL4 MiMIC lines at each site and the response of the oviducts to optogenetic (ChR2-induced) stimulation are indicated.
ND, not determined.
Peripheral cell bodies express OA receptors
Some processes from Octβ2R and OAMB are likely to be derived from neurons in the VNC (McKinney et al. 2020), and labeled processes for all 6 OA receptors can be detected in the medal abdominal nerve (MAN) that connects the VNC to the reproductive tract (data not shown). We have previously reported that a small number of peripheral neurons intrinsic to the reproductive tract also express Octβ2R and OAMB, some of which localize to the uterus (Deshpande et al. 2022). Similarly, we detect a small cluster of cell bodies in the anterior uterus that expresses Octα2R and Octβ3R (Fig. 4a and b). We do not detect Octβ1R(+) cells in the anterior uterus (Fig. 1c), but at least 1 Octβ1R(+) cell localizes to the posterior uterus (Fig. 4e). For comparison, we show subsets of cells that express Octβ2R in the anterior uterus (Fig. 4f) and OAMB in the posterior uterus (Fig. 4g) as previously reported (Deshpande et al. 2022). OA receptor–expressing cell bodies are also present within the nerves that descend from the AbG and that connect the anterior uterus to the base of the ovaries (indicated as free-hanging in Table 2 and Fig. 4). These include cells present within bilaterally symmetric nerves near the oviducts that express Octα2R, Oct-TyrR, Octβ1R, and Octβ3R (Fig. 4a–d).
Fig. 4.

Peripheral cell bodies in the reproductive tract. a–g) MiMIC Gal4 lines for the indicated receptors were used to express GFP (green) and colabeled for phalloidin (magenta) as in Figs. 1–3. Cell bodies in the anterior uterus (Ant Ut) express Octα2R (a) and Octβ3R (b), and at least 1 cell body expressing Octβ1R is present in the posterior uterus (e). All other cell bodies shown here that express Octα2R, Octβ3R, Octβ1R, and Oct-TyrR are contained within or attached to free-hanging nerves that connect the MAN to the reproductive tract or connect the anterior and posterior regions of the reproductive tract (a–d). Previously reported cells expressing Octβ2R in the anterior uterus (f) and OAMB in the posterior uterus are shown for comparison (g). Boxed insets show the indicated cells at a higher magnification. Scale bars: (a–e) 100 μm; (f and g and boxed insets in a–d) 10 μm.
We have previously shown that both OAMB and Octβ2R are coexpressed in neurons that express ppk1.0-LexA (Deshpande et al. 2022), a marker for mechanosensitive neurons that regulate CO contractions and postmating behavior (Gou et al. 2014) (Hasemeyer et al. 2009). We find that subsets of cells in the anterior uterus that express Octα2R or Octβ3R coexpress ppk1.0-LexA (Fig. 5a and c). In addition, at least 1 Octβ1(+) cell in the posterior uterus expresses ppk1.0-LexA (Fig. 5b). The coexpression of Octβ2R and OAMB with ppk1.0-LexA is shown for comparison (Fig. 5d and e).
Fig. 5.

A subset of peripheral cell bodies expressing OA receptors colabel with the marker ppk1.0-LexA. a–e) MiMIC Gal4 lines for the indicated receptors were used to express GFP (green) and colabeled with ppk1.0-LexA expressing RFP (magenta). The boxed regions indicated in a–e are shown at a higher magnification in a′–e′ and a″−e″. Cells labeled for the receptor alone (white arrowhead), ppk1.0 alone (black arrowhead), or both the receptor and ppk1.0 (double white + black arrowheads) are indicated. Scale bars: (a–e) 100 μm; (a′–e′, a″−e″) 10 μm.
OAMB is expressed in the sperm storage organs and accessory glands
OA has been shown to regulate sperm storage, and RNA knock-down experiments have implicated OAMB in this process (Avila et al. 2012). We observe expression of the OAMB-Gal4 MiMIC in the seminal receptacle, the spermatheca, and the parovarian glands, consistent with a previous report using a custom Ab and additional tissue-specific Gal4 lines (Lee et al. 2003) (Fig. 6). We do not detect any additional OA receptors in the bulb of the spermathecae or the “cap” of the parovarian glands (data not shown). The location of the OAMB(+) cells in the bulb of the spermathecae is consistent with that of previously described secretory cells (Schnakenberg et al. 2011) (Allen and Spradling 2008). The identity of the cells that express OAMB in the lumen of the spermathecae and seminal receptacle is not clear; however, their location and shape suggest the possibility that they are a component of an epithelial like-layer, perhaps similar to that which lines the oviducts.
Fig. 6.

OAMB is expressed in the seminal receptacle and spermathecae. a–c) OAMB(+) cells were marked with GFP and colabeled with phalloidin (magenta). Cells in the seminal receptacle (a–a″, SR, white arrows), the spermathecae (b–b′′′, Sp), and the parovarion glands (c–c″, Pa) express OAMB. In the spermathecae (b–b′′′), cells were labeled in both the bulb (white arrowheads) and the lumen of the stalk (black arrowheads). Scale bars: (a–c) 50 μm; (a–a″, b–b′′′, and c′) 10 μm.
Optogenetic stimulation of OA receptor-expressing cells drives lateral oviduct muscle contractions
We have previously shown that optogenetic stimulation of cells that express Octβ2R induces lateral oviduct contractions (Deshpande et al. 2022). To determine whether other OA receptor–expressing cells might also promote oviduct contractility, we expressed the channelrhodopsin variant ChR2-XXM (Dawydow et al. 2014) (Scholz et al. 2017) using other OA receptor MiMIC Gal4 lines. In addition to Octβ2R(+) cells, we find that optogenetic activation of Octβ1R(+) and Oct-TyrR(+) cells induce lateral oviduct contractions that are statistically different from the control. Stimulation of 2 of 5 preparations using the Octβ3R MiMIC driver was followed by lateral oviduct contractions (Fig. 7), but this did not statistically differ from the control. One presumably spontaneous contraction was seen during one of the stimulation periods for each of the controls that expressed Octβ1R-MiMIC-Gal4, Oct3R-MiMIC-Gal4, or UAS-ChR2-XXM alone (Fig. 7).
Fig. 7.

Optogenetic stimulation of OA receptor–expressing cells drives lateral oviduct contractions. a) Fly abdomens were severed from the throrax, dissected to expose the reproductive organs, and optogenetically stimulated (b). The number of LO contractions in response to each of 3 successive stimulations per fly was quantified. The number of flies per genotype is indicated in parentheses (n). The data were analyzed using Dunnett's test to compare multiple conditions to the negative control UAS-ChRXXM without a Gal4 driver. The control differed significantly (***P < 0.001) from flies expressing UAS-ChRXXM with Octβ1R-Gal4 or Oct-TyrR-Gal4 but not Octβ3R-Gal4 or flies expressing a Gal4 driver in the absence of UAS-ChRXXM.
Although the oviduct epithelium expresses both Octβ2R and OAMB, we have previously shown that optogenetic stimulation of epithelial cells has no detectable effects on oviduct muscle contractions (Deshpande et al. 2022). In addition, we do not detect expression of any OA receptors in the muscles of the reproductive tract (Figs. 1–4 and Deshpande et al. (2022)). We therefore conclude that cells expressing Octβ1R and Oct-TyrR that respond to optogenetic stimulation are likely to be neurons, although it is not possible to rule out the possibility that receptor expression undetectable by the MiMIC lines but expressed at low levels in muscle could contribute to this response.
Bath applied OA elicits calcium changes in sperm storage organs
Bath applied OA induces muscle contractions and calcium transients within the muscle cells of both the ovaries and the lateral oviducts in Drosophila (Middleton et al. 2006) (Deshpande et al. 2022) (Meiselman et al. 2018). Muscle cells labeled with the 24B-Gal4 driver (Martinez-Azorin et al. 2013) also surround the lumen of the seminal receptacle. To determine if OA could stimulate the activity of muscles in the seminal receptacle, we used 24B-Gal4 to express the calcium sensor UAS-RCaMP1b (Fig. 8a–c). We observed frequent spontaneous calcium transients at baseline in the absence of OA (Supplementary Video 1) and an increase in calcium within seconds of applying of OA (Fig. 8a and b and Supplementary Video 1). The calcium transients appeared as waves, both at baseline and in the presence of OA (Supplementary Video 1). Dose response experiments indicate that the seminal receptacle muscle cells show a maximal response to OA concentrations as low as 0.01 μM (10 nM).
OAMB regulates calcium transients in secretory cells within the spermatheca
The secretory cells in the spermathecae have been suggested to be regulated by OAMB (Avila et al. 2012) and previously shown to be involved in sperm storage (Schnakenberg et al. 2011) (Allen and Spradling 2008). As shown above (Fig. 6), we find that OAMB is expressed in the spermathecal bulb at a site consistent with previously described secretory cells (Schnakenberg et al. 2011) (Allen and Spradling 2008). To determine the acute effects of OA on the presumptive secretory cells of the spermathecae, we expressed UAS-RCaMP1b using the driver 40B09-Gal4 (Fig. 8d) and bath applied OA (or vehicle) to preparations of the reproductive tract that had been dissected out of the abdomen and both the CNS and all other surrounding tissue removed as described (Deshpande et al. 2022) (“isolated preparation,” see Materials and methods). We observed a robust increase in cytosolic calcium (Fig. 8d) that returned to a value near baseline within ∼80 s (Fig. 8e). Dose response experiments show that the secretory cells responded to OA at concentrations as low as 0.1–0.5 μM (100–500 nM; Fig. 8f), although the response was variable across preparations.
To directly assess the contribution of OAMB to calcium transients in spermathecal cells, we used a previously tested RNAi transgene directed against OAMB mRNA (Perkins et al. 2015). We tested both virgin and mated flies since a number of physiological changes occur after mating (White et al. 2021a) (Findlay et al. 2014) (Chapman et al. 2003). We again used the 40B09-Gal4 driver to express UAS-RCaMP1b and coexpressed UAS-OAMB-RNAi and UAS-dicer2 to knock down OAMB. The controls for these experiments used 40B09-Gal4 to express UAS-RCaMP1b and UAS-dicer2 in the absence of UAS-OAMB-RNAi. OAMB knock-down significantly blunted the maximum amplitude of the RCaMP signal seen in response to OA in the spermathecae of mated flies compared with controls; this effect was not statistically significant for virgin flies (Fig. 8e). Both mated and virgin flies showed a decrease in the duration of the calcium signal seen in response to OA (Fig. 8f, shown as the time from the onset of the increase until the maximum). In virgin, but not mated flies, the baseline RCaMP signal also appeared to be lower when OAMB was knocked down (Fig. 8i); however, we acknowledge the difficulty in assessing baseline fluorescence across different preparations. In sum, our data indicate that OAMB regulates the calcium response to OA of spermatheca secretory cells, consistent with the previously proposed role for OAMB in regulating sperm storage and suggesting that OAMB calcium-mediated pathways may be responsible for its effects on sperm storage (Avila et al. 2012) (Chen et al. 2022).
Discussion
To gain insight into the mechanisms by which OA and cells that express OA receptors may regulate egg-laying, we have used a panel of high-fidelity Gal4 “MiMIC” lines to map expression in the reproductive tract of all the known Drosophila OA receptors. We also show that a subset of OA receptor–expressing cells may influence oviduct muscle contractions and that OAMB regulates the acute response to OA of a sperm storage organ.
We have previously shown that OAMB and Octβ2R are expressed in neuronal processes and peripheral neurons within the reproductive tract (Deshpande et al. 2022). Here, we show that multiple cells that localize to the reproductive tract also express Octα2R, Octβ1R, Octβ3R, and Oct-TyrR. Coexpression with the neuronal marker ppk1.0-LexA indicates that the cells that express Octα2R, Octβ1R, and Octβ3R in the reproductive tract are neurons. While we have not demonstrated colocalization of Oct-TyrR(+) with ppk1.0, the morphology of Oct-TyrR(+) cells and their processes within the reproductive tract strongly suggest that they, too, are neurons.
Potential differences between the functions of neurons that express each OA receptor subtype are highlighted by differences in the degree to which they appear to innervate each organ within the reproductive tract. For example, Octβ1R, Octβ3R, and Oct-TyrR innervate the base of the ovaries, suggesting the possibility that they could influence egg maturation. OAMB, Octα2R, and Oct-TyrR(+) processes innervate the SR, suggesting that both Octα2R and Oct-TyrR might play a role in sperm storage in addition to the established role for OAMB (Avila et al. 2012). All 6 OA receptors innervate the lateral and common oviducts, but the morphology of boutons at each site show subtle differences, and this too may influence their function. The morphological differences between glutamatergic, octopaminergic, and peptidergic boutons at the larval NMJ are well characterized (Jia et al. 1993) (Keshishian et al. 1996). The relationship between bouton morphology and neurochemical identity at other sites in the fly has received relatively little attention.
Nonneuronal cells that express OAMB in the reproductive tract include follicle cells that surround the developing oocyte and epithelial cells that line the lumen of the oviducts (Deady and Sun 2015) (Lee et al. 2003). We have confirmed that cells within the SR also express OAMB (Lee et al. 2003) and show that Ca2+ levels in the muscle of the SR are sensitive to OA. It possible that OAMB(+) cells within the lumen of the SR contribute to the response of the surrounding muscle, similar to the mechanism previously proposed for epithelial cells in the oviducts (Lee et al. 2009) (Lim et al. 2014). However, it is also possible that neuronal processes that express OAMB are responsible for this effect and further experiments will be needed to distinguish between these possibilities.
Previous results indicate that loss of Octβ2R blocks contraction of the lateral oviducts and optogenetic activation of Octβ2R-expressing neurons can induce lateral oviduct contractions (Deshpande et al. 2022). We find that optogenetic activation of Octβ1R- and Oct-TyrR-expressing cells can also induce lateral oviduct contractions. Since mutation of Octβ2R essentially blocks contractions caused by bath applied OA (Deshpande et al. 2022), the possibility that multiple, equally important, parallel pathways within the reproductive tract mediate oviduct contraction seems unlikely. Rather, we hypothesize that signaling pathways upstream of Octβ2R are responsible for these effects seen when activating Octβ1R(+) or Oct-TyrR(+) cells.
Previous studies have demonstrated a requirement for OA in the regulation of sperm storage by the spermathecae (Avila et al. 2012) (Chen et al. 2022), but the more acute effects of octopaminergic signaling at this site have been less clear. We show that OA induces calcium transients in secretory cells of the spermathecae and that this effect is blocked by knock-down of OAMB within these cells. These data suggest that calcium-mediated pathways may be responsible for the effects of OAMB on sperm storage. Since a secretory cell driver was used to knock-down OAMB, these effects would appear to occur via a cell autonomous mechanism within the secretory cells themselves rather than another cell type.
Most previous studies on reproduction in the fly have focused on the function of OA receptors in nonneuronal tissues, such as the epithelium and follicle cells (Li et al. 2015) (Lee et al. 2009) (Lim et al. 2014) (Deady and Sun 2015) (White et al. 2021b). Similarly, as noted above, we believe that nonneuronal cells regulate calcium transients in the secretory cells of the spermathecae and the oviduct epithelium as well as follicle cell rupture. Conversely, for the optogenetic stimulation of oviduct contraction, we suggest that effects we observe result from the activation of neurons or neuronal processes that express OA receptors. Both OAMB and Octβ2R are expressed in epithelial cells in the reproductive tract, and their expression in the epithelium is required for fertility (Lee et al. 2009) (Lim et al. 2014). However, expression of ChR2-XXM in epithelial cells does not induce oviduct contractions (Deshpande et al. 2022). In addition, we do not detect any of the 6 OA receptors in muscle cells using the GAL4 drivers (Deshpande et al. 2022) and we do not detect Octα2R, Octβ1R, Octβ3R, or Oct-TyrR in the epithelium, follicle cells, secretory cells, or any other identified nonneuronal cell types in the reproductive system using the MiMIC Gal4 lines. We therefore conclude that at least some of the effects we have observed when activating or inhibiting OA receptor–expressing cells are likely to be neuronal in origin. We anticipate that further experiments targeting subsets of OA expressing neurons will allow us to more determine the identity of these cells and thus increase our understanding of the complex neuronal network that regulates egg-laying.
Supplementary Material
Acknowledgments
We thank the labs of Dr. Lawrence Zipursky and Dr. Kelsey Martin at UCLA for graciously sharing their confocal microscopes. We thank Hugo J. Bellen, Pei-Tseng Lee and other members of the Bellen lab for generating the MiMIC lines and the GAL4 conversions, especially Yuchun He, Wen-Wen Lin, and Drs. Sonal Nagarkar-Jaiswal and Oguz Kanca as part of their efforts to generate “A comprehensive resource for manipulating the Drosophila genome with swappable insertion cassettes,” funded by NIH R24 OD031447, to Hugo J. Bellen and Oguz Kanca. We thank Jianjun Sun (University of Connecticut) for his work screening relevant Gal4 expression patterns. We thank the following people for generously supplying fly lines: Dr. Kyung-An Han (University of Texas, El Paso), Dr. Bing Ye (University of Michigan), and Dr. Robert Kittel (University of Würzberg).
Contributor Information
Ethan W Rohrbach, Interdepartmental Program in Neuroscience, Brain Research Institute, Gonda (Goldschmied) Neuroscience and Genetics Research Center, UCLA, Los Angeles, CA 90095, USA.
Elizabeth M Knapp, Department of Psychiatry and Biobehavioral Sciences, David Geffen School of Medicine, UCLA, Los Angeles, CA 90095, USA.
Sonali A Deshpande, Department of Psychiatry and Biobehavioral Sciences, David Geffen School of Medicine, UCLA, Los Angeles, CA 90095, USA.
David E Krantz, Department of Psychiatry and Biobehavioral Sciences, David Geffen School of Medicine, UCLA, Los Angeles, CA 90095, USA.
Data availability
Fly strains are available upon request and/or from the Bloomington Stock Center as indicated. The authors affirm that all data necessary for confirming the conclusions of the article are present within the article, figures, and tables. Full data sets are available on request.
Supplemental material available at G3 online.
Funding
This work was supported by the National Institutes of Health including grants R01MH107390 and R01MH114017 (D.E.K.), and the training grants T32DA024635 (E.W.R.) and F32NS123014 (E.M.K.).
Literature cited
- Allen AK, Spradling AC. 2008. The Sf1-related nuclear hormone receptor Hr39 regulates Drosophila female reproductive tract development and function. Development. 135(2):311–321. doi: 10.1242/dev.015156. [DOI] [PubMed] [Google Scholar]
- Andreatta G, Kyriacou CP, Flatt T, Costa R. 2018. Aminergic signaling controls ovarian dormancy in Drosophila. Sci Rep. 8(1):2030. doi: 10.1038/s41598-018-20407-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Avila FW, Bloch Qazi MC, Rubinstein CD, Wolfner MF. 2012. A requirement for the neuromodulators octopamine and tyramine in Drosophila melanogaster female sperm storage. Proc Natl Acad Sci U S A. 109(12):4562–4567. doi: 10.1073/pnas.1117689109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balfanz S, Strunker T, Frings S, Baumann A. 2005. A family of octopamine [corrected] receptors that specifically induce cyclic AMP production or Ca2+ release in Drosophila melanogaster. J Neurochem. 93(2):440–451. doi: 10.1111/j.1471-4159.2005.03034.x. [DOI] [PubMed] [Google Scholar]
- Chapman T, Bangham J, Vinti G, Seifried B, Lung O, Wolfner MF, Smith HK, Partridge L. 2003. The sex peptide of Drosophila melanogaster: female post-mating responses analyzed by using RNA interference. Proc Natl Acad Sci U S A. 100(17):9923–9928. doi: 10.1073/pnas.1631635100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen DS, Clark AG, Wolfner MF. 2022. Octopaminergic/tyraminergic Tdc2 neurons regulate biased sperm usage in female Drosophila melanogaster. Genetics. 221(4):iyac096. doi: 10.1093/genetics/iyac096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawydow A, Gueta R, Ljaschenko D, Ullrich S, Hermann M, Ehmann N, Gao S, Fiala A, Langenhan T, Nagel G, et al. 2014. Channelrhodopsin-2-XXL, a powerful optogenetic tool for low-light applications. Proc Natl Acad Sci U S A. 111(38):13972–13977. doi: 10.1073/pnas.1408269111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deady LD, Sun J. 2015. A follicle rupture assay reveals an essential role for follicular adrenergic signaling in Drosophila ovulation. PLoS Genet. 11(10):e1005604. doi: 10.1371/journal.pgen.1005604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deshpande SA, Rohrbach EW, Asuncion JD, Harrigan J, Eamani A, Schlingmann EH, Suto DJ, Lee P-T, Schweizer FE, Bellen HJ, et al. 2022. Regulation of Drosophila oviduct muscle contractility by octopamine. iScience. 25(8):104697. doi: 10.1016/j.isci.2022.104697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Kholy S, Stephano F, Li Y, Bhandari A, Fink C, Roeder T. 2015. Expression analysis of octopamine and tyramine receptors in Drosophila. Cell Tissue Res. 361(3):669–684. doi: 10.1007/s00441-015-2137-4. [DOI] [PubMed] [Google Scholar]
- Evans PD, Maqueira B. 2005. Insect octopamine receptors: a new classification scheme based on studies of cloned Drosophila G-protein coupled receptors. Invert Neurosci. 5(3–4):111–118. doi: 10.1007/s10158-005-0001-z. [DOI] [PubMed] [Google Scholar]
- Findlay GD, Sitnik JL, Wang W, Aquadro CF, Clark NL, Wolfner MF. 2014. Evolutionary rate covariation identifies new members of a protein network required for Drosophila melanogaster female post-mating responses. PLoS Genet. 10(1):e1004108. doi: 10.1371/journal.pgen.1004108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gou B, Liu Y, Guntur AR, Stern U, Yang CH. 2014. Mechanosensitive neurons on the internal reproductive tract contribute to egg-laying-induced acetic acid attraction in Drosophila. Cell Rep. 9(2):522–530. doi: 10.1016/j.celrep.2014.09.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gramates LS, Agapite J, Attrill H, Calvi BR, Crosby MA, dos Santos G, Goodman JL, Goutte-Gattat D, Jenkins VK, Kaufman T, et al. 2022. FlyBase: a guided tour of highlighted features. Genetics. 220(4):iyac035. doi: 10.1093/genetics/iyac035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasemeyer M, Yapici N, Heberlein U, Dickson BJ. 2009. Sensory neurons in the Drosophila genital tract regulate female reproductive behavior. Neuron. 61(4):511–518. doi: 10.1016/j.neuron.2009.01.009. [DOI] [PubMed] [Google Scholar]
- Jia XX, Gorczyca M, Budnik V. 1993. Ultrastructure of neuromuscular junctions in Drosophila: comparison of wild type and mutants with increased excitability. J Neurobiol. 24(8):1025–1044. doi: 10.1002/neu.480240804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keshishian H, Broadie K, Chiba A, Bate M. 1996. The Drosophila neuromuscular junction: a model system for studying synaptic development and function. Annu Rev Neurosci. 19(1):545–575. doi: 10.1146/annurev.ne.19.030196.002553. [DOI] [PubMed] [Google Scholar]
- Kim J, Hyun M, Hibi M, You YJ. 2021. Maintenance of quiescent oocytes by noradrenergic signals. Nat Commun. 12(1):6925. doi: 10.1038/s41467-021-26945-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee HG, Rohila S, Han KA. 2009. The octopamine receptor OAMB mediates ovulation via Ca2+/calmodulin-dependent protein kinase II in the Drosophila oviduct epithelium. PLoS One. 4(3):e4716. doi: 10.1371/journal.pone.0004716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee HG, Seong CS, Kim YC, Davis RL, Han KA. 2003. Octopamine receptor OAMB is required for ovulation in Drosophila melanogaster. Dev Biol. 264(1):179–190. doi: 10.1016/j.ydbio.2003.07.018. [DOI] [PubMed] [Google Scholar]
- Li Y, Fink C, El-Kholy S, Roeder T. 2015. The octopamine receptor octβ2R is essential for ovulation and fertilization in the fruit fly Drosophila melanogaster. Arch Insect Biochem Physiol. 88(3):168–178. doi: 10.1002/arch.21211. [DOI] [PubMed] [Google Scholar]
- Lim J, Sabandal PR, Fernandez A, Sabandal JM, Lee HG, Evans P, Han K-A. 2014. The octopamine receptor octβ2R regulates ovulation in Drosophila melanogaster. PLoS One. 9(8):e104441. doi: 10.1371/journal.pone.0104441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maqueira B, Chatwin H, Evans PD. 2005. Identification and characterization of a novel family of Drosophila beta-adrenergic-like octopamine G-protein coupled receptors. J Neurochem. 94(2):547–560. doi: 10.1111/j.1471-4159.2005.03251.x. [DOI] [PubMed] [Google Scholar]
- Martinez-Azorin F, Calleja M, Hernandez-Sierra R, Farr CL, Kaguni LS, Garesse R. 2013. Muscle-specific overexpression of the catalytic subunit of DNA polymerase gamma induces pupal lethality in Drosophila melanogaster. Arch Insect Biochem Physiol. 83(3):127–137. doi: 10.1002/arch.21101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKinney HM, Sherer LM, Williams JL, Certel SJ, Stowers RS. 2020. Characterization of Drosophila octopamine receptor neuronal expression using MiMIC-converted Gal4 lines. J Comp Neurol. 528(13):2174–2194. doi: 10.1002/cne.24883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meiselman MR, Kingan TG, Adams ME. 2018. Stress-induced reproductive arrest in Drosophila occurs through ETH deficiency-mediated suppression of oogenesis and ovulation. BMC Biol. 16(1):18. doi: 10.1186/s12915-018-0484-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Middleton CA, Nongthomba U, Parry K, Sweeney ST, Sparrow JC, Elliott CJH. 2006. Neuromuscular organization and aminergic modulation of contractions in the Drosophila ovary. BMC Biol. 4(1):17. doi: 10.1186/1741-7007-4-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkins LA, Holderbaum L, Tao R, Hu Y, Sopko R, McCall K, Yang-Zhou D, Flockhart I, Binari R, Shim H-S, et al. 2015. The transgenic RNAi project at Harvard Medical School: resources and validation. Genetics. 201(3):843–852. doi: 10.1534/genetics.115.180208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi YX, Xu G, Gu GX, Mao F, Ye GY, Liu W, Huang J. 2017. A new Drosophila octopamine receptor responds to serotonin. Insect Biochem Mol Biol. 90:61–70. doi: 10.1016/j.ibmb.2017.09.010. [DOI] [PubMed] [Google Scholar]
- Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, et al. 2012. Fiji: an open-source platform for biological-image analysis. Nat Methods. 9(7):676–682. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnakenberg SL, Matias WR, Siegal ML. 2011. Sperm-storage defects and live birth in Drosophila females lacking spermathecal secretory cells. PLoS Biol. 9(11):e1001192. doi: 10.1371/journal.pbio.1001192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholz N, Guan C, Nieberler M, Grotemeyer A, Maiellaro I, Gao S, Beck S, Pawlak M, Sauer M, Asan E, et al. 2017. Mechano-dependent signaling by latrophilin/CIRL quenches cAMP in proprioceptive neurons. eLife. 6:e28360. doi: 10.7554/eLife.28360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White MA, Bonfini A, Wolfner MF, Buchon N. 2021a. Drosophila melanogaster sex peptide regulates mated female midgut morphology and physiology. Proc Natl Acad Sci U S A. 118(1):e2018112118. doi: 10.1073/pnas.2018112118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White MA, Chen DS, Wolfner MF. 2021b. She’s got nerve: roles of octopamine in insect female reproduction. J Neurogenet. 35(3):132–153. doi: 10.1080/01677063.2020.1868457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshinari Y, Ameku T, Kondo S, Tanimoto H, Kuraishi T, Shimada-Niwa Y, Niwa R. 2020. Neuronal octopamine signaling regulates mating-induced germline stem cell increase in female Drosophila melanogaster. eLife. 9:e57101. doi: 10.7554/eLife.57101. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Fly strains are available upon request and/or from the Bloomington Stock Center as indicated. The authors affirm that all data necessary for confirming the conclusions of the article are present within the article, figures, and tables. Full data sets are available on request.
Supplemental material available at G3 online.