

Abstract
目的
沙眼衣原体(Chlamydia trachomatis,Ct)泌尿生殖道感染引起输卵管积水和纤维化的机制尚不完全清楚。肠道微生物群除引起肠道疾病外,还与心脑血管疾病、肝硬化、呼吸道感染、多囊卵巢综合征、子宫内膜异位和细菌性阴道炎等肠外疾病的发生、发展密切相关。鼠李糖乳杆菌GG(Lactobacillus rhamnosus GG,LGG)是一种被广泛研究和应用的肠道益生菌,其益处包括治疗胃肠道疾病和调节机体免疫,它还可以缓解过敏反应和腹泻,并抑制各种呼吸道疾病和泌尿生殖系统病原体所致的疾病。本研究以致病性和Ct高度相似的鼠衣原体(Chlamydia muridarium,Cm)感染小鼠作为动物模型,初步探讨LGG对小鼠Cm上生殖道感染的调节作用及机制,为了解Ct泌尿生殖道感染的致病机制提供实验依据,并为Ct感染的治疗提供新的思路。
方法
将6周龄C57BL/6J雌性小鼠分为实验组和对照组,其中实验组小鼠口服5×108菌落形成单位的LGG 19 d,对照组小鼠口服PBS。在两组小鼠口服LGG或PBS的第9天皮下注射2.5 mg 孕酮,第14天经阴道感染1×105包涵体形成单位的Cm。小鼠感染Cm后,每周取阴道拭子和直肠拭子,将拭子系列稀释后接种HeLa细胞,24 h后进行间接免疫荧光染色,在显微镜下计数衣原体包涵体。于Cm感染后第14天和第63天分别处死小鼠,分离上、下生殖道,将组织匀浆接种HeLa细胞培养24 h后,计数两组小鼠双侧子宫角、输卵管卵巢及宫颈阴道组织中的Cm载量;用流式细胞术检测Cm感染后第14天小鼠脾T淋巴细胞胞内IFN-γ、IL-5和IL-17的分泌;观察感染后第63天小鼠双侧子宫角和输卵管的病理损伤,并将生殖道各部分进行病理切片和HE染色,评价输卵管管腔扩张和炎性细胞浸润程度;采用间接ELISA检测Cm感染后第14天和第63天小鼠血清特异性IgM和IgG水平。
结果
Cm经阴道感染小鼠后,两组小鼠衣原体在生殖道的清除速度、感染后第14天双侧子宫角和输卵管的上行感染量、Cm从生殖道向肠道转移和定植量差异均无统计学意义(均P>0.05);Cm感染后第14天,实验组小鼠脾T淋巴细胞内IL-17的分泌水平较对照组小鼠显著降低(t=2.486,P<0.05),但两组小鼠脾分泌IFN-γ和IL-5的CD4+T淋巴细胞比例及血清Cm特异性IgM和IgG水平差异均无统计学意义(均P>0.05);Cm感染后第63天,实验组小鼠双侧子宫角和输卵管的炎症程度与对照组差异无统计学意义(P>0.05),但输卵管水肿程度显著低于对照组(P<0.05)。
结论
口服LGG不影响Cm阴道感染小鼠病原体生殖道的上行感染、清除及其向肠道的转移和定植,亦未显著影响小鼠血清特异性IgM和IgG水平;口服LGG可减少Cm感染后小鼠脾Th17细胞应答水平,能减轻小鼠Ct感染引起的输卵管纤维化和输卵管积水。
Keywords: 鼠李糖乳杆菌GG, 鼠衣原体, 输卵管积水, 白细胞介素-17
Abstract
Objective
Chlamydia trachomatis is a pathogen which can cause hydrosalpinx and tubal fibrosis when infecting the urogenital tract. However, the mechanism is still not clear. There is growing evidence that the gut microbiota is associated with the pathogenesis of both intestinal and extra-intestinal disorders, such as cardiovascular disease, hepatocirrhosis, allergy, respiratory tract infection, polycystic ovary syndrome, endometriosis, and bacterial vaginitis. Lactobacillus rhamnosus GG (LGG) is one of the most extensively studied and widely used probiotic bacteria, the benefits of LGG including the treatment in gastrointestinal disorders and immunomodulation are well demonstrated, and it can also alleviate hypersensitivity reaction and diarrhoea, inhibit a variety of respiratory and urogenital diseases. Chlamydia muridarium (Cm) infection is a good model for the study on human Chlamydia pathogenicity in genitourinary tract. The mice infected with Cm were used as animal models to preliminarily explore the mechanism for the effect of LGG on upper reproductive tract infection in the mice, and to provide experimental basis for the pathogenesis of Chlamydia trachomatis genitourinary tract infection and the new idea for the treatment of Chlamydia trachomatis infection.
Methods
Five to six weeks-old C57BL/6J mice were divided into 2 groups: An experimental group and a control group. The experimental group were administrated with 5×108 colony forming units (CFU) LGG for 19 consecutive days, while the control group were feed PBS. The mice in the 2 group were subcutaneously injected with 2.5 mg progesterone on Day 9 and infected with 1×105 inclusion body forming unit of Cm via the vaginal tract on Day 14. Vaginal and rectal swabs were taken every 7 days to infect HeLa cells for 24 hours, then the indirect immunofluorescence assay was used and the number of inclusion bodies of Chlamydia were calculated. Mice were euthanized on Day 14 and Day 63 after Cm inoculation, the vaginal tracts were dissected, and the tissue homogenates were prepared to culture the pathogens for 24 hours. The Cm bearing capacity in the bilateral uterine horn, tubal ovary, and cervical vaginal tissues in the 2 groups were calculated. The spleen cells were harvested to assay the intracellular IFN-γ, IL-5, and IL-17 by flow cytometry. On Day 63 after the Chlamydia infection, the pathology injury in the bilateral uterine horn and oviduct was observed, and the pathological sections and HE staining in the various part of genital tract were performed. The inflammatory cell infiltration and lumen dilatation was assessed. The specific IgM and IgG in sera were detected by indirect ELISA on Day 14 and 63 after infection.
Results
There was no effect of LGG on the clearing of Cm from the urogenital tract, the Chlamydia ascending to fallopian tube or the uterine horn, and the organism dissemination and colonization to the gastrointestinal tract (all P>0.05). On Day 14 after Cm infection via the vagina, the IL-17 expression level in the experimental group was significant decreased than that in the control group (t=2.486, P<0.05), but there was no significant difference between the 2 groups in the CD4+ T rate in spleen and IgM and IgG levels in serum after Cm intravaginal infection (all P>0.05). On Day 63 after Cm infection, there was no difference in the severity of inflammation in the uterine horns and fallopian tubes between the 2 groups (P>0.05), but the dilation of the fallopian tubes and hydrosalpinx was attenuated in the experimental group compared with the control group (P<0.05).
Conclusion
Oral administration of LGG has no effect on inhibiting Cm ascending to upper genital tract and preventing the dissemination and colonization of Cm to the gastrointestinal tract, which also cannot affect the secretion of specific IgM and IgG in sera. Oral administration of LGG can suppress the production of IL-17 in the spleen cells and attenuate hydrosalpinx development when following Cm intravaginal infection in mice.
Keywords: Lactobacillus rhamnosus GG, Chlamydial muridarium, hydrosalpinx, interleukin-17
沙眼衣原体(Chlamydia trachomatis,Ct)泌尿生殖道感染可引起非淋球菌性尿道炎,并可导致慢性输卵管炎、盆腔炎和输卵管性不孕等多种并发症和后遗症[1],研究Ct致病机制和寻找有效的治疗措施有重要意义。鼠衣原体(Chlamydia muridarium,Cm)是一种新的Ct鼠生物型衣原体,其引起的鼠泌尿生殖道感染和Ct导致的人类泌尿生殖道感染非常相似,故Cm鼠生殖道感染模型目前被广泛应用于探索Ct的病理损伤和免疫保护作用的研究[2]。
在正常机体肠道中栖息着一定种类和数量的微生物群,在维持肠道的稳态和宿主健康中发挥十分重要的作用。肠道菌群失调除引起炎性肠病等肠道疾病外,还与心脑血管疾病、肝硬化、肥胖、多囊卵巢综合征、子宫内膜异位和细菌性阴道炎等肠外疾病的发病密切相关[3-6]。而补充乳酸杆菌、干酪乳杆菌、鼠李糖乳杆菌等益生菌有助于维持菌群稳态、调节宿主免疫功能,促进机体健康[7-8]。肠道添加益生菌,除可辅助治疗急性胃肠炎等多种肠道疾病外,还可减轻非酒精性脂肪性肝病和急性呼吸道感染等[9]。此外,研究[6, 10]报道:粪菌移植、肠道添加鼠李糖乳杆菌HN001和嗜酸乳酸杆菌La-14也可减轻女性生殖道疾病。
鼠李糖乳杆菌GG(Lactobacillus rhamnosus GG,LGG)是经国家卫生部认证的可放心食用的6类儿童益生菌之一,可调节机体免疫、缓解超敏反应的发生,对腹泻和湿疹等有很好的调理作用,并抑制HIV的传播和复制,减轻大肠杆菌泌尿道感染和肠道沙门菌致病作用等[11]。本研究通过Cm感染小鼠模型,探讨LGG对小鼠Cm生殖道感染的调节作用及机制,旨在为Ct泌尿生殖道感染的疾病机制提供实验依据,并为Ct泌尿生殖道感染的治疗提供新的思路。
1. 材料与方法
1.1. 材料
1.1.1. 实验动物和菌株
5~6周龄雌性C57BL/6J小鼠26只,由中南大学实验动物部提供和饲养。野生型Cm菌株(CM G13.32.1)由美国德州大学圣安东尼奥健康与生命中心钟光明教授提供。LGG购自中国微生物菌种保藏管理委员会普通微生物中心。HeLa细胞由中南大学湘雅附属第三医院临床实验中心保存。
1.1.2. 试剂
PE抗鼠CD4抗体、FITC抗鼠CD8a抗体、PE-Cy7抗鼠CD3e抗体、eFluor® 506抗鼠IL-17抗体、PerCPCy5.5抗鼠IL-4抗体和APC抗鼠IFN-γ抗体均购自美国Bdbio Sciences公司;HRP标记的羊抗鼠IgM和HRP标记的羊抗鼠IgG购自美国Thermo Fisher Scientific公司;胎牛血清购自北京沃比森科技有限公司。
1.2. 方法
1.2.1. 小鼠口服LGG和经阴道感染Cm
26只雌性C57BL/6J小鼠随机分为实验组和对照组,实验组小鼠每只每天口服5×108菌落形成单位的LGG,共19 d[12];对照组小鼠口服PBS。口服LGG后第9天,两组小鼠均予皮下注射2.5 mg孕酮以同步小鼠生理周期和促进小鼠对Cm的易感性。LGG口服后第14天,两组小鼠均经阴道接种1×105包涵体形成单位(inclusion forming unit,IFU)的Cm原体(elemeniary body,EB)。Cm感染后每间隔7 d采集小鼠阴道和直肠拭子。Cm感染后第14天每组各处死5只小鼠,第63天每组处死8只小鼠,分离生殖道,并收集血清和脾细胞。
1.2.2. 小鼠生殖道Cm带菌量检测
将拭子收集的小鼠阴道和直肠脱落细胞和Cm感染后第14天的阴道和宫颈、双侧卵巢及输卵管匀浆用2 mL蔗糖磷酸谷氨酸溶液(sucrose-phosphate-glutamate,SPG)按1꞉5、1꞉10、1꞉20、1꞉40、1꞉80稀释后感染HeLa细胞,在CO2培养箱中培养24 h后弃去培养液,用PBS洗涤2次,然后用4%多聚甲醛固定30 min,0.5% Triton X-100处理10 min,用含10%血清的DMEM培养基于室温下封闭1 h,再加入1꞉1 000稀释的兔抗Cm EB(Chlamydia muridarum elementary body,克隆号1064)于37 ℃下孵育1 h,充分洗涤后加入Cy2标记的羊抗兔IgG(1꞉300稀释)和Hoechst(Hoechest 33258)(1꞉1 000稀释),于37 ℃下避光孵育45 min,用PBS洗涤5次。在荧光显微镜下,每个孔随机选取5个视野,计数平均每一视野中的IFU。若在某些视野下IFU数量小于5个,则计数整个细胞的IFU以对衣原体进行定量。
1.2.3. 小鼠生殖道病理损伤
采用大体标本观察小鼠子宫、卵巢和输卵管病变。并用10%甲醛固定、切片、HE染色,采用双盲法对管腔扩张和炎症反应进行评分,判定上生殖道病变。63 d后处死小鼠,分离生殖道组织(包括阴道、子宫角、输卵管、卵巢),肉眼对输卵管积水程度进行评分并拍照保存。评分标准如下:0分为无明显水肿;1分为输卵管直径小于卵巢直径,需用放大镜观察;2分为有明显水肿,但输卵管直径仍小于卵巢直径;3分为输卵管直径等于卵巢直径;4分为输卵管直径大于卵巢直径。
1.2.4. 流式细胞术检测细胞内细胞因子
小鼠感染Cm后第14天两组各处死5只小鼠,无菌分离脾细胞,氯化钾铵裂解液(ammonium-chloride-potassium lysing buffer,ACK)裂解红细胞后,在96孔细胞培养板中每孔加入200 μL 2×106的脾细胞,用终浓度为20 ng/mL佛波醇(phorbol-12-myristate-13-acetate,PMA)、1 μg/mL离子霉素和2 μmol/mL布雷菲德菌素A(Brefeldin A,BFA)于37 ℃下刺激5 h,以PBS洗涤后用CD16/CD32单克隆抗体(monoclonal antibody,mAb)在冰上避光孵育30 min封闭抗体的可结晶片段(fragment crystallizable,Fc)受体,然后进行表面膜分子和细胞内细胞因子染色,采用流式细胞术检测分泌IFN-γ、IL-5和IL-17的细胞比例。
1.2.5. ELISA检测Cm特异性IgG和IgM
纯化的Cm将100 μL1×包被缓冲液(0.05 mmol/L碳酸盐缓冲液,pH9.6)加入酶标板过夜,第2天用含吐温-20的磷酸盐缓冲液(phosphate buffered saline with Tween-20,PBST)洗涤3次后加入3% BSA,再于4 ℃下封闭过夜。以PBST洗涤后加入用1% BSA稀释的小鼠血清于37 ℃下孵育1 h。以PBST洗涤,分别加入HRP标记的羊抗鼠IgM和IgG,于37 ℃下孵育1 h后用PBST洗涤,加入2, 2'-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐[2, 2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)]显色剂,于37 ℃下避光反应30 min。用10%十二烷基硫酸钠终止反应,酶标仪检测405 nm处的吸光度值。收集Cm感染后第14天和第63天小鼠血清,将其1꞉1 600稀释后分析Cm特异性IgM和IgG水平。
1.3. 统计学处理
用双侧t检验分析两组小鼠上生殖道Cm的载量、细胞因子和抗体水平;用Wilcoxon Rank Sum Test比较各组阴道和肠道带菌量及输卵管积水的病理评分。用Fisher's确切概率法计算肉眼输卵管积水的发生率。以P<0.05为差异有统计学意义。
2. 结 果
2.1. 口服LGG未改变小鼠下生殖道衣原体清除
小鼠在感染Cm第3~14天后,Cm在阴道发生显著增殖,阴道脱落细胞中活菌量在感染Cm后第21~35天减少,第42天后阴道拭子中未分离到Cm(图1)。两组小鼠阴道Cm的带菌量差异无统计学意义(P>0.05),口服LGG后,小鼠下生殖道衣原体的清除没有受到明显影响。
图1.

Cm 感染后小鼠下生殖道衣原体载量
Figure 1 Loading capacity of Chlamydia after Cm infection in lower genital tract in mice
2.2. 口服LGG未影响 Cm 在生殖道的上行感染
小鼠感染Cm后第14天,两组小鼠上生殖道衣原体载量差异均无统计学意义(P>0.05,图2),口服LGG未影响Cm上行到子宫角和输卵管。
图2.

Cm 感染后第14天小鼠生殖道衣原体载量
Figure 2 Loading capacity in genital tract on the 14th day after Cm infection in mice
C+V: Vagina and cervix; L-UH: Left uterine horn; R-UH: Right uterine horn; L-OV: Left oviduct; R-OV: Right oviduct.
2.3. 口服LGG不改变 Cm 在小鼠肠道的扩散和定植
肛门直肠拭子Cm培养结果显示:Cm感染第7天开始肠道可检测到活菌,第28天Cm检出量最高,第56天检测到的活菌数量减少,但两组小鼠肠道定植的Cm量差异无统计学意义(P>0.05,图3)。
图3.
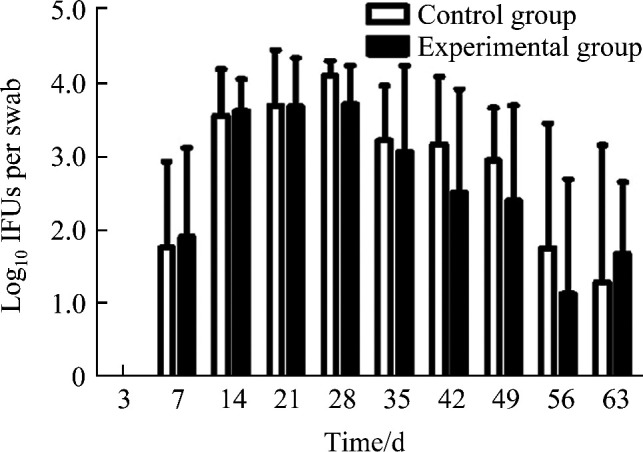
Cm 感染后小鼠肠道的衣原体载量
Figure 3 Loading capacity of Chlamydia after Cm infection in gastrointestinal tract in mice
2.4. 口服LGG减轻 Cm 感染引起的上生殖道病变
小鼠感染Cm后第63天,肉眼观察输卵管和子宫角水肿、积水及积水导致的管腔扩张(图4A)。对照组小鼠输卵管积水发生率为87.5%(7/8),评分为4.88±3.00;实验组小鼠的输卵管积水发生率为62.5%(5/8),评分为2.25±2.25,其输卵管积水评分较对照组明显降低(P<0.05,图4B)。
图4.

口服LGG对 Cm 感染小鼠生殖道病理损伤的影响
Figure 4 Effect of oral administration of LGG on genital tract pathology in mice after intravaginal Cm infection
A: Gross pathology of genital tract. The white arrow indicates the oviducts with hydrosalpinges. B: Hydrosalpinx score. *P<0.05 vs the control group.
2.5. 口服LGG小鼠Th17细胞应答水平降低
Cm感染后第14天,实验组小鼠和对照组小鼠脾淋巴细胞中分泌IFN-γ的CD4+T和CD8+T细胞百分比及分泌IL-5的CD4+T细胞差异均无统计学意义(均P>0.05;图5A~5C)。但实验组小鼠脾淋巴细胞中2.33%±0.75%的CD4+T细胞分泌IL-17,而对照组中分泌IL-17的CD4+T细胞的百分比为3.49%±0.86%,实验组小鼠Th17细胞应答水平显著低于对照组(t=2.486,P<0.05;图5D)。
图5.

口服LGG对 Cm 感染后第14天小鼠脾脏T淋巴分泌细胞因子的调节
Figure 5 Regulation of oral LGG to cytokines secretion in mouse spleen T lymphocytes after intravaginal Cm infection
A: Intracellular IFN-γ in CD4+ T cells; B: Intracellular IFN-γ in CD8+ T cells; C: Intracellular IL-5 in CD4+ T cells; D: Intracellular IL-17 in CD4+ T cells. *P<0.05 vs the control group.
2.6. 口服LGG未改变 Cm 特异性IgM和IgG的分泌
结果显示两组血清中的IgM和IgG表达量差异均无统计学意义(均P>0.05,图6)。
图6.

口服LGG对 Cm 感染小鼠特异性IgM和IgG表达水平的影响
Figure 6 Effect of oral LGG on specific IgG and IgM expression in sera after intravaginal Cm infection
3. 讨 论
Ct感染所致的慢性输卵管炎、盆腔炎和输卵管性不孕的机制目前尚不清楚。以往认为这些并发症和后遗症是Ct在生殖道持续感染所致,但近年来越来越多的研究[13-14]发现Ct感染后期,病变输卵管组织基本检测不到衣原体。Ct感染引起的上生殖道病变可能是由免疫病理反应所致。一方面,生殖道Cm感染引起的急性期炎症反应诱生的促炎细胞因子,在清除衣原体同时也可能导致病理损伤[15-16];另一方面,衣原体经生殖道感染后可进入肠道,并在肠道长期持续性存在[17-18]。虽然经生殖道、呼吸道和静脉感染的衣原体均可进入肠道,但经肠道接种的衣原体不进入生殖道[19-20]。此外,研究[21]显示衣原体可长期定植于肠道,并诱发较强的免疫应答,对宿主产生特异、持续性的免疫刺激,亦可能与输卵管积水和纤维化的形成有关。
LGG能在肠道定植并发挥免疫调节作用。本结果显示:经阴道感染Cm后,两组小鼠在第3~14天衣原体发生显著增殖,第21~35天衣原体活菌量减少,第42天后阴道拭子中未分离到Cm。这与以往研究[22]中阴道感染Cm后,导致输卵管积水的生殖道衣原体在感染后第28天左右被清除的情况一致。另根据研究[23-24]报道:小鼠经阴道感染Cm后第7天和第14天,其双侧子宫角和输卵管中均可检测到Cm,且载量最高,并相对稳定;第14天后双侧子宫角和输卵管中Cm载量逐渐减少。第14天的结果也提示衣原体上行到上生殖道,但肠道补充LGG未改变小鼠生殖道衣原体的清除速度,且口服LGG未影响经阴道感染的Cm向子宫和输卵管的上行感染。此外,本实验结果也表明口服LGG未影响Cm向肠道的扩散和定植。但有意思的是,虽然肠道补充LGG未影响衣原体在生殖道和肠道的增殖,但可显著减轻衣原体生殖道感染引起的输卵管扩张和输卵管积水。
为探究口服LGG减轻Cm感染诱发的上生殖道病变的原因,本研究分析了衣原体感染后小鼠脾淋巴细胞诱生细胞因子水平及血清特异性抗体滴度,发现口服LGG未显著改变衣原体感染后T淋巴细胞IFN-γ 和IL-5的分泌及特异性IgM和IgG表达水平,提示LGG对Cm感染诱导的Th1和Th2型免疫应答的调节作用较弱,亦不影响衣原体抗体的分泌。但口服LGG能使小鼠脾淋巴细胞中衣原体诱生的IL-17的表达显著减少。
IL-17是CD4+T细胞分泌的一种促炎细胞因子,在机体抗感染中发挥重要作用。此外,IL-17还参与免疫应答的调节,其高表达与多种自身免疫性疾病和肿瘤的发生、发展密切相关。Ct和Cm感染诱生的IL-17可增加IFN-γ的分泌,协助Th1细胞应答,促进体内衣原体的清除;但越来越多的研究[16, 25-26]表明IL-17的高表达与衣原体感染所致输卵管积水的形成密切相关。已有研究[25, 27-29]表明肠道LGG可有效调节Th17/调节性T细胞应答水平,抑制细胞IL-17的分泌。因此,口服LGG减轻Cm感染后期输卵管积水的形成可能与LGG抑制Th17细胞应答有关。
Ct感染引起上生殖道病变是多因素综合作用的结果,除IL-17A外,遗传背景、衣原体生殖道感染量、TNF-α、IL-13和调节性T细胞应答等均可影响其形成和发展。此外,LGG可通过多种机制调节免疫应答,如诱导机体产生抗微生物肽和TNF-α、促进IL-10的分泌等;除抑制IL-17分泌外,口服LGG是否存在其他机制影响衣原体生殖道感染引起的病变还有待于进一步研究。
基金资助
国家自然科学基金(31800774)。
This work was supported by the National Natural Science Foundation of China (31800774).
利益冲突声明
作者声称无任何利益冲突。
原文网址
http://xbyxb.csu.edu.cn/xbwk/fileup/PDF/2021111187.pdf
参考文献
- 1. Elwell C, Mirrashidi K, Engel J. Chlamydia cell biology and pathogenesis[J]. Nat Rev Microbiol, 2016, 14(6): 385-400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Pal S, Peterson EM, de la Maza LM. New murine model for the study of Chlamydia trachomatis genitourinary tract infections in males[J]. Infect Immun, 2004, 72(7): 4210-4216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Petersen C, Round JL. Defining dysbiosis and its influence on host immunity and disease[J]. Cell Microbiol, 2014, 16(7): 1024-1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lindheim L, Bashir M, Münzker J, et al. Alterations in gut microbiome composition and barrier function are associated with reproductive and metabolic defects in women with polycystic ovary syndrome (PCOS): a pilot study[J]. PLoS One, 2017, 12(1): e0168390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Kobayashi H, Higashiura Y, Shigetomi H, et al. Pathogenesis of endometriosis: the role of initial infection and subsequent sterile inflammation[J]. Mol Med Rep, 2014, 9(1): 9-15. [DOI] [PubMed] [Google Scholar]
- 6. Jang SY, Jeong JJ, Choi SY, et al. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 attenuate gardnerella vaginalis-infected bacterial vaginosis in mice[J]. Nutrients, 2017, 9(6): 531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Zhang Z, Lv J, Pan L, et al. Roles and applications of probiotic Lactobacillus strains[J]. Appl Microbiol Biotechnol, 2018, 102(19): 8135-8143. [DOI] [PubMed] [Google Scholar]
- 8. Marchesi Julian R, David HA, Francesca F, et al. The gut microbiota and host health: a new clinical frontier[J]. Gut, 2016, 65(2): 330-339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Suez J, Zmora N, Segal E, et al. The pros, cons, and many unknowns of probiotics[J]. Nat Med, 2019, 25(5): 716-729. [DOI] [PubMed] [Google Scholar]
- 10. Quaranta G, Sanguinetti M, Masucci L. Fecal microbiota transplantation: A potential tool for treatment of human female reproductive tract diseases[J]. Front Immunol, 2019, 10: 2653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Capurso L. Thirty Years of Lactobacillus rhamnosus GG: A review[J]. J Clin Gastroenterol, 2019, 53(Suppl 1): S1-S41. [DOI] [PubMed] [Google Scholar]
- 12. Kawase M, He F, Kubota A, et al. Oral administration of lactobacilli from human intestinal tract protects mice against influenza virus infection[J]. Lett Appl Microbiol, 2010, 51(1): 6-10. [DOI] [PubMed] [Google Scholar]
- 13. Ahmadi MH, Mirsalehian A, Bahador A. Association of Chlamydia trachomatis with infertility and clinical manifestations: a systematic review and meta-analysis of case-control studies[J]. Infect Dis (Lond), 2016, 48(7): 517-523. [DOI] [PubMed] [Google Scholar]
- 14. Hafner LM. Pathogenesis of fallopian tube damage caused by Chlamydia trachomatis infections[J]. Contraception, 2015, 92(2): 108-115. [DOI] [PubMed] [Google Scholar]
- 15. Agrawal SK, Rawre J, Khanna N, et al. Increase in Chlamydia trachomatis genital and extra-genital infections in Indian males[J]. Ind J Med Microbiol, 2019, 37(2): 285-286. [DOI] [PubMed] [Google Scholar]
- 16. Fang S, Akash HV, Amy V, et al. Combined blockade of TNF-α and IL-17A alleviates progression of collagen-induced arthritis without causing serious infections in mice[J]. J Immunol, 2019, 202(7): 2017-2026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Chen J, Zhang H, Zhou Z, et al. Chlamydial induction of hydrosalpinx in 11 strains of mice reveals multiple host mechanisms for preventing upper genital tract pathology[J]. PLoS One, 2014, 9(4): e95076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. 邵丽丽, 马璟玥, 练婷婷, 等. 不同途径接种衣原体后小鼠多脏器衣原体检测分析 [J]. 中华皮肤科杂志, 2019, 52(8): 554-560. [Google Scholar]; SHAO Lili, MA Jingyue, LIAN Tingting, et al. Detection and analysis of multiple organ chlamydia in mice after chlamydia inoculated by different routes[J]. Chinese Journal of Dermatology, 2019, 52(8): 554-560. [Google Scholar]
- 19. Wang L, Zhang Q, Zhang T, et al. The Chlamydia muridarum organisms fail to auto-inoculate the mouse genital tract after colonization in the gastrointestinal tract for 70 days[J/OL]. PLoS One, 2016, 11(5): e0155880[2021-03-28]. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0155880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wang L, Zhu C, Zhang T, et al. Nonpathogenic colonization with Chlamydia in the gastrointestinal tract as oral vaccination for inducing transmucosal protection[J/OL]. Infect Immun, 2018, 86(2): e00630-17[2021-03-28]. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5778366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Tian Q, Zhou Z, Wang L, et al. Gastrointestinal coinfection promotes Chlamydial pathogenicity in the genital tract[J/OL]. Infect Immun, 2020, 88(4): e00905-19[2021-03-28]. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7093119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. de la Maza LM, Pal S, Khamesipour A, et al. Intravaginal inoculation of mice with the Chlamydia trachomatis mouse pneumonitis biovar results in infertility[J]. Infect Immun, 1994, 62(5): 2094-2097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Gratrix J, Singh AE, Bergman J, et al. Prevalence and characteristics of rectal chlamydia and gonorrhea cases among men who have sex with men after the introduction of nucleic acid amplification test screening at 2 Canadian sexually transmitted infection clinics[J]. Sex Transm Dis, 2014, 41(10): 589-591. [DOI] [PubMed] [Google Scholar]
- 24. van Liere GA, Dirks JA, Hoebe CJ, et al. Anorectal chlamydia trachomatis load is similar in men who have sex with men and women reporting anal sex[J/OL]. PLoS One, 2015, 10(8): e134991. [2021-03-28]. 10.1371/journal.pone.0134991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. 陈彦波, 吴移谋. Th17细胞在衣原体感染中的作用研究[J]. 微生物学免疫学进展, 2015, 43(2): 45-48. [Google Scholar]; CHEN Yanbo, WU Yimou. Study on the role of Th17 cells in Chlamydia infection[J]. Progress in Microbiology and Immunology, 2015, 43(2): 45-48. [Google Scholar]
- 26. Li Y, Wei C, Xu H, et al. The immunoregulation of Th17 in host against intracellular bacterial infection[J]. Mediators Inflamm, 2018, 51(20): 6587296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Wu C, Lin F, Lee Y, et al. Effect of Lactobacillus rhamnosus GG immunopathologic changes in chronic mouse asthma model[J]. J Microbiol Immunol Infect, 2019, 52(6): 911-919. [DOI] [PubMed] [Google Scholar]
- 28. Chen R, Xu M, Du J, et al. Lactobacillus rhamnosus GG supernatant promotes intestinal barrier function, balances Treg and TH17 cells and ameliorates hepatic injury in a mouse model of chronic-binge alcohol feeding[J]. Toxicol Lett, 2016, 241: 103-110. [DOI] [PubMed] [Google Scholar]
- 29. Ghiamati Yazdi F, Zakeri A, van Ark I, et al. Crude turmeric extract improves the suppressive effects of Lactobacillus rhamnosus GG on allergic inflammation in a murine model of house dust mite-induced asthma[J]. Front Immunol, 2020, 11: 1092. [DOI] [PMC free article] [PubMed] [Google Scholar]