

Abstract
Errors in embryonic cardiac development are a leading cause of congenital heart defects (CHDs), including morphological abnormalities of the heart that are often detected after birth. In the past few decades, an emerging role for cilia in the pathogenesis of CHD has been identified, but this topic still largely remains an unexplored area. Mouse forward genetic screens and whole exome sequencing analysis of CHD patients have identified enrichment for de novo mutations in ciliary genes or non‐ciliary genes, which regulate cilia‐related pathways, linking cilia function to aberrant cardiac development. Key events in cardiac morphogenesis, including left–right asymmetric development of the heart, are dependent upon cilia function. Cilia dysfunction during left–right axis formation contributes to CHD as evidenced by the substantial proportion of heterotaxy patients displaying complex CHD. Cilia‐transduced signaling also regulates later events during heart development such as cardiac valve formation, outflow tract septation, ventricle development, and atrioventricular septa formation. In this review, we summarize the role of motile and non‐motile (primary cilia) in cardiac asymmetry establishment and later events during heart development.
Keywords: cardiac asymmetry, cardiac development, cilia, congenital heart defects, laterality
1. INTRODUCTION
The heart is one of the first organs to form during embryogenesis, functioning to ensure that an adequate supply of nutrients and oxygen moves throughout the developing embryo (Buckingham et al., 2005; C. M. J. Tan & Lewandowski, 2020). Heart development is a complex process that requires precise spatiotemporal gene expression to orchestrate the accurate formation of a four‐chambered organ with a connected circulatory system (Bruneau, 2013; Buijtendijk et al., 2020; Houyel & Meilhac, 2021). Errors during these precisely controlled processes can lead to structural malformations known as congenital heart defects (CHDs; Althali & Hentges, 2022; Morton et al., 2022; Samsa et al., 2015). CHDs are present in nearly 1% of live births, thereby making CHDs the most common birth defect (Althali & Hentges, 2022; van der Linde et al., 2011). Several genetic and non‐genetic factors contribute to the development of CHD (S. S. Patel & Burns, 2013; Peng et al., 2019; Shiaulou Yuan et al., 2013). Understanding the etiology of CHD can improve diagnostic tools and help affected families to understand the severity of the disease (Dodge‐Khatami, 2016; Houyel & Meilhac, 2021; Pierpont et al., 2018). This knowledge is valuable because recent advances in CHD diagnosis and treatment have substantially increased the survival of CHD patients into adulthood (Mandalenakis et al., 2020).
In the past few decades, several studies have highlighted the importance of hair‐like cellular structures called cilia during cardiac development. Defects in cilia structure or function contribute to a variety of CHDs resulting from errors in cardiac asymmetry establishment, morphological arrangement, and valve development (Clement et al., 2009; Y. Li et al., 2015; X. Li et al., 2020; Slough et al., 2008; Toomer et al., 2019; Watanabe et al., 2003; Willaredt et al., 2012). In this review, we will summarize the current knowledge revealing how motile and non‐motile cilia at the left–right (L‐R) organizer (LRO) influence cardiac asymmetric morphogenesis and discuss the role of cardiac primary cilia in heart development and CHDs.
2. THE ARCHITECTURE OF CILIA
Cilia are evolutionarily conserved, unique, sensory antennae‐like structures of several micrometers in length that protrude from the apical surfaces of most vertebrate cells (Drummond, 2012; Long & Huang, 2019; Malicki & Johnson, 2017; Narasimhan & Roy, 2015; Satir & Christensen, 2007; Tasouri & Tucker, 2011; Willaredt et al., 2012). Cilia sense external environmental queues including light, low‐molecular‐weight chemicals, proteins, and mechanical stimuli and transfer the biochemical information to the cell to regulate cellular signaling pathways. Cilia also secrete ectosomes that signal to recipient cells to regulate cell– communication (Ferreira et al., 2019; Koefoed et al., 2014; Long & Huang, 2019; Malicki & Johnson, 2017; Toomer et al., 2019). Cilia function is critical for embryonic patterning, organogenesis, and adult tissue/organ homeostasis (Kim et al., 2023). During the mid‐20th century, the detailed structure of cilia was revealed using transmission electron microscopy (Fawcett & Porter, 1954; Gibbons, 1961; Rieder et al., 1979). Structurally, cilia have a nine parallel doublet microtubule‐based cytoskeleton core called the axoneme, surrounded by a ciliary membrane that is continuous with the cell membrane (Fawcett & Porter, 1954; Figure 1a). The ciliary axoneme is anchored at the base by the basal body, which is a modified centriole. The basal body is tethered to the plasma membrane via distinct appendages called transition fibers, establishing a selective gating system that controls protein trafficking between the cytoplasm and cilia (May et al., 2021; Miller et al., 2013; for more detail about cilia structure, see reviews Mill et al., 2023; Mirvis et al., 2018).
FIGURE 1.
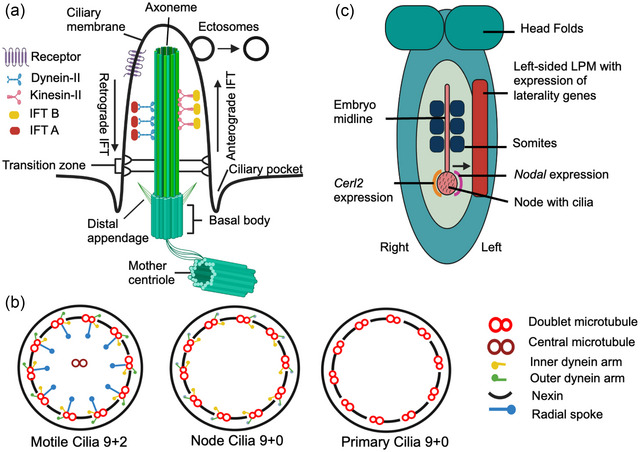
Mouse embryonic node, cilia structure, and doublet microtubule arrangements in motile and primary cilia. (a) Structurally, the cilium consists of a basal body, an axoneme, and dynein or kinesin proteins transporting the IFT complex proteins along the axoneme. (b) The axoneme microtubule arrangement. Motile cilium showing the 9 + 2 arrangement of microtubules along with inner dynein arms, outer dynein arms and radial spokes. The node motile cilia lack the central microtubule pair (9 + 0) and radial spokes. The primary cilia lack the motility components including the central microtubule pair (9 + 0). (c) Mouse embryo showing the cilia present at the embryonic node. The anticlockwise movement of motile cilia at the embryonic node generates leftward fluid flow and activates laterality gene expression and signaling on the left of the embryo in the lateral plate mesoderm. Figure created with Biorender.com and Microsoft Paints.
Depending upon their motility and axonemal architecture, cilia can be classified as either motile or non‐motile (primary cilia; Ishikawa, 2017; Kiesel et al., 2020; Sun et al., 2019). Motile cilia are found at the embryonic node (Alten et al., 2012), brain ventricles (Olstad et al., 2019), oviducts (Shuiqiao Yuan et al., 2021), and airway epithelium (Shah et al., 2009). The motile cilia axoneme usually has a 9 + 2 arrangement of microtubules, equipped with dynein arms and radial spokes, which generate beats via dynein arm‐driven adenosine triphosphate (ATP) hydrolysis, to produce fluid movement or propel gametes (Figure 1b; Hale & Sadoshima, 2022; Hjeij et al., 2014; Koefoed et al., 2014; Reiter & Leroux, 2017). However, node motile cilia are an exception because they lack the central microtubule pair and radial spokes (9 + 0 with dynein arms) but can generate rotational movement that results in fluid flow (Figure 1b; Huang et al., 2009; Ishikawa, 2017; Shinohara et al., 2015). Non‐motile or primary cilia have a 9 + 0 microtubule arrangement, missing the central microtubule pair, dynein arms, and radial spokes (Sun et al., 2019; Figure 1b). A single primary cilium protrudes from the surface of most vertebrate cell types. Primary cilia were previously considered as vestigial evolutionary remnants with no function (Toomer et al., 2019). However, primary cilia have been discovered to be dynamic structures that function as mechanosensors and signaling hubs that are fundamental to the development and maintenance of many tissues and organs (Fulmer et al., 2020; Miller et al., 2013; Tasouri & Tucker, 2011).
Recent findings have identified that cilia possess local protein translation machinery to maintain ultrastructure and function (Hao et al., 2021). However, for the transport of cytoplasmic proteins across the axoneme, cilia rely on the intraflagellar transport system (IFT). The IFT is a specialized evolutionarily conserved bidirectional microtubule motor‐based transport system, required for the growth and maintenance of both motile and primary cilia (Fry et al., 2014; Rosenbaum & Witman, 2002). The IFT complexes (IFT‐A retrograde transport and IFT‐B anterograde transport), along with kinesins, dynein motor proteins, and adapter molecules, ferry proteins and other cargo across the axoneme and in and out of the cilia (Koefoed et al., 2014; May et al., 2021; K. Patel & Smith, 2023; Reiter & Leroux, 2017; Tasouri & Tucker, 2011; Toomer et al., 2019; Willaredt et al., 2012; Figure 1). Mutations in the IFT complex affect the entire cilia transport system. Mutations in IFT‐B members result in short or absent cilia, while with IFT‐A, defects typically result in distorted cilia with a distended tip due to the accumulation of stranded IFT cargo (Pigino et al., 2009; Y. Zhang et al., 2018; for more detail about IFT system, see reviews Lechtreck, 2015; Taschner & Lorentzen, 2016).
Cilia possess complex repositories of channels and membrane‐spanning receptors that transduce mechanical, electrical, and chemical signals from the extracellular environment to the cytoplasm in a tissue‐specific and time‐dependent context (Christensen et al., 2007; Nishimura et al., 2018; Pala et al., 2017). The co‐localization of multiple channels and signaling components within the cilia raises the possibility that cilia coordinate the cross‐talk between signaling pathways (Nachury, 2014). These pathways include G protein‐coupled receptors (GPCRs; Brewer et al., 2023; Mykytyn & Askwith, 2017), receptor tyrosine kinase (Christensen et al., 2017), hedgehog (Hh; Bangs & Anderson, 2017; Tasouri & Tucker, 2011), Wingless‐related integration site (Wnt) (Kyun et al., 2020; Wallingford & Mitchell, 2011), platelet‐derived growth factor receptor (PDGFR; Schmid et al., 2018; Schneider et al., 2005), transforming growth factor‐beta (TGF‐β)/bone morphogenic protein (BMP; Álvarez‐Satta et al., 2021; Clement et al., 2013; Gencer et al., 2017), planar cell polarity (PCP; Borovina et al., 2010; Park et al., 2008; Ross et al., 2005; Song et al., 2010), and extracellular matrix (ECM; Battini et al., 2006; McGlashan et al., 2006). Of these cilia‐regulated pathways, several are known to be important for cardiac development (Gabriel et al., 2021). Defects in cilia structure and aberrant signaling via the cilium result in a variety of cilia‐associated diseases, collectively known as ciliopathies (Baker & Beales, 2009; Hale & Sadoshima, 2022; Long & Huang, 2019; Reiter & Leroux, 2017; Toomer et al., 2019). There are nearly 200 distinct ciliopathies (Rao Damerla et al., 2014). First‐order ciliopathies are caused by ciliary gene disruption, while second‐order ciliopathies result from defects in non‐ciliary genes required for cilia function (Reiter & Leroux, 2017; for more detail regarding first‐order and second‐order cilia genes and associated ciliopathies, see Vasquez et al., 2021).
3. LRO: THE BODY'S SYMMETRY BREAKER
Vertebrate visceral organs and their associated vasculature are characteristically arranged asymmetrically inside the body in an orientation known as situs solitus (Fujinaga, 1997; Nöthe‐Menchen et al., 2019; Sampaio et al., 2014; Shapiro et al., 2014; Stevens et al., 2010; Watanabe et al., 2003). Organ asymmetry defects and associated diseases can cause life‐threatening complications (Saba et al., 2022). Failure to establish the characteristic internal organ asymmetry can lead to laterality defects such as heterotaxy (random orientation of organs) or situs inversus (mirror image reversal of organs; Chen et al., 2019; Eitler et al., 2022; Saba et al., 2022). In several vertebrates, bilateral symmetry is broken at a transient embryonic structure known as the LRO (Blum et al., 2007). The LRO name varies between species; in mice, it is called the node (Nonaka et al., 1998, 2002; Figure 1c), in frog, it is the gastrocoel roof plate (Sáenz‐Ponce et al., 2012; Schweickert et al., 2007), while in zebrafish, it is the Kupffer's vesicle (KV; Kramer‐Zucker et al., 2005). The motile cilia at embryonic LRO generate extracellular fluid flow (nodal flow), causing the expression of asymmetric genes in the lateral plate mesoderm (LPM) and activation of downstream signaling effectors, which co‐ordinates the asymmetric morphogenesis of the internal organs (Hirokawa et al., 2012; Kawasumi et al., 2011; Larkins et al., 2012; Watanabe et al., 2003).
The mouse embryonic node is a tear‐shaped, mesodermal‐derived, epithelial pit at the posterior end of the notochord containing monociliated cells (Hirokawa et al., 2009; Sulik et al., 1994; Yamanaka et al., 2007). The cilia on the node cells are tilted posteriorly, which produces leftward flow when the cilia are motile (Nonaka et al., 2005). Defects in PCP pathways, which regulate the tilt direction of node motile cilia, result in laterality defects (Mahaffey et al., 2013; Song et al., 2010). The motile cilia at the embryonic node beat radially in an anticlockwise direction to perpetuate a leftward flow, while non‐motile or sensory cilia, which extend from the crown cells surrounding the pit, sense the flow (McGrath et al., 2003; Yoshiba et al., 2012). This flow sensing triggers a conserved molecular cascade by initiating laterality gene expression in the LPM, which subsequently culminates in the correct positioning of the visceral organs (McGrath et al., 2003; Nonaka et al., 2002; Sampaio et al., 2014). In mouse, a total of nearly 200–300 motile cilia at the embryonic node generate nodal flow; however, as few as two motile cilia are enough to break embryonic internal bilateral symmetry (Shinohara et al., 2012).
At the molecular level, embryonic leftward fluid flow activates the nodal signaling cascade (Brennan et al., 2002). Nodal, a TGF‐β family signaling molecule, is asymmetrically expressed at the node and activates left–right patterning gene expression in the LPM (Figure 1c; Collignon et al., 1996; Norris & Robertson, 1999; Saijoh et al., 2003; Sakuma et al., 2002). Nodal asymmetrical expression is restricted to the left side of the node by the Nodal antagonist Cerl2. Cerl2 is initially expressed bilaterality around the node at the early headfold stage; however, leftward nodal flow initiates Cerl2 mRNA decay on the left side, which is further enhanced by a Wnt3–Cerl2 interlinked feedback loop (Belo et al., 2017; Inácio et al., 2013; Marques et al., 2004; Nakamura et al., 2012; Schweickert et al., 2010). Nodal activity on the left side of the node triggers the initiation of the laterality gene expression program (Brennan et al., 2002; Norris et al., 2002). Cells that receive nodal signaling adopt a left‐sided cell fate (Yamamoto et al., 2003). Nodal expression is further maintained by the downstream transcription factors lefty and Pitx2 (Adachi et al., 1999; Logan et al., 1998; Oki et al., 2009; Piedra et al., 1998; Yoshioka et al., 1998). Once Nodal expression has ceased, asymmetric Pitx2 expression in the heart, foregut, cardinal vein, and umbilical vein is maintained by the transcription factor Nkx2‐5 (Shiratori et al., 2001). These findings suggest that node cilia‐induced laterality gene expression and activation of downstream signaling play a decisive role in asymmetrical development and patterning of the internal organs including the heart.
4. PRESENCE OF CILIA IN CARDIAC DEVELOPMENT
Cardiac organogenesis in vertebrates is a highly complex, tightly coordinated series of gene expression and signaling events that require the migration of different types of progenitor cells to form the heart (Koefoed et al., 2014; Münsterberg & Yue, 2008). The earliest known cardiac progenitor cells express the transcription factor Mesp1 and are derived from mesodermal cells present in the anterior primitive streak region (Kitajima et al., 2000; Saga et al., 1996, 1999). These cardiac progenitor cells from the anterior primitive streak region migrate under the embryonic headfolds and form a bilaterally symmetrical crescent‐shaped heart field at the midline (Figure 2a; Ivanovitch et al., 2017; Kuhn & Wu, 2010). In the mouse, the heart starts to form after E7.5, when cells from the two crescent‐shaped heart fields migrate toward the ventral midline and converge to form a linear heart tube (Figure 2b; Kelly, 2012; Willaredt et al., 2012; M. Wu, 2018). The linear heart tube undergoes dextral looping to form an asymmetric four‐chambered heart (Figure 2c). The cardiac looping process is controlled by several factors including nodal signaling at the LRO, which specifies left identity in myocardial precursor cells (Desgrange et al., 2020).
FIGURE 2.
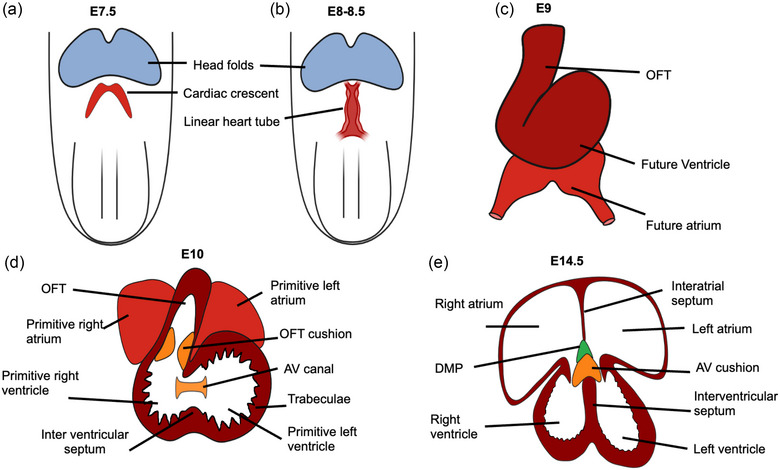
The stages of mouse heart development. (a) Mouse heart development at Embryonic Day 7.5, when cardiac progenitor cells migrate toward the midline to form a crescent‐shaped heart field. (b) The crescent‐shaped heart field cells converge at the midline to form a linear heart tube. (c) The linear heart tube undergoes dextral looping to shape the future four‐chambered heart. (d) The ventricular wall of the looped heart undergoes trabeculation, and endocardial cushion cells protrude into the heart tube to start to form the cardiac valves. (e) The four‐chambered heart is formed around E14.5 with the presence of atrial and ventricular septa. AV, atrioventricular; DMP, dorsal mesenchymal protrusion; OFT, outflow tract. Figure created with Biorender.com and Microsoft Paints.
Once the linear heart tube loops, the endocardial cushions (cardiac valve precursor structures located at the atrioventricular [AV] junction and within the outflow tract [OFT]) start to swell. These precursor cells then undergo epithelial‐to‐mesenchymal transition (EMT) and secrete ECM resulting in the formation of the heart valves (O'Donnell & Yutzey, 2020; Person et al., 2005; Figure 2d). Additionally, following looping, the heart wall forms a network of luminal projections known as trabeculae. The presence of cardiac trabeculae increases cardiac output and facilitates nutrient and oxygen exchange in the embryonic myocardium prior to the establishment of the coronary vasculature. After the establishment of coronary vasculature, the trabeculae undergo extensive remodeling and coalesce together with the cardiac wall to form a thick muscular layer (Lai et al., 2010; Qu et al., 2022; Samsa et al., 2013, 2015; M. Wu, 2018; Figure 2e).
Two extra cardiac cell sources, the cardiac neural crest cells (Schleiffarth et al., 2007; Stefanovic et al., 2021) and epicardial cells (J. Li et al., 2017; Ridge et al., 2017) also contribute to the development of the heart. The cardiac neural crest cells migrate from the hindbrain region including rhombomeres 6‐8 and contribute to the development of the OFT, cardiac valves, cardiac septa, and arteries of the heart (George et al., 2020; Hutson & Kirby, 2007; Schleiffarth et al., 2007; Stefanovic et al., 2021; Yan et al., 2021). In the mouse, around E10.5, the cardiac OFT then undergoes septation to form the aorta and pulmonary artery, which are required to carry oxygenated and deoxygenated blood in and out of the heart (Schleiffarth et al., 2007; Stefanovic et al., 2021). Epicardial cells migrate from the pro‐epicardial organ (located at the venous pole of the heart) and attach to the surface of heart around E9.5 in the mouse. These cells underdo EMT and contribute to different cell types in the heart, facilitating establishment of the coronary circulation (J. Li et al., 2017; Ridge et al., 2017; see fig. 3 in the review from Clowes et al., 2014). The process of four‐chambered heart formation is completed around E14.5 in mouse embryos (Savolainen et al., 2009).
Primary cilia have been reported to appear throughout the embryonic heart development (Slough et al., 2008; Willaredt et al., 2012). Ultrastructural analysis of cardiac cilia using transmission electron microscopy shows the presence of cilia with nine pairs of microtubules, lacking the central microtubule pair and cilia motility machinery, confirming the presence of primary cilia (Slough et al., 2008; Toomer et al., 2019; Willaredt et al., 2012). Slough et al. (2008) illustrated the presence of primary cilia on mouse embryonic heart cells as early as E9.5 of development (Slough et al., 2008). During cardiac development, cilia are found on all cell types in the heart including cardiomyocytes, endocardial cells, epicardial cells, and cardiac cushion cells (Gerhardt et al., 2013; Myklebust et al., 1977; Slough et al., 2008). However, using the cilia cytoskeleton marker acetylated α‐tubulin and the basal body marker pericentrin, Gerhardt et al. (2013) observed a lack of cilia on ventricular septa and ventricular cells close to the septum between E10.5 and E12.5 of heart development. Cilia on cardiac cushion mesenchymal cells are present in a ciliary pocket of varying depth with a random orientation. Endocardial cell primary cilia are orientated toward the lumen of the inflow and OFTs, while atrial and ventricular primary cilia are oriented toward the blood‐filled cardiac chambers, and epicardium primary cilia are oriented toward the pericardial space (Diguet et al., 2015; Slough et al., 2008; Willaredt et al., 2012). The presence and length of cilia in the mitral valves correlate with the type of ECM produced during heart development. For example, in early development, the valves have abundant proteoglycans but little collagen and express cilia on nearly all cells. As development progresses, by Postnatal Day 0, there is an increase in collagen expression in the valves concomitant with a reduction in the number and length of primary cilia. These cilia are mainly localized to regions with low collagen expression (Toomer et al., 2019).
A growing body of work has also identified the importance of primary cilia in cardiac development (Burnicka‐Turek et al., 2016; Fulmer et al., 2019; Gerhardt et al., 2013; Hartill et al., 2018; Y. Li et al., 2015; Slough et al., 2008; Willaredt et al., 2012). Using a forward genetic screen in the mouse, Y. Li et al. (2015) identified 61 genes, in which pathogenic mutations could produce echocardiographically identifiable CHDs. Of these genes, 35 genes encoded either motile or primary cilia proteins, with many of the remaining genes being involved in cilia‐related signaling (16 genes) or vesicular trafficking (10 genes), which are required for cilia formation and function (Y. Li et al., 2015).
5. LRO CILIA AND CARDIAC ASYMMETRY
Cilia function at the LRO is critical for the establishment of cardiac left–right (L‐R) asymmetry. Both motile and sensory cilia (primary cilia) at the mouse embryonic node are indispensable for the establishment of left–right asymmetry (McGrath et al., 2003; Nonaka et al., 1998; Yoshiba et al., 2012), which is important for the early stages of heart morphogenesis and appropriate connections to the vasculature (Koefoed et al., 2014). In mice, the first morphological sign of left–right asymmetry is the dynamic right‐sided looping of the primitive heart tube, followed by embryonic turning that converts the embryo from a lordotic to a fetal position (Chatterjee et al., 2007; Honda et al., 2020). Heart asymmetry is critical for efficient oxygenation of the blood and establishment of the systemic and pulmonary circulation (Francis et al., 2012). Defects in laterality establishment significantly increase the risk of CHD. Nearly 3% of all CHD results from heterotaxy, and complex CHD is often associated with heterotaxy (57% in heterotaxy patients vs. 1% in the general population; Agarwal et al., 2021; Burnicka‐Turek et al., 2016; Djenoune et al., 2022; Merklin & Varano, 1963; Slough et al., 2008). Here, we will discuss some of the known LRO‐related cilia components that regulate cardiac asymmetry establishment.
5.1. Nodal cilia motility component defects and cardiac asymmetry
The primary cilia dyskinesias (PCDs; OMIM 244400) are a set of diseases associated with motile cilia dysfunction (Y. Li et al., 2016; Reiter & Leroux, 2017). PCD is often associated with situs inversus, heterotaxy, and complex CHD (Best et al., 2019; Francis et al., 2012; Harrison et al., 2016). Y. Li et al. (2015) reported that most PCD‐causing motile cilia genes are also known to cause CHD. The dysfunction of the motile cilia outer dynein arms (ODA), which generate motor force for ciliary beating (Zimmermann et al., 2023), is the major cause of PCD (Wallmeier et al., 2016). ODA genes including Dnaic1 (Francis et al., 2012) (see Table 1 for the list of cilia genes mentioned in this review), DNAH5 (Ibañez‐Tallon et al., 2002; Nöthe‐Menchen et al., 2019; S. Y. Tan et al., 2007), DNAH11 (S. Liu et al., 2019; Xia et al., 2021), or ODA‐docking genes like ARMC4 (Hjeij et al., 2013; Onoufriadis et al., 2014), TTC25 (Wallmeier et al., 2016), Ccdc39 (Solomon et al., 2017), DNAH10 (C. Liu et al., 2018), and MNS1 (Ta‐Shma et al., 2018) are frequently associated with motile cilia dysfunction, left–right patterning defects,, and cardiac asymmetry defects. Hjeij et al. (2014) have identified a mutation in the ODA docking gene CCDC151 in PCD individuals with dextrocardia. CCDC151 mutant cilia fail to assemble with the ODA component DNAH5 and the ODA‐docking complex. This altered complex formation results in cilia structural changes with complete loss of ODA and impaired ciliary beating, leading to a spectrum of situs defects associated with complex heart defects (Hjeij et al., 2014). Mutations in inner dynein arm (IDA) genes including DNAH6 (Y. Li et al., 2016) or DNAH7 (Y. Z. Zhang et al., 2002) impair the formation of the IDA structure and central microtubule pair, which are required for cilia motility. Knockdown of dnah6 in zebrafish embryos causes a constellation of heterotaxy phenotypes with reduced cilia length at the KV, disruption of left‐sided southpaw (zebrafish Nodal homolog) expression, abnormal body curvature, and altered orientation of heart and gut looping (Y. Li et al., 2016). Likewise, disruption of the coiled‐coil domain containing‐40 (Ccdc40) gene, which regulates the assembly of the IDA and the dynein regulatory complexes (Becker‐Heck et al., 2011), drastically reduces the cilia length at the mouse node, affecting cilia motility and compromising nodal flow, resulting in situs inversus or heterotaxia with cardiac looping defects (Becker‐Heck et al., 2011; Sugrue & Zohn, 2017).
TABLE 1.
List of cilia structure, function, and signaling genes (mentioned in the article), in alphabetical order, that cause cardiac defects when mutated.
| Gene symbol | Gene name | First/second‐order cilia gene | Mutant phenotype | Reference |
|---|---|---|---|---|
| ARMC4 | Armadillo repeat containing 4 | First order | Laterality defects, cardiac looping defects | (Hjeij et al., 2013; Onoufriadis et al., 2014) |
| CCDC 40 | Coiled‐coil domain containing 40 | First order | Laterality defects, cardiac looping defects | (Becker‐Heck et al., 2011; Sugrue & Zohn, 2017) |
| CCDC151 | Coiled‐coil domain containing 151 | First order | Laterality defects, dextrocardia, ventricular septal defects | (Hjeij et al., 2014) |
| CCDC39 | Coiled‐coil domain containing 39 | First order | Laterality defects, cardiac looping defects | (Solomon et al., 2017) |
| CRELD1 | Cysteine Rich With EGF Like Domains 1 | Second order | Atrioventricular septal defects (AVSDs), valve defects | (Beckert et al., 2021, Burnicka‐Turek et al., 2016) |
| DHH | Desert hedgehog | Second order | Mitral valve prolapse (MVP) | (Fulmer et al., 2020) |
| DNAAF1 | Dynein axonemal assembly factor 1 | First order | Laterality defects, cardiac looping defects | (Hartill et al., 2018) |
| DNAAF3 | Dynein axonemal assembly factor 3 | First order | Laterality defects, cardiac looping defects | (Mitchison et al., 2012) |
| DNAH5 | Dynein axonemal heavy chain 5 | First order | Laterality defects, cardiac looping defects | (Nöthe‐Menchen et al., 2019) |
| DNAH6 | Dynein axonemal heavy chain 6 | First order | Laterality defects, cardiac looping defects | (Y. Li et al., 2016) |
| DNAH7 | Dynein axonemal heavy chain 7 | First order | Laterality defects, cardiac looping defects | (Y. J. Zhang et al., 2002) |
| DNAH 10 | Dynein axonemal heavy chain 10 | First order | Laterality defects, cardiac looping defects | (C. Liu et al., 2018) |
| DNAH11 | Dynein axonemal heavy chain 11 | First order | Laterality defects, AVSDs | (Bartoloni et al., 2002; Burnicka‐Turek et al., 2016; Dougherty et al., 2016; S. Liu et al., 2019; Xia et al., 2021) |
| DNAIC1 | Dynein Axonemal Intermediate Chain 1 | First order | Laterality defects, cardiac looping defects | (Francis et al., 2012) |
| DYX1C1(DNAAF4) | Dyslexia Susceptibility 1 Candidate 1 | First order | Laterality defects, cardiac looping defects | (Tarkar et al., 2013) |
| DZIP1 | DAZ interacting zinc finger protein 1 | First order | MVP | (Toomer et al., 2019) |
| EXOC5 | Exocyst complex component 5 | First order | Bicuspid aortic valve disease and aortic stenosis | (Fulmer et al., 2019) |
| FTM (Rpgrip1l) | Fantom | First order | Laterality defects, cardiac looping defects, AVSD | (Gerhardt et al., 2013; Vierkotten et al., 2007) |
| GPR22 | G protein‐coupled receptor (GPCR) 22 | First order | Laterality defects, cardiac looping defects, cardiac edema | (Verleyen et al., 2014) |
| GRK5 | GPCR kinase 5 | Second order | Laterality defects, cardiac looping defects, valve development defects | (Burkhalter et al., 2013; Casar Tena et al., 2015) |
| IFT20 | Intraflagellar transport 20 | First order | Proepicardial organ and myocardial tissue size defects | (Peralta et al., 2020) |
| IFT46 | Intraflagellar transport 46 | First order | Laterality defects, cardiac looping defects | (Lee et al., 2015) |
| IFT54 (TRAF3IP1) | Intraflagellar transport 54(TRAF3 interacting protein 1) | First order | Proepicardial organ and myocardial tissue size defects | (Peralta et al., 2020) |
| IFT57 | Intraflagellar transport 57 | First order | Laterality defects, cardiac looping defects | (Houde et al., 2006) |
| IFT74 | Intraflagellar transport 74 | First order | Laterality defects, cardiac looping defects, AVSD, hypoplastic left heart | (Bakey et al., 2023) |
| IFT88 | Intraflagellar transport 88 | First order | Laterality defects, cardiac looping defects, outflow tract defects, ventricular trabeculation defects, cardiac cushion EMT defects, valves defects, proepicardial organ and myocardial tissue size defects | (Burns et al., 2019; Clement et al., 2009; Murcia et al., 2000; Peralta et al., 2020; Toomer et al., 2019; Willaredt et al., 2012) |
| IFT172 | Intraflagellar transport 172 | First order | Laterality defects, cardiac looping defects | (Gorivodsky et al., 2009) |
| INVS | Inversin | First order | Laterality defects, cardiac looping defects | (Lowe et al., 1996, Okada et al., 1999; Watanabe et al., 2003; Yokoyama et al., 1993) |
| KIF3A | kinesin family member 3A | First order | Laterality defects, cardiac looping defects | (Takeda et al., 1999) |
| KIF3B | kinesin family member 3B | First order | Laterality defects, cardiac looping defects | (Nonaka et al., 1998) |
| MEGF8 | Multiple Epidermal Growth Factor‐like Domains 8 | Second order | Laterality defects, cardiac looping defects | (Y. Li et al., 2015; Z. Zhang et al., 2009) |
| MKS1 | MKS Transition Zone Complex Subunit 1 | First order | AVSDs | (Burnicka‐Turek et al., 2016; Cui et al., 2011) |
| MNS1 | Meiosis specific nuclear structural 1 | First order | Laterality defects | (Ta‐Shma et al., 2018) |
| PDGFR‐α | Platelet‐derived growth factor receptor‐alpha | Second order | MVP | (Moore et al., 2021) |
| PKD1 | Polycystin 1, transient receptor potential channel interacting | First order | AVSDs, myocardial wall thinning, double‐outlet right ventricle, cardiac valves defects | (Boulter et al., 2001; Juan et al., 2023) |
| PKD1L1 | Polycystin 1 like 1, transient receptor potential channel interacting | First order | Laterality defects, cardiac looping defects, cardiac valve defects | (Field et al., 2011; Juan et al., 2023) |
| PKD2 | Polycystin 2, transient receptor potential cation channel | First order | Laterality defects, cardiac looping defects, cardiac septation defects, cardiac valve defects | (Juan et al., 2023; Pennekamp et al., 2002; G. Wu et al., 2000) |
| TBC1D32 | TBC1 domain family member 32 | First order | Laterality defects, cardiac looping defects | (Y. Li et al., 2015) |
| TCTN2 | Tectonic family member 2 | First order | Ventricular septal defects | (Sang et al., 2011) |
| TTC25 | Tetratricopeptide repeat domain 25 | First order | Laterality defects, cardiac looping defects | (Wallmeier et al., 2016) |
Note: First‐order genes are those that encode proteins required to form the cilia, cilia motility, or for cilia cargo transport. Second‐order genes are those that participate in signaling at the cilium or have undefined roles in the cilium.
Several other cilia genes like DNAAF1 (Hartill et al., 2018), DNAAF3 (Mitchison et al., 2012), and DYX1C1(DNAAF4) (Tarkar et al., 2013), which are required for dynein heavy chain assembly and cilia motility, also display PCD with heterotaxy and complex CHD when mutated. Mutations in the central microtubule pair and radial spoke genes are often associated with PCD but not heterotaxy (Best et al., 2019), as node motile cilia lack these structures (Figure 1b).
5.2. Nodal cilia non‐motility component defects and cardiac asymmetry
The non‐motile structural components of cilia such as centrosomal proteins, IFTs, and transition zone components also affect cardiac left–right asymmetry establishment (Gorivodsky et al., 2009; Houde et al., 2006; Lee et al., 2015; Murcia et al., 2000; Shylo et al., 2020; C. Wu et al., 2014). Takeda et al. (1999) and Nonaka et al. (1998) showed that cilia at the node are indispensable for breaking bilateral symmetry by deleting the Kinesin superfamily proteins Kif3a (Takeda et al., 1999) and Kif3b (Nonaka et al., 1998), respectively, resulting in loss of node cilia, symmetry defects, and randomization of cardiac looping (Marszalek et al., 1999; McGrath et al., 2003; Nonaka et al., 1998; Takeda et al., 1999). The restoration of Kif3a expression only in crown cells (which possess sensory cilia) in Kif3a mutant embryos allows a response to artificially induced fluid flow and rescues Nodal and Pitx2c expression in the LPM. However, the rescue of cardiac looping defects was not analyzed (Yoshiba et al., 2012). Mice embryos with a faulty IFT system, such as mutants for the IFT complex B gene Ift46, die around E10–E10.5, with neural tube and heart defects. Pericardial edema and cardiac looping defects were observed, as well as a lack of cilia, at the embryonic node. The absence of nodal flow in these mutants results in bilateral Lefty1 expression in the LPM and cardiac looping defects (Lee et al., 2015). Other IFT genes like Ift57 (Houde et al., 2006), Ift88 (Murcia et al., 2000), Ift172 (Gorivodsky et al., 2009), and Ift74 (Bakey et al., 2023) also showed defects in node cilia structure and function with randomized heart looping in homozygous mutants.
Several studies where cilia genes were genetically manipulated revealed normal but immobile node cilia with abnormal left–right cardiac asymmetry (Slough et al., 2008). Mice mutant for the protein Inversin, which localizes at the proximal end of the cilium near the basal body, show no obvious defects in node monocilia but have disrupted nodal flow and right‐sided Nodal expression in the LPM. Inversin mutant mice consistently display situs inversus with heart looping defects (Lowe et al., 1996; Okada et al., 1999; Watanabe et al., 2003; Yokoyama et al., 1993).
5.3. Defects in genes associated with cilia signaling function and cardiac asymmetry
Several genes that regulate signaling pathways that function at the cilium have been identified as having a role in the establishment of cardiac asymmetry (Y. Li et al., 2015). The fluid flow at the node induces movement of the primary cilia on crown cells, provoking an increase in calcium concentration in the crown cells. This change in calcium concentration stimulates the nodal signaling cascade on the left side of the node, which is translated into activation of laterality gene expression (McGrath et al., 2003; Pennekamp et al., 2002). Disruption of the calcium gradient across the node results in ambiguous expression of Nodal pathway genes, generating symmetry in L‐R development (Takao et al., 2013; Shiaulou Yuan et al., 2015), and cardiac looping defects (Pennekamp et al., 2002). The calcium‐permeable channel Polycystin‐2 (Pkd2) and its binding partner Pkd1l1 form a complex, which localizes to all node cilia (Field et al., 2011; Kamura et al., 2011; Yoshiba & Hamada, 2014). This complex has mechanosensing properties and plays a critical role in calcium gradient establishment and activation of nodal signaling in the LPM (Field et al., 2011; Kamura et al., 2011; McGrath et al., 2003; Pennekamp et al., 2002; Schottenfeld et al., 2007). Pkd2 (Kamura et al., 2011; Pennekamp et al., 2002) or Pkd1l1 (Field et al., 2011) mutant embryos show laterality defects with abnormal heart looping, and altered embryonic turning, but no structural or functional defects in node motile cilia. The specific restoration of Pkd2 in perinodal crown cells rescues the symmetry defects in Pkd2 mutants (Yoshiba et al., 2012). The application of mechanical forces to immotile cilia at the LRO triggers intraciliary calcium ion transients (Djenoune et al., 2023; Katoh et al., 2023), confirming the mechanosensory role of non‐motile cilia at the LRO in the establishment of left–right asymmetry.
The Hh pathway is known to play an important role in cilia signaling (Goetz & Anderson, 2010). In a screen for CHD phenotypes, an enrichment in genes associated with Hh signaling was reported (Y. Li et al., 2015). Hh pathway‐associated genes begin to be expressed during the early stages of laterality establishment (Hu et al., 2017) and are required for heart tube asymmetry (Tsiairis & McMahon, 2009; X. M. Zhang et al., 2001). The negative regulators of Hh signaling, Tbc1d32 and Megf8, cause heterotaxy with CHD in mouse mutants (Y. Li et al., 2015). Hh signaling‐induced heart looping abnormalities are associated with defects in myocardial differentiation and the failure to upregulate expression of the cardiac transcription factor Nkx2.5 (X. M. Zhang et al., 2001). The knockdown or overexpression of the GPCR gpr22 in zebrafish also results in changes in cilia length and structure with defective L‐R pattering and randomized cardiac looping leading to heart edema (Verleyen et al., 2014). Dysregulation of the mammalian target of rampamycin (mTOR) signaling pathway results in altered nodal cilia length, with symmetry defects (Burkhalter et al., 2019; Shiaulou Yuan et al., 2012). Knockdown of grk5 in zebrafish augments mTORC1 signaling and fails to break the cardiac symmetry (Burkhalter et al., 2013; Casar Tena et al., 2015), also affecting the expression of genes in the heart, which are important for valve development (Burkhalter et al., 2013).
6. PRIMARY CILIA, HEART DEVELOPMENT, AND CHDs
Research on primary cilia has been ongoing since the 1960s, when primary cilia were initially distinguished as structurally different from motile cilia and found to exist in the majority of mammalian cells (Myklebust et al., 1977). Primary cilia play a crucial role in cell differentiation and embryonic development (May et al., 2021). Disorders linked to primary cilia have been identified across various organ systems (Hale & Sadoshima, 2022). Our understanding of the role of primary cilia in heart development is still in its infancy, but there is an increasing recognition of primary cilia as important biomechanical and molecular regulators of cardiac development (Toomer et al., 2019).
6.1. Primary cilia and associated signaling defects in cardiac valve development
Cilia‐regulated signaling and responses to changes in shear stress are important for cardiac valve development (Gabriel et al., 2021). Unlike other cilia‐related CHDs, valve diseases are more frequently identified in adults than in infants. However, the early signs of disease can be observed during cardiac development with changes in the valve ECM (Fulmer et al., 2020; Morningstar et al., 2021; Toomer et al., 2019). Bicuspid aortic valve (BAV) and mitral valve prolapse (MVP) are cardiac valve defects (LaHaye et al., 2014) commonly identified in syndromic diseases associated with cilia defects (Karp et al., 2012). Cardiac valve diseases often include complications such as cardiac arrhythmias, heart failure, and sudden cardiac death, which sometimes require surgical intervention (Coutsoumbas & Di Pasquale, 2021). Previous studies have shown that genetic ablation of cilia‐related genes disturbs cardiac valve ECM expression causing highly penetrant myxomatous phenotypes like BAV (Fulmer et al., 2019; Toomer et al., 2017) and MVP (Toomer et al., 2019). Primary cilia regulate aortic valve development by directly or indirectly altering the production of critical ECM components (Toomer et al., 2017). A genome‐wide association study (GWAS) using a cohort of BAV and control patients identified single nucleotide polymorphisms in several exocyst complex genes (EXOC4, EXOC6, EXOC8) that are important in regulating ciliogenesis through cargo shuttling to the membrane (Fulmer et al., 2019). The authors further verified the role of the exocyst complex in BAV by knocking down a key linker protein, Exoc5, in both mouse and zebrafish, resulting in a ciliogenesis defect with a BAV phenotype (Fulmer et al., 2019).
Recent studies have identified the role of primary cilia in MVP (Fulmer et al., 2020; Toomer et al., 2019). MVP is characterized by the mechanical incompetence of mitral valve leaflets with increased proteoglycan production and collagen and elastin fragmentation (Fry et al., 2014; Fulmer et al., 2020; Morningstar et al., 2021; Toomer et al., 2019). The known MVP‐causal genes DCHS1 (Durst et al., 2015) and FLNA (Kyndt et al., 2007) also show reduced cilia length in the mitral valve in knockout models, confirming a role for cilia in MVP (Toomer et al., 2019). Toomer et al. (2019) conditionally deleted the ciliary IFT‐B gene Ift88 using an endocardial cell‐specific NfatC1Cre . This results in loss of the cilia axoneme from the endocardial cell‐derived valve mesenchyme, yielding significantly decreased valve interstitial cell density. Additionally, there was robust activation of ECM gene pathways in the anterior mitral leaflets, an indicator of early‐stage myxomatous degeneration resulting in adult myxomatous valve pathology (Toomer et al., 2019). GWAS of MVP cases have identified significant enrichment of MVP‐associated variants in cilia genes. Whole exome sequencing of MVP patients has identified variants in DZIP1 (Toomer et al., 2019). DZIP1 is a cilia‐related gene, which regulates ciliogenesis or Hh signaling (Lapart et al., 2019; Wang et al., 2013; B. Zhang et al., 2015). Knock‐in mice of a human DZIP1 mutation proved to be a genetically accurate model for non‐syndromic MVP with adult myxomatous mitral valves and functional MVP with a reduction in cilia length. At the transcriptome level, the comparison between human DZIP1 mutation knock‐in mice and endocardial cell‐specific Ift88 deletion showed similar ECM pathway changes (Toomer et al., 2019).
The cilia‐regulated pathway ligand, desert Hh (dhh), which is expressed within the cardiac valves, regulates cytoskeleton organization during valve leaflet development. The dhh signal originates from the endocardium, resulting in the paracrine cross‐talk between the endocardium and ciliated valve interstitial cells to shape the valve formation. Conditional deletion of dhh using either NfatC1enCre (specifically expressed in valve endocardial cells that do not undergo EMT) or Tie2Cre (expressed in endothelial or endocardial cells) resulted in an MVP phenotype with no change in cilia length, confirming endocardial cilia‐mediated paracrine cross‐talk in valve development (Fulmer et al., 2020). Conditional deletion of PDGFRα receptor (which also localizes along the ciliary axoneme) with NfatC1enCre , resulted in enlarged anterior valve leaflets with myxomatous MVP‐like phenotypes. PDGFRα suppresses EMT in a subset of valve endothelial cells by regulating the serine/threonine kinase (protein kinase B)/Extracellular signal‐regulated kinase (AKT/ERK) pathway that stabilizes the valve endocardium and prevents a disease phenotype (Moore et al., 2021).
6.2. Primary cilia and associated signaling defects in cardiac atria, ventricle, and other heart structures
AV septal defects (AVSDs) are CHDs commonly associated with heterotaxy syndrome (Francis et al., 2012; Icardo & Sanchez de Vega, 1991; Kathiriya & Srivastava, 2000; Kennedy et al., 2007; Seo et al., 1992; S. Y. Tan et al., 2007). AV septation and dorsal mesenchymal protrusion structure development have been attributed to the migration of second heart field (SHF) progenitor cells (Deepe et al., 2020). Cilia‐mediated Hh signaling is required for the SHF progenitor cell migration and OFT septation (Burnicka‐Turek et al., 2016; Goddeeris et al., 2008; Hoffmann et al., 2009; Washington Smoak et al., 2005). As previously mentioned, cilia‐regulated Hh signaling also plays an important role in laterality establishment (Hu et al., 2017), suggesting a link between AVSD and heterotaxy syndrome. This mechanistic link was further verified with the identification of the first human AVSD gene CRELD1, which is also a component of cilia (Beckert et al., 2021; Burnicka‐Turek et al., 2016).
Turek et al. (2016) identified recessive mutant alleles in two other cilia genes, Dnah11 and Mks1. The Dnah11 mutation results in AVSD with no disturbance in SHF Hh signaling (Burnicka‐Turek et al., 2016). DNAH11 is an ODA component required for cilia motility, and disruption of its function is known to cause laterality defects (Bartoloni et al., 2002; Dougherty et al., 2016). The AVSD observed in Dnah11 mutants could be the result of disturbances in early situs establishment events (Burnicka‐Turek et al., 2016). Additionally, Mks1, a component of the ciliary basal body, causes AVSD when mutated, with downregulation of SHF Hh expression, which is independent of laterality defects (Burnicka‐Turek et al., 2016). Another Mks1 mutant recovered by Cui et al. (2011) also showed CHDs, polycystic kidneys, and randomization of left–right patterning (Cui et al., 2011). The tectonic protein Tctn2, which resides in the transition zone of cilia, interacts with Msk1 and regulates Hh signaling. The Tctn2 knockout mouse also shows phenotypes characteristic of cilia‐mediated Hh defects including cleft palate, polydactyly, VSD, and right‐sided stomach placement (Sang et al., 2011). Another cilia gene, Ftm (also called Rpgrip1l), localizes at the base of cilia and regulates Hh signaling. Ftm mutant embryos show left–right asymmetry defects with randomized heart looping (Vierkotten et al., 2007). Later during heart development, Ftm null hearts show perimembranous VSDs along with muscular ventral septa defects, diminished ventricle wall thickness, and decreases in cilia length, which correlate with reduced cell proliferation. This was attributed to reduced sonic Hh (Shh) and Pdgfra signaling in the ventricles. The Ftm mutant atria and atrial septa showed no defects during heart development and no signs of altered Shh signaling. Since Hh signaling is associated with the development of the atrial septa but no atrial septa developmental defects were observed in Ftm mutants, it is proposed that there are different mechanisms by which Hh signaling regulates atrial and ventricular development. Because no cilia were observed on ventricle septa cells, the defective ventricular septa development in Ftm null mice suggests that the signal for ventricle septal development originates from cilia‐bearing cells in ventricle walls or that ventricular wall cell proliferation contributes to the formation of ventricular septa (Gerhardt et al., 2013).
Loss of function mutants of IFT genes also display cardiac malformations. For example, Ift88 mutant mice show defects in cardiac OFT septation, ventricular trabeculation, AVSD, and cardiac cushion EMT, with no localization of the Hh pathway transcription factor Gli2 in cardiac cilia (Burns et al., 2019; Clement et al., 2009; Willaredt et al., 2012). In a previous study, Washington Smoak et al. (2005) reported that Shh deletion results in abnormal migration of neural crest cells, contributing to arch artery and OFT septation defects. Using an Ift88 hypomorphic allele generated by N‐ethyl‐N‐nitrosourea (ENU) mutagenesis, Willaredt et al. (2012) identified no defects in the migration pattern of cardiac neural crest cells (CNCC) into the heart, but CNCC already migrated into the pharyngeal arches lacked cilia and displayed defective Shh and Bmp2/4 signaling, which are required for OFT septation. Mutant embryos for the gene Kif3a, which is essential for anterograde intraflagellar transport, also show laterality defects along with defective development of the endocardial cushions and compact myocardium (Slough et al., 2008). Recent studies by Peralta et al. (2020) have identified a cilia‐independent, non‐canonical, role of IFT complex B proteins (Ift88, Ift54, and Ift20) in modulating the Hippo pathway effector YAP1 and restricting proepicardial and myocardial tissue size during development (Peralta et al., 2020).
6.3. The mechanosensory role of primary cilia in heart development
Primary cilia are known to have a role as fluid shear stress sensor at node (McGrath et al., 2003) and in the kidney (Nauli et al., 2006; Xu et al., 2007, 2009); however, the exact mechanism by which biomechanical forces regulate gene expression is not fully understood. Yet, the mechanosensory function of primary cilia during development cannot be ignored (Djenoune et al., 2023; Katoh et al., 2023). Extensive tissue remodeling during heart development dramatically changes cardiac fluid shear stress patterns (Garoffolo & Pesce, 2019). In a correlative study of primary cilia distribution on endothelial and endocardium cells in chicken, Van der Heiden et al. (2006) revealed an inverse relation between primary cilia distribution and expression of the high shear stress marker Klf2. Endothelial cells have higher shear stress, resulting in higher expression of stress marker Klf2 with a decreased presence of cilia; likewise in endocardial cells where shear stress is low, Klf2 is not detected, and primary cilia are present. The primary cilia on endothelial and endocardial cells sense shear stress forces and transmit them to the cytoskeleton, triggering a response. In theory, these shear stresses could potentially play a role in shaping the structural organization of the heart chambers (Van der Heiden et al., 2006).
The primary cilia gene Pkd2, responsible for encoding an integral membrane glycoprotein that bears resemblance to subunits of calcium channels, is recognized for its role as a mechanosensor in both blood vessels (MacKay et al., 2020) and at the node (Yoshiba et al., 2012). Pkd2 mutant embryos retained the presence of primary cilia in the heart (Slough et al., 2008). G. Wu et al. (2000) demonstrated that Pkd2 mutant mice die in utero with renal failure and cardiac septation defects (G. Wu et al., 2000). The mutant hearts showed decreased endocardial cushion cellularity and a thin compact myocardium, compared to stage‐matched wild‐type hearts (Slough et al., 2008). Boulter et al. (2001) described mice carrying a targeted mutation in the polycystin‐1 gene (Pkd1), which also encodes an integral membrane protein that localizes to the primary cilium and interacts with Pkd2. Pkd1 mutant mice showed AVSD with disorganization and thinning of the myocardial wall and double‐outlet right ventricle (Boulter et al., 2001). A recent study in zebrafish has also identified a synergistic role for the pkd genes (pkd1, pkd2, and pkd1l1) in blood flow‐driven valve development by repressing the expression of klf2a and klf2b (Juan et al., 2023). During cardiac trabeculation, cardiac contraction and hemodynamic forces exert mechanical stresses, which are detected by primary cilia on ventricular endocardial cells and decoded by the flow‐responsive transcription factor Klf2a. This leads to the activation of notch1b‐efnb2a‐nrg1 pathway, which regulates the cross‐talk between the endocardium and myocardium during cardiac trabeculation. Using ift88 morphant zebrafish, Samsa et al. (2015) noted that primary cilia on endocardial cells are required for notch1b activation. However, notch1 activation is independent of ciliary Hh signaling but is required for functional primary cilia (Samsa et al., 2015). This suggests that primary cilia might serve as sensors of shear stress, and this function could contribute to cardiac valve and chamber development.
7. CILIOPATHIES, CHDs, AND CLINICAL RELEVANCE
CHDs are often found in patients with clinically recognized syndromes and developmental disorders affecting cilia function (Barisic et al., 2015; Elbedour et al., 1994; Engesaeth et al., 1993). For instance, in motile ciliopathies, also known as PCD, nearly half of the patients exhibit abnormal situs, and approximately 3.5%–6% manifest CHD as a part of their clinical profile (Best et al., 2019; Kennedy et al., 2007; Noone et al., 2004; Shapiro et al., 2014). Autosomal dominant polycystic kidney disease (ADPKD), a ciliopathy, often presents with significant heart‐related complications, contributing to increased morbidity (Rahman et al., 2009). Notably, around a quarter of ADPKD patients receive a diagnosis of MVP, in addition to other cardiac anomalies (Toomer et al., 2019). Dysregulations of ECM in both MVP and ADPKD impair molecular architecture and function, contributing to the disease phenotype (Toomer et al., 2019; Wilson et al., 1992). In Bardet–Biedl syndrome (BBS), another ciliopathy, individuals exhibit a 170‐fold higher prevalence of laterality defects, compared to the general population, although this occurrence remains lower than in patients with primary ciliary dyskinesia (PCD) (Olson et al., 2019). Furthermore, individuals with BBS also display defects such as AVSDs, vascular anomalies, and dilated cardiomyopathy (Niederlova et al., 2019; Yadav et al., 2013). Trisomy 21, a chromosomal copy number disorder, is associated not only with neurological abnormalities but also with CHD. Disruption of primary cilia formation and signaling in Trisomy 21 is attributed to elevated expression levels of the centrosomal protein Pericentrin, a gene located on chromosome 21 in humans. This excess Pericentrin disrupts ciliary protein trafficking and leads to defective Shh signaling as observed in murine Trisomy 21 models (Galati et al., 2018; Jewett et al., 2023). Pathogenic mutations in cilia gene CRELD1 have also been identified in Trisomy 21 patients, resulting in AVSD defects (Asim et al., 2018).
To date, nearly 51 genes associated with PCD have been identified (Zhao et al., 2021). The majority of mutations fall into the category of loss‐of‐function variants (Knowles et al., 2013); however, copy number variants have also been detected in heterotaxy patients (Fakhro et al., 2011). Most of the affected PCD genes encode proteins associated with the cilia motility machinery (Y. Li et al., 2016; Reiter & Leroux, 2017). Nevertheless, alterations in genes that are active within the cytoplasm and participate in the preliminary formation of cilia have also been associated with PCD (Horani et al., 2012, 2013; Knowles et al., 2013). For instance, Nakhleh et al. (2012) observed that a notable percentage of individuals with heterotaxy and congenital heart disease exhibited impaired cilia function with an irregular ciliary beat pattern, although the ciliary structure itself remained intact. This phenomenon might signify a modified manifestation of PCD (Nakhleh et al., 2012).
8. CONCLUDING REMARKS AND PERSPECTIVES
Advances in molecular biology, disease modeling, microscopy and genome sequencing have illuminated the pivotal role of the minuscule cellular organelle, the cilium, in the establishment of cardiac asymmetry and heart function. Investigations employing patient‐specific cohort studies for CHD or forward genetic screens in mice have uncovered novel de novo mutations in cilia‐related genes. Furthermore, mutations in genes involved in pathways signaling through cilia localization have also emerged as contributors to cardiac developmental abnormalities. Experiments conducted in lower vertebrate animal models and human cell lines have further deepened our understanding of the multifaceted functions of cilia in development and homeostasis. The ongoing discovery of new cilia genes associated with CHD holds the promise for enhancing clinical awareness of the genetic causes of CHD, leading to guidance for family planning, improved prognosis, and the development of prenatal genetic screening tests for complex CHD cases requiring urgent intervention. Nonetheless, our knowledge regarding the precise involvement of cilia in regulating cardiac development remains in its nascent stages. Additional research is imperative to elucidate the diverse functions of motile and non‐motile primary cilia in cardiac development and function.
AUTHOR CONTRIBUTIONS
Wasay Mohiuddin Shaikh Qureshi performed research and wrote the manuscript. Kathryn E. Hentges wrote the manuscript, edited the manuscript, and obtained funding.
ACKNOWLEDGMENTS
We thank Nouf Althali for assistance with the creation of figures in BioRender.com.
Shaikh Qureshi, W. M. , & Hentges, K. E. (2024). Functions of cilia in cardiac development and disease. Annals of Human Genetics, 88, 4–26. 10.1111/ahg.12534
DATA AVAILABILITY STATEMENT
The data that support the findings of this study are openly available in Pubmed at https://pubmed.ncbi.nlm.nih.gov.
REFERENCES
- Adachi, H. , Saijoh, Y. , Mochida, K. , Ohishi, S. , Hashiguchi, H. , Hirao, A. , & Hamada, H. (1999). Determination of left/right asymmetric expression of nodal by a left side‐specific enhancer with sequence similarity to a lefty‐2 enhancer. Genes & Development, 13(12), 1589–1600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agarwal, R. , Varghese, R. , Jesudian, V. , & Moses, J. (2021). The heterotaxy syndrome: Associated congenital heart defects and management. Indian Journal of Thoracic and Cardiovascular Surgery, 37(1), 67–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alten, L. , Schuster‐Gossler, K. , Beckers, A. , Groos, S. , Ulmer, B. , Hegermann, J. , Ochs, M. , & Gossler, A. (2012). Differential regulation of node formation, nodal ciliogenesis and cilia positioning by Noto and Foxj1. Development (Cambridge, England), 139(7), 1276–1284. [DOI] [PubMed] [Google Scholar]
- Althali, N. J. , & Hentges, K. E. (2022). Genetic insights into non‐syndromic Tetralogy of Fallot. Frontiers in Physiology, 13, 1012665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Álvarez‐Satta, M. , Lago‐Docampo, M. , Bea‐Mascato, B. , Solarat, C. , Castro‐Sánchez, S. , Christensen, S. T. , & Valverde, D. (2021). ALMS1 regulates TGF‐β signaling and morphology of primary cilia. Frontiers in Cell and Developmental Biology, 9, 623829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asim, A. , Agarwal, S. , Panigrahi, I. , Sarangi, A. N. , Muthuswamy, S. , & Kapoor, A. (2018). CRELD1 gene variants and atrioventricular septal defects in Down syndrome. Gene, 641, 180–185. [DOI] [PubMed] [Google Scholar]
- Baker, K. , & Beales, P. L. (2009). Making sense of cilia in disease: The human ciliopathies. American Journal of Medical Genetics Part C, Seminars in Medical Genetics, 151C(4), 281–295. [DOI] [PubMed] [Google Scholar]
- Bakey, Z. , Cabrera, O. A. , Hoefele, J. , Antony, D. , Wu, K. , Stuck, M. W. , Micha, D. , Eguether, T. , Smith, A. O. , Van Der Wel, N. N. , Wagner, M. , Strittmatter, L. , Beales, P. L. , Jonassen, J. A. , Thiffault, I. , Cadieux‐Dion, M. , Boyes, L. , Sharif, S. , Tüysüz, B. , & Pazour, G. J. (2023). IFT74 variants cause skeletal ciliopathy and motile cilia defects in mice and humans. Plos Genetics, 19(6), e1010796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bangs, F. , & Anderson, K. V. (2017). Primary cilia and mammalian hedgehog signaling. Cold Spring Harbor Perspectives in Biology, 9(5), a028175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barisic, I. , Boban, L. , Loane, M. , Garne, E. , Wellesley, D. , Calzolari, E. , Dolk, H. , Addor, M.‐C. , Bergman, J. E. , Braz, P. , Draper, E. S. , Haeusler, M. , Khoshnood, B. , Klungsoyr, K. , Pierini, A. , Queisser‐Luft, A. , Rankin, J. , Rissmann, A. , & Verellen‐Dumoulin, C. (2015). Meckel–Gruber Syndrome: A population‐based study on prevalence, prenatal diagnosis, clinical features, and survival in Europe. European Journal of Human Genetics, 23(6), 746–752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartoloni, L. , Blouin, J.‐L. , Pan, Y. , Gehrig, C. , Maiti, A. K. , Scamuffa, N. , Rossier, C. , Jorissen, M. , Armengot, M. , Meeks, M. , Mitchison, H. M. , Chung, E. M. K. , Delozier‐Blanchet, C. D. , Craigen, W. J. , & Antonarakis, S. E. (2002). Mutations in the DNAH11 (axonemal heavy chain dynein type 11) gene cause one form of situs inversus totalis and most likely primary ciliary dyskinesia. PNAS, 99(16), 10282–10286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battini, L. , Fedorova, E. , Macip, S. , Li, X. , Wilson, P. D. , & Gusella, G. L. (2006). Stable knockdown of polycystin‐1 confers integrin‐alpha2beta1‐mediated anoikis resistance. Journal of the American Society of Nephrology, 17(11), 3049–3058. [DOI] [PubMed] [Google Scholar]
- Becker‐Heck, A. , Zohn, I. E. , Okabe, N. , Pollock, A. , Lenhart, K. B. , Sullivan‐Brown, J. , Mcsheene, J. , Loges, N. T. , Olbrich, H. , Haeffner, K. , Fliegauf, M. , Horvath, J. , Reinhardt, R. , Nielsen, K. G. , Marthin, J. K. , Baktai, G. , Anderson, K. V. , Geisler, R. , Niswander, L. , & Burdine, R. D. (2011). The coiled‐coil domain containing protein CCDC40 is essential for motile cilia function and left‐right axis formation. Nature Genetics, 43(1), 79–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beckert, V. , Rassmann, S. , Kayvanjoo, A. H. , Klausen, C. , Bonaguro, L. , Botermann, D. S. , Krause, M. , Moreth, K. , Spielmann, N. , Da Silva‐Buttkus, P. , Fuchs, H. , Gailus‐Durner, V. , De Angelis, M. H. , Händler, K. , Ulas, T. , Aschenbrenner, A. C. , Mass, E. , & Wachten, D. (2021). Creld1 regulates myocardial development and function. Journal of Molecular and Cellular Cardiology, 156, 45–56. [DOI] [PubMed] [Google Scholar]
- Belo, J. A. , Marques, S. , & Inácio, J. M. (2017). The role of Cerl2 in the establishment of left‐right asymmetries during axis formation and heart development. Journal of Cardiovascular Development and Disease, 4(4), 23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Best, S. , Shoemark, A. , Rubbo, B. , Patel, M. P. , Fassad, M. R. , Dixon, M. , Rogers, A. V. , Hirst, R. A. , Rutman, A. , Ollosson, S. , Jackson, C. L. , Goggin, P. , Thomas, S. , Pengelly, R. , Cullup, T. , Pissaridou, E. , Hayward, J. , Onoufriadis, A. , O'callaghan, C. , & Hogg, C. (2019). Risk factors for situs defects and congenital heart disease in primary ciliary dyskinesia. Thorax, 74(2), 203–205. [DOI] [PubMed] [Google Scholar]
- Blum, M. , Andre, P. , Muders, K. , Schweickert, A. , Fischer, A. , Bitzer, E. , Bogusch, S. , Beyer, T. , Van Straaten, H. W. M. , & Viebahn, C. (2007). Ciliation and gene expression distinguish between node and posterior notochord in the mammalian embryo. Differentiation, 75(2), 133–146. [DOI] [PubMed] [Google Scholar]
- Borovina, A. , Superina, S. , Voskas, D. , & Ciruna, B. (2010). Vangl2 directs the posterior tilting and asymmetric localization of motile primary cilia. Nature Cell Biology, 12(4), 407–412. [DOI] [PubMed] [Google Scholar]
- Boulter, C. , Mulroy, S. , Webb, S. , Fleming, S. , Brindle, K. , & Sandford, R. (2001). Cardiovascular, skeletal, and renal defects in mice with a targeted disruption of the Pkd1 gene. PNAS, 98(21), 12174–12179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brennan, J. , Norris, D. P. , & Robertson, E. J. (2002). Nodal activity in the node governs left‐right asymmetry. Genes & Development, 16(18), 2339–2344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brewer, K. M. , Engle, S. E. , Bansal, R. , Brewer, K. K. , Jasso, K. R. , Mcintyre, J. C. , Vaisse, C. , Reiter, J. F. , & Berbari, N. F. (2023). Physiological condition‐dependent changes in ciliary GPCR localization in the brain. Eneuro, 10(3), ENEURO.0360–22.2023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruneau, B. G. (2013). Signaling and transcriptional networks in heart development and regeneration. Cold Spring Harbor Perspectives in Biology, 5(3), a008292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckingham, M. , Meilhac, S. , & Zaffran, S. (2005). Building the mammalian heart from two sources of myocardial cells. Nature Reviews Genetics, 6(11), 826–835. [DOI] [PubMed] [Google Scholar]
- Buijtendijk, M. F. J. , Barnett, P. , & Van Den Hoff, M. J. B. (2020). Development of the human heart. American Journal of Medical Genetics Part C, Seminars in Medical Genetics, 184(1), 7–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burkhalter, M. D. , Fralish, G. B. , Premont, R. T. , Caron, M. G. , & Philipp, M. (2013). Grk5l controls heart development by limiting mTOR signaling during symmetry breaking. Cell Reports, 4(4), 625–632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burkhalter, M. D. , Sridhar, A. , Sampaio, P. , Jacinto, R. , Burczyk, M. S. , Donow, C. , Angenendt, M. , Hempel, M. , Walther, P. , Pennekamp, P. , Omran, H. , Lopes, S. S. , Ware, S. M. , & Philipp, M. (2019). Imbalanced mitochondrial function provokes heterotaxy via aberrant ciliogenesis. Journal of Clinical Investigation, 129(7), 2841–2855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burnicka‐Turek, O. , Steimle, J. D. , Huang, W. , Felker, L. , Kamp, A. , Kweon, J. , Peterson, M. , Reeves, R. H. , Maslen, C. L. , Gruber, P. J. , Yang, X. H. , Shendure, J. , & Moskowitz, I. P. (2016). Cilia gene mutations cause atrioventricular septal defects by multiple mechanisms. Human Molecular Genetics, 25(14), 3011–3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns, T. A. , Deepe, R. N. , Bullard, J. , Phelps, A. L. , Toomer, K. A. , Hiriart, E. , Norris, R. A. , Haycraft, C. J. , & Wessels, A. (2019). A novel mouse model for cilia‐associated cardiovascular anomalies with a high penetrance of total anomalous pulmonary venous return. Anatomical Record (Hoboken), 302(1), 136–145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casar Tena, T. , Burkhalter, M. D. , & Philipp, M. (2015). Left‐right asymmetry in the light of TOR: An update on what we know so far. Biologie Cellulaire, 107(9), 306–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chatterjee, B. , Richards, K. , Bucan, M. , & Lo, C. (2007). Nt mutation causing laterality defects associated with deletion of rotatin. Mammalian Genome, 18(5), 310–315. [DOI] [PubMed] [Google Scholar]
- Chen, W. , Zhang, Y. , Yang, S. , Shi, Z. , Zeng, W. , Lu, Z. , & Zhou, X. (2019). Bi‐allelic mutations in NUP205 and NUP210 are associated with abnormal cardiac left‐right patterning. Circulation: Genomic and Precision Medicine, 12(7), e002492. [DOI] [PubMed] [Google Scholar]
- Christensen, S. T. , Morthorst, S. K. , Mogensen, J. B. , & Pedersen, L. B. (2017). Primary cilia and coordination of receptor tyrosine kinase (RTK) and transforming growth factor β (TGF‐β) signaling. Cold Spring Harbor Perspectives in Biology, 9(6), a028167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen, S. T. , Pedersen, L. B. , Schneider, L. , & Satir, P. (2007). Sensory cilia and integration of signal transduction in human health and disease. Traffic (Copenhagen, Denmark), 8(2), 97–109. [DOI] [PubMed] [Google Scholar]
- Clement, C. A. , Ajbro, K. D. , Koefoed, K. , Vestergaard, M. L. , Veland, I. R. , Henriques De Jesus, M. P. R. , Pedersen, L. B. , Benmerah, A. , Andersen, C. Y. , Larsen, L. A. , & Christensen, S. T. (2013). TGF‐β signaling is associated with endocytosis at the pocket region of the primary cilium. Cell Reports, 3(6), 1806–1814. [DOI] [PubMed] [Google Scholar]
- Clement, C. A. , Kristensen, S. G. , MøllgåRd, K. , Pazour, G. J. , Yoder, B. K. , Larsen, L. A. , & Christensen, S. T. (2009). The primary cilium coordinates early cardiogenesis and hedgehog signaling in cardiomyocyte differentiation. Journal of Cell Science, 122(17), 3070–3082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clowes, C. , Boylan, M. G. S. , Ridge, L. A. , Barnes, E. , Wright, J. A. , & Hentges, K. E. (2014). The functional diversity of essential genes required for mammalian cardiac development. Genesis, 52(8), 713–737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collignon, J. , Varlet, I. , & Robertson, E. J. (1996). Relationship between asymmetric nodal expression and the direction of embryonic turning. Nature, 381(6578), 155–158. [DOI] [PubMed] [Google Scholar]
- Coutsoumbas, G. V. , & Di Pasquale, G. (2021). Mitral valve prolapse with ventricular arrhythmias: Does it carries a worse prognosis? European Heart Journal Supplements, 23, (Suppl E), E77–E82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui, C. , Chatterjee, B. , Francis, D. , Yu, Q. , Sanagustin, J T. , Francis, R. , Tansey, T. , Henry, C. , Wang, B. , Lemley, B. , Pazour, G. J. , & Lo, C. W. (2011). Disruption of Mks1 localization to the mother centriole causes cilia defects and developmental malformations in Meckel–Gruber syndrome. Disease Models & Mechanisms, 4(1), 43–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deepe, R. , Fitzgerald, E. , Wolters, R. , Drummond, J. , Guzman, K. , Hoff, M. J. B. V. D. , & Wessels, A. (2020). The mesenchymal cap of the atrial septum and atrial and atrioventricular septation. Journal of Cardiovascular Development and Disease, 7(4), 50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desgrange, A. , Le Garrec, J.‐F. , Bernheim, S. , Bønnelykke, T. H. , & Meilhac, S. M. (2020). Transient nodal signaling in left precursors coordinates opposed asymmetries shaping the heart loop. Developmental Cell, 55(4), 413–431.e6. [DOI] [PubMed] [Google Scholar]
- Diguet, N. , Le Garrec, J. F. , Lucchesi, T. , & Meilhac, S. M. (2015). Imaging and analyzing primary cilia in cardiac cells. Methods in Cell Biology, 127, 55–73. [DOI] [PubMed] [Google Scholar]
- Djenoune, L. , Berg, K. , Brueckner, M. , & Yuan, S. (2022). A change of heart: New roles for cilia in cardiac development and disease. Nature Reviews Cardiology, 19(4), 211–227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Djenoune, L. , Mahamdeh, M. , Truong, T. V. , Nguyen, C. T. , Fraser, S. E. , Brueckner, M. , Howard, J. , & Yuan, S. (2023). Cilia function as calcium‐mediated mechanosensors that instruct left‐right asymmetry. Science, 379(6627), 71–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dodge‐Khatami, A. (2016). Advances and research in congenital heart disease. Translational Pediatrics, 5(3), 109–111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dougherty, G. W. , Loges, N. T. , Klinkenbusch, J. A. , Olbrich, H. , Pennekamp, P. , Menchen, T. , Raidt, J. , Wallmeier, J. , Werner, C. , Westermann, C. , Ruckert, C. , Mirra, V. , Hjeij, R. , Memari, Y. , Durbin, R. , Kolb‐Kokocinski, A. , Praveen, K. , Kashef, M. A. , Kashef, S. , & Omran, H. (2016). DNAH11 localization in the proximal region of respiratory cilia defines distinct outer dynein arm complexes. American Journal of Respiratory Cell and Molecular Biology, 55(2), 213–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drummond, I. A. (2012). Cilia functions in development. Current Opinion in Cell Biology, 24(1), 24–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durst, R. , Sauls, K. , Peal, D. S. , Devlaming, A. , Toomer, K. , Leyne, M. , Salani, M. , Talkowski, M. E. , Brand, H. , Perrocheau, M. , Simpson, C. , Jett, C. , Stone, M. R. , Charles, F. , Chiang, C. , Lynch, S. N. , Bouatia‐Naji, N. , Delling, F. N. , Freed, L. A. , & Slaugenhaupt, S. A. (2015). Mutations in DCHS1 cause mitral valve prolapse. Nature, 525(7567), 109–113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eitler, K. , Bibok, A. , & Telkes, G. (2022). Situs inversus totalis: A clinical review. International Journal of General Medicine, 15, 2437–2449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elbedour, K. , Zucker, N. , Zalzstein, E. , Barki, Y. , & Carmi, R. (1994). Cardiac abnormalities in the Bardet–Biedl syndrome: Echocardiographic studies of 22 patients. American Journal of Medical Genetics, 52(2), 164–169. [DOI] [PubMed] [Google Scholar]
- Engesaeth, V. G , Warner, J. O. , & Bush, A. (1993). New associations of primary ciliary dyskinesia syndrome. Pediatric Pulmonology, 16(1), 9–12. [DOI] [PubMed] [Google Scholar]
- Fakhro, K. A. , Choi, M. , Ware, S. M. , Belmont, J. W. , Towbin, J. A. , Lifton, R. P. , Khokha, M. K. , & Brueckner, M. (2011). Rare copy number variations in congenital heart disease patients identify unique genes in left‐right patterning. PNAS, 108(7), 2915–2920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fawcett, D. W. , & Porter, K. R. (1954). A study of the fine structure of ciliated epithelia. Journal of Morphology, 94(2), 221–281. [Google Scholar]
- Ferreira, R. R. , Fukui, H. , Chow, R. , Vilfan, A. , & Vermot, J. (2019). The cilium as a force sensor‐myth versus reality. Journal of Cell Science, 132(14), jcs213496. [DOI] [PubMed] [Google Scholar]
- Field, S. , Riley, K.‐L. , Grimes, D T. , Hilton, H. , Simon, M. , Powles‐Glover, N. , Siggers, P. , Bogani, D. , Greenfield, A. , & Norris, D. P. (2011). Pkd1l1 establishes left‐right asymmetry and physically interacts with Pkd2. Development (Cambridge, England), 138(6), 1131–1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis, R. J. B. , Christopher, A. , Devine, W. A. , Ostrowski, L. , & Lo, C. (2012). Congenital heart disease and the specification of left‐right asymmetry. American Journal of Physiology. Heart and Circulatory Physiology, 302(10), H2102–H2111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fry, A. M. , Leaper, M. J. , & Bayliss, R. (2014). The primary cilium: Guardian of organ development and homeostasis. Organogenesis, 10(1), 62–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujinaga, M. (1997). Development of sidedness of asymmetric body structures in vertebrates. International Journal of Developmental Biology, 41(2), 153–186. [PubMed] [Google Scholar]
- Fulmer, D. , Toomer, K. , Guo, L. , Moore, K. , Glover, J. , Moore, R. , Stairley, R. , Lobo, G. , Zuo, X. , Dang, Y. , Su, Y. , Fogelgren, B. , Gerard, P. , Chung, D. , Heydarpour, M. , Mukherjee, R. , Body, S. C. , Norris, R. A. , & Lipschutz, J. H. (2019). Defects in the exocyst‐cilia machinery cause bicuspid aortic valve disease and aortic stenosis. Circulation, 140(16), 1331–1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulmer, D. , Toomer, K. A. , Glover, J. , Guo, L. , Moore, K. , Moore, R. , Stairley, R. , Gensemer, C. , Abrol, S. , Rumph, M. K. , Emetu, F. , Lipschutz, J. H. , Mcdowell, C. , Bian, J. , Wang, C. , Beck, T. , Wessels, A. , Renault, M.‐A. , & Norris, R. A. (2020). Desert hedgehog‐primary cilia cross talk shapes mitral valve tissue by organizing smooth muscle actin. Developmental Biology, 463(1), 26–38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabriel, G. C. , Young, C. B. , & Lo, C. W. (2021). Role of cilia in the pathogenesis of congenital heart disease. Seminars in Cell & Developmental Biology, 110, 2–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galati, D. F. , Sullivan, K. D. , Pham, A. T. , Espinosa, J. M. , & Pearson, C. G. (2018). Trisomy 21 represses cilia formation and function. Developmental Cell, 46(5), 641–650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garoffolo, G. , & Pesce, M. (2019). Mechanotransduction in the cardiovascular system: From developmental origins to homeostasis and pathology. Cells, 8(12), 1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gencer, S. , Oleinik, N. , Kim, J. , Panneer Selvam, S. , De Palma, R. , Dany, M. , Nganga, R. , Thomas, R. J. , Senkal, C. E. , Howe, P. H. , & Ogretmen, B. (2017). TGF‐β receptor I/II trafficking and signaling at primary cilia are inhibited by ceramide to attenuate cell migration and tumor metastasis. Science Signaling, 10(502), eaam7464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- George, R. M. , Maldonado‐Velez, G. , & Firulli, A. B. (2020). The heart of the neural crest: Cardiac neural crest cells in development and regeneration. Development (Cambridge, England), 147(20), dev188706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerhardt, C. , Lier, J. M. , Kuschel, S. , & Rüther, U. (2013). The ciliary protein Ftm is required for ventricular wall and septal development. PLoS ONE, 8(2), e57545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gibbons, I. R. (1961). The relationship between the fine structure and direction of beat in gill cilia of a lamellibranch mollusc. The Journal of Biophysical and Biochemical Cytology, 11(1), 179–205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goddeeris, M. M. , Rho, S. , Petiet, A. , Davenport, C. L. , Johnson, G. A , Meyers, E. N. , & Klingensmith, J. (2008). Intracardiac septation requires hedgehog‐dependent cellular contributions from outside the heart. Development (Cambridge, England), 135(10), 1887–1895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goetz, S. C. , & Anderson, K. V. (2010). The primary cilium: A signalling centre during vertebrate development. Nature Reviews Genetics, 11(5), 331–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorivodsky, M. , Mukhopadhyay, M. , Wilsch‐Braeuninger, M. , Phillips, M. , Teufel, A. , Kim, C. , Malik, N. , Huttner, W. , & Westphal, H. (2009). Intraflagellar transport protein 172 is essential for primary cilia formation and plays a vital role in patterning the mammalian brain. Developmental Biology, 325(1), 24–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hale, Z. E. , & Sadoshima, J. (2022). Primary cilia and their role in acquired heart disease. Cells, 11(6), 960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hao, K. , Chen, Y. , Yan, X. , & Zhu, X. (2021). Cilia locally synthesize proteins to sustain their ultrastructure and functions. Nature Communications, 12(1), 6971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison, M. J. , Shapiro, A. J. , & Kennedy, M. P. (2016). Congenital heart disease and primary ciliary dyskinesia. Paediatric Respiratory Reviews, 18, 25–32. [DOI] [PubMed] [Google Scholar]
- Hartill, V. L. , Van De Hoek, G. , Patel, M. P. , Little, R. , Watson, C. M. , Berry, I. R. , Shoemark, A. , Abdelmottaleb, D. , Parkes, E. , Bacchelli, C. , Szymanska, K. , Knoers, N. V. , Scambler, P. J. , Ueffing, M. , Boldt, K. , Yates, R. , Winyard, P. J. , Adler, B. , & Johnson, C. A. (2018). DNAAF1 links heart laterality with the AAA+ ATPase RUVBL1 and ciliary intraflagellar transport. Human Molecular Genetics, 27(3), 529–545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirokawa, N. , Tanaka, Y. , & Okada, Y. (2009). Left‐right determination: Involvement of molecular motor KIF3, cilia, and nodal flow. Cold Spring Harbor Perspectives in Biology, 1(1), a000802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirokawa, N. , Tanaka, Y. , & Okada, Y. (2012). Cilia, KIF3 molecular motor and nodal flow. Current Opinion in Cell Biology, 24(1), 31–39. [DOI] [PubMed] [Google Scholar]
- Hjeij, R. , Lindstrand, A. , Francis, R. , Zariwala, M A. , Liu, X. , Li, Y. , Damerla, R. , Dougherty, G. W. , Abouhamed, M. , Olbrich, H. , Loges, N. T. , Pennekamp, P. , Davis, E. E. , Carvalho, C. M. B. , Pehlivan, D. , Werner, C. , Raidt, J. , Köhler, G. , Häffner, K. , & Omran, H. (2013). ARMC4 mutations cause primary ciliary dyskinesia with randomization of left/right body asymmetry. American Journal of Human Genetics, 93(2), 357–367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hjeij, R. , Onoufriadis, A. , Watson, C. M. , Slagle, C. E. , Klena, N. T. , Dougherty, G. W. , Kurkowiak, M. , Loges, N. T. , Diggle, C. P. , Morante, N. F. C. , Gabriel, G. C. , Lemke, K. L. , Li, Y. , Pennekamp, P. , Menchen, T. , Konert, F. , Marthin, J. K. , Mans, D. A. , & Mitchison, H. M. (2014). CCDC151 mutations cause primary ciliary dyskinesia by disruption of the outer dynein arm docking complex formation. American Journal of Human Genetics, 95(3), 257–274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann, A. D. , Peterson, M. A. , Friedland‐Little, J. M. , Anderson, S. A. , & Moskowitz, I. P. (2009). Sonic hedgehog is required in pulmonary endoderm for atrial septation. Development (Cambridge, England), 136(10), 1761–1770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honda, H. , Abe, T. , & Fujimori, T. (2020). The chiral looping of the embryonic heart is formed by the combination of three axial asymmetries. Biophysical Journal, 118(3), 742–752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horani, A. , Druley, T. E. , Zariwala, M. A. , Patel, A. C. , Levinson, B. T. , Van Arendonk, L. G. , Thornton, K. C. , Giacalone, J. C. , Albee, A. J. , Wilson, K. S. , Turner, E. H. , Nickerson, D. A. , Shendure, J. , Bayly, P. V. , Leigh, M. W. , Knowles, M. R. , Brody, S. L. , Dutcher, S. K. , & Ferkol, T. W. (2012). Whole‐exome capture and sequencing identifies HEATR2 mutation as a cause of primary ciliary dyskinesia. American Journal of Human Genetics, 91(4), 685–693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horani, A. , Ferkol, T. W. , Shoseyov, D. , Wasserman, M. G. , Oren, Y. S. , Kerem, B. , Amirav, I. , Cohen‐Cymberknoh, M. , Dutcher, S. K. , Brody, S. L. , Elpeleg, O. , & Kerem, E. (2013). LRRC6 mutation causes primary ciliary dyskinesia with dynein arm defects. PLoS ONE, 8(3), e59436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houde, C. , Dickinson, R. J. , Houtzager, V. M. , Cullum, R. , Montpetit, R. , Metzler, M. , Simpson, E. M. , Roy, S. , Hayden, M. R. , Hoodless, P. A. , & Nicholson, D. W. (2006). Hippi is essential for node cilia assembly and Sonic hedgehog signaling. Developmental Biology, 300(2), 523–533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houyel, L. , & Meilhac, S. M. (2021). Heart development and congenital structural heart defects. Annual Review of Genomics and Human Genetics, 22(1), 257–284. [DOI] [PubMed] [Google Scholar]
- Hu, G. , Li, G. , Wang, H. , & Wang, Y. (2017). Hedgehog participates in the establishment of left‐right asymmetry during amphioxus development by controlling Cerberus expression. Development (Cambridge, England), 144(24), 4694–4703. [DOI] [PubMed] [Google Scholar]
- Huang, S. , Hirota, Y. , & Sawamoto, K. (2009). Various facets of vertebrate cilia: Motility, signaling, and role in adult neurogenesis. Proceedings of the Japan Academy, Ser. B, Physical and Biological Sciences, 85(8), 324–336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hutson, M. R. , & Kirby, M. L. (2007). Model systems for the study of heart development and disease. Cardiac neural crest and conotruncal malformations. Seminars in Cell & Developmental Biology, 18(1), 101–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibanez‐Tallon, I. (2002). Loss of function of axonemal dynein Mdnah5 causes primary ciliary dyskinesia and hydrocephalus. Human Molecular Genetics, 11(6), 715–721. [DOI] [PubMed] [Google Scholar]
- Icardo, J. M. , & Sanchez De Vega, M. J. (1991). Spectrum of heart malformations in mice with situs solitus, situs inversus, and associated visceral heterotaxy. Circulation, 84(6), 2547–2558. [DOI] [PubMed] [Google Scholar]
- Inácio, J. M. , Marques, S. , Nakamura, T. , Shinohara, K. , Meno, C. , Hamada, H. , & Belo, J. A. (2013). The dynamic right‐to‐left translocation of Cerl2 is involved in the regulation and termination of Nodal activity in the mouse node. PLoS ONE, 8(3), e60406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishikawa, T. (2017). Axoneme structure from motile cilia. Cold Spring Harbor Perspectives in Biology, 9(1), a028076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanovitch, K. , Temiño, S. , & Torres, M. (2017). Live imaging of heart tube development in mouse reveals alternating phases of cardiac differentiation and morphogenesis. Elife, 6, e30668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jewett, C. E. , Mccurdy, B. L. , O'toole, E. T. , Stemm‐Wolf, A. J. , Given, K. S. , Lin, C. H. , Olsen, V. , Martin, W. , Reinholdt, L. , Espinosa, J. M. , Sullivan, K. D. , Macklin, W. B. , Prekeris, R. , & Pearson, C. G. (2023). Trisomy 21 induces pericentrosomal crowding delaying primary ciliogenesis and mouse cerebellar development. Elife, 12, e78202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juan, T. , Ribeiro Da Silva, A. , Cardoso, B. , Lim, S. , Charteau, V. , & Stainier, D. Y. R. (2023). Multiple pkd and piezo gene family members are required for atrioventricular valve formation. Nature Communications, 14(1), 214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamura, K. , Kobayashi, D. , Uehara, Y. , Koshida, S. , Iijima, N. , Kudo, A. , Yokoyama, T. , & Takeda, H. (2011). Pkd1l1 complexes with Pkd2 on motile cilia and functions to establish the left‐right axis. Development (Cambridge, England), 138(6), 1121–1129. [DOI] [PubMed] [Google Scholar]
- Karp, N. , Grosse‐Wortmann, L. , & Bowdin, S. (2012). Severe aortic stenosis, bicuspid aortic valve and atrial septal defect in a child with Joubert Syndrome and Related Disorders (JSRD)—A case report and review of congenital heart defects reported in the human ciliopathies. European Journal of Medical Genetics, 55(11), 605–610. [DOI] [PubMed] [Google Scholar]
- Kathiriya, I. S. , & Srivastava, D. (2000). Left‐right asymmetry and cardiac looping: Implications for cardiac development and congenital heart disease. American Journal of Medical Genetics, 97(4), 271–279. [DOI] [PubMed] [Google Scholar]
- Katoh, T. A. , Omori, T. , Mizuno, K. , Sai, X. , Minegishi, K. , Ikawa, Y. , Nishimura, H. , Itabashi, T. , Kajikawa, E. , Hiver, S. , Iwane, A. H. , Ishikawa, T. , Okada, Y. , Nishizaka, T. , & Hamada, H. (2023). Immotile cilia mechanically sense the direction of fluid flow for left‐right determination. Science, 379(6627), 66–71. [DOI] [PubMed] [Google Scholar]
- Kawasumi, A. , Nakamura, T. , Iwai, N. , Yashiro, K. , Saijoh, Y. , Belo, J. A. , Shiratori, H. , & Hamada, H. (2011). Left–right asymmetry in the level of active Nodal protein produced in the node is translated into left–right asymmetry in the lateral plate of mouse embryos. Developmental Biology, 353(2), 321–330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly, R. G. (2012). The second heart field. Current Topics in Developmental Biology, 100, 33–65. [DOI] [PubMed] [Google Scholar]
- Kennedy, M. P. , Omran, H. , Leigh, M. W. , Dell, S. , Morgan, L. , Molina, P. L. , Robinson, B. V. , Minnix, S. L. , Olbrich, H. , Severin, T. , Ahrens, P. , Lange, L. , Morillas, H. N. , Noone, P. G. , Zariwala, M. A. , & Knowles, M. R. (2007). Congenital heart disease and other heterotaxic defects in a large cohort of patients with primary ciliary dyskinesia. Circulation, 115(22), 2814–2821. [DOI] [PubMed] [Google Scholar]
- Kiesel, P. , Alvarez Viar, G. , Tsoy, N. , Maraspini, R. , Gorilak, P. , Varga, V. , Honigmann, A. , & Pigino, G. (2020). The molecular structure of mammalian primary cilia revealed by cryo‐electron tomography. Nature Structural & Molecular Biology, 27(12), 1115–1124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, S.‐E. , Nechipurenko, I. , & Christensen, S. T. (2023). Editorial: Signaling by primary cilia in development and disease. Frontiers in Cell and Developmental Biology, 11, 1186367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kitajima, S. , Takagi, A. , Inoue, T. , & Saga, Y. (2000). MesP1 and MesP2 are essential for the development of cardiac mesoderm. Development (Cambridge, England), 127(15), 3215–3226. [DOI] [PubMed] [Google Scholar]
- Knowles, M. R. , Daniels, L. A. , Davis, S. D. , Zariwala, M. A. , & Leigh, M. W. (2013). Primary ciliary dyskinesia. Recent advances in diagnostics, genetics, and characterization of clinical disease. American Journal of Respiratory and Critical Care Medicine, 188(8), 913–922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knowles, M. R. , Ostrowski, L. E. , Loges, N. T. , Hurd, T. , Leigh, M. W. , Huang, L. , Wolf, W. E. , Carson, J. L. , Hazucha, M. J. , Yin, W. , Davis, S. D. , Dell, S. D. , Ferkol, T. W. , Sagel, S. D. , Olivier, K. N. , Jahnke, C. , Olbrich, H. , Werner, C. , Raidt, J. , & Zariwala, M. A. (2013). Mutations in SPAG1 cause primary ciliary dyskinesia associated with defective outer and inner dynein arms. American Journal of Human Genetics, 93(4), 711–720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koefoed, K. , Veland, I. R. , Pedersen, L. B. , Larsen, L. A. , & Christensen, S. T. (2014). Cilia and coordination of signaling networks during heart development. Organogenesis, 10(1), 108–125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kramer‐Zucker, A. G. , Olale, F. , Haycraft, C. J. , Yoder, B. K. , Schier, A. F. , & Drummond, I. A. (2005). Cilia‐driven fluid flow in the zebrafish pronephros, brain and Kupffer's vesicle is required for normal organogenesis. Development (Cambridge, England), 132(8), 1907–1921. [DOI] [PubMed] [Google Scholar]
- Kuhn, E. N. , & Wu, S. M. (2010). Origin of cardiac progenitor cells in the developing and postnatal heart. Journal of Cellular Physiology, 225(2), 321–325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyndt, F. , Gueffet, J.‐P. , Probst, V. , Jaafar, P. , Legendre, A. , Le Bouffant, F. , Toquet, C. , Roy, E. , Mcgregor, L. , Lynch, S. A. , Newbury‐Ecob, R. , Tran, V. , Young, I. , Trochu, J.‐N. , Le Marec, H. , & Schott, J.‐J. (2007). Mutations in the gene encoding filamin A as a cause for familial cardiac valvular dystrophy. Circulation, 115(1), 40–49. [DOI] [PubMed] [Google Scholar]
- Kyun, M.‐L. , Kim, S.‐O. , Lee, H. G. , Hwang, J.‐A. , Hwang, J. , Soung, N.‐K. , Cha‐Molstad, H. , Lee, S. , Kwon, Y. T. , Kim, B. Y. , & Lee, K. H. (2020). Wnt3a stimulation promotes primary ciliogenesis through β‐catenin phosphorylation‐induced reorganization of centriolar satellites. Cell Reports, 30(5), 1447–1462. [DOI] [PubMed] [Google Scholar]
- Lahaye, S. , Lincoln, J. , & Garg, V. (2014). Genetics of valvular heart disease. Current Cardiology Reports, 16(6), 487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai, D. , Liu, X. , Forrai, A. , Wolstein, O. , Michalicek, J. , Ahmed, I. , Garratt, A. N. , Birchmeier, C. , Zhou, M. , Hartley, L. , Robb, L. , Feneley, M. P. , Fatkin, D. , & Harvey, R. P. (2010). Neuregulin 1 sustains the gene regulatory network in both trabecular and nontrabecular myocardium. Circulation Research, 107(6), 715–727. [DOI] [PubMed] [Google Scholar]
- Lapart, J.‐A. , Gottardo, M. , Cortier, E. , Duteyrat, J.‐L. , Augière, C. , Mangé, A. , Jerber, J. , Solassol, J. , Gopalakrishnan, J. , Thomas, J. , & Durand, B. (2019). Dzip1 and Fam92 form a ciliary transition zone complex with cell type specific roles in Drosophila . Elife, 8, e49307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larkins, C. E. , Bushey Long, A. , & Caspary, T. (2012). Defective Nodal and Cerl2 expression in the Arl13b(hnn) mutant node underlie its heterotaxia. Developmental Biology, 367(1), 15–24. [DOI] [PubMed] [Google Scholar]
- Lechtreck, K. F. (2015). IFT‐cargo interactions and protein transport in cilia. Trends in Biochemical Sciences, 40(12), 765–778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee, M.‐S. , Hwang, K.‐S. , Oh, H.‐W. , Ji‐Ae, K. , Kim, H.‐T. , Cho, H.‐S. , Lee, J.‐J. , Yeong Ko, J. , Choi, J.‐H. , Jeong, Y.‐M. , You, K.‐H. , Kim, J. , Park, D.‐S. , Nam, K.‐H. , Aizawa, S. , Kiyonari, H. , Shioi, G. , Park, J.‐H. , Zhou, W. , & Kim, C.‐H. (2015). IFT46 plays an essential role in cilia development. Developmental Biology, 400(2), 248–257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, J. , Miao, L. , Zhao, C. , Shaikh Qureshi, W. M. , Shieh, D. , Guo, H. , Lu, Y. , Hu, S. , Huang, A. , Zhang, L. , Cai, C.‐L. , Wan, L. Q. , Xin, H. , Vincent, P. , Singer, H. A. , Zheng, Y. , Cleaver, O. , Fan, Z.‐C. , & Wu, M. (2017). CDC42 is required for epicardial and pro‐epicardial development by mediating FGF receptor trafficking to the plasma membrane. Development, 144(9), 1635–1647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, X. , Lu, Q. , Peng, Y. , Geng, F. , Shao, X. , Zhou, H. , Cao, Y. , & Zhang, R. (2020). Primary cilia mediate Klf2‐dependant Notch activation in regenerating heart. Protein Cell, 11(6), 433–445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y. , Klena, N. T. , Gabriel, G. C. , Liu, X. , Kim, A. J. , Lemke, K. , Chen, Y. , Chatterjee, B. , Devine, W. , Damerla, R. R. , Chang, C. , Yagi, H. , San Agustin, J. T. , Thahir, M. , Anderton, S. , Lawhead, C. , Vescovi, A. , Pratt, H. , Morgan, J. , & Lo, C. W. (2015). Global genetic analysis in mice unveils central role for cilia in congenital heart disease. Nature, 521(7553), 520–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y. , Yagi, H. , Onuoha, E. O. , Damerla, R. R. , Francis, R. , Furutani, Y. , Tariq, M. , King, S M. , Hendricks, G. , Cui, C. , Saydmohammed, M. , Lee, D. M. , Zahid, M. , Sami, I. , Leatherbury, L. , Pazour, G. J. , Ware, S. M. , Nakanishi, T. , Goldmuntz, E. , & Lo, C. W. (2016). DNAH6 and its interactions with PCD genes in heterotaxy and primary ciliary dyskinesia. Plos Genetics, 12(2), e1005821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, C. , Cao, R. , Xu, Y. , Li, T. , Li, F. , Chen, S. , Xu, R. , & Sun, K. (2018). Rare copy number variants analysis identifies novel candidate genes in heterotaxy syndrome patients with congenital heart defects. Genome Medicine, 10(1), 40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, S. , Chen, W. , Zhan, Y. , Li, S. , Ma, X. , Ma, D. , Sheng, W. , & Huang, G. (2019). DNAH11 variants and its association with congenital heart disease and heterotaxy syndrome. Scientific Reports, 9(1), 6683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logan, M. , Pagán‐Westphal, S. M. , Smith, D. M. , Paganessi, L. , & Tabin, C. J. (1998). The transcription factor Pitx2 mediates situs‐specific morphogenesis in response to left‐right asymmetric signals. Cell, 94(3), 307–317. [DOI] [PubMed] [Google Scholar]
- Long, H. , & Huang, K. (2019). Transport of ciliary membrane proteins. Frontiers in Cell and Developmental Biology, 7, 381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lowe, L. A. , Supp, D. M. , Sampath, K. , Yokoyama, T. , Wright, C. V. E. , Potter, S. S , Overbeek, P. , & Kuehn, M. R. (1996). Conserved left‐right asymmetry of nodal expression and alterations in murine situs inversus. Nature, 381(6578), 158–161. [DOI] [PubMed] [Google Scholar]
- Mackay, C. E. , Leo, M. D. , Fernández‐Peña, C. , Hasan, R. , Yin, W. , Mata‐Daboin, A. , Bulley, S. , Gammons, J. , Mancarella, S. , & Jaggar, J. H. (2020). Intravascular flow stimulates PKD2 (polycystin‐2) channels in endothelial cells to reduce blood pressure. Elife, 9, e56655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahaffey, J. P. , Grego‐Bessa, J. , Liem, K. F. , & Anderson, K. V. (2013). Cofilin and Vangl2 cooperate in the initiation of planar cell polarity in the mouse embryo. Development (Cambridge, England), 140(6), 1262–1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malicki, J. J. , & Johnson, C. A. (2017). The cilium: Cellular antenna and central processing unit. Trends in Cell Biology, 27(2), 126–140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandalenakis, Z. , Giang, K. W. , Eriksson, P. , Liden, H. , Synnergren, M. , Wåhlander, H. , Fedchenko, M. , Rosengren, A. , & Dellborg, M. (2020). Survival in children with congenital heart disease: Have we reached a peak at 97%? Journal of the American Heart Association, 9(22), e017704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marques, S. , Borges, A. C. , Silva, A. C. , Freitas, S. , Cordenonsi, M. , & Belo, J. A. (2004). The activity of the Nodal antagonist Cerl‐2 in the mouse node is required for correct L/R body axis. Genes & Development, 18(19), 2342–2347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marszalek, J. R. , Ruiz‐Lozano, P. , Roberts, E. , Chien, K. R. , & Goldstein, L. S. B. (1999). Situs inversus and embryonic ciliary morphogenesis defects in mouse mutants lacking the KIF3A subunit of kinesin‐II. PNAS, 96(9), 5043–5048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- May, E. A. , Sroka, T. J. , & Mick, D. U. (2021). Phosphorylation and ubiquitylation regulate protein trafficking, signaling, and the biogenesis of primary cilia. Frontiers in Cell and Developmental Biology, 9, 664279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mcglashan, S. R. , Jensen, C. G. , & Poole, C. A. (2006). Localization of extracellular matrix receptors on the chondrocyte primary cilium. Journal of Histochemistry and Cytochemistry, 54(9), 1005–1014. [DOI] [PubMed] [Google Scholar]
- Mcgrath, J. , Somlo, S. , Makova, S. , Tian, X. , & Brueckner, M. (2003). Two populations of node monocilia initiate left‐right asymmetry in the mouse. Cell, 114(1), 61–73. [DOI] [PubMed] [Google Scholar]
- Merklin, R. J. , & Varano, N. R. (1963). SITUS INVERSUS AND CARDIAC DEFECTS: A study of 111 cases of reversed asymmetry. The Journal of Thoracic and Cardiovascular Surgery, 45(3), 334–342. [PubMed] [Google Scholar]
- Mill, P. , Christensen, S. T. , & Pedersen, L. B. (2023). Primary cilia as dynamic and diverse signalling hubs in development and disease. Nature Reviews Genetics, 24(7), 421–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller, K. A. , Ah‐Cann, C. J. , Welfare, M. F. , Tan, T. Y. , Pope, K. , Caruana, G. , Freckmann, M.‐L. , Savarirayan, R. , Bertram, J. F. , Dobbie, M. S. , Bateman, J. F. , & Farlie, P. G. (2013). Cauli: A mouse strain with an Ift140 mutation that results in a skeletal ciliopathy modelling Jeune syndrome. Plos Genetics, 9(8), e1003746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mirvis, M. , Stearns, T. , & James Nelson, W. (2018). Cilium structure, assembly, and disassembly regulated by the cytoskeleton. Biochemical Journal, 475(14), 2329–2353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchison, H. M. , Schmidts, M. , Loges, N. T. , Freshour, J. , Dritsoula, A. , Hirst, R. A. , O'callaghan, C. , Blau, H. , Al Dabbagh, M. , Olbrich, H. , Beales, P. L. , Yagi, T. , Mussaffi, H. , Chung, E. M. K. , Omran, H. , & Mitchell, D. R. (2012). Mutations in axonemal dynein assembly factor DNAAF3 cause primary ciliary dyskinesia. Nature Genetics, 44(4), 381–389, s1–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore, K. , Fulmer, D. , Guo, L. , Koren, N. , Glover, J. , Moore, R. , Gensemer, C. , Beck, T. , Morningstar, J. , Stairley, R. , & Norris, R. A. (2021). PDGFRα: Expression and function during mitral valve morphogenesis. Journal of Cardiovascular Development and Disease, 8(3), 28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morningstar, J. E. , Gensemer, C. , Moore, R. , Fulmer, D. , Beck, T. C. , Wang, C. , Moore, K. , Guo, L. , Sieg, F. , Nagata, Y. , Bertrand, P. , Spampinato, R. A. , Glover, J. , Poelzing, S. , Gourdie, R. G. , Watts, K. , Richardson, W. J. , Levine, R. A. , Borger, M. A. , & Norris, R. A. (2021). Mitral valve prolapse induces regionalized myocardial fibrosis. Journal of the American Heart Association, 10(24), e022332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morton, S. U. , Quiat, D. , Seidman, J. G. , & Seidman, C. E. (2022). Genomic frontiers in congenital heart disease. Nature Reviews Cardiology, 19(1), 26–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Münsterberg, A. , & Yue, Q. (2008). Cardiac progenitor migration and specification: The dual function of Wnts. Cell Adhesion & Migration, 2(2), 74–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murcia, N. S. , Richards, W. G. , Yoder, B. K. , Mucenski, M. L. , Dunlap, J. R. , & Woychik, R. P. (2000). The Oak Ridge Polycystic Kidney (orpk) disease gene is required for left–right axis determination. Development (Cambridge, England), 127(11), 2347–2355. [DOI] [PubMed] [Google Scholar]
- Myklebust, R. , Engedal, H. , Saetersdal, T. S. , & Ulstein, M. (1977). Primary 9 + 0 cilia in the embryonic and the adult human heart. Anatomy and Embryology, 151(2), 127–139. [DOI] [PubMed] [Google Scholar]
- Mykytyn, K. , & Askwith, C. (2017). G‐protein‐coupled receptor signaling in cilia. Cold Spring Harbor Perspectives in Biology, 9(9), a028183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nachury, M. V. (2014). How do cilia organize signalling cascades? Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 369(1650), 20130465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura, T. , Saito, D. , Kawasumi, A. , Shinohara, K. , Asai, Y. , Takaoka, K. , Dong, F. , Takamatsu, A. , Belo, J. A. , Mochizuki, A. , & Hamada, H. (2012). Fluid flow and interlinked feedback loops establish left‐right asymmetric decay of Cerl2 mRNA. Nature Communications, 3, 1322. [DOI] [PubMed] [Google Scholar]
- Nakhleh, N. , Francis, R. , Giese, R. A. , Tian, X. , Li, Y. , Zariwala, M. A. , Yagi, H. , Khalifa, O. , Kureshi, S. , Chatterjee, B. , Sabol, S. L. , Swisher, M. , Connelly, P. S. , Daniels, M. P. , Srinivasan, A. , Kuehl, K. , Kravitz, N. , Burns, K. , Sami, I. , & Lo, C. W. (2012). High prevalence of respiratory ciliary dysfunction in congenital heart disease patients with heterotaxy. Circulation, 125(18), 2232–2242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narasimhan, V. , & Roy, S. (2015). Cilia: Organelles at the heart of heart disease. Current Biology, 25(13), R559–R562. [DOI] [PubMed] [Google Scholar]
- Nauli, S. M. , Rossetti, S. , Kolb, R. J. , Alenghat, F. J. , Consugar, M. B. , Harris, P. C. , Ingber, D. E. , Loghman‐Adham, M. , & Zhou, J. (2006). Loss of polycystin‐1 in human cyst‐lining epithelia leads to ciliary dysfunction. Journal of the American Society of Nephrology, 17(4), 1015–1025. [DOI] [PubMed] [Google Scholar]
- Niederlova, V. , Modrak, M. , Tsyklauri, O. , Huranova, M. , & Stepanek, O. (2019). Meta‐analysis of genotype‐phenotype associations in Bardet‐Biedl syndrome uncovers differences among causative genes. Human Mutation, 40(11), 2068–2087. [DOI] [PubMed] [Google Scholar]
- Nishimura, Y. , Kasahara, K. , Shiromizu, T. , Watanabe, M. , & Inagaki, M. (2018). Primary cilia as signaling hubs in health and disease. Advanced Science, 6, 1801138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonaka, S. , Shiratori, H. , Saijoh, Y. , & Hamada, H. (2002). Determination of left‐right patterning of the mouse embryo by artificial nodal flow. Nature, 418(6893), 96–99. [DOI] [PubMed] [Google Scholar]
- Nonaka, S. , Tanaka, Y. , Okada, Y. , Takeda, S. , Harada, A. , Kanai, Y. , Kido, M. , & Hirokawa, N. (1998). Randomization of left‐right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell, 95(6), 829–837. [DOI] [PubMed] [Google Scholar]
- Nonaka, S. , Yoshiba, S. , Watanabe, D. , Ikeuchi, S. , Goto, T. , Marshall, W. F. , & Hamada, H. (2005). De novo formation of left‐right asymmetry by posterior tilt of nodal cilia. Plos Biology, 3(8), e268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noone, P. G. , Leigh, M. W. , Sannuti, A. , Minnix, S. L. , Carson, J. L. , Hazucha, M. , Zariwala, M. A. , & Knowles, M. R. (2004). Primary ciliary dyskinesia: Diagnostic and phenotypic features. American Journal of Respiratory and Critical Care Medicine, 169(4), 459–467. [DOI] [PubMed] [Google Scholar]
- Norris, D. P. , Brennan, J. , Bikoff, E. K. , & Robertson, E. J. (2002). The Foxh1‐dependent autoregulatory enhancer controls the level of Nodal signals in the mouse embryo. Development (Cambridge, England), 129(14), 3455–3468. [DOI] [PubMed] [Google Scholar]
- Norris, D. P. , & Robertson, E. J. (1999). Asymmetric and node‐specific nodal expression patterns are controlled by two distinct cis‐acting regulatory elements. Genes & Development, 13(12), 1575–1588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nöthe‐Menchen, T. , Wallmeier, J. , Pennekamp, P. , Höben, I. M. , Olbrich, H. , Loges, N. T. , Raidt, J. , Dougherty, G. W. , Hjeij, R. , Dworniczak, B. , Omran, H. , Amirav, I. , Biebach, L. , Fabricius, D. , Griese, M. , Große‐Onnebrink, J. , Häffner, K. , Hector, A. , & Zariwala, M. A. (2019). Randomization of left‐right asymmetry and congenital heart defects: The role of DNAH5 in humans and mice. Circulation: Genomic and Precision Medicine, 12, e002686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'donnell, A. , & Yutzey, K. E. (2020). Mechanisms of heart valve development and disease. Development (Cambridge, England), 147(13), dev183020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okada, Y. , Nonaka, S. , Tanaka, Y. , Saijoh, Y. , Hamada, H. , & Hirokawa, N. (1999). Abnormal nodal flow precedes situs inversus in iv and inv mice. Molecular Cell, 4(4), 459–468. [DOI] [PubMed] [Google Scholar]
- Oki, S. , Kitajima, K. , Marques, S. , Belo, J. A. , Yokoyama, T. , Hamada, H. , & Meno, C. (2009). Reversal of left‐right asymmetry induced by aberrant Nodal signaling in the node of mouse embryos. Development (Cambridge, England), 136(23), 3917–3925. [DOI] [PubMed] [Google Scholar]
- Olson, A. J. , Krentz, A. D. , Finta, K. M. , Okorie, U. C. , & Haws, R. M. (2019). Thoraco‐abdominal abnormalities in Bardet‐Biedl Syndrome: Situs inversus and heterotaxy. Journal of Pediatrics, 204, 31–37. [DOI] [PubMed] [Google Scholar]
- Olstad, E. W. , Ringers, C. , Hansen, J. N. , Wens, A. , Brandt, C. , Wachten, D. , Yaksi, E. , & Jurisch‐Yaksi, N. (2019). Ciliary beating compartmentalizes cerebrospinal fluid flow in the brain and regulates ventricular development. Current Biology, 29(2), 229–241.e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onoufriadis, A. , Shoemark, A. , Munye, M. M. , James, C. T. , Schmidts, M. , Patel, M. , Rosser, E. M. , Bacchelli, C. , Beales, P. L. , Scambler, P. J. , Hart, S. L. , Danke‐Roelse, J. E. , Sloper, J. J. , Hull, S. , Hogg, C. , Emes, R. D. , Pals, G. , Moore, A. T. , Chung, E. M. K. , & Mitchison, H. M. (2014). Combined exome and whole‐genome sequencing identifies mutations in ARMC4 as a cause of primary ciliary dyskinesia with defects in the outer dynein arm. Journal of Medical Genetics, 51(1), 61–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pala, R. , Alomari, N. , & Nauli, S. (2017). Primary cilium‐dependent signaling mechanisms. International Journal of Molecular Sciences, 18(11), 2272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park, T. J. , Mitchell, B. J. , Abitua, P. B. , Kintner, C. , & Wallingford, J. B. (2008). Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells. Nature Genetics, 40(7), 871–879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel, K. , & Smith, N. J. (2023). Primary cilia, A‐kinase anchoring proteins and constitutive activity at the orphan G protein‐coupled receptor GPR161: A tale about a tail. British Journal of Pharmacology, 48, 560. [DOI] [PubMed] [Google Scholar]
- Patel, S. S. , & Burns, T. L. (2013). Nongenetic risk factors and congenital heart defects. Pediatric Cardiology, 34(7), 1535–1555. [DOI] [PubMed] [Google Scholar]
- Peng, J. , Meng, Z. , Zhou, S. , Zhou, Y. , Wu, Y. , Wang, Q. , Wang, J. , & Sun, K. (2019). The non‐genetic paternal factors for congenital heart defects: A systematic review and meta‐analysis. Clinical Cardiology, 42(7), 684–691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pennekamp, P. , Karcher, C. , Fischer, A. , Schweickert, A. , Skryabin, B. , Horst, J. , Blum, M. , & Dworniczak, B. (2002). The ion channel polycystin‐2 is required for left‐right axis determination in mice. Current Biology, 12(11), 938–943. [DOI] [PubMed] [Google Scholar]
- Peralta, M. , Ortiz Lopez, L. , Jerabkova, K. , Lucchesi, T. , Vitre, B. , Han, D. , Guillemot, L. , Dingare, C. , Sumara, I. , Mercader, N. , Lecaudey, V. , Delaval, B. , Meilhac, S M. , & Vermot, J. (2020). Intraflagellar transport complex B proteins regulate the hippo effector Yap1 during cardiogenesis. Cell Reports, 32(3), 107932. [DOI] [PubMed] [Google Scholar]
- Person, A. D. , Klewer, S. E. , & Runyan, R. B. (2005). Cell biology of cardiac cushion development. International Review of Cytology, 243, 287–335. [DOI] [PubMed] [Google Scholar]
- Piedra, M. E , Icardo, J. M. , Albajar, M. , Rodriguez‐Rey, J. C. , & Ros, M. A. (1998). Pitx2 participates in the late phase of the pathway controlling left‐right asymmetry. Cell, 94(3), 319–324. [DOI] [PubMed] [Google Scholar]
- Pierpont, M. E. , Brueckner, M. , Chung, W. K. , Garg, V. , Lacro, R. V. , McGuire, A. L. , Mital, S. , Priest, J. R. , Pu, W. T. , Roberts, A. , Ware, S. M. , Gelb, B. D. , & Russell, M. W. (2018). Genetic basis for congenital heart disease: Revisited: A scientific statement from the American Heart Association. Circulation, 138(21), e653–e711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pigino, G. , Geimer, S. , Lanzavecchia, S. , Paccagnini, E. , Cantele, F. , Diener, D. R. , Rosenbaum, J. L. , & Lupetti, P. (2009). Electron‐tomographic analysis of intraflagellar transport particle trains in situ. Journal of Cell Biology, 187(1), 135–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu, X. , Harmelink, C. , & Baldwin, H. S (2022). Endocardial‐myocardial interactions during early cardiac differentiation and trabeculation. Frontiers in Cardiovascular Medicine, 9, 857581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahman, E. , Niaz, F. A. , Al‐Suwaida, A. , Nahrir, S. , Bashir, M. , Rahman, H. , & Hammad, D. (2009). Analysis of causes of mortality in patients with autosomal dominant polycystic kidney disease: A single center study. Saudi Journal of Kidney Diseases and Transplantation, 20(5), 806–810. [PubMed] [Google Scholar]
- Rao Damerla, R. , Gabriel, G. C. , Li, Y. , Klena, N. T. , Liu, X. , Chen, Y. , Cui, C. , Pazour, G. J. , & Lo, C. W. (2014). Role of cilia in structural birth defects: Insights from ciliopathy mutant mouse models. Birth Defects Research. Part C, Embryo Today, 102(2), 115–125. [DOI] [PubMed] [Google Scholar]
- Reiter, J. F. , & Leroux, M. R. (2017). Genes and molecular pathways underpinning ciliopathies. Nature Reviews Molecular Cell Biology, 18(9), 533–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridge, L. A. , Mitchell, K. , Al‐Anbaki, A. , Shaikh Qureshi, W. M. , Stephen, L. A. , Tenin, G. , Lu, Y. , Lupu, I.‐E. , Clowes, C. , Robertson, A. , Barnes, E. , Wright, J. A. , Keavney, B. , Ehler, E. , Lovell, S. C. , Kadler, K. E. , & Hentges, K. E. (2017). Non‐muscle myosin IIB (Myh10) is required for epicardial function and coronary vessel formation during mammalian development. Plos Genetics, 13(10), e1007068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rieder, C. L. , Jensen, C. G. , & Jensen, L. C. W. (1979). The resorption of primary cilia during mitosis in a vertebrate (PtK1) cell line. Journal of Ultrastructure Research, 68(2), 173–185. [DOI] [PubMed] [Google Scholar]
- Rosenbaum, J. L. , & Witman, G. B. (2002). Intraflagellar transport. Nature Reviews Molecular Cell Biology, 3(11), 813–825. [DOI] [PubMed] [Google Scholar]
- Ross, A. J. , May‐Simera, H. , Eichers, E. R. , Kai, M. , Hill, J. , Jagger, D. J. , Leitch, C. C. , Chapple, J. P. , Munro, P. M. , Fisher, S. , Tan, P. L. , Phillips, H. M. , Leroux, M. R. , Henderson, D. J. , Murdoch, J. N. , Copp, A. J. , Eliot, M.‐M. , Lupski, J. R. , Kemp, D. T. , & Beales, P. L. (2005). Disruption of Bardet‐Biedl syndrome ciliary proteins perturbs planar cell polarity in vertebrates. Nature Genetics, 37(10), 1135–1140. [DOI] [PubMed] [Google Scholar]
- Saba, T. G. , Geddes, G. C. , Ware, S. M. , Schidlow, D. N. , Del Nido, P. J. , Rubalcava, N. S. , Gadepalli, S. K. , Stillwell, T. , Griffiths, A. , Bennett Murphy, L. M. , Barber, A. T. , Leigh, M. W. , Sabin, N. , & Shapiro, A. J. (2022). A multi‐disciplinary, comprehensive approach to management of children with heterotaxy. Orphanet Journal of Rare Diseases, 17(1), 351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sáenz‐Ponce, N. , Santillana‐Ortiz, J.‐D. , & Del Pino, E. M. (2012). The gastrocoel roof plate in embryos of different frogs. Differentiation, 83(2), S62–S66. [DOI] [PubMed] [Google Scholar]
- Saga, Y. , Hata, N. , Kobayashi, S. , Magnuson, T. , Seldin, M. F. , & Taketo, M. M. (1996). MesP1: A novel basic helix‐loop‐helix protein expressed in the nascent mesodermal cells during mouse gastrulation. Development (Cambridge, England), 122(9), 2769–2778. [DOI] [PubMed] [Google Scholar]
- Saga, Y. , Miyagawa‐Tomita, S. , Takagi, A. , Kitajima, S. , Miyazaki, J.‐I. , & Inoue, T. (1999). MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development (Cambridge, England), 126(15), 3437–3447. [DOI] [PubMed] [Google Scholar]
- Saijoh, Y. , Oki, S. , Ohishi, S. , & Hamada, H. (2003). Left‐right patterning of the mouse lateral plate requires nodal produced in the node. Developmental Biology, 256(1), 161–173. [DOI] [PubMed] [Google Scholar]
- Sakuma, R. , Ohnishi, Y.‐I. , Meno, C. , Fujii, H. , Juan, H. , Takeuchi, J. , Ogura, T. , Li, E. , Miyazono, K. , & Hamada, H. (2002). Inhibition of Nodal signalling by Lefty mediated through interaction with common receptors and efficient diffusion. Genes to Cells, 7(4), 401–412. [DOI] [PubMed] [Google Scholar]
- Sampaio, P. , Ferreira, R. R. , Guerrero, A. , Pintado, P. , Tavares, B. , Amaro, J. , Smith, A. A. , Montenegro‐Johnson, T. , Smith, D. J. , & Lopes, S. S. (2014). Left‐right organizer flow dynamics: How much cilia activity reliably yields laterality? Developmental Cell, 29(6), 716–728. [DOI] [PubMed] [Google Scholar]
- Samsa, L. A. , Givens, C. , Tzima, E. , Stainier, D. Y. R. , Qian, L. , & Liu, J. (2015). Cardiac contraction activates endocardial Notch signaling to modulate chamber maturation in zebrafish. Development (Cambridge, England), 142(23), 4080–4091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samsa, L. A. , Yang, B. , & Liu, J. (2013). Embryonic cardiac chamber maturation: Trabeculation, conduction, and cardiomyocyte proliferation. American Journal of Medical Genetics Part C, Seminars in Medical Genetics, 163(3), 157–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sang, L. , Miller, J. J. , Corbit, K. C. , Giles, R. H. , Brauer, M. J. , Otto, E. A. , Baye, L. M. , Wen, X. , Scales, S. J. , Kwong, M. , Huntzicker, E. G. , Sfakianos, M. K. , Sandoval, W. , Bazan, J. F , Kulkarni, P. , Garcia‐Gonzalo, F. R. , Seol, A. D. , O'toole, J. F. , Held, S. , & Jackson, P. K. (2011). Mapping the NPHP‐JBTS‐MKS protein network reveals ciliopathy disease genes and pathways. Cell, 145(4), 513–528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satir, P. , & Christensen, S. T. (2007). Overview of structure and function of mammalian cilia. Annual Review of Physiology, 69, 377–400. [DOI] [PubMed] [Google Scholar]
- Savolainen, S. M. , Foley, J. F. , & Elmore, S. A. (2009). Histology atlas of the developing mouse heart with emphasis on E11.5 to E18.5. Toxicologic Pathology, 37(4), 395–414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schleiffarth, J. R. , Person, A. D. , Martinsen, B. J. , Sukovich, D. J. , Neumann, A. , Baker, C. V. H. , Lohr, J. L. , Cornfield, D. N. , Ekker, S. C. , & Petryk, A. (2007). Wnt5a is required for cardiac outflow tract septation in mice. Pediatric Research, 61(4), 386–391. [DOI] [PubMed] [Google Scholar]
- Schmid, F. M. , Schou, K. B. , Vilhelm, M. J. , Holm, M. S. , Breslin, L. , Farinelli, P. , Larsen, L. A. , Andersen, J. S. , Pedersen, L. B. , & Christensen, S. T. (2018). IFT20 modulates ciliary PDGFRα signaling by regulating the stability of Cbl E3 ubiquitin ligases. Journal of Cell Biology, 217(1), 151–161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider, L. , Clement, C. A. , Teilmann, S. C. , Pazour, G. J. , Hoffmann, E. K. , Satir, P. , & Christensen, S. T. (2005). PDGFRalphaalpha signaling is regulated through the primary cilium in fibroblasts. Current Biology, 15(20), 1861–1866. [DOI] [PubMed] [Google Scholar]
- Schottenfeld, J. , Sullivan‐Brown, J. , & Burdine, R. D. (2007). Zebrafish curly up encodes a Pkd2 ortholog that restricts left‐side‐specific expression of southpaw. Development (Cambridge, England), 134(8), 1605–1615. [DOI] [PubMed] [Google Scholar]
- Schweickert, A. , Vick, P. , Getwan, M. , Weber, T. , Schneider, I. , Eberhardt, M. , Beyer, T. , Pachur, A. , & Blum, M. (2010). The nodal inhibitor Coco is a critical target of leftward flow in Xenopus. Current Biology, 20(8), 738–743. [DOI] [PubMed] [Google Scholar]
- Schweickert, A. , Weber, T. , Beyer, T. , Vick, P. , Bogusch, S. , Feistel, K. , & Blum, M. (2007). Cilia‐driven leftward flow determines laterality in Xenopus. Current Biology, 17(1), 60–66. [DOI] [PubMed] [Google Scholar]
- Seo, J. W. , Brown, N. A. , Ho, S. Y. , & Anderson, R. H. (1992). Abnormal laterality and congenital cardiac anomalies. Relations of visceral and cardiac morphologies in the iv/iv mouse. Circulation, 86(2), 642–650. [DOI] [PubMed] [Google Scholar]
- Shah, A. S. , Ben‐Shahar, Y. , Moninger, T. O. , Kline, J. N. , & Welsh, M. J. (2009). Motile cilia of human airway epithelia are chemosensory. Science, 325(5944), 1131–1134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shapiro, A. J. , Davis, S. D. , Ferkol, T. , Dell, S. D. , Rosenfeld, M. , Olivier, K. N. , Sagel, S. D. , Milla, C. , Zariwala, M. A. , Wolf, W. , Carson, J. L. , Hazucha, M. J. , Burns, K. , Robinson, B. , Knowles, M. R. , & Leigh, M. W. (2014). Laterality defects other than situs inversus totalis in primary ciliary dyskinesia: Insights into situs ambiguus and heterotaxy. Chest, 146(5), 1176–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinohara, K. , Chen, D. , Nishida, T. , Misaki, K. , Yonemura, S. , & Hamada, H. (2015). Absence of radial spokes in mouse node cilia is required for rotational movement but confers ultrastructural instability as a trade‐off. Developmental Cell, 35(2), 236–246. [DOI] [PubMed] [Google Scholar]
- Shinohara, K. , Kawasumi, A. , Takamatsu, A. , Yoshiba, S. , Botilde, Y. , Motoyama, N. , Reith, W. , Durand, B. , Shiratori, H. , & Hamada, H. (2012). Two rotating cilia in the node cavity are sufficient to break left‐right symmetry in the mouse embryo. Nature Communications, 3, 622. [DOI] [PubMed] [Google Scholar]
- Shiratori, H. , Sakuma, R. , Watanabe, M. , Hashiguchi, H. , Mochida, K. , Sakai, Y. , Nishino, J. , Saijoh, Y. , Whitman, M. , & Hamada, H. (2001). Two‐step regulation of left‐right asymmetric expression of Pitx2: Initiation by nodal signaling and maintenance by Nkx2. Molecular Cell, 7(1), 137–149. [DOI] [PubMed] [Google Scholar]
- Shylo, N. A. , Emmanouil, E. , Ramrattan, D. , & Weatherbee, S. D. (2020). Loss of ciliary transition zone protein TMEM107 leads to heterotaxy in mice. Developmental Biology, 460(2), 187–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slough, J. , Cooney, L. , & Brueckner, M. (2008). Monocilia in the embryonic mouse heart suggest a direct role for cilia in cardiac morphogenesis. Developmental Dynamics, 237(9), 2304–2314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Solomon, G. M. , Francis, R. , Chu, K. K. , Birket, S. E. , Gabriel, G. , Trombley, J. E. , Lemke, K. L. , Klena, N. , Turner, B. , Tearney, G. J. , Lo, C. W. , & Rowe, S. M. (2017). Assessment of ciliary phenotype in primary ciliary dyskinesia by micro‐optical coherence tomography. JCI Insight, 2(5), e91702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song, H. , Hu, J. , Chen, W. , Elliott, G. , Andre, P. , Gao, B. , & Yang, Y. (2010). Planar cell polarity breaks bilateral symmetry by controlling ciliary positioning. Nature, 466(7304), 378–382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefanovic, S. , Etchevers, H. C. , & Zaffran, S. (2021). Outflow tract formation‐embryonic origins of conotruncal congenital heart disease. Journal of Cardiovascular Development and Disease, 8(4), 42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens, J. , Ermakov, A. , Braganca, J. , Hilton, H. , Underhill, P. , Bhattacharya, S. , Brown, N. A. , & Norris, D. P. (2010). Analysis of the asymmetrically expressed Ablim1 locus reveals existence of a lateral plate Nodal‐independent left sided signal and an early, left‐right independent role for nodal flow. BMC Developmental Biology, 10(1), 54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugrue, K. F. , & Zohn, I. E. (2017). Mechanism for generation of left isomerism in Ccdc40 mutant embryos. PLoS ONE, 12(2), e0171180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sulik, K. , Dehart, D. B. , Inagaki, T. , Carson, J. L. , Vrablic, T. , Gesteland, K. , & Schoenwolf, G. C. (1994). Morphogenesis of the murine node and notochordal plate. Developmental Dynamics, 201(3), 260–278. [DOI] [PubMed] [Google Scholar]
- Sun, S. , Fisher, R. L. , Bowser, S. S. , Pentecost, B. T. , & Sui, H. (2019). Three‐dimensional architecture of epithelial primary cilia. Proceedings of the National Academy of Sciences, 116(19), 9370–9379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takao, D. , Nemoto, T. , Abe, T. , Kiyonari, H. , Kajiura‐Kobayashi, H. , Shiratori, H. , & Nonaka, S. (2013). Asymmetric distribution of dynamic calcium signals in the node of mouse embryo during left‐right axis formation. Developmental Biology, 376(1), 23–30. [DOI] [PubMed] [Google Scholar]
- Takeda, S. , Yonekawa, Y. , Tanaka, Y. , Okada, Y. , Nonaka, S. , & Hirokawa, N. (1999). Left‐right asymmetry and kinesin superfamily protein KIF3A: New insights in determination of laterality and mesoderm induction by kif3A‐/‐ mice analysis. Journal of Cell Biology, 145(4), 825–836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan, C. M. J. , & Lewandowski, A. J. (2020). The transitional heart: From early embryonic and fetal development to neonatal life. Fetal Diagnosis and Therapy, 47(5), 373–386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan, S. Y. , Rosenthal, J. , Zhao, X. Q. , Francis, R. J. , Chatterjee, B. , Sabol, S. L. , Linask, K. L. , Bracero, L. , Connelly, P. S. , Daniels, M. P. , Yu, Q. , Omran, H. , Leatherbury, L. , & Lo, C. W. (2007). Heterotaxy and complex structural heart defects in a mutant mouse model of primary ciliary dyskinesia. Journal of Clinical Investigation, 117(12), 3742–3752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarkar, A. , Loges, N. T. , Slagle, C. E. , Francis, R. , Dougherty, G. W. , Tamayo, J. V. , Shook, B. , Cantino, M. , Schwartz, D. , Jahnke, C. , Olbrich, H. , Werner, C. , Raidt, J. , Pennekamp, P. , Abouhamed, M. , Hjeij, R. , Köhler, G. , Griese, M. , Li, Y. , & Omran, H. (2013). DYX1C1 is required for axonemal dynein assembly and ciliary motility. Nature Genetics, 45(9), 995–1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taschner, M. , & Lorentzen, E. (2016). The intraflagellar transport machinery. Cold Spring Harbor Perspectives in Biology, 8(10), a028092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ta‐Shma, A. , Hjeij, R. , Perles, Z. , Dougherty, G. W. , Abu Zahira, I. , Letteboer, S. J. F. , Antony, D. , Darwish, A. , Mans, D. A. , Spittler, S. , Edelbusch, C. , Cindrić, S. , Nöthe‐Menchen, T. , Olbrich, H. , Stuhlmann, F. , Aprea, I. , Pennekamp, P. , Loges, N. T. , Breuer, O. , & Omran, H. (2018). Homozygous loss‐of‐function mutations in MNS1 cause laterality defects and likely male infertility. Plos Genetics, 14(8), e1007602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tasouri, E. , & Tucker, K. L. (2011). Primary cilia and organogenesis: Is Hedgehog the only sculptor? Cell and Tissue Research, 345(1), 21–40. [DOI] [PubMed] [Google Scholar]
- Toomer, K. A. , Fulmer, D. , Guo, L. , Drohan, A. , Peterson, N. , Swanson, P. , Brooks, B. , Mukherjee, R. , Body, S. , Lipschutz, J. H. , Wessels, A. , & Norris, R. A. (2017). A role for primary cilia in aortic valve development and disease. Developmental Dynamics, 246(8), 625–634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toomer, K. A. , Yu, M. , Fulmer, D. , Guo, L. , Moore, K. S. , Moore, R. , Drayton, K. L. D. , Glover, J. , Peterson, N. , Ramos‐Ortiz, S. , Drohan, A. , Catching, B. J. , Stairley, R. , Wessels, A. , Lipschutz, J. H. , Delling, F. N. , Jeunemaitre, X. , Dina, C. , Collins, R. L. , & Norris, R. A. (2019). Primary cilia defects causing mitral valve prolapse. Science Translational Medicine, 11(493), eaax0290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsiairis, C. D. , & Mcmahon, A. P. (2009). An Hh‐dependent pathway in lateral plate mesoderm enables the generation of left/right asymmetry. Current Biology, 19(22), 1912–1917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Der Heiden, K. , Groenendijk, B. C. W. , Hierck, B. P. , Hogers, B. , Koerten, H. K. , Mommaas, A. M , Gittenberger‐De Groot, A. C. , & Poelmann, R. E. (2006). Monocilia on chicken embryonic endocardium in low shear stress areas. Developmental Dynamics, 235(1), 19–28. [DOI] [PubMed] [Google Scholar]
- Van Der Linde, D. , Konings, E. E. M. , Slager, M. A. , Witsenburg, M. , Helbing, W. A. , Takkenberg, J. J. M. , & Roos‐Hesselink, J. W. (2011). Birth prevalence of congenital heart disease worldwide: A systematic review and meta‐analysis. Journal of the American College of Cardiology, 58(21), 2241–2247. [DOI] [PubMed] [Google Scholar]
- Vasquez, S. S. V. , Van Dam, J. , & Wheway, G. (2021). An updated SYSCILIA gold standard (SCGSv2) of known ciliary genes, revealing the vast progress that has been made in the cilia research field. Molecular Biology of the Cell, 32(22), br13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verleyen, D. , Luyten, F. P. , & Tylzanowski, P. (2014). Orphan G‐protein coupled receptor 22 (Gpr22) regulates cilia length and structure in the zebrafish Kupffer's vesicle. PLoS ONE, 9(10), e110484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vierkotten, J. , Dildrop, R. , Peters, T. , Wang, B. , & RüTher, U. (2007). Ftm is a novel basal body protein of cilia involved in Shh signalling. Development (Cambridge, England), 134(14), 2569–2577. [DOI] [PubMed] [Google Scholar]
- Wallingford, J. B. , & Mitchell, B. (2011). Strange as it may seem: The many links between Wnt signaling, planar cell polarity, and cilia. Genes & Development, 25(3), 201–213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallmeier, J. , Shiratori, H. , Dougherty, G. W. , Edelbusch, C. , Hjeij, R. , Loges, N. T. , Menchen, T. , Olbrich, H. , Pennekamp, P. , Raidt, J. , Werner, C. , Minegishi, K. , Shinohara, K. , Asai, Y. , Takaoka, K. , Lee, C. , Griese, M. , Memari, Y. , Durbin, R. , & Omran, H. (2016). TTC25 deficiency results in defects of the outer dynein arm docking machinery and primary ciliary dyskinesia with left‐right body asymmetry randomization. American Journal of Human Genetics, 99(2), 460–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang, C. , Low, W.‐C. , Liu, A. , & Wang, B. (2013). Centrosomal protein DZIP1 regulates Hedgehog signaling by promoting cytoplasmic retention of transcription factor GLI3 and affecting ciliogenesis. Journal of Biological Chemistry, 288(41), 29518–29529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Washington Smoak, I. , Byrd, N. A. , Abu‐Issa, R. , Goddeeris, M. M. , Anderson, R. , Morris, J. , Yamamura, K. , Klingensmith, J. , & Meyers, E. N. (2005). Sonic hedgehog is required for cardiac outflow tract and neural crest cell development. Developmental Biology, 283(2), 357–372. [DOI] [PubMed] [Google Scholar]
- Watanabe, D. , Saijoh, Y. , Nonaka, S. , Sasaki, G. , Ikawa, Y. , Yokoyama, T. , & Hamada, H. (2003). The left‐right determinant Inversin is a component of node monocilia and other 9+0 cilia. Development (Cambridge, England), 130(9), 1725–1734. [DOI] [PubMed] [Google Scholar]
- Willaredt, M. A. , Gorgas, K. , Gardner, H. A. R. , & Tucker, K. L. (2012). Multiple essential roles for primary cilia in heart development. Cilia, 1(1), 23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson, P. D. , Hreniuk, D. , & Gabow, P. A. (1992). Abnormal extracellular matrix and excessive growth of human adult polycystic kidney disease epithelia. Journal of Cellular Physiology, 150(2), 360–369. [DOI] [PubMed] [Google Scholar]
- Wu, C. , Yang, M. , Li, J. , Wang, C. , Cao, T. , Tao, K. , & Wang, B. (2014). Talpid3‐binding centrosomal protein Cep120 is required for centriole duplication and proliferation of cerebellar granule neuron progenitors. PLoS ONE, 9(9), e107943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu, G. , Markowitz, G. S. , Li, L. , D'agati, V. D. , Factor, S. M. , Geng, L. , Tibara, S. , Tuchman, J. , Cai, Y. , Hoon Park, J. , Van Adelsberg, J. , Hou, H. , Kucherlapati, R. , Edelmann, W. , & Somlo, S. (2000). Cardiac defects and renal failure in mice with targeted mutations in Pkd2. Nature Genetics, 24(1), 75–78. [DOI] [PubMed] [Google Scholar]
- Wu, M. (2018). Mechanisms of trabecular formation and specification during cardiogenesis. Pediatric Cardiology, 39(6), 1082–1089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xia, H. , Huang, X. , Deng, S. , Xu, H. , Yang, Y. , Liu, X. , Yuan, L. , & Deng, H. (2021). DNAH11 compound heterozygous variants cause heterotaxy and congenital heart disease. PLoS ONE, 16(6), e0252786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, C. , Rossetti, S. , Jiang, L. , Harris, P. C. , Brown‐Glaberman, U. , Wandinger‐Ness, A. , Bacallao, R. , & Alper, S. L. (2007). Human ADPKD primary cyst epithelial cells with a novel, single codon deletion in the PKD1 gene exhibit defective ciliary polycystin localization and loss of flow‐induced Ca2+ signaling. American Journal of Physiology. Renal Physiology, 292(3), F930–F945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu, C. , Shmukler, B E. , Nishimura, K. , Kaczmarek, E. , Rossetti, S. , Harris, P. C. , Wandinger‐Ness, A. , Bacallao, R. L. , & Alper, S. L. (2009). Attenuated, flow‐induced ATP release contributes to absence of flow‐sensitive, purinergic Cai2+ signaling in human ADPKD cyst epithelial cells. American Journal of Physiology. Renal Physiology, 296(6), F1464–F1476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yadav, D. K. , Beniwal, M. K. , & Jain, A. (2013). Bardet‐Biedl syndrome a rare cause of cardiomyopathy. Indian Pediatrics, 50(6), 599–601. [PubMed] [Google Scholar]
- Yamamoto, M. , Mine, N. , Mochida, K. , Sakai, Y. , Saijoh, Y. , Meno, C. , & Hamada, H. (2003). Nodal signaling induces the midline barrier by activating Nodal expression in the lateral plate. Development (Cambridge, England), 130(9), 1795–1804. [DOI] [PubMed] [Google Scholar]
- Yamanaka, Y. , Tamplin, O. J. , Beckers, A. , Gossler, A. , & Rossant, J. (2007). Live imaging and genetic analysis of mouse notochord formation reveals regional morphogenetic mechanisms. Developmental Cell, 13(6), 884–896. [DOI] [PubMed] [Google Scholar]
- Yan, S. , Lu, J. , & Jiao, K. (2021). Epigenetic regulation of cardiac neural crest cells. Frontiers in Cell and Developmental Biology, 9, 678954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokoyama, T. , Copeland, N. G. , Jenkins, N. A. , Montgomery, C. A. , Elder, F. F. B. , & Overbeek, P. A. (1993). Reversal of left‐right asymmetry: A situs inversus mutation. Science, 260(5108), 679–682. [DOI] [PubMed] [Google Scholar]
- Yoshiba, S. , & Hamada, H. (2014). Roles of cilia, fluid flow, and Ca2+ signaling in breaking of left‐right symmetry. Trends in Genetics, 30(1), 10–17. [DOI] [PubMed] [Google Scholar]
- Yoshiba, S. , Shiratori, H. , Kuo, I. Y. , Kawasumi, A. , Shinohara, K. , Nonaka, S. , Asai, Y. , Sasaki, G. , Belo, J. A. , Sasaki, H. , Nakai, J. , Dworniczak, B. , Ehrlich, B. E. , Pennekamp, P. , & Hamada, H. (2012). Cilia at the node of mouse embryos sense fluid flow for left‐right determination via Pkd2. Science, 338(6104), 226–231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshioka, H. , Meno, C. , Koshiba, K. , Sugihara, M. , Itoh, H. , Ishimaru, Y. , Inoue, T. , Ohuchi, H. , Semina, E. V. , Murray, J. C. , Hamada, H. , & Noji, S. (1998). Pitx2, a bicoid‐type homeobox gene, is involved in a lefty‐signaling pathway in determination of left‐right asymmetry. Cell, 94(3), 299–305. [DOI] [PubMed] [Google Scholar]
- Yuan, S. , Li, J. , Diener, D. R. , Choma, M. A. , Rosenbaum, J. L. , & Sun, Z. (2012). Target‐of‐rapamycin complex 1 (Torc1) signaling modulates cilia size and function through protein synthesis regulation. PNAS, 109(6), 2021–2026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan, S. , Wang, Z. , Peng, H. , Ward, S. M. , Hennig, G. W. , Zheng, H. , & Yan, W. (2021). Oviductal motile cilia are essential for oocyte pickup but dispensable for sperm and embryo transport. PNAS, 118(22), e2102940118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan, S. , Zaidi, S. , & Brueckner, M. (2013). Congenital heart disease: Emerging themes linking genetics and development. Current Opinion in Genetics & Development, 23(3), 352–359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan, S. , Zhao, L. , Brueckner, M. , & Sun, Z. (2015). Intraciliary calcium oscillations initiate vertebrate left‐right asymmetry. Current Biology, 25(5), 556–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, B. , Zhang, T. , Wang, G. , Wang, G. , Chi, W. , Jiang, Q. , & Zhang, C. (2015). GSK3β‐Dzip1‐Rab8 cascade regulates ciliogenesis after mitosis. Plos Biology, 13(4), e1002129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, X. M. , Ramalho‐Santos, M. , & Mcmahon, A. P. (2001). Smoothened mutants reveal redundant roles for Shh and Ihh signaling including regulation of L/R asymmetry by the mouse node. Cell, 105(6), 781–792. [PubMed] [Google Scholar]
- Zhang, Y. , Liu, H. , Li, W. , Zhang, Z. , Zhang, S. , Teves, M. E. , Stevens, C. , Foster, J. A. , Campbell, G. E. , Windle, J. J. , Hess, R. A. , Pazour, G. J. , & Zhang, Z. (2018). Intraflagellar transporter protein 140 (IFT140), a component of IFT‐A complex, is essential for male fertility and spermiogenesis in mice. Cytoskeleton (Hoboken), 75(2), 70–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang, Y. J. , O'neal, W. K. , Randell, S. H. , Blackburn, K. , Moyer, M. B. , Boucher, R. C. , & Ostrowski, L. E. (2002). Identification of dynein heavy chain 7 as an inner arm component of human cilia that is synthesized but not assembled in a case of primary ciliary dyskinesia. Journal of Biological Chemistry, 277(20), 17906–17915. [DOI] [PubMed] [Google Scholar]
- Zhang, Z. , Alpert, D. , Francis, R. , Chatterjee, B. , Yu, Q. , Tansey, T. , Sabol, S. L. , Cui, C. , Bai, Y. , Koriabine, M. , Yoshinaga, Y. , Cheng, J.‐F. , Chen, F. , Martin, J. , Schackwitz, W. , Gunn, T. M. , Kramer, K. L. , De Jong, P. J. , Pennacchio, L. A. , & Lo, C. W. (2009). Massively parallel sequencing identifies the gene Megf8 with ENU‐induced mutation causing heterotaxy. PNAS, 106(9), 3219–3224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao, X. , Bian, C. , Liu, K. , Xu, W. , Liu, Y. , Tian, X. , Bai, J. , Xu, K.‐F. , & Zhang, X. (2021). Clinical characteristics and genetic spectrum of 26 individuals of Chinese origin with primary ciliary dyskinesia. Orphanet Journal of Rare Diseases, 16(1), 293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmermann, N. , Noga, A. , Obbineni, J. M. , & Ishikawa, T. (2023). ATP‐induced conformational change of axonemal outer dynein arms revealed by cryo‐electron tomography. EMBO Journal, 42(12), e112466. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are openly available in Pubmed at https://pubmed.ncbi.nlm.nih.gov.