

Summary
The initial step in estrogen-regulated transcription is the binding of a ligand to its cognate receptors, named estrogen receptors (ERα and ERβ). Phytochemicals present in foods and environment can compete with endogenous hormones to alter physiological responses. We screened 224 flavonoids in our engineered biosensor ERα and ERβ PRL-array cell lines to characterize their activity on several steps of the estrogen signaling pathway. We identified 83 and 96 flavonoids that can activate ERα or ERβ, respectively. While most act on both receptors, many appear to be subtype-selective, including potent flavonoids that activate ER at sub-micromolar concentrations. We employed an orthogonal assay using a transgenic zebrafish in vivo model that validated the estrogenic potential of these compounds. To our knowledge, this is the largest study thus far on flavonoids and the ER pathway, facilitating the identification of a new set of potential endocrine disruptors acting on both ERα and ERβ.
Subject areas: Biological sciences, Endocrinology, endocrine regulation, Phytochemistry
Graphical abstract
Highlights
-
•
Screen of 224 flavonoid compounds for estrogenic activity
-
•
Discovery of many sub-micromolar estrogenic flavonoids
-
•
Estrogenic flavonoids show receptor subtype selectivity and specificity
-
•
Estrogenic flavonoids can increase growth of breast cancer cells and activate zebrafish EREs
Biological sciences; Endocrinology; Endocrine regulation; Phytochemistry
Introduction
Estrogen receptor is a member of the nuclear receptor (NR) superfamily of transcription factors that regulate gene expression in response to a variety of stimuli. Upon hormone binding, ER interacts with its DNA response elements, recruits coregulators, and either increases (with agonists like 17β-estradiol, E2) or decreases (with antagonists like 4-hydroxytamoxifen, 4HT; or ICI182,780, ICI) transcription of target genes. Estrogen receptors include two subtypes, estrogen receptor-α (ERα) and estrogen receptor-β (ERβ). Estrogen signaling plays a vital role in multiple tissues such as the bone,1 mammary gland,2 cardiovascular,3 and endometrium.4 Overactivation or disruption of this pathway can lead to diseases such as osteoporosis5 and breast cancer,6,7 highlighting the importance of estrogen receptor research. Modulations to the pathway can come in the form of endocrine disrupting compounds (EDCs) acting on estrogen receptors at inappropriate times.8 Previous studies by the ToxCast initiative using a combination of 18 high-throughput assays for ER activity found 111 out of 1,812 chemicals as active and, by using a set of 45 chemicals (“EPA45 library”), the authors were able to predict in vivo activity using only in vitro assays.9 Furthermore, since the early 2000s, studies have shown the ability of several plant chemicals, termed phytoestrogens, to activate human estrogen receptors. Clearly, phytoestrogens and many industrial chemicals such as bisphenol A (BPA), DDT, and genistein have been shown to bind ER, alter normal ER physiology, and also increase the proliferation of breast cancer cells.10
Within the large family of phytoestrogens, flavonoids are a subfamily of polyphenolic plant compounds, some of which are consumed in human diets (i.e., soy, tea, wine, fruits, and vegetables11). Flavonoids generally have a 15-carbon skeleton composed of two phenyl and one heterocyclic ring. The compounds can be found in many different plant sources including berries, tea, chocolate, and spices.12 In plants, flavonoids produce the pigment for flower coloration and are involved in UV filtration and nitrogen fixation. In humans, flavonoids can affect multiple physiological pathways including angiogenesis,13 bone regeneration,13,14,15 and the inflammatory pathway.16,17 Flavonoids have also been investigated as therapeutic agents for diseases such as atherosclerosis,18 Alzheimer’s disease,19,20 depression,21,22 and several types of cancer,23,24,25,26,27 as they have potential anti-oxidant and anti-inflammatory properties.
Previous studies have investigated the ability of flavonoids to activate ER, finding mixed results in physiology and disease. One of the most studied flavonoids, the soy phytoestrogen genistein,28 has been shown to bind to both ERs and activate classical signaling cascades,29,30 with a preference for ERβ. Studies suggest that genistein can be neuroprotective in Alzheimer’s disease without activating cancer pathways that are common from E2 treatment.31 Meta analysis has shown that isoflavonoid compounds, a subfamily of flavonoids, exert a beneficial effect against estrogen-deficient bone loss.32 On the other hand, overuse of licorice root, a plant with many flavonoids,33 can lead to irregular menstrual cycles in women and reproductive problems in men,34 both of which can be due to increased estrogen levels.35,36 While many studies exist that examine flavonoid actions through ER binding, these studies are not typically robust, only focusing on a single or small set of compounds including genistein, daidzein,37 apigenin,38 naringenin,39 equol,40 and quercetin.41 Interestingly, studies have also shown differences in binding affinity for ERα and ERβ for flavonoid compounds such as corylifol A (ERα-selective)42 and 9-prenylapigenin (ERβ-selective).43 Given our routine use of highly multiplexed high-content analysis assays for receptors (ERα,44 AR,45 PR46), we sought to investigate a wider range of flavonoids and determine their effects on both estrogen receptor subtypes.
In this study, we utilized the previously described and highly validated multiparametric PRL-HeLa array biosensor cell line backbone44 to test 224 flavonoids for their estrogenic activity on both ERs. We found that 37% of flavonoids in the library (83/224) caused ERα DNA binding at concentrations of 10μM or lower, while 43% (96/224) induced ERβ DNA binding. When comparing chemical structures, the highest enrichment for hits was in the isoflavone and chalcone groups, with over 50% of the compounds inducing ER DNA binding as visualized by high throughput imaging at the engineered, readily visible ERE-rich locus. We further analyzed two compounds, isobavachin and phenoxodiol, that activate ERα at low nanomolar concentrations (EC50 of 1.28nM and 13.97nM, respectively); their impact was further demonstrated on ER signaling and growth of MCF-7 breast cancer cells. We also identified three potent compounds showing ERα/β agonist activity. Finally, we investigate ER-subtype selective compounds in our PRL-array cell culture models and in an orthogonal assay involving a transgenic zebrafish model47 of ER activity. Our mechanistic HT screening with ER biosensor models and follow-on results in an orthogonal animal model support the notion that continued inquiry of flavonoid action on both estrogen receptors holds promise to better understand their environmental impact in the food supply, and potentially identify new therapeutic moieties.
Results
Profiling 224 flavonoid compounds for ER activity via single cell high content analysis
In an effort to add high throughput assays for EDCs characterization, we have previously profiled the EPA45 library,9 a reference set of 45 ER agonists, antagonists, and inactive chemicals, using the PRL-array ERα models,48 and in MCF7 cells measuring ERα levels and activity.44,49 Within the EPA45 set there are five flavonoid compounds, four of which modulated ERα. Our data, along with numerous reports that show that flavonoids can act as phytoestrogens, inspired us to test a larger number of compounds within this chemical class for estrogenic activity using our sensitive, high throughput microscopy-based imaging assay. We acquired a library of 224 flavonoid compounds from Selleckchem to test in our PRL-array cells expressing either ERα or ERβ.50 This library is comprised of a wide range of flavonoid structures, including flavones, isoflavones, chalcones, and some glycosylated versions, as well as a few non-standard flavone compounds (Figure S1A illustrates examples of flavonoids structures in comparison with the ER agonist 17β-estradiol, E2). We used NR-ToxPred (a support vector machine-based web server51) to predict the activity of the flavonoids in the library on ER and other nuclear receptors (Figure S1B), with ERα having the highest predicted activity with the compound set, followed closely by progesterone receptor (PR) and ERβ.
We tested the library in both ERα- and ERβ-expressing biosensor cells lines ERα-GFP:PRL-HeLa and GFP-ERβ:PRL-HeLa. These cell lines contain a tandem array of ER binding sites taken from the prolactin gene controlling a dsRed2 reporter gene (Figure S2A) stably multimerized ∼100X into the genome of HeLa cells. This stable line, combined with tet-inducible ERα-GFP (Figure S2B) or GFP-ERβ (Figure S2B), allows for visualization and quantitation of estrogenic activity of compounds including agonists (E2) and antagonists (4HT). While the fluorescent dsRed2 reporter protein is functional, it takes 24–48 h to visualize accumulation; as a result, we utilize smRNA FISH (small molecule fluorescence in situ hybridization) of the dsRed2 mRNA as a transcriptional readout on the same timescale as array formation (30-120m). The screening focused on identifying flavonoids that were able to induce GFP-ER signal over the PRL-array locus (representing ER binding to the engineered ERE-rich genomic locus), which is low in DMSO control treatment, and then rapidly (∼minutes) increases following E2 (Figures 1A and 1E for ERα-GFP:PRL-HeLa and GFP-ERβ:PRL-HeLa, respectively), and treatment with other known ER ligands. For ERα-GFP:PRL-HeLa, negative and positive controls (DMSO and 1nM E2) showed significant separation in the fraction of cells with a visible array within the assay plates, with 21% of DMSO-treated cells and 70% of E2 cells having arrays, respectively (Figure 1B). We tested the library at an initial concentration of 10μM for 1 h and identified 85 compounds (using a Z score>3 cutoff, the list of hits is provided in Table S1) that induced ERα-GFP DNA binding (green box, Figure 1C). Intriguingly, 35 of the 85 hits (43.2%) have no published literature suggesting they are potential ERα ligands (based on reviews,52 PubChem, and PubMed searches; Table S1). Within the primary screen, we simultaneously measured transcriptional output using an RNA FISH probe to the dsRed2 reporter gene as reported previously.53 Only one compound (P276-00) that induced ER-loading onto the PRL array was shown to suppress gene transcription, whereas all 84 other hits increased gene transcription when compared to DMSO. 82 of the flavonoids that increased FISH signals were between 60 and 120% of E2 induction (green dot, Figure 1D).
Figure 1.

Screen of 224 flavonoid compounds in PRL-ERα and PRL-ERβ cell lines
(A) Representative images of ERα-GFP and dsRed2 FISH under DMSO and 1nM E2 treatment for 1 h.
(B) Fraction of cells with a visible array in DMSO and E2 1nM control wells in ERα-GFP:PRL-HeLa screening plates.
(C) Fraction of cells showing a visible array after treatment for 1h with 224 flavonoids at 10μM in ERα-GFP:PRL-HeLa cells. Green box represents 3 Z scores above DMSO.
(D) For the compounds considered a hit in ERα-GFP:PRL-HeLa, the dsRed2 FISH intensity is calculated relative to E2. The red dot is 10nM 4HT, the yellow dot is 1nM E2.
(E) Representative images of GFP-ERβ and dsRed2 FISH following 1h DMSO or 1nM E2 treatment.
(F) Fraction of GFPL-ERβ:PRL-HeLa cells with a visible array in DMSO and E2 1nM control wells from screening plates.
(G) Fraction of cells showing a visible array after treatment for 1h with 224 flavonoids at 10μM in GFPL-ERβ:PRL-HeLa. Green box represents 3 Z score above DMSO.
(H) For the compounds considered a hit in GFPL-ERβ:PRL-HeLa, the dsRed2 FISH intensity is represented relative to E2. The red dot is 10nM 4HT, the yellow dot is 1nM E2. ∗ p value <0.05. Errors bars represent standard deviation.
For GFP-ERβ:PRL-HeLa, again, control treatments (DMSO and 1nM E2) showed significant separation in the fraction of cells with a visible array within the assay plates, with 25% of DMSO-treated cells having array and 75% of E2 cells having arrays (Figure 1F). The GFP-ERβ:PRL-HeLa has been known (Abbott et al., submitted) to have a higher ligand independent DNA binding than the ERα counterpart. Using the same 3 Z score cutoff, we found that 96/244 compounds increased array formation more than DMSO (Figure 1G). 37 of the 96 hits (38.5%) have no published literature suggesting they are potential ERβ ligands (Table S1). Unlike the ERα-GFP:PRL-HeLa cells, most of the array inducing flavonoids in the GFP-ERβ:PRL-HeLa cells line produced less FISH signal than E2, with only 4 compounds inducing more reporter gene transcription (Figure 1H).
As shown in ToxPred, other nuclear receptors could also be modulated by flavonoids. For that reason, we tested the flavonoid library in androgen receptor (GFP-AR)45 (Figures S1C and S1E) and progesterone receptor (GFP-PR)46 (Figures S1D and S1F) PRL array biosensor cell lines, which have similar endpoints as the ER cell lines. As control for GFP-AR, we use 100nM dihydrotestosterone (DHT), the natural ligand for androgen receptor. As control for GFP-PR, we use 100nM progesterone (P4), the natural ligand for progesterone receptor and 100nM R5020, a synthetic ligand. While the controls successfully induced array formation for their target receptors, none of the flavonoids induced array formation in either cell line at 10μM after 2 h of treatment.
We then chose to validate the hits from the primary screen at two lower doses (100nM and 1μM), to determine the relative potency of these compounds to induce ER array loading and transcriptional output (Figure 2A). From this approach, we identified six compounds that induced ERα DNA binding and transcription (±-equol, equol, genistein, isobavachin, phenoxodiol, and neodiosmin) at the 100nM dose, with four compounds (equol, genistein, isobavachin, and phenoxodiol) also active in the GFP-ERβ:PRL-HeLa cells at the same dose. 23 compounds induced ERα-GFP:PRL-HeLa activity at 1μM, with 36 compounds inducing GFP-ERβ:PRL-HeLa activity. We next examined the hits by chemical classes. Figure 2B shows the entire library fractionated by chemical structure similarity, with flavones and glycosylated flavones being the most dominant class. When evaluating which compound class had the highest enrichment of hits, we found that chalcones (55%) and isoflavones (57%) were the most frequent for ERα (Figure 2C), with isoflavones accounting for the most compounds active at 100nM. For ERβ, chalcones (60%) and glycosylated chalcones (56%) were the most enriched for active compounds.
Figure 2.

Extended dose response library breakdown for hits from flavonoid screen
(A) Heatmap representing DNA binding (proportion of cells with array) and dsRed2 FISH intensity at three concentrations for both ERα-GFP:PRL-HeLa and GFPL-ERβ:PRL-HeLa cell lines. The scale represents each features Z score compared to DMSO.
(B) Percentage breakdown of the flavonoid library by chemical class.
(C) Percent of compounds from each library that hit in each cell line.
(D) Normalized dsRed2 FISH intensity for compounds in the screen that reduced FISH signal. Compounds with an asterisk also formed an array. ∗p < 0.05. Error bars represent standard deviation.
Six flavonoids (Figure 2D) within the screen suppressed transcriptional output to a lower level than DMSO (only one of which, P276-00, also induced array formation) which suggests inhibition of basal transcription. These included flavopiridol and flavopiridol HCl, two known transcriptional inhibitors. Collectively, these data reveal the breadth of activity of flavonoid-related compounds in modulating estrogen receptor function.
Isobavachin and phenoxodiol are potent activators of ERα and ERβ
During the screen, we identified two compounds, isobavachin (IBV) and phenoxodiol (PHX), that induced ERα and ERβ PRL-array targeting (DNA binding) and transcription (mRNA FISH) at the 100nM dose. There is a paucity of information on these compounds which sparked our interest in following them up further. A paper from 2006 showed isobavachin as an activator of ERα54 at similar doses. PHX is a semi-synthetic metabolite of daidzein that displaced E2 from ERα in a rat uterine cytosol assay,55 although at much higher concentrations than used in our assay. Neither compound has been shown to be an ERβ activator. We performed extended dose responses for both active flavonoids and E2 and measured ERα-GFP:PRL-HeLa array formation (Figure 3A) and transcriptional induction (Figure 3D), and GFP-ERβ:PRL-HeLa array formation (Figure 3G) and transcriptional activity (Figure 3J). We determined EC50 for DNA binding in ERα-GFP:PRL-HeLa for E2, IBV, and PHX to be 0.023nM, 1.28nM, and 17.76nM, respectively (Figure 3M). For GFP-ERβ:PRL-HeLa, the EC50 for E2, IBV, and PHX were determined to be 0.017nM, 0.22nM, and 14.31nM, respectively. These two compounds were found to have the lowest EC50 of any flavonoid compound in the screen and induced transcription equivalent to E2 in terms of RNA FISH intensity at the PRL locus.
Figure 3.

Isobavachin and phenoxodiol are potent activators of estrogen receptor
Dose-response curves in ERα-GFP:PRL-HeLa measuring proportion of cells with an array for (A) E2, isobavachin, and phenoxodiol, (B) isobavachin, bavachin, isobavachalcone, neobavaisoflavone, or (C) phenoxodiol, genistein, ±equol, and daidzein. Dose-response curves in ERα-GFP:PRL-HeLa measuring average dsRed2 FISH intensity for (D) E2, isobavachin, and phenoxodiol, (E) isobavachin, bavachin, isobavachalcone, neobavaisoflavone, or (F) phenoxodiol, genistein, ±equol and daidzein. Dose-response curves in GFPL-ERβ:PRL-HeLa measuring proportion of cells with an array for (G) E2, isobavachin, and phenoxodiol, (H) isobavachin, bavachin, isobavachalcone, neobavaisoflavone, or (I) phenoxodiol, genistein, ±equol and daidzein. Dose-response curves in PRL-ERβ measuring average dsRed2 FISH intensity for (J) E2, isobavachin, and phenoxodiol, (K) isobavachin, bavachin, isobavachalcone, neobavaisoflavone, or (L) phenoxodiol, genistein, ±equol and daidzein.
(M) Table including compounds structures and calculated EC50. Error bars represent standard deviation.
We next compared these hits to structurally related compounds within the library. For IBV, we performed extended dose responses on the closely related compounds bavachin, isobavachalcone, and neobavaisoflavone, and determined the DNA binding and transcription data for ERα-GFP:PRL-HeLa (Figures 3B and 3E, respectively) and GFP-ERβ:PRL-HeLa (Figures 3H and 3K, respectively). IBV had a 4,000-fold lower EC50 for ERα (and 1,000-fold lower for ERβ) than its closely related compounds, perhaps due to its non-linear structure when compared to the others (Figure 3M). IBV and its related compounds were all more potent in the GFP-ERβ:PRL-HeLa model.
For comparison to PHX, we performed dose responses on the closely related compounds genistein, ±equol, and daidzein, and evaluated DNA binding and transcription for ERα-GFP:PRL-HeLa (Figures 3C and 3F, respectively), and PRL-ERβ (Figures 3I and 3L, respectively). PHX had a 10-fold lower EC50 than its parent compound, daidzein, for both ERα and ERβ, with slightly lower EC50 than the other isoflavones (Figure 3M). These results also confirm previously published data discussing estrogen receptor subtype-selectivity for flavonoids as IBV, bavachin, isobavachalcone, neobavaisoflavone, and ±equol show greater than a 5-fold selectivity for either estrogen receptor.
IBV and PHX activate ERα target genes and proliferation in MCF-7 cells
Next, to validate our results from the engineered PRL-array model in an orthogonal assay, we sought to determine if IBV and PHX had similar estrogenic actions in an endogenous setting. First, we determined whether the flavonoid hits could activate the well-studied ERα target gene, GREB1,53 in the ERα-positive cell line MCF-7. Using smRNA FISH with probes designed to hybridize to either the introns or exons of GREB1 (representative images in Figure S3A 56), we tested two concentrations of the compound for 24 h of treatment. We measured significant induction of transcriptionally active GREB1 alleles following: (1) E2 treatment (1nM), (2) IBV treatment at both 10 and 100nM, and (3) PHX at 100nM (Figure S3B). Transcriptionally active alleles are defined as an overlapping of intron and exon probes at a defined subnuclear location (for more information see Mistry R.M. et al.56). Additionally, we also utilized qPCR to evaluate whether these compounds induced GREB1 steady state mRNA expression. Figure S3C shows activation of GREB1 transcription as compared to DMSO after 24 h of E2 (1nM), IBV (10nM), PHX, genistein, ±equol (100nM), neobavaisoflavone, bavachin, isobavachalcone, and daidzein (1μM). Furthermore, we sought to determine if IBV and PHX could stimulate proliferation of MCF-7 cells similarly to E2. MCF-7 cells (Figure S3D) were treated with different doses of several of the compounds for 120 h, and then cell number was counted by HT imaging. All compounds tested increased the number of MCF-7 cells (around 150% increase in cell number) compared with DMSO with E2 being the most potent, followed by ± equol, then IBV, and PHX. Taken together, these data demonstrate that IBV and PHX are powerful estrogenic compounds in a physiologically relevant system.
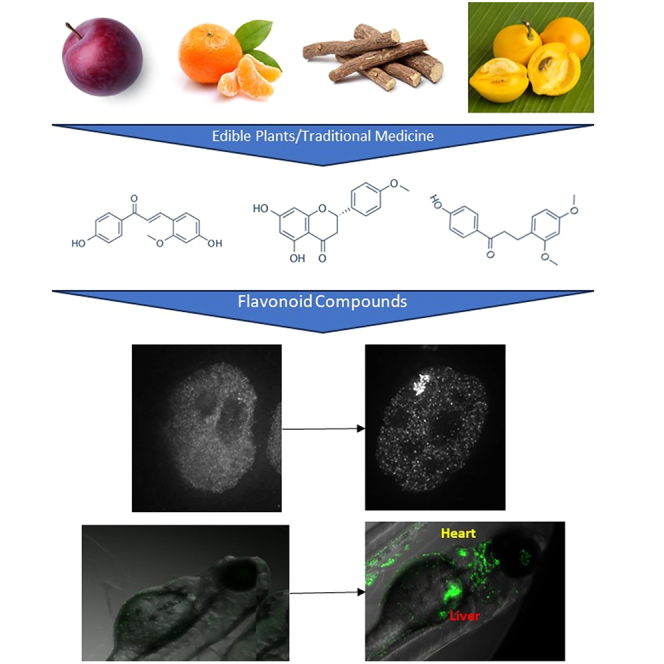
Isosakuranetin, echinatin, and loureirin A are activators of ER
In the primary screen, we identified several flavonoids with no previously reported connection to estrogen receptors. Three of the most potent were isosakuranetin, echinatin, and loureirin A. Isosakuranetin is a flavone found in plums and mandarin oranges,57 echinatin is a chalcone found in Chinese licorice root, and loureirin A, a chalcone, is found in mundu fruit (Garcinia dulcis),58 a sour orange-like fruit in tropical Asian countries, and dragon’s blood (Dracaena cinnabari), a tree used for dyes and medicine in the Mediterranean.59 Therefore, we performed an extended dose response for these three compounds in our ERα-GFP:PRL-HeLa and GFP-ERβ:PRL-HeLa cell lines. Echinatin was able to induce increased array formation (Figure 4A) and accumulation of reporter gene RNA (Figure 4D) in both ERα and ERβ models with EC50 values of 173.59nM and 69.48nM, respectively. Isosakuranetin had similar EC50 for induced array formation (Figure 4B) for ERα (147.88nM) and ERβ (143.12nM), but, interestingly, the EC50 for activation of the RNA FISH (Figure 4E) was over 10-fold lower for ERα when compared to ERβ. However, of the three compounds tested, Isosakuranetin was the least capable of inducing dsRed2 reporter gene. For loureirin A, similar EC50 measurements for array induction (Figure 4C) were for ERα (50.05nM) and ERβ (76.18nM); however, again, ERα RNA FISH induction occurred at a lower concentration than ERβ (Figure 4F). We next sought to determine effects of these compounds on GREB1 gene expression (Figure 4G) and proliferation (Figure 4H) in MCF-7 cells. Although all three compounds significantly increased GREB1 expression when compared to DMSO, only echinatin induced expression to the same level as E2. All three compounds were also able to induce cell proliferation, but loureirin A (50% increase) was not able to reach the level of E2 (150% increase), while isosakuranetin and echinatin were able to do so. Altogether these data show evidence that these three flavonoids are potent activators of both ERα and ERβ.
Figure 4.

Echinatin, isosakuranetin, and loureirin A as activators of estrogen receptors
Dose-response curves for ERα-GFP:PRL-HeLa and GFPL-ERβ:PRL-HeLa showing the proportion of cells with a visible array for (A) echinatin, (B) isosakuranetin, and (C) loureirin A. Dose-response curves for ERα-GFP:PRL-HeLa and GFPL-ERβ:PRL-HeLa for normalized dsRed2 FISH intensity for (D) echinatin, (E) isosakuranetin, and (F) loureirin A.
(G) GREB1 gene expression in MCF-7 cells with E2 at 10nM and isosakuranetin, echinatin, and loureirin A at 1μM after 24 h of treatment.
(H) Proliferation assay in MCF-7 cells. ∗ p value <0.05. Error bars represent standard deviation.
Subtype-selective flavonoid activators of ER
Performing the screen in both the ERα-GFP:PRL-HeLa and GFP-ERβ:PRL-HeLa cells lines afforded us the ability to determine subtype specificity and selectivity of the flavonoid compounds. From the primary screen, we found 14 compounds that only induced array formation in the ERα-GFP:PRL-HeLa and 24 compounds that only induced array formation in the GFP-ERβ:PRL-HeLa (Figure 5A). It is important to note that all of these compounds had EC50 > 1μM (as seen in Table S1), and it is possible that had we used a higher concentration than 10μM, more of these compounds would have been hits for both receptors. To illustrate, we plotted the EC50 for each hit compound (Figure 5B) to better understand subtype-specific compounds. In addition to IBV, neobavaisoflavone, bavachin, and isobavachalcone, licochalcone A, a chalcone found in licorice (Glycyrrhiza glabra), is another compound with ERβ-selectivity is,. Lichochalcone A shows a 5-fold preference (Figure 5C) for ERβ (EC50 92.1nM) compared to ERα (EC50 508.23nM). Interestingly, the highest dose tested (10μM) of licochalcone A did not activate RNA FISH expression in the ERα-GFP:PRL-HeLa cell line (Figure 5D), but it did activate the dsRed2 reporter gene in GFP-ERβ:PRL-HeLa.
Figure 5.

Subtype selectivity of compounds in the flavonoid screen
(A) Venn diagram of hits in the ERα-GFP:PRL-HeLa and GFPL-ERβ:PRL-HeLa cell lines.
(B) Scatterplot looking at hit compounds ERα-GFP:PRL-HeLa and GFPL-ERβ:PRL-HeLa cell lines. Compounds that were not a hit in a cell line were set at 100000nM dose. Dose-response curve for licochalcone A for (C) proportion of cells with array and (D) normalized dsRed2 FISH intensity.
(E) Example images of zebrafish following different treatments.
(F) Heatmap showing percent of zebrafish embryos with GFP-positive heart or liver upon a given treatment. n ≥ 18 zebrafish embryos per condition. Error bars represent standard deviation.
Utilizing an orthogonal organismal assay to assess flavonoids activity in vivo
To further test the activity of flavonoid compounds in an in vivo model, we utilized a previously published transgenic zebrafish model47 that is capable of differentiating between ERα and ERβ activities. In the zebrafish model, compounds that activate the ERE-GFP reporter in the liver do so by binding to the zebrafish homolog of ERα (zebrafish esr2a and esr2b), while ERE-GFP in the heart valves is an indicator of ERβ activity (zebrafish esr1). Figure 5E shows example images of the zebrafish embryos treated with DMSO or 100nM E2 for 3 days post fertilization, with yellow arrows identifying ERE-GFP signal in the liver and red arrows indicating ERE-GFP signal in the heart. We found that E2, IBV, PHX, and ±equol, activate ER signal in both the heart and liver at the highest nonlethal doses tested (Figure 5F, images in Figures S4B–S4F). We tested the flavonoid compounds with concentrations up to 100μM, and all caused embryo death, while the same amount of DMSO was not lethal (examples in Figure S4A). Within the zebrafish assay, we see subtype selective effects with liquiritigenin (Figure S4C), which at 10μM only activates the ERE-GFP in the zebrafish heart (e.g., ERβ). IBV (Figure S4E), at 1μM, resulted in GFP-positive livers in 83.3% of the zebrafish but GFP-positive heart valves in only 41.6%. Neither isosakuranetin, echinatin, nor loureirin A activated the ERE-GFP in the zebrafish at the nonlethal doses tested (<100μM). We hypothesize that this is likely due to the previously characterized reduced sensitivity of the zebrafish assay.47 Even with the positive control E2, the ERE-activity in the zebrafish models does not appear until 10nM, meaning this assay is much less sensitive (∼2 logs) than our PRL-array lines, likely reflecting the role of ligands to be absorbed through the skin. Even so, we could measure subtype selectivity and dose responsiveness further lending evidence that the newly characterized flavonoids compounds are being ER activators in a subtype-selective manner.
Discussion
In this study, we present a primary screen and follow-on results designed to determine how a library of 224 flavonoid compounds affected mechanistic steps (DNA binding and transcription induction) associated with ER transcription across multiple cell-based assays. Some flavonoid compounds have previously been evaluated in multiple assays due to their bioactivity and presence in highly consumed dietary sources. Many flavonoids have been shown to have antioxidant activity and other beneficial effects, including their effectiveness as cancer treatments. A few studies8,32,60 have revealed some flavonoids possess an ability to activate ERα and ERβ transcription and induce proliferation; indeed, many flavonoids are termed phytoestrogens, but only a small number of their large chemical family has routinely been investigated. A recent study showed how a flavonoid, isosinensetin (used at low micromolar concentrations, not present in our library screen), can rescue estrogen deficiency-induced osteoporosis through ER pathway activation in murine bone assays.61 In our high throughput, high content screen, we confirmed many known ER-active compounds (genistein, daidzein, apigenin), and determined that within our 85 hits, 49 (57.6%) had no scientific literature attributing the compound to ERα activation, thus justifying the screening approach with an HT amenable mechanistic HCA-based screen. Interestingly, out of all the hits only two of the compounds induced ER recruitment to the PRL array but suppressed transcription (P276-00 and irigenin). This is as opposed to compounds such as bisphenols, where our group found that >50% of bisphenols tested could act antagonistically in these cells.62 We identified, for the first time, multiple ERα-active flavonoids that are found in human-consumable plants used in everyday life including rice (epicatechin gallate), sage/rosemary (4,7-demethoxy-5-hydroxyflavone, eriodictyol, hesperetin, and many more), mandarin oranges (acacetin), raspberry (isoquercitrin), and cucumbers (vitexin and isovitexin), thus markedly increasing human population exposure to estrogenic compounds through food consumption.63,64 While we have focused our efforts on the rapid, genomic signaling of E2, flavonoids that did not activate the PRL-array locus could possibly activate ER through non-canonical pathways, such as membrane ER or GPER.
Many of the flavonoids show specificity for one receptor over the other, with 21 compounds showing at least a 5-fold preference. It is well documented that ERβ has a smaller and narrower ligand binding pocket than ERα with only two amino acids different in the ligand binding cavity.65 These changes lead to the design of ligands with receptor subtype specificity like PPT (ERα) and DPN (ERβ).66 Agonist-induced activation of the estrogen receptors requires the movement of helix 12 to allow for the recruitment of coactivators (such as SRC-3, p300) and finally, recruitment of RNA polymerase to activate transcription.67 We previously discovered compounds within the EPA45 library (progesterone, dibutyl phthalate) [45] that could induce array formation without significantly inducing the dsRed2 FISH signal above DMSO. We hypothesize these compounds, upon binding, change the conformation to allow for DNA binding while not changing the position of helix 12 enough to recruit coactivators (like true agonists) or corepressors (like true antagonists).
Intriguingly, IBV and PHX were the two most potent flavonoids in the library that activated ERα, with EC50 of 1.28nM and 17.76nM, respectively. IBV, found in Psoralea corylifolia, a plant used in Indian and Chinese traditional medicine (used to treat the skin condition, lichen planus68), was the most potent compound in the screen. A single previous article from 200654 has shown IBV and its related compounds as activators of ERα, and a related article showed IBV could induce proliferation in MCF-7 cells.60 Further evidence of IBV effects on the ERα pathway comes from the finding that it is protective against osteoporosis69 where IBV treatment could reverse the effect of estrogen deficiency in bone repair assays. Although IBV has not been shown before to be an ERβ ligand, we found it to be ∼5-fold more selective for ERβ. PHX, a semi-synthetic derivative of genistein and daidzein, was the second most potent compound tested and has previously been shown to displace E2 from ERα in rat uteri (EC50 of ∼500nM).55 PHX, also known as idroxinil and haginin E, has been studied for its ability to sensitize colon cancer cells to 5-fluorouracil and oxiplatin through induction of caspase-3 RNA levels and activity70 though clinical use has not been successful due to low bioavailability.71 However, the concentrations used in these studies generally were much higher (1-10μM) than the observed EC50 we determined for ERα activation (<20nM), comparable to what we observed in the zebrafish model.
We also characterized three new flavonoids with no previously published literature on ERα or ERβ. Isosakuranetin, echinatin, and loureirin A all showed sub-micromolar EC50 for both ERα and ERβ array formation and MCF-7 growth assay. Differences in EC50 for the assays presented in this article can be related to time of ligand exposure (1 h for array binding, 72 h for zebrafish, 120 h for MCF7 growth) as well as compound uptake and metabolism in the zebrafish assay. Isosakuranetin, found in plums and mandarin oranges, is able to relieve chronic pain in rats72 and has been shown to be a potent inhibitor of TRPM3 channels in neurons.73 Echinatin can protect against arterial stiffening74 and induce apoptosis in colorectal cancer cells lines HT116 and HT29.75 Loureirin A, found in dragon’s blood76 was used to promote blood circulation and can relieve arthritis pain in mice.77 The potency of these compounds in our model makes them potentially interesting candidates for estrogen replacement therapies/supplements. Although this study is the largest screen of flavonoids tested for ERα and ERβ activity to date, there remain ∼6,000 compounds in the flavonoid class that contribute to pigments of fruits, vegetables, and medicinal plants,78 that could potentially show a similar scale of wide ranging subtype-selectivity. If the hit percentage (40%) we found in our library holds true for the larger set of flavonoid compounds, that means there could be as many as 2,400 estrogenic flavonoids many of which could show subtype-selectivity that remain to be characterized. The ERβ-selectivity of the bavachin compounds (bavachin, isobavachalcone, neobavaisoflavone, and IBV) provides a core of compounds to help determine the flavonoid chemical structure required for ER subtype selectively and an excellent starting point for the creation of synthetic subtype-selective compounds. These studies showcase how we are just scratching the surface of flavonoid-ER biology with potent estrogenic flavonoids like IBV being underutilized in estrogen research and therapies.
Limitations of the study
A limitation of this study includes the reduced sensitivity of the ERE-GFP reporter in transgenic zebrafish to estrogen. Due to the required aquatic addition of hormone and necessity to diffuse through the skin, the assay required 10nM E2 to activate the reporter in the zebrafish model, while our cellular assay responds to 100pM E2. Also, most flavonoid compounds were lethal >10μM to zebrafish embryos; nonetheless, we were able to confirm four hits that were among the most potent in our cellular assays. With 10μM concentrations of flavonoids, it is possible that some compounds may be less subtype-specific at concentrations higher than 10μM.
STAR★Methods
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Chemicals, peptides, and recombinant proteins | ||
| Flavonoid Compound Library | Selleckchem | L7700 |
| Estradiol | SigmaAldrich | |
| Isobavachin | TargetMol | T3931 |
| Phenoxodiol | TargetMol | T16522 |
| Bavachin | TargetMol | T3846 |
| Isobavachalcone | TargetMol | T3861 |
| Neobavaisoflavone | TargetMol | T6S0139 |
| Daidzein | TargetMol | T1238 |
| ±Equol | TargetMol | T4000 |
| Genistein | TargetMol | T1737 |
| Echinatin | TargetMol | T3926 |
| Isosakuranetin | TargetMol | T5S0229 |
| Loureirin A | TargetMol | T5S0896 |
| Lichochalcone A | TargetMol | T3600 |
| Liquiritigenin | TargetMol | T3325 |
| Experimental models: Cell lines | ||
| ERα-GFP:PRL-HeLa | Mancini Lab | |
| GFP-ERβ:PRL-HeLa | Mancini Lab | |
| GFP-AR(DBDswap):PRL-HeLa | Mancini Lab | |
| GFP-PR(DBDswap):PRL-HeLa | Mancini Lab | |
| MCF-7 | Mancini Lab | |
| Experimental models: Organisms/Strains | ||
| Transgenic Zebrafish | Daniel Gorelick Lab | |
| Software and algorithms | ||
| Pipeline Pilot Image Analysis software | Biovia | https://www.3ds.com/products-services/biovia/products/data-science/pipeline-pilot/ |
| Orange Software | Demsar et al. (2003) | https://orangedatamining.com |
| AAT Bio EC50 Calculator | AAT Bio | https://www.aatbio.com/tools/ec50-calculator |
| Other | ||
| dsRED2 FISH Probe | LGC Biosearch | Custom |
Resource availability
Lead contact
Further information for resources and reagents should be directed to and will be fulfilled by the lead contact, Michael Mancini (mancini@bcm.edu).
Materials availability
This study did not generate new unique reagents.
Data and code availability
Data reported in this paper will be shared by the lead contact upon request. This paper does not report any original code. Sources are listed in the key resources table for all analysis programs utilized. Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
Experimental model and study participant details
Cell experiments were performed with ERα-GFP:PRL-HeLa, GFP-ERβ:PRL-HeLa, GFP-ARDBDswap:PRL-HeLa, GFP-PRDBDswap:PRL-HeLa, or MCF-7 cell lines. The PRL-HeLa cell lines are under doxycycline control of their nuclear receptor. All cell lines are maintained in phenol-red free Dulbecco’s modified eagle’s media (DMEM) containing 10% fetal bovine serum (FBS), with the GFP cell line media containing 200 μg/ml Hygromycin and 100μg/ml G418, and the MCF-7 cell media containing 1X Penn/Strep. Cells were plated for 48hrs in 5% charcoal-stripped FBS, phenol-red free DMEM before treatment on a Greiner 384-well optical bottom plate. To initiate receptor expression, cells were treated with doxycycline (1μg/ml) 24 hours before experimental treatment.
Transgenic zebrafish were obtained and handled through the BCM Advanced Technology Zebrafish Core. Adult zebrafish were raised on a 14-h light, 10-h dark cycle in the BCM Zebrafish Research Facility on a Tecniplast recirculating water system (Tecniplast S.p.A., Buguggiate, Italy). All zebrafish used for experiments were Tg(5xERE:GFP)c262 (37,38). All zebrafish experiments were approved by the BCM Institutional Animal Care and Use Committee.
Adult zebrafish were allowed to spawn naturally in groups for one hour, after which embryos were harvested. Embryos were collected in 60 cm2 Petri dishes in E2B with methylene blue (7.5 mM NaCl, 0.25 mM KCl, 0.5 mM MgSO4, 75 μM KH2PO4, 25 μM Na2HPO4, 0.5 mM CaCl2, NaHCO3, 0.00005% methylene blue) before being sorted into densities of no more than 100 embryos in new 60 cm2 Petri dishes containing E3B (5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, 0.00002% methylene blue), and then stored in an incubator at 28.5°C on a 14-h light, 10-h dark cycle until treatment. Embryos (still within their chorions) were plated into a 96-well plate that was preloaded with the chemicals of interest. Using a 1000 μl pipet tip with the narrow end cut off, embryos were collected 3 at a time in 200 μl E3B for each well of the plate. Completed plates were then stored in a completely dark incubator at 28.5°C until embryos were ready to be imaged. Embryos were allowed to hatch naturally from their chorions.
Zebrafish were anesthetized using tricaine at 25 μg/mL. Plates were then imaged (brightfield and GFP channels) with a Yokogawa CV8000 microscope using an Olympus 4x/0.16 objective scanning the whole well. A z-stack of 500 μm was collected at 50 μm steps, converted into maximum intensity projections for image analysis. Each image was qualitatively evaluated by visual inspection and scored for number of live zebrafish, those appearing dead or under stress (stressed zebrafish have a curved spine/dead zebrafish can be seen in varying states of decomposition), GFP signal in the heart valves, or GFP signal in the liver.
Method details
Fluorescence in situ hybridization
Cells in a 384-well plate were fixed in EM-grade 4% formaldehyde in RNAse-free phosphate-buffered saline for 20 min and then permeabilized with 70% ethanol in RNAse-free water at 4°C overnight. Cells were washed in wash buffer (WB, 2X SSC and 10% formamide), followed by hybridization buffer (0.1g dextran sulfate, 1ml of 20X SSC buffer, 1ml of formamide and 8ml of nuclease-free water) containing a dsRED2 RNA probe (Q670, LGC Biosearch Technologies, diluted 1:1000) or GREB1 intron (A647N) and exon (Q570) probes (LGC Biosearch Technologies, diluted 1:500) overnight at 37°C. Following probe hybridization, cells were washed with WB for 30 minutes, followed by 2X SSC buffer containing DAPI for 30 min. Cells were left in 2X SSC for imaging.
High throughput imaging & analysis
PRL-Array cells were imaged on an ImageXpress Micro (Molecular Devices) high throughput confocal microscope with a 20X/0.75 NA lens including seven z-stacks at 1μm intervals in the GFP and FISH channels without image binning and 4 fields per well. Image analysis was performed using a custom workflow developed with the imaging collection within the Pipeline Pilot GUI-based programming platform (PLP, BIOVIA) as described previously.79 In brief, images and metadata data are first read into PLP and then background corrected. Next, the ER-GFP and FISH z-stacked images are projected using the “stack focuser” plug-in integrated into PLP. The DAPI signal is then used to define nuclear regions that are subsequently expanded by 8μm to include analysis of the perinuclear cytoplasm. Array segmentation is performed using a local subtraction (per nuclear area), peak-finding algorithm followed by user-defined filtering to minimize false-positives. Count, morphometric and intensity-based features are subsequently measured for each channel and mask combination.
For GREB1 smFISH analysis in MCF-7 cells, images were taken on a Cytivia DeltaVision deconvolution microscope using a 60X/1.4NA oil immersion lens from glass coverslips. Further technical details can be found in a smFISH protocol paper published by our group.56
qPCR and proliferation
For qPCR, MCF7 cells were plated for 48h in 5% charcoal-stripped FBS, phenol red free Dulbecco’s modified Eagle’s medium before treatment in a 6-well plate. Cells were treated with compound for 24 hours before media was removed and TriReagent was added. Total RNA was isolated using the Directzol RNA miniprep kit (Zymoresearch) using the manufacturer’s instructions, including a DNAse treatment. One microgram of total RNA was reverse transcribed using Moloney Murine Leukemia VVirus (M-MuLV) reverse transcriptase, random hexamers and dNTPs (New England Biolabs). The cDNA was analyzed with real-time PCR using 1x SYBR iTaq Supermix (BioRad) with primers synthesized by IDT. Reactions were carried out in an Quantstudio 6 Flex (ThermoFisher) for 40 cycles. The fold change in expression was calculated using the ΔΔCt comparative threshold cycle method,79 with ribosomal protein 36B4 mRNA as an internal control.
For proliferation studies, before treatment MCF7 cells were plated in a 384-well for 120h in 5% charcoal-stripped FBS phenol red free Dulbecco’s modified Eagle’s medium. Compounds were added to MCF7 cells for 120h followed by fixation and DAPI staining. Nuclei were counted automatically using PLP. Each compound/dose was performed in quadruplicate wells.
Quantification and statistical analysis
Graphics & statistical analysis
All graphics were made using Microsoft Excel unless otherwise stated. The heat map in Figure 2 was made in the data analysis software Orange.80 p-value was determined using student’s t-test for single comparisons or one-way ANOVA for independent samples and a Tukey HSD post-hoc test for comparisons across multiple samples. EC50’s were calculated using the AAT Bio EC50 calculator.
Acknowledgments
M.A.M, F.S., and M.G.M. are funded in part by NIEHS (P42ES027704). M.A.M., M.J.B., J.O., P.S., F.S., K.S., D.H.A. are supported via the CPRIT-funded GCC Center for Advanced Microscopy and Image Informatics (RP170719). P.S. and M.A.M. are also supported in part by the CPRIT-funded Combinatorial Drug Discovery Program (RP200668). M.A.M. is also partially funded by the GCC Center for Precision Environmental Health (P30ES030285). D.A.G. and K.A.N. are supported by NIH ES030285.
Author contributions
Conceptualization, M.J.B., F.S., and M.A.M.; investigation, M.J.B. and J.O.; methodology, M.J.B., J.O., P.S., K.S., D.H.A., K.A.N., and D.A.G.; resources, M.G.M, P.S., and D.A.G., writing – original draft, M.J.B.; writing – review & editing, M.J.B., F.S., and M.A.M.; visualization, M.J.B.; supervision, F.S. and M.A.M.; funding acquisition, M.A.M.
Declaration of interests
The authors declare no competing interests.
Published: February 20, 2024
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.isci.2024.109275.
Supplemental information
References
- 1.Khalid A.B., Krum S.A. Estrogen receptors alpha and beta in bone. Bone. 2016;87:130–135. doi: 10.1016/j.bone.2016.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Feng Y., Manka D., Wagner K.U., Khan S.A. Estrogen receptor-alpha expression in the mammary epithelium is required for ductal and alveolar morphogenesis in mice. Proc. Natl. Acad. Sci. USA. 2007;104:14718–14723. doi: 10.1073/pnas.0706933104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Murphy E. Estrogen signaling and cardiovascular disease. Circ. Res. 2011;109:687–696. doi: 10.1161/CIRCRESAHA.110.236687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Yu K., Huang Z.Y., Xu X.L., Li J., Fu X.W., Deng S.L. Estrogen Receptor Function: Impact on the Human Endometrium. Front. Endocrinol. 2022;13 doi: 10.3389/fendo.2022.827724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gennari L., Merlotti D., Nuti R. Selective estrogen receptor modulator (SERM) for the treatment of osteoporosis in postmenopausal women: focus on lasofoxifene. Clin. Interv. Aging. 2010;5:19–29. doi: 10.2147/cia.s6083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Porras L., Ismail H., Mader S. Positive Regulation of Estrogen Receptor Alpha in Breast Tumorigenesis. Cells. 2021;10 doi: 10.3390/cells10112966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Martin E.M., Orlando K.A., Yokobori K., Wade P.A. The estrogen receptor/GATA3/FOXA1 transcriptional network: lessons learned from breast cancer. Curr. Opin. Struct. Biol. 2021;71:65–70. doi: 10.1016/j.sbi.2021.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Darbre P.D. Endocrine disrupting chemicals and breast cancer cells. Adv. Pharmacol. 2021;92:485–520. doi: 10.1016/bs.apha.2021.04.006. [DOI] [PubMed] [Google Scholar]
- 9.Judson R.S., Magpantay F.M., Chickarmane V., Haskell C., Tania N., Taylor J., Xia M., Huang R., Rotroff D.M., Filer D.L., et al. Integrated Model of Chemical Perturbations of a Biological Pathway Using 18 In Vitro High-Throughput Screening Assays for the Estrogen Receptor. Toxicol. Sci. 2015;148:137–154. doi: 10.1093/toxsci/kfv168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Williams G.P., Darbre P.D. Low-dose environmental endocrine disruptors, increase aromatase activity, estradiol biosynthesis and cell proliferation in human breast cells. Mol. Cell. Endocrinol. 2019;486:55–64. doi: 10.1016/j.mce.2019.02.016. [DOI] [PubMed] [Google Scholar]
- 11.Waheed Janabi A.H., Kamboh A.A., Saeed M., Xiaoyu L., BiBi J., Majeed F., Naveed M., Mughal M.J., Korejo N.A., Kamboh R., et al. Flavonoid-rich foods (FRF): A promising nutraceutical approach against lifespan-shortening diseases. Iran. J. Basic Med. Sci. 2020;23:140–153. doi: 10.22038/IJBMS.2019.35125.8353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Egert S., Rimbach G. Which sources of flavonoids: complex diets or dietary supplements? Adv. Nutr. 2011;2:8–14. doi: 10.3945/an.110.000026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shanmugavadivu A., Balagangadharan K., Selvamurugan N. Angiogenic and osteogenic effects of flavonoids in bone regeneration. Biotechnol. Bioeng. 2022;119:2313–2330. doi: 10.1002/bit.28162. [DOI] [PubMed] [Google Scholar]
- 14.Munguía L., Ortiz M., González C., Portilla A., Meaney E., Villarreal F., Nájera N., Ceballos G. Beneficial Effects of Flavonoids on Skeletal Muscle Health: A Systematic Review and Meta-Analysis. J. Med. Food. 2022;25:465–486. doi: 10.1089/jmf.2021.0054. [DOI] [PubMed] [Google Scholar]
- 15.Ramesh P., Jagadeesan R., Sekaran S., Dhanasekaran A., Vimalraj S. Flavonoids: Classification, Function, and Molecular Mechanisms Involved in Bone Remodelling. Front. Endocrinol. 2021;12 doi: 10.3389/fendo.2021.779638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Al-Khayri J.M., Sahana G.R., Nagella P., Joseph B.V., Alessa F.M., Al-Mssallem M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules. 2022;27 doi: 10.3390/molecules27092901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hamsalakshmi, Alex A.M., Arehally Marappa M., Joghee S., Chidambaram S.B. Therapeutic benefits of flavonoids against neuroinflammation: a systematic review. Inflammopharmacology. 2022;30:111–136. doi: 10.1007/s10787-021-00895-8. [DOI] [PubMed] [Google Scholar]
- 18.Li R.L., Wang L.Y., Liu S., Duan H.X., Zhang Q., Zhang T., Peng W., Huang Y., Wu C. Natural Flavonoids Derived From Fruits Are Potential Agents Against Atherosclerosis. Front. Nutr. 2022;9 doi: 10.3389/fnut.2022.862277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Biswas M., Das A., Basu S. Flavonoids: The Innocuous Agents Offering Protection against Alzheimer's Disease Through Modulation of Proinflammatory and Apoptotic Pathways. Curr. Top. Med. Chem. 2022;22:769–789. doi: 10.2174/1568026622666220330011645. [DOI] [PubMed] [Google Scholar]
- 20.Medrano-Jiménez E., Meza-Sosa K.F., Urbán-Aragón J.A., Secundino I., Pedraza-Alva G., Pérez-Martínez L. Microglial activation in Alzheimer's disease: The role of flavonoids and microRNAs. J. Leukoc. Biol. 2022;112:47–77. doi: 10.1002/JLB.3MR1021-531R. [DOI] [PubMed] [Google Scholar]
- 21.Pannu A., Sharma P.C., Thakur V.K., Goyal R.K. Emerging Role of Flavonoids as the Treatment of Depression. Biomolecules. 2021;11 doi: 10.3390/biom11121825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ali S., Corbi G., Maes M., Scapagnini G., Davinelli S. Exploring the Impact of Flavonoids on Symptoms of Depression: A Systematic Review and Meta-Analysis. Antioxidants. 2021;10 doi: 10.3390/antiox10111644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Li C., Li X., Jiang Z., Wang D., Sun L., Li J., Han Y. Flavonoids Inhibit Cancer by Regulating the Competing Endogenous RNA Network. Front. Oncol. 2022;12 doi: 10.3389/fonc.2022.842790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang M., Yu F., Zhang Y., Chang W., Zhou M. The Effects and Mechanisms of Flavonoids on Cancer Prevention and Therapy: Focus on Gut Microbiota. Int. J. Biol. Sci. 2022;18:1451–1475. doi: 10.7150/ijbs.68170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Berk Ş., Kaya S., Akkol E.K., Bardakçı H. A comprehensive and current review on the role of flavonoids in lung cancer-Experimental and theoretical approaches. Phytomedicine. 2022;98 doi: 10.1016/j.phymed.2022.153938. [DOI] [PubMed] [Google Scholar]
- 26.Slika H., Mansour H., Wehbe N., Nasser S.A., Iratni R., Nasrallah G., Shaito A., Ghaddar T., Kobeissy F., Eid A.H. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed. Pharmacother. 2022;146 doi: 10.1016/j.biopha.2021.112442. [DOI] [PubMed] [Google Scholar]
- 27.Zhai K., Mazurakova A., Koklesova L., Kubatka P., Büsselberg D. Flavonoids Synergistically Enhance the Anti-Glioblastoma Effects of Chemotherapeutic Drugs. Biomolecules. 2021;11 doi: 10.3390/biom11121841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dutta B., Park J.E., Qing I.T.Y., Kon O.L., Sze S.K. Soy-Derived Phytochemical Genistein Modifies Chromatome Topology to Restrict Cancer Cell Proliferation. Proteomics. 2018;18 doi: 10.1002/pmic.201700474. [DOI] [PubMed] [Google Scholar]
- 29.Yuseran H., Hartoyo E., Nurseta T., Kalim H. Molecular docking of genistein on estrogen receptors, promoter region of BCLX, caspase-3, Ki-67, cyclin D1, and telomere activity. J. Taibah Univ. Med. Sci. 2019;14:79–87. doi: 10.1016/j.jtumed.2018.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chan K.K.L., Siu M.K.Y., Jiang Y.X., Wang J.J., Leung T.H.Y., Ngan H.Y.S. Estrogen receptor modulators genistein, daidzein and ERB-041 inhibit cell migration, invasion, proliferation and sphere formation via modulation of FAK and PI3K/AKT signaling in ovarian cancer. Cancer Cell Int. 2018;18:65. doi: 10.1186/s12935-018-0559-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Duan X., Li Y., Xu F., Ding H. Study on the neuroprotective effects of Genistein on Alzheimer's disease. Brain Behav. 2021;11 doi: 10.1002/brb3.2100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lambert M.N.T., Hu L.M., Jeppesen P.B. A systematic review and meta-analysis of the effects of isoflavone formulations against estrogen-deficient bone resorption in peri- and postmenopausal women. Am. J. Clin. Nutr. 2017;106:801–811. doi: 10.3945/ajcn.116.151464. [DOI] [PubMed] [Google Scholar]
- 33.Song Y.Q., Guan X.Q., Weng Z.M., Liu J.L., Chen J., Wang L., Cui L.T., Fang S.Q., Hou J., Ge G.B. Discovery of hCES2A inhibitors from Glycyrrhiza inflata via combination of docking-based virtual screening and fluorescence-based inhibition assays. Food Funct. 2021;12:162–176. doi: 10.1039/d0fo02140g. [DOI] [PubMed] [Google Scholar]
- 34.Licorice. 2022. https://www.drugs.com/mtm/licorice.html#side-effects
- 35.Patel S., Homaei A., Raju A.B., Meher B.R. Estrogen: The necessary evil for human health, and ways to tame it. Biomed. Pharmacother. 2018;102:403–411. doi: 10.1016/j.biopha.2018.03.078. [DOI] [PubMed] [Google Scholar]
- 36.Chen T., Wu F., Wang X., Ma G., Xuan X., Tang R., Ding S., Lu J. Different levels of estradiol are correlated with sexual dysfunction in adult men. Sci. Rep. 2020;10 doi: 10.1038/s41598-020-69712-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yu M., Qi H., Gao X. Daidzein promotes milk synthesis and proliferation of mammary epithelial cells via the estrogen receptor alpha-dependent NFkappaB1 activation. Anim. Biotechnol. 2022;33:43–52. doi: 10.1080/10495398.2020.1763376. [DOI] [PubMed] [Google Scholar]
- 38.Pham T.H., Page Y.L., Percevault F., Ferrière F., Flouriot G., Pakdel F. Apigenin, a Partial Antagonist of the Estrogen Receptor (ER), Inhibits ER-Positive Breast Cancer Cell Proliferation through Akt/FOXM1 Signaling. Int. J. Mol. Sci. 2021;22 doi: 10.3390/ijms22010470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pellegrini M., Bulzomi P., Galluzzo P., Lecis M., Leone S., Pallottini V., Marino M. Naringenin modulates skeletal muscle differentiation via estrogen receptor alpha and beta signal pathway regulation. Genes Nutr. 2014;9:425. doi: 10.1007/s12263-014-0425-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nishimura Y., Mabuchi K., Takano A., Hara Y., Negishi H., Morimoto K., Ueno T., Uchiyama S., Takamata A. S-equol Exerts Estradiol-Like Anorectic Action with Minimal Stimulation of Estrogen Receptor-alpha in Ovariectomized Rats. Front. Endocrinol. 2017;8:281. doi: 10.3389/fendo.2017.00281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pang X.G., Cong Y., Bao N.R., Li Y.G., Zhao J.N. Quercetin Stimulates Bone Marrow Mesenchymal Stem Cell Differentiation through an Estrogen Receptor-Mediated Pathway. BioMed Res. Int. 2018;2018 doi: 10.1155/2018/4178021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xin D., Wang H., Yang J., Su Y.F., Fan G.W., Wang Y.F., Zhu Y., Gao X.M. Phytoestrogens from Psoralea corylifolia reveal estrogen receptor-subtype selectivity. Phytomedicine. 2010;17:126–131. doi: 10.1016/j.phymed.2009.05.015. [DOI] [PubMed] [Google Scholar]
- 43.Hajirahimkhan A., Mbachu O., Simmler C., Ellis S.G., Dong H., Nikolic D., Lankin D.C., van Breemen R.B., Chen S.N., Pauli G.F., et al. Estrogen Receptor (ER) Subtype Selectivity Identifies 8-Prenylapigenin as an ERbeta Agonist from Glycyrrhiza inflata and Highlights the Importance of Chemical and Biological Authentication. J. Nat. Prod. 2018;81:966–975. doi: 10.1021/acs.jnatprod.7b01070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bolt M.J., Singh P., Obkirchner C.E., Powell R.T., Mancini M.G., Szafran A.T., Stossi F., Mancini M.A. Endocrine disrupting chemicals differentially alter intranuclear dynamics and transcriptional activation of estrogen receptor-alpha. iScience. 2021;24 doi: 10.1016/j.isci.2021.103227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Szafran A.T., Bolt M.J., Obkirchner C.E., Mancini M.G., Helsen C., Claessens F., Stossi F., Mancini M.A. A Mechanistic High-Content Analysis Assay Using a Chimeric Androgen Receptor That Rapidly Characterizes Androgenic Chemicals. SLAS Discov. 2020;25:695–708. doi: 10.1177/2472555220922917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Treviño L.S., Bolt M.J., Grimm S.L., Edwards D.P., Mancini M.A., Weigel N.L. Differential Regulation of Progesterone Receptor-Mediated Transcription by CDK2 and DNA-PK. Mol. Endocrinol. 2016;30:158–172. doi: 10.1210/me.2015-1144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Gorelick D.A., Lucia C., Hao R., Karim S., Bondesson M. Use of Reporter Genes to Analyze Estrogen Response: The Transgenic Zebrafish Model. Methods Mol. Biol. 2022;2418:173–185. doi: 10.1007/978-1-0716-1920-9_11. [DOI] [PubMed] [Google Scholar]
- 48.Szafran A.T., Mancini M.G., Stossi F., Mancini M.A. Sensitive image-based chromatin binding assays using inducible ERalpha to rapidly characterize estrogenic chemicals and mixtures. iScience. 2022;25 doi: 10.1016/j.isci.2022.105200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stossi F., Singh P.K., Mistry R.M., Johnson H.L., Dandekar R.D., Mancini M.G., Szafran A.T., Rao A.U., Mancini M.A. Quality Control for Single Cell Imaging Analytics Using Endocrine Disruptor-Induced Changes in Estrogen Receptor Expression. Environ. Health Perspect. 2022;130 doi: 10.1289/EHP9297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Abbott D.A., Mancini M.G., Bolt M.J., Szafran A.T., Neugebauer K.A., Stossi F., Gorelick D.A., Mancini M.A. A novel ERbeta high throughput microscopy platform for testing endocrine disrupting chemicals. Heliyon. 2023;10 doi: 10.1016/j.heliyon.2023.e23119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ramaprasad A.S.E., Smith M.T., McCoy D., Hubbard A.E., La Merrill M.A., Durkin K.A. Predicting the binding of small molecules to nuclear receptors using machine learning. Brief. Bioinform. 2022;23 doi: 10.1093/bib/bbac114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kiyama R. Estrogenic flavonoids and their molecular mechanisms of action. J. Nutr. Biochem. 2023;114 doi: 10.1016/j.jnutbio.2022.109250. [DOI] [PubMed] [Google Scholar]
- 53.Stossi F., Dandekar R.D., Mancini M.G., Gu G., Fuqua S.A.W., Nardone A., De Angelis C., Fu X., Schiff R., Bedford M.T., et al. Estrogen-induced transcription at individual alleles is independent of receptor level and active conformation but can be modulated by coactivators activity. Nucleic Acids Res. 2020;48:1800–1810. doi: 10.1093/nar/gkz1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wang Z.Q., Weber N., Lou Y.J., Proksch P. Prenylflavonoids as nonsteroidal phytoestrogens and related structure-activity relationships. ChemMedChem. 2006;1:482–488. doi: 10.1002/cmdc.200500089. [DOI] [PubMed] [Google Scholar]
- 55.Tilley A.J., Zanatta S.D., Qin C.X., Kim I.K., Seok Y.M., Stewart A., Woodman O.L., Williams S.J. 2-Morpholinoisoflav-3-enes as flexible intermediates in the synthesis of phenoxodiol, isophenoxodiol, equol and analogues: vasorelaxant properties, estrogen receptor binding and Rho/RhoA kinase pathway inhibition. Bioorg. Med. Chem. 2012;20:2353–2361. doi: 10.1016/j.bmc.2012.02.008. [DOI] [PubMed] [Google Scholar]
- 56.Mistry R.M., Singh P.K., Mancini M.G., Stossi F., Mancini M.A. Single Cell Analysis Of Transcriptionally Active Alleles By Single Molecule FISH. J. Vis. Exp. 2020 doi: 10.3791/61680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Luo Y., Zeng W., Huang K.E., Li D.X., Chen W., Yu X.Q., Ke X.H. Discrimination of Citrus reticulata Blanco and Citrus reticulata 'Chachi' as well as the Citrus reticulata 'Chachi' within different storage years using ultra high performance liquid chromatography quadrupole/time-of-flight mass spectrometry based metabolomics approach. J. Pharm. Biomed. Anal. 2019;171:218–231. doi: 10.1016/j.jpba.2019.03.056. [DOI] [PubMed] [Google Scholar]
- 58.Khamthong N., Hutadilok-Towatana N. Phytoconstituents and Biological Activities of Garcinia dulcis (Clusiaceae): A Review. Nat. Prod. Commun. 2017;12:453–460. [PubMed] [Google Scholar]
- 59.Hao H.Z., He A.D., Wang D.C., Yin Z., Zhou Y.J., Liu G., Liang M.L., Da X.W., Yao G.Q., Xie W., et al. Antiplatelet activity of loureirin A by attenuating Akt phosphorylation: In vitro studies. Eur. J. Pharmacol. 2015;746:63–69. doi: 10.1016/j.ejphar.2014.10.059. [DOI] [PubMed] [Google Scholar]
- 60.Dong X., Fan Y., Yu L., Hu Y. Synthesis of four natural prenylflavonoids and their estrogen-like activities. Arch. Pharm. 2007;340:372–376. doi: 10.1002/ardp.200700057. [DOI] [PubMed] [Google Scholar]
- 61.Qin Y., Song D., Liao S., Chen J., Xu M., Su Y., Lian H., Peng H., Wei L., Chen K., et al. Isosinensetin alleviates estrogen deficiency-induced osteoporosis via suppressing ROS-mediated NF-kappaB/MAPK signaling pathways. Biomed. Pharmacother. 2023;160 doi: 10.1016/j.biopha.2023.114347. [DOI] [PubMed] [Google Scholar]
- 62.Stossi F., Bolt M.J., Ashcroft F.J., Lamerdin J.E., Melnick J.S., Powell R.T., Dandekar R.D., Mancini M.G., Walker C.L., Westwick J.K., Mancini M.A. Defining estrogenic mechanisms of bisphenol A analogs through high throughput microscopy-based contextual assays. Chem. Biol. 2014;21:743–753. doi: 10.1016/j.chembiol.2014.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ku Y.S., Ng M.S., Cheng S.S., Lo A.W.Y., Xiao Z., Shin T.S., Chung G., Lam H.M. Understanding the Composition, Biosynthesis, Accumulation and Transport of Flavonoids in Crops for the Promotion of Crops as Healthy Sources of Flavonoids for Human Consumption. Nutrients. 2020;12 doi: 10.3390/nu12061717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lee E.R., Kang G.H., Cho S.G. Effect of flavonoids on human health: old subjects but new challenges. Recent Pat. Biotechnol. 2007;1:139–150. doi: 10.2174/187220807780809445. [DOI] [PubMed] [Google Scholar]
- 65.Paterni I., Granchi C., Katzenellenbogen J.A., Minutolo F. Estrogen receptors alpha (ERalpha) and beta (ERbeta): subtype-selective ligands and clinical potential. Steroids. 2014;90:13–29. doi: 10.1016/j.steroids.2014.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kraichely D.M., Sun J., Katzenellenbogen J.A., Katzenellenbogen B.S. Conformational changes and coactivator recruitment by novel ligands for estrogen receptor-alpha and estrogen receptor-beta: correlations with biological character and distinct differences among SRC coactivator family members. Endocrinology. 2000;141:3534–3545. doi: 10.1210/endo.141.10.7698. [DOI] [PubMed] [Google Scholar]
- 67.Katzenellenbogen J.A., Muthyala R., Katzenellenbogen B.S. Nature of the ligand-binding pocket of estrogen rceptor alpha and beta: The search for subtype-selective ligands and implications for the prediciton of estorgenic activity. Pure Appl. Chem. 2003;75:2397–2403. [Google Scholar]
- 68.Atzmony L., Reiter O., Hodak E., Gdalevich M., Mimouni D. Treatments for Cutaneous Lichen Planus: A Systematic Review and Meta-Analysis. Am. J. Clin. Dermatol. 2016;17:11–22. doi: 10.1007/s40257-015-0160-6. [DOI] [PubMed] [Google Scholar]
- 69.Li W.D., Yan C.P., Wu Y., Weng Z.B., Yin F.Z., Yang G.M., Cai B.C., Chen Z.P. Osteoblasts proliferation and differentiation stimulating activities of the main components of Fructus Psoraleae corylifoliae. Phytomedicine. 2014;21:400–405. doi: 10.1016/j.phymed.2013.09.015. [DOI] [PubMed] [Google Scholar]
- 70.Yaylaci E., Onen H.I., Yar Saglam A.S. Phenoxodiol sensitizes metastatic colorectal cancer cells to 5-fluorouracil- and oxaliplatin-induced apoptosis through intrinsic pathway. EXCLI J. 2020;19:936–949. doi: 10.17179/excli2020-2042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Porter K., Fairlie W.D., Laczka O., Delebecque F., Wilkinson J. Idronoxil as an Anticancer Agent: Activity and Mechanisms. Curr. Cancer Drug Targets. 2020;20:341–354. doi: 10.2174/1568009620666200102122830. [DOI] [PubMed] [Google Scholar]
- 72.Jia S., Zhang Y., Yu J. Antinociceptive Effects of Isosakuranetin in a Rat Model of Peripheral Neuropathy. Pharmacology. 2017;100:201–207. doi: 10.1159/000478986. [DOI] [PubMed] [Google Scholar]
- 73.Vangeel L., Benoit M., Miron Y., Miller P.E., De Clercq K., Chaltin P., Verfaillie C., Vriens J., Voets T. Functional expression and pharmacological modulation of TRPM3 in human sensory neurons. Br. J. Pharmacol. 2020;177:2683–2695. doi: 10.1111/bph.14994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhang J., Xie S.A., Wang J., Liu J., Liu Y., Zhou S., Li X., Han L., Pang W., Yao W., et al. Echinatin maintains glutathione homeostasis in vascular smooth muscle cells to protect against matrix remodeling and arterial stiffening. Matrix Biol. 2023;119:1–18. doi: 10.1016/j.matbio.2023.03.007. [DOI] [PubMed] [Google Scholar]
- 75.Kwak A.W., Lee J.Y., Lee S.O., Seo J.H., Park J.W., Choi Y.H., Cho S.S., Yoon G., Lee M.H., Shim J.H. Echinatin induces reactive oxygen species-mediated apoptosis via JNK/p38 MAPK signaling pathway in colorectal cancer cells. Phytother Res. 2023;37:563–577. doi: 10.1002/ptr.7634. [DOI] [PubMed] [Google Scholar]
- 76.Li Y., Zhang Y., Wang R., Wei L., Deng Y., Ren W. Metabolic profiling of five flavonoids from Dragon's Blood in human liver microsomes using high-performance liquid chromatography coupled with high resolution mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017;1052:91–102. doi: 10.1016/j.jchromb.2017.03.022. [DOI] [PubMed] [Google Scholar]
- 77.Hu S.L., Wang K., Shi Y.F., Shao Z.X., Zhang C.X., Sheng K.W., Ge Z.D., Chen J.X., Wang X.Y. Downregulating Akt/NF-kappaB signaling and its antioxidant activity with Loureirin A for alleviating the progression of osteoarthritis: In vitro and vivo studies. Int. Immunopharmacol. 2020;78 doi: 10.1016/j.intimp.2019.105953. [DOI] [PubMed] [Google Scholar]
- 78.Panche A.N., Diwan A.D., Chandra S.R. Flavonoids: an overview. J. Nutr. Sci. 2016;5:e47. doi: 10.1017/jns.2016.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Bolt M.J., Stossi F., Newberg J.Y., Orjalo A., Johansson H.E., Mancini M.A. Coactivators enable glucocorticoid receptor recruitment to fine-tune estrogen receptor transcriptional responses. Nucleic Acids Res. 2013;41:4036–4048. doi: 10.1093/nar/gkt100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Demsar J C.T., Erjavec A., Gorup C., Hocevar T., Multinovic M., Mozina M., Polajnar M., Toplk M., Staric A., Stajdohar M., et al. Orange: Data Mining in Python. J. Mach. Learn. Res. 2013;14:2349–2353. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data reported in this paper will be shared by the lead contact upon request. This paper does not report any original code. Sources are listed in the key resources table for all analysis programs utilized. Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.