

Abstract
Following the commodity risk assessment of bonsai plants (Pinus parviflora grafted on Pinus thunbergii) from China performed by EFSA, the EFSA Plant Health Panel performed a pest categorisation of Pyrrhoderma noxium, a clearly defined plant pathogenic basidiomycete fungus of the order Hymenochaetales and the family Hymenochaetaceae. The pathogen is considered as opportunistic and has been reported on a wide range of hosts, mainly broad‐leaved and coniferous woody plants, causing root rots. In addition, the fungus was reported to live saprophytically on woody substrates and was isolated as an endophyte from a few plant species. This pest categorisation focuses on the hosts that are relevant for the EU (e.g. Citrus, Ficus, Pinus, Prunus, Pyrus, Quercus and Vitis vinifera). Pyrrhoderma noxium is present in Africa, Central and South America, Asia and Oceania. It has not been reported in the EU. Pyrrhoderma noxium is not included in Commission Implementing Regulation (EU) 2019/2072. Plants for planting (excluding seeds), bark and wood of host plants as well as soil and other growing media associated with plant debris are the main pathways for the entry of the pathogen into the EU. Host availability and climate suitability factors occurring in parts of the EU are favourable for the establishment and spread of the pathogen. The introduction and spread of the pathogen into the EU are expected to have an economic and environmental impact in parts of the territory where hosts are present. Phytosanitary measures are available to prevent the introduction and spread of the pathogen into the EU. Pyrrhoderma noxium satisfies all the criteria that are within the remit of EFSA to assess for this species to be regarded as potential Union quarantine pest.
Keywords: avocado, camellia, Diospyros, eucalyptus, Musa, Phellinus noxius, root rot
1. INTRODUCTION
1.1. Background and Terms of Reference as provided by the requestor
1.1.1. Background
The new Plant Health Regulation (EU) 2016/2031, on the protective measures against pests of plants, is applying from 14 December 2019. Conditions are laid down in this legislation in order for pests to qualify for listing as Union quarantine pests, protected zone quarantine pests or Union regulated non‐quarantine pests. The lists of the EU regulated pests together with the associated import or internal movement requirements of commodities are included in Commission Implementing Regulation (EU) 2019/2072. Additionally, as stipulated in the Commission Implementing Regulation 2018/2019, certain commodities are provisionally prohibited to enter in the EU (high risk plants, HRP). EFSA is performing the risk assessment of the dossiers submitted by exporting to the EU countries of the HRP commodities, as stipulated in Commission Implementing Regulation 2018/2018. Furthermore, EFSA has evaluated a number of requests from exporting to the EU countries for derogations from specific EU import requirements.
In line with the principles of the new plant health law, the European Commission with the Member States are discussing monthly the reports of the interceptions and the outbreaks of pests notified by the Member States. Notifications of an imminent danger from pests that may fulfil the conditions for inclusion in the list of the Union quarantine pest are included. Furthermore, EFSA has been performing horizon scanning of media and literature.
As a follow‐up of the above‐mentioned activities (reporting of interceptions and outbreaks, HRP, derogation requests and horizon scanning), a number of pests of concern have been identified. EFSA is requested to provide scientific opinions for these pests, in view of their potential inclusion by the risk manager in the lists of Commission Implementing Regulation (EU) 2019/2072 and the inclusion of specific import requirements for relevant host commodities, when deemed necessary by the risk manager.
1.1.2. Terms of Reference
EFSA is requested, pursuant to Article 29(1) of Regulation (EC) No 178/2002, to provide scientific opinions in the field of plant health.
EFSA is requested to deliver 53 pest categorisations for the pests listed in Annex 1A, 1B, 1D and 1E (for more details see mandate M‐2021‐00027 on the Open.EFSA portal). Additionally, EFSA is requested to perform pest categorisations for the pests so far not regulated in the EU, identified as pests potentially associated with a commodity in the commodity risk assessments of the HRP dossiers (Annex 1C; for more details see mandate M‐2021‐00027 on the Open.EFSA portal). Such pest categorisations are needed in the case where there are not available risk assessments for the EU.
When the pests of Annex 1A are qualifying as potential Union quarantine pests, EFSA should proceed to phase 2 risk assessment. The opinions should address entry pathways, spread, establishment, impact and include a risk reduction options analysis.
Additionally, EFSA is requested to develop further the quantitative methodology currently followed for risk assessment, in order to have the possibility to deliver an express risk assessment methodology. Such methodological development should take into account the EFSA Plant Health Panel Guidance on quantitative pest risk assessment and the experience obtained during its implementation for the Union candidate priority pests and for the likelihood of pest freedom at entry for the commodity risk assessment of High Risk Plants.
1.2. Interpretation of the Terms of Reference
Phellinus noxius is one of a number of pests listed in Annex 1 to the Terms of Reference (ToR) to be subject to pest categorisation to determine whether it fulfils the criteria of a potential Union quarantine pest for the area of the EU excluding Ceuta, Melilla and the outermost regions of Member States referred to in Article 355(1) of the Treaty on the Functioning of the European Union (TFEU), other than Madeira and the Azores, and so inform EU decision‐making as to its appropriateness for potential inclusion in the lists of pests of Commission Implementing Regulation (EU) 2019/2072. If a pest fulfils the criteria to be potentially listed as a Union quarantine pest, risk reduction options will be identified.
Considering the nomenclature provided by Index Fungorum (https://www.indexfungorum.org/; accessed on 1 September 2023) according to which Pyrrhoderma noxium is the current name of Phellinus noxius on the basis of both morphological and phylogenetic evidence (Zhou et al., 2018) (see Section 3.1.1 on Identity and Taxonomy), the Panel chose to use the name Pyrrhoderma noxium instead of Phellinus noxius throughout the pest categorisation.
1.3. Additional information
This pest categorisation was initiated following the commodity risk assessment of bonsai plants (Pinus parviflora grafted on Pinus thunbergii) from China performed by EFSA (EFSA PLH Panel, 2022), in which P. noxium was identified as a relevant non‐regulated EU pest, which could potentially enter the EU on bonsai plants.
2. DATA AND METHODOLOGIES
2.1. Data
2.1.1. Information on pest status from NPPOs
In the context of the current mandate, EFSA is preparing pest categorisations for new/emerging pests that are not yet regulated in the EU. When official pest status is not available in the European and Mediterranean Plant Protection Organization (EPPO) Global Database (EPPO, online), EFSA consults the NPPOs of the relevant MSs.
2.1.2. Literature search
A literature search on P. noxium was conducted at the beginning of the categorisation in the ISI Web of Science bibliographic database, using the scientific name of the pest as search term. Papers relevant for the pest categorisation were reviewed, and further references and information were obtained from experts, as well as from citations within the references and grey literature. Pest information on hosts and distribution was retrieved through the systematic literature search, using the EPPO Global Database and CABI (2022) as complementary sources.
2.1.3. Database search
Data about the import of commodity types that could potentially provide a pathway for the pest to enter the EU and about the area of hosts grown in the EU were obtained from EUROSTAT (Statistical Office of the European Communities).
The Europhyt and TRACES databases were consulted for pest‐specific notifications on interceptions and outbreaks. Europhyt is a web‐based network run by the Directorate General for Health and Food Safety (DG SANTÉ) of the European Commission as a subproject of PHYSAN (Phyto‐Sanitary Controls) specifically concerned with plant health information. TRACES is the European Commission's multilingual online platform for sanitary and phytosanitary certification required for the importation of animals, animal products, food and feed of non‐animal origin and plants into the European Union, and the intra‐EU trade and EU exports of animals and certain animal products. Up until May 2020, the Europhyt database managed notifications of interceptions of plants or plant products that do not comply with EU legislation, as well as notifications of plant pests detected in the territory of the Member States and the phytosanitary measures taken to eradicate or avoid their spread. The recording of interceptions switched from Europhyt to TRACES in May 2020.
GenBank was searched to determine whether it contained any nucleotide sequences for P. noxium which could be used as reference material for molecular diagnosis. GenBank® (www.ncbi.nlm.nih.gov/genbank/) is a comprehensive publicly available database that as of August 2019 (release version 227) contained over 6.25 trillion base pairs from over 1.6 billion nucleotide sequences for 450,000 formally described species (Sayers et al., 2020).
2.2. Methodologies
The Panel performed the pest categorisation for P. noxium, following guiding principles and steps presented in the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018), the EFSA guidance on the use of the weight of evidence approach in scientific assessments (EFSA Scientific Committee, 2017) and the International Standards for Phytosanitary Measures No. 11 (FAO, 2013).
The criteria to be considered when categorising a pest as a potential Union quarantine pest (QP) is given in Regulation (EU) 2016/2031 Article 3 and Annex I, Section 1 of the Regulation. Table 1 presents the Regulation (EU) 2016/2031 pest categorisation criteria on which the Panel bases its conclusions. In judging whether a criterion is met the Panel uses its best professional judgement (EFSA Scientific Committee, 2017) by integrating a range of evidence from a variety of sources (as presented above in Section 2.1.) to reach an informed conclusion as to whether or not a criterion is satisfied.
TABLE 1.
Pest categorisation criteria under evaluation, as derived from Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column).
| Criterion of pest categorisation | Criterion in Regulation (EU) 2016/2031 regarding union quarantine pest (article 3) |
|---|---|
| Identity of the pest (Section 3.1 ) | Is the identity of the pest clearly defined, or has it been shown to produce consistent symptoms and to be transmissible? |
| Absence/presence of the pest in the EU territory (Section 3.2 ) |
Is the pest present in the EU territory? If present, is the pest in a limited part of the EU or is it scarce, irregular, isolated or present infrequently? If so, the pest is considered to be not widely distributed. |
| Pest potential for entry, establishment and spread in the EU territory (Section 3.4 ) | Is the pest able to enter into, become established in, and spread within, the EU territory? If yes, briefly list the pathways for entry and spread. |
| Potential for consequences in the EU territory (Section 3.5 ) | Would the pests' introduction have an economic or environmental impact on the EU territory? |
| Available measures (Section 3.6 ) | Are there measures available to prevent pest entry, establishment, spread or impacts? |
| Conclusion of pest categorisation (Section 4 ) | A statement as to whether (1) all criteria assessed by EFSA above for consideration as a potential quarantine pest were met and (2) if not, which one(s) were not met |
The Panel's conclusions are formulated respecting its remit and particularly with regard to the principle of separation between risk assessment and risk management (EFSA founding regulation (EU) No 178/2002); therefore, instead of determining whether the pest is likely to have an unacceptable impact, deemed to be a risk management decision, the Panel will present a summary of the observed impacts in the areas where the pest occurs, and make a judgement about potential likely impacts in the EU. While the Panel may quote impacts reported from areas where the pest occurs in monetary terms, the Panel will seek to express potential EU impacts in terms of yield and quality losses and not in monetary terms, in agreement with the EFSA guidance on quantitative pest risk assessment (EFSA PLH Panel, 2018). Article 3 (d) of Regulation (EU) 2016/2031 refers to unacceptable social impact as a criterion for quarantine pest status. Assessing social impact is outside the remit of the Panel.
3. PEST CATEGORISATION
3.1. Identity and biology of the pest
3.1.1. Identity and taxonomy
Is the identity of the pest clearly defined, or has it been shown to produce consistent symptoms and/or to be transmissible?
Yes, the identity of Pyrrhoderma noxium is clearly defined and the pathogen has been shown to produce consistent symptoms and to be transmissible.
Pyrrhoderma noxium (Corner) L.W. Zhou & Y.C. Dai is a basidiomycete plant pathogenic fungus of the order Hymenochaetales and family Hymenochaetaceae (Index Fungorum; accessed on 1 September 2023).
This pathogen was first described as Fomes noxius by Corner in 1932 (Corner, 1932). It was then renamed as Phellinus noxius (Cunningham, 1965), which is the predominant name found in the literature, and thereafter as Phellinidium noxium (Bondartseva et al., 1992). More recently, morphological and phylogenetic molecular analysis reassigned Phellinidium noxium to the genus Pyrrhoderma, as Pyrrhoderma noxium (Zhou et al., 2018). Although this is the current name of the fungus (Index Fungorum; accessed on 1 September 2023), the phylogeny and taxonomy of this species should be further analysed (Stewart et al., 2020; Zhou et al., 2018). Indeed, recent phylogenetic analyses based on ITS (internal transcribed spacer) and LSU (28S nuclear large sub‐unit) rDNA genes suggest that Pyrrhoderma noxium may represent several distinct genetic groups (Garfinkel et al., 2020). Moreover, Stewart et al. (2020) based on their phylogenetic study suggested that Phellinus noxius isolates from eastern Asia and Oceania are distinct from Pyrrhoderma noxium and that Phellinus noxius may represent one or more cryptic species. According to the same authors, these are preliminary results and further analyses were recommended to better resolve P. noxium species boundaries. Indeed, the LSU and ITS markers used in the work of Stewart et al. (2020) yield different clustering patterns, with the LSU showing a greater distinction between Phellinus noxius and Pyrrhoderma noxium than the ITS marker, suggesting that further analyses are needed to better resolve P. noxium species boundaries. The EPPO Global Database (EPPO, online) provides taxonomic identification only for the previous name Phellinus noxius, as followed:
Preferred name: Phellinus noxius (Corner) G. Cunningham
Order: Hymenochaetales
Family: Hymenochaetaceae
Genus: Phellinus
Species: Phellinus noxius
Nevertheless, in this pest categorisation, the Panel adopted the nomenclature provided by Index Fungorum (https://www.indexfungorum.org/; accessed on 1 September 2023) according to which Pyrrhoderma noxium is the current name of Phellinus noxius on the basis of both morphological and phylogenetic evidence (Zhou et al., 2018). As explained in Section 1.2, the Panel chose thus to use the name Pyrrhoderma noxium to refer to the pathogen throughout the pest categorisation.
Synonyms: Fomes noxius Corner (EPPO, online). Additional synonyms listed in Index Fungorum (accessed on 1 September 2023) include Phellinidium noxium (Corner) Bondartseva & S. Herrera, and Phellinus noxius (Corner) G. Cunn.
The EPPO code 1 (EPPO, 2019; Griessinger & Roy, 2015) for this species is PHELNO (EPPO, online).
3.1.2. Biology of the pest
Pyrrhoderma noxium is a facultative pathogen that causes brown root‐rot on a wide range of plant species, including mostly woody species but also herbaceous ones (see Section 3.1.3). It can also live saprophytically on woody substrates in soil and become a parasite when both favourable environmental conditions and hosts are present (Chang, 1996). In addition, this species has been also isolated as an endophyte from a few plant species (Chen et al., 2011), including rice (Absalan et al., 2023).
Infected roots, stumps and woody debris in the soil are reported as the main source of inoculum for infection and as substrates for the long‐term (up to 10 years) survival of P. noxium (Chang, 1996). The fungus does not produce any long‐lived survival structures, such as chlamydospores (Chang, 1996), and it has limited ability to grow in the soil under field conditions without the presence of woody plant debris (Wu et al., 2020). Therefore, soil without infected woody debris cannot be considered as a long‐term reservoir of P. noxium inoculum (Wu et al., 2020). Nevertheless, artificial inoculation experiments showed that basidiospores, arthrospores (formed by fragmentation of the mycelium) and mycelia of P. noxium can survive in soil up to 4.5 months, 3.5 months and 10 weeks, respectively (Chang, 1996).
Although both basidiospores and arthrospores are apparently not suitable structures for long‐term survival, they may play an important role in the long‐distance dispersal of P. noxium via air currents (Chung et al., 2015). This role is mostly ascribed to basidiospores (Chung et al., 2015), since arthrospores have never been observed in nature (Bolland, 1984), but only in axenic culture (Leung et al., 2020; Sahashi et al., 2012). In contrast, basidiocarps and basidiospores of P. noxium have been observed under natural conditions on dead and fallen trees (Chung et al., 2015; Hsiao et al., 2019). Basidiospores of P. noxium can be produced on two different types of fructifications, a flat (resupinate) type and a bracket type, with the flat type being the most frequently found in nature (Hsiao et al., 2019). In addition, these basidiospores can be disseminated by wind (Cannon et al., 2022; Chung et al., 2015). Although the majority of basidiospores travel only few metres from the basidiocarps, it is likely that the effective dispersal range of a low number of basidiospores can be higher, reaching several kilometres (Chung et al., 2015). Basidiospores of P. noxium can directly infect stumps and lower stem/trunk (less than two metres from the ground) through wounds (Ann et al., 2002; Bolland, 1984; Hsiao et al., 2019).
However, infections by means of P. noxium basidiospores can also occur indirectly by first germinating and colonising plant debris in the soil, from which the mycelium grows to infect the lateral and taproots of a neighbouring host plant (Ann et al., 2002; Chung et al., 2015). Indeed, the most common way of infection of P. noxium is either between roots of a living tree and infected debris in the soil or root‐to‐root contact between infected trees and healthy adjacent ones (Ann et al., 2002; Chung et al., 2015). Pyrrhoderma noxium can remain viable in fragments of contaminated roots up to 2 years, and inside the root system of dead trees up to 10 years (Chang, 1996).
Infection by P. noxium usually begins in the roots. This fungus colonises the root system, grows towards the trunk and covers the stem base and the root collar of the tree with a dark brown to blackish mycelial crust (Bolland, 1984). The growth of this mycelial crust can reach heights up to 5 m, but it is more commonly found in the first 0.3–0.9 m from the tree trunk base (Cannon et al., 2022). During the colonisation, P. noxium secretes enzymes that break down the cellulose, haemicellulose and lignin in the woody root tissues, leading to the decay of the root structure (Cannon et al., 2022). This results in a significant reduction in physical support and in disruption of the flow of water and nutrients to the tree, further leading to its decline (Ann et al., 2002). Basidiocarps can develop on standing trees (bracket‐like sporocarp) or trees blown down (resupinate‐like sporocarp) (Bolland, 1984), especially under warm and humid weather conditions (Wu et al., 2020). Both basidiocarps produce airborne spores that can spread around and infect tree stumps and wounds in living trees, leading to the establishment of new infection sites (Ann et al., 2002; Hsiao et al., 2019; Wu et al., 2020).
Plants of all ages are susceptible to P. noxium infection (Ann et al., 2002). Nevertheless, the progression of the disease is generally faster in young trees than in old ones (Gray, 2017). The time from the initial infection to the appearance of visible signs and symptoms of the disease can vary widely depending on various factors, including the host plant species, environmental conditions and virulence of P. noxium genotypes (Cannon et al., 2022). In most tree species, it takes 1–2 months from the initial infection for symptoms to become apparent on the host, causing its death within 2–3 months (quick decline) (Ann et al., 2002; Ann, Lee, & Tsai, 1999). However, in some cases, symptoms of P. noxium‐caused brown root rot disease may occur over periods of a year or more, generally culminating in tree death within 2–3 years (slow decline) (Ann et al., 2002; Ann, Lee, & Tsai, 1999).
In laboratory conditions, and in potato dextrose agar medium, P. noxium can grow at temperatures between 12°C and 36°C, with the optimal temperature near 30°C (Ann, Lee, & Huang, 1999), at which growth rate can reach 35 mm/day (Ann et al., 2002; Ann, Lee, & Huang, 1999). The fungus does not grow below 8°C or above 36°C (Ann, Lee, & Huang, 1999). Pyrrhoderma noxium prefers acid soils, since it has been shown to grow at pH ranging from 3.5 to 7.0, whereas at pH above 7.5, its growth is inhibited in potato dextrose broth (Ann, Lee, & Huang, 1999).
Genomic and transcriptomic studies on P. noxium revealed that this pathogen has at least 488 genes that encode plant cell wall‐degrading enzymes, with slight differences on their expression during colonisation of different wood substrates (Ibarra Caballero et al., 2020). According to the same authors, these genomic features of P. noxium might be responsible for its wide host range and its capacity to quickly kill tree hosts.
3.1.3. Host range/species affected
Pyrrhoderma noxium is an opportunistic pathogen reported to infect a wide range of plant species (Ann, Lee, & Huang, 1999; Chang, 1995b), including more than 430 species, from 246 genera and 85 families (Appendix A). The list of host plants encompasses mostly broad‐leaved and coniferous woody species, and to a lesser extent herbaceous species (Appendix A). The most diversified (with most genera) families among its host range are Fabaceae, Moraceae, Lauraceae, Malvaceae, Euphorbiaceae, Myrtaceae, Meliaceae, Rosaceae, Arecaceae, Rubiaceae, Pinaceae and Rutaceae; while the most diversified (with most species) genera are Ficus, Cinnamomum, Macaranga, Prunus, Acacia, Bauhinia, Eucalyptus, Citrus, Diospyros and Pinus.
As shown in Appendix A, P. noxium has been mostly associated with tropical and subtropical plant species. However, P. noxium hosts also include many plant species with high relevance for the EU, namely Acacia spp. (Ann et al., 2002), Bauhinia spp. (e.g. Ann et al., 2002), Citrus spp. (Stewart et al., 2020; Tsai et al., 2017), Camellia spp. (Ann et al., 2002), Diospyros spp., including D. kaki (Ann et al., 2002; Ann, Lee, & Huang, 1999; Tsai et al., 2017), Eucalyptus spp. (Agustini et al., 2014; Ann et al., 2002; Glen et al., 2014; Hsiao et al., 2019), Ficus spp. (Ann et al., 2002; Brooks, 2002; Gray, 2017; Hsiao et al., 2019; Tsai et al., 2017), Musa spp. (Ivory & Daruhi, 1993; Stewart et al., 2020), Persea americana (Ann et al., 2002; Stewart et al., 2020), Pinus spp. (Abe et al., 1995; Ann et al., 2002), Prunus spp., including P. armeniaca (Tsai et al., 2017) and P. persica (Akiba et al., 2015; Ann et al., 2002; Tsai et al., 2017), Pyrus spp., including P. communis (Ann et al., 2002; Tsai et al., 2017), Quercus sp. (Chung et al., 2015) and Vitis vinifera (Ann et al., 2002; Tsai et al., 2017).
Given that the symptoms of P. noxium in the belowground tree parts and lower stem are rather specific and distinct from those of other root rot pathogens (see Section 3.1.5), there is less uncertainty about its host range than for other pathogens where the identification of hosts cannot be based on visual symptoms and molecular methods are needed to confirm host status (e.g. Pestalotiopsis microspora). However, it is likely that P. noxium can infect other plant species, given the ability of this pathogen to produce a great variety of plant cell wall‐degrading enzymes (see Section 3.1.2), and thus enhancing its capacity to infect diverse hosts.
3.1.4. Intraspecific diversity
Different molecular techniques, such as DNA sequencing and genotyping, have been used to analyse the genetic diversity within and among populations of P. noxium. All those studies found high levels of genetic diversity within populations. For example, population genetic analyses of P. noxium isolates from Taiwan and Japan, using simple sequence repeat (SSR) markers (Akiba et al., 2015; Chung et al., 2015) and whole‐genome sequencing (Chung et al., 2017), revealed a high diversity of genotypes at population level. Similarly, distinct lineages were identified within 95 P. noxium isolates from geographically diverse locations across eastern Asia and Oceania, based on sequences of four nuclear DNA loci (Stewart et al., 2020). In contrast to the nuclear genome, the mitochondrial genome of P. noxium is nearly identical among isolates at protein‐coding regions, but differs greatly in the length of the non‐coding regions (i.e. intergenic sequences) (Lee et al., 2019).
There is evidence of mitochondrial exchange between P. noxium individual fungal cells, possibly occurring during hyphal fusion and mating (Lee et al., 2019). This exchange of mitochondrial material may lead to the formation of recombinant mitotypes (i.e. new combinations of mitochondrial DNA), potentially giving rise to novel genotypes (Lee et al., 2019). The ability of P. noxium to reproduce sexually (Chung et al., 2015, 2017) can also contribute to the increase of genetic diversity within populations. This may have implications on the plasticity and adaptation of the different P. noxium genotypes to various adverse environmental conditions, including fungicide exposure. It can also have important implications for P. noxium virulence. Indeed, differences in virulence have been detected among isolates of P. noxium obtained from either the same or from different host species (Nandris et al., 1987; Sahashi et al., 2010).
3.1.5. Detection and identification of the pest
Are detection and identification methods available for the pest?
Yes, there are methods available for the detection and identification of Pyrrhoderma noxium.
Symptoms and signs
Pyrrhoderma noxium is a pathogen that primarily infects the root system of various tree species, causing brown root‐rot (Ann et al., 2002). Trees infected by P. noxium often exhibit reduced plant growth, yellowing and wilting of leaves, defoliation, branch dieback, leading eventually to plant death within a few months (quick decline) to several years (slow decline) (Ann et al., 2002; Ann, Lee, & Tsai, 1999; Sahashi et al., 2012). In general, young trees showed more rapid death than older trees (Sahashi et al., 2012). However, these aboveground symptoms can vary greatly depending on the tree species, the age and environmental conditions and are visible only at a later stage of infection (Sahashi et al., 2012). In addition, most of those symptoms are similar to those caused by many root rot pathogens (Sahashi et al., 2012). In contrast, the symptoms of P. noxium in the belowground tree parts and lower stem are rather specific and distinct from those of other root rot pathogens. They are typically characterised by a dark brown‐blackish thick mycelial sheath or crust formed on the surface of the roots and lower stem of infected trees (Sahashi et al., 2012). This crust/sheath may also have soil particles and small stones stuck to it when P. noxium is growing in contact with soil (Sahashi et al., 2012). The internal root tissue is brown at first and then turns white and soft, with a network of dark brown lines all over (Chung et al., 2015). In the advanced stages of decay, a thin white to brown mycelial mat forms between the bark and wood (Sahashi et al., 2012). The presence of P. noxium basidiocarps on the basal trunk or exposed roots can also be a sign of brown root‐rot disease (Cannon et al., 2022). However, basidiocarps are not always present in natural conditions, especially during dry periods.
Morphology
Pyrrhoderma noxium can be easily isolated on culture media from roots or lower parts of basal stems exhibiting symptoms (Sahashi et al., 2012). A selective medium for P. noxium was developed by Chang (1995a), using malt extract agar as a basal medium amended with a set of antibiotics and fungicides. When growing in culture, P. noxium colonies exhibit certain characteristics that may aid in its identification. In potato sucrose agar, the mycelial colonies are at first white, turning to brown, with irregular dark brown lines or patches (Sahashi et al., 2012). In potato dextrose agar, the fungus produces brown mycelial colonies with irregular dark brown lines or patches permeating the culture (Ann et al., 2002). Nevertheless, the absence of clamp connections formed during mitosis leading to mycelial growth and the presence of trichocysts (small, hair‐like projection on the surface of fungal cell) and arthrospores in culture are the main typical features of P. noxium (Figure 1) (Ann, Lee, & Huang, 1999; Sahashi et al., 2012). Other species of Pyrrhoderma rarely produce arthrospores in culture (Chung et al., 2015). On the other hand, on sawdust medium, P. noxium produces typical basidiocarps, after 3–4 months (Ann, Lee, & Huang, 1999). They are thin (with about 0.5–2.0 cm thick), hard and uneven, first yellowish‐brown with a white margin and later become brown‐dark grey (Ann et al., 2002; Ann, Lee, & Huang, 1999). Some morphological characteristics of the hymenium and hyphal construction of the basidiocarp can also be used to distinguish P. noxium from its closely related species Pyrrhoderma lamaoense (Ann et al., 2002). Pyrrhoderma noxium can be distinguished from P. lamaoense by having wide setal hyphae (specialised structures distinguished from the vegetative hyphae mostly by thickened walls) and the absence of setae (thick‐walled cystidia) in the hymenium (see Figure 2 in Leung et al., 2020 for morphology pictures of P. noxium examined by light microscopy and scanning electron microscopy) (Abe et al., 1995; Ann et al., 2002; Leung et al., 2020). The basidiospores of P. noxium are smooth, hyaline and ovoid to broadly ellipsoid, averaging from 4.0 × 3.8 μm to 6.0 × 4.8 μm (Ann, Lee, & Huang, 1999). A more detailed morphological description of P. noxium is provided by Abe et al. (1995) and CABI (2022).
FIGURE 1.
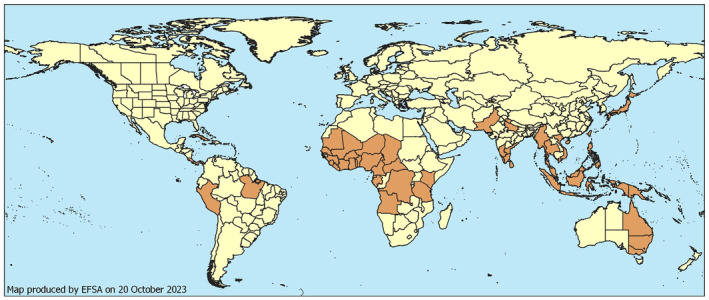
Global distribution of Pyrrhoderma noxium. Sources: systematic literature review (Section 2.1.2) and CABI (2022) (see Appendix B).
DNA‐based identification
The molecular techniques available for the identification of P. noxium are mostly based on the sequencing of the internal transcribed spacers (ITS) of genomic rDNA, in particular the region ITS1–5.8S–ITS2, the nuclear large subunit rDNA (nrLSU) and protein‐coding genes like the translation elongation factor 1‐alpha (EF1‐α) or the second largest subunits of RNA polymerase II (RPB2) (Leung et al., 2020; Stewart et al., 2020; Tsai et al., 2017). As for other fungi, the combined use of these genetic markers, often increase the accuracy of the identification and provides the resolution needed to separate P. noxium from closely related species. Moreover, species‐specific primers based on the ITS region of rDNA were developed for P. noxium (Tsai et al., 2007; Wu et al., 2009). The presence and abundance of P. noxium in root tissues can also be evaluated by quantitative real‐time PCR (Liu et al., 2022) using the specific primers G1F (5’‐GCCCTTTCCTCCGCTTATTG‐3′) and G1R2 (5’‐ATTGGACTTGGGGACTGC‐3′) targeting the ITS region (228 bp) developed by Wu et al. (2011). Nucleotide sequences and whole genome of P. noxium are available in GenBank (www.ncbi.nlm.nih.gov/genbank; 815 sequences retrieved on 5 October 2023) and could be used as reference material for molecular diagnosis.
More recently, loop‐mediated isothermal amplification (LAMP) was developed as a diagnostic tool to detect and identify P. noxium in culture (mycelium) and in wood chips (Zhang et al., 2022).
No EPPO Standard is available for the detection and identification of P. noxium.
3.2. Pest distribution
3.2.1. Pest distribution outside the EU
Pyrrhoderma noxium has been reported to be present in Central (Costa Rica, Cuba, Panama, Puerto Rico) and South America (Brazil, Peru), Africa (Angola, Benin, Burkina Faso, Cameroon, Central African Republic, Chad, Democratic Republic of the Congo, Gabon, Ghana, Guinea, Ivory Coast, Kenya, Liberia, Mali, Mauritania, Nigeria, Senegal, Sierra Leone, Tanzania, Togo, and Uganda), Asia (China, Hong Kong, India, Indonesia, Japan, Malaysia, Myanmar, Pakistan, Philippines, Singapore, Sri Lanka, Taiwan, Thailand, and Vietnam) and Oceania [American Samoa, Australia, Federated Stated of Micronesia (Chuuk, Kosrae, Pohnpei, Yap), Fiji, French Polynesia, Guam, Mariana Island, Niue, Papua New Guinea, Republic of Palau, Republic of Vanuatu, Rota Island, Saipan, Samoa and Solomon Islands]. The current geographical distribution of P. noxium is shown in Figure 1. A list of the countries and states/provinces from where the fungus has been reported is included in Appendix B. The records are based on the systematic literature search (Section 2.1.2), including information from CABI (2022).
3.2.2. Pest distribution in the EU
Is the pest present in the EU territory? If present, is the pest in a limited part of the EU or is it scarce, irregular, isolated or present infrequently? If so, the pest is considered to be not widely distributed.
No. Pyrrhoderma noxium is not known to be present in the EU.
3.3. Regulatory status
3.3.1. Commission Implementing Regulation 2019/2072
Pyrrhoderma noxium is not listed in Annex II of Commission Implementing Regulation (EU) 2019/2072, an implementing act of Regulation (EU) 2016/2031, or in any emergency plant health legislation.
3.3.2. Hosts or species affected that are prohibited from entering the union from third countries
A list of commodities included in Annex VI of Commission Implementing Regulation (EU) 2019/2072 is provided in Table 2. Some of the hosts relevant to the EU, Persea americana, Diospyros spp., Prunus spp. and Quercus spp., are included in the Commission Implementing Regulation (EU) 2018/2019 on high‐risk plants.
TABLE 2.
List of plants, plant products and other objects that are Pyrrhoderma noxium hosts whose introduction into the Union from certain third countries is prohibited (Source: Commission Implementing Regulation (EU) 2019/2072, Annex VI).
| List of plants, plant products and other objects whose introduction into the union from certain third countries is prohibited | |||
|---|---|---|---|
| Description | CN code | Third country, group of third countries or specific area of third country | |
| 1. | Plants of […] Pinus L., […] other than fruit and seeds |
ex 0602 20 20 ex 0602 20 80 ex 0602 90 41 ex 0602 90 45 ex 0602 90 46 ex 0602 90 47 ex 0602 90 50 ex 0602 90 70 ex 0602 90 99 ex 0604 20 20 ex 0604 20 40 |
Third countries other than: Albania, Andorra, Armenia, Azerbaijan, Belarus, Bosnia and Herzegovina, Canary Islands, Faeroe Islands, Georgia, Iceland, Liechtenstein, Moldova, Monaco, Montenegro, North Macedonia, Norway, Russia (only the following parts: Central Federal District (Tsentralny federalny okrug), Northwestern Federal District (Severo‐Zapadny federalny okrug), Southern Federal District (Yuzhny federalny okrug), North Caucasian Federal District (Severo‐Kavkazsky federalny okrug) and Volga Federal District (Privolzhsky federalny okrug)), San Marino, Serbia, Switzerland, Türkiye and Ukraine |
| 2. | Plants of […] and Quercus L., with leaves, other than fruit and seeds |
ex 0602 10 90 ex 0602 20 20 ex 0602 20 80 ex 0602 90 41 ex 0602 90 45 ex 0602 90 46 ex 0602 90 48 ex 0602 90 50 ex 0602 90 70 ex 0602 90 99 ex 0604 20 90 ex 1404 90 00 |
Third countries other than: Albania, Andorra, Armenia, Azerbaijan, Belarus, Bosnia and Herzegovina, Canary Islands, Faeroe Islands, Georgia, Iceland, Liechtenstein, Moldova, Monaco, Montenegro, North Macedonia, Norway, Russia (only the following parts: Central Federal District (Tsentralny federalny okrug), Northwestern Federal District (Severo‐Zapadny federalny okrug), Southern Federal District (Yuzhny federalny okrug), North Caucasian Federal District (Severo‐Kavkazsky federalny okrug) and Volga Federal District (Privolzhsky federalny okrug)), San Marino, Serbia, Switzerland, Turkey and Ukraine |
| 5. |
Isolated bark of Quercus L., other than Quercus suber L. |
ex 1404 90 00 ex 4401 40 90 |
Canada, Mexico, United States |
| 9. | Plants for planting of […] Prunus L. and Pyrus L. and their hybrids […], other than seeds |
ex 0602 10 90 ex 0602 20 20 ex 0602 90 30 ex 0602 90 41 ex 0602 90 45 ex 0602 90 46 ex 0602 90 48 ex 0602 90 50 ex 0602 90 70 ex 0602 90 91 ex 0602 90 99 |
Third countries other than Albania, Algeria, Andorra, Armenia, Australia, Azerbaijan, Belarus, Bosnia and Herzegovina, Canada, Canary Islands, Egypt, Faeroe Islands, Georgia, Iceland, Israel, Jordan, Lebanon, Libya, Liechtenstein, Moldova, Monaco, Montenegro, Morocco, New Zealand, North Macedonia, Norway, Russia (only the following parts: Central Federal District (Tsentralny federalny okrug), Northwestern Federal District (Severo‐ Zapadny federalny okrug), Southern Federal District (Yuzhny federalny okrug), North Caucasian Federal District (Severo‐ Kavkazsky federalny okrug) and Volga Federal District (Privolzhsky federalny okrug)), San Marino, Serbia, Switzerland, Syria, Tunisia, Turkey, Ukraine, the United Kingdom (1) and United States other than Hawaii |
| 10. | Plants of Vitis L., other than fruits |
ex 0602 10 10 ex 0602 20 10 ex 0604 20 90 ex 1404 90 00 |
Third countries other than Switzerland |
| 11. | Plants of Citrus L., [………], and their hybrids, other than fruits and seeds |
ex 0602 10 90 ex 0602 20 20 ex 0602 20 30 ex 0602 20 80 ex 0602 90 45 ex 0602 90 46 ex 0602 90 47 ex 0602 90 50 ex 0602 90 70 ex 0602 90 91 ex 0602 90 99 ex 0604 20 90 ex 1404 90 00 |
All third countries |
| 20. | Growing medium as such, other than soil, consisting in whole or in part of solid organic substances, other than that composed entirely of peat or fibre of Cocos nucifera L., previously not used for growing of plants or for any agricultural purposes |
ex 2530 10 00 ex 2530 90 00 ex 2703 00 00 ex 3101 00 00 ex 3824 99 93 |
Third countries other than Switzerland |
3.4. Entry, establishment and spread in the EU
3.4.1. Entry
Is the pest able to enter into the EU territory? If yes, identify and list the pathways.
Yes. Pyrrhoderma noxium could potentially enter the EU, mainly via host plants for planting (excluding seeds for sowing), parts of host plants (e.g. branches, bark, wood), and soil/plant growing media associated with debris of host plants.
Comment on plants for planting as a pathway.
Plants for planting are a main pathway of entry of the pathogen into the EU.
The Panel identified the following main pathways for the entry of P. noxium into the EU territory:
host plants for planting (excluding seeds for sowing),
bark and wood (timber, logs, sawdust, wooden pallets) of host plants, and
soil and other plant growing media associated with infected host plant debris, all originating in infested third countries.
Pyrrhoderma noxium is reported to infect the roots, base stem/trunk and root collar of plants (Section 3.1.2 Biology of the pest). Thus, the pathogen could potentially enter into the EU territory on plant parts (e.g. stems). However, this is considered a minor pathway for the entry of the pathogen into the EU.
Although there are no data available, basidiospores (and arthrospores if they are produced in natural conditions) of the pathogen may also be present as contaminants on other substrates or objects (e.g. second hand agricultural machinery and equipment, crates, fresh fruit, etc.) imported into the EU from infested countries. Nevertheless, these are considered minor pathways for the entry of the pathogen into the EU.
A list of all the potential pathways for the entry of the pathogen into the EU is included in Table 3.
TABLE 3.
Potential pathways for entry of Pyrrhoderma noxium into the EU.
| Pathways (e.g. host/intended use/source) | Life stage | Relevant mitigations (e.g. prohibitions [Annex VI], special requirements [Annex VII] or phytosanitary certificates [Annex XI] within Implementing Regulation 2019/2072) |
|---|---|---|
| Host plants for planting, other than seeds | Mycelium, basidiospores, arthrospores (if any) | Several main hosts identified in Section 3.1.3 are included in Commission Implementing Regulation 2019/2072. There is a temporary prohibition for high‐risk plants (Regulation 2018/2019) |
| Parts of host plants, other than fruits and seeds (stems, roots) | Mycelium, basidiospores, arthrospores (if any) | A phytosanitary certificate is required for the introduction into the Union from third countries other than Switzerland, of parts of host plants other than fruits and seeds (Annex XI, Part B of Commission Implementing Regulation (EU) 2019/2072) |
| Soil as such not attached or associated with plants for planting | Mycelium, basidiospores, arthrospores (if any) | The introduction into the Union from third countries, other than Switzerland, of soil as such consisting in part of solid organic substances is banned (Annex VI (19) of Commission Implementing Regulation (EU) 2019/2072) |
| Growing medium as such, other than soil not attached or associated with plants for planting | Mycelium, basidiospores, arthrospores (if any) | A phytosanitary certificate is required for the introduction into the Union from third countries, other than Switzerland, of growing medium attached to or associated with plants, intended to sustain the vitality of the plants (Annex XI, Part A (1) of Commission Implementing Regulation (EU) 2019/2072). Special requirements also exist for this commodity (Annex VII (1) of Commission Implementing Regulation (EU) 2019/2072) |
| Growing medium, attached to or associated with host and non‐host plants for planting carrying infected plant debris, with the exception of sterile medium of in vitro plants | Mycelium, basidiospores, arthrospores (if any) | A phytosanitary certificate is required for the introduction into the Union from third countries, other than Switzerland, of growing medium attached to or associated with plants, intended to sustain the vitality of the plants (Annex XI, Part A (1) of Commission Implementing Regulation (EU) 2019/2072). Special requirements also exist for this commodity (Annex VII (1) of Commission Implementing Regulation (EU) 2019/2072) |
| Machinery and vehicles with contaminated soil and/or infected debris of host plants | Mycelium, basidiospores, arthrospores (if any) | A phytosanitary certificate is required for the introduction into the Union from third countries, other than Switzerland, of machinery and vehicles (Annex XI, Part A (1) of Commission Implementing Regulation (EU) 2019/2072). Special requirements also exist for this commodity (Annex VII (2) of Commission Implementing Regulation (EU) 2019/2072) |
The quantity of fresh produce of main hosts imported into the EU from countries where P. noxium is present is provided in Table 4.
TABLE 4.
EU annual imports of fresh produce of main hosts from countries where P. noxium is present, 2016–2020 (in 100 kg) Source: Eurostat, accessed November 2023.
| Commodity | HS code | 2016 | 2017 | 2018 | 2019 | 2020 |
|---|---|---|---|---|---|---|
| Live forest trees | 0602 90 | 56 | 98 | 243 | 138 | 88 |
Notifications of interceptions of harmful organisms began to be compiled in Europhyt in May 1994 and in TRACES in May 2020. As of Nov 2023, there were no record of interception of Pyrrhoderma noxium in the Europhyt and TRACES databases.
3.4.2. Establishment
Is the pest able to become established in the EU territory?
Yes. Both the biotic (host availability) and abiotic (climate suitability) factors occurring in the EU suggest that P. noxium could establish in parts of the EU where hosts are grown.
Following its entry into the EU, P. noxium could establish in parts of the EU where hosts are grown and the climatic conditions are conducive to completing its life cycle. Based on its biology (see Section 3.1.2), P. noxium could potentially be transferred from the pathways of entry to the host plants grown in the EU by root‐to‐root contact, wind, water (irrigation, rain) splash, soil or other plant‐growing media associated with infected plant debris, and possibly insects, as well as with birds and small mammals (see Section 3.4.3). The frequency of this transfer depends on the volume and frequency of the imported commodities, their destination (e.g. nurseries, retailers, packinghouses) and proximity to the hosts, as well as on the management of plant debris.
Climatic mapping is the principal method for identifying areas that could provide suitable conditions for the establishment of a pest taking key abiotic factors into account (Baker, 2002). Availability of hosts is considered in Section 3.4.2.1. Climatic factors are considered in Section 3.4.2.2.
3.4.2.1. EU distribution of main host plants
As noted above and shown in Appendix A, P. noxium has a wide host range, also considering that it is able to colonise several of those plant species endophytically or to survive as a saprophyte in dead plant debris. Some of its main hosts (e.g. Citrus spp., Prunus spp., Pyrus spp., including P. communis, Quercus sp., and Vitis vinifera; see Section 3.1.3) are widely distributed in the EU, both in commercial production (nurseries, open fields, orchards) and in home gardens, parks or forests. The harvested area of most of the main hosts of P. noxium cultivated in the EU in recent years is shown in Table 5.
TABLE 5.
Harvested area of some Pyrrhoderma noxium hosts relevant for the EU, 2017–2021 (1000 ha). Source: EUROSTAT (accessed November 2023).
| Crop | Code | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|---|
| Grapes | W1000 | 3133.32 | 3135.50 | 3155.20 | 3146.24 | 3120.22 |
| Citrus fruits | T0000 | 502.84 | 508.99 | 512.83 | 522.10 | 519.96 |
| Peaches/nectarines | F1210 F1220 | 221.64 | 214.97 | 206.87 | 203.32 | 194.01 |
| Cherries | F1240 | 173.37 | 175.49 | 176.30 | 178.61 | 175.71 |
| Pears | F1120 | 113.81 | 113.54 | 110.66 | 108.29 | 106.96 |
| Apricots | F1230 | 72.23 | 72.57 | 73.22 | 76.13 | 73.48 |
| Avocados | F2300 | 12.72 | 13.22 | 17.50 | 19.58 | 22.86 |
| Bananas | F2400 | 18.91 | 17.94 | 18.27 | 22.11 | 22.01 |
3.4.2.2. Climatic conditions affecting establishment
Based on the data available in the literature on the geographical coordinates of the locations from where P. noxium has been reported, the pathogen is present in non‐EU areas with BSh, Cfa and Cfb, Köppen–Geiger climate zones. These climate zones also occur in the EU, where hosts of P. noxium are grown (Appendix D). Appendix D also provides a Koeppen–Geiger map based on all P. noxium records, also those without local coordinates. For a more accurate description of geographical areas suitable for establishment, a hardiness zone map was generated. This map is based on the 30‐year average absolute minimum temperature and suggests that P. noxium would be able to establish in the EU, although in a restricted area along the Mediterranean (Figure 2). In addition, soil temperature maps (Figures 3 and 4) based on the temperature range (12–36°C) that allow P. noxium to grow (Ann, Lee, & Huang, 1999) also suggest that there are areas in the EU (coastal areas in Mediterranean countries) suitable for the establishment of this pathogen.
FIGURE 2.
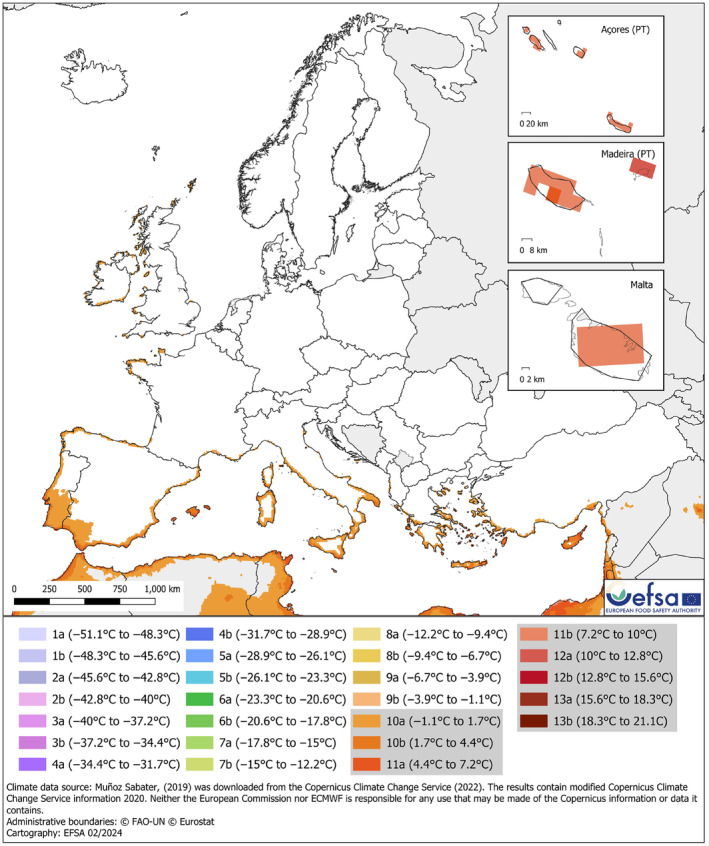
Hardiness zone map based on the average annual minimum temperature for the period 1993–2022. The map highlights the hardiness zones in Europe and some neighbouring areas where the average minimum temperature is higher or equal to the minimum value sampled using the pathogen occurrence. This value is included in the hardiness zones ≥ 10a (highlighted in grey in the legend). The map is based on the implementation of the USDA Plant Hardiness Zones (USDA, 2023).
FIGURE 3.
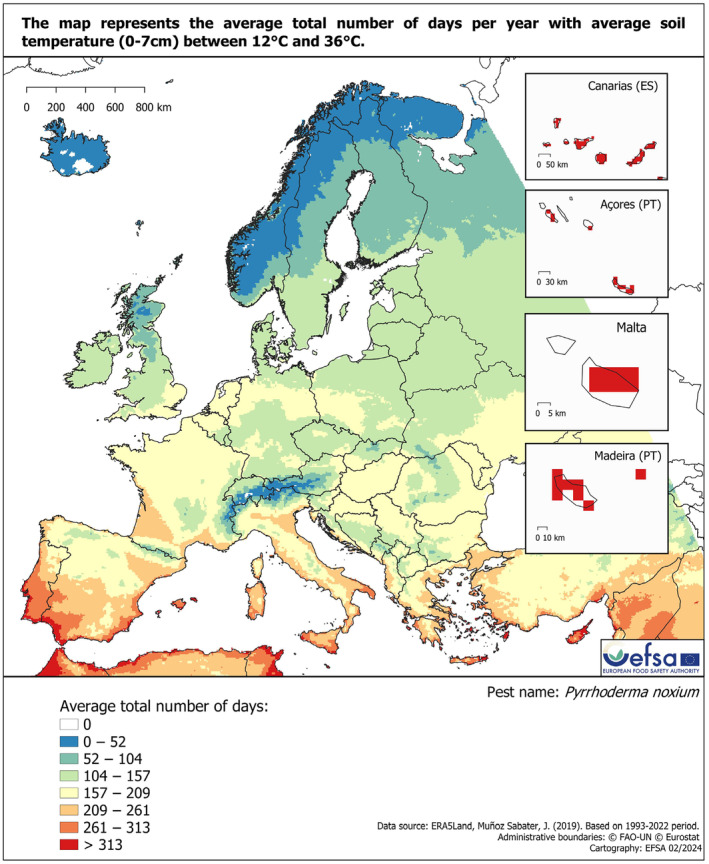
Average total number of days with average soil temperature (0–7 cm) between 12°C and 36°C for Europe and neighbouring areas, with days in seven classes. Data Source: Muñoz‐Sabater et al. (2021). Based on the 1993–2022 period.
FIGURE 4.
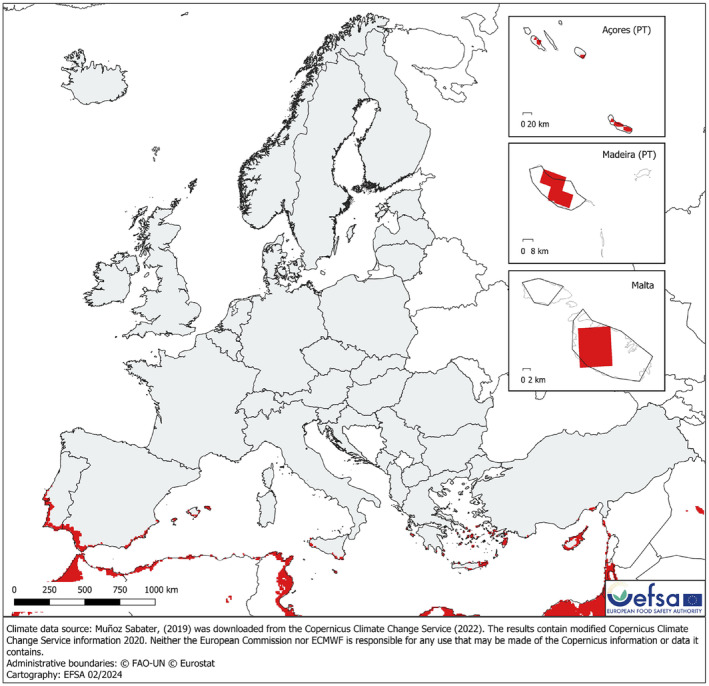
Average total number of days with average soil temperature (0–7 cm) between 12°C and 36°C including only the areas in Europe and neighbouring areas that have average total number of days greater than a threshold of 315 days, obtained from the points of observation. Same data source as Figure 4. Please note that the pest distribution locations from remote islands may not have available information for soil temperature (0–7 cm). This may have an effect on the threshold extracted for this pest.
Regarding the interpretation of the soil temperature map, it should be kept in mind that:
P. noxium is not a strict soilborne pathogen (freely living in the soil), but it is rather associated with plant/wood debris incorporated in the soil;
The temperature data refer to the top layer of soil (0–7 cm), while most ot the roots of trees, and therefore pathogen inoculum, are expected to be present deeper in the soil, where temperatures may be different;
Mycelial growth temperatures could be important to estimate the likelihood of establishment in this case (e.g. root‐to‐root transmission), but temperatures allowing the development of fruiting bodies and sporulation would be even more important. However, in this last case, the air temperature rather than the soil temperature would play a role, but this could be covered by the hardiness zone map (Figure 3).
3.4.3. Spread
Describe how the pest would be able to spread within the EU territory following establishment.
Following its establishment in the EU, Pyrrhoderma noxium could potentially spread within the EU by both natural and human‐assisted means.
Host plants for planting are a main means of spread of P. noxium within the EU.
Pyrrhoderma noxium could potentially spread within the EU by natural and human‐assisted means.
Spread by natural means. The rate of root‐to‐root spread of P. noxium is variable. Under optimal climatic conditions (warm temperature and wet weather), a growth of 6 m per year of the mycelium along a row of susceptible trees is reported (Cannon et al., 2022). This growth rate is likely to be lower under dry conditions and on resistant host trees (Cannon et al., 2022). The survival and short‐distance dispersion of P. noxium depends on infected plant tissues in the soil close to host plants; while basidiospores are more likely to be involved in long‐distance dispersal of the fungus (Hsiao et al., 2019). Although it has not been studied in the case of P. noxium, wind, wind‐driven rain, insects and small animals may also contribute to the dispersal of basidiospores and arthrospores.
Spread by human‐assisted means. The pathogen can spread over long distances through the movement of infected host plants for planting (e.g. rootstocks, grafted plants, scions), including dormant plants, as well as contaminated soil/plant‐growing media associated with plant debris and agricultural machinery, tools, etc. Similarly, infected wood (e.g. timber, logs, sawdust) and wooden components (e.g. pallets), can serve as potential carriers for the spread of P. noxium (Cannon et al., 2022).
3.5. Impacts
Would the pests' introduction have an economic or environmental impact on the EU territory?
Yes, the introduction into and spread within the EU of Pyrrhoderma noxium is expected to have economic and environmental impacts where hosts are grown.
The brown root rot caused by P. noxium has been reported as one of the most serious diseases of trees in the tropics and subtropics (Ann et al., 2002). Indeed, P. noxium is a destructive and fast‐growing pathogen that frequently causes rapid death of a wide range of woody species (Cannon et al., 2022). In Japan, up to 41% tree mortality/decline due to P. noxium was recorded in plantations, encompassing mostly Casuarina equisetifolia, Calophyllum inophyllum, Podocarpus macrophyllus, Garcinia subelliptica, Delonix regia and Erythrina variegata (Abe et al., 1995). In the Ivory Coast, rubber tree (Hevea brasiliensis) plantations, around 25% incidence of trees infected by P. noxium was reported, with 63% mortality (Nandris et al., 1988). Severe P. noxium damage on several woody tree species has also been reported from Taiwan (Chang, 1995b), Mariana Islands (Hodges & Tenorio, 1984), Malaysia (Farid et al., 2005, 2009) and Australia (Bolland, 1984).
Besides plantations and native forest trees, P. noxium also caused significant yield losses in some fruit crop species. For example, in avocado (Persea americana) orchards in Taiwan (Ann et al., 2002) and Australia (Dann et al., 2009), P. noxium was reported to cause considerable losses to avocado growers by killing the trees. In Australia, there are reports of mortality rate of 10% in several avocado orchards, with an estimated economic loss of $AUD5400 per hectare (Dann et al., 2009; Everett & Siebert, 2018). Moreover, P. noxium can persist in the soil, even after infected plants are removed (see Section 3.1.2). This long‐term presence in the environment can continue to affect subsequent crops, leading to continuing yield losses if not properly managed. In tropical countries where P. noxium has been noticed for a long time, it has established a reputation of being a very aggressive pathogen (e.g. Ann, Lee, & Huang, 1999; Chang, 1995b; Farid et al., 2009).
Based on the above, it is expected that the introduction into and spread within the EU of P. noxium would potentially have an economic and environmental impact where hosts are grown. Damage in those areas may be significant with mortality of established trees, and failure of replanting. Pyrrhoderma noxium is a polyphagous pathogen (Ann, Lee, & Huang, 1999; Chang, 1995b), so it may infect many plant species growing in the EU, whose susceptibility to P. noxium is unknown. Should these species be suitable hosts of P. noxium, the economic and environmental impacts are likely to be high as the pathogen can kill host plants.
3.6. Available measures and their limitations
Are there measures available to prevent pest entry, establishment, spread or impacts such that the risk becomes mitigated?
Yes. Although not specifically targeted against Pyrrhoderma noxium, existing phytosanitary measures (see Sections 3.3.2 and 3.4.1) mitigate the likelihood of the pathogen's entry into the EU territory on certain host plants. Potential additional measures are also available to further mitigate the risk of entry, establishment, spread and impacts of the pathogen in the EU (see Section 3.6.1).
3.6.1. Identification of potential additional measures
Phytosanitary measures (prohibitions) are currently applied to some host plants for planting (see Section 3.3.2).
Additional potential risk reduction options and supporting measures are shown in Sections 3.6.1.1 and 3.6.1.2.
3.6.1.1. Additional potential risk reduction options
Potential additional control measures are listed in Table 6.
TABLE 6.
Selected control measures (a full list is available in EFSA PLH Panel, 2018) for pest entry/establishment/spread/impact in relation to currently unregulated hosts and pathways. Control measures are measures that have a direct effect on pest abundance.
| Control measure/risk reduction option (Blue underline = Zenodo doc, Blue = WIP) | RRO summary | Risk element targeted (entry/establishment/spread/impact) |
|---|---|---|
| Require pest freedom | Plants, plant products and other objects must come from a pest‐free country or a pest‐free area or a pest‐free place of production | Entry/Spread |
| Growing plants in isolation |
Description of possible exclusion conditions that could be implemented to isolate the crop from pests and if applicable relevant vectors. E.g. a dedicated structure such as glass or plastic greenhouses Growing nursery plants in isolation may represent an effective control measure |
Entry/Establishment/Spread |
| Managed growing conditions | Proper field drainage, plant distancing, use of pathogen‐free agricultural tools (e.g. pruning scissors, saws and grafting blades), and removal of infected plants and plant debris in the nursery/field/orchard could potentially mitigate the likelihood of infection at origin as well as the spread of the pathogen | Entry/Spread/Impact |
| Crop rotation, associations and density, weed/volunteer control |
Crop rotation, associations and density, weed/volunteer control are used to prevent problems related to pests and are usually applied in various combinations to make the habitat less favourable for pests The measures deal with (1) allocation of crops to field (over time and space) (multi‐crop, diversity cropping) and (2) to control weeds and volunteers as hosts of pests/vectors Although P. noxium has been isolated either as an endophyte or as a pathogen from a wide range of hosts (Appendix A), crop rotation (wherever feasible) may represent an effective means to reduce inoculum sources and potential survival of the pathogen (Ann et al., 2002) |
Establishment/Spread/Impact |
| Use of resistant and tolerant plant species/varieties |
Resistant plants are used to restrict the growth and development of a specified pest and/or the damage they cause when compared to susceptible plant varieties under similar environmental conditions and pest pressure
An approach to control root rot induced by P. noxium is to replant the infested areas with resistant species (Ann, Lee, & Huang, 1999) |
Entry/Establishment/Impact |
| Roguing and pruning |
Roguing is defined as the removal of infested plants and/or uninfested host plants in a delimited area, whereas pruning is defined as the removal of infested plant parts only without affecting the viability of the plant. P. noxium survives as a saprophyte or colonises as an endophyte infected attached plant organs, which can act as inoculum sources. Thus, roguing of host plants may be an effective measure for reducing the inoculum sources and the spread capacity of the pathogen in the field |
Entry/Spread/Impact |
| Biological control, Biopesticides and behavioural manipulation |
Biological control of P. noxium has been investigated at the laboratory scale only on leaf oils obtained from the plant Cinnamomum osmophloeum (Cheng et al., 2018), antagonistic fungi such as Trichoderma asperellum (Chou et al., 2019), bacteria such as Streptomyces sp. and Bacillus sp. (Leung et al., 2020) The potential for biocontrol in the rhizosphere has been demonstrated, particularly with species of Trichoderma (Jacob et al., 1991; Kothandaraman et al., 1991) |
Entry/Impact |
| Chemical treatments on crops including reproductive material | Various fungicides have been found to have activity against the pathogen (Lim et al., 1990; Mappes and Hiepko, 1984), but routine field treatments with these fungicides are not economical | Entry/Establishment/Impact |
| Chemical treatments on consignments or during processing |
Use of chemical compounds that may be applied to plants or to plant products after harvest, during process or packaging operations and storage The treatments addressed in this information sheet are:
As an example, the treatment of pallets can be mentioned |
Entry/Spread |
| Physical treatments on consignments or during processing |
This information sheet deals with the following categories of physical treatments: irradiation/ionisation; mechanical cleaning (brushing, washing); sorting and grading, and; removal of plant parts (e.g. debarking wood). This information sheet does not address: heat and cold treatment (information sheet 1.14); roguing and pruning (information sheet 1.12) Physical treatments (irradiation, mechanical cleaning, sorting, etc.) may reduce or mitigate the risk of entry/spread of P. noxium although no specific information is available for this fungal species |
Entry/Spread |
| Cleaning and disinfection of facilities, tools and machinery |
The physical and chemical cleaning and disinfection of facilities, tools, machinery, transport means, facilities and other accessories (e.g. boxes, pots, pallets, palox, supports, hand tools). The measures addressed in this information sheet are washing, sweeping and fumigation P. noxium infects its host plants mostly through contact of the inoculum with roots. Therefore, and although no specific information is available on this species, cleaning and surface sterilisation of soil tilling tools as well as of equipment and facilities (including premises, storage areas) are good cultural and handling practices employed in the production and marketing of any commodity and may mitigate the likelihood of entry or spread of the pathogen |
Entry/Spread |
| Limits on soil | P. noxium survives in the soil and on plant debris in or on the soil surface. Therefore, plants, plant products and other objects (e.g. used farm machinery) should be free from soil to ensure freedom from P. noxium | Entry/Spread |
| Soil treatment |
The control of soil organisms by chemical and physical methods listed below:
Many soil treatments have been tested and numerous experiments have been performed to find an effective way of eliminating such inoculum. Currently, the most efficient method of destroying the residual inoculum is by flooding the field, and the most practical way is to fumigate the infested soil with ammonia generated from urea amended in soil under alkaline conditions (Ann and Ko, 1994; Chang, 1996; Chang & Chang, 1999) |
Entry/Establishment/Impact |
| Use of non‐contaminated water |
Chemical and physical treatment of water to eliminate waterborne microorganisms. The measures addressed in this information sheet are chemical treatments (e.g. chlorine, chlorine dioxide, ozone); physical treatments (e.g. membrane filters, ultraviolet radiation, heat); ecological treatments (e.g. slow sand filtration) Considering that P. noxium may spread via contaminated irrigation water, physical or chemical treatment of irrigation water may be applied in nurseries and greenhouses. However, also disinfected water, once used to clean plant material, can transfer inoculum from a source to other plants |
Entry/Spread |
| Waste management |
Waste management in authorised facilities and official restriction on its movement may prevent the pathogen from escaping in the environment. On‐site proper management of roguing residues is also recommended as an efficient measure |
Establishment/Spread |
| Heat and cold treatments |
Controlled temperature treatments aimed to kill or inactivate pests without causing any unacceptable prejudice to the treated material itself. The measures addressed in this information sheet are autoclaving; steam; hot water; hot air; cold treatment In laboratory conditions, the fungus does not grow below 8°C or above 36°C (Ann, Lee, & Huang, 1999). Kiln drying of wood and heat treatment of pallets are relevant measures here, although specific data on effectiveness against P. noxium are lacking |
Entry/Spread |
| Conditions of transport |
Specific requirements for mode and timing of transport of commodities to prevent escape of the pest and/or contamination.
If plant material, potentially infected or contaminated with P. noxium (including waste material) must be transported, specific transport conditions (type of packaging/protection, transport means) should be defined to prevent the pathogen from escaping. These may include, albeit not exclusively: physical protection, sorting prior to transport, sealed packaging, etc. |
Entry/Spread |
| Post‐entry quarantine and other restrictions of movement in the importing country |
This information sheet covers post‐entry quarantine (PEQ) of relevant commodities; temporal, spatial and end‐use restrictions in the importing country for import of relevant commodities; Prohibition of import of relevant commodities into the domestic country ‘Relevant commodities’ are plants, plant parts and other materials that may carry pests, either as infection, infestation or contamination Recommended for plant species known to be hosts of P. noxium. This measure does not apply to fruits of host plants |
Establishment/Spread |
3.6.1.2. Additional supporting measures
Potential additional supporting measures are listed in Table 7.
TABLE 7.
Selected supporting measures (a full list is available in EFSA PLH Panel, 2018) in relation to currently unregulated hosts and pathways. Supporting measures are organisational measures or procedures supporting the choice of appropriate risk reduction options that do not directly affect pest abundance.
| Supporting measure | Summary | Risk element targeted (entry/establishment/spread/impact) |
|---|---|---|
| Inspection and trapping | Due to its possible endophytic lifestyle, P. noxium may remain quiescent or latent within asymptomatic host tissues. On symptomatic plants, the symptoms caused by P. noxium are similar to those caused by other root rot pathogens. Therefore, it is unlikely that P. noxium could be detected based on visual inspection only | Entry/Establishment/Spread |
| Laboratory testing | Macroscopic examination of the basidiocarps and of symptoms at the below ground parts of plants, microscopic examination of morphological features such mycelium, trichocysts and basidispores and DNA‐based identification allow the reliable detection and identification of P. noxium (see Section 3.1.5) | Entry/Establishment/Spread |
| Sampling | Necessary as part of other risk reduction options | Entry/Establishment/Spread |
| Phytosanitary certificate and plant passport | Recommended for plant species known to be hosts of P. noxium, including plant parts, but excluding seeds for sowing | Entry/Spread |
| Certified and approved premises | Certified and approved premises may reduce the likelihood of the plants and plant products originating in those premises to be infected by P. noxium | Entry/Spread |
| Certification of reproductive material (voluntary/official) | The risk of entry and/or spread of P. noxium is reduced if host plants for planting, excluding seeds for sowing, are produced under an approved certification scheme and tested free of the pathogen | Entry/Spread |
| Delimitation of Buffer zones | Delimitation of a buffer zone around an outbreak area can prevent spread of the pathogen and maintain a pest‐free area, site or place of production | Spread |
| Surveillance | Surveillance to guarantee that plants and plant products originate from a pest‐free area could be an option | Entry/Establishment/Spread |
3.6.1.3. Biological or technical factors limiting the effectiveness of measures
Latently infected (asymptomatic) host plants and plant products are unlikely to be detected by visual inspection.
The similarity of symptoms of P. noxium with those of other root rot pathogens poses a serious challenge to the detection and identification of the pathogen based solely on visual inspection of the above ground parts of hosts.
The wide host range of the pathogen and its ability to survive endophytically on asymptomatic plants limit the possibility to develop standard diagnostic protocols for all potential hosts.
3.7. Uncertainty
No key uncertainty was identified.
4. CONCLUSIONS
Pyrrhoderma noxium satisfies all the criteria that are within the remit of EFSA to assess for this species to be regarded as potential Union quarantine pest (Table 8).
TABLE 8.
The Panel's conclusions on the pest categorisation criteria defined in Regulation (EU) 2016/2031 on protective measures against pests of plants (the number of the relevant sections of the pest categorisation is shown in brackets in the first column).
| Criterion of pest categorisation | Panel's conclusions against criterion in regulation (EU) 2016/2031 regarding union quarantine pest | Key uncertainties |
|---|---|---|
| Identity of the pest (Section 3.1 ) | The identity of Pyrrhoderma noxium is clearly defined. The pathogen has been shown to produce consistent symptoms and to be transmissible | None |
| Absence/presence of the pest in the EU (Section 3.2 ) | Pyrrhoderma noxium is not known to be present in the EU | None |
| Pest potential for entry, establishment and spread in the EU (Section 3.4 ) | Pyrrhoderma noxium could potentially enter, established in and spread within the EU. The main pathways for entry of the pathogen into the EU are: (i) host plants for planting (ii) bark and wood of host plants and (iii) soil and other plant growing media containing plant debris, all originating in infested third countries. Both the biotic (host availability) and abiotic (climate suitability) factors occurring in parts of the EU where hosts are grown are favourable for the establishment of the pathogen. Following its establishment, the pathogen could spread within the EU by both natural and human‐assisted means | None |
| Potential for consequences in the EU (Section 3.5 ) | Pyrrhoderma noxium introduction into and spread within the EU may have an economic and environmental impact where hosts are grown | None |
| Available measures (Section 3.6 ) | Although not specifically targeted against P. noxium, existing phytosanitary measures mitigate the likelihood of the pathogen's entry, establishment and spread in the EU territory. Potential additional measures also exist to further mitigate the risk of introduction and spread of the pathogen in the EU | None |
| Conclusion (Section 4 ) | Pyrrhoderma noxium satisfies all the criteria that are within the remit of EFSA to assess for this species to be regarded as potential Union quarantine pest | None |
| Aspects of assessment to focus on/scenarios to address in future if appropriate: | Further phylogenetic analyses would make it possible to better resolve P. noxium species boundaries | |
ABBREVIATIONS
- EPPO
European and Mediterranean Plant Protection Organization
- FAO
Food and Agriculture Organization
- HRP
high‐risk plants
- IPPC
International Plant Protection Convention
- ISPM
International Standards for Phytosanitary Measures
- MS
Member State
- PLH
EFSA Panel on Plant Health
- PZ
Protected Zone
- TFEU
Treaty on the Functioning of the European Union
- ToR
Terms of Reference
GLOSSARY
- Containment (of a pest)
Application of phytosanitary measures in and around an infested area to prevent spread of a pest (FAO, 2022).
- Control (of a pest)
Suppression, containment or eradication of a pest population (FAO, 2022).
- Entry (of a pest)
Movement of a pest into an area where it is not yet present, or present but not widely distributed and being officially controlled (FAO, 2022).
- Eradication (of a pest)
Application of phytosanitary measures to eliminate a pest from an area (FAO, 2022).
- Establishment (of a pest)
Perpetuation, for the foreseeable future, of a pest within an area after entry (FAO, 2022).
- Greenhouse
A walk‐in, static, closed place of crop production with a usually translucent outer shell, which allows controlled exchange of material and energy with the surroundings and prevents release of plant protection products (PPPs) into the environment.
- Hitchhiker
An organism sheltering or transported accidentally via inanimate pathways including with machinery, shipping containers and vehicles; such organisms are also known as contaminating pests or stowaways (Toy & Newfield, 2010).
- Impact (of a pest)
The impact of the pest on the crop output and quality and on the environment in the occupied spatial units.
- Introduction (of a pest)
The entry of a pest resulting in its establishment (FAO, 2022).
- Pathway
Any means that allows the entry or spread of a pest (FAO, 2022).
- Phytosanitary measures
Any legislation, regulation or official procedure having the purpose to prevent the introduction or spread of quarantine pests, or to limit the economic impact of regulated non‐quarantine pests (FAO, 2022).
- Quarantine pest
A pest of potential economic importance to the area endangered thereby and not yet present there, or present but not widely distributed and being officially controlled (FAO, 2022).
- Risk reduction option (RRO)
A measure acting on pest introduction and/or pest spread and/or the magnitude of the biological impact of the pest should the pest be present. A RRO may become a phytosanitary measure, action or procedure according to the decision of the risk manager.
- Spread (of a pest)
Expansion of the geographical distribution of a pest within an area (FAO, 2022).
CONFLICT OF INTEREST
If you wish to access the declaration of interests of any expert contributing to an EFSA scientific assessment, please contact interestmanagement@efsa.europa.eu.
REQUESTOR
European Commission
QUESTION NUMBER
EFSA‐Q‐2024‐00349
MAP DISCLAIMER
The designations employed and the presentation of material on any maps included in this scientific output do not imply the expression of any opinion whatsoever on the part of the European Food Safety Authority concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries.
COPYRIGHT FOR NON‐EFSA CONTENT
EFSA may include images or other content for which it does not hold copyright. In such cases, EFSA indicates the copyright holder and users should seek permission to reproduce the content from the original source.
PANEL MEMBERS
Claude Bragard, Paula Baptista, Elisavet Chatzivassiliou, Francesco Di Serio, Paolo Gonthier, Josep Anton Jaques Miret, Annemarie Fejer Justesen, Alan MacLeod, Christer Sven Magnusson, Panagiotis Milonas, Juan A. Navas‐Cortes, Stephen Parnell, Roel Potting, Philippe L. Reignault, Emilio Stefani, Hans‐Hermann Thulke, Wopke Van der Werf, Antonio Vicent Civera, Jonathan Yuen, and Lucia Zappalà.
APPENDIX A. Pyrrhoderma noxium host plants/species affected
A.1.
| Host status | Host name | Plant family | Common name | Reference |
|---|---|---|---|---|
| Cultivated hosts | Acacia auriculiformis | Fabaceae | Earleaf acacia | Lewis and Arentz (1988) |
| Acacia confusa | Fabaceae | Taiwan acacia | Ann et al. (2002), Akiba et al. (2015), Ann, Lee, and Tsai (1999), Hsiao et al. (2019) | |
| Acacia crassicarpa | Fabaceae | Northern Wattle | Ivory and Daruhi (1993) | |
| Acacia excelsa | Fabaceae | Ironwood | Cannon et al. (2022) | |
| Acacia mangium | Fabaceae | Brown salwood | Stewart et al. (2020) | |
| Acacia nilotica | Fabaceae | Gum arabic tree | Kutama et al. (2012) | |
| Acaciella glauca (Syn. Leucaena glauca) | Fabaceae | Amourette, Redwood | Heubel (1939) | |
| Acer saccharum | Aceraceae | Sugar Maple | Ibarra Caballero et al. (2020) | |
| Actinodaphne pedicellata | Lauraceae | Litsea | Ann et al. (2002) | |
| Adenanthera pavonina | Fabaceae | Sandalwood Tree, red‐bead tree | Brooks (2002), Cannon et al. (2022) | |
| Agathis macrophylla | Araucariaceae | Fijian kauri | Neil (1988) | |
| Albizia falcataria (Syn. Falcataria falcata) | Fabaceae | Albizia | Singh and Pandey (1989) | |
| Albizia lebbeck | Fabaceae | Siris tree, Woman's Tongue, East Indian Walnut | Hodges and Tenorio (1984) | |
| Albizia spp. | Fabaceae | Ram (1975) | ||
| Aleurites fordii (Syn. Vernicia fordii) | Euphorbiaceae | Tung oil tree | Ann et al. (2002) | |
| Aleurites moluccanus | Euphorbiaceae | Candle nut tree | Wu et al. (2011) | |
| Aleurites montanus (Syn. Vernicia montana) | Euphorbiaceae | Wood oil tree | Bertus (1935) | |
| Alstonia scholaris | Apocynaceae | Blackboard tree | Ann et al. (2002) | |
| Amherstia nobilis | Fabaceae | Ann, Lee, and Tsai (1999) | ||
| Anacardium occidentale | Anacardiaceae | Cashew nut | Supriadi et al. (2004) | |
| Annona montana | Annonaceae | Mountain soursop | Ann et al. (2002) | |
| Annona squamosa | Annonaceae | Custard apple, Sugar apple | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Annona squamosa × A. cherimola | Annonaceae | Atimoya | Ann et al. (2002) | |
| Antiaris toxicaria | Moraceae | Bark cloth tree | Neil (1988) | |
| Antirhea chinensis | Rubiaceae | Bois goudron |
Wu et al. (2011) |
|
| Aralia elata | Araliaceae | Angelica tree | Sahashi et al. (2007) | |
| Araucaria cunninghamii | Araucariaceae | Hoop pine | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Araucaria heterophylla | Araucariaceae | Norfolk Island pine | Chang and Yang (1998), Ann et al. (2002) | |
| Araucaria hunsteinii | Araucariaceae | Klinki Pine | Singh and Pandey (1989) | |
| Ardisia sieboldii | Primulaceae | Akiba et al. (2015), Sahashi et al. (2015) | ||
| Areca catechu | Arecaceae | Areca‐nut palm | Farr et al. (2021) | |
| Areca triandra | Arecaceae | Singh (1966) | ||
| Argusia argentea | Boraginaceae | Velvet soldierbush | Sahashi et al. (2012) | |
| Artocarpus | Moraceae | Riggenbach (1958) | ||
| Artocarpus altilis | Moraceae | Breadfruit | Hsiao et al. (2019) | |
| Artocarpus heterophyllus | Moraceae | Jack fruit | Ann et al. (2002) | |
| Averrhoa carambola | Oxalidaceae | Carambola, Star fruit | Ann, Lee, and Huang (1999), Ann et al. (2002), Wu et al. (2011), Tsai et al. (2017) | |
| Azadirachta excelsa | Meliaceae | Sentang | Stewart et al. (2020) | |
| Barringtonia asiatica | Lecythidaceae | Sea poison tree | Brooks (2002), Cannon et al. (2022) | |
| Barringtonia petiolata | Lecythidaceae | Ivory and Daruhi (1993) | ||
| Barringtonia samoensis | Lecythidaceae | Brooks (2002), Cannon et al. (2022) | ||
| Bauhinia × blakeana | Fabaceae | Hsiao et al. (2019), Stewart et al. (2020) | ||
| Bauhinia × hybrid | Fabaceae | Butterfly tree | Ann et al. (2002), Tsai et al. (2017) | |
| Bauhinia acuminata | Fabaceae | Dwarf white orchid tree | Farr et al. (2021) | |
| Bauhinia purpurea | Fabaceae | Purple bauhinia | Ann et al. (2002) | |
| Bauhinia racemosa | Fabaceae | Abe et al. (1995) | ||
| Bauhinia sp. | Fabaceae | Ann, Lee, and Tsai (1999) | ||
| Bauhinia variegata | Fabaceae | Orchid tree | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Betula papyrifera | Betulaceae | Paper birch | Nicole et al. (1995) | |
| Bidens pilosa | Asteraceae | Beggartick | Chen et al. (2023) | |
| Bischofia javanica | Phyllanthaceae | Autumn maple tree | Ann et al. (2002), Sahashi et al. (2010, 2015) | |
| Boehmeria nivea | Urticaceae | Ramio, China‐grass | McIntosh (1951) | |
| Bomba × ceiba | Malvaceae | Silk cotton | Ann et al. (2002) | |
| Bridelia monoica | Phyllanthaceae | Pop gun seed | Wu et al. (2011) | |
| Broussonetia kazinoki | Moraceae | Small paper mulberry | Ann et al. (2002) | |
| Broussonetia papyrifera | Moraceae | Paper mulberry | Ann et al. (2002) | |
| Burckella thurstonii | Sapotaceae | Singh et al. (1980) | ||
| Buxus bodinieri | Buxaceae | Sahashi et al. (2012) | ||
| Caesalpinia ferrea | Fabaceae | Brazilian ironwood | Stewart et al. (2020) | |
| Caesalpinia gilliesii | Fabaceae | Bird‐of‐paradise shrub | Adra et al. (2022) | |
| Cajanus cajan | Fabaceae | Bengal pea | Stewart et al. (2020) | |
| Callicarpa japonica | Lamiaceae | Japanese beautyberry | Sahashi et al. (2012) | |
| Calocedrus formosana | Cupressaceae | Taiwan incense cedar | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Calophyllum inophyllum | Calophyllaceae | Indian poon beauty leaf | Abe et al. (1995), Ann et al. (2002), Akiba et al. (2015), Sahashi et al. (2010, 2015) | |
| Calophyllum neoebudicum | Calophyllaceae | Brooks (2002), Cannon et al. (2022) | ||
| Camellia japonica | Theaceae | Camellia | Ann et al. (2002) | |
| Camellia sinensis | Theaceae | Tea | Ann et al. (2002) | |
| Cananga odorata | Annonaceae | Ylang‐ylang | Brooks (2002), Cannon et al. (2022) | |
| Canarium harveyi | Burseraceae | Brooks (2002), Cannon et al. (2022) | ||
| Carica papaya | Caricaceae | Papaya | Neil (1988) | |
| Cassia fistula | Fabaceae | Yellow golden shower tree | Ann et al. (2002), Hsiao et al. (2019) | |
| Cassia grandis | Fabaceae | Horse cassia | Farr et al. (2021) | |
| Cassia siamea | Fabaceae | Cassia tree | Hsiao et al. (2019) | |
| Casuarina equisetifolia | Casuarinaceae | Ironwood tree | Abe et al. (1995), Ann et al. (2002), Sahashi et al. (2010, 2015), Wu et al. (2011), Akiba et al. (2015), Tsai et al. (2017) | |
| Casuarina spp. | Casuarinaceae | Hodges and Tenorio (1984) | ||
| Casuarina torulosa | Casuarinaceae | Australian mahogany | Farr et al. (2021) | |
| Cedrela odorata | Meliaceae | Bastard cedar | Farr et al. (2021) | |
| Cedrela spp. | Meliaceae | Mallet et al. (1985) | ||
| Ceiba pentandra | Malvaceae | Kapok | Ann et al. (2002), Stewart et al. (2020) | |
| Ceiba speciosa | Malvaceae | Chorisia | Akiba et al. (2015) | |
| Celtis boninensis | Cannabaceae | Akiba et al. (2015), Sahashi et al. (2015) | ||
| Celtis sinensis | Cannabaceae | Chinese elm | Tsang et al. (2020) | |
| Celtis sp. | Cannabaceae | Steyaert (1948) | ||
| Cerbera manghas | Apocynaceae | Odollam cerberus tree | Ann et al. (2002), Cannon et al. (2022) | |
| Chamaecyparis formosensis | Cupressaceae | Taiwan red cypress | Ann et al. (2002) | |
| Chamaecyparis obtusa | Cupressaceae | Japanese false cypress | Sahashi et al. (2014) | |
| Chionanthus retusus | Oleaceae | Chinese fringe tree | Chen et al. (2023) | |
| Chorisia speciosa | Malvaceae | Floss silk tree | Ann et al. (2002), Hsiao et al. (2019) | |
| Chrysalidocarpus lutescens | Arecaceae | Yellow areca palm | Ann et al. (2002), Hsiao et al. (2019) | |
| Cinchona spp. | Rubiaceae | Riggenbach (1958) | ||
| Cinnamomum burmannii | Lauraceae | Cinnamon |
Wu et al. (2011), Stewart et al. (2020) |
|
| Cinnamomum camphora | Lauraceae | Camphor | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019), Tsang et al. (2020) | |
| Cinnamomum cassia | Lauraceae | Padang cassia | Chung et al. (2015) | |
| Cinnamomum doederleinii | Lauraceae | Akiba et al. (2015) | ||
| Cinnamomum insularimontanum | Lauraceae | Hsiao et al. (2019) | ||
| Cinnamomum japonicum | Lauraceae | Japanese cinnamon | Sahashi et al. (2010) | |
| Cinnamomum kanehirae | Lauraceae | Stout camphor | Ann et al. (2002), Chung et al. (2015), Hsiao et al. (2019) | |
| Cinnamomum osmophloeum | Lauraceae | Taiwan cinnamon | Tsai et al. (2017) | |
| Cinnamomum pseudopedunculatum | Lauraceae | Akiba et al. (2015), Sahashi et al. (2015) | ||
| Cinnamomum yabunikkei | Lauraceae | Akiba et al. (2015) | ||
| Cinnamomum zeylanicum (Syn. Cinnamomum verum) | Lauraceae | Ceylon cinnamon | Ann et al. (2002) | |
| Cinnamomum kotoense | Lauraceae | Botel tobago cinnamon tree | Tsai et al. (2017) | |
| Citrus limon | Rutaceae | Lemon | Tsai et al. (2017) | |
| Citrus limonia | Rutaceae | Mandarin lime | Stewart et al. (2020) | |
| Citrus spp. | Rutaceae | Farr et al. (2021) | ||
| Cleyera japonica | Pentaphylacaceae | Japanese cleyera | Chung et al. (2015) | |
| Cocos nucifera | Arecaceae | Coconut | Farr et al. (2021) | |
| Codiaeum variegatum | Euphorbiaceae | Croton | Ann et al. (2002) | |
| Coffea arabica | Rubiaceae | Coffee | Ann et al. (2002) | |
| Coffea liberica | Rubiaceae | Mallamaire (1935) | ||
| Cola nitida | Malvaceae | Kola | Adebayo (1975) | |
| Cordia alliodora | Boraginaceae | Ecuador laurel | Neil (1986) | |
| Cordia aspera | Cordiaceae | Brooks (2002), Cannon et al. (2022) | ||
| Cordia dichotoma | Boraginaceae | Cordia | Ann et al. (2002) | |
| Corymbia citriodora (Syn. Eucalyptus citriodora) | Myrtaceae | Lemon gum | Ann et al. (2002), Tsai et al. (2017) | |
| Crossostylis biflora | Rhizophoraceae | Brooks (2002), Cannon et al. (2022) | ||
| Cryptocarya concinnai | Lauraceae | Konishi crytocarya | Ann et al. (2002) | |
| Cryptomeria japonica | Cupressaceae | Japanese cedar | Sahashi et al. (2014) | |
| Cyathea lunulata (Syn. Sphaeropteris lunulata) | Cyatheaceae | Singh et al. (1980) | ||
| Cyathocaly × sp. | Annonaceae | Singh et al. (1980) | ||
| Cycas taitungensis | Cycadaceae | Chang and Yang (1998), Ann, Lee, and Huang (1999) | ||
| Cycas taiwaniana | Cycadaceae | Taiwan cycas | Chang and Yang (1998), Ann et al. (2002) | |
| Dalbergia sissoo | Fabaceae | Sissoo tree | Ann et al. (2002) | |
| Delonix regia | Fabaceae | Flame tree | Abe et al. (1995), Ann et al. (2002), Tsai et al. (2017) | |
| Dillenia biflora | Dilleniaceae |
Singh et al. (1980) |
||
| Dimocarpus longan | Sapindaceae | Longan | Ann et al. (2002), Tsai et al. (2017) | |
| Diospyros decandra | Ebenaceae | Ann, Lee, and Tsai (1999) | ||
| Diospyros egbert‐walkeri (Syn. Diospyros ferrea, D. vera) | Ebenaceae | Sea ebony | Akiba et al. (2015) | |
| Diospyros ferrea var. buxifolia | Ebenaceae | Philippine ebony persimmon | Ann et al. (2002) | |
| Diospyros kaki | Ebenaceae | Persimmon | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Diospyros oldhamii | Ebenaceae | Oldham persimmon | Ann et al. (2002) | |
| Diospyros samoensis | Ebenaceae | Brooks (2002), Cannon et al. (2022) | ||
| Distylium gracile | Hamamelidaceae | Hsiao et al. (2019) | ||
| Distylium lepidotum | Hamamelidaceae | Akiba et al. (2015), Shashi et al. (2015) | ||
| Distylium racemosum | Hamamelidaceae | Isu tree | Akiba et al. (2015) | |
| Dracaena draco | Asparagaceae | Dragon tree | Stewart et al. (2020) | |
| Dracaena fragrans | Asparagaceae | Dracena |
Stewart et al. (2020) |
|
| Dracontomelon vitiense | Anacardiaceae |
Neil (1988) |
||
| Duranta repens (Syn. D. erecta) | Verbenaceae | Creeping sky flower | Ann et al. (2002), Tsai et al. (2017) | |
| Dypsis lutescens | Arecaceae | Yellow butterfly palm | Huang et al. (2015) | |
| Dysoxylum amooroides | Meliaceae |
Ivory and Daruhi (1993) |
||
| Dysoxylum richii | Meliaceae |
Singh et al. (1980) |
||
| Dysoxylum samoense | Meliaceae | Brooks (2002), Cannon et al. (2022) | ||
| Ehretia dichotoma | Ehretiaceae |
Sahashi et al. (2012) |
||
| Ehretia philippinensis | Ehretiaceae | Akiba et al. (2015) | ||
| Elaeagnus rotundata | Elaeagnaceae | Sahashi et al. (2015) | ||
| Elaeis guineensis | Arecaceae | African oil palm | Pinruan et al. (2010) | |
| Elaeis sp. | Arecaceae | Riggenbach (1958) | ||
| Elaeocarpus decipiens (Syn. Elaeocarpus zollingeri) | Elaeocarpaceae | Japanese blueberry tree | Akiba et al. (2015) | |
| Elaeocarpus kambi | Elaeocarpaceae |
Singh et al. (1980) |
||
| Elaeocarpus serratus | Elaeocarpaceae | Ceylon olive | Ann et al. (2002) | |
| Elaeocarpus sylvestris | Elaeocarpaceae | Chung et al. (2015) | ||
| Elateriospermum tapos | Euphorbiaceae | Tapos |
Stewart et al. (2020) |
|
| Elattostachys falcata | Sapindaceae | Brooks (2002), Cannon et al. (2022) | ||
| Endospermum macrophyllum | Euphorbiaceae |
Singh et al. (1980) |
||
| Eriobotrya japonica | Rosaceae | Loquat | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Eriodendron | Malvaceae | Riggenbach (1958) | ||
| Erythrina | Fabaceae | Hodges and Tenorio (1984) | ||
| Erythrina lithosperma (Syn. Erythrina subumbrans) | Fabaceae | Ram (1975) | ||
| Erythrina variegata (Syn. E. indica) | Fabaceae | Indian coral tree | Abe et al. (1995), Akiba et al. (2015), Shashi et al. (2015) | |
| Erythrospermum acuminatissimum | Achariaceae |
Singh et al. (1980) |
||
| Eucalyptus camaldulensis | Myrtaceae | Murray red gum eucalyptus | Ann et al. (2002) | |
| Eucalyptus deglupta | Myrtaceae |
Ivory and Daruhi (1993) |
||
| Eucalyptus grandis | Myrtaceae | Maiden eucalyptus | Ann et al. (2002) | |
| Eucalyptus pellita | Myrtaceae | red mahogany | Agustini et al. (2014), Glen et al. (2014) | |
| Eucalyptus robusta | Myrtaceae | Beakpod eucalyptus | Hsiao et al. (2019) | |
| Eucalyptus spp. | Myrtaceae |
Sahashi et al. (2012), |
||
| Eucalyptus urophylla | Myrtaceae | Ivory and Daruhi (1993) | ||
| Eugenia uniflora | Myrtaceae | Surinam cherry, pitanga | Akiba et al. (2015) | |
| Euonymus boninensis | Celastraceae |
Sahashi et al. (2015) |
||
| Euphorbia pulcherrima | Euphorbiaceae | Ann, Tsai, Wang, and Hsien (1999) | ||
| Ficus benghalensis | Moraceae | Banyan | ||
| Ficus benjamina | Moraceae | Weeping fig | Hsiao et al. (2019) | |
| Ficus carica | Moraceae | Common fig | Cannon et al. (2022) | |
| Ficus drupacea | Moraceae | Chen et al. (2023) | ||
| Ficus elastica | Moraceae | Rubber plant | Ann et al. (2002); Akiba et al. (2015), Sahashi et al. (2015), Hsiao et al. (2019) | |
| Ficus hanceana (Syn. Ficus pumila) | Moraceae | Creeping fig | Chang and Yang (1998) | |
| Ficus macrocarpa | Moraceae | Small‐leafed banyan | Ann et al. (2002) | |
| Ficus macrophylla | Moraceae | Moreton Bay fig | Gray (2017) | |
| Ficus microcarpa | Moraceae | Curtain fig | Akiba et al. (2015), Tsai et al. (2017), Tsang et al. (2020) | |
| Ficus obliqua | Moraceae | Small‐leaved fig | Brooks (2002), Cannon et al. (2022) | |
| Ficus pumila var. awkeotsang (Syn. Ficus awkeotsang) | Moraceae | Jellyfig | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Ficus punctata | Moraceae | Ann, Lee, and Tsai (1999) | ||
| Ficus religiosa | Moraceae | Bo tree fig | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Ficus septica | Moraceae |
Neil (1988) |
||
| Ficus simplicissima | Moraceae | Asian fig | Wu et al. (2011) | |
| Ficus sp. | Moraceae | Brooks (2002), Cannon et al. (2022) | ||
| Ficus superba | Moraceae | Deciduous fig | Chung et al. (2015) | |
| Ficus tinctoria | Moraceae | Dye fig | Brooks (2002), Akiba et al. (2015), Cannon et al. (2022) | |
| Ficus variegata | Moraceae | Red‐stem fig | Wu et al. (2011) | |
| Ficus virgata | Moraceae | Akiba et al. (2015) | ||
| Firmiana simplex | Sterculiaceae | Chinese parasol | Ann et al. (2002) | |
| Flindersia brayleyana | Rutaceae | Singh and Pandey (1989) | ||
| Flemingia macrophylla | Fabaceae | Wild hop | Farr et al. (2021) | |
| Flueggea flexuosa | Phyllanthaceae | Brooks (2002), Cannon et al. (2022) | ||
| Fraxinus formosana | Oleaceae | Formosan ash | Ann et al. (2002), Hsiao et al. (2019) | |
| Garcinia mangostana | Clusiaceae | Mangosteen | CABI (2022) | |
| Garcinia myrtifolia | Clusiaceae |
Singh et al. (1980) |
||
| Garcinia subelliptica | Clusiaceae | Happiness tree, Fukugi tree | Abe et al. (1995), Sahashi et al. (2010), Akiba et al. (2015) | |
| Gardenia jasminoides | Rubiaceae | Cape jasmine | Ann et al. (2002) | |
| Garuga floribunda | Burseraceae |
Neil (1988) |
||
| Gleditsia fera | Fabaceae |
Stewart et al. (2020) |
||
| Gliricidia sepium | Fabaceae | Quick stick |
Neil (1988) |
|
| Glochidion obovatum | Euphorbiaceae | Akiba et al. (2015) | ||
| Glochidion ramiflorum | Phyllanthaceae | Brooks (2002), Cannon et al. (2022) | ||
| Gmelina arborea | Lamiaceae | Snapdragon tree | Brooks (2002) | |
| Grevillea | Proteaceae | Riggenbach (1958) | ||
| Grevillea robusta | Proteaceae | Silver oak | ||
| Guamia mariannae (Syn. Meiogyne cylindrocarpa) | Annonaceae |
Stewart et al. (2020) |
||
| Heliotropium foertherianum | Heliotropiaceae | Tree heliotrope, Argusia | Akiba et al. (2015) | |
| Heritiera littoralis | Malvaceae | Looking glass tree | Chen et al. (2023) | |
| Heritiera sp. | Malvaceae |
Singh et al. (1980) |
||
| Hernandia nymphaeifolia | Hernandiaceae | Sea hearse | Brooks (2002), Cannon et al. (2022) | |
| Hevea brasiliensis | Euphorbiaceae | Rubber tree | Brooks (2002) | |
| Hevea sp. | Euphorbiaceae | Riggenbach (1958) | ||
| Hibiscus glaber | Malvaceae | Dwarf Mahoe | Sahashi et al. (2015) | |
| Hibiscus rosa‐sinensis | Malvaceae | Hibiscus | Abe et al. (1995), Ann et al. (2002), Akiba et al. (2015) | |
| Hibiscus schizopetalus | Malvaceae | Fringed hibiscus | Ann et al. (2002) | |
| Hibiscus tiliaceus | Malvaceae | Linden hibiscus | Ann et al. (2002), Akiba et al. (2015), Cannon et al. (2022) | |
| Hydrangea chinensis | Hydrangeaceae | Chinese hydrangea | Ann et al. (2002) | |
| Ilex mertensii | Aquifoliaceae | Sahashi et al. (2015) | ||
| Ilex rotunda | Aquifoliaceae | Round‐leaf holly | Sahashi et al. (2007) | |
| Inocarpus fagifer | Fabaceae | Brooks (2002), Cannon et al. (2022) | ||
| Intsia bijuga | Fabaceae | Moluccan ironwood | Brooks (2002), Cannon et al. (2022) | |
| Ixora chinensis | Rubiaceae | Jungle flame | Chung et al. (2015) | |
| Ixora x williamsii | Hybrid | Chung et al. (2015) | ||
| Jacaranda sp. | Bignoniaceae | Jacaranda | Adra et al. (2022) | |
| Jatropha integerrima (Syn. J. pandurifolia) | Euphorbiaceae | Peregrina | Shashi et al. (2015) | |
| Juniperus chinensis var. kaizuka | Cupressaceae | Dragon juniper | Tsai et al. (2017) | |
| Keteleeria davidiana var. formosana | Pinaceae | Taiwan keteleeria | Ann et al. (2002) | |
| Khaya ivorensis | Meliaceae | Singh and Pandey (1989) | ||
| Khaya senegalensis | Meliaceae | African mahogany | Burcham et al. (2015), Wang et al. (2016) | |
| Kigelia pinnata (Syn. Kigelia africana) | Bignoniaceae | Sausage tree | Ann et al. (2002), Tsai et al. (2017) | |
| Kleinhovia hospita | Malvaceae | Farr et al. (2021) | ||
| Koelreuteria elegans | Sapindaceae | Flamegold tree |
Stewart et al. (2020) |
|
| Koelreuteria elegans var. formosana (Syn. Koelreuteria henryi) | Sapindaceae | Flame gold rain tree | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Koelreuteria paniculata | Sapindaceae | Golden rain tree | CABI (2022) | |
| Lagerstroemia micrantha | Lythraceae | Chung et al. (2015) | ||
| Lagerstroemia speciosa | Lythraceae | Queen's crepe myrtle | Ann et al. (2002) | |
| Lagerstroemia subcostata | Lythraceae | Sahashi et al. (2007) | ||
| Lagerstroemia turbinate | Lythraceae | Crepe myrtle | Ann et al. (2002) | |
| Lannea coromandelica | Anacardiaceae | Supriadi et al. (2004) | ||
| Lantana camara | Verbenaceae | Lantana | Ann et al. (2002) | |
| Larix kaempferi | Pinaceae | Sahashi et al. (2014) | ||
| Leptopetalum grayi | Rubiaceae | Sahashi et al. (2015) | ||
| Leucaena leucocephala | Fabaceae | White popinac | Ann et al. (2002); Akiba et al. (2015), Sahashi et al. (2015) | |
| Leucaena sp. | Fabaceae | Cannon et al. (2022) | ||
| Ligustrum japonicum | Oleaceae | Japanese privet |
Sahashi et al. (2012) |
|
| Ligustrum micranthum | Oleaceae | Akiba et al. (2015), Shashi et al. (2015) | ||
| Liquidambar formosana | Hamamelidaceae | Formosa sweet gum | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Litchi chinensis | Sapindaceae | Litchi | Ann, Lee, and Huang (1999); Ann et al. (2002); Akiba et al. (2015), Tsai et al. (2017) | |
| Litsea glutinosa | Lauraceae | Indian laurel | Ann et al. (2002) | |
| Litsea hypophaea | Lauraceae | Mountain pepper | Ann et al. (2002) | |
| Litsea japonica | Lauraceae | Litsea | Akiba et al. (2015) | |
| Litsea monopetala | Lauraceae | Many flowered litsea |
Wu et al. (2011) |
|
| Livistona chinensis | Arecaceae | Chinese fan palm |
Wu et al. (2011) |
|
| Lophostemon confertus | Myrtaceae | Zhang et al. (2022) | ||
| Macaranga graeffeana | Euphorbiaceae |
Singh et al. (1980) |
||
| Macaranga harveyana | Euphorbiaceae | Brooks (2002), Cannon et al. (2022) | ||
| Macaranga mappa | Euphorbiaceae | Cannon et al. (2022) | ||
| Macaranga sp. | Euphorbiaceae | Liao et al. (2023) | ||
| Macaranga stipulosa | Euphorbiaceae | Brooks (2002), Cannon et al. (2022) | ||
| Macaranga tanarius | Euphorbiaceae | Macaranga | Ann et al. (2002), Hsiao et al. (2019) | |
| Macaranga tanarius var. tomentosa | Euphorbiaceae | Akiba et al. (2015) | ||
| Macaranga thompsonii | Euphorbiaceae | Pengua tree |
Stewart et al. (2020) |
|
| Machilus chekiangensis | Lauraceae |
Zhang et al. (2022) |
||
| Machilus kobu | Lauraceae | Sahashi et al. (2015) | ||
| Machilus thunbergii (Syn. Persea thunbergii) | Lauraceae | Japanese bay tree, red machilus, tabunoki | Sahashi et al. (2007, 2010), Akiba et al. (2015) | |
| Machilus zuihoensis (Syn. Persea zuihoensis) | Lauraceae | Incense machilus | Ann et al. (2002) | |
| Maesa tenera | Myrsinaceae | Taiwan maesa | Ann et al. (2002) | |
| Mallotus paniculatus | Euphorbiaceae | Turn in the wind | Ann et al. (2002), Tsang et al. (2020) | |
| Malpighia emarginata | Malpighiaceae | Acerola | Sahashi et al. (2015) | |
| Mangifera indica | Anacardiaceae | Mango | Akiba et al. (2015), Sahashi et al. (2015, 2017), Hsiao et al. (2019) | |
| Maytenus diversifolia | Celastraceae |
Sahashi et al. (2012) |
||
| Melaleuca bracteata | Myrtaceae | Black tea tree | Tsai et al. (2017) | |
| Melaleuca leucadendron | Myrtaceae | Weeping paperbark | Ann et al. (2002) | |
| Melia azedarach | Meliaceae | China berry | Ann et al. (2002), Akiba et al. (2015), Hsiao et al. (2019) | |
| Melia azedarach var. subtripinnata | Meliaceae | Sahashi et al. (2010, 2015) | ||
| Melodinus angustifolius | Apocynaceae | Narrow leafed melodinus | Ann et al. (2002) | |
| Metroxylon sp. | Arecaceae |
Neil (1988) |
||
| Michelia compressa | Magnoliaceae | Formosan michelia | Ann et al. (2002) | |
| Michelia figo | Magnoliaceae | Banana magnolia | Ann et al. (2002) | |
| Michelia sp. | Magnoliaceae | Ann, Lee, and Tsai (1999) | ||
| Microcos paniculata | Malvaceae | Garden mint |
Wu et al. (2011) |
|
| Morinda citrifolia | Rubiaceae | Indian mulberry | Brooks (2002), Cannon et al. (2022) | |
| Morus australis | Moraceae | Korean mulberry | Akiba et al. (2015), Sahashi et al. (2015) | |
| Muntingia calabura | Muntingiaceae | Indian cherry | Ann et al. (2002) | |
| Murraya paniculata | Rutaceae | Orange jasmine | Ann et al. (2002), Tsai et al. (2017), Hsiao et al. (2019) | |
| Musa acuminata x Musa balbisiana | Musaceae | Banana |
Stewart et al. (2020) |
|
| Musa spp. | Musaceae |
Ivory and Daruhi (1993) |
||
| Musa textilis | Musaceae | Berwick (1949) | ||
| Myristica castaneifolia | Myristicaceae |
Singh et al. (1980) |
||
| Myristica fatua (Syn. Virola surinamensis) | Myristicaceae | Banak | Brooks (2002), Cannon et al. (2022) | |
| Nageia nagi | Podocarpaceae | Nagi | Chung et al. (2015) | |
| Nandina domestica | Berberidaceae | Orange jessamine | Akiba et al. (2015) | |
| Neolitsea parvigemma | Lauraceae | Small bud neolitsea | Ann et al. (2002) | |
| Neolitsea sericea | Lauraceae | Sahashi et al. (2015) | ||
| Neonauclea forsteri | Rubiaceae | Brooks (2002), Cannon et al. (2022) | ||
| Nerium oleander | Apocynaceae | Oleander | Ann et al. (2002) | |
| Ochroma lagopus | Malvaceae | Balsa tree | Farr et al. (2021) | |
| Ochrosia mariannensis | Apocynaceae | Fago |
Stewart et al. (2020) |
|
| Ochrosia nakaiana | Apocynaceae | Sahashi et al. (2015) | ||
| Oncidium Gower Ramsey | Orchidaceae | Tsai et al. (2017) | ||
| Osmanthus fragrans | Oleaceae | Sweet Osmanthus | Ann et al. (2002), Tsai et al. (2017) | |
| Osmanthus insularis | Oleaceae | Akiba et al. (2015), Sahashi et al. (2015) | ||
| Osmoxylon sp. | Araliaceae |
Neil (1988) |
||
| Pachira macrocarpa (Syn. P. aquatica) | Malvaceae | Malabar chestnut | Ann et al. (2002); Sahashi et al. (2015), Hsiao et al. (2019) | |
| Palaquium formosanum | Sapotaceae | Formosan nato tree | Ann et al. (2002) | |
| Palaquium hornei | Sapotaceae | Singh et al. (1980) | ||
| Pandanus boninensis | Pandanaceae | Sahashi et al. (2015) | ||
| Pangium edule | Achariaceae | Ivory and Daruhi (1993) | ||
| Parinari insularum | Chrysobalanaceae | Singh et al. (1980) | ||
| Parinari laurina (Syn. Atuna excelsa subsp. racemosa) | Chrysobalanaceae | Atuna | Singh et al. (1980) | |
| Passiflora edulis | Passifloraceae | Purple Passion Fruit | Ann, Tsai, Wang, and Hsien (1999) | |
| Persea americana | Lauraceae | Avocado | Ann et al. (2002), Stewart et al. (2020) | |
| Persea kobu (Syn. Machilus kobu) | Lauraceae | Sahashi et al. (2015) | ||
| Phyllanthus myrtifolius | Phyllanthaceae | Ceylon mirtle | Leung et al. (2020) | |
| Pinus caribaea | Pinaceae | Almonicar (1992) | ||
| Pinus elliottii | Pinaceae | Farr et al. (2021) | ||
| Pinus luchuensis | Pinaceae | Luchu pine | Abe et al. (1995) | |
| Pinus ponderosa | Pinaceae | Ponderosa pine | Ibarra Caballero et al. (2020) | |
| Pinus spp. | Pinaceae | Ibarra Caballero et al. (2020) | ||
| Pinus thunbergii (syn. Pinus thunbergiana) | Pinaceae | Black pine | Ann et al. (2002) | |
| Piper nigrum | Pinaceae | Black Pepper | Farr et al. (2021) | |
| Pipturus argenteus | Urticaceae |
Brooks (2002), Cannon et al. (2022) |
||
| Pistacia chinensis | Anacardiaceae | Chinese pistache | Ann et al. (2002) | |
| Pittosporum tobira | Pittosporaceae | Japanese pittosporum | Akiba et al. (2015) | |
| Planchonella grayana | Sapotaceae |
Brooks (2002), Cannon et al. (2022) |
||
| Planchonella obovata | Sapotaceae | Sea gutta | Akiba et al. (2015), Sahashi et al. (2015) | |
| Planchonella samoensis | Sapotaceae |
Brooks (2002), Cannon et al. (2022) |
||
| Planchonella torricellensis | Sapotaceae | Brooks (2002) | ||
| Podocarpus macrophyllus | Podocarpaceae | Yew | Abe et al. (1995), Ann et al. (2002), Sahashi et al. (2010), Tsai et al. (2017) | |
| Pometia pinnata | Sapindaceae | Brooks (2002), Cannon et al. (2022) | ||
| Pongamia pinnata | Fabaceae | Pongamia | Ann et al. (2002) | |
| Populus deltoides | Salicaceae | Poplar | Singh and Pandey (1989) | |
| Prunus armeniaca | Rosaceae | Apricot | Tsai et al. (2017) | |
| Prunus campanulata | Rosaceae | Taiwan cherry | Tsai et al. (2017) | |
| Prunus cerasoides var. campanulata (Syn. Cerasus campanulata) | Rosaceae | Bell‐flowered cherry | Akiba et al. (2015), Sahashi et al. (2015) | |
| Prunus mahaleb | Rosaceae | Mahaleb cherry | Ibarra Caballero et al. (2020) | |
| Prunus mume | Rosaceae | Japanese apricot, plum | Ann et al. (2002), Tsai et al. (2017) | |
| Prunus persica (syn. Amygdalus persica) | Rosaceae | Peach | Ann et al. (2002), Tsai et al. (2017), Akiba et al. (2015) | |
| Prunus serrulata | Rosaceae | Oriental cherry | Chung et al. (2015) | |
| Prunus spp. | Rosaceae | Ibarra Caballero et al. (2020) | ||
| Psidium cattleianum f. lucidum | Myrtaceae | Strawberry guava | Sahashi et al. (2015) | |
| Psidium guajava | Myrtaceae | Guava | Tsai et al. (2017) | |
| Pterocarpus indicus | Fabaceae | Rose wood | Ann et al. (2002), Tsai et al. (2017) | |
| Pterospermum acerifolium | Malvaceae | Chen et al. (2023) | ||
| Pyrus communis | Rosaceae | European pear | Ann, Lee, and Huang (1999) | |
| Pyrus pyrifolia | Rosaceae | Pear | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Pyrus sp. | Rosaceae | Chung et al. (2015) | ||
| Quercus sp. | Fagaceae | Chung et al. (2015) | ||
| Reevesia formosana | Malvaceae | Taiwan Reevesia | Fu and Cheung (2023) | |
| Rhaphiolepis indica var. umbellata | Rosaceae | India hawthorne | Akiba et al. (2015), Sahashi et al. (2015) | |
| Rhaphiolepis umbellata (Syn. Laurus umbellate) | Rosaceae | Japanese‐hawthorn | Sahashi et al. (2010) | |
| Rhododendron obtusum | Ericaceae | Rhododendron | Ann et al. (2002) | |
| Rhododendron simsii | Ericaceae | Sims's azalea | Chung et al. (2015) | |
| Rhus succedanea (Syn. Toxicodendron succedaneum) | Anacardiaceae | Japanese wax tree | Sahashi et al. (2015) | |
| Rhus taitensis | Anacardiaceae | Brooks (2002), Cannon et al. (2022) | ||
| Roystonea regia | Arecaceae | Royal palm | Ann et al. (2002) | |
| Salix babylonica | Salicaceae | Willow | Ann et al. (2002) | |
| Salix nigra | Salicaceae | Black willow | Ibarra Caballero et al. (2020) | |
| Salix spp. | Salicaceae | Ibarra Caballero et al. (2020) | ||
| Samanea saman | Fabaceae | Rain tree | Brooks (2002), Cannon et al. (2022) | |
| Santalum album | Santalaceae | Burgess et al. (2018) | ||
| Schefflera octophylla | Araliaceae | Scheffera | Ann et al. (2002) | |
| Schima mertensiana (Syn. Schima boninensis) | Theaceae | Akiba et al. (2015), Sahashi et al. (2015) | ||
| Schinus terebinthifolia | Anacardiaceae | Brazilian peppertree | Tsai et al. (2017) | |
| Scolopia saeva | Salicaceae | Stewart et al. (2020) | ||
| Sonneratia sp. | Lythraceae | Stewart et al. (2020) | ||
| Spathodea campanulata | Bignoniaceae | African tulip tree | Tsai et al. (2017), Hsiao et al. (2019) | |
| Spondias dulcis | Anacardiaceae | Otaheite apple | Sahashi et al. (2015), Cannon et al. (2022) | |
| Stachytarpheta jamaicensis | Verbenaceae | Jamaica vervain | Sahashi et al. (2015) | |
| Sterculia foetida | Sterculiaceae | Hazel sterculia | Ann et al. (2002) | |
| Sterculia lanceolata | Malvaceae |
Wu et al. (2011), Huang et al. (2016) |
||
| Sterculia nobilis | Malvaceae | Ping pong | Ann et al. (2002), Tsai et al. (2017) | |
| Swietenia macrophylla | Meliaceae | Big leaved mahogany | Hsiao et al. (2019) | |
| Swietenia mahagoni | Meliaceae | Mahogany | Ann et al. (2002) | |
| Syzygium inophylloides | Myrtaceae | Brooks (2002), Cannon et al. (2022) | ||
| Syzygium samarangense | Myrtaceae | Wax apple | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Syzygium sp. | Myrtaceae | Brooks (2002), Cannon et al. (2022) | ||
| Tabebuia chrysantha | Bignoniaceae | Yellow golden bell tree | Ann et al. (2002) | |
| Taiwania cryptomerioides | Chrysomelidae | Taiwania | Ann et al. (2002) | |
| Tamarindus indica | Fabaceae | Tamarind tree | Hsiao et al. (2019) | |
| Taxodium distichum | Cupressaceae | Deciduous cypress | Chung et al. (2015) | |
| Tectona grandis | Lamiaceae | Teak | Hsiao et al. (2019), Stewart et al. (2020) | |
| Tephrosia vogelii | Fabaceae | Beeley (1938) | ||
| Terminalia boivinii | Combretaceae | Bovinii | Ann et al. (2002) | |
| Terminalia calamansanai | Combretaceae | Neil (1988) | ||
| Terminalia catappa | Combretaceae | Indian almond | Ann et al. (2002); Akiba et al. (2015), Sahashi et al. (2015), Tsai et al. (2017) | |
| Terminalia ivorensis | Combretaceae | Ivory Coast almond | Cannon et al. (2022) | |
| Terminalia richii | Combretaceae | Brooks (2002), Cannon et al. (2022) | ||
| Theobroma | Malvaceae | Riggenbach (1958) | ||
| Theobroma cacao | Malvaceae | Cocoa | Brooks (2002) | |
| Thevetia peruviana (Syn. Cascabela thevetia) | Apocynaceae | Lucky‐nut, Milk tree | Ann, Tsai, Wang, and Hsien (1999) | |
| Toona australis | Meliaceae | Neil (1988) | ||
| Toona sinensis (Syn. Cedrela sinensis) | Meliaceae | Chinese cedar | Tsai et al. (2017) | |
| Tournefortia argentea | Boraginaceae | Tree heliotrope | Hsiao et al. (2019) | |
| Trachelospermum asiaticum | Apocynaceae | Asiatic jasmine | Sahashi et al. (2015) | |
| Trema orientalis | Ulmaceae | Charcoal tree | Akiba et al. (2015), Sahashi et al. (2015), Hsiao et al. (2019) | |
| Ulmus parvifolia | Ulmaceae | Chinese elm | Ann et al. (2002) | |
| Veitchia spp. | Arecaceae | Neil (1988) | ||
| Vitis sp. | Vitaceae | Chung et al. (2015) | ||
| Vitis vinifera | Vitaceae | Grape | Ann, Lee, and Huang (1999), Ann et al. (2002), Tsai et al. (2017) | |
| Wikstroemia pseudoretusa | Thymelaeaceae | Sahashi et al. (2015) | ||
| Zanthoxylum ailanthoides var. boninshimae | Rutaceae | Japanese prickly ash | Sahashi et al. (2015) | |
| Zelkova formosana | Ulmaceae | Chung et al. (2015) | ||
| Zelkova serrata | Ulmaceae | Zelkova | Ann et al. (2002), Hsiao et al. (2019) | |
| Zelkova sp. | Ulmaceae | Ann, Lee, and Tsai (1999) | ||
| Ziziphus mauritiana | Rhamnaceae | Indian jujube | Tsai et al. (2017) | |
| Wild weed hosts | Artemisia capillaris | Asteraceae | Wormwood | Ann et al. (2002) |
| Artemisia princeps | Asteraceae | Mugwort | Ann et al. (2002) | |
| Dendrocnide sp. | Urticaceae | Neil (1988) | ||
| Digitaria ciliaris | Poaceae | Chen et al. (2023) | ||
| Ipomoea pes‐caprae | Convolvulaceae | Morning glory | Ann et al. (2002) | |
| Lactuca indica | Asteraceae | Wild lettuce | Chang and Yang (1998), Ann et al. (2002) | |
| Melicope merrilli | Rutaceae | Melicope | Ann et al. (2002) | |
| Oplismenus compositus | Poaceae | Basket grass | Chen et al. (2023) | |
| Paspalum distichum | Poaceae | Chen et al. (2023) | ||
| Saurauia oldhamii (Syn. Saurauia tristyla) | Actinidiaceae | Ann et al. (2002) | ||
| Typhonium blumei | Araceae | Chen et al. (2023) | ||
| Urena lobata | Malvaceae | Cadillo | Ann et al. (2002) | |
| Zoysia matrella | Poaceae | Manila grass | Chen et al. (2023) | |
| Artificial/experimental host | Calophyllum vexans | Calophyllaceae | Abe et al. (1995) | |
| Cassia bicapsularis (Syn. Senna bicapsularis) | Fabaceae | Christmas bush | Ann, Tsai, Wang, and Hsien (1999) | |
| Citrus grandis (Syn. Citrus maxima) | Rutaceae | Pampelmuse | Ann, Tsai, Wang, and Hsien (1999) | |
| Citrus reticulata | Rutaceae | Mandarin orange | Ann, Tsai, Wang, and Hsien (1999) | |
| Citrus sinensis | Rutaceae | Sweet orange | Ann, Tsai, Wang, and Hsien (1999) | |
| Euphoria longana (Syn. Dimocarpus longan) | Sapindaceae | Longan | Ann, Lee, and Huang (1999) | |
| Fagus crenata | Fagaceae | Siebold's beech, Japanese beech | Abe et al. (1995) | |
| Jasminum sambac | Oleaceae | Arabian jasmine | Ann, Tsai, Wang, and Hsien (1999) | |
| Paspalum conjugatum | Poaceae | Chen et al. (2023) | ||
| Quercus acutissima | Fagaceae | Abe et al. (1995) | ||
| Thespesia populnea | Malvaceae | Portia tree | Hodges and Tenorio (1984) |
APPENDIX B. Distribution of Pyrrhoderma noxium
B.1.
| Region | Country | Sub‐national (e.g. state) | Status | References |
|---|---|---|---|---|
| North America | Cuba | Present, no details | CABI (2022) | |
| Puerto Rico | Present, no details | CABI (2022) | ||
| Central America | Costa Rica | Present, no details | CABI (2022) | |
| Panama | Present, no details | Zhou et al. (2018) | ||
| South America | Brazil | Present, no details | Mendes & Urben (2023) | |
| Peru | Present, no details | CABI (1969) | ||
| Africa | Angola | Present, no details | CABI (2022) | |
| Benin | Present, no details | CABI (2022) | ||
| Burkina Faso | Present, no details | CABI (2022) | ||
| Cameroon | Present, no details | Nicole et al. (1985), CABI (2022) | ||
| Central African Republic | Present, no details | CABI (2022) | ||
| Chad | Present, no details | CABI (1969) | ||
| Democratic Republic of the Congo | Present, no details | CABI (2022) | ||
| Gabon | Present, no details | CABI (2022) | ||
| Ghana | Present, no details | CABI (2022) | ||
| Guinea | Present, no details | CABI (1969) | ||
| Ivory Cost | Present, no details | CABI (2022) | ||
| Kenya | Present, no details | CABI (2022) | ||
| Liberia | Present, no details | Nicole et al. (1985) | ||
| Mali | Present, no details | CABI (1969) | ||
| Mauritania | Present, no details | CABI (1969) | ||
| Nigeria | Cross‐River, Ondo, Ikom | Present, no details | CABI (2022) | |
| Senegal | Present, no details | CABI (1969) | ||
| Sierra Leone | Present, no details | CABI (2022) | ||
| Tanzania | Present, no details | CABI (2022) | ||
| Togo | Present, no details | CABI (2022) | ||
| Uganda | Present, no details | CABI (2022) | ||
| Asia | China | Present, no details | Zhou et al. (2018) | |
| Hong Kong | Present, no details | Stewart et al. (2020) | ||
| India | Present, no details | CABI (2022) | ||
| Indonesia | Java, Sumatra | Present, no details | CABI (2022) | |
| Japan | Present, no details | Akiba et al. (2015) | ||
| Malaysia | Present, no details | Stewart et al. (2020) | ||
| Myanmar | Present, no details | CABI (2022) | ||
| Pakistan | Present, no details | CABI (2022) | ||
| Philippines | Present, no details | Glen et al. (2014) | ||
| Singapore | Present, no details | CABI (2022) | ||
| Sri Lanka | Present, no details | Silva et al. (2017) | ||
| Taiwan | Present, no details | Hsiao et al. (2019), Ann et al. (2002) | ||
| Thailand | Present, no details | Samseemoung et al. (2011), Sunthudlakhar et al. (2022) | ||
| Vietnam | Present, no details | CABI (2022) | ||
| Oceania | American Samoa | Present, no details | Brooks (2002), Cannon et al. (2022) | |
| Australia | Present, no details | Stewart et al. (2020) | ||
| Federated Stated of Micronesia | Chuuk, Kosrae, Pohnpei, Yap | Present, no details | Akiba et al. (2015), Cannon et al. (2022) | |
| Fiji | Present, no details | CABI (2022) | ||
| French Polynesia | Tahiti | Present, no details | Mallet et al. (1985) | |
| Guam | Present, no details | Cannon et al. (2022) | ||
| Mariana Island | Present, no details | Cannon et al. (2022) | ||
| Niue | Present, no details | CABI (2022) | ||
| Papua New Guinea | Present, no details | CABI (2022) | ||
| Republic of Palau | Present, no details | Cannon et al. (2022) | ||
| Republic of Vanuatu | Present, no details | CABI (2022) | ||
| Rota Island | Present, no details | Cannon et al. (2022) | ||
| Saipan | Present, no details | Cannon et al. (2022) | ||
| Samoa | Present, no details | CABI (2022) | ||
| Solomon Islands | Present, no details | CABI (2022) |
APPENDIX C. Harvested area of Pyrrhoderma noxium main hosts in the EU MS
C.1.
Harvested area of Pyrrhoderma noxium main hosts in the EU MS, 2017–2021 (1000 ha). Source: EUROSTAT (accessed November 2023).
| Crop | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| Grapes (W1000) | 3133.32 | 3135.50 | 3155.20 | 3146.24 | 3120.22 |
| Belgium | 0.24 | 0.30 | 0.38 | 0.49 | 0.56 |
| Bulgaria | 34.11 | 34.11 | 30.05 | 28.74 | 28.53 |
| Czechia | 15.81 | 15.94 | 16.08 | 16.14 | 16.36 |
| Denmark | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Germany | : | : | : | : | : |
| Estonia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ireland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Greece | 101.75 | 100.34 | 101.85 | 104.21 | 89.84 |
| Spain | 937.76 | 939.92 | 936.89 | 931.63 | 929.39 |
| France | 750.46 | 750.62 | 755.47 | 759.59 | 757.83 |
| Croatia | 21.90 | 20.51 | 19.82 | 21.45 | 21.21 |
| Italy | 670.09 | 675.82 | 697.91 | 703.90 | 702.67 |
| Cyprus | 5.93 | 6.67 | 6.67 | 6.18 | 6.16 |
| Latvia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lithuania | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Luxembourg | 1.26 | 1.25 | 1.24 | 1.24 | 1.23 |
| Hungary | 67.08 | 66.06 | 64.92 | 59.63 | 59.07 |
| Malta | 0.68 | 0.42 | 0.42 | 0.45 | 0.46 |
| Netherlands | 0.16 | 0.17 | 0.16 | 0.17 | 0.19 |
| Austria | 46.33 | 46.50 | 46.36 | 46.16 | 42.84 |
| Poland | 0.67 | 0.73 | 0.74 | 1.00 | 1.00 |
| Portugal | 178.95 | 179.25 | 175.65 | 175.67 | 175.62 |
| Romania | 175.32 | 172.80 | 176.34 | 165.60 | 163.61 |
| Slovenia | 15.86 | 15.65 | 15.57 | 15.29 | 14.90 |
| Slovakia | 8.47 | 8.01 | 7.92 | 7.73 | 7.75 |
| Finland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Sweden | 0.04 | 0.05 | 0.05 | 0.08 | 0.09 |
| Citrus fruits (T0000) | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| EU | 502.84 | 508.99 | 512.83 | 522.10 | 519.96 |
| Greece | 43.47 | 46.26 | 44.23 | 45.62 | 41.16 |
| Spain | 294.26 | 297.62 | 296.48 | 297.97 | 300.50 |
| France | 4.27 | 4.39 | 4.61 | 6.80 | 6.77 |
| Croatia | 2.06 | 1.97 | 2.20 | 2.10 | 2.14 |
| Italy | 135.36 | 134.64 | 140.74 | 145.10 | 144.70 |
| Cyprus | 2.92 | 3.05 | 3.20 | 3.03 | 2.91 |
| Malta | 0.00 | 0.00 | 0.00 | 0.00 | 0.11 |
| Portugal | 20.51 | 21.07 | 21.37 | 21.48 | 21.68 |
| Peaches (F1210) & nectarines (F1220) | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| EU | 221.64 | 214.97 | 206.87 | 203.32 | 194.01 |
| Belgium | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Bulgaria | 3.89 | 3.52 | 3.21 | 2.78 | 2.67 |
| Czechia | 0.37 | 0.38 | 0.34 | 0.34 | 0.32 |
| Denmark | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Germany | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Estonia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ireland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Greece | 41.38 | 42.65 | 41.41 | 45.43 | 38.55 |
| Spain | 84.22 | 80.31 | 77.69 | 72.13 | 72.06 |
| France | 9.32 | 9.10 | 9.03 | 11.66 | 11.77 |
| Croatia | 0.96 | 0.86 | 0.88 | 0.78 | 0.80 |
| Italy | 67.02 | 64.30 | 60.44 | 58.68 | 56.54 |
| Cyprus | 0.30 | 0.29 | 0.31 | 0.34 | 0.35 |
| Latvia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lithuania | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Luxembourg | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hungary | 5.62 | 5.20 | 5.06 | 4.17 | 4.14 |
| Malta | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Netherlands | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Austria | 0.16 | 0.18 | 0.18 | 0.18 | 0.18 |
| Poland | 2.13 | 2.12 | 2.15 | 0.80 | 1.00 |
| Portugal | 3.90 | 3.74 | 3.78 | 3.80 | 3.76 |
| Romania | 1.77 | 1.70 | 1.79 | 1.69 | 1.37 |
| Slovenia | 0.28 | 0.26 | 0.25 | 0.25 | 0.24 |
| Slovakia | 0.32 | 0.36 | 0.35 | 0.31 | 0.28 |
| Finland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Sweden | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Crop | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| Cherries (F1240) | 173.37 | 175.49 | 176.30 | 178.61 | 175.71 |
| Belgium | 1.40 | 1.14 | 1.14 | 1.12 | 1.13 |
| Bulgaria | 10.06 | 11.23 | 12.16 | 11.73 | 11.93 |
| Czechia | 2.11 | 2.07 | 2.16 | 2.15 | 2.12 |
| Denmark | 0.66 | 0.56 | 0.53 | 0.61 | 0.56 |
| Germany | 7.96 | 7.94 | 7.94 | 7.89 | 7.81 |
| Estonia | 0.01 | 0.00 | 0.00 | 0.01 | 0.00 |
| Ireland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Greece | 15.83 | 16.21 | 16.24 | 20.70 | 16.93 |
| Spain | 27.59 | 27.50 | 27.60 | 27.91 | 29.61 |
| France | 8.01 | 8.13 | 8.03 | 7.51 | 7.50 |
| Croatia | 3.53 | 2.94 | 2.85 | 3.12 | 3.20 |
| Italy | 29.27 | 29.16 | 29.21 | 29.01 | 28.06 |
| Cyprus | 0.23 | 0.22 | 0.23 | 0.23 | 0.22 |
| Latvia | 0.10 | 0.10 | 0.12 | 0.10 | 0.10 |
| Lithuania | 0.73 | 0.76 | 0.77 | 0.77 | 0.76 |
| Luxembourg | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hungary | 15.65 | 15.88 | 15.93 | 16.62 | 16.79 |
| Malta | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Netherlands | 0.81 | 0.79 | 0.78 | 0.79 | 0.77 |
| Austria | 0.25 | 0.30 | 0.30 | 0.30 | 0.29 |
| Poland | 36.44 | 36.91 | 37.29 | 35.20 | 35.00 |
| Portugal | 6.30 | 6.14 | 6.50 | 6.49 | 6.41 |
| Romania | 6.02 | 7.06 | 6.09 | 5.94 | 6.12 |
| Slovenia | 0.19 | 0.20 | 0.21 | 0.22 | 0.22 |
| Slovakia | 0.19 | 0.21 | 0.20 | 0.16 | 0.13 |
| Finland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Sweden | 0.03 | 0.03 | 0.03 | 0.04 | 0.05 |
| Pears (F1120) | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| EU | 113.81 | 113.54 | 110.66 | 108.29 | 106.96 |
| Belgium | 10.02 | 10.15 | 10.37 | 10.66 | 10.45 |
| Bulgaria | 0.45 | 0.57 | 0.70 | 0.50 | 0.55 |
| Czechia | 0.71 | 0.75 | 0.80 | 0.83 | 0.80 |
| Denmark | 0.30 | 0.29 | 0.30 | 0.30 | 0.30 |
| Germany | 2.14 | 2.14 | 2.14 | 2.14 | 2.14 |
| Estonia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ireland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Greece | 4.07 | 4.41 | 4.34 | 5.42 | 4.37 |
| Spain | 21.89 | 21.33 | 20.62 | 20.22 | 20.02 |
| France | 5.25 | 5.24 | 5.25 | 5.90 | 5.89 |
| Croatia | 0.71 | 0.80 | 0.86 | 0.73 | 0.75 |
| Italy | 31.73 | 31.34 | 28.71 | 26.60 | 26.79 |
| Cyprus | 0.07 | 0.06 | 0.06 | 0.07 | 0.08 |
| Latvia | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Lithuania | 0.82 | 0.82 | 0.82 | 0.85 | 0.85 |
| Luxembourg | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 |
| Hungary | 2.90 | 2.84 | 2.81 | 2.62 | 2.74 |
| Malta | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Netherlands | 9.70 | 10.00 | 10.09 | 10.00 | 10.07 |
| Austria | 0.46 | 0.49 | 0.50 | 0.54 | 0.55 |
| Poland | 7.26 | 7.30 | 7.22 | 5.80 | 5.60 |
| Portugal | 11.54 | 11.21 | 11.33 | 11.33 | 11.16 |
| Romania | 3.12 | 3.10 | 3.08 | 3.09 | 3.17 |
| Slovenia | 0.20 | 0.21 | 0.21 | 0.22 | 0.23 |
| Slovakia | 0.11 | 0.12 | 0.11 | 0.10 | 0.09 |
| Finland | 0.04 | 0.05 | 0.04 | 0.05 | 0.05 |
| Sweden | 0.12 | 0.11 | 0.10 | 0.11 | 0.11 |
| Apricots (F1230) | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| EU | 72.23 | 72.57 | 73.22 | 76.13 | 73.48 |
| Belgium | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Bulgaria | 2.90 | 2.55 | 2.91 | 1.84 | 3.06 |
| Czechia | 1.10 | 1.15 | 1.15 | 1.17 | 1.12 |
| Denmark | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Germany | 0.23 | 0.23 | 0.23 | 0.23 | 0.23 |
| Estonia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ireland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Greece | 7.31 | 7.94 | 8.35 | 12.24 | 8.96 |
| Spain | 21.00 | 20.57 | 20.24 | 19.78 | 19.44 |
| France | 12.20 | 12.27 | 12.28 | 12.08 | 11.88 |
| Croatia | 0.28 | 0.27 | 0.26 | 0.29 | 0.31 |
| Italy | 17.36 | 17.81 | 17.91 | 17.81 | 17.74 |
| Cyprus | 0.19 | 0.18 | 0.18 | 0.20 | 0.21 |
| Latvia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lithuania | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Luxembourg | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hungary | 4.97 | 5.04 | 4.99 | 5.94 | 6.05 |
| Malta | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Netherlands | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Austria | 0.79 | 0.83 | 0.82 | 0.83 | 0.86 |
| Poland | 0.96 | 0.97 | 1.06 | 0.90 | 0.90 |
| Portugal | 0.56 | 0.56 | 0.54 | 0.52 | 0.53 |
| Romania | 2.11 | 1.97 | 2.04 | 2.03 | 1.92 |
| Slovenia | 0.08 | 0.08 | 0.08 | 0.09 | 0.09 |
| Slovakia | 0.19 | 0.16 | 0.18 | 0.20 | 0.19 |
| Finland | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Sweden | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Crop | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| Avocados (F2300) | 12.72 | 13.22 | 17.50 | 19.58 | 22.86 |
| Greece | 0.60 | 0.72 | 1.08 | 1.10 | 1.93 |
| Spain | 11.81 | 12.16 | 14.10 | 15.85 | 18.06 |
| France | 0.23 | 0.24 | 0.24 | 0.13 | 0.13 |
| Cyprus | 0.08 | 0.10 | 0.10 | 0.16 | 0.17 |
| Portugal | 0.00 | 0.00 | 1.98 | 2.34 | 2.57 |
| Crop | 2017 | 2018 | 2019 | 2020 | 2021 |
|---|---|---|---|---|---|
| Bananas (F2400) | 18.91 | 17.94 | 18.27 | 22.11 | 22.01 |
| Greece | 0.09 | 0.09 | 0.10 | 0.10 | 0.10 |
| Spain | 9.08 | 9.09 | 9.06 | 9.10 | 9.10 |
| France | 8.49 | 7.50 | 7.78 | 11.58 | 11.48 |
| Cyprus | 0.21 | 0.22 | 0.21 | 0.22 | 0.21 |
| Portugal | 1.04 | 1.05 | 1.12 | 1.12 | 1.12 |
APPENDIX D. Köppen–Geiger maps
D.1.
Figure D.1 shows the Köppen–Geiger map for P. noxium based on report locations with geographical coordinates only. Figure D.2 shows the Köppen–Geiger map for P. noxium based on report locations with and without geographical coordinates.
FIGURE D.1.
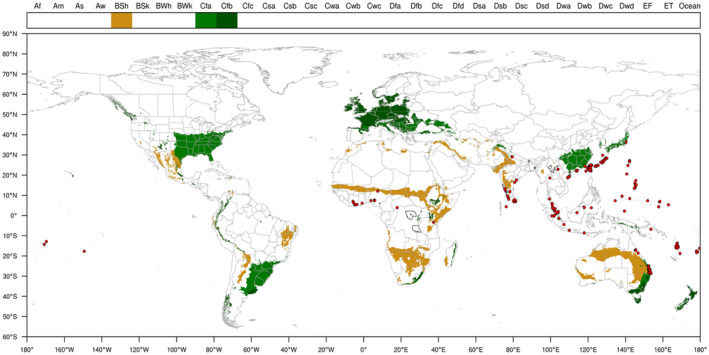
Distribution of three Köppen–Geiger climate types, i.e. BSh, Cfa and Cfb, that occur in the EU and in third countries where Pyrrhoderma noxium has been reported (locations with geographical coordinates only). The legend shows the list of Köppen–Geiger climates. Red dots indicate point locations where P. noxium was reported.
FIGURE D.2.
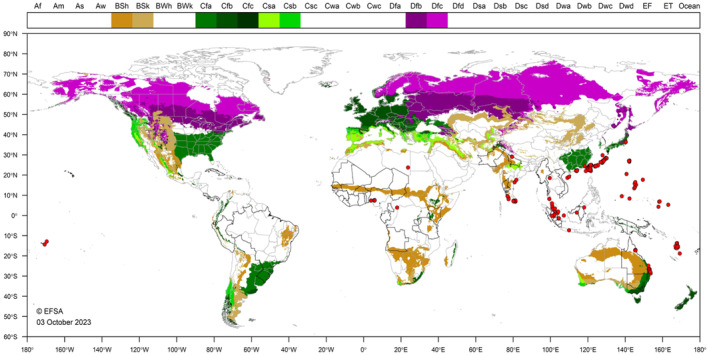
Koeppen–Geiger map based on the pathogen records with local geographical coordinates (as Figure 3), but also on those records without local coordinates, which were thus assigned to large administrative areas (e.g. Pakistan, Japan), and thus include several climate types.
EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C. , Baptista, P. , Chatzivassiliou, E. , Di Serio, F. , Gonthier, P. , Jaques Miret, J. A. , Justesen, A. F. , MacLeod, A. , Magnusson, C. S. , Milonas, P. , Navas‐Cortes, J. A. , Parnell, S. , Potting, R. , Stefani, E. , Thulke, H.‐H. , Van der Werf, W. , Vicent Civera, A. , Yuen, J. , Zappalà, L. … Reignault, P. L. (2024). Pest categorisation of Pyrrhoderma noxium . EFSA Journal, 22 (3), e8667. 10.2903/j.efsa.2024.8667
Adopted: 22 February 2024
Notes
An EPPO code, formerly known as a Bayer code, is a unique identifier linked to the name of a plant or plant pest important in agriculture and plant protection. Codes are based on genus and species names. However, if a scientific name is changed, the EPPO code remains the same. This provides a harmonised system to facilitate the management of plant and pest names in computerised databases, as well as data exchange between IT systems (EPPO, 2019; Griessinger & Roy, 2015).
REFERENCES
- Abe, Y. , Kobayashi, T. , Ohnuki, M. , Hattori, T. , & Tsurumachi, M. (1995). Brown root rot of trees caused by Phellinus noxius in windbreaks on Ishigaki Island, Japan‐incidence of disease, pathogen, and artificial inoculation. Annals of the Phytopathological Society of Japan, 61, 425–433. [Google Scholar]
- Absalan, S. , Jayawardena, R. S. , Bundhun, D. , Jungkhun, N. , & Lumyong, S. (2023). First report of Pyrrhoderma noxium from rice in northern Thailand. Plant Pathology & Quarantine, 13(1), 63–70. 10.5943/ppq/13/1/6 [DOI] [Google Scholar]
- Adebayo, A. A. (1975). Studies on the Fomes root diseases of Kola Cola nitida . Ghana Journal of Agricultural Science, 11–15. [Google Scholar]
- Adra, C. , Panchalingam, H. , & Kurtböke, D. İ. (2022). Termite‐gut‐associated actinomycetes as biological control agents against phytopathogen Pyrrhoderma noxium . Microbiology Australia, 43(4), 190–193. 10.1071/MA22052 [DOI] [Google Scholar]
- Agustini, L. , Francis, A. , Glen, M. , Indrayadi, H. , & Mohammed, C. L. (2014). Signs and identification of fungal root‐rot pathogens in tropical Eucalyptus pellita plantations. Forest Pathology, 44(6), 486–495. 10.1111/efp.12145 [DOI] [Google Scholar]
- Akiba, M. , Ota, Y. , Tsai, I. J. , Hattori, T. , Sahashi, N. , & Kikuchi, T. (2015). Genetic differentiation and spatial structure of Phellinus noxius, the causal agent of brown root rot of woody plants in Japan. PLoS One, 10(10), e0141792. 10.1371/journal.pone.0141792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almonicar, R. S. (1992). Two types of root rot diseases affecting Acacia mangium . Nitrogen Fixing Tree Research Reports, 10, 94–95. [Google Scholar]
- Ann, P. J. , Chang, T. T. , & Ko, W. H. (2002). Phellinus noxius brown root rot of fruit and ornamental trees in Taiwan. Plant Disease, 86, 820–826. 10.1094/PDIS.2002.86.8.820 [DOI] [PubMed] [Google Scholar]
- Ann, P. J. , & Ko, W. H. (1994). Studies on ecology of brown root rot of fruit trees caused by Phellinus noxius and disease control. Plant Pathology Bulletin, 3, 69 (Abstract). [Google Scholar]
- Ann, P. J. , Lee, H. L. , & Huang, T. C. (1999). Brown root rot of 10 species of fruit trees caused by Phellinus noxius in Taiwan. Plant Disease, 83, 746–750. [DOI] [PubMed] [Google Scholar]
- Ann, P. J. , Lee, H. L. , & Tsai, J. N. (1999). Survey of brown root disease of fruit and ornamental trees caused by Phellinus noxius in Taiwan. Plant Pathology Bulletin, 8, 51–60. [Google Scholar]
- Ann, P. J. , Tsai, J. N. , Wang, I. T. , & Hsien, M. L. (1999). Response of fruit trees and ornamental plants to brown root rot disease by artificial inoculation with Phellinus noxius . Plant Pathology Bulletin, 8, 61–66. [Google Scholar]
- Baker, R. H. A. (2002). Predicting the limits to the potential distribution of alien crop pests. In Hallman G. J. & Schwalbe C. P. (Eds.), Invasive arthropods in agriculture: Problems and solutions (pp. 207–241). Science Publishers Inc. [Google Scholar]
- Beeley F, 1938. Annual Report. Pathological Division. Report Rubber Research Institute of Malaya. pp. 128–156.
- Bertus, L. S. (1935). Report on the work of the mycological division. Administration Report Director Agricultural Ceylon, D53–D60. [Google Scholar]
- Berwick, E. J. H. (1949). The removal of standing rubber on coastal clays. Malayan Agricultural Journal, 32, 298–303. [Google Scholar]
- Bolland, L. (1984). Phellinus noxius: Cause of a significant root‐rot in Queensland hoop pine plantations. Australian Forestry, 47, 2–10. 10.1080/00049158.1984.10675972 [DOI] [Google Scholar]
- Bondartseva, M. A. , Herrera, S. D. , & Sandoval, F. C. (1992). Taxonomical problems of the Cuban Hymenochaetaceous fungi. Mikologiia I Fitopatologiia, 26, 1–13. [Google Scholar]
- Brooks, F. E. (2002). Brown root rot disease in American Samoa's tropical rain forests. Pacific Science, 56(4), 377–387. [Google Scholar]
- Burcham, D. C. , Wong, J. Y. , Ali, M. I. M. , Abarrientos, N. V. , Fong, Y. K. , & Schwarze, F. W. M. R. (2015). Characterization of host‐fungus interactions among wood decay fungi associated with Khaya senegalensis (Desr.) a. Juss (Meliaceae) in Singapore. Forest Pathology, 45(6), 492–504. [Google Scholar]
- Burgess, T. I. , Howard, K. , Steel, E. , & Barbour, E. L. (2018). To prune or not to prune; pruning induced decay in tropical sandalwood. Forest Ecology and Management, 430, 204–218. [Google Scholar]
- CABI . (1969). Fomes noxius Corner [on Hevea brasiliensis and other species]. Distribution maps of plant diseases No 104. CAB International. [Google Scholar]
- CABI . (2022). Phellinus noxius (brown tea root disease). CABI Compendium. 10.1079/cabicompendium.40154 [DOI]
- Cannon, P. G. , Klopfenstein, N. B. , Kim, M.‐S. , Stewart, J. E. , & Chung, C.‐L. (2022). Brown Root Rot Disease Caused by Phellinus noxius in U.S.‐Affiliated Pacific Islands. Forest Service U.S. Department of Agriculture, General Technical Report PNW‐GTR‐1006, September 2022. https://www.fs.usda.gov/pnw/pubs/pnw_gtr1006.pdf
- Chang, T. T. (1995a). A selective medium for Phellinus noxius . European Journal of Forest Pathology, 25, 185–190. [Google Scholar]
- Chang, T. T. (1995b). Decline of nine tree species associated with brown root rot caused by Phellinus noxius in Taiwan. Plant Disease, 79, 962–965. [Google Scholar]
- Chang, T.‐T. (1996). Survival of Phellinus noxius in soil and in the roots of dead host plants. Phytopathology, 86, 272–276. [Google Scholar]
- Chang, T. T. , & Chang, R. J. (1999). Generation of volatile ammonia from urea fungicidal to Phellinus noxius in infested wood in soil under controlled conditions. Plant Pathology, 48, 337–344. [Google Scholar]
- Chang, T. T. , & Yang, W. W. (1998). Phellinus noxius in Taiwan: Distribution, host plants and the pH and texture of the rhizosphere soils of infected hosts. Mycological Research, 102(9), 1085–1088. [Google Scholar]
- Chen, C. Y. , Wu, Z. C. , Liu, T. Y. , Yu, S. S. , Tsai, J. N. , Tsai, Y. C. , Tsai, I. J. , & Chung, C. L. (2023). Investigation of asymptomatic infection of Phellinus noxius in herbaceous plants. Phytopathology, 113, 460–469. 10.1094/PHYTO-08-22-0281-R [DOI] [PubMed] [Google Scholar]
- Chen, X. Y. , Qi, Y. D. , Wei, J. H. , Zhang, Z. , Wang, D. L. , Feng, J. D. , & Gan, B. C. (2011). Molecular identification of endophytic fungi from medicinal plant Huperzia serrata based on rDNA ITS analysis. World Journal of Microbiology and Biotechnology, 27(3), 495–503. 10.1007/s11274-010-0480-x [DOI] [Google Scholar]
- Cheng, S. S. , Lin, C. Y. , Chung, M. J. , Chen, Y. J. , & Chang, S. T. (2018). Potential source of environmentally benign antifungal agents from Cinnamomum osmophloeum leaves against Phellinus noxius . Plant Protection Science, 55(1), 43–53. [Google Scholar]
- Chou, H. , Xiao, Y. T. , Tsai, J. N. , Li, T. T. , Wu, H. Y. , Liu, L. Y. , Tzeng, D. S. , & Chung, C. L. (2019). In vitro and in planta evaluation of Trichoderma asperellum TA as a biocontrol agent against Phellinus noxius, the cause of brown root rot disease of trees. Plant Disease, 103(11), 2733–2741. [DOI] [PubMed] [Google Scholar]
- Chung, C.‐L. , Huang, S.‐Y. , Huang, Y.‐C. , Tzean, S.‐S. , Ann, P.‐J. , Tsai, J.‐N. , Yang, C.‐C. , Lee, H.‐H. , Huang, T.‐W. , Huang, H.‐Y. , Chang, T.‐T. , Lee, H.‐L. , & Liou, R.‐F. (2015). The genetic structure of Phellinus noxius and dissemination pattern of Brown Root Rot Disease in Taiwan. PLoS ONE, 10(10), e0139445. 10.1371/journal.pone.0139445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung, C. L. , Lee, T. J. , Akiba, M. , Lee, H. H. , Kuo, T. H. , Liu, D. , Ke, H. M. , Yokoi, T. , Roa, M. B. , Lu, M. J. , Chang, Y. Y. , Ann, P. J. , Tsai, J. N. , Chen, C. Y. , Tzean, S. S. , Ota, Y. , Hattori, T. , Sahashi, N. , Liou, R. F. , … Tsai, I. J. (2017). Comparative and population genomic landscape of Phellinus noxius: A hypervariable fungus causing root rot in trees. Molecular Ecology, 26, 6301–6316. 10.1111/mec.14359 [DOI] [PubMed] [Google Scholar]
- Copernicus Climate Change Service , 2022. ERA5‐Land hourly data from 1950 to present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). 10.24381/cds.e2161bac [cited within Fig. 4]. [DOI]
- Corner, E. J. H. (1932). The identification of the brown root fungus. Gardens' Bulletin, Strait Settlements, 5(5), 317–350. [Google Scholar]
- Cunningham, G. H. (1965). Polyporaceae of New Zealand. New Zealand Department of Scientific and Industrial Research Bulletin, 164, 221–222. [Google Scholar]
- Dann, E. , Smith, L. , Pegg, K. , Grose, M. , & Pegg, G. (2009). Report on Phellinus noxius, the cause of brown rot, in Australian avocados. Talking Avocados, 20, 28–34. [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Bragard, C. , Baptista, P. , Chatzivassiliou, E. , Di Serio, F. , Jaques Miret, J. A. , Justesen, J.‐A. F. , MacLeod, A. , Magnusson, C. S , Milonas, P. , Navas‐Cortes, J. A. , … Gonthier, P. (2022). Commodity risk assessment of bonsai plants from China consisting of Pinus parviflora grafted on Pinus thunbergii. EFSA Journal, 20(2), 7077. 10.2903/j.efsa.2022.7077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA PLH Panel (EFSA Panel on Plant Health) , Jeger, M. , Bragard, C. , Caffier, D. , Candresse, T. , Chatzivassiliou, E. , Dehnen‐Schmutz, K. , Gregoire, J.‐C. , Jaques Miret, J. A. , MacLeod, A. , Navajas Navarro, M. , Niere, B. , Parnell, S. , Potting, R. , Rafoss, T. , Rossi, V. , Urek, G. , Van Bruggen, A. , Van Der Werf, W. , … Gilioli, G. (2018). Guidance on quantitative pest risk assessment. EFSA Journal, 16(8), 5350. 10.2903/j.efsa.2018.5350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EFSA Scientific Committee , Hardy, A. , Benford, D. , Halldorsson, T. , Jeger, M. J. , Knutsen, H. K. , More, S. , Naegeli, H. , Noteborn, H. , Ockleford, C. , Ricci, A. , Rychen, G. , Schlatter, J. R. , Silano, V. , Solecki, R. , Turck, D. , Benfenati, E. , Chaudhry, Q. M. , Craig, P. , … Younes, M. (2017). Scientific opinion on the guidance on the use of the weight of evidence approach in scientific assessments. EFSA Journal, 15(8), 4971. 10.2903/j.efsa.2017.4971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- EPPO (European and Mediterranean Plant Protection Organization) . (2019). EPPO codes. https://www.eppo.int/RESOURCES/eppo_databases/eppo_codes
- EPPO (European and Mediterranean Plant Protection Organization) . (online). EPPO Global Database. https://gd.eppo.int
- Everett, K. R. , & Siebert, B. (2018). Exotic plant disease threats to the New Zealand avocado industry and climatic suitability: A review. New Zealand Plant Protection, 71, 25–38. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations) . (2013). ISPM (International Standards for Phytosanitary Measures) 11—Pest risk analysis for quarantine pests. FAO, Rome, 36 pp. https://www.ippc.int/sites/default/files/documents/20140512/ispm_11_2013_en_2014‐04‐30_201405121523‐494.65%20KB.pdf
- FAO (Food and Agriculture Organization of the United Nations) . (2022). International Standards for Phytosanitary Measures. ISPM 5 Glossary of phytosanitary terms. FAO, Rome. https://www.fao.org/3/mc891e/mc891e.pdf
- Farid, A. M. , Lee, S. S. , Maziah, Z. , & Patahayah, M. (2009). Pathogenicity of Rigidoporus microporus and Phellinus noxius against four major plantation tree species in peninsular Malaysia. Journal of Tropical Forest Science, 21(4), 289–298. [Google Scholar]
- Farid, A. M. , Lee, S. S. , Maziah, Z. , Rosli, H. , & Norwati, M. (2005). Basal root rot, a new disease of teak (Tectona grandis) in Malaysia caused by Phellinus noxius . Malaysian Journal of Microbiology, 1(2), 40–45. [Google Scholar]
- Farr, D. F. , Rossman, A. Y. , & Castlebury Lisa, A. (2021). United States National Fungus Collections Fungus‐Host Dataset. Ag Data Commons. 10.15482/USDA.ADC/1524414 [DOI] [Google Scholar]
- Fu, C.‐H. , & Cheung, W.‐Y. (2023). The first report of brown root rot disease caused by Phellinus noxius on Reevesia formosana which raises concerns on the conservation of R. Formosana and Formotosena seebohmi (Hemiptera: Cicadoidea) in Taiwan. Taiwania, 68(3), 377–382. [Google Scholar]
- Garfinkel, A. R. , Cannon, P. G. , Klopfenstein, N. B. , Stewart, J. E. , & Kim, M.‐S. (2020). Identification of genetic groups within the invasive brown root rot pathogen, Pyrrhoderma noxium (formerly Phellinus noxius). In Reynolds G. J., Wilhelmi N. P., & Palacios P. (Eds.), Proceeding of the 66th Western International Forest Disease Work Conference; 3‐7 June 2019; Estes Park, CO. WIFDWC (pp. 141–145). www.wifdwc.org [Google Scholar]
- Glen, M. , Yuskianti, V. , Puspitasari, D. , Francis, A. , Agustini, L. , Rimbawanto, A. , Indrayadi, H. , Gafur, A. , & Mohammed, C. L. (2014). Identification of basidiomycete fungi in Indonesian hardwood plantations by DNA barcoding. Forest Pathology, 44(6), 496–508. [Google Scholar]
- Gray, P. (2017). Brown root rot Phellinus noxius. pp. 1–7. https://www.northerntreecare.com.au/wp‐content/uploads/2021/09/Brown‐Root‐Rot.pdf
- Griessinger, D. , & Roy, A.‐S. (2015). EPPO codes: a brief description. https://www.eppo.int/media/uploaded_images/RESOURCES/eppo_databases/A4_EPPO_Codes_2018.pdf
- Heubel, G. A. (1939). A concise report on the plantation crops in the south Sumatra area during 1938. Bergcultures, 13, 768–782. [Google Scholar]
- Hodges, C. S. , & Tenorio, J. A. (1984). Root disease of Delonix regia and associated tree species in the Mariana Islands caused by Phellinus noxius. Plant Disease, 68(4), 334–336. 10.1094/PD-69-334 [DOI] [Google Scholar]
- Hsiao, W.‐W. , Hung, T.‐H. , & Sun, E.‐J. (2019). Newly discovered basidiocarps of Phellinus noxius on 33 tree species with brown root rot disease in Taiwan and the basidiospore variations in growth rate. Taiwania, 64, 263–268. [Google Scholar]
- Huang, H. , Sun, L. H. , Bi, K. , Zhong, G. H. , & Hu, M. Y. (2016). The effect of phenazine‐1‐carboxylic acid on the morphological, physiological, and molecular characteristics of Phellinus noxius . Molecules, 21(5), 613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, Y. , Chang, T. , Chung, C. , & Liou, R. (2015). Genetic diversity of Phellinus noxius from Taipei and Yilan of Taiwan. Plant Pathology Bulletin, 24(2), 77–88. [Google Scholar]
- Ibarra Caballero, J. R. , Ata, J. P. , Leddy, K. A. , Glenn, T. C. , Kieran, T. J. , Klopfenstein, N. B. , Kim, M.‐S. , & Stewart, J. E. (2020). Genome comparison and transcriptome analysis of the invasive brown root rot pathogen, Phellinus noxius, from different geographic regions reveals potential enzymes associated with degradation of different wood substrates. Fungal Biology, 124(2), 144–154. 10.1016/j.funbio.2019.12.007 [DOI] [PubMed] [Google Scholar]
- Ivory, M. H. , & Daruhi, G. (1993). Outbreaks and new records: Vanuatu: New host records for Phellinus noxius in Vanuatu. FAO (Food and Agriculture Organization of the United Nations) Plant Protection Bulletin, 41, 37–38. [Google Scholar]
- Jacob, C. K. , Annajutty, J. , & Jayarathnam, K. (1991). Effect of fungal antagonists on Phellinus noxius causing brown root disease of Hevea. Indian Journal of Natural Rubber Research, 4(2), 142–145. [Google Scholar]
- Kothandaraman, R. , Kochuthresiamma, J. , Mathew, J. , & Rajalakshmi, V. K. (1991). Actinomycete population in the rhizosphere of Hevea and its inhibitory effect on Phellinus noxius . Indian Journal of Natural Rubber Research, 4(2), 150–152. [Google Scholar]
- Kutama, A. S. , Saratu, A. O. , & Asmu, A. U. (2012). Occurrence of wood‐and root‐rot basidiomycetes on trees in Bayero university, Kano, Nigeria. Bayero Journal of Pure and Applied Sciences, 5, 26–30. 10.4314/bajopas.v5i2.4 [DOI] [Google Scholar]
- Lee, H.‐H. , Ke, H.‐M. , Lin, C.‐Y. I. , Lee, T. J. , Chung, C.‐L. , & Tsai, I. J. (2019). Evidence of extensive intraspecific noncoding reshuffling in a 169‐kb mitochondrial genome of a Basidiomycetous fungus. Genome Biology and Evolution, 11(10), 2774–2788. 10.1093/gbe/evz181 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung, K. T. , Chen, C. Y. , You, B. J. , Lee, M. H. , & Huang, J. W. (2020). Brown root rot disease of Phyllanthus myrtifolius: The causal agent and two potential biological control agents. Plant Disease, 104(11), 3043–3053. 10.1094/PDIS-02-20-0412-RE [DOI] [PubMed] [Google Scholar]
- Lewis, D. J. , & Arentz, F. (1988). Technical note on a computer intensive analysis method. Klinkii Lp, Papua New Guinea. Forestry Society of Papua new Guinea University of Technology, 3(4), 60–61. [Google Scholar]
- Liao, T. Z. , Chen, Y. H. , Tsai, J. N. , Chao, C. , Huang, T. P. , Hong, C. F. , Wu, Z. C. , Tsai, I. J. , Lee, H. H. , Klopfenstein, N. B. , Kim, M. S. , Stewart, J. E. , Atibalentja, N. , Brooks, F. E. , Cannon, P. G. , Farid, A. M. , Hattori, T. , Kwan, H. S. , Lam, R. Y. C. , … Chung, C. L. (2023). Translocation of fungicides and their efficacy in controlling Phellinus noxius, the cause of Brown root rot disease. Plant Disease, 107, 2039–2053. [DOI] [PubMed] [Google Scholar]
- Lim, T. K. , Hamm, R. T. , & Mohamad, R. B. (1990). Persistency and volatile behaviour of selected chemicals in treated soil against three basidiomycetous root disease pathogens. International Journal of Pest Management, 36(1), 23–26. [Google Scholar]
- Liu, T. Y. , Chen, C. H. , Yang, Y. L. , Tsai, I. J. , Ho, Y. N. , & Chung, C. L. (2022). The brown root rot fungus Phellinus noxius affects microbial communities in different root‐associated niches of Ficus trees. Environmental Microbiology, 24(1), 276–297. 10.1111/1462-2920.15862 [DOI] [PubMed] [Google Scholar]
- Mallamaire, A. (1935). On some root rots in the Ivory Coast. Revue de Botanique Appliquee, 15, 603–608. [Google Scholar]
- Mallet, B. , Geiger, J. P. , Nandris, D. , Nicole, M. , Renard, J. L. , & Van Canh, T. (1985). Root rot fungi in West Africa. Agronomie (Paris), 5, 553–554. [Google Scholar]
- Mappes, D. , & Hiepko, G. (1984). New possibilities for controlling root diseases of plantation crops. Mededelingen Van de Faculteit Landbouwwetenschappen Rijksuniversiteit Gent, 49(2a), 283–292. [Google Scholar]
- McIntosh, A. E. S. (1951). Annual report of the Department of Agriculture, Malaya, for the Year 1949, 87.
- Mendes, M. A. S. , & Urben, A. F. (2023). Fungos relatados em plantas no Brasil, Laboratório de Quarentena Vegetal. Embrapa Recursos Genéticos e Biotecnologia. https://pragawall.cenargen.embrapa.br/aiqweb/michtml/fgbanco01.asp [Google Scholar]
- Muñoz Sabater, J. (2019). ERA5‐Land hourly data from 1950 to present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). 10.24381/cds.e2161bac [DOI]
- Muñoz‐Sabater, J. , Dutra, E. , Agustí‐Panareda, A. , Albergel, C. , Arduini, G. , Balsamo, G. , Boussetta, S. , Choulga, M. , Harrigan, S. , Hersbach, H. , Martens, B. , Miralles, D. G. , Piles, M. , Rodríguez‐Fernández, N. J. , Zsoter, E. , Buontempo, C. , & Thépaut, J.‐N. (2021). ERA5‐land: A state‐of‐the‐art global reanalysis dataset for land applications. Earth System Science Data, 13(9), 4349–4383. 10.5194/essd-13-4349-2021 [DOI] [Google Scholar]
- Nandris, D. , Nicole, M. , & Geiger, J. P. (1987). Variation in virulence among Rigidoporus lignosus and Phellinus noxius isolates from West Africa. European Journal of Forest Pathology, 17(4/5), 271–281. 10.1111/j.1439-0329.1987.tb01026.x [DOI] [Google Scholar]
- Nandris, D. , Nicole, M. , & Geiger, J. P. (1988). Root‐rot diseases of the rubber tree in the Ivory Coast. 1. Severity, dynamics, and characterization of epidemics. Canadian Journal of Forest Research, 18(10), 1248–1254. [Google Scholar]
- Neil, P. E. (1986). A preliminary note on Phellinus noxius root rot of Cordia alliodora plantings in Vanuatu. European Journal of Plant Pathology, 16, 274–280. [Google Scholar]
- Neil, P. E. (1988). Root disease (Phellinus noxius (Corner) G. H. Cunn.) of Cordia alliodora in Vanuatu. Commonwealth Forestry Review, 67, 363–372. [Google Scholar]
- Nicole, M. , Chamberland, H. , Rioux, D. , Xixuan, X. , Blanchette, R. A. , Geiger, J. P. , & Ouellette, G. B. (1995). Wood degradation by Phellinus noxius: Ultrastructure and cytochemistry. Canadian Journal of Microbiology, 41, 253–265. [Google Scholar]
- Nicole, M. , Nandris, D. , Geiger, J. P. , & Rio, B. (1985). Variability among African populations of Rigidoporus lignosus and Phellinus noxius . European Journal of Forest Pathology, 15, 293–300. [Google Scholar]
- Pinruan, U. , Rungjindamai, N. , Choeyklin, R. , Lumyong, S. , Hyde, K. D. , & Jones, E. B. G. (2010). Occurrence and diversity of basidiomycetous endophytes from the oil palm, Elaeis guineensis in Thailand. Fungal Diversity, 41(1), 71–88. [Google Scholar]
- Ram, C. S. V. (1975). Brown root disease of tea. Planters' Chronicle, 70, 217–218. [Google Scholar]
- Riggenbach, A. (1958). Fomes noxius, a thiamine‐deficient fungus. Nature, 182, 1390–1391. [DOI] [PubMed] [Google Scholar]
- Sahashi, N. , Akiba, M. , Ishihara, M. , Abe, Y. , & Morita, S. (2007). First report of the brown root rot disease caused by Phellinus noxius, its distribution and newly recorded host plants in the Amami Islands, southern Japan. Forest Pathology, 37(3), 167–173. [Google Scholar]
- Sahashi, N. , Akiba, M. , Ishihara, M. , Miyazaki, K. , & Kanzaki, N. (2010). Cross inoculation tests with Phellinus noxius isolates from nine different host plants in the Ryukyu Islands, southwestern Japan. Plant Disease, 94, 358–360. [DOI] [PubMed] [Google Scholar]
- Sahashi, N. , Akiba, M. , Ishihara, M. , Ota, Y. , & Kanzaki, N. (2012). Brown root rot of trees caused by Phellinus noxius in the Ryukyu Islands, subtropical areas of Japan. Forest Pathology, 42, 353–361. [Google Scholar]
- Sahashi, N. , Akiba, M. , Ota, Y. , Masuya, H. , Hattori, T. , Mukai, A. , Shimada, R. , Ono, T. , & Sato, T. (2015). Brown root rot caused by Phellinus noxius in the Ogasawara (Bonin) islands, southern Japan‐ current status of the disease and its host plants. Australasian Plant Disease Notes, 10, 33. 10.1007/s13314-015-0183-0 [DOI] [Google Scholar]
- Sahashi, N. , Akiba, M. , Takemoto, S. , Yokoi, T. , Ota, Y. , & Kanzaki, N. (2014). Phellinus noxius causes brown root rot on four important conifer species in Japan. European Journal of Plant Pathology, 140, 869–873. [Google Scholar]
- Samseemoung, G. , Jayasuriya, H. P. W. , & Soni, P. (2011). Oil palm pest infestation monitoring and evaluation by helicopter‐mounted, low altitude remote sensing platform. Journal of Applied Remote Sensing, 5(1), 3540. [Google Scholar]
- Sayers, E. W. , Cavanaugh, M. , Clark, K. , Ostell, J. , Pruitt, K. D. , & Karsch‐Mizrachi, I. (2020). Genbank. Nucleic Acids Research, 48(Database issue), D84–D86. 10.1093/nar/gkz956 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silva, M. K. R. , Fernando, T. H. P. S. , Wijesundara, R. L. C. , Nanayakkara, C. M. , & Tennakoon, B. I. (2017). Development of Brown root disease in Sri Lankan rubber plantations: Possible involvement of other tree species. Journal of the Rubber Research Institute of Sri Lanka, 97, 47–57. [Google Scholar]
- Singh, B. (1966). Timber decay due to five species of Fomes as new records in India. Indian Forester, 92, 653–655. [Google Scholar]
- Singh, S. , Bola, I. , & Kumar, J. (1980). Diseases of plantation trees in Fiji Islands I. Brown root rot of mahogany (Swietenia macrophylla king). Indian Forester, 106, 526–532. 10.36808/if/1980/v106i8/11215 [DOI] [Google Scholar]
- Singh, S. , & Pandey, P. C. (1989). Brown root‐rot of poplars. Indian Forester, 115(9), 661–669. [Google Scholar]
- Stewart, J. E. , Kim, M.‐S. , Ota, Y. , Sahashi, N. , Hanna, J. W. , Akiba, M. , Ata, J. P. , Atibalentja, N. , Brooks, F. , Chung, C.‐L. , Dann, E. K. , Mohd Farid, A. , Hattori, T. , Lee, S. S. , Otto, K. , Pegg, G. S. , Schlub, R. L. , Shuey, L. S. , Tang, A. M. C. , … Klopfenstein, N. B. (2020). Phylogenetic and population genetic analyses reveal three distinct lineages of the invasive brown root‐rot pathogen, Phellinus noxius, and bioclimatic modeling predicts differences in associated climate niches. European Journal of Plant Pathology, 156, 751–766. 10.1007/s10658-019-01926-5 [DOI] [Google Scholar]
- Steyaert, R. L. (1948). A contribution to the study of the plant parasites of the Belgian Congo. Bulletin de la Societe Royale de Botanique de Belgique, 80, 11–58. [Google Scholar]
- Sunthudlakhar, P. , Sithisarn, P. , Rojsanga, P. , & Jarikasem, S. (2022). HPLC quantitative analysis of protocatechuic acid contents in 11 Phellinus mushroom species collected in Thailand. Brazilian Journal of Pharmaceutical Sciences, 58, e20656. 10.1590/s2175-97902022e20656 [DOI] [Google Scholar]
- Supriadi, A. E. M. , Wahyuno, D. , Rahayuningsih, S. , Karyani, N. , & Dahsyat, M. (2004). Brown root rot disease of cashew in West Nusa Tenggara: Distribution and its causal organism. Indonesian Journal of Agricultural Science, 5(1), 32–36. [Google Scholar]
- Toy, S. J. , & Newfield, M. J. (2010). The accidental introduction of invasive animals as hitchhikers through inanimate pathways: A New Zealand perspective. Revue Scientifique et Technique (International Office of Epizootics)., 29(1), 123–133. [DOI] [PubMed] [Google Scholar]
- Tsai, J. N. , Ann, P. J. , Liou, R. F. , Hsieh, W. H. , & Ko, W. H. (2017). Phellinus noxius: Molecular diversity among isolates from Taiwan and ITS phylogenetic relationship with other species of Phellinus based on sequences of the ITS region. Botanical Studies, 58(1), 9. 10.1186/s40529-017-0162-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai, J. N. , Hsieh, W. H. , Ann, P. J. , & Yang, C. M. (2007). Development of specific primers for Phellinus noxius . Plant Pathology Bulletin, 16, 193–202. (Only the abstract in English). [Google Scholar]
- Tsang, K. S. W. , Cheung, M. K. , Lam, R. Y. C. , & Kwan, H. S. (2020). A preliminary examination of the bacterial, archaeal, and fungal rhizosphere microbiome in healthy and Phellinus noxius‐infected trees. Microbiology, 9(10), e1115. 10.1002/mbo3.1115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- USDA . (2023). USDA Plant Hardiness Zone Map., U.S. Department of Agriculture, Agricultural Research Service. https://planthardiness.ars.usda.gov/
- Wang, Y.‐F. , Meng, H. , Gu, V. W. , & Gu, J.‐D. (2016). Molecular diagnosis of the brown root rot disease agent Phellinus noxius on trees and in soil by rDNA ITS analysis. Applied Environmental Biotechnology, 1(1), 81–91. [Google Scholar]
- Wu, J. , Peng, S. L. , Zhao, H. B. , Tang, M. H. , Li, F. R. , & Chen, B. M. (2011). Selection of species resistant to the wood rot fungus Phellinus noxius . European Journal of Plant Pathology, 130, 463–467. [Google Scholar]
- Wu, M. L. , Chang, T. T. , Jaung, L. M. , Hung, T. H. , Chen, C. H. , & Lin, L. D. (2009). Establishment of PCR rapid detection technique for tree brown root rot disease. Quarterly Journal of Chinese Forestry, 42(2), 239–247. (Only the abstract in English). [Google Scholar]
- Wu, M. L. , Chen, C. H. , Chang, T. T. , Jaung, L. M. , & Hung, T. H. (2011). Establishment of the SOP of PCR detection techniques for brown root rot disease. Quarterly Journal of Chinese Forestry, 44, 7–17. (Only the abstract in English). [Google Scholar]
- Wu, Z.‐C. , Chang, Y.‐Y. , Lai, Q.‐J. , Lin, H.‐A. , Tzean, S.‐S. , Liou, R.‐F. , Tsai, I. J. , & Chung, C.‐L. (2020). Soil is not a reservoir for Phellinus noxius . Phytopathology, 110(2), 362–369. [DOI] [PubMed] [Google Scholar]
- Zhang, H. , Tze, K. N. , Kai, C. L. , Zoen, W. L. , Wai, F. Y. , & Wai, S. W. (2022). Development and evaluation of loop‐mediated isothermal amplification (LAMP) as a preliminary diagnostic tool for Brown root rot disease caused by Phellinus noxius (Corner) G.H. H. Cunningham in Hong Kong Urban Tree Management. Sustainability, 14, 9708. 10.3390/su14159708 [DOI] [Google Scholar]
- Zhou, L.‐W. , Ji, X.‐H. , Vlasák, J. , & Dai, Y.‐C. (2018). Taxonomy and phylogeny of Pyrrhoderma: A redefinition, the segregation of Fulvoderma, gen. nov., and identifying four new species. Mycologia, 110(5), 872–889. 10.1080/00275514.2018.1474326 [DOI] [PubMed] [Google Scholar]