

Abstract
Pre-clinical and cell culture evidence supports the role of the ketone beta-hydroxybutyrate (BHB) as an immunomodulatory molecule that may inhibit inflammatory signalling involved in several chronic diseases such as type 2 diabetes (T2D), but studies in humans are lacking. Therefore, we investigated the anti-inflammatory effect of BHB in humans across three clinical trials. To investigate if BHB suppressed pro-inflammatory cytokine secretion, we treated LPS-stimulated leukocytes from overnight-fasted adults at risk for T2D with BHB (Study 1). Next (Study 2), we investigated if exogenously raising BHB acutely in vivo by ketone monoester supplementation (KME) in adults with T2D would suppress pro-inflammatory plasma cytokines. In Study 3, we investigated the effect of BHB on inflammation via ex vivo treatment of LPS-stimulated leukocytes with BHB and in vivo thrice-daily pre-meal KME for 14 days in adults with T2D. Ex vivo treatment with BHB suppressed LPS-stimulated IL-1β, TNF-α, and IL-6 secretion and increased IL-1RA and IL-10 (Study 1). Plasma IL-10 increased by 90 min following ingestion of a single dose of KME in T2D, which corresponded to peak blood BHB (Study 2). Finally, 14 days of thrice-daily KME ingestion did not significantly alter plasma cytokines or leukocyte subsets including monocyte and T-cell polarization (Study 3). However, direct treatment of leukocytes with BHB modulated TNF-α, IL-1β, IFN-γ, and MCP-1 secretion in a time- and glucose-dependent manner (Study 3). Therefore, BHB appears to be anti-inflammatory in T2D, but this effect is transient and is modulated by the presence of disease, glycaemia, and exposure time.
Keywords: beta-hydroxybutyrate, type 2 diabetes, cytokines, ketones
In this series of three human studies, we found that both in vivo and ex vivo treatment of leukocytes with the ketone body beta-hydroxybutyrate induces an anti-inflammatory cytokine milieu. However, this effect is modulated by prolonged (24-h) hyperglycaemia. Furthermore, in vivo this anti-inflammatory effect does not persist once BHB has returned to basal levels, even after repeated exposure to BHB across 14 days.
Graphical Abstract
Graphical Abstract.
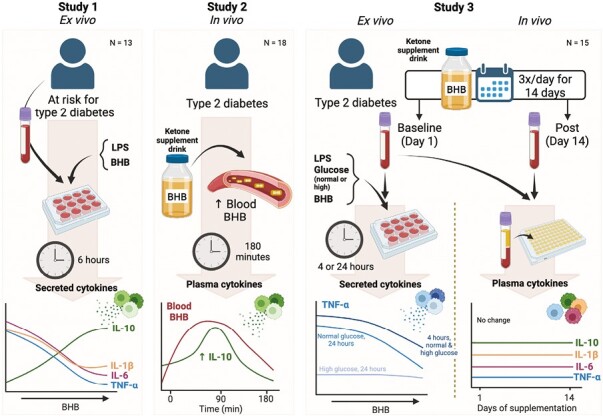
Introduction
The ketone body beta-hydroxybutyrate (BHB) is purported to possess anti-inflammatory and immunomodulatory properties. BHB is produced endogenously by the liver from free fatty acids mobilized under fasted and/or calorie-/carbohydrate-restricted conditions and serves as an important fuel source for the brain, heart, and kidneys [1]. Cell culture and animal studies have produced convincing evidence that BHB has additional immunomodulatory properties, including suppression of inflammatory signalling implicated in chronic low-grade inflammatory diseases such as gout, Alzheimer’s disease, Parkinson’s disease, and obesity [2–5]. Indeed, BHB has been shown to: (i) suppress activation of the NLR-family pyrin domain containing 3 (NLRP3) inflammasome and secretion of the downstream pro-inflammatory cytokine IL-1β [6], thereby preventing gout flares [2]; (ii) increase the expression of anti-oxidant enzymes by acting as a histone deacetylase inhibitor [2, 7]; and (iii) suppress hepatic endoplasmic reticulum stress caused by ageing and hyperlipidaemia [8, 9]. Therefore, BHB may have the potential to attenuate inflammatory signalling involved in the pathogenesis of various inflammatory diseases.
People living with type 2 diabetes (T2D) exhibit widespread immune dysfunction, as immune cells exhibit a persistent state of low-level activation in the basal state resulting in chronic low-grade inflammation [10]. Paradoxically, even though circulating and tissue-resident immune cells are more polarized to pro-inflammatory phenotypes in T2D [10], these cells exhibit impaired defensive immune functions resulting in greater frequency and severity of infections [11, 12]. In addition, classic work implicates the pro-inflammatory cytokine tumour necrosis factor (TNF)-α not only as a marker of chronic inflammation but also as a causal factor in the pathogenesis of insulin resistance [13–17]. There is ongoing interest in using anti-inflammatory therapies to treat T2D and other diseases [18–20], including promising results with the drug anakinra [20], which inhibits interleukin (IL)-1β signalling via the same pathway suppressed by BHB in cell and rodent models [6]. However, the targeted nature of many anti-inflammatory drugs can adversely affect acute defensive immune functions [18, 21, 22]. Therefore, a molecule that can suppress basal inflammation while preserving or enhancing immune defense may be beneficial and complementary to conventional treatment of T2D.
Recent advances in developing oral ketone supplements that raise blood BHB in the absence of dietary changes have enabled researchers to test the direct effects of BHB in humans [23]. Although evidence from cell and animal studies is convincing, the evidence from human studies is somewhat equivocal and nuanced. Previous work by our group found that acute ingestion of a ketone monoester or ketone salt supplement to exogenously raise blood BHB increased markers of NLRP3 activation in young, healthy individuals [24], and Thomsen et al. observed a similar effect with BHB infusion [25]. However, 14 days of repeated ketone monoester supplementation (KME) in individuals at risk for T2D decreased markers of LPS-stimulated, but not unstimulated, NLRP3 inflammasome activation despite no changes in circulating plasma cytokines [5]. Whether the presence of T2D or the level of baseline inflammation influences the immunomodulatory impact of BHB remains to be determined, but it is reasonable to speculate that the anti-inflammatory effects of BHB may be modified under cardiometabolic disease states characterized by chronic inflammation.
To our knowledge, no studies have tested if: (i) immune cells from individuals with T2D respond to BHB in a similar manner to other chronic inflammatory states, (ii) BHB suppresses low-grade inflammation in T2D, and (iii) key immune functions will be preserved or improved following BHB administration in T2D. To test these important questions, we conducted a series of three clinical trials aiming to investigate the direct effect of BHB in separate cohorts of individuals at risk for T2D and those diagnosed with T2D using ex vivo treatment of whole blood cultures (Study 1 and 3), single-dose oral BHB supplementation in vivo (Study 2), and repeated in vivo treatment with oral BHB supplementation across a 14-day period (Study 3).
Results
Study 1: BHB suppresses inflammatory cytokine release from blood leukocytes from individuals at risk for T2D
Previously, our group found that raising blood BHB in vivo for 14 days via KME in individuals at risk for T2D decreased markers of LPS-stimulated, but not unstimulated, NLRP3 inflammasome activation. Additionally, fasted plasma cytokines were unchanged suggesting that an inflammatory stimulus is required for BHB to be anti-inflammatory [26]. Therefore, we hypothesized that ex vivo treatment of LPS-stimulated leukocytes with BHB would suppress the NLRP3-mediated cytokines, IL-1β and IL-18, and other inflammatory cytokines. To test this, we treated whole blood from individuals at risk for T2D with LPS and increasing but physiologically relevant concentrations of BHB (2–10 mM; Fig. 1); a ketogenic diet, short-term fasting, and exogenous ketone supplementation can modestly raise blood BHB to approximately 2 mM or as high as 5 mM depending on the length of the fast or dose of exogenous ketone supplement, and previous rodent studies have shown the most robust effects at 10 mM. IL-1β secretion into culture was decreased with 5 and 10 mM BHB but not 2 mM BHB (main effect of BHB, P < 0.001; 0 vs 2 mM BHB, P = 0.401; 0 vs 5 mM BHB, P < 0.001; 0 vs 10 mM BHB, P = 0.002; Fig. 1A). Accordingly, secretion of IL-1RA mirrored the pattern of IL-1β secretion (main effect of BHB, P = 0.003; 0 vs 2 mM BHB, P = 0.101; 0 vs 5 mM BHB, P = 0.006; 0 vs 10 mM BHB, P < 0.001; Fig. 1B). Interestingly, IL-10 secretion was increased with 5 and 10 mM BHB but not 2 mM BHB (main effect of BHB, P < 0.001; 0 vs 2 mM BHB, P = 0.183; 0 vs 5 mM BHB, P < 0.001; 0 vs 10 mM BHB, P < 0.001; Fig. 1C). This coincided with suppression of IL-6 and TNF-α secretion by increasing BHB concentrations (IL-6: main effect of BHB, P < 0.001, 0 vs 2 mM BHB, P = 0.029, 0 vs 5 mM BHB, P < 0.001, 0 vs 10 mM BHB, P < 0.001; Fig. 1D; TNF-α: main effect of BHB, P = 0.013, 0 vs 2 mM BHB, P = 0.039, 0 vs 5 mM BHB, P = 0.089, 0 vs 10 mM BHB, P = 0.001; Fig. 1E). Collectively, these data suggest that physiological concentrations of BHB were sufficient to suppress pro-inflammatory cytokine secretion while simultaneously enhancing the secretion of the anti-inflammatory cytokines IL-1RA and IL-10 in leukocytes from individuals at risk for T2D under experimental endotoxemia ex vivo.
Figure 1.
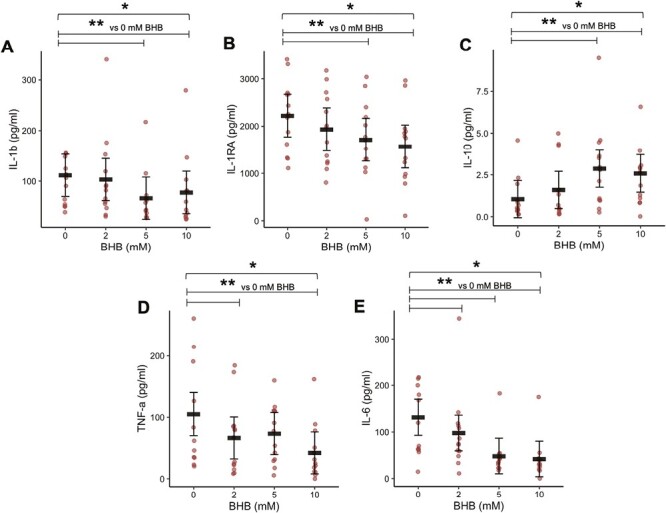
Cytokines secreted from whole blood cultures (Study 1). Cells were treated with LPS (10 ng/ml) and increasing concentrations of BHB for 6 h. Black horizontal bars are estimated marginal means ± 95% CI derived from a linear mixed effects model. Red circles represent individual data. *Main effect of BHB, P < 0.05. **Within-factor comparisons vs 0 mM BHB, P < 0.05. BHB, beta-hydroxybutyrate. N = 13
Study 2: BHB acutely enhances in vivo circulating IL-10 in T2D
Given the apparent need for an inflammatory stimulus for BHB to be anti-inflammatory, we tested the effect of BHB in T2D—a metabolic disease characterized by chronic low-grade inflammation [10]. Therefore, we hypothesized that elevating BHB in vivo via KME ingestion would be anti-inflammatory in T2D under basal conditions.
To test this, we conducted a randomized, placebo-controlled crossover trial in which individuals with T2D consumed a single dose of KME to acutely elevate blood BHB in an attempt to test the direct effects of raising BHB in vivo. Plasma TNF-α, IL-1β, IL-18, IL-1RA, and IL-6 were unchanged by KME (Fig. 2A, C–F). However, consistent with our earlier observations of enhanced IL-10 secretion with BHB ex vivo, plasma IL-10 concentrations were elevated 90 min following consumption of a single dose of KME compared to placebo, with IL-10 returning to baseline within 3 h of the dose (condition × time interaction, P = 0.046; Fig. 2B). This elevation in IL-10 coincided with the peak in blood BHB concentration [mean (SD), baseline, 0.2 (0.1) mM; 90 min, 1.7 (0.6) mM; 180 min, 0.8 (0.4) mM] resulting from KME (full BHB data reported by Falkenhain et al. [27]). It must be noted that an influential data point at 90 min in the KME condition was identified by a Cook’s distance value above the defined threshold. When this data point was removed from the data set, the certainty of the condition × time interaction was reduced such that P = 0.069, and statistical significance was lost. Due to the fact that the trend for an interaction remained, we feel that this influential data point likely represented a true but somewhat exaggerated response such that this data point remains represented in Fig. 2B and the estimated marginal means reported in Supplementary Table S2. Therefore, while these data should be interpreted with a degree of caution, it is likely that elevated BHB acutely increased the anti-inflammatory cytokine IL-10 in circulation in vivo in humans with T2D suggesting that BHB could exert modest anti-inflammatory effects under inflammatory states.
Figure 2.
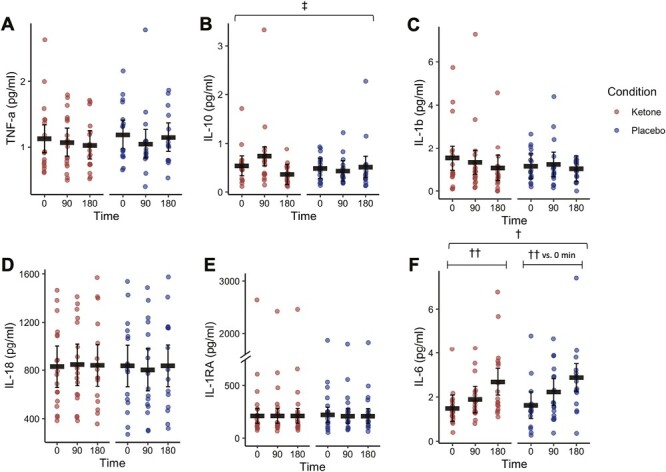
(A–F) Circulating plasma cytokines following an acute dose of a KME or placebo supplement (Study 2). Red circles represent individual data from the KME condition. Blue circles represent individual data from the placebo condition. Black horizontal bars represent estimated marginal means ± 95% CI derived from a linear mixed effects model. ‡Interaction effect, glucose × BHB, P < 0.05. †Main effect of time, P < 0.05. ††Within-factor comparison (time vs. 0 minutes, P < 0.05
Study 3: Effect of ex vivo treatment with BHB in normo- and hyperglycaemia
Effect of BHB and glucose on the IL-10-TNF-α signalling axis in an ex vivo model
Given that T2D is defined by chronically elevated blood glucose levels, we investigated how the acute anti-inflammatory effect of BHB might interact with glucose concentrations in LPS-stimulated leukocytes. Whole blood samples collected from participants with T2D prior to any interventions were incubated for either 4 or 24 h in the presence of either normal (5 mM) or high (15 mM) glucose concentrations and stimulated with LPS. Following 4 h of treatment, LPS-stimulated TNF-α secretion was suppressed by BHB (main effect of BHB, P < 0.001; 1 vs 0 mM BHB, P = 0.670; 2 vs 0 mM BHB, P = 0.815; 5 vs 0 mM BHB, P = 0.052; Fig. 3A) which was similar to the findings from the Study 1 culture (Fig. 1D). Interestingly, the anti-inflammatory effect of BHB was differentially impacted by 24 h of exposure to normal or high glucose (main effect of glucose, P < 0.001; BHB × glucose interaction, P = 0.051; main effect of BHB, P = 0.058; Fig. 3B). In the presence of normal glucose, 24 h of treatment with BHB suppressed LPS-stimulated TNF-α. However, 24 h of exposure to high glucose appeared to impair the secretion of LPS-stimulated TNF-α relative to normal glucose in the absence of BHB, such that there was no further suppression by BHB (Fig. 3B). Interestingly, these effects on LPS-stimulated TNF-α occurred in the absence of any changes in IL-10 secretion following both 4- and 24-h treatment (Fig. 3Cand D), suggesting that the effects of BHB on LPS-stimulated TNF-α were not mediated by increased secretion of LPS-stimulated IL-10. Although it is possible that the ability of IL-10 to inhibit TNF-α secretion (i.e. IL-10 “action”) may have been modulated independent of IL-10 concentrations [28, 29], we did not measure this outcome in these cultures and this remains speculative.
Figure 3.
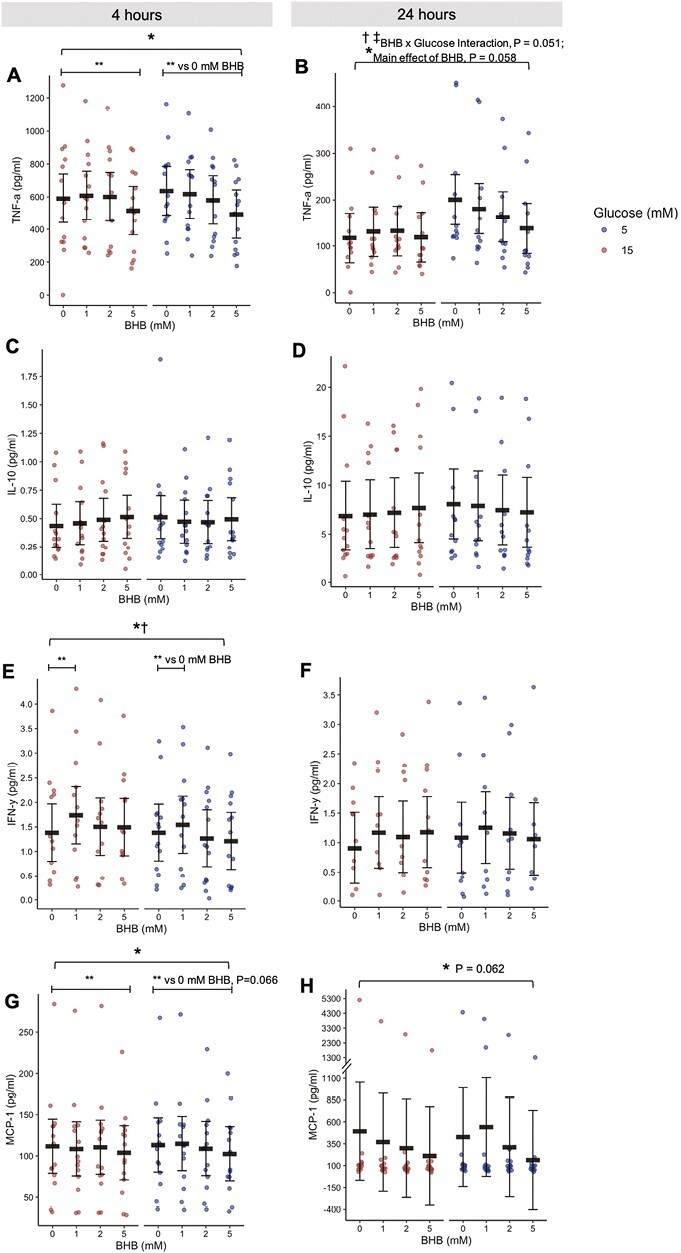
(A–H) Cytokines secreted into culture after 4 (A, C, E, & G) or 24 (B, D, F, & H) h of treatment with BHB (0, 1, 2, or 5 mM) and 15 mM glucose (representing hyperglycemia) or 5 mM glucose (representing normoglycemia) (Study 3). Red circles represent 15 mM glucose. Blue circles represent 5 mM glucose. Circles are individual data. Black horizontal bars represent estimated marginal means ± 95% CI derived from a linear mixed effects model. *Main effect of BHB, P < 0.05. †Main effect of glucose, P < 0.05. ‡Interaction effect, glucose × BHB, P < 0.05. **Within-factor comparison (BHB vs. 0 mM BHB), P < 0.05. Data are estimated marginal means. BHB, beta-hydroxybutyrate. N = 14
IFN-γ was increased in the presence of 1 mM BHB (main effect of BHB, P = 0.020; 1 mM BHB vs 0, P = 0.017; Fig. 3E), but not in the presence of 2 or 5 mM BHB. Additionally, LPS-stimulated IFN-γ was higher in the presence of high glucose (main effect of glucose, P = 0.017) although the effects of both BHB and glucose were lost after 24 h (Fig. 3F). We also observed that secretion of LPS-stimulated monocyte chemoattractant protein (MCP)-1, a chemokine implicated in insulin resistance via promotion of tissue infiltration by monocytes [10], was suppressed by 4 h of BHB treatment (main effect of BHB, P = 0.011; 1 vs 0 mM BHB, P = 0.439; 2 vs 0 mM BHB, 0.768; 5 vs 0 mM BHB, P = 0.066; Fig. 3G) but this difference was less pronounced following 24 h (main effect of BHB, P = 0.062; 1 vs 0 mM BHB, P = 0.445; 2 vs 0 mM BHB, 0.232; 5 vs 0 mM BHB, P = 0.084; Fig. 3H). Taken together, these cytokine data support the notion that BHB is an immunomodulatory signal which can acutely suppress LPS-stimulated inflammatory signalling in the presence of normoglycaemia and acute but not chronic hyperglycaemia.
Effect of BHB and glucose on defensive immune functions
Paradoxically, immune cells in T2D exhibit deficits in their ability to mount an effective defensive response to pathogens, as evidenced by defects in NADPH oxidase-mediated oxidative burst [30–32] and blunted phagocytosis [33–35]. Therefore, we aimed to determine if BHB would improve these immune functions and if glucose would modulate any potential effect. To test this, we used the same culture conditions as used for the cytokine cultures to test the acute and more prolonged effect of normal and high glucose and physiological concentrations of BHB.
Following 4 h of treatment granulocytes generated a more robust oxidative burst response with increasing BHB (main effect of BHB, P < 0.001; 5 mM BHB vs 0; P < 0.001; Fig. 4D), although the monocyte response was unchanged (Fig. 4C). Following 24 h of treatment this effect on granulocytes was somewhat lost, however, this appeared due to a differential effect of glucose on oxidative burst such that increasing BHB may have enhanced oxidative burst in the presence of hyper- but not normoglycaemia (main effect of glucose, P = 0.028; Fig. 4H). Interestingly, after 24 h BHB decreased the amount of ROS produced by oxidative burst monocytes in the presence of normoglycaemia, but slightly increased it in the presence of hyperglycaemia (BHB × glucose interaction, P = 0.034; Fig. 4G). These effects occurred despite no changes in the proportions of monocytes and granulocytes performing oxidative bursts (Fig. 4A and B, E and F).
Figure 4.
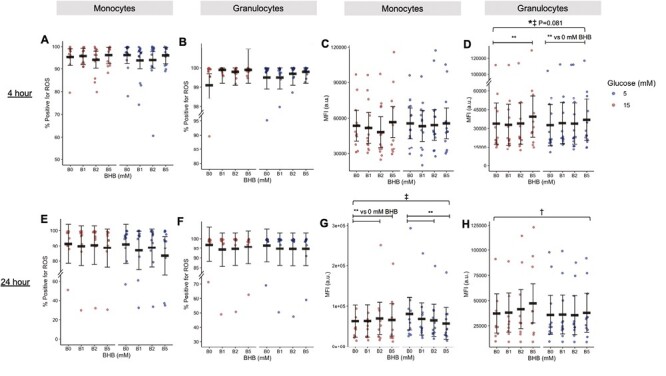
(A–H) Oxidative burst quantified by flow cytometry following 4 (A–D) or 24 h (E–H) of treatment with BHB (0, 1, 2, or 5 mM) and 15 mM glucose (representing hyperglycaemia) or 5 mM glucose (representing normoglycemia) in monocytes (A, C, E, and G) or granulocytes (B, D, F, and H) (Study 3). Blue circles represent 5 mM glucose. Circles are individual data. Black horizontal bars represent estimated marginal means ±95% CI derived from a linear mixed effects model. *Main effect of BHB, P < 0.05. †Main effect of glucose, P < 0.05. ‡Interaction effect, glucose × BHB, P < 0.05. **Within-factor comparison (BHB vs. 0 mM BHB), P < 0 .05. Data are estimated marginal means. MFI, median fluorescence intensity; BHB, beta-hydroxybutyrate. N = 13
Exposure of monocytes to 24 h of hyperglycaemia did suppress the proportion of cells that could effectively perform phagocytosis relative to monocytes in normoglycaemic conditions with no apparent effects of BHB (main effect of glucose, P < 0.001; Fig. 5E). However, treatment with BHB was unable to induce an improvement in the phagocytic capacity of monocytes or granulocytes regardless of treatment duration or BHB concentration (Fig. 5A–D, F–H). Our results indicate that BHB may exert a modest immune function-enhancing effect on oxidative burst but cannot rescue hyperglycaemia-blunted phagocytic function.
Figure 5.
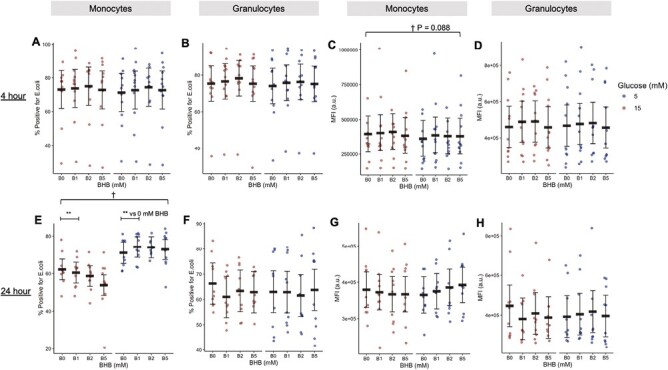
Phagocytosis quantified by flow cytometry following 4 (A–D) or 24 h (E–H) of treatment with BHB (0, 1, 2, or 5 mM) and 15 mM glucose (representing hyperglycaemia) or 5 mM glucose (representing normoglycaemia) in monocytes (A, C, E, and G) or granulocytes (B, D, F, and H) (Study 3). Red circles represent 15 mM glucose. Blue circles represent 5 mM glucose. Circles are individual data. Black horizontal bars represent estimated marginal means ± 95% CI derived from a linear mixed effects model. †Main effect of glucose, P < .05. **Within-factor comparison (BHB vs. 0 mM BHB), P < 0.05. Data are estimated marginal means. MFI, median fluorescence intensity; BHB, beta-hydroxybutyrate. N = 12
Study 3: Effect of repeated in vivo treatment with BHB in T2D
Next, we asked if the effects of BHB that were found in ex vivo cultures would be replicable in vivo when individuals with T2D were exposed to repeated increases in blood BHB via ingestion of KME over a longer period of time. To test this, we conducted a randomized, placebo-controlled crossover trial in which individuals with T2D consumed a KME or placebo thrice daily for 14 days. A single dose of KME increased blood BHB to 1.4 (1.1) mM [mean (SD)] within 15 min and to 1.8 (0.7) mM within 30 min.
Leukocyte subsets
First, we looked at the monocyte and T-cell subsets present in circulation as these are the major producers of the cytokines that we planned to quantify. However, we did not find any differences in the proportions of classical, non-classical, or intermediate monocytes, pan-T cells, CD4+ Th cells, Treg cells, or Th subsets (Supplementary Table S6). Therefore, repeated KME ingestion across 14 days did not influence basal immune cell polarization.
Fasting plasma cytokines
Next, we quantified fasting plasma cytokines. Similar to Walsh et al. [5], we also found that fasting plasma cytokines (TNF-α, IFN-γ, IL-1RA, IL-1β, IL-6, and MCP-1) and CRP following 14 days of KME were largely unchanged (Supplementary Table S7). Therefore, our results indicate that in vivo treatment with BHB in T2D may acutely increase IL-10 as long as BHB is elevated (Study 2), but this effect was not preserved in the fasted state even following repeated in vivo exposure to BHB (Study 3).
IL-10 Function
Plasma cytokines do not always capture cytokine action at the cellular level [28, 29]. Given the ex vivo impact of BHB treatment of whole blood cultures of reducing TNF-α (Fig. 1D, 3A, 3B), and the in vivo effect of raising BHB augmenting IL10 (Fig. 2), we wondered if BHB may be directly modulating IL-10 function. IL-10 function was characterized as the ability to increase concentrations of ex vivo IL-10 treatment to phosphorylate STAT3 (the key intracellular mediator of IL-10 action; main effect of IL-10, P < 0.001; Fig. 6A and B) and suppress LPS-stimulated TNF-α secretion in whole blood cultures from fasting blood samples before and after 14 days of KME or placebo supplementation (main effect of IL-10, P < 0.001; Fig. 6C).
Figure 6.
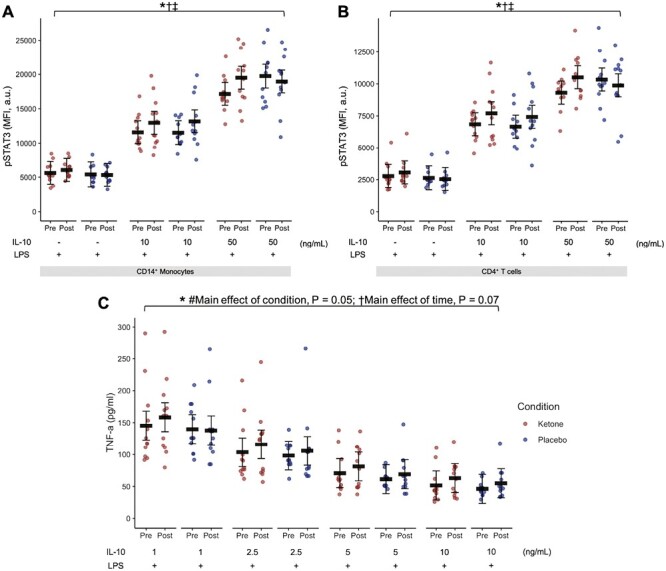
(A and B) Median fluorescence intensity (MFI) of phosphorylated STAT3 (pSTAT3) by IL-10 in LPS-stimulated whole blood cultures collected before and after 14 days of in vivo KME or placebo in the fasted state (Study 3). (C): TNF-α secretion in response to IL-10 treatment in LPS-stimulated whole blood cultures collected before and after 14 days of in vivo KME or placebo in the fasted state. Red circles represent the KME condition. Blue circles represent the placebo condition. Circles represent individual data. Black horizontal bars are estimated marginal means ± 95% CI derived from a linear mixed effects model. *Main effect of IL-10, P < 0.05. †Main effect of time, P = 0.07. #Main effect of condition, P = 0.05. ‡Interaction effect, condition × time, P < 0.05. BHB, beta-hydroxybutyrate; a.u., arbitrary units. N = 12
Consistent with our hypothesis of a dissociation between plasma cytokine concentrations and cellular cytokine action, CD14+ monocyte pSTAT3 in response to IL-10 treatment ex vivo was enhanced by KME (condition × time interaction, P = 0.049; Fig. 6A) despite no clear changes in plasma IL-10 concentration (Supplementary Table S7) or changes in IL-10 receptor expression on CD14+ monocytes (Supplementary Table S9). Similarly, pSTAT3 was also increased in response to ex vivo IL-10 treatment in CD4+ T cells (condition × time interaction, P = 0.007; Fig. 6B) in the absence of changes in IL-10 receptor expression on CD4+ T cells (Supplementary Table S9). These findings indicate that BHB may have increased the ability of IL-10 to activate intracellular signalling in both innate and adaptive immune cells, potentially contributing to the overall immunomodulatory effects of BHB.
As expected, IL-10 treatment of whole blood cultures inhibited TNF-α secretion (main effect of IL-10, P < 0.001; Fig. 6C). Interestingly, this improvement in IL-10 sensitivity did not translate to a reduction in LPS-stimulated TNF-α secretion mediated by KME supplementation (condition × time interaction, P = 0.505, Fig. 6; Supplementary Table S10). Therefore, KME supplementation increased IL-10 sensitivity as quantified by IL-10-mediated phosphorylation of STAT3, however, this did not impact TNF-α secretion in the presence of experimental endotoxemia.
Basal immune functions
Finally, we asked if measures of immune function would be preserved or enhanced following repeated in vivo exposure to BHB. However, BHB did not impact LPS-stimulated oxidative burst following 14 days of exogenous BHB supplementation (Supplementary Table S6). Additionally, repeated KME did not impact the proportion of cells performing phagocytosis (Supplementary Table S6), but KME increased the number of Escherichia coli that were phagocytosed by monocytes (condition × time interaction, P = 0.034; Supplementary Table S6). A similar trend occurred in granulocytes although this effect did not reach statistical significance (condition × time, P = 0.079; Supplementary Table S6). In combination with the ex vivo cultures, these data suggest that any immune-enhancing effect of BHB is primarily acute in nature, and likely more effective at higher physiological concentrations.
Discussion
The ketone body BHB is a purported immunomodulatory molecule. Cell and animal studies have shown evidence that BHB suppresses inflammatory signalling present in chronic, low-grade inflammation that is involved in certain disease states including gout and obesity [2, 5]. Here, we report evidence that direct ex vivo treatment with physiological concentrations of BHB robustly suppresses the secretion of the pro-inflammatory cytokines TNF-α, IL-1b, and IL-6 while concurrently increasing the secretion of the anti-inflammatory cytokines IL-10 and IL-1RA in immune cells from humans at risk for T2D (Study 1, Fig. 1), but these apparent anti-inflammatory effects of BHB appear to be muted in T2D (Study 3, Fig. 3). We also show that direct in vivo treatment with BHB in T2D acutely increases plasma IL-10 (Study 2, Fig. 2) but any anti-inflammatory effects do not appear to persist after 14 days of repeated in vivo exposure in T2D (Study 3, Supplementary Table S7. Finally, these data show that despite any anti-inflammatory effect of BHB, in vivo and ex vivo treatment with BHB does not compromise or may even enhance certain markers of immune function (Study 3, Figs. 4 and 5).
Inflammation is involved in the development of insulin resistance even prior to the clinical presentation of T2D [10]. Indeed, classic work has demonstrated the causal and detrimental effect of the pro-inflammatory cytokine, TNF-α, and its downstream signalling components on glycemic control and effective insulin signalling [13–15], such that neutralization of TNF-α results in an immediate increase in glucose infusion rate during a hyperinsulinemic–euglycemic clamp [13]. Therefore, we were interested to observe that BHB suppressed TNF-α and other inflammatory cytokine secretion in ex vivo cultures of immune cells derived from individuals with obesity at high risk for T2D (Fig. 1). Additionally, physiologically relevant concentrations of BHB increased IL-10, a potent anti-inflammatory cytokine that directly antagonizes TNF-α [36, 37]. Furthermore, when these cultures were repeated with immune cells derived from individuals with T2D, a similar, albeit less robust, effect of BHB on TNF-α secretion was observed. We were also unable to replicate an anti-inflammatory effect of BHB on other cytokines with the exception of MCP-1 and IFN-γ (Fig. 3). Therefore, we speculate that immune cells from individuals with T2D may be more resistant to the anti-inflammatory effects of BHB, with data comparing 5 mM to 15 mM glucose culture conditions suggesting that high glucose—or the resultant changes in immune cell phenotype arising from chronic exposure to hyperglycaemia in T2D—partially moderates this effect.
Previous work by our group did not observe any effect of repeated in vivo exposure to BHB for 14 days on plasma cytokines in individuals with obesity [5]. However, whole blood cultures of NLRP3 inflammasome activation from this previous study suggested that augmented basal inflammation might be needed to be able to detect a potential effect [5]. Basal inflammation tends to be higher in T2D, therefore we hypothesized that BHB may be more anti-inflammatory in vivo in this population. In line with our hypothesis, acutely elevated BHB increased plasma IL-10 concentrations in T2D (Fig. 2). However, this anti-inflammatory effect was short-lived, such that IL-10 decreased concurrently with the decrease in blood BHB at 180 minutes post-supplement ingestion. In line with this interpretation, repeated in vivo exposure to BHB for 14 days in individuals with T2D did not exert any persistent effect on fasting plasma cytokines (Supplementary Table S7). The apparent absence of a robust and preserved anti-inflammatory effect in people with T2D despite improvements in cytokine secretion profiles in people with obesity suggests that BHB may be more effective at lowering inflammation preceding the development of T2D as opposed to reversing disease-related inflammation. Future studies should test this speculation in other populations ‘at risk’ for the development of T2D or other chronic inflammatory states to identify the ideal time-point to maximize BHB’s therapeutic potential. Taken together, these results suggest that any in vivo anti-inflammatory effect of BHB—as captured by changes in circulating cytokine concentrations—is subtle and short-lived.
Given the importance of inflammation in T2D pathology, anti-inflammatory drugs have been investigated as an alternative approach to treating T2D to address underlying dysfunctions in the immune system [19]. Treatment with IL-1 receptor blockade, IL-1β antagonists, and salsalate, which inhibit IκB kinase (IKK)β and NF-κB signalling, have generally been found to lower A1c and fasting plasma glucose [19]. However, a problematic side effect of anti-inflammatory drugs, particularly those targeting the IL-1 pathway, includes greater susceptibility to infections. A meta-analysis of the safety of IL-1 antagonist in patients with rheumatoid arthritis found an 8.9% risk of infection [18] while a safety trial of LY2189102 in patients with T2D found that 26.6% of patients acquired infections throughout the 12-week trial compared to 18.5% of patients allocated to the placebo group [21]. Additionally, the CANTOS trial reported a higher incidence of fatal infection with Canakinumab treatment [22]. Therefore, we tested the effects of BHB on select immune functions known to be defective in T2D. Ex vivo treatment with BHB appeared to enhance oxidative burst in granulocytes, although the effect on monocytes was less clear (Fig. 4). Repeated in vivo treatment with BHB for 14 days did not enhance or impair oxidative burst. However, we observed a more robust oxidative burst response following both conditions likely due to the improved quality of the controlled diet provided to participants throughout both interventions. Ex vivo exposure to hyperglycaemia for 24 h impaired phagocytosis in line with previous reports [30, 31]. This effect may be due to the glycation of complement or Fcγ receptors in the presence of hyperglycaemia [34, 35]. However, BHB did not rescue this or significantly alter phagocytic capacity in any other conditions (Fig. 5). Interestingly, repeated in vivo treatment with BHB for 14 days did enhance the phagocytic capacity of immune cells (Supplementary Table S6). Taken together, these results indicate that the anti-inflammatory effect of BHB does not impair select immune functions, and may, in fact, enhance oxidative burst. If there are potential anti-inflammatory effects of supplementing with BHB or ketogenic interventions in humans with T2D, these data are important as they indicate that innate immune function does not appear to be impaired by BHB.
The studies reported in the present paper are the first, to our knowledge, to directly test the immunomodulatory effects of BHB in the absence of any other dietary changes in individuals with obesity and T2D. In addition, the ex vivo BHB treatments of the same immune cells conducted in parallel to the clinical trials provide additional insight into the nuanced in vivo effects of BHB and highlight the complexity of translating highly controlled in vitro and animal studies into human trials. An important limitation of our 14-day in vivo trial was the lack of collection of blood BHB concentrations resulting from the KME throughout the duration of the trial. Therefore, we cannot be certain that participants experienced exposure to BHB comparable to the concentrations used in our ex vivo cultures. However, this limitation was necessary to maintain researcher and participant blinding throughout the trial and we did confirm compliance by logbook and return of empty supplement bottles. To circumvent this, we characterized the acute blood BHB response to a single dose of the KME prior to randomization, demonstrating that the supplement effectively induced moderate ketosis (1.8 (0.7) mM) within 30 min of ingestion. In addition, the BHB kinetics of the KME are well-characterized and repeatable across populations [38–43].
In summary, these data support the notion that BHB exerts nuanced and diverse immunomodulatory functions in human immune cells and demonstrate that these effects can be elucidated with in vivo and ex vivo experiments. Moreover, these data indicate that any immunomodulatory effects of BHB in vivo appear lost once BHB concentrations return to basal levels and that the presence of an inflammatory stimulus may be necessary to observe the influence of BHB on immune cells. Considering the seemingly differential impact of BHB depending on the presence of obesity, T2D, and normo- or hyperglycaemia, future research should aim to further investigate the mediating factors responsible for these differences in humans. The impact of BHB may depend on T2D status, glucose levels, and the presence of (other) inflammatory stimuli. Finally, it remains to be determined if the immunomodulatory effects of BHB occur through a central signalling or metabolic pathway, or if diverse mechanisms are responsible for these differential effects.
Materials and methods
Study 1: treatment of whole blood cultures with BHB in individuals at risk for T2D
This paper reports tertiary outcomes from a previously published randomized, double-blind, placebo-controlled crossover trial in which the primary purpose was to assess the effect of thrice-daily pre-meal KME for 14 days on glycemic control in individuals at risk for T2D (registered on ClinicalTrials.gov, NCT03817749; approved by the University of British Columbia Clinical Research Ethics Board; H18-02930). The primary and secondary outcomes and associated methods from this trial are published in detail elsewhere [5, 26]. Here we report tertiary outcomes on the effect of ex vivo treatment with BHB on inflammatory cytokine secretion in whole blood cultures using the fasted baseline samples from individuals at risk for T2D.
Participants
Participants at risk for T2D (N = 14; 4 males, 10 females) were recruited between March 2019 to January 2020 and screened by phone for eligibility. Eligible volunteers were invited to visit the laboratory for further information about the study. The inclusion and exclusion criteria are reported in Supplementary Materials. Participant characteristics are reported in Table 1.
Table 1.
Participant baseline characteristics from Studies 1 to 3
| Study 1 | Study 2 | Study 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Total | Female | Male | Total | Female | Male | Total | Female | Male | |
| N (%) | 14 (100) | 10 (71) | 4 (29) | 18 (100) | 7 (39) | 11 (61) | 15 (100) | 6 (40) | 9 (60) |
| Age (years) | 56 (12) | 59 (8) | 54 (18.4) | 62 (8) | 61 (8) | 63 (8) | 56.5 (9.1) | 57.5 (8.5) | 55.8 (9.9) |
| Anthropometric characteristics | |||||||||
| Body weight (kg) | 95.1 (22.2) | 91.5 (25.2) | 99.7 (8.8) | 94.0 (25.4) | 93.2 (30.8) | 94.4 (23) | 99.8 (16.7) | 94.4 (12.8) | 103.3 (18.7) |
| Fat mass (%) | 40.8 (9.5) | 44.8 (8.1) | 32.6 (1.2) | 33.3 (9.7) | 40.3 (7.5) | 29.6 (14.7) | 38.3 (8.9) | 47.4 (2.5) | 32.3 (5.7) |
| BMI (kg/m2) | 33.8 (6.9) | 34.1 (8.2) | 32.7 (1.5) | 32.4 (8.1) | 33.6 (9.9) | 29.6 (8.9) | 34.5 (4.8) | 36.8 (5.0) | 32.9 (4.3) |
| Waist circumference (cm) | 109.2 (17.1) | 108.4 (20.4) | 108.8 (1.7) | 109.8 (17.7) | 110.0 (16.7) | 109.8 (19.1) | 112.6 (12.2) | 110.7 (8.9) | 113.9 (14.4) |
| Systolic blood pressure (mmHg) | 133 (18) | 134 (19) | 137 (17) | 130 (16) | 133 (17) | 128 (16) | 143 (18) | 135 (25) | 147 (13) |
| Diastolic blood pressure (mmHg) | 81 (11) | 84 (10) | 83 (14) | 77 (8) | 75 (8) | 78 (9) | 90 (9) | 89 (12) | 90 (8) |
| Clinical characteristics | |||||||||
| Duration of T2D (years) | N/A | N/A | N/A | 8.5 (3.6) | 9.0 (3.5) | 8.2 (3.7) | 6.5 (5.3) | 6.1 (4.8) | 6.7 (5.9) |
| HbA1c (%) | N/A | N/A | N/A | 7.0 (0.6) | 7.0 (0.7) | 7.0 (0.5) | 7.6 (1.8) | 7.3 (1.3) | 7.8 (2.1) |
Abbreviations: BMI: body mass index; HbA1c: haemoglobin A1c; T2D: type 2 diabetes.
Data are means (SD).
Study visit procedures
Prospective participants were invited to visit the laboratory for familiarization with the study procedures and confirmation of eligibility. Participants arrived at the laboratory in the morning for their baseline and post-testing visits having fasted overnight (≥10 h), having consumed a standardized diet, avoided alcohol, and avoided exercise for the previous 24 h. Anthropometrics were measured, and a blood draw was taken from the antecubital vein by standard venipuncture.
Cytokine secretion from lipopolysaccharide (LPS)-stimulated whole blood cultures
EDTA blood (BD Vacutainer) from baseline was diluted 1:2 with warm RPMI (Cat.No.10-0430-CV; Corning; final concentration 5 mM glucose). LPS (10 μg/ml; Cat. No. L5418; Sigma–Aldrich) and BHB ((R)-3-hydroxybutyric acid; Cat. No. 54920-1G-F; Sigma–Aldrich) or RPMI vehicle control were aliquoted and warm diluted blood was added (final concentrations: LPS, 10 ng/ml; BHB, 2, 5, or 10 mM). Following incubation (37°C; 5% CO2) for 6 h, supernatants were collected and stored at −70°C. Supernatants were batch analyzed in technical replicates for TNF-α, IL-10, IL-1β, IL-18, IL-1 receptor agonist (IL-1RA), and IL-6 using a U-PLEX Metabolic Group 1 human assay kit (MesoScale Discovery). The lower and upper limits of detection (LLOD; ULOD) are available on the manufacturer’s website.
Study 2: acute ketone supplementation in T2D
This paper reports secondary outcomes from a randomized, double-blind, placebo-controlled crossover trial assessing the acute effect of one dose of KME on blood glucose in individuals with T2D (registered on ClinicalTrials.gov, NCT04194450; approved by the University of British Columbia Clinical Research Ethics Board; H19-02947). The primary outcomes from this trial have been reported elsewhere [27]. Here we report the in vivo cytokine response to elevated blood BHB by ingestion of one dose of KME. This trial involved three lab visits: one familiarization visit and two experimental trials (placebo or KME) which were conducted in a randomized counterbalanced double-blind fashion. Following the familiarization visit, participants were randomly assigned to the KME or placebo condition using the online generator (‘Sealed Envelope’).
Participants
Participants with T2D (N = 18; 11 males, 7 females) were recruited between December 2019 to November 2021 and screened by phone for eligibility. Eligible volunteers were invited to visit the laboratory for further information. The inclusion and exclusion criteria are reported in Supplementary Materials. Participant characteristics are reported in Table 1.
Study visit procedures
Prospective participants were invited to visit the laboratory for familiarization with the study procedures and confirmation of eligibility. On the morning of the experimental visit, participants arrived and fasted at the laboratory (≥10 h overnight) and had consumed a standardized diet, avoided alcohol, and avoided exercise for the previous 24 h. Anthropometrics were measured and an intravenous catheter was inserted into the antecubital vein by standard venipuncture for repeated blood draws. Blood samples were collected (0 min), following which participants ingested a dose of KME or placebo. Blood samples were collected again at 30, 60, 90, 120, 150, and 180 min with plasma cytokines quantified at 0-, 90- and 180-min timepoints.
Supplement details
The KME contained a single 0.3 g/kg dose of (R)-3-hydroxybutyl (R)-3-hydroxybutyrate ketone monoester (ΔG®, HVMN; 0.4 g/ml BHB), natural flavouring, and stevia. The flavour-matched placebo supplement was formulated based on a recipe provided by HVMN and included the same natural flavouring, stevia, Bittrex stock (denatonium benzoate powder in water), and arrowroot powder to match the viscosity of the active supplement. The KME and placebo supplements were aliquoted into opaque 30 ml bottles by the researchers and labelled A or B by a researcher who was not involved in the study. The independent researcher maintained the blinding code for the duration of the trial such that the researchers involved in conducting the trial were blinded throughout the entirety of data collection and statistical analyses.
Plasma cytokines
Blood collected into EDTA tubes (BD Vacutainer) was centrifuged, aliquoted, and stored at −70°C. Plasma samples were batch analyzed in technical replicates for TNF-α, IL-10, IL-1β, IL-18, IL-1RA, and IL-6 using the V-PLEX Proinflammatory Panel 1 human assay kit (MesoScale Discovery). The LLOD’s and ULOD’s are publicly available on the manufacturer’s website.
Study 3: 14-day ketone trial in T2D
This paper reports secondary outcomes from a larger randomized, double-blind, placebo-controlled crossover trial in which the primary purpose was to assess the effect of thrice-daily pre-meal KME for 14 days on glycemic control in individuals with T2D (registered on ClinicalTrials.gov, NCT05155410; approved by the University of British Columbia Clinical Research Ethics Board; H21-01762). The primary outcomes of this trial have been reported elsewhere [27]. Here we report the immunomodulatory effect of BHB in vivo by use of repeated KME, and the direct effect of BHB in isolation on cultured blood leukocytes in an ex vivo model. This trial involved 29 days, including five lab visits and two 14-day supplementation periods (KME or placebo).
Participants
Participants with T2D (N = 15; 9 males, 6 females) were recruited between February and October 2022. Interested volunteers were screened by phone, and eligible volunteers were invited to visit the laboratory for further information about the study. The inclusion and exclusion criteria are reported in Supplementary Materials. Participant characteristics are reported in Table 1. Following the familiarization visit, participants were randomly assigned to the KME or placebo condition using variable permuted block sizes of 2 and 4 in R and revealed to the researchers in a sealed envelope as A-B or B-A.
Study visit procedures
Eligible participants were invited to visit the laboratory for familiarization with the study procedures and confirmation of eligibility. Participants arrived at the laboratory in the morning for their baseline and post-testing visits having fasted overnight (≥10 h), having consumed a standardized diet, avoided alcohol, and avoided exercise for the previous 24 h. Anthropometrics were measured, and a baseline blood draw was taken from the antecubital vein by standard venipuncture. Supplements (KME or placebo) were consumed 15 min prior to each meal for 14 days. Following the completion of the 14-day intervention, participants returned to the lab in the fasted state the following morning during which all ‘Post’ measures were collected. Researchers connected with participants regularly via email, phone call, or text to address participant concerns/questions and to ensure compliance.
Dietary control
Participants followed a controlled diet for each study condition. Energy intake was estimated using the Harris–Benedict equation (activity factor of 1.4) and diets were designed to meet the criteria of the Canadian dietary guidelines for T2D with approximately 45–65% of energy derived from carbohydrates, 20–35% from fat, and 10–35% from protein. All food was provided by a local meal preparation company.
Supplement details
The KME used in this trial was an (R)-3-hydroxybutyl (R)-3-hydroxybutyrate ketone monoester (KE4, KetoneAid) and contained 15 g of BHB per 30 ml dose (0.5 g/ml), water, allulose, stevia, natural flavouring, and potassium sorbate. The flavour-matched placebo supplement was formulated based on a recipe provided by KetoneAid and included the same natural flavouring, stevia, denatonium benzoate powder (0.125 mg/ml), allulose, and potassium sorbate. The change in the KME used for Study 3 was made to improve the palatability of the supplement and consequently the adherence to the intervention. However, the active ingredient in the supplement used in Study 2 and 3 were the same. The KME and placebo supplements were aliquoted into opaque 30 ml bottles by the researchers and labelled A or B by a researcher who was not involved in the study. The independent researcher maintained the blinding code for the duration of the trial such that the researchers involved in conducting the trial were blinded throughout the entirety of data collection and statistical analyses.
Immunological outcomes
Whole blood cultures
To explore cytokine responses to elevated BHB and glucose, EDTA blood from the baseline of the first condition was collected and diluted 1:2 with warm RPMI 1640 (Cat.No.10-0430-CV; Corning) in 5 or 15 mM glucose, which were chosen to mimic the glycemic conditions that leukocytes might regularly be exposed to in non-T2D and T2D. BHB (100 mM; (R)-3-Hydroxybutyric acid; Cat. No. 54920-1G-F; Sigma–Aldrich) and warm diluted blood were added to make final concentrations of 1, 2, or 5 mM BHB, similar to what would be experienced during nutritional ketosis. Following incubation in 37°C and 5% CO2 for 2 or 22 h, LPS (2000 ng/ml; Cat. No. L5418; Sigma–Aldrich) was added (final concentration, 50 ng/ml). Cultures were incubated for 2 h. Culture supernatants were centrifuged, aliquoted, and stored at −70°C.
To determine oxidative burst and phagocytosis in response to elevated BHB and glucose, sodium heparin blood (BD Vacutainer) from the baseline of the first condition was collected and diluted 1:2 with warm RPMI 1640 (Cat. No. 10-0430-CV; Corning; 5 or 15 mM glucose). BHB (100 mM; (R)-3-hydroxybutyric acid; Cat. No. 54920-1G-F; Sigma–Aldrich) and warm diluted blood were added to make final concentrations of 1, 2, or 5 mM BHB. Following incubation at 37°C and 5% CO2 for 4 or 24 h, samples were centrifuged and processed for quantification of oxidative burst.
Quantification of cytokines
Cytokines were quantified in serum and culture supernatants. For quantification of serum cytokines, blood drawn (Serum Separator Tubes, BD Vacutainer) and allowed to coagulate for 1 h and then centrifuged. Aliquots were stored at −70°C. Following completion of the study, serum samples were batch analyzed in technical replicates for TNF-α, IL-10, IFN-γ, IL-1β, IL-6, using the V-PLEX Proinflammatory Panel 1 human assay kit (MesoScale Discovery), for monocyte chemoattractant (MCP)-1 using the V-PLEX Chemokine Panel 1 human assay kit (MesoScale Discovery), for IL-1RA using the V-PLEX Cytokine Panel 2 human assay kit (MesoScale Discovery), and high sensitivity C-reactive protein (hs-CRP) using the V-PLEX CRP human assay kit (MesoScale Discovery).
Culture supernatants were centrifuged and then frozen at −70°C. Following completion of the study, culture supernatants were batch analyzed in technical replicates for TNF-α, IL-10, IFN-γ, IL-1β, IL-18, IL-4, IL-6, and MCP-1 using the U-PLEX Custom Biomarker human assay kit (MesoScale Discovery). The LLOD’s and ULOD’s are publicly available on the manufacturer’s website.
Quantification of oxidative burst
The same procedure was used for quantification of oxidative burst in the basal state and following 4- and 24-h cultures. Fresh or cultured sodium heparin blood was treated with dihydrorhodamine (DHR)123 (5 μg/ml final concentration; Cat. No. 85100-5; Cayman Chemical) for 15 min in a 37°C water bath in the dark. LPS (Cat. No. L5418; Sigma–Aldrich) was added (50 ng/ml final concentration) and samples were incubated for 45 min. RBCs were lysed (1x final concentration; Cat. No. 130-094-183; Miltenyi Biotec) and washed twice in PBS (SH30256.02; HyClone). Samples were immediately analyzed on a CytoFLEX S (Beckman Coulter). DHR123 fluorescence was quantified on the 530 nm filter. The hierarchical gating strategy is pictured in Supplementary Fig. S1.
Quantification of phagocytosis
The same procedure was used for quantification of phagocytosis in the basal state and following 4- and 24-h cultures with different glucose and BHB concentrations. Phagocytosis was quantified by flow cytometry using the E. coli phagocytosis assay kit (Cayman Chemical; Cat. No. 601370). Fresh or cultured sodium heparin blood was treated with FITC-labelled E. coli for 10 min in a 37°C water bath in the dark. Samples were promptly put on ice and cold quenching solution was added. Samples were washed twice in cold PBS (SH30256.02; HyClone). Samples were lysed and fixed in 1x BD Phosflow lyse/fix buffer (Cat. No. 558049; BD), washed twice in PBS and immediately analyzed by flow cytometry. Intracellular E. coli were quantified on the 530 nm filter. The hierarchical gating strategy is pictured in Supplementary Fig. S2.
Quantification of IL-10 function
IL-10 function was determined using the combination of two ex vivo approaches: (i) measurement of IL-10 mediated phosphorylated STAT3 using flow cytometry; and (ii) IL-10-mediated inhibition of TNF-α secretion in LPS-stimulated whole blood cultures, as described previously [29]. Briefly, EDTA blood was stimulated with increasing concentrations of IL-10 (1, 10, 50, 100, 1000 ng/ml) and pSTAT3 median fluorescence intensity (MFI) was quantified in CD14+ monocytes and CD4+ lymphocytes using flow cytometry. Parallel whole-blood cultures stimulated with LPS (1 ng/ml) in the absence or presence of increasing doses of IL-10 (1, 2.5, 5, and 10 ng/ml) were incubated for 4 h at 37°C and 5% CO2, after which the culture supernatants were stored at −70°C until batch analysis of TNF-α concentrations using a Duo-Set ELISA.
Haematology and leukocyte subsets
Fresh EDTA blood was analyzed on a DXH520 haematology analyzer (Beckman Coulter) for a complete blood count. Further characterization of specific monocyte and T-cell subsets were quantified by flow cytometry. Whole EDTA blood was stained with fluorescent-labelled antibodies (Miltenyi Biotec). RBCs were lysed (1X final concentration; Cat. No. 130-094-183; Miltenyi Biotec) and washed twice in PBS (SH30256.02; HyClone). Samples were resuspended in PBS, stained with a viability dye (7-aminoactinomycin D (7-AAD) or propidium iodide), and analyzed on a CytoFLEX S (Beckman Coulter). The antibodies and hierarchical gating strategies used for acquisition are reported in detail in Supplementary Material and Supplementary Figs. 3–5.
Study approval
All clinical investigation was conducted in accordance with the Declaration of Helsinki. Written informed consent was obtained prior to enrolment of participants for all three trials. All trials were registered on ClinicalTrials.gov (NCT03817749; NCT04194450; NCT05155410) and approved by the University of British Columbia Clinical Research Ethics Board (H18-02930; H19-02947; H21-01762).
Statistical analyses
All statistical analyses for all studies were conducted by researchers blinded to condition. Once data analyses were complete, researchers were unblinded. Statistical analyses for the outcomes reported in the present paper were performed in RStudio (Version 4.0.3 ‘Bunny-Wunnies Freak Out’) using a linear mixed model (LMM). The sjPlot, lme4, lmerTest, and emmeans packages were used for analyses [44–47]. First, data were visually inspected to determine if they met model assumptions for normality and linearity of data using a Q–Q plot, and homoscedasticity by plotting residuals against fitted values. Data were assessed for influential data points using Cook’s distance. A data point was considered influential if Cook’s distance was above the cut-off, which was calculated as 4/N. Those data points were then checked but only excluded if there was sufficient reason to do so (i.e. technical error or inclusion significantly changed the outcome or interpretation). Data are reported as estimated marginal means with 95% confidence intervals derived from the LMM unless indicated otherwise. All estimated marginal means, 95% confidence intervals, and main and interaction effects are reported in Supplementary Tables 1–10. Significant within-factor comparisons are only reported if the main effect was significant.
For Study 1, The LMM was built with a fixed effect of BHB concentration and a random slope for participant. Significance was set at P < 0.05.
For pre/post outcomes from Study 2, the LMM was built with fixed effects for time and condition and a random slope for participant. Significance was set at P < 0.05.
Three models were used for Study 3. For analysis of pSTAT3 and TNF-α secretion into IL-10-treated cultures, the LMM was built with fixed effects of time, condition, and IL-10 treatment concentration and a random slope for participant. For outcomes derived from BHB-treated ex vivo cultures, the LMM was built with a fixed effect of BHB concentration and a random slope for participant. The remaining pre/post outcomes were analyzed with a LMM was built with fixed effects for time and condition (KME or placebo) and a random slope for participant. Significance was set at P < 0.05.
Supplementary Material
Acknowledgements
The authors would like to thank the dedicated study volunteers for their time, effort, and interest with these studies, the undergraduate volunteers who helped with data collection, Mr. Morgan Decksheimer and the team at Meal Prep for You (MP4U) for their seamless meal preparation and flexible coordination of meal pickups for participants, and Ms. Kara Crampton and Mr. Ali Daraei for creating and maintaining study blinding. The graphical abstract was created on BioRender.com.
Glossary
Abbreviations
- BHB
beta-hydroxybutyrate
- CD
cluster of differentiation
- CRP
C-reactive protein
- EDTA
ethylenediaminetetraacetic acid
- ELISA
enzyme-linked immunosorbent assay
- IFN
interferon gamma
- IL
interleukin
- IL-1RA
interleukin 1 receptor antagonist
- KME
ketone monoester supplement
- LLOD
lower limit of detection
- LMM
linear mixed effects model
- LPS
lipopolysaccharide
- MCP-1
monocyte chemoattractant protein 1
- MFI
median fluorescence intensity
- NLRP3
NLR-family pyrin domain containing 3
- PBS
phosphate-buffered saline
- pSTAT3
phosphorylated signal transducer and activator of transcription 3
- RBC
red blood cell
- ROS
reactive oxygen species
- RPMI
Roswell Park Memorial Institute medium
- SD
standard deviation
- T2D
type 2 diabetes
- Th cell
T helper cell
- TNF
tumour necrosis factor alpha
- Treg cell
T regulatory cell
- ULOD
upper limit of detection.
Contributor Information
Helena Neudorf, University of British Columbia Okanagan, School of Health and Exercise Sciences, Kelowna, BC, Canada.
Hashim Islam, University of British Columbia Okanagan, School of Health and Exercise Sciences, Kelowna, BC, Canada.
Kaja Falkenhain, University of British Columbia Okanagan, School of Health and Exercise Sciences, Kelowna, BC, Canada.
Barbara Oliveira, University of British Columbia Okanagan, School of Health and Exercise Sciences, Kelowna, BC, Canada.
Garett S Jackson, University of British Columbia Okanagan, School of Health and Exercise Sciences, Kelowna, BC, Canada.
Alfonso Moreno-Cabañas, University of Castilla-La Mancha, Department of Sport Sciences, Exercise Physiology Lab at Toledo, Spain.
Kenneth Madden, University of British Columbia, Department of Medicine, Centre of Aging SMART, Vancouver, BC, Canada.
Joel Singer, University of British Columbia, School of Population and Public Health, Vancouver, BC, Canada.
Jeremy J Walsh, McMaster University, Department of Kinesiology, Hamilton, ON, Canada.
Jonathan P Little, University of British Columbia Okanagan, School of Health and Exercise Sciences, Kelowna, BC, Canada.
Ethical Approval
All clinical investigation was conducted in accordance with the Declaration of Helsinki. Written informed consent was obtained prior to enrolment of participants for all three trials. All trials were approved by the University of British Columbia Clinical Research Ethics Board (H18-02930; H19-02947; H21-01762).
Conflict of Interests
J.P.L. is Chief Scientific Officer for the not-for-profit Institute for Personalized Therapeutic Nutrition. J.P.L. holds founder shares in Metabolic Insights Inc., a for-profit company that developed non-invasive metabolic monitoring devices. No other authors declare conflict of interest.
Funding
This work was funded by grants from the Canadian Institutes of Health Research (CIHR) (PJ9-166217, PJT-169116) and Natural Sciences and Engineering Research Council (NSERC; RGPIN-2019-05204) of Canada to J.P.L. J.P.L. was supported by a Michael Smith Foundation for Health Research (MSFHR) Scholar Award (16890) and a Killam Accelerator Research Fellowship during the conduct of these studies. H.N. was supported by a Natural Sciences and Engineering Research Council of Canada award (CGSD3-547438-2020).
Data Availability
Data from these trials will be made available upon reasonable request to the corresponding author.
Author Contributions
J.P.L., J.W., J.S., and K.M. conceived of the study and secured funding for the trials. J.P.L. and J.W. designed Study 1 and H.N. provided input and designed the whole blood cultures. J.W. and H.N. conducted data collection for Study 1. J.P.L. and B.O. designed Study 2 with input from K.F. K.F., B.O., and H.I. conducted data collection for Study 2. J.P.L., K.F., and H.N. designed Study 3. K.F., H.N., H.I., A.M.-C., and G.S.J. conducted data collection for Study 3. H.N. performed all statistical analyses with input from K.F. and oversight from J.PL. H.N. wrote the manuscript with assistance from K.F., H.I., and J.L. All authors contributed to revising the figures and manuscript. All authors approved the final version of this manuscript.
Permission to Reproduce
All data presented in this manuscript is original data generated by the authors.
Clinical Trial Registration
All clinical trials were registered on ClinicalTrials.gov (NCT03817749; NCT04194450; NCT05155410).
References
- 1. Laffel L. Ketone bodies: a review of physiology, pathophysiology, and application of monitoring diabetes. Diabetes Metab Res Rev 1999, 15, 412–26. doi: 10.1002/(sici)1520-7560(199911/12)15:6<412::aid-dmrr72>3.0.co;2-8 [DOI] [PubMed] [Google Scholar]
- 2. Goldberg EL, Asher JL, Molony RD, Shaw AC, Zeiss CJ, Wang C, et al. β-Hydroxybutyrate deactivates neutrophil NLRP3 inflammasome to relieve gout flares. Cell Rep 2017, 18, 2077–87. doi: 10.1016/j.celrep.2017.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Shippy DC, Wilhelm C, Viharkumar PA, Raife TJ, Ulland TK.. Beta-hydroxybutyrate inhibits inflammasome activation to attenuate Alzheimer’s disease pathology. J Neuroinflammation 2020, 17, 280. doi: 10.1186/s12974-020-01948-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Jiang Z, Yin X, Wang M, Wang Y, Li F, Gao Y, et al. Beta-hydroxybutyrate alleviates pyroptosis in MPP+/MPTP-induced Parkinson’s disease models via inhibiting STAT3/NLRP3/GSDMD pathway. Int Immunopharmacol 2022, 113, 109451. doi: 10.1016/j.intimp.2022.109451 [DOI] [PubMed] [Google Scholar]
- 5. Walsh JJ, Neudorf H, Little JP.. 14-Day ketone supplementation lowers glucose and improves vascular function in obesity: a randomized crossover trial. J Clin Endocrinol Metab 2021, 106, e1738–54. doi: 10.1210/clinem/dgaa925 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Youm Y-H, Nguyen KY, Grant RW, Goldberg EL, Bodogai M, Kim D, et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat Med 2015, 21, 263–9. doi: 10.1038/nm.3804 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Shimazu T, Hirschey MD, Newman J, He W, Shirakawa K, Moan NL, et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 2013, 339, 211–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Bae HR, Kim DH, Park MH, Lee B, Kim MJ, Lee EK, et al. β-hydroxybutyrate suppresses inflammasome formation by ameliorating endoplasmic reticulum stress via AMPK activation. Oncotarget 2016, 7, 66444–54. doi: 10.18632/oncotarget.12119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Lee AK, Kim DH, Bang E, Choi YJ, Chung HY.. β-hydroxybutyrate suppresses lipid accumulation in aged liver through GPR109A-mediated signaling. Aging Dis 2020, 11, 777–90. doi: 10.14336/AD.2019.0926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Rohm TV, Meier DT, Olefsky JM, Donath MY.. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. doi: 10.1016/j.immuni.2021.12.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Benfield T, Benfield T, Jensen JS, Jensen JS, Nordestgaard BG, Nordestgaard BG.. Influence of diabetes and hyperglycaemia on infectious disease hospitalisation and outcome. Diabetologia 2007, 50, 549–54. [DOI] [PubMed] [Google Scholar]
- 12. Martins M, Boavida JM, Raposo JF, Froes F, Nunes B, Ribeiro RT, et al. Diabetes hinders community-acquired pneumonia outcomes in hospitalized patients. BMJ Open Diabetes Res Care 2016, 4, e000181–e000181. doi: 10.1136/bmjdrc-2015-000181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Hotamisligil GS, Shargill NS, Spiegelman BM.. Adipose expression of tumor necrosis factor-α: direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. doi: 10.1126/science.7678183 [DOI] [PubMed] [Google Scholar]
- 14. Feingold KR, Soued M, Staprans I, Gabin LA, Donahue ME, Huang B-J, et al. Effect of tumor necrosis factor (TNF) on lipid metabolism in the diabetic rat: evidence that inhibition of adipose tissue lipoprotein lipase activity is not required for TNF-induced hyperlipidemia. JCI 1989, 83, 1116–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Yuan M, Konstantopoulos N, Lee J, Hansen L, Li ZW, Karin M, et al. Reversal of obesity- and diet-induced insulin resistance with salicylates or targeted disruption of Ikkbeta. Science 2001, 293, 1673–7. doi: 10.1126/science.1061620 [DOI] [PubMed] [Google Scholar]
- 16. Arkan MC, Hevener AL, Greten FR, Maeda S, Li Z-W, Long JM, et al. IKK-[beta] links inflammation to obesity-induced insulin resistance. Nat Med 2005, 11, 191–8. doi: 10.1038/nm1185 [DOI] [PubMed] [Google Scholar]
- 17. Hirosumi J, Tuncman G, Chang L, Görgün CZ, Uysal KT, Maeda K, et al. A central role for JNK in obesity and insulin resistance. Nature 2002, 420, 333–6. doi: 10.1038/nature01137 [DOI] [PubMed] [Google Scholar]
- 18. Nikfar S, Saiyarsarai P, Tigabu BM, Abdollahi M.. Efficacy and safety of interleukin-1 antagonists in rheumatoid arthritis: a systematic review and meta-analysis. Rheumatol Int 2018, 38, 1363–83. doi: 10.1007/s00296-018-4041-1 [DOI] [PubMed] [Google Scholar]
- 19. Donath MY. Targeting inflammation in the treatment of type 2 diabetes: time to start. Nat Rev Drug Discov 2014, 13, 465–76. doi: 10.1038/nrd4275 [DOI] [PubMed] [Google Scholar]
- 20. Cavalli G, Dinarello CA.. Anakinra therapy for non-cancer inflammatory diseases. Front Pharmacol 2018, 9, 1157–1157. doi: 10.3389/fphar.2018.01157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Sloan-Lancaster RJ, Abu-Raddad E, Polzer J, Miller J, Scherer J, De Gaetano A, et al. Double-blind, randomized study evaluating the glycemic and anti-inflammatory effects of subcutaneous LY2189102, a neutralizing IL-1β antibody, in patients with type 2 diabetes. Diabetes Care 2013, 36, 2239–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ridker PM, Everett BM, Thuren T, MacFadyen JG, Chang WH, Ballantyne C, et al.; CANTOS Trial Group. Anti-inflammatory therapy with canakinumab for atherosclerotic disease. N Engl J Med 2017, 377, 1119–31. doi: 10.1056/NEJMoa1707914 [DOI] [PubMed] [Google Scholar]
- 23. Falkenhain K, Islam H, Little JP.. Exogenous ketone supplementation: an emerging tool for physiologists with potential as a metabolic therapy. Exp Physiol 2023, 108, 177–87. doi: 10.1113/EP090430 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Neudorf H, Durrer C, Myette‐Cote E, Makins C, O’Malley T, Little JP.. Oral ketone supplementation acutely increases markers of NLRP3 inflammasome activation in human monocytes. Mol Nutr Food Res 2019, 63, e1801171-n/a. [DOI] [PubMed] [Google Scholar]
- 25. Thomsen HH, Rittig N, Johannsen M, Møller AB, Jørgensen JO, Jessen N, et al. Effects of 3-hydroxybutyrate and free fatty acids on muscle protein kinetics and signaling during LPS-induced inflammation in humans: anticatabolic impact of ketone bodies. Am J Clin Nutr 2018, 108, 857–67. doi: 10.1093/ajcn/nqy170 [DOI] [PubMed] [Google Scholar]
- 26. Walsh JJ, Myette-Côté E, Neudorf H, Little JP.. Potential therapeutic effects of exogenous ketone supplementation for type 2 diabetes: a review. Curr Pharm Des 2020, 26, 958–969. doi: 10.2174/1381612826666200203120540 [DOI] [PubMed] [Google Scholar]
- 27. Falkenhain K, Oliveira BF, Islam H, Neudorf H, Cen HH, Johnson JD, et al. The effect of acute and 14-day exogenous ketone supplementation on glycaemic control in adults with type 2 diabetes: two randomized controlled trials. Am J Physiol Endo Metab 2023, 326, E61–E72. doi: 10.1152/ajpendo.00332.2023 [DOI] [PubMed] [Google Scholar]
- 28. Barry JC, Shakibakho S, Durrer C, Simtchouk S, Jawanda KK, Cheung ST, et al. Hyporesponsiveness to the anti-inflammatory action of interleukin-10 in type 2 diabetes. Sci Rep 2016, 6, 21244–21244. doi: 10.1038/srep21244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Islam H, Jackson GS, Yoon JSJ, Cabral-Santos C, Lira FS, Mui AL, et al. Sex differences in IL-10’s anti-inflammatory function: greater STAT3 activation and stronger inhibition of TNF-α production in male blood leukocytes ex vivo. Am J Physiol Cell Physiol 2022, 322, C1095–104. doi: 10.1152/ajpcell.00091.2022 [DOI] [PubMed] [Google Scholar]
- 30. Joshi MB, Lad A, Bharath Prasad AS, Balakrishnan A, Ramachandra L, Satyamoorthy K.. High glucose modulates IL-6 mediated immune homeostasis through impeding neutrophil extracellular trap formation. FEBS Lett 2013, 587, 2241–6. doi: 10.1016/j.febslet.2013.05.053 [DOI] [PubMed] [Google Scholar]
- 31. Joshi MB, Ahamed R, Hegde M, Nair AS, Ramachandra L, Satyamoorthy K.. Glucose induces metabolic reprogramming in neutrophils during type 2 diabetes to form constitutive extracellular traps and decreased responsiveness to lipopolysaccharides. Biochim Biophys Acta Mol Basis Dis 2020, 1866, 165940–165940. doi: 10.1016/j.bbadis.2020.165940 [DOI] [PubMed] [Google Scholar]
- 32. Perner A, Nielsen SE, Rask-Madsen J.. High glucose impairs superoxide production from isolated blood neutrophils. Intensive Care Med 2003, 29, 642–5. doi: 10.1007/s00134-002-1628-4 [DOI] [PubMed] [Google Scholar]
- 33. Restrepo BI, Twahirwa M, Rahbar MH, Schlesinger LS.. Phagocytosis via complement or Fc-gamma receptors is compromised in monocytes from type 2 diabetes patients with chronic hyperglycemia. PLoS One 2014, 9, e92977–e92977. doi: 10.1371/journal.pone.0092977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Hair PS, Echague CG, Rohn RD, Krishna NK, Nyalwidhe JO, Cunnion KM.. Hyperglycemic conditions inhibit C3-mediated immunologic control of Staphylococcus aureus. J Transl Med 2012, 10, 35–35. doi: 10.1186/1479-5876-10-35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Mauriello CT, Hair PS, Rohn RD, Rister NS, Krishna NK, Cunnion KM.. Hyperglycemia inhibits complement-mediated immunological control of s. aureus in a rat model of peritonitis. J Diabetes Res 2014, 2014, 762051–762051. doi: 10.1155/2014/762051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Cassatella MA, Meda L, Bonora S, Ceska M, Constantin G.. Interleukin 10 (IL-10) inhibits the release of proinflammatory cytokines from human polymorphonuclear leukocytes Evidence for an autocrine role of tumor necrosis factor and IL-1β in mediating the production of IL-8 triggered by lipopolysaccharide. J Exp Med 1993, 178, 2207–11. doi: 10.1084/jem.178.6.2207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Shin D-I, Banning U, Kim Y-M, Verheyen J, Hannen M, Bönig H, et al. Interleukin 10 inhibits TNF-alpha production in human monocytes independently of interleukin 12 and interleukin 1 beta. Immunol Invest 1999, 28, 165–75. doi: 10.3109/08820139909061145 [DOI] [PubMed] [Google Scholar]
- 38. Stubbs BJ, Cox PJ, Evans R, Santer P, Miller JJ, Faull OK, et al. On the metabolism of exogenous ketones in humans. Front Physiol 2017, 8, 848. doi: 10.3389/fphys.2017.00848 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Falkenhain K, Daraei A, Forbes SC, Little JP.. Effects of exogenous ketone supplementation on blood glucose: a systematic review and meta-analysis. Adv Nutr 2022, 13, 1697–714. doi: 10.1093/advances/nmac036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Soto-Mota A, Vansant H, Evans RD, Clarke K.. Safety and tolerability of sustained exogenous ketosis using ketone monoester drinks for 28 days in healthy adults. Regul Toxicol Pharmacol 2019, 109, 104506. doi: 10.1016/j.yrtph.2019.104506 [DOI] [PubMed] [Google Scholar]
- 41. Soto‐Mota A, Norwitz NG, Evans R, Clarke K, Barber TM.. Exogenous ketosis in patients with type 2 diabetes: Safety, tolerability and effect on glycaemic control. Endocrinol Diabetes Metab J 2021, 4, e00264-n/a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Myette‐Côté E, Neudorf H, Rafiei H, Clarke K, Little JP.. Prior ingestion of exogenous ketone monoester attenuates the glycaemic response to an oral glucose tolerance test in healthy young individuals. J Physiol 2018, 596, 1385–95. doi: 10.1113/jp275709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Myette-Côté E, Caldwell HG, Ainslie PN, Clarke K, Little JP.. A ketone monoester drink reduces the glycemic response to an oral glucose challenge in individuals with obesity: a randomized trial. Am J Clin Nutr 2019, 110, 1491–1501. doi: 10.1093/ajcn/nqz232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Lüdecke D. sjPlot: Data Visualization for Statistics in Social Science. 2023. https://CRAN.R-project.org/package=sjPlot
- 45. Bates D, Maechler M, Bolker B, Walker S.. Fitting linear mixed-effects models using lme4. J Stat Softw 2015, 67, 1–48. [Google Scholar]
- 46. Kuznetsova A, Brockhoff PB, Christensen RHB.. lmertest package: tests in linear mixed effects models. J Stat Softw 2017, 82, 1–26. [Google Scholar]
- 47. Lenth R, Buerkner P, Herve M, Love J, Riebl H, Singmann H.. Estimated Marginal Means, aka Least-Squares Means. 2021. https://github.com/rvlenth/emmeans [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data from these trials will be made available upon reasonable request to the corresponding author.