

Summary
Wildfires are a global crisis, but current fire models fail to capture vegetation response to changing climate. With drought and elevated temperature increasing the importance of vegetation dynamics to fire behavior, and the advent of next generation models capable of capturing increasingly complex physical processes, we provide a renewed focus on representation of woody vegetation in fire models. Currently, the most advanced representations of fire behavior and biophysical fire effects are found in distinct classes of fine‐scale models and do not capture variation in live fuel (i.e. living plant) properties. We demonstrate that plant water and carbon dynamics, which influence combustion and heat transfer into the plant and often dictate plant survival, provide the mechanistic linkage between fire behavior and effects. Our conceptual framework linking remotely sensed estimates of plant water and carbon to fine‐scale models of fire behavior and effects could be a critical first step toward improving the fidelity of the coarse scale models that are now relied upon for global fire forecasting. This process‐based approach will be essential to capturing the influence of physiological responses to drought and warming on live fuel conditions, strengthening the science needed to guide fire managers in an uncertain future.
Keywords: carbon dynamics, fire behavior, fire effects, fire modeling, plant physiology, remote sensing, vegetation–fire interactions, water dynamics
| Contents | ||
|---|---|---|
| Summary | 953 | |
| I. | Introduction | 953 |
| II. | Existing models of fire behavior and effects | 955 |
| III. | Physiological controls of fire behavior and effects | 956 |
| IV. | Integrating fine‐scale physiology with fire models | 961 |
| V. | Remote sensing of plant water and carbon to inform process modeling | 963 |
| VI. | Conclusions and future directions | 964 |
| Acknowledgements | 965 | |
| References | 966 |
I. Introduction
Wildland fire burns hundreds of millions of hectares of forests, woodlands, and grasslands annually (Giglio et al., 2013), shaping terrestrial ecosystems (Bond & van Wilgen, 1996; Bond & Keeley, 2005; Bond, 2021), and their impacts on global carbon (Bowman et al., 2009) and water (Li & Lawrence, 2017) cycles. To accurately predict fire behavior and resulting effects, we need to understand the influences of vegetation structure and physiology on combustion (e.g. Byram, 1959; Weise & Wright, 2013) and the microenvironment (e.g. Finnigan, 2000; Banerjee et al., 2020; Atchley et al., 2021). Recent studies demonstrate that burned area shows greater increases in forests and shrublands where vegetation moisture is more sensitive to water limitation (Rao et al., 2022), and that hydraulic traits of woody evergreen species are responsible for up to 3.6‐fold variation in live fuel moisture content (LFMC, ratio of biomass water content to oven‐dry biomass, Table 1; Scarff et al., 2021). These findings emphasize the importance of accounting for ecophysiological controls on woody vegetation in wildfire forecasting and prescribed fire planning, where fuel effects dominate under less extreme fire danger conditions (Cruz et al., 2022). This will be particularly important in fire‐susceptible ecosystems which experience significant drought and LFMC declines below thresholds that drive increases in fire behavior (Pimont et al., 2019a). As atmospheric aridity continues to increase with rising temperatures, compounding plant water stress (Grossiord et al., 2020), the number of regions for which live fuels (i.e. living plants) are important in determining the behavior of fires and their ecological outcomes is likely to expand (Resco de Dios et al., 2021).
Table 1.
Glossary of pyro‐ecophysiology terms.
| Canopy bulk density (CBD, mass per unit volume) | Measure of how closely canopy fuels are packed, reflects likelihood that fire can move through the forest canopy |
| Carbon starvation | Plant mortality resulting from inability of NSC to meet metabolic demands |
| Cavitation | Process by which excessive water tension causes expansion of dissolved air to form bubbles; in plants, this causes a break in the water column and a decrease in hydraulic conductivity |
| Conduction | Heat transfer through a material from a region of higher temperature to a region of lower temperature |
| Consumption (mass per unit area) | Amount of biomass consumed during fire |
| Convection | Aeat transfer by the movement of a gas or liquid |
| Embolism | Blockage of a vessel by a mass; in plants, caused by air bubbles formed in xylem via cavitation |
| Equivalent water thickness (EWT, g m−2) | Measure of leaf water content |
| Fire radiative energy density (FRED, MJ m−2) | Measure of the intensity of radiative energy released from fuel during a fire |
| Hydraulic conductivity (water mass or volume per unit time per unit area) | Measure of a system's ability to transport water |
| Hydraulic failure | Plant mortality resulting from failure of the water column by exceeding PLC thresholds |
| Leaf mass per area (LMA, g m−2) | Measure of leaf thickness and density; inverse of specific leaf area; also called dry matter content (DMC) in remote sensing applications |
| Live fuel moisture content (LFMC, %) | Ratio of water mass to dry mass in living plants; controlled largely by physiological mechanisms, rather than weather |
| Non‐structural carbohydrates (NSC) | Plant carbon used for functions other than building structural biomass, such as growth, metabolism, osmoregulation, transport, storge, and defense |
| Osmotic potential (MPa) | Potential of water molecules to move from a less concentrated to a more concentrated solution across a semi‐permeable membrane |
| Percent loss of conductivity (PLC) | A measure of xylem vulnerability to cavitation at a given water potential |
| Phenology | Biological cycles resulting from seasonal or interannual climate variations |
| Physiology | Bynamic chemical and physical processes that govern function |
| Pyro‐ecophysiology | Ecophysiology‐based approach to live fuel research that considers how plant water and carbon cycles independently and collectively interact at the leaf and whole plant level to regulate flammability and subsequent fire behavior (Jolly & Johnson, 2018) |
| Radiation | Heat transfer through a gas or vacuum other than by heating of the intervening space |
| Relative water content (RWC, %) | Plant water content relative to its fully hydrated state |
| Senescence | Process of biological aging; can be stress‐induced or developmental |
| Traits | Morphological, physiological or phenological features measurable at the individual level (Violle et al., 2007) |
| Transpiration | Loss of water vapor from a living body; in plants, evaporation via stomata |
| Vapor pressure deficit (VPD, kPa) | The difference between the amount of moisture in the air and how much moisture the air can hold when saturated; measure of atmospheric aridity |
| Water potential (MPa) | Pressure potential required to remove a water molecule from its matrix (e.g. xylem); a measure of plant water status |
Historically, representation of live fuels in fire behavior models has been limited to static fuel models (Table 2) that generalize vegetation into classes, such as grass, shrub, and timber (Albini, 1976; Scott & Burgan, 2005; but see Rothermel & Philpot, 1973; Hough & Albini, 1978 for seasonally‐ and age‐dynamic fuel models). With recent advances in process‐based modeling and remote sensing of both fuels and fires, there is now opportunity to capture more realistic fuel heterogeneities, including the physiological dynamics that determine live fuel conditions (‘pyro‐ecophysiology’, Jolly & Johnson, 2018). This will allow exploration of their influence on fire behavior, ecological outcomes (i.e. plant injury, mortality, and recovery; hereafter, ‘fire effects’) and cascading hazards. Recent work has called for modeling efforts to improve integration of fire behavior and its effects (e.g. Hood et al., 2018; O'Brien et al., 2018; Kleynhans et al., 2021) to better capture fire–vegetation–environment feedbacks. This integration is particularly important in the context of low‐intensity and prescribed fire. While fuel heterogeneity has little effect in a high‐intensity fire environment (Atchley et al., 2021; Cruz et al., 2022), changes in phenology can make the difference between containment and escape for prescribed fires, where combustion dynamics are engineered to achieve desired biological and physical outcomes (e.g. species‐specific vegetation survival or mortality, fire risk reduction, soil protection, and smoke management (Hiers et al., 2020)) with implications for future fire and landscape dynamics (Mitchell et al., 2009; Gallagher et al., 2021). Plant water and carbon cycles have been recognized as important to both fire behavior (Nelson, 2001; Macias Fauria et al., 2011; Jolly et al., 2014; Jolly & Johnson, 2018) and ecological fire effects (Michaletz & Johnson, 2007; Hood et al., 2018; Bär et al., 2019). Yet, they have not been considered as an integrating framework mechanistically linking the two.
Table 2.
Glossary of models and remote sensing methods.
| Models | |
| 3D Fuel frameworks | Models that generate synthetic 3D fuels as inputs for CFD fire models |
| Computational fluid dynamics (CFD) | Physics‐based models that simulate the interaction of liquids and gases based on fluid mechanics principles; used in simulation of fine‐scale fire behavior and fire–atmosphere interactions within individual stands on timescales of minutes to hours |
| Ecosystem process | Mechanistic models based on theoretical understanding of ecological processes; for fire applications, can represent both fire behavior and effects on regional to global scales |
| Empirical fire effects | Statistical models that predict tree status (live, dead) as a function of tree characteristics and observed fire injury, often estimated based on flame length |
| Fire danger rating systems | Broad‐scale assessments of fire ignition, spread, and hazard potential based on current and antecedent weather, fuels, and topography |
| Functional structural plant models (FSPMs) | Mechanistic models of 3D plant structure, environment, and physiological response, from gene to community scales |
| Fuel models | Stylized set of fuel bed characteristics used in fire models |
| Landscape fire succession | Spatial simulation models of fire and vegetation dynamics at stand to landscape scales |
| Operational fire | Computationally inexpensive models that rely on empirical representations of fire behavior and fixed, stylized fuel models to determine flame length and resulting mortality |
| Process‐based fire effects | Models which predict injury to different plant compartments based on heat transfer |
| Vegetation demography | Models which capture dynamic, size‐structured vegetation |
| Remote sensing | |
| Hyperspectral | Acquired in narrow, contiguous wavelength bands; high spectral resolution allows for material characterization, inference of chemical/biological processes, and novel signature identification |
| Lidar | Light detection and ranging; active optical sensing method using return time of a pulsed laser to measure distance |
| Microwave | Detects backscattering of actively transmitted radiation in the microwave (1 cm–1 m) portion of the electromagnetic spectrum; longer wavelengths allow penetration of clouds, rain, and surfaces |
| Multispectral | Acquired in broad, often discontinuous, wavelength bands; low spectral resolution reduces cost and complexity for monitoring known signatures |
| Optical | Detects reflected radiation in the visible (VIS, 380–780 nm), near‐ (NIR, 780–1000 nm) and short‐wave infrared (SWIR, 1000–2500 nm) portions of the electromagnetic spectrum; primarily passive |
| Thermal | Detects emitted radiation in the thermal infrared (TIR, 780 nm–1 mm) portions of the electromagnetic spectrum; primarily passive |
We argue that plant water and carbon cycles drive live fuel moisture and dry mass dynamics, which influence heat transfer into the plant during a fire and subsequent postfire survival and recovery. This focus on plant water and carbon dynamics provides a mechanistic framework that links plant physiology to fine‐scale fire behavior and fire‐induced effects on plant tissues, addressing a gap in current approaches to modeling fire and vegetation (Fig. 1). We focus in this review on the interactions of woody plants and fire, due to the availability of literature and to maintain focus on the underlying argument. However, many of the needs and underlying connections that we recognize have importance to nonwoody systems. Understanding how within‐plant physiology and fire interact will allow exploration of mechanisms important to ensuing plant, stand, and landscape‐level vegetation dynamics, which influence subsequent fire behavior. To fully capture fire‐ecosystem feedbacks, we propose exploring new ways to bring together fire behavior and effects models at fine scales where both fire and physiological processes can be represented in sufficient detail to define important biophysical mechanisms, with heat transfer to vegetation providing the mechanistic linkage from physiology to fire behavior and subsequent effects (Varner et al., 2021). A fundamental understanding of how fire and plants interact at the fine scale is needed to constrain simulations used at management‐relevant scales. Under unprecedented future climate conditions, for which we do not have past or present analogs, biophysical process modeling will be essential to understand response to conditions that fall outside the range of variability captured by current empirical models. Linking remotely sensed estimates of plant water and carbon status to models of fire behavior and effects offers a mechanistic approach critical to capturing the influence of physiological responses to drought and warming on live fuel conditions under global change.
Fig. 1.
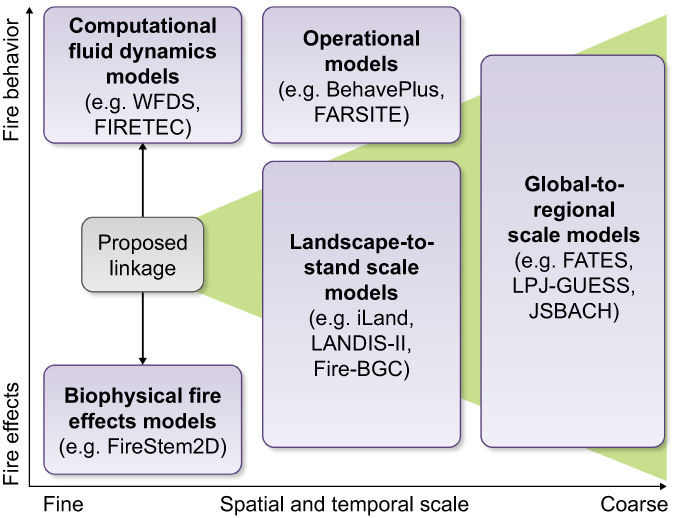
Model classification according to scale and process representation. In this conceptual figure, we show the classification applied to different models of fire behavior and effects discussed in this review. We also highlight our proposed linkage between fine‐scale computational fluid dynamics (CFD) and biophysical fire effects models.
II. Existing models of fire behavior and effects
There are many existing fire behavior and effects models under use and continuous development. Rather than focusing on any one model or group of models, here we focus on commonly applied scales and model characteristics. We classify these as global to regional‐scale models, landscape to stand‐scale models, operational models, computational fluid dynamics models, and fine‐scale biophysical fire effects models.
On global to regional scales (Fig. 1), fire behavior and effects have been combined within ecosystem process models using various approaches to simulate ecosystem dynamics with fire disturbance (see Rabin et al., 2017 for in‐depth review). Simulations commonly span decades to centuries and may include the entire globe. In these models, fire behavior and effects are commonly simulated explicitly at the computational grid level (c. 0.1–1°) or implicitly at a subgrid level. Fire spread is a function of fuel characteristics (loading, size distribution, and moisture), ignition patterns, weather, and latent suppression. These models often capture a dimension of vegetation growth and/or demographics which can dynamically inform fuel loads and fuel availability. Fire behavior is usually represented empirically based on experimental and modeling studies by Rothermel (1972) and Van Wagner (1973). Fire effects are determined by total energy release and thresholds for mortality by either individuals, cohorts, or by fractioning existing vegetation. For example, recent advances in vegetation demography modeling include the addition of plant hydraulics in FATES (Christoffersen et al., 2016), allowing decoupling of water and carbon dynamics for more direct determination of live fuel vulnerability to fire at multiple timescales. Live fuel moisture content dynamics are captured by simulating water‐ and carbon‐cycle processes directly at subdaily time scales, allowing for assessment of climate impacts (e.g. change in CO2, temperature, and precipitation) on future LFMC trends under different climate scenarios (Ma et al., 2021). However, simplified representation of fire precludes mechanistic understanding of fire–physiology interactions.
At stand to landscape scales, a large number of models were developed for spatial plant ecology and fire occurrence at the scale relevant to forest management (10–100 km2; e.g. FIRE‐BGC (Keane et al., 2004), LANDIS‐II (Sturtevant et al., 2009; Scheller et al., 2019)). These models are applied to decadal to century‐long simulations. Observed or modeled fire dynamics account for stand‐to‐stand fire spread probabilistically or through the estimation of mechanistic processes. These models simulate cohorts or representative sets of individual plants/trees to estimate vegetation development and demography which inform fuel loading and availability. While this class of models can be leveraged to explore fire behavior and effects, they do not mechanistically link the two (Keane et al., 2004). Within the stand (1–10 km2), fire effects are usually homogenous or stochastically based on homogenous traits of the stand (Furniss et al., 2022). For example, FIRE‐BGCv2 (Keane et al., 2011) links biogeochemical processes with stand‐scale fire behavior and effects based on live and dead fuel loading, leveraging FARSITE (Finney, 1998), but does not address within‐stand variability. Keane et al. (2004) and Sturtevant & Fortin (2021) provide extensive reviews of landscape to stand‐scale models.
Operational fire models such as BehavePlus (Andrews, 2013), FARSITE, FLAMMAP (Finney, 1998), or FFE‐FVS (Reinhardt & Crookston, 2003) also represent fire behavior and effects. However, these models, which need to be computationally inexpensive to be useful, rely on empirical representations of fire behavior (Rothermel, 1972), and use fixed, stylized fuel models (Scott & Burgan, 2005) to determine flame length and resulting mortality. Their linkage between fire behavior and effects is thus very simple, and largely does not account for dynamic plant physiology or demography.
At the substand scale (< 1 m–1 km), the most advanced representations of fire behavior and effects are currently found in distinct classes of models. Fire behavior is best captured with Computational Fluid Dynamics (CFD) models such as FIRETEC (Linn et al., 2002) or the Wildland–Urban Interface Fire Dynamics Simulator (WFDS, Mell et al., 2009). These models simulate individual fire events, or portions thereof, on short timescales of minutes to hours. Fuel parameters are static, with exception of moisture mass and dry mass of the fuel, which may decrease through dehydration and consumption, respectively. These models do not represent fire effects, as they do not explicitly distinguish between live and dead fuels, or between different vegetation species or functional groups. In absence of explicit vegetation response to changing meteorological conditions, they cannot resolve changes in LFMC or the sensitivity of fire behavior to live fuel moisture dynamics (Jolly, 2007).
Prediction of fine‐scale ecological fire effects (postfire vegetation injury, mortality, and recovery) has been approached using both empirical and process‐based models, with fire intensity provided as input from other models or measurements. Most postfire mortality models rely on empirical correlations (Woolley et al., 2012; Hood et al., 2018) which predict vegetation status (live or dead) as a function of plant characteristics (e.g. species, bark thickness, height, and diameter) and observed fire injury (e.g. crown scorch, and bark char), often estimated based on flame scorch height (Van Wagner, 1973). A number of process‐based biophysical fire effects models predict injury to different woody plant compartments (e.g. stem (FireStem2D, Chatziefstratiou et al., 2013) or crown (Michaletz & Johnson, 2006) heating models). Few, however, have considered injuries to more than one part of the plant (Michaletz & Johnson, 2008), and none address the influence of interacting injuries across multiple woody plant compartments on postfire plant function.
Direct coupling of fine‐scale fire behavior and effects models remains challenging due to the different temporal scales that are relevant for both. As an alternative to solve this challenge, we propose exploring nested modeling frameworks (Gettelman et al., 2022; Shuman et al., 2022) which can transfer information across models at scales where both fire and physiological processes can be represented in sufficient detail to define important biophysical mechanisms. Recent work has made strides in mechanistically linking fire behavior to ecological effects through plant traits (Zylstra, 2021), but without mechanistic determination of vegetation density and moisture content resulting from plant water and carbon dynamics. By incorporating fine‐scale physiology, we can capture dynamic water and carbon to represent these properties more realistically.
III. Physiological controls of fire behavior and effects
1. Evidence for the role of plant water and carbon in fire behavior
Fire behavior is broadly controlled by complex interactions among fuel density and moisture content, topography, and the atmosphere (Countryman, 1966). These factors not only interact, but also can be highly variable in space and time, resulting in dynamic fire environments. Live fuel moisture content is a key fuel property governing fire behavior (Fig. 2a). Declining LFMC is associated with an increase in area burned (Dennison & Moritz, 2009; Nolan et al., 2016; Pimont et al., 2019b), and LFMC below 100% sharply increases fire rate of spread (ROS, Pimont et al., 2019a) and the probability of large fires (Martin‐StPaul et al., 2018) (Fig. 2a). Given its impact on fire behavior, LFMC has been incorporated into fire danger rating systems (e.g. Deeming et al., 1977; Stocks et al., 1989) and fire behavior models (e.g. Rothermel, 1972).
Fig. 2.
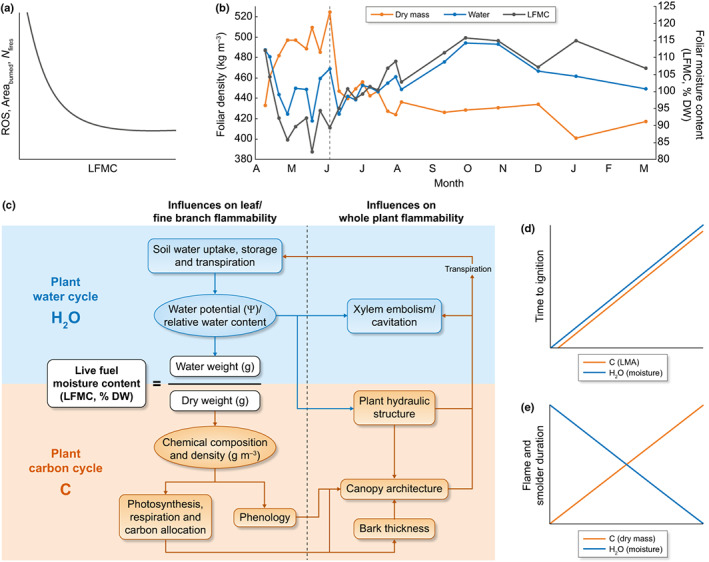
Live fuel moisture content (LFMC, black), an important driver of landscape‐scale fire behavior (a), is a function of plant carbon (C, orange) and water (H2O, blue) (b, c), which can have opposing influences on fire behavior in leaf level burn experiments (d, e). (a) Rate of spread (ROS), area burned, and fire occurrence (N fires) as a function of LFMC, as shown by Dennison & Moritz (2009), Martin‐StPaul et al. (2018), Pimont et al. (2019a,b), and others. (b) Monthly variation in foliar dry mass (orange) and water (blue) density, and foliar moisture content (dry mass/water mass, LFMC; black) extracted from Jolly et al. (2016) with WebPlotDigitizer (Rohatgi, 2021). Dashed vertical line indicates divergence between LFMC and water density driven by increasing dry mass. (c) Conceptual model of LFMC as a function of plant water and carbon cycles redrawn from Jolly & Johnson (2018). (d) Time to ignition as a function of leaf carbon (leaf mass per area, LMA) and water (moisture) as shown in Grootemaat et al. (2015) and Bianchi et al. (2019). (e) Flame and smolder duration as a function of leaf carbon (dry mass) and water (moisture), as shown in Grootemaat et al. (2015) and Bianchi et al. (2019).
The water and dry mass components of LFMC are outcomes of plant water‐ and carbon‐cycle processes, respectively, and are controlled by environmental conditions, soil properties, and plant physiology (Fig. 2b,c; Macias Fauria et al., 2011; Jolly & Johnson, 2018; Ruffault et al., 2018). Plant water and carbon can vary independently in both space and time, with opposing effects on LFMC. For example, a sharp prebudburst (early June) increase in foliar dry mass of older, live pine needles has been shown to reduce LFMC despite increasing foliar water mass (Fig. 2b; Jolly et al., 2014, 2016). Based on these findings, Jolly & Johnson (2018) proposed a framework decomposing LFMC into plant water and carbon cycle processes (Fig. 2c), where the numerator is determined by whole plant hydraulics (e.g. transpiration; relative water content; and xylem embolism), and the denominator by carbon allocation (e.g. chemical composition/density; phenology; and canopy architecture). These processes influence fire behavior both by changing tissue‐ and canopy‐level properties that influence heat transfer, and by altering the proportion of dead biomass in fuels and on the ground surface. Seasonal or drought‐induced senescence and shedding are examples of physiological processes that alter leaf, fine branch, and whole plant flammability, transforming live fuels into dead fuels and increasing litter accumulation. Tissue senescence is associated with shifts in osmolytes and water content (Milla et al., 2007), which can increase or decrease LFMC. Tissue mortality also increases the dead to live fuel ratio, causing a strong increase in fire spread and intensity (e.g. Cruz et al., 2015; Sieg et al., 2017; Balaguer‐Romano et al., 2020). When senesced tissues are shed, litter accumulates in the surface fuel‐bed and may increase the likelihood of surface fire.
The importance of both plant water‐ and carbon‐cycle processes for fire behavior becomes clear when examining their effects on heat transfer and combustion (Michaletz & Johnson, 2007; Bär et al., 2019; Dietenberger et al., 2020; Kleynhans et al., 2021). Preheating, the process by which heat transfer evaporates water, dries, and decomposes carbon‐containing polymers in unburned fuels, occurs via conduction, convection, and radiation. Thermal conductivity varies with material density, temperature, and water content, while convection depends on geometry and orientation of the fuel surface. The proportion of radiation absorbed by unburned fuel depends on both its thermal absorptance as well as geometry and orientation. Fuel chemistry and water content are therefore important to both conductive and radiative heat transfer through their influence on thermal conductivity and absorptance, while convection and radiative heat transfer to the fuel surface depend on vegetation geometry and orientation, including leaf, branch, crown, or canopy structure (Michaletz & Johnson, 2006; Pausas & Moreira, 2012). The rate at which the temperature increases in response to heat transfer depends on the mass and specific heat of the biomass, which depend on plant tissue water content and composition (Boardman et al., 2021). Water content also determines how much energy is required to heat the fuel to evaporation and combustion temperatures (Yashwanth et al., 2016), while the content of carbon‐based polymers determines the amount and rate of thermal decomposition, influencing the rate of preheating and time to combustion (Kilzer & Broido, 1965). Fuel chemistry and water content are also critical in determining whether combustion reactions are self‐sustaining and how they will contribute to fire intensity and rate of spread (Quintiere, 2006; Matt et al., 2020). During combustion, the content of organic polymers determines how much energy can be released, and water content can determine whether flaming or smoldering combustion occurs. Higher rates of fire spread and probability of flaming combustion, which occurs at high temperature and/or low moisture, are therefore more likely when live fuels are water stressed and the ratio of water mass to dry mass (LFMC) is lower.
Results from leaf‐level burn experiments support the influence of plant water and carbon processes on preheating. Time to ignition (Fig. 2d) has been shown to increase with moisture (Grootemaat et al., 2015; Bianchi et al., 2019; but see Fletcher et al., 2007) and leaf mass per area (LMA, inverse of specific leaf area, Grootemaat et al., 2015), which is associated with accumulation of total structural carbohydrates and lignin (Poorter et al., 2009). Flame and smolder duration (Fig. 2e) also increase with dry mass (Grootemaat et al., 2015), but decrease with moisture (Bianchi et al., 2019). These contrasting effects of leaf‐level water and carbon on different fire behavior characteristics underscore the importance of capturing LFMC components independently. Despite evidence from Alam et al. (2020) that certain leaf and shoot flammability metrics (e.g. ignitibility and combustibility) are decoupled, the same study showed a positive relationship of leaf dry matter content with shoot and species‐level flammability, suggesting that effects of leaf water and carbon on fire behavior are scalable. At canopy scales, Jolly et al. (2016) found that increased leaf density, which accounts for changes in dry mass associated with phenological change in foliar chemistry and carbon allocation, increased modeled crown fire propagation and area burned. Additionally, canopy bulk density, a measure of canopy architecture influenced by water‐ and carbon‐cycle processes via growth rate, environment, and phenology, has long been recognized as an important driver of spread rate in active crown fires (Van Wagner, 1977; Resco de Dios, 2020). The independent relationships of water‐ and carbon‐cycle processes with fire behavior, and their complex interdependence, highlight the need for a more mechanistic representation of LFMC that accounts for underlying physiology and allows for dynamic change.
2. Evidence for the role of plant water and carbon in fire effects
First‐order effects of fire on vegetation are the direct result of combustion and heat transfer to plant tissues (Michaletz & Johnson, 2007), and can be modulated by plant physiological characteristics. The same traits that influence fire behavior through controls on heat transfer also influence fire effects by modulating tissue temperatures and exposure times, particularly in low and mixed‐severity fire regimes where vegetation is not combusted and immediately killed (O'Brien et al., 2018; Varner et al., 2021). Percent mortality has been demonstrated to increase with fire radiative energy density (Smith et al., 2017; Steady et al., 2019), or time‐integrated radiative flux density from ignition to cessation of the fire. Since absorbed fire radiative energy is tied to flame and smolder duration (O'Brien et al., 2016), which increases with dry mass and decreases with moisture (Fig. 2e), we expect first‐order mortality to increase with dry mass and decrease with moisture content (Fig. 3a). This is consistent with significant increases in simulated canopy fuel consumption with reduced canopy fuel moisture, particularly under low‐wind scenarios (Sieg et al., 2017). While the physical relationship between moisture and flame and smolder duration should hold across species and ecosystems, further research is required to validate the relationship between moisture content and first‐order mortality, as tissues with higher moisture contents can also be more heat sensitive (Wright & Bailey, 1982).
Fig. 3.
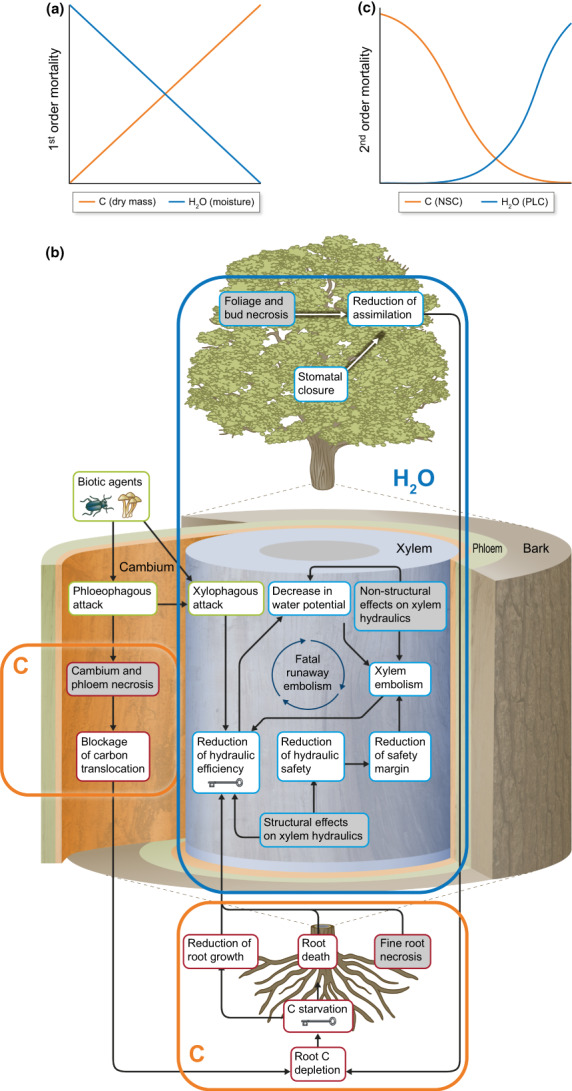
Plant carbon (C, orange) and water (H2O, blue) determine fire effects. (a) First‐order effects are the direct result of heat transferred to plant tissues (Michaletz & Johnson, 2007), as modulated by plant tissue properties (water and carbon content). Percent mortality increases with time integrated from ignition to cessation of fire (fire radiative energy density (FRED); Smith et al., 2017; Steady et al., 2019), which should approximate flame and smolder duration (Fig. 1e), which increase with leaf carbon (dry mass) and decrease with leaf water (moisture) content (Grootemaat et al., 2015; Bianchi et al., 2019). (b) Conceptual diagram illustrating the cascade of potential physiological responses to post‐fire injuries in plant roots, stems, and crowns adapted from Bär et al. (2019). Blue frame captures processes impacting the plant water cycle leading to reduction in hydraulic efficiency and ultimate hydraulic failure (fatal runaway embolism). Orange frames capture processes impacting the plant carbon cycle, leading to carbon starvation. (c) Second‐order effects are modulated by post‐fire water and carbon availability, along with integrity of the water and carbon uptake and transport systems (Hood et al., 2018; Bär et al., 2019). Mortality thresholds for hydraulic failure and carbon starvation are defined based on percent loss of conductivity (PLC, Hammond et al., 2019) and non‐structural carbohydrate (NSC) concentrations (Barker Plotkin et al., 2021), both of which are impacted by fire (Varner et al., 2009; Michaletz et al., 2012; West et al., 2016).
When vegetation is not consumed, fire‐induced injuries, including hydraulic dysfunction, cambium and apical meristem necrosis, leaf and fine root necrosis, are proposed to influence whole‐plant carbon and water budgets resulting in hydraulic failure and/or carbon starvation (Fig. 3b; Midgley et al., 2011; Michaletz et al., 2012; Hood et al., 2018; Michaletz, 2018; Silva et al., 2018; Bär et al., 2019; Berenguer et al., 2021). This framework suggests that second‐order effects on live vegetation are driven, in part, by postfire water and carbon availability, along with integrity of the water and carbon uptake and transport systems (Fig. 3b,c; Michaletz & Johnson, 2007; Hood et al., 2018; Michaletz, 2018; Bär et al., 2019). Postfire environmental conditions and species‐specific traits determine whether resulting functional and growth limitations will lead to vegetation recovery or mortality (Bär et al., 2019; Sayer et al., 2020; Hood, 2021; Ruswick et al., 2021). For instance, traits such as water stress resistance are observed to differ between disturbance‐dependent and obligate sprouters in Mediterranean‐type climate regions (Pratt et al., 2012), with implications for postfire community composition. Franco et al. (2014) note that in neotropical savannas, where fire is common and accelerated fire frequencies and prolonged droughts are expected under climate change, the interplay between temperature optima and CO2 fertilization effects on photosynthesis, photorespiration, and respiration will define how much carbon is available for postfire plant growth and resprouting, determining the acclimation potential of a given species, plant community or ecosystem. Indeed, from Mediterranean to Tropical ecosystems, the balance between forest expansion and retreat, between seedling recruitment and mortality, has been attributed to species differences in drought sensitivity (e.g. embolism resistance, rooting depth, and stomatal regulation) and carbon balance (e.g. stomatal regulation, osmotic adjustment, and carbon allocation) (Keeley, 1998; Franco et al., 2014).
Drought and herbivory research, where tree mortality has a long history of study, has defined mortality thresholds for hydraulic failure in terms of percent loss of hydraulic conductivity (PLC; Hammond et al., 2019), and for carbon starvation in terms of nonstructural carbohydrate (NSC) depletion (Barker Plotkin et al., 2021). Evidence of increased PLC from laboratory experiments (Michaletz et al., 2012; West et al., 2016; Partelli‐Feltrin et al., 2021), as well as from forest fires (Bär et al., 2018), indicates that fire increases vulnerability to cavitation (Midgley et al., 2011; Hood, 2021). In addition, fire can decrease root NSC (Varner et al., 2009), reducing stores available to support metabolism, growth, and subsequent stress response, resulting in mortality independent of canopy damage. Together, this research suggests similar relationships for second‐order fire mortality (Fig. 3c) as for mortality from drought and herbivory, where sufficiently high PLC or low NSC results in mortality. This is consistent with recent work linking mortality and recovery from low‐intensity fire to prefire water status (van Mantgem et al., 2018; Partelli‐Feltrin et al., 2020) and concentrations of NSC (Zhu et al., 2012; Sayer et al., 2020). In addition, smoldering consumption can impact water uptake directly through root loss (O'Brien et al., 2010). In conifers, trade‐offs have also been documented between resistance to cavitation and to fire, likely mediated by allocation of carbon to either building thick bark or dense xylem, but not both (Resco de Dios et al., 2018). These trade‐offs have implications for woody vegetation recovery from fire under climate change, where drought conditions can predispose woody vegetation to hydraulic (Partelli‐Feltrin et al., 2020) and carbon (Sayer et al., 2020) limitations, impacting capacity to recover from fire‐induced injury and/or resprout. Indeed, intense postfire drought can also cause significant resprout mortality resulting from simultaneous loss of hydraulic conductivity and depletion of root starch (Pratt et al., 2014).
3. Importance of plant water and carbon dynamics
Given the demonstrated role of plant water and carbon status in both fire behavior (Fig. 2) and effects (Fig. 3), accurately capturing their dynamics (seasonal variation, differences across species or functional types, with topography, and natural disturbance) will lead to improvements in fire behavior and effects models. Carbon and water status of live fuels change dynamically through time with environmental conditions and seasonal phenology (Baffoin et al., 2021). Variation in plant dry matter is driven by changes in organic molecules, largely NSC and lipids synthesized in the leaf and allocated to different plant organs (i.e. roots, stems, leaves, and reproductive structures) as needed for respiration and storage (Chapin et al., 1990). Diurnal variation results from daily synthesis and export patterns, while seasonal variation is largely driven by re‐allocation to meet seasonal demands associated with phenology or source limitation (e.g. drought‐induced reduction in photosynthesis; Martínez‐Vilalta et al., 2016). Changes in water content are driven by the interplay between soil water availability, physiological and morphological regulation, and atmospheric vapor pressure deficit through the soil–plant‐atmosphere continuum (Nelson, 2001; Macias Fauria et al., 2011). Water and carbon status differ across organs and over time as water moves from root to leaf via transpiration and as carbon is assimilated and transported across organs via the phloem (Fig. 4a, Zhou et al., 2020), and these dynamics differ by species and/or plant functional type. For instance, while evergreen conifers show strong seasonal variation in leaf NSC and dampened variation in stem NSC and water deficit (∆W), the opposite is true for evergreen angiosperms (Fig. 4b, Sánchez‐Costa et al., 2015; Martínez‐Vilalta et al., 2016). Topographic variation (microclimates, water availability, and soil resources available for growth) can also modify physiology, and fuel availability and loading, and has been shown to govern fire behavior and effects (Knapp et al., 1993; Krawchuk et al., 2016; Swann et al., 2022). Different functional strategies in relation to seasonal water deficit and disturbance, such as those defined by leaf lifespan (deciduous vs evergreen) and minimum water potential (Ackerly, 2004), will determine how these water and carbon dynamics manifest across species. Even within an individual species, plants can modulate the effects of variability in water resources by modifying leaf area, rooting depth, and/or stomatal conductance (Rambal, 1993). Among co‐occurring chaparral shrubs, differences in rooting depth can drive differential responses of leaf water potential to seasonal drought (Davis & Mooney, 1986; Smith & Richardson, 1990), and those with similar responses may use different modes of seasonal osmotic adjustment to accomplish changes in osmotic potential (i.e. shift in water volume vs solute concentration; Bowman & Roberts, 1985), with strong implications for LFMC. Species differences in drought response and associated leaf senescence and shedding have implications for litter accumulation and fuel‐bed flammability as well due to species variation in litter flammability and nonadditive effects in multi‐species litter mixtures (de Magalhães & Schwilk, 2012, 2021).
Fig. 4.
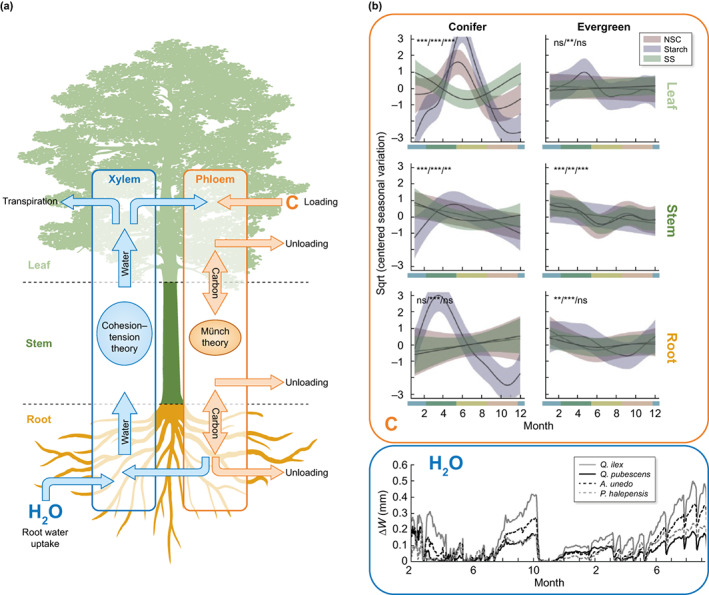
Plant carbon (C, orange) and water (H2O, blue) are constantly varying across organs (a), species and time (b). (a) Plant water (H2O, blue) and carbon (C, orange) exchange across organs (leaf, root, stem) by cohesion‐tension and Münch flow through xylem and phloem adapted from Zhou et al. (2020). (b, Carbon) Global patterns of seasonal variation (centered smooths, square‐root transformed mg g−1 dry mass) in total non‐structural carbohydrates (NSC), starch, and soluble sugars (SS) as a function of month, functional type (columns; conifer, evergreen angiosperm), and organ (rows; leaf, stem, root) adapted from Martínez‐Vilalta et al. (2016). (b, H2O) Seasonal course of tree water deficit (∆W, de‐trended stem diameter variation) for four species representing three different Mediterranean forest functional types: evergreen sclerophyllous (Quercus ilex L., Arbutus unedo L.); winter deciduous (Quercus pubescens Willd.); early successional, drought‐adapted conifer (Pinus halepensis Mill.) (adapted from Sánchez‐Costa et al., 2015).
Climate and phenology drive seasonal change in these fire‐relevant traits, regulating water and NSC balances. Under future climate conditions, drought and phenological shifts can affect greenup, senescence, and susceptibility to mortality, all of which impact fuel load and flammability (Jolly et al., 2016). For instance, prefire drought increases the likelihood of mortality (van Mantgem et al., 2018; Partelli‐Feltrin et al., 2020) and of resprouting failure (Karavani et al., 2018; Resco de Dios et al., 2020). Topographic variation can be an important control on drought induced mortality and resulting fuel loads. For example, increased water availability in concave areas can mediate the increase in hydraulic stress typical at lower elevations (Tai et al., 2017). Prefire drought and heat stress may also diminish flowering and seed production, negatively impacting species with fire‐cued recruitment (Nolan et al., 2021). Heat susceptibility of tissues (Bär et al., 2021) and probability of vegetation survival can also be influenced by plant phenological stage during a burn (Trabaud, 1991; Knapp et al., 2009; Ruckman et al., 2012; Pratt et al., 2014), highlighting the importance of capturing seasonal variation in prediction of mortality (Bond & van Wilgen, 1996). Beyond drought, other natural disturbances can cause physical damage to vegetation (e.g. pathogens and herbivory), change in fuel loads and availability due to rapid mortality (e.g. bark beetles and tropical cyclones), or increased resource availability for plant growth (e.g. windfall), all of which can be incorporated into plant physiology models to understand their feedbacks on fire behavior and effects (Karp et al., 2021; Rouet‐Leduc et al., 2021; Fettig et al., 2022; Ibanez et al., 2022; Lee et al., 2022). Acknowledging dynamic physiology recognizes that fire behavior and its effects will vary, within and across species, with plant water status (Nolan et al., 2018), carbon dynamics (photosynthesis, respiration, allocation), phenology (Bär et al., 2021), species (McAllister & Weise, 2017; Nolan et al., 2018; Resco de Dios, 2020), and time (Fig. 4).
IV. Integrating fine‐scale physiology with fire models
The wildland fire research community has called for the use of process‐based models to explore the potential mechanisms and interactions driving fire dynamics and effects and to conduct virtual experiments that allow for consideration of no‐analog future climate conditions (Michaletz et al., 2013; Hoffman et al., 2018; O'Brien et al., 2018). The demonstrated importance of plant water and carbon to fire behavior and effects, and their variability across species and time, underscore the value in applying plant carbon and water process models to explore the biophysical mechanisms linking vegetation to fire behavior and effects under present and future conditions.
Recent advances in whole‐plant modeling offer a promising framework for linking simulations of plant water‐ and carbon‐related processes to finer‐scale models of fire behavior and effects. Nolan et al. (2018, 2020) suggested a physiological basis to model LFMC for fire behavior applications using leaf water potential as a proxy. This approach allows for species‐specific variation with environmental drivers, while acknowledging the importance of capturing variation in leaf mass to account for decoupling of seasonal changes in the water and carbon contents that comprise LFMC. Mechanistic functional structural plant models (FSPMs) (Vos et al., 2010; Louarn & Song, 2020; de Vries, 2021), of 3D plant structure, environment, and physiological response, from gene to community scales, offer a potential solution to capture both water and carbon cycle processes. Functional structural plant models, which can simulate water and carbon flows across 3D plant compartments (root, stem, and leaf) in response to environmental conditions, have been demonstrated for use in simulation of both small plants and trees and include open‐source platforms that enable coupling with external modeling tools (e.g. Zhou et al., 2020).
Such a model could be coupled to fine‐scale fire behavior models using fuel modeling frameworks such as Fuel3D (Parsons et al., 2011), FuelManager (Pimont et al., 2016), STANDFIRE (Parsons et al., 2018), or FastFuels (Parsons et al., 2020). These frameworks can generate the 3D structure of canopy, mid‐, and under‐story vegetation for input to physics‐based fire models, such as FIRETEC or WFDS, allowing examination of within‐stand 3D fuel–fire interactions. Currently, meteorological or remotely sensed data (see the Remote sensing of plant water and carbon to inform process modeling section) can be used to set the moisture properties in these fuel models. Moisture is most often assigned a fixed value by the user for the course of the simulation across minutes to hours, and only FuelManager allows for assignment of alive vs dead status. As an advance to enable mechanistic evaluation of fine‐scale interactions between physiological and heat transfer processes and their influence on fire behavior and effects, we propose incorporation of a physiological process model to determine fuel condition based on physiological descriptions (plant water and carbon‐related processes) of individual plants (e.g. Cochard et al., 2021; Ruffault et al., 2022a,b). This could be achieved through use of a FSPM with remotely sensed parameterization (Fig. 5a,b) to inform 3D fuel properties (Fig. 5c). These properties would provide the necessary inputs to parameterize fire behavior models (Fig. 5d), the outputs of which could inform predictions of subsequent plant water and carbon dynamics (Fig. 5a). Model representations of these postfire physiological processes could then inform second‐order fire effects (Fig. 5e) through a process‐based mortality model (e.g. Michaletz & Johnson, 2008; Butler & Dickinson, 2010) and subsequent 3D fuel properties (Fig. 5c). This approach will require parallel advances in mechanistic modeling of whole‐plant fire injury processes to determine whether interacting injuries result in ultimate recovery, mortality, or functional limitations (Hood et al., 2018; Bär et al., 2019), which will be essential to determining subsequent fuel properties.
Fig. 5.
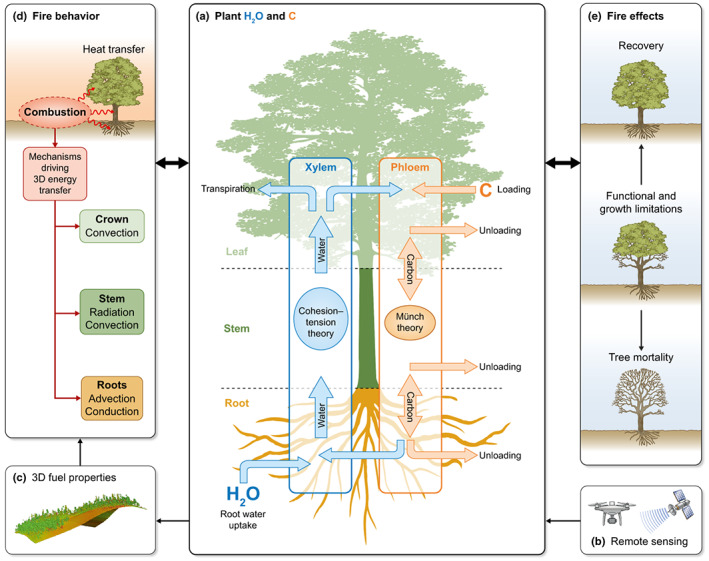
Plant carbon (C) and water (H2O) dynamics provide the mechanistic linkage between fire behavior and effects. In our conceptual framework, plant carbon and water process models (a; adapted from Zhou et al., 2020), which can be parameterized by remotely sensed estimates of canopy carbon and water contents (b; e.g. Dahlin et al., 2013; Singh et al., 2015), provide input to 3D fuel models (c; e.g. Pimont et al., 2016; Parsons et al., 2018). Detailed fuel properties are then exported to physics‐based fire models (d; adapted from O'Brien et al., 2018; Bär et al., 2019) that provide heat transfer estimates for input to the plant water and carbon model (a), which feeds into simulation of fire effects (e; adapted from Bär et al., 2019). These effects, along with remotely sensed estimates (b), drive subsequent plant water and carbon (a), feeding 3D fuels (c), and the cycle repeats.
The individual plant (or within‐plant) detail of this approach will enable further exploration of the plant traits most critical to the process of, and sensitivity to, heat transfer, allowing for determination of scale‐relevant parameters. A model sensitivity approach varying plant physiological input parameters could be used to define functional groupings, based on species traits (response of tissue‐level water and carbon to environmental drivers) that influence fire behavior and effects, which may allow for more generalized response functions to reduce computational resource requirements for use at larger scales.
By directly linking combustion, physiological process, and fire‐injury plant‐mortality models to hydrologic and plant succession models in a spatially explicit way, this framework could potentially be extended to simulate landscape‐scale ecohydrology, and its response to fire disturbance under present and future climate conditions. By calculating transient heterogenous fuel moistures and temperatures based on incident radiation, heat fluxes, and topographic variation, this model extension could capture species‐ (or functional type‐) dependent responses to soil moisture at different depths. Landscape‐scale models of ecohydrology could use available downscaled climate reconstructions and models and spatially explicit maps of soil characteristics, topography, and aspect (e.g. Thornton et al., 2014; Soil Survey Staff, 2022) to simulate plant and fire response to landscape heterogeneity. This would allow plant water and carbon to vary spatially, influencing both fire behavior and vegetation mortality, which feedback to alter hydrology and plant succession, influencing ecosystem trajectory and response to subsequent fire disturbance. Such a framework could also allow for exploration of the role of other natural disturbance, alongside species composition and competitive interactions, in determining fuel load and the likelihood of future fire.
Another way to link fine‐scale mechanistic models to coarser scale outcomes is through metamodels and nested model design. These techniques aim to find the crucial aspects of the mechanism that would be propagated at a higher level of spatial or temporal coarseness. Traditional or machine‐learning metamodels can emulate nonlinear processes at scales where simulation would be unfeasible (Sparks et al., 2011; Huang et al., 2016). These can be integrated into existing coarse‐scale models to propagate fine‐scale understanding in a more computationally efficient manner (Lu & Ricciuto, 2019). A nested model design can allow for experimentation at relevant scales of physiology and mechanism to constrain coarser models or metamodels. For example, species trait‐based functional types of fire behavior and effects can be created and used at coarser scales; while separately simulating component species at scales relevant to physiology, combustion, or fire injury (Neilson et al., 2005). Bayesian methods can further allow for integrating multiple nested models with observational data to constrain the performance of coarse‐scale models along with the propagation of uncertainty (Talluto et al., 2016).
V. Remote sensing of plant water and carbon to inform process modeling
Remote sensing of vegetation exploits the biophysical links between spectral information contained in the electromagnetic radiation reflected or emitted from the earth's surface and vegetation state, function, and dynamics. Optical data have been used to map LFMC using multispectral sensors at local (Marino et al., 2020), continental (Yebra et al., 2018a), and global (Quan et al., 2021) scales, producing mapping products at spatial resolutions from tens to thousands of meters. Visible spectrum reflectance is collected by many remote sensing platforms for vegetation monitoring purposes; but alone is insufficient for the prediction of LFMC. Addition of spectral data from the shortwave infrared (SWIR) region of the electromagnetic spectrum, currently collected by only a limited number of platforms, increases the accuracy of optical LFMC estimations (Yebra et al., 2018a). One challenge with direct optical estimation of fuel moisture is that satellite and airborne platforms output reflectance data as two‐dimensional coverages. In forested areas, these 2D data typically capture canopy conditions while obscuring understory characteristics (Yebra et al., 2018b), which are likely to differ widely from the overstory due to compositional and physiological differences. Similarly, these data are confounded by the presence of clouds, which are transient, yet exceedingly common, in many fire‐frequent areas of the world.
Acknowledgment of these challenges has led to research on the application of microwave remote sensing for direct and indirect fuel moisture content estimation (Fan et al., 2018; Wang et al., 2019). Typically, LFMC is estimated using radar‐derived vegetation optical depth, a proxy for vegetation water content (Moesinger et al., 2020). Radar systems operate in the microwave portion of the electromagnetic spectrum and can penetrate cloud cover and the upper canopy to detect the volume of liquid water in vegetation, offering an advantage relative to optical data. Nonetheless, a problem shared by direct microwave and passive optical measurements of LFMC in forests is the discrimination of signals related to the forest canopy, understory, surface litter, and soil (Gale et al., 2021). Merging microwave and optical data is a promising way forward. Rao et al. (2020) demonstrated that radar backscatter observations (from Sentinel‐1) directly enhanced LFMC predictability in comparison with only using optical reflectance (from Landsat‐8) for foliar, herbaceous and fine woody fuels. However, LFMC of denser woody fuels can only be estimated using longer wavelengths (i.e. L‐band radar; Tanase et al., 2015).
Despite these advances in mapping LFMC, little has been done to provide independent measures of dry mass and water content in the estimation of LFMC. To this end LMA (g m−2; also called dry matter content for remote sensing applications) and equivalent water thickness (EWT, g m−2) are appropriate target traits for optical sensing, capturing the effects of changing carbon allocation and moisture, respectively. Detection of EWT using optical sensors has a strong physical basis because water absorbs near infrared and short‐wave infrared radiation (Yebra et al., 2013). At the leaf scale, variation in LMA has a moderate influence on reflectance in the short‐wave infrared wavelengths (Feret et al., 2008) given the absorption features of structural properties found in leaves (Curran, 1989). For example, estimation of morphological and structural biochemical properties, including LMA, using reflectance spectroscopy of dried leaf material has been shown to be highly effective (e.g. Serbin et al., 2014). However, the presence of water in hydrated leaves can make the spectral estimation of dry matter content more challenging because water absorption masks the contribution of dry matter to the spectral response (Bowyer & Danson, 2004; Riano et al., 2005). Despite these challenges, the estimation of LMA at leaf and canopy scales has been shown to have reasonable accuracy using hyperspectral instruments (Ely et al., 2019; Chlus et al., 2020; Kamoske et al., 2021), likely due to covariance between EWT and LMA, as well as between leaf‐level functional properties and plant and canopy growth form and structure (Ollinger, 2011). Radiative transfer models have also been inverted using multispectral optical data to simultaneously estimate LMA and EWT and, therefore, LFMC (Yebra et al., 2013).
At smaller scales, techniques exist that allow for high‐temporal resolution vegetation carbon and water inventories. For instance, novel spectroscopic approaches leveraging high‐resolution spectra‐trait modeling (Serbin & Townsend, 2020; Burnett et al., 2021) provide the opportunity to more directly link spectral signatures with underlying physiology and plant condition (e.g. water and carbon content) in 2D (e.g. Dahlin et al., 2013; Singh et al., 2015) and 3D (Chlus et al., 2020; Kamoske et al., 2021). Likewise, dual‐wavelength lidar shows promise in moisture content estimation due to its ability to distinguish forest layers through ranging (Gale et al., 2021). These approaches are currently not available on space‐based platforms, limiting spatial coverage. However, these systems can be used together with ground‐based systems (e.g. imaging spectroscopy) to calibrate airborne and spaceborne hyperspectral (e.g. EnMAP, Guanter et al., 2016; SBG, Cawse‐Nicholson et al., 2021; OzFuel, ANU Institute for Space, 2021), thermal (ECOSTRESS, Anderson et al., 2021), and lidar (GEDI, Rishmawi et al., 2021) missions.
While current remote sensing LFMC products lack immediate predictive capability beneficial for fire management, they can be used to improve next‐generation process models directly (parameterization) or indirectly (benchmarking predictions). This will lead to improved simulations of fire behavior and spread under changing climate conditions. A challenge posed by the availability of remote sensing data is ensuring integration of data from disparate sources with existing databases for fuel characteristics and LFMC. New ‘big’ data systems are needed for standardized fusion of data and scalable dynamic updates to fuel data in a changing environment.
VI. Conclusions and future directions
Fire behavior and effects are intrinsically associated with plant physiology through water and carbon cycles. Integration of plant water and carbon process models with fine‐scale fire models will allow for the representation of process‐level feedbacks between fire behavior and effects, enabling the use of virtual experiments to explore vegetation responses to global change scenarios (increased temperature and vapor pressure deficit, change in precipitation) and resulting fire outcomes. Recent advances in remote sensing, in combination with upcoming sensor deployments, will enable high‐resolution mapping of plant water and carbon status across regions and seasons, providing parameterization and benchmarking for next‐generation models. Several areas are ripe for development across the disciplines of plant physiology, fire modeling, and remote sensing to fully enable these advances in understanding and simulation of vegetation–fire interactions. Furthermore, while the research presented here focuses on woody plants (trees and shrubs), additional efforts are needed to understand the interactions between climate, fire, and physiology of other vegetation types that contribute to fire regimes (Knapp, 1985; D'Antonio & Vitousek, 1992; Taylor et al., 2014; Simpson et al., 2016). Parallel advances on all frontiers will be critical to meet the urgency of the wildfire crisis.
With respect to fire behavior, we need to better understand the impacts of plant carbon and water on heat transfer and their interaction with fuel structure beyond the leaf scale. For fire effects, more studies are necessary to disentangle the physiological impacts of fire on tissue‐level water and carbon status, along with interactions across organs to enable scaling to whole plant function and mortality (Hood et al., 2018; Michaletz, 2018; Kleynhans et al., 2021). We need benchmark datasets of physiological mortality mechanisms across organs, species, size class, life history, geographic region, season, and under climate change scenarios (Hood et al., 2018). New terrestrial lidar approaches for evaluating fuels and fire effects that are based on structure and visual‐spectrum imagery may be a useful approach for determining these benchmarks and understanding vertical heterogeneity in fire impacts on plant organs at these scales (Gallagher et al., 2021; Pokswinski et al., 2021).
Fire model implementation needs to address the nonlinear biophysical‐processes of whole‐plant heat transfer and resulting fire behavior and effects based on dynamic physiology, including refined fire‐effects predictions stemming from an improved understanding of mechanisms (Kleynhans et al., 2021). Advances in machine learning can provide estimations of unknown process and enhance computational efficiency but require us to understand underlying processes and diagnose cases of overfitting. Directly computing many of these dynamics may be increasingly tractable as computation progresses. We additionally need to understand how the interaction among these dynamics affects fire and tree mortality at scales relevant to human decision‐making. Using the inherent advantage of each scale of fire models, nested model design can work to incorporate new understanding at multiple scales (for example, plant physiology, stand management of plant density, landscape fire spread and suppression, and global carbon balance). Developing and validating nested model designs will be crucial to understand how each scale of organization influences the larger scale of organization.
Advancements in model implementation require finer spatial and temporal resolution observations of important processes to fire behavior and effects. Concerning remotely sensed drivers, we need to disentangle the numerous contributions to foliar reflectance signatures, along with advanced techniques for detection of subcanopy vegetation (Gale et al., 2021). Improved spatial and spectral resolution will reduce uncertainty in characterization of water and carbon status across regions and seasons, and better statistical methods will make outcomes more robust across larger areas and disparate biomes (Yebra et al., 2013). Advancements in data reduction and/or model ingestion will be needed to allow effective use of this higher resolution data. Ground‐based experiments and observations can be used to better describe vegetation–fire interactions, and every prescribed burn provides an opportunity for data collection and model validation.
Nelson (2001) suggested that ‘the complexity of the task may have precluded all attempts to develop a reasonably complete physics‐ and physiology‐based model’ of live fuel moisture. With an increasing wealth of drought physiology research, the advent of next‐generation models, and new spaceborne sensor deployments on the horizon, we are finally poised to tackle the problem of providing a dynamic and mechanistic description of fire behavior and effects through vegetation processes. Doing so will better equip us and the ecosystems we depend on to survive and thrive in a future made uncertain by global change.
Competing interests
None declared.
Author contributions
This manuscript was the outcome of a three‐day workshop hosted by Los Alamos National Laboratory in January 2021. LTD, AKJ and RRL planned and designed the workshop. ADC facilitated the workshop. LTD, AKJ, ALA, MRG, JKH, CMH, SMH, ELL, WMJ, RHN, JJO, RAP, FP, JR, SPS, SS, JKS, NSS, DRW, CX and NY gave presentations at the workshop. J‐LD, VRD, and MY contributed to presentations but were unable to attend. LTD, AKJ, and ADC moderated Q&A sessions. LTD, AKJ, RRL, ALA, JKH, WMJ, ELL, JJO and SPS moderated breakout sessions. LTD, AKJ, RRL, IA, ALA, ADC, MRG, JKH, CMH, SMH, MDH, WMJ, AJ, ELL, WM, STM, RHN, JJO, RAP, RP‐F, FP, JR, KAS, ES‐F, SPS, SS, JKS, CHS, NSS, DRW, MW, CX and NY participated in the workshop. LTD and AKJ led manuscript development and writing. AB and ZJR were consulted for subject matter expertise on the manuscript. All authors contributed to manuscript writing and editing.
Acknowledgements
We would like to thank three anonymous reviewers for their time and effort in providing valuable feedback which greatly improved the quality of this manuscript. We also acknowledge all participants of Los Alamos National Laboratory's January 2021 Workshop for their contributions to this effort. LTD, AJ, RRL and SS were supported by the Los Alamos National Laboratory (LANL) through its Center for Space and Earth Science (CSES). Center for Space and Earth Science is funded by LANL's Laboratory Directed Research and Development (LDRD) program under project no. 20210528CR. AJ and ZJR received additional funding from LANL LDRD under project no. 20210689ECR. RPF and STM were supported by SERDP project RC18‐1346 and an NSERC Discovery Grant. AB acknowledges funding from the Austrian Science Fund (FWF, project P32203) and from the University of Innsbruck (Early‐Stage Funding, grant W‐171705). MY receives funding from the Australian Research Council, the Australian Research Data Commons, The SmartSat Cooperative Research Centre and Singtel Optus Pty Limited. JKS was supported by the National Center for Atmospheric Research, a major facility sponsored by the National Science Foundation (NSF) under Cooperative Agreement no. 1852977, with additional support from NASA Arctic Boreal Vulnerability Experiment Grant 80NSSC19M0107. JKS, SPS and CX were also supported as part of the Next‐Generation Ecosystem Experiments – Tropics, funded by the US Department of Energy, Office of Science, Office of Biological and Environmental Research. SPS was also partially supported by the NASA Surface Biology and Geology Mission Study (NNG20OB24A) and through the United States Department of Energy contract no. DE‐SC0012704 to Brookhaven National Laboratory. CMH acknowledges US Department of Defense (DoD) Strategic Environmental Research and Development Program (SERDP) Project RC19‐1119. IA declares support from NSF WIFIRE Commons under grants 2040676 and 2134904. VRD acknowledges funding from MICINN projects RTI2018‐094691‐B‐C31; EU H2020 (grant agreements 101003890). RAP acknowledges support from US Department of Defense Strategic Environmental Research and Development Program's Closing Gaps Project RC20‐1025. USDA Forest Service personnel were supported by annual Forest Service appropriations. This document has been approved for unlimited release under LA‐UR‐22‐20430.
References
- Ackerly D. 2004. Functional strategies of chaparral shrubs in relation to seasonal water deficit and disturbance. Ecological Monographs 74: 25–44. [Google Scholar]
- Alam MA, Wyse SV, Buckley HL, Perry GLW, Sullivan JJ, Mason NWH, Buxton R, Richardson SJ, Curran TJ. 2020. Shoot flammability is decoupled from leaf flammability, but controlled by leaf functional traits. Journal of Ecology 108: 641–653. [Google Scholar]
- Albini FA. 1976. Estimating wildfire behavior and effects. General technical report, INT‐30. Ogden, UT, USA: USDA Forest Service, Intermountain Forest and Range Experiment Station. [Google Scholar]
- Anderson MC, Yang Y, Xue J, Knipper KR, Yang Y, Gao F, Hain CR, Kustas WP, Cawse‐Nicholson K, Hulley G et al. 2021. Interoperability of ECOSTRESS and Landsat for mapping evapotranspiration time series at sub‐field scales. Remote Sensing of Environment 252: 112189. [Google Scholar]
- Andrews PL. 2013. Current status and future needs of the BehavePlus Fire Modeling System. International Journal of Wildland Fire 23: 21–33. [Google Scholar]
- ANU Institute for Space . 2021. OzFuel pre‐phase A study: Australian forest fuel monitoring from space . [WWW document] URL inspace.anu.edu.au/activity/missions/ozfuel [accessed 13 January 2022].
- Atchley AL, Linn R, Jonko A, Hoffman C, Hyman JD, Pimont F, Sieg C, Middleton RS. 2021. Effects of fuel spatial distribution on wildland fire behaviour. International Journal of Wildland Fire 30: 179. [Google Scholar]
- Baffoin R, Charrier G, Bouchardon A‐E, Bonhomme M, Améglio T, Lacointe A. 2021. Seasonal changes in carbohydrates and water content predict dynamics of frost hardiness in various temperate tree species. Tree Physiology 41: 1583–1600. [DOI] [PubMed] [Google Scholar]
- Balaguer‐Romano R, Díaz‐Sierra R, Madrigal J, Voltas J, Resco de Dios V. 2020. Needle senescence affects fire behavior in Aleppo Pine (Pinus halepensis Mill.) stands: a simulation study. Forests 11: 1054. [Google Scholar]
- Banerjee T, Heilman W, Goodrick S, Hiers JK, Linn R. 2020. Effects of canopy midstory management and fuel moisture on wildfire behavior. Scientific Reports 10: 17312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bär A, Michaletz ST, Mayr S. 2019. Fire effects on tree physiology. New Phytologist 223: 1728–1741. [DOI] [PubMed] [Google Scholar]
- Bär A, Nardini A, Mayr S. 2018. Post‐fire effects in xylem hydraulics of Picea abies, Pinus sylvestris and Fagus sylvatica . New Phytologist 217: 1484–1493. [DOI] [PubMed] [Google Scholar]
- Bär A, Schröter DM, Mayr S. 2021. When the heat is on: high temperature resistance of buds from European tree species. Plant, Cell & Environment 44: 2593–2603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barker Plotkin A, Blumstein M, Laflower D, Pasquarella VJ, Chandler JL, Elkinton JS, Thompson JR. 2021. Defoliated trees die below a critical threshold of stored carbon. Functional Ecology 35: 2156–2167. [Google Scholar]
- Berenguer E, Lennox GD, Ferreira J, Malhi Y, Aragão LEOC, Barreto JR, Del Bon Espírito‐Santo F, Figueiredo AES, França F, Gardner TA et al. 2021. Tracking the impacts of El Niño drought and fire in human‐modified Amazonian forests. Proceedings of the National Academy of Sciences, USA 118: e2019377118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bianchi LO, Oddi FJ, Muñoz M, Defossé GE. 2019. Comparison of leaf moisture content and ignition characteristics among native species and exotic conifers in northwestern Patagonia, Argentina. Forest Science 65: 375–386. [Google Scholar]
- Boardman CR, Dietenberger MA, Weise DR. 2021. Specific heat capacity of wildland foliar fuels to 434°C. Fuel 292: 120396. [Google Scholar]
- Bond WJ. 2021. Out of the shadows: ecology of open ecosystems. Plant Ecology & Diversity 14: 205–222. [Google Scholar]
- Bond WJ, Keeley JE. 2005. Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends in Ecology & Evolution 20: 387–394. [DOI] [PubMed] [Google Scholar]
- Bond WJ, van Wilgen BW. 1996. Fire and plants. London, UK: Chapman & Hall. [Google Scholar]
- Bowman DMJS, Balch JK, Artaxo P, Bond WJ, Carlson JM, Cochrane MA, D'Antonio CM, DeFries RS, Doyle JC, Harrison SP et al. 2009. Fire in the Earth system. Science 324: 481–484. [DOI] [PubMed] [Google Scholar]
- Bowman WD, Roberts SW. 1985. Seasonal and diurnal water relations adjustments in three evergreen chaparral shrubs. Ecology 66: 738–742. [Google Scholar]
- Bowyer P, Danson FM. 2004. Sensitivity of spectral reflectance to variation in live fuel moisture content at leaf and canopy level. Remote Sensing of Environment 92: 297–308. [Google Scholar]
- Burnett AC, Anderson J, Davidson KJ, Ely KS, Lamour J, Li Q, Morrison BD, Yang D, Rogers A, Serbin SP. 2021. A best‐practice guide to predicting plant traits from leaf‐level hyperspectral data using partial least squares regression. Journal of Experimental Botany 72: 6175–6189. [DOI] [PubMed] [Google Scholar]
- Butler BW, Dickinson MB. 2010. Tree injury and mortality in fires: developing process‐based models. Fire Ecology 6: 55–79. [Google Scholar]
- Byram GM. 1959. Combustion of forest fuels. In: Davis KP, ed. Forest fire: control and use. New York, NY, USA: McGraw‐Hill, 61–89. [Google Scholar]
- Cawse‐Nicholson K, Townsend PA, Schimel D, Assiri AM, Blake PL, Buongiorno MF, Campbell P, Carmon N, Casey KA, Correa‐Pabón RE et al. 2021. NASA's surface biology and geology designated observable: a perspective on surface imaging algorithms. Remote Sensing of Environment 257: 112349. [Google Scholar]
- Chapin FSI, Schulze E‐D, Mooney HA. 1990. The ecology and economics of storage in plants. Annual Review of Ecology and Systematics 2: 423–447. [Google Scholar]
- Chatziefstratiou EK, Bohrer G, Bova AS, Subramanian R, Frasson RPM, Scherzer A, Butler BW, Dickinson MB. 2013. FireStem2D – a two‐dimensional heat transfer model for simulating tree stem injury in fires. PLoS ONE 8: e70110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chlus A, Kruger EL, Townsend PA. 2020. Mapping three‐dimensional variation in leaf mass per area with imaging spectroscopy and lidar in a temperate broadleaf forest. Remote Sensing of Environment 250: 112043. [Google Scholar]
- Christoffersen BO, Gloor M, Fauset S, Fyllas NM, Galbraith DR, Baker TR, Kruijt B, Rowland L, Fisher RA, Binks OJ et al. 2016. Linking hydraulic traits to tropical forest function in a size‐structured and trait‐driven model (TFS v.1‐Hydro). Geoscientific Model Development 9: 4227–4255. [Google Scholar]
- Cochard H, Pimont F, Ruffault J, Martin‐StPaul N. 2021. SurEau: a mechanistic model of plant water relations under extreme drought. Annals of Forest Science 78: 55. [Google Scholar]
- Countryman CM. 1966. The concept of the fire environment. Fire Control Notes 27: 8–10. [Google Scholar]
- Cruz MG, Alexander ME, Fernandes PM. 2022. Evidence for lack of a fuel effect on forest and shrubland fire rates of spread under elevated fire danger conditions: implications for modelling and management. International Journal of Wildland Fire 31: 471–479. [Google Scholar]
- Cruz MG, Gould JS, Kidnie S, Bessell R, Nichols D, Slijepcevic A, Cruz MG, Gould JS, Kidnie S, Bessell R et al. 2015. Effects of curing on grassfires: II. Effect of grass senescence on the rate of fire spread. International Journal of Wildland Fire 24: 838–848. [Google Scholar]
- Curran PJ. 1989. Remote sensing of foliar chemistry. Remote Sensing of Environment 30: 271–278. [Google Scholar]
- Dahlin KM, Asner GP, Field CB. 2013. Environmental and community controls on plant canopy chemistry in a Mediterranean‐type ecosystem. Proceedings of the National Academy of Sciences, USA 110: 6895–6900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Antonio CM, Vitousek PM. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23: 63–87. [Google Scholar]
- Davis SD, Mooney HA. 1986. Water use patterns of four co‐occurring chaparral shrubs. Oecologia 7: 172–177. [DOI] [PubMed] [Google Scholar]
- Deeming JE, Cohen JD, Burgan RE. 1977. The National fire‐danger rating system – 1978. General technical report, INT‐GTR‐39. Ogden, UT, USA: US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. [Google Scholar]
- Dennison PE, Moritz MA. 2009. Critical live fuel moisture in chaparral ecosystems: a threshold for fire activity and its relationship to antecedent precipitation. International Journal of Wildland Fire 18: 1021–1027. [Google Scholar]
- Dietenberger MA, Boardman CR, Shotorban B, Mell W, Weise DR. 2020. Thermal degradation modeling of live vegetation for fire dynamic simulator. In: Proceedings, 2020 Spring technical meeting, Central States Section of the Combustion Institute, 1–20.
- Ely KS, Burnett AC, Lieberman‐Cribbin W, Serbin SP, Rogers A. 2019. Spectroscopy can predict key leaf traits associated with source–sink balance and carbon–nitrogen status. Journal of Experimental Botany 70: 1789–1799. [DOI] [PubMed] [Google Scholar]
- Fan L, Wigneron J‐P, Xiao Q, Al‐Yaari A, Wen J, Martin‐StPaul N, Dupuy J‐L, Pimont F, Al Bitar A, Fernandez‐Moran R et al. 2018. Evaluation of microwave remote sensing for monitoring live fuel moisture content in the Mediterranean region. Remote Sensing of Environment 205: 210–223. [Google Scholar]
- Feret J‐B, François C, Asner GP, Gitelson AA, Martin RE, Bidel LPR, Ustin SL, le Maire G, Jacquemoud S. 2008. PROSPECT‐4 and 5: advances in the leaf optical properties model separating photosynthetic pigments. Remote Sensing of Environment 112: 3030–3043. [Google Scholar]
- Fettig CJ, Runyon JB, Homicz CS, James PMA, Ulyshen MD. 2022. Fire and insect interactions in North American forests. Current Forestry Reports 8: 301–316. [Google Scholar]
- Finney MA. 1998. Farsite: fire area simulator‐model development and evaluation. Res. Pap. RMRS‐RP‐4, Revised 2004. Ogden, UT, USA: US Department of Agriculture, Forest Service, Rocky Mountain Research Station. [Google Scholar]
- Finnigan J. 2000. Turbulence in plant canopies. Annual Review of Fluid Mechanics 32: 519–571. [Google Scholar]
- Fletcher TH, Pickett BM, Smith SG, Spittle GS, Woodhouse MM, Haake E, Weise DR. 2007. Effects of moisture on ignition behavior of moist California chaparral and Utah leaves. Combustion Science and Technology 179: 1183–1203. [Google Scholar]
- Franco AC, Rossatto DR, de Carvalho Ramos Silva L, da Silva Ferreira C. 2014. Cerrado vegetation and global change: the role of functional types, resource availability and disturbance in regulating plant community responses to rising CO2 levels and climate warming. Theoretical and Experimental Plant Physiology 26: 19–38. [Google Scholar]
- Furniss TJ, Hessburg PF, Povak NA, Salter RB, Wigmosta MS. 2022. Predicting future patterns, processes, and their interactions: benchmark calibration and validation procedures for forest landscape models. Ecological Modelling 473: 110099. [Google Scholar]
- Gale MG, Cary GJ, Van Dijk AIJM, Yebra M. 2021. Forest fire fuel through the lens of remote sensing: review of approaches, challenges and future directions in the remote sensing of biotic determinants of fire behaviour. Remote Sensing of Environment 255: 112282. [Google Scholar]
- Gallagher MR, Maxwell AE, Guillén LA, Everland A, Loudermilk EL, Skowronski NS. 2021. Estimation of plot‐level burn severity using terrestrial laser scanning. Remote Sensing 13: 4168. [Google Scholar]
- Gettelman A, Geer AJ, Forbes RM, Carmichael GR, Feingold G, Posselt DJ, Stephens GL, van den Heever SC, Varble AC, Zuidema P. 2022. The future of Earth system prediction: advances in model‐data fusion. Science Advances 8: eabn3488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giglio L, Randerson JT, van der Werf GR. 2013. Analysis of daily, monthly, and annual burned area using the fourth‐generation global fire emissions database (GFED4). Journal of Geophysical Research: Biogeosciences 118: 317–328. [Google Scholar]
- Grootemaat S, Wright IJ, van Bodegom PM, Cornelissen JHC, Cornwell WK. 2015. Burn or rot: leaf traits explain why flammability and decomposability are decoupled across species. Functional Ecology 29: 1486–1497. [Google Scholar]
- Grossiord C, Buckley TN, Cernusak LA, Novick KA, Poulter B, Siegwolf RTW, Sperry JS, McDowell NG. 2020. Plant responses to rising vapor pressure deficit. New Phytologist 226: 1550–1566. [DOI] [PubMed] [Google Scholar]
- Guanter L, Kaufmann H, Foerster S, Brosinsky A, Wulf H, Bochow M, Boesche N, Brell M, Buddenbaum H, Chabrillat S et al. 2016. EnMap science plan. Potsdam, Germany: GFZ Data Services. [Google Scholar]
- Hammond WM, Yu K, Wilson LA, Will RE, Anderegg WRL, Adams HD. 2019. Dead or dying? Quantifying the point of no return from hydraulic failure in drought‐induced tree mortality. New Phytologist 223: 1834–1843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiers JK, O'Brien JJ, Varner JM, Butler BW, Dickinson M, Furman J, Gallagher M, Godwin D, Goodrick SL, Hood SM et al. 2020. Prescribed fire science: the case for a refined research agenda. Fire Ecology 16: 11. [Google Scholar]
- Hoffman CM, Sieg CH, Linn RR, Mell W, Parsons RA, Ziegler JP, Hiers JK. 2018. Advancing the science of wildland fire dynamics using process‐based models. Fire 1: 32. [Google Scholar]
- Hood SM. 2021. Physiological responses to fire that drive tree mortality. Plant, Cell & Environment 44: 692–695. [DOI] [PubMed] [Google Scholar]
- Hood SM, Varner JM, van Mantgem P, Cansler CA. 2018. Fire and tree death: understanding and improving modeling of fire‐induced tree mortality. Environmental Research Letters 13: 113004. [Google Scholar]
- Hough WA, Albini FA. 1978. Predicting fire behavior in palmetto‐gallberry fuel complexes. Research paper SE‐RP‐174. Asheville, NC, USA: USDA Forest Service, Southeastern Forest Experiment Station, 48 p. [Google Scholar]
- Huang M, Ray J, Hou Z, Ren H, Liu Y, Swiler L. 2016. On the applicability of surrogate‐based Markov chain Monte Carlo‐Bayesian inversion to the Community Land Model: case studies at flux tower sites. Journal of Geophysical Research: Atmospheres 121: 7548–7563. [Google Scholar]
- Ibanez T, Platt WJ, Bellingham PJ, Vieilledent G, Franklin J, Martin PH, Menkes C, Pérez‐Salicrup DR, Russell‐Smith J, Keppel G. 2022. Altered cyclone–fire interactions are changing ecosystems. Trends in Plant Science 27: 1218–1230. [DOI] [PubMed] [Google Scholar]
- Jolly WM. 2007. Sensitivity of a surface fire spread model and associated fire behaviour fuel models to changes in live fuel moisture. International Journal of Wildland Fire 16: 503–509. [Google Scholar]
- Jolly WM, Hadlow AM, Huguet K, Jolly WM, Hadlow AM, Huguet K. 2014. De‐coupling seasonal changes in water content and dry matter to predict live conifer foliar moisture content. International Journal of Wildland Fire 23: 480–489. [Google Scholar]
- Jolly WM, Hintz J, Linn RL, Kropp RC, Conrad ET, Parsons RA, Winterkamp J. 2016. Seasonal variations in red pine (Pinus resinosa) and jack pine (Pinus banksiana) foliar physio‐chemistry and their potential influence on stand‐scale wildland fire behavior. Forest Ecology and Management 373: 167–178. [Google Scholar]
- Jolly WM, Johnson DM. 2018. Pyro‐ecophysiology: shifting the paradigm of live wildland fuel research. Fire 1: 8. [Google Scholar]
- Kamoske AG, Dahlin KM, Serbin SP, Stark SC. 2021. Leaf traits and canopy structure together explain canopy functional diversity: an airborne remote sensing approach. Ecological Applications 31: e02230. [DOI] [PubMed] [Google Scholar]
- Karavani A, Boer MM, Baudena M, Colinas C, Díaz‐Sierra R, Pemán J, de Luis M, Enríquez‐de‐Salamanca Á, Resco de Dios V. 2018. Fire‐induced deforestation in drought‐prone Mediterranean forests: drivers and unknowns from leaves to communities. Ecological Monographs 88: 141–169. [Google Scholar]
- Karp AT, Faith JT, Marlon JR, Staver AC. 2021. Global response of fire activity to late Quaternary grazer extinctions. Science 374: 1145–1148. [DOI] [PubMed] [Google Scholar]
- Keane RE, Cary GJ, Davies ID, Flannigan MD, Gardner RH, Lavorel S, Lenihan JM, Li C, Rupp TS. 2004. A classification of landscape fire succession models: spatial simulations of fire and vegetation dynamics. Ecological Modelling 179: 3–27. [Google Scholar]
- Keane RE, Loehman RA, Holsinger LM. 2011. The FireBgcv2 landscape fire and succession model: a research simulation platform for exploring fire and vegetation dynamics. Gen. Tech. Rep. RMRS‐GTR‐255. Fort Collins, CO, USA: US Department of Agriculture, Forest Service, Rocky Mountain Research Station. [Google Scholar]
- Keeley JE. 1998. Coupling demography, physiology and evolution in chaparral shrubs. In: Rundel PW, Montenegro G, Jaksic FM, eds. Ecological studies. Landscape disturbance and biodiversity in Mediterranean‐type ecosystems. Berlin, Heidelberg, Germany: Springer, 257–264. [Google Scholar]
- Kilzer FJ, Broido A. 1965. Speculations on the nature of cellulose pyrolysis. Pyrodynamics 2: 151–163. [Google Scholar]
- Kleynhans EJ, Atchley AL, Michaletz ST. 2021. Chapter Eleven – Modeling fire effects on plants: from organs to ecosystems. In: Johnson EA, Miyanishi K, eds. Plant disturbance ecology, 2nd edn. San Diego, CA, USA: Academic Press, 383–421. [Google Scholar]
- Knapp AK. 1985. Effect of fire and drought on the ecophysiology of Andropogon gerardii and Panicum virgatum in a tallgrass prairie. Ecology 66: 1309–1320. [Google Scholar]
- Knapp AK, Fahnestock JT, Hamburg SP, Statland LB, Seastedt TR, Schimel DS. 1993. Landscape patterns in soil–plant water relations and primary production in tallgrass prairie. Ecology 74: 549–560. [Google Scholar]
- Knapp EE, Estes BL, Skinner CN. 2009. Ecological effects of prescribed fire season: a literature review and synthesis for managers. Gen. Tech. Rep. PSW‐GTR‐224. Albany, CA, USA: US Department of Agriculture, Forest Service, Pacific Southwest Research Station. [Google Scholar]
- Krawchuk MA, Haire SL, Coop J, Parisien M‐A, Whitman E, Chong G, Miller C. 2016. Topographic and fire weather controls of fire refugia in forested ecosystems of northwestern North America. Ecosphere 7: e01632. [Google Scholar]
- Lee EH, Beedlow PA, Brooks JR, Tingey DT, Wickham C, Rugh W. 2022. Physiological responses of Douglas‐fir to climate and forest disturbances as detected by cellulosic carbon and oxygen isotope ratios. Tree Physiology 42: 5–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li F, Lawrence DM. 2017. Role of fire in the global land water budget during the twentieth century due to changing ecosystems. Journal of Climate 30: 1893–1908. [Google Scholar]
- Linn R, Reisner J, Colman J, Winterkamp J. 2002. Studying wildfire behavior using Firetec . International Journal of Wildland Fire 11: 233–246. [Google Scholar]
- Louarn G, Song Y. 2020. Two decades of functional–structural plant modelling: now addressing fundamental questions in systems biology and predictive ecology. Annals of Botany 126: 501–509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu D, Ricciuto D. 2019. Efficient surrogate modeling methods for large‐scale Earth system models based on machine‐learning techniques. Geoscientific Model Development 12: 1791–1807. [Google Scholar]
- Ma W, Zhai L, Pivovaroff A, Shuman J, Buotte P, Ding J, Christoffersen B, Knox R, Moritz M, Fisher RA et al. 2021. Assessing climate change impacts on live fuel moisture and wildfire risk using a hydrodynamic vegetation model. Biogeosciences 18: 4005–4020. [Google Scholar]
- Macias Fauria M, Michaletz ST, Johnson EA. 2011. Predicting climate change effects on wildfires requires linking processes across scales. WIREs Climate Change 2: 99–112. [Google Scholar]
- de Magalhães RMQ, Schwilk DW. 2012. Leaf traits and litter flammability: evidence for non‐additive mixture effects in a temperate forest. Journal of Ecology 100: 1153–1163. [Google Scholar]
- de Magalhães RQ, Schwilk DW. 2021. Moisture absorption and drying alter nonadditive litter flammability in a mixed conifer forest. Journal of Ecology 109: 234–245. [Google Scholar]
- van Mantgem PJ, Falk DA, Williams EC, Das AJ, Stephenson NL. 2018. Pre‐fire drought and competition mediate post‐fire conifer mortality in western U.S. National Parks. Ecological Applications 28: 1730–1739. [DOI] [PubMed] [Google Scholar]
- Marino E, Yebra M, Guillén‐Climent M, Algeet N, Tomé JL, Madrigal J, Guijarro M, Hernando C. 2020. Investigating live fuel moisture content estimation in fire‐prone shrubland from remote sensing using empirical modelling and RTM simulations. Remote Sensing 12: 2251. [Google Scholar]
- Martínez‐Vilalta J, Sala A, Asensio D, Galiano L, Hoch G, Palacio S, Piper FI, Lloret F. 2016. Dynamics of non‐structural carbohydrates in terrestrial plants: a global synthesis. Ecological Monographs 86: 495–516. [Google Scholar]
- Martin‐StPaul N, Ruffault J, Pimont F, Dupuy J‐L. 2018. Live fuel moisture content: variability, predictability and impact on fire behavior and activity. In: Viegas DX, ed. Advances in forest fire research. Coimbra, Portugal: Imprensa da Universidade de Coimbra, 246–253. [Google Scholar]
- Matt FJ, Dietenberger MA, Weise DR. 2020. Summative and ultimate analysis of live leaves from Southern U.S. forest plants for use in fire modeling. Energy & Fuels 34: 4703–4720. [Google Scholar]
- McAllister S, Weise DR. 2017. Effects of season on ignition of live wildland fuels using the forced ignition and flame spread test apparatus. Combustion Science and Technology 189: 231–247. [Google Scholar]
- Mell W, Maranghides A, McDermott R, Manzello SL. 2009. Numerical simulation and experiments of burning Douglas fir trees. Combustion and Flame 156: 2023–2041. [Google Scholar]
- Michaletz ST. 2018. Xylem dysfunction in fires: towards a hydraulic theory of plant responses to multiple disturbance stressors. New Phytologist 217: 1391–1393. [DOI] [PubMed] [Google Scholar]
- Michaletz ST, Johnson EA. 2006. Foliage influences forced convection heat transfer in conifer branches and buds. New Phytologist 170: 87–98. [DOI] [PubMed] [Google Scholar]
- Michaletz ST, Johnson EA. 2007. How forest fires kill trees: a review of the fundamental biophysical processes. Scandinavian Journal of Forest Research 22: 500–515. [Google Scholar]
- Michaletz ST, Johnson EA. 2008. A biophysical process model of tree mortality in surface fires. Canadian Journal of Forest Research 38: 2013–2029. [Google Scholar]
- Michaletz ST, Johnson EA, Mell WE, Greene DF. 2013. Timing of fire relative to seed development may enable non‐serotinous species to recolonize from the aerial seed banks of fire‐killed trees. Biogeosciences 10: 5061–5078. [Google Scholar]
- Michaletz ST, Johnson EA, Tyree MT. 2012. Moving beyond the cambium necrosis hypothesis of post‐fire tree mortality: cavitation and deformation of xylem in forest fires. New Phytologist 194: 254–263. [DOI] [PubMed] [Google Scholar]
- Midgley JJ, Kruger LM, Skelton R. 2011. How do fires kill plants? The hydraulic death hypothesis and Cape Proteaceae ‘fire‐resisters’. South African Journal of Botany 77: 381–386. [Google Scholar]
- Milla R, Palacio S, Maestro‐Martínez M, Montserrat‐Martí G. 2007. Leaf exchange in a Mediterranean shrub: water, nutrient, non‐structural carbohydrate and osmolyte dynamics. Tree Physiology 27: 951–960. [DOI] [PubMed] [Google Scholar]
- Mitchell SR, Harmon ME, O'Connell KEB. 2009. Forest fuel reduction alters fire severity and long‐term carbon storage in three Pacific Northwest ecosystems. Ecological Applications 19: 643–655. [DOI] [PubMed] [Google Scholar]
- Moesinger L, Dorigo W, de Jeu R, van der Schalie R, Scanlon T, Teubner I, Forkel M. 2020. The global long‐term microwave vegetation optical depth climate archive (VODCA). Earth System Science Data 12: 177–196. [Google Scholar]
- Neilson RP, Pitelka LF, Solomon AM, Nathan R, Midgley GF, Fragoso JMV, Lischke H, Thompson K. 2005. Forecasting regional to global plant migration in response to climate change. Bioscience 55: 749–759. [Google Scholar]
- Nelson RM. 2001. Chapter 4 – Water relations of forest fuels. In: Johnson EA, Miyanishi K, eds. Forest fires. San Diego, CA, USA: Academic Press, 79–149. [Google Scholar]
- Nolan RH, Blackman CJ, de Dios VR, Choat B, Medlyn BE, Li X, Bradstock RA, Boer MM. 2020. Linking forest flammability and plant vulnerability to drought. Forests 11: 779. [Google Scholar]
- Nolan RH, Boer MM, Resco de Dios V, Caccamo G, Bradstock RA. 2016. Large‐scale, dynamic transformations in fuel moisture drive wildfire activity across southeastern Australia. Geophysical Research Letters 43: 4229–4238. [Google Scholar]
- Nolan RH, Collins L, Leigh A, Ooi MKJ, Curran TJ, Fairman TA, Resco de Dios V, Bradstock R. 2021. Limits to post‐fire vegetation recovery under climate change. Plant, Cell & Environment 44: 3471–3489. [DOI] [PubMed] [Google Scholar]
- Nolan RH, Hedo J, Arteaga C, Sugai T, Resco de Dios V. 2018. Physiological drought responses improve predictions of live fuel moisture dynamics in a Mediterranean forest. Agricultural and Forest Meteorology 263: 417–427. [Google Scholar]
- O'Brien JJ, Hiers JK, Varner JM, Hoffman CM, Dickinson MB, Michaletz ST, Loudermilk EL, Butler BW. 2018. Advances in mechanistic approaches to quantifying biophysical fire effects. Current Forestry Reports 4: 161–177. [Google Scholar]
- O'Brien JJ, Kevin Hiers J, Mitchell RJ, Varner JM, Mordecai K. 2010. Acute physiological stress and mortality following fire in a long‐unburned longleaf pine ecosystem. Fire Ecology 6: 1–12. [Google Scholar]
- O'Brien JJ, Loudermilk EL, Hiers JK, Pokswinski SM, Hornsby B, Hudak AT, Strother D, Rowell E, Bright BC. 2016. Canopy‐derived fuels drive patterns of in‐fire energy release and understory plant mortality in a longleaf pine (Pinus palustris) sandhill in Northwest Florida, USA. Canadian Journal of Remote Sensing 42: 489–500. [Google Scholar]
- Ollinger SV. 2011. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytologist 189: 375–394. [DOI] [PubMed] [Google Scholar]
- Parsons RA, Mell WE, McCauley P. 2011. Linking 3D spatial models of fuels and fire: effects of spatial heterogeneity on fire behavior. Ecological Modelling 222: 679–691. [Google Scholar]
- Parsons RA, Pimont F, Wells L, Cohn G, Jolly WM, de Coligny F, Rigolot E, Dupuy J‐L, Mell W, Linn RR. 2018. Modeling thinning effects on fire behavior with STANDFIRE. Annals of Forest Science 75: 7. [Google Scholar]
- Parsons RA, Wells L, Linn R, Hiers K, Marcozzi A, Rosales D. 2020. FastFuels: 3D fuels for next generation fire models . Missoula Fire Sciences Laboratory, MT, USA. [WWW document] URL https://www.firelab.org/project/fastfuels‐3d‐fuels‐next‐generation‐fire‐models [accessed 16 January 2022].
- Partelli‐Feltrin R, Johnson DM, Sparks AM, Adams HD, Kolden CA, Nelson AS, Smith AMS. 2020. Drought increases vulnerability of Pinus ponderosa saplings to fire‐induced mortality. Fire 3: 56. [Google Scholar]
- Partelli‐Feltrin R, Smith AMS, Adams HD, Kolden CA, Johnson DM. 2021. Short‐ and long‐term effects of fire on stem hydraulics in Pinus ponderosa saplings. Plant, Cell & Environment 44: 696–705. [DOI] [PubMed] [Google Scholar]
- Pausas JG, Moreira B. 2012. Flammability as a biological concept. New Phytologist 194: 610–613. [DOI] [PubMed] [Google Scholar]
- Pimont F, Parsons R, Rigolot E, de Coligny F, Dupuy J‐L, Dreyfus P, Linn RR. 2016. Modeling fuels and fire effects in 3D: model description and applications. Environmental Modelling & Software 80: 225–244. [Google Scholar]
- Pimont F, Ruffault J, Martin‐StPaul NK, Dupuy J‐L, Pimont F, Ruffault J, Martin‐StPaul NK, Dupuy J‐L. 2019a. Why is the effect of live fuel moisture content on fire rate of spread underestimated in field experiments in shrublands? International Journal of Wildland Fire 28: 127–137. [Google Scholar]
- Pimont F, Ruffault J, Martin‐StPaul NK, Dupuy J‐L, Pimont F, Ruffault J, Martin‐StPaul NK, Dupuy J‐L. 2019b. A cautionary note regarding the use of cumulative burnt areas for the determination of fire danger index breakpoints. International Journal of Wildland Fire 28: 254–258. [Google Scholar]
- Pokswinski S, Gallagher MR, Skowronski NS, Loudermilk EL, Hawley C, Wallace D, Everland A, Wallace J, Hiers JK. 2021. A simplified and affordable approach to forest monitoring using single terrestrial laser scans and transect sampling. MethodsX 8: 101484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poorter H, Niinemets Ü, Poorter L, Wright IJ, Villar R. 2009. Causes and consequences of variation in leaf mass per area (LMA): a meta‐analysis. New Phytologist 182: 565–588. [DOI] [PubMed] [Google Scholar]
- Pratt RB, Jacobsen A, Jacobs S, Esler K. 2012. Xylem transport safety and efficiency differ among fynbos shrub life history types and between two sites differing in mean rainfall. International Journal of Plant Sciences 173: 474–483. [Google Scholar]
- Pratt RB, Jacobsen AL, Ramirez AR, Helms AM, Traugh CA, Tobin MF, Heffner MS, Davis SD. 2014. Mortality of resprouting chaparral shrubs after a fire and during a record drought: physiological mechanisms and demographic consequences. Global Change Biology 20: 893–907. [DOI] [PubMed] [Google Scholar]
- Quan X, Yebra M, Riaño D, He B, Lai G, Liu X. 2021. Global fuel moisture content mapping from MODIS. International Journal of Applied Earth Observation and Geoinformation 101: 102354. [Google Scholar]
- Quintiere JG. 2006. Fundamentals of fire phenomena. Chichester, UK: Wiley. [Google Scholar]
- Rabin SS, Melton JR, Lasslop G, Bachelet D, Forrest M, Hantson S, Kaplan JO, Li F, Mangeon S, Ward DS et al. 2017. The Fire Modeling Intercomparison Project (FireMIP), phase 1: Experimental and analytical protocols with detailed model descriptions. Geoscientific Model Development 10: 1175–1197. [Google Scholar]
- Rambal S. 1993. The differential role of mechanisms for drought resistance in a Mediterranean evergreen shrub: a simulation approach. Plant, Cell & Environment 16: 35–44. [Google Scholar]
- Rao K, Williams AP, Diffenbaugh NS, Yebra M, Konings AG. 2022. Plant–water sensitivity regulates wildfire vulnerability. Nature Ecology & Evolution 6: 332–339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao K, Williams AP, Flefil JF, Konings AG. 2020. SAR‐enhanced mapping of live fuel moisture content. Remote Sensing of Environment 245: 111797. [Google Scholar]
- Reinhardt E, Crookston NL. 2003. The fire and fuels extension to the forest vegetation simulator. Gen. Tech. Rep. RMRS‐GTR‐116. Ogden, UT, USA: US Department of Agriculture, Forest Service, Rocky Mountain Research Station. [Google Scholar]
- Resco de Dios V. 2020. Plant–fire interactions: applying ecophysiology to wildfire management. Cham, Switzerland: Springer. [Google Scholar]
- Resco de Dios V, Arteaga C, Hedo J, Gil‐Pelegrín E, Voltas J. 2018. A trade‐off between embolism resistance and bark thickness in conifers: are drought and fire adaptations antagonistic? Plant Ecology & Diversity 11: 253–258. [Google Scholar]
- Resco de Dios V, Arteaga C, Peguero‐Pina JJ, Sancho‐Knapik D, Qin H, Zveushe OK, Sun W, Williams DG, Boer MM, Voltas J et al. 2020. Hydraulic and photosynthetic limitations prevail over root non‐structural carbohydrate reserves as drivers of resprouting in two Mediterranean oaks. Plant, Cell & Environment 43: 1944–1957. [DOI] [PubMed] [Google Scholar]
- Resco de Dios V, Hedo J, Cunill Camprubí À, Thapa P, Martínez del Castillo E, Martínez de Aragón J, Bonet JA, Balaguer‐Romano R, Díaz‐Sierra R, Yebra M et al. 2021. Climate change induced declines in fuel moisture may turn currently fire‐free Pyrenean mountain forests into fire‐prone ecosystems. Science of the Total Environment 797: 149104. [DOI] [PubMed] [Google Scholar]
- Riano D, Vaughan P, Chuvieco E, Zarco‐Tejada PJ, Ustin SL. 2005. Estimation of fuel moisture content by inversion of radiative transfer models to simulate equivalent water thickness and dry matter content: analysis at leaf and canopy level. IEEE Transactions on Geoscience and Remote Sensing 43: 819–826. [Google Scholar]
- Rishmawi K, Huang C, Zhan X. 2021. Monitoring key forest structure attributes across the conterminous United States by integrating GEDI LiDAR measurements and VIIRS data. Remote Sensing 13: 442. [Google Scholar]
- Rohatgi A. 2021. WebPlotDigitizer, Pacifica, CA, USA, v.4.5. [WWW document] URL https://automeris.io/WebPlotDigitizer [accessed 29 August 2021].
- Rothermel RC. 1972. A mathematical model for predicting fire spread in wildland fuels. Res. Pap. INT‐115. Ogden, UT, USA: US Department of Agriculture, Intermountain Forest and Range Experiment Station. [Google Scholar]
- Rothermel RC, Philpot CW. 1973. Predicting changes in chaparral flammability. Journal of Forestry 71: 640–643. [Google Scholar]
- Rouet‐Leduc J, Pe'er G, Moreira F, Bonn A, Helmer W, Shahsavan Zadeh SAA, Zizka A, van der Plas F. 2021. Effects of large herbivores on fire regimes and wildfire mitigation. Journal of Applied Ecology 58: 2690–2702. [Google Scholar]
- Ruckman EM, Schwinning S, Lyons KG. 2012. Effects of phenology at burn time on post‐fire recovery in an invasive C4 grass. Restoration Ecology 20: 756–763. [Google Scholar]
- Ruffault J, Limousin J‐M, Pimont F, Dupuy J‐L, De Càceres M, Cochard H, Mouillot F, Blackman CJ, Torres‐Ruiz JM, Parsons RA et al. 2022a. Plant hydraulic modelling of leaf and canopy fuel moisture content reveals increasing vulnerability of a Mediterranean forest to wildfires under extreme drought. New Phytologist 237: 1256–1269. [DOI] [PubMed] [Google Scholar]
- Ruffault J, Martin‐StPaul N, Pimont F, Dupuy J‐L. 2018. How well do meteorological drought indices predict live fuel moisture content (LFMC)? An assessment for wildfire research and operations in Mediterranean ecosystems. Agricultural and Forest Meteorology 262: 391–401. [Google Scholar]
- Ruffault J, Pimont F, Cochard H, Dupuy J‐L, Martin‐StPaul N. 2022b. SurEau‐Ecos v.2.0: a trait‐based plant hydraulics model for simulations of plant water status and drought‐induced mortality at the ecosystem level. Geoscientific Model Development 15: 5593–5626. [Google Scholar]
- Ruswick SK, O'Brien JJ, Aubrey DP. 2021. Carbon starvation is absent regardless of season of burn in Liquidambar styraciflua L. Forest Ecology and Management 479: 118588. [Google Scholar]
- Sánchez‐Costa E, Poyatos R, Sabaté S. 2015. Contrasting growth and water use strategies in four co‐occurring Mediterranean tree species revealed by concurrent measurements of sap flow and stem diameter variations. Agricultural and Forest Meteorology 207: 24–37. [Google Scholar]
- Sayer MAS, Tyree MC, Kuehler EA, Jackson JK, Dillaway DN. 2020. Physiological mechanisms of foliage recovery after spring or fall crown scorch in young longleaf pine (Pinus palustris Mill.). Forests 11: 208. [Google Scholar]
- Scarff FR, Lenz T, Richards AE, Zanne AE, Wright IJ, Westoby M. 2021. Effects of plant hydraulic traits on the flammability of live fine canopy fuels. Functional Ecology 35: 835–846. [Google Scholar]
- Scheller RM, Kretchun A, Hawbaker T, Henne PD. 2019. A landscape model of variable social‐ecological fire regimes. Ecological Modelling 401: 8593. [Google Scholar]
- Scott JH, Burgan RE. 2005. Standard fire behavior fuel models: a comprehensive set for use with Rothermel's surface fire spread model. Gen. Tech. Rep. RMRS‐GTR‐153. Fort Collins, CO, USA: US Department of Agriculture, Forest Service, Rocky Mountain Research Station. [Google Scholar]
- Serbin SP, Singh A, McNeil BE, Kingdon CC, Townsend PA. 2014. Spectroscopic determination of leaf morphological and biochemical traits for northern temperate and boreal tree species. Ecological Applications 24: 1651–1669. [DOI] [PubMed] [Google Scholar]
- Serbin SP, Townsend PA. 2020. Scaling functional traits from leaves to canopies. In: Cavender‐Bares J, Gamon JA, Townsend PA, eds. Remote sensing of plant biodiversity. Cham, Switzerland: Springer International, 43–82. [Google Scholar]
- Shuman JK, Balch JK, Barnes RT, Higuera PE, Roos CI, Schwilk DW, Stavros EN, Banerjee T, Bela MM, Bendix J et al. 2022. Reimagine fire science for the anthropocene. PNAS Nexus 1: pgac115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sieg CH, Linn RR, Pimont F, Hoffman CM, McMillin JD, Winterkamp J, Baggett LS. 2017. Fires following bark beetles: factors controlling severity and disturbance interactions in ponderosa pine. Fire Ecology 13: 1–23. [Google Scholar]
- Silva CVJ, Aragão LEOC, Barlow J, Espirito‐Santo F, Young PJ, Anderson LO, Berenguer E, Brasil I, Brown IF, Castro B et al. 2018. Drought‐induced Amazonian wildfires instigate a decadal‐scale disruption of forest carbon dynamics. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 373: 20180043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson KJ, Ripley BS, Christin P‐A, Belcher CM, Lehmann CER, Thomas GH, Osborne CP. 2016. Determinants of flammability in savanna grass species. Journal of Ecology 104: 138–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh A, Serbin SP, McNeil BE, Kingdon CC, Townsend PA. 2015. Imaging spectroscopy algorithms for mapping canopy foliar chemical and morphological traits and their uncertainties. Ecological Applications 25: 2180–2197. [DOI] [PubMed] [Google Scholar]
- Smith AMS, Talhelm AF, Johnson DM, Sparks AM, Kolden CA, Yedinak KM, Apostol KG, Tinkham WT, Abatzoglou JT, Lutz JA et al. 2017. Effects of fire radiative energy density dose on Pinus contorta and Larix occidentalis seedling physiology and mortality. International Journal of Wildland Fire 26: 82. [Google Scholar]
- Smith RE, Richardson DM. 1990. Comparative post‐fire water relations of selected reseeding and resprouting fynbos plants in the Jonkershoek Valley, Cape Province, South Africa. South African Journal of Botany 56: 683–694. [Google Scholar]
- Soil Survey Staff . 2022. Gridded National Soil Survey Geographic (gNATSGO) database for the conterminous United States . United States Department of Agriculture, Natural Resources Conservation Service. [WWW document] URL https://nrcs.app.box.com/v/soils [accessed 12 December 2022].
- Sparks AH, Forbes GA, Hijmans RJ, Garrett KA. 2011. A metamodeling framework for extending the application domain of process‐based ecological models. Ecosphere 2: art90. [Google Scholar]
- Steady WD, Partelli Feltrin R, Johnson DM, Sparks AM, Kolden CA, Talhelm AF, Lutz JA, Boschetti L, Hudak AT, Nelson AS et al. 2019. The survival of Pinus ponderosa saplings subjected to increasing levels of fire behavior and impacts on post‐fire growth. Fire 2: 23. [Google Scholar]
- Stocks BJ, Lawson BD, Alexander ME, Van Wagner CE, McAlpine RS, Lynham TJ, Dubé DE. 1989. The Canadian forest fire danger rating system: an overview. Forestry Chronicle 65: 450–457. [Google Scholar]
- Sturtevant B, Fortin M‐J. 2021. Understanding and modeling forest disturbance interactions at the landscape level. Frontiers in Ecology and Evolution 9: 653647. [Google Scholar]
- Sturtevant BR, Scheller RM, Miranda BR, Shinneman D, Syphard A. 2009. Simulating dynamic and mixed‐severity fire regimes: a process‐based fire extension for LANDIS‐II. Ecological Modelling 220: 3380–3393. [Google Scholar]
- Swann DEB, Bellingham PJ, Martin PH. 2022. Resilience of a tropical montane pine forest to fire and severe droughts. Journal of Ecology 111: 90–109. [Google Scholar]
- Tai X, Mackay DS, Anderegg WRL, Sperry JS, Brooks PD. 2017. Plant hydraulics improves and topography mediates prediction of aspen mortality in southwestern USA. New Phytologist 213: 113–127. [DOI] [PubMed] [Google Scholar]
- Talluto MV, Boulangeat I, Ameztegui A, Aubin I, Berteaux D, Butler A, Doyon F, Drever CR, Fortin M‐J, Franceschini T et al. 2016. Cross‐scale integration of knowledge for predicting species ranges: a metamodelling framework. Global Ecology and Biogeography 25: 238–249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanase MA, Panciera R, Lowell K, Aponte C, Tanase MA, Panciera R, Lowell K, Aponte C. 2015. Monitoring live fuel moisture in semiarid environments using L‐band radar data. International Journal of Wildland Fire 24: 560–572. [Google Scholar]
- Taylor K, Brummer T, Rew LJ, Lavin M, Maxwell BD. 2014. Bromus tectorum response to fire varies with climate conditions. Ecosystems 17: 960–973. [Google Scholar]
- Thornton PE, Thornton MM, Mayer BW, Wilhelmi N, Wei Y, Devarakonda R, Cook RB. 2014. Daymet: daily surface weather data on a 1‐km grid for North America, v.2. Oak Ridge, TN, USA: ORNL Distributed Active Archive Center. doi: 10.3334/ORNLDAAC/1219. [DOI] [Google Scholar]
- Trabaud L. 1991. Fire regimes and phytomass growth dynamics in a Quercus coccifera garrigue. Journal of Vegetation Science 2: 307–314. [Google Scholar]
- Van Wagner CE. 1973. Height of crown scorch in forest fires. Canadian Journal of Forest Research 3: 373–378. [Google Scholar]
- Van Wagner CE. 1977. Conditions for the start and spread of crown fire. Canadian Journal of Forest Research 7: 23–34. [Google Scholar]
- Varner JM, Hood SM, DougP A, Yedinak K, Hiers JK, Jolly WM, Shearman TM, McDaniel JK, O'Brien JJ, Rowell EM. 2021. Tree crown injury from wildland fires: causes, measurement and ecological and physiological consequences. New Phytologist 231: 1676–1685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varner JM, Putz FE, Mitchell RJ, Hiers JK, O'Brien JJ, Gordon DR. 2009. Post‐fire tree stress and growth following smoldering duff fires. Forest Ecology and Management 258: 2467–2474. [Google Scholar]
- Violle C, Navas M‐L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. 2007. Let the concept of trait be functional! Oikos 116: 882–892. [Google Scholar]
- Vos J, Evers JB, Buck‐Sorlin GH, Andrieu B, Chelle M, de Visser PHB. 2010. Functional–structural plant modelling: a new versatile tool in crop science. Journal of Experimental Botany 61: 2101–2115. [DOI] [PubMed] [Google Scholar]
- de Vries J. 2021. Using evolutionary functional–structural plant modelling to understand the effect of climate change on plant communities. in silico Plants 3: diab029. [Google Scholar]
- Wang L, Quan X, He B, Yebra M, Xing M, Liu X. 2019. Assessment of the dual polarimetric Sentinel‐1A data for forest fuel moisture content estimation. Remote Sensing 11: 1568. [Google Scholar]
- Weise DR, Wright CS. 2013. Wildland fire emissions, carbon and climate: characterizing wildland fuels. Forest Ecology and Management 317: 26–40. [Google Scholar]
- West AG, Nel JA, Bond WJ, Midgley JJ. 2016. Experimental evidence for heat plume‐induced cavitation and xylem deformation as a mechanism of rapid post‐fire tree mortality. New Phytologist 211: 828–838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolley T, Shaw DC, Ganio LM, Fitzgerald S. 2012. A review of logistic regression models used to predict post‐fire tree mortality of western North American conifers. International Journal of Wildland Fire 21: 1–35. [Google Scholar]
- Wright HA, Bailey AW. 1982. Fire ecology: United States and Southern Canada. New York, NY, USA: Wiley. [Google Scholar]
- Yashwanth BL, Shotorban B, Mahalingam S, Lautenberger CW, Weise D. 2016. A numerical investigation of the influence of radiation and moisture content on pyrolysis and ignition of a leaf‐like fuel element. Combustion and Flame 163: 301–316. [Google Scholar]
- Yebra M, Dennison PE, Chuvieco E, Riaño D, Zylstra P, Hunt ER, Danson FM, Qi Y, Jurdao S. 2013. A global review of remote sensing of live fuel moisture content for fire danger assessment: moving towards operational products. Remote Sensing of Environment 136: 455–468. [Google Scholar]
- Yebra M, Dijk AV, Cary G. 2018a. Evaluation of the feasibility and benefits of operational use of alternative satellite data in the Australian Flammability Monitoring System to ensure long‐term data continuity . Bushfire & Natural Hazards CRC. [WWW document] URL https://www.bnhcrc.com.au/sites/default/files/managed/downloads/evaluation_fms_feasibility.pdf [accessed 16 January 2022].
- Yebra M, Quan X, Riaño D, Rozas Larraondo P, van Dijk AIJM, Cary GJ. 2018b. A fuel moisture content and flammability monitoring methodology for continental Australia based on optical remote sensing. Remote Sensing of Environment 212: 260–272. [Google Scholar]
- Zhou X‐R, Schnepf A, Vanderborght J, Leitner D, Lacointe A, Vereecken H, Lobet G. 2020. CPlantBox, a whole‐plant modelling framework for the simulation of water‐ and carbon‐related processes. in silico Plants 2: diaa001. [Google Scholar]
- Zhu W‐Z, Xiang J‐S, Wang S‐G, Li M‐H. 2012. Resprouting ability and mobile carbohydrate reserves in an oak shrubland decline with increasing elevation on the eastern edge of the Qinghai–Tibet Plateau. Forest Ecology and Management 278: 118–126. [Google Scholar]
- Zylstra P. 2021. Linking fire behaviour and its ecological effects to plant traits, using FRaME in R. Methods in Ecology and Evolution 12: 1365–1378. [Google Scholar]