

Abstract
Broad topics of the plastisphere in various environments are reviewed, including its methodologies, diversity, functionality, and outlook.
Due to their durability, malleability, and low cost, plastics have become an integral material of industrial and consumer products [1]. Global plastic production has enormously grown from 1.5 million metric tonnes in 1950 and 367 million metric tonnes in 2020 [2]. Despite the remarkable benefits and convenience of plastics to human lives, awareness about the negative environmental impacts has increased because of the vast amount of plastic waste. It is estimated that approximately 12,000 metric tonnes of plastic waste will be accumulated globally by 2050 [3]. Under the action of external forces, for example, solar radiation, physical abrasion, and biological degradation, large pieces of plastics can break down into smaller particles, generating microplastics (diameter < 5 mm) [4, 5, 6]. Accumulating studies show that microplastics are ubiquitous throughout various environments, including marine, freshwater, atmospheric, and soil ecosystems, even in the remotest areas of the planet [7, 8, 9, 10, 11]. Their presence and persistence in environments have raised significant concerns as microplastics can be ingested and subsequently transmitted to higher trophic‐level organisms through the food chain, and thus harm biodiversity and ecosystems [12, 13, 14].
As an exogenous and hydrophobic substrate, microplastic surfaces can provide a unique niche for the growth and proliferation of a diversity of microorganisms, constituting a distinct ecological habitat called the “plastisphere” [15, 16, 17]. Following the development of modern molecular methods and emerging bioinformatics tools, the application of high‐throughput DNA sequencing is increasing our understanding of the diverse microorganisms that inhabit the plastisphere [18, 19]. However, the consensus is lacking in the scientific community on the characteristics of the plastisphere. For instance, previous studies dominantly reported that the plastisphere microbial communities showed different compositional structures compared with the microbial communities living in surrounding environments, whereas several studies observed similar microbial communities on microplastics [20, 21]. The microbial diversity in the plastisphere could be higher or lower than that in the surrounding communities [22, 23]. Additionally, similar or distinct microbial functions may be observed within the plastisphere [24, 25]. Plastisphere studies focused initially on the marine environment and gradually extended to the freshwater and soil environments, and even indoor house environments [26, 27, 28, 29, 30]. The various environmental matrices also limited the definitive answers on the characteristics of the plastisphere. Several critical questions concerning the plastisphere remain unclear. Currently, whether microplastics really recruit specific microbial communities is unknown, and few studies have clearly illustrated the ecological processes shaping the microbial assembly on microplastics. How stochastic and deterministic processes influence the plastisphere communities remain unclear, and knowledge gaps exist concerning the adverse effects that the plastisphere may pose to the environment and ecosystem. Therefore, for this review, peer‐reviewed journal articles that investigated the plastisphere on microplastics were searched using the Web of Science (http://apps.webofknowledge.com/) with the keywords “microplastic(s)” and “plastisphere” (Figure S1), and the study designs and characterization methods exploring the plastisphere in different environments are consequently discussed. The composition, diversity, and underlying ecological mechanisms of the plastisphere were also estimated. We end by considering important unanswered questions in this field and future research priorities. This review provides a comprehensive interpretation of the plastisphere microbial communities.
PLASTISPHERE STUDIES: FROM MARINE TO LAND
Dr. Erik Zettler and colleagues first established the concept of the “plastisphere” in 2013 (Figure S1). High‐throughput sequencing method was used in their study to characterize the diverse microbial community of heterotrophs, autotrophs, predators, and symbionts on marine plastic debris collected from the North Atlantic [16]. Following this study, the plastisphere in the marine environment was extensively investigated [19]. Studies on the plastisphere in the freshwater ecosystem started in 2014 as researchers gradually recognized that marine microplastics originated from freshwater systems. In that year, Hoellein and colleagues published two papers on the plastisphere in freshwater ecosystems [17, 31]. One study used high‐throughput sequencing method to describe the bacterial assemblages colonizing microplastics sampled from an urbanized river in Chicago, Illinois, USA. The other study incubated plastispheres in three freshwater ecosystems (river, pond, and artificial stream, respectively). Community compositions on microplastics were observed to significantly differ with the respective planktonic communities. In comparison with marine and freshwater systems, soil matrices are more complex, containing variable proportions of mineral and organic matter. Tracing a solid analyte in a sample composed of solids is more difficult, and this delayed the analysis of soil plastisphere to a later period. In 2019, Huang and colleagues applied farmland field sampling and laboratory soil incubation, respectively, to estimate the characteristics of the soil plastisphere [32, 33]. In the same year, Puglisi et al. investigated the plastic‐associated biofilm in constructed solid waste disposal systems (landfill sites) [34]. These studies found that the plastisphere communities showed distinct microbial diversities, community compositions, and co‐occurrence interactions compared with those in the surrounding environmental microbial communities. The plastisphere in the atmosphere environment was investigated in the most recent years. In 2022, Wang and colleagues estimated the plastisphere in an indoor environment via artificial incubation [35]. In summary, plastisphere studies followed the detection of microplastics in different environments, which originates in marine environments, extends to freshwater ecosystems, and finally, to terrestrial habitats and other artificial environments.
EXPERIMENTAL METHODS FOR STUDYING PLASTISPHERE
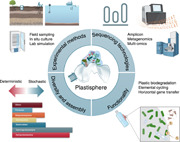
The methodology to estimate the characteristics of the plastisphere is manifold. In general, plastisphere samples are collected via three routes: field sampling, in situ field culture, and laboratory‐simulated experimentation (Tables S1–S3, Figure 1). The proportions of the three methods in studying the aquatic plastispheres are approximately the same, whereas the laboratory‐simulated experiments dominate the soil plastisphere studies. For the field sampling studies, the researchers collect the microplastics from aquatic or terrestrial ecosystems, pick up the microplastics, and analyze the microbial communities on the microplastics. The sampling and sorting of microplastics from liquid and solid samples have different details. In the aquatic environments, microplastic sampling methods are similar for both fresh and seawater samples. Manta or plankton nets are commonly used, and the collected particles are identified and selected on the spot [20, 36, 37]. For sampling in sediment or soil, solid grab samplers are often used; the collected bulk solid samples are sieved, and the putative microplastics are extracted [38, 39, 40]. The sampled microplastics are transferred to the laboratories on dry ice, and stored at −80°C before analysis. For studies using in situ field culture, microplastics are selected with certain polymer types and shapes and are then incubated in the field environments under natural conditions. After a specific duration, the microplastics are collected, and the microbial communities on their surface are analyzed [41, 42, 43]. For studies using simulated laboratory experiments, microplastics are incubated under controlled conditions (e.g., pH, nutrients, and temperature) [27, 44, 45]. For studies simulating aquatic environments, a biofilm incubation system coupled with a continuous flow device is commonly used [46]. For laboratory soil microcosm experiments, microplastics are usually incubated in sterilized glass containers under specific conditions [28, 47].
Figure 1.

The typical experimental approaches to study plastisphere in aquatic and terrestrial environments.
These methods have both advantages and disadvantages. The field sampling method can reveal the realistic ecological characteristics of the plastisphere. However, the selection of microplastics needs to be performed immediately after the sampling in case the biofilms change. Direct visual examination by the naked eyes or with dissecting microscopes is usually employed, which would inevitably introduce non‐polymer particles (e.g., mineral particles, oil residues, or chitin‐like materials) and influence further analyses [48]. Additionally, plastisphere DNA is usually extracted from all the collected polymer particles; thus, it is difficult for the field sampling studies to identify the differences between polymer types. For instance, Li et al. [29] collected microplastics and their surrounding water samples in freshwater and seawater ecosystems and analyzed the microbial composition, functions, assembly processes, and interaction networks but did not identify the polymer types of the collected microplastics. Similarly, Luo et al. [24] sampled the film residues from 55 plastic‐mulching croplands in subtropical areas of China but did not report the polymer types of these film residues. These studies can provide the characteristics of plastisphere communities and compare the results with those of the free‐living communities. For further studies, other natural materials can be collected from the sampling sites, which would be helpful for studying the specific microbial species in the plastisphere.
For the in situ incubation studies, microplastics with certain features can be selected. Therefore, in situ incubation studies using different polymers, shapes, or colors were frequently performed. For example, Pinnell and Turner [49] evaluated the microbial communities on conventional (polyethylene terephthalate [PET]) and biodegradable (polyhydroxyalkanoate [PHA]) microplastic substrates after incubation at the Laguna Madre lagoon in the northern Gulf of Mexico for 28 days. They intended to observe the distinct compositional structure between the biodegradable and conventional plastispheres. Wen et al. [50] studied the community structure and functional diversity of the microbial communities on microplastics of different colors by incubating the microplastics in a freshwater pond in the Yangtze Estuary, China. They reported that the bacterial community and structure of the plastisphere were plastic color‐dependent. Additionally, the in situ incubation method is also used to compare the biofilm on microplastics with that on other natural substrates. Oberbeckmann et al. set up an in situ experiment with polyethylene (PE) and polystyrene (PS) pellets and wooden beads along a coastal‐to‐estuarine gradient in the Baltic Sea, Germany [51]. However, this method requires sufficient time for biofilm growth, and it is not possible to maintain constant environmental conditions during the growing period. Thus, this method faces difficulties in quantifying the influences of environmental conditions on plastisphere characteristics. Additionally, to ensure the recovery of cultured microplastics, mesh bags are often used for the studies in aquatic environments. Recently, several studies also employed this method in terrestrial environments [28, 52, 53, 54, 55]. For example, Zhu et al. [28] used nylon mesh bags (mesh size: 50 μm) as microplastic containers and buried them in the soil to investigate the microbial community of the soil plastisphere. However, this operation may introduce heterogeneity in the observed plastisphere as the contact distances between microplastic and soil particles in the system may differ. Even though in situ incubation has several limitations, the incorporation of the environmental conditions provides insights into the effects of realistic environmental variables on plastisphere communities.
In comparison with the in situ incubation method, laboratory simulation experiments can study the influences of single or multiple environmental variables on the plastisphere characteristics under controlled conditions. Yang et al. [56] used a biofilm reactor to incubate a plastisphere under laboratory conditions and evaluate the effects of incubation time on the microbial community structure. Li et al. [44] reported that the soil pH, rather than heavy metals, showed a stronger influence on the plastisphere bacterial communities. However, this method has several limitations. One major concern is the microplastic concentration used in the laboratory incubation test. In principle, the laboratory incubation should maximumly simulate the field conditions. Therefore, the microplastic concentrations used in laboratory incubation should be similar to their environmental concentrations. However, even though all the published papers declared that the microplastic concentrations used in their studies were based on the detected concentrations, the values used are generally higher [57]. Moreover, it is unrealistic to perform experiments on time scales close to that experienced for microplastics in the real environment. For example, microplastics collected from marine environments or landfill sites may have aged for years [58]. It is nearly impossible to duplicate field processes in a laboratory setting, e.g., temperature, pH, redox, light intensity, and indigenous microorganisms. Laboratory simulations take advantage of controlled conditions, which significantly contribute to our understanding of the mechanisms shaping the plastisphere community. In our opinion, a combination of the above methods would be optimal for a comprehensive understanding of the formation of the plastisphere and the contributing factors.
CHARACTERIZATION METHODS: FROM AMPLICON SEQUENCING TO OMICS RESEARCH
Imaging technologies, such as scanning electron microscopy (SEM) and combinatorial labeling and spectral imaging‐fluorescence in situ hybridization (CLASI‐FISH), have demonstrated that the plastisphere can be a crowded, surface‐based micro‐ecosystem that includes a diverse range of microorganisms [59, 60, 61]. Over recent years, next‐generation sequencing (NGS) technologies have gained enormous popularity in the analysis of microbial communities [18, 19]. Most plastisphere microbiome surveys have used amplicon and metagenomic sequencing. Polymerase chain reaction (PCR) coupled with the sequencing of the taxonomic marker gene can uncover the taxonomic profiles of microbial communities at low cost, marking this omnipresent in microbiome research (Figure 2). The 16S rRNA gene is the most common marker used for identifying bacterial communities in the plastisphere. Wang et al. used 16S rRNA gene sequencing to analyze the bacterial communities on microplastics collected from two urban rivers in China [62]. Ogonowski et al. explored the Baltic bacterioplankton and plastisphere by Illumina sequencing of 16S rRNA gene libraries and reported substrate‐driven selection [63]. In comparison, only a handful of studies have focused on eukaryotes in plastisphere communities [64, 65, 66]. By implementing internal transcribed spacer2 metabarcoding on plastic debris, De Tender et al. performed the first study to identify and characterize fungal genera on marine microplastics [67].
Figure 2.

Amplicon and shotgun metagenomic sequencing used for studying plastisphere.
Although the second‐generation sequencing technology has yielded substantial data concerning the plastisphere composition, it is still limited in the taxonomic resolution and coverage. This technology only produces short reads, forcing researchers to choose short barcoding regions and thus have a lower barcoding resolution [68]. Hence, long‐read sequencing technology, such as the third‐generation Pacific Biosystems SMRT and Oxford Nanopore sequencing, has been used in studying the plastisphere composition and provide improved classification of taxa [69, 70]. For instance, Davidov et al. identified 61 plastic‐associated microorganisms at the species level from the Mediterranean Sea using DNA metabarcoding with Nanopore MinION [69]. Several studies have applied shotgun metagenomic sequencing to estimate the whole genomic DNA in the plastisphere [49, 71, 72], as this technology can simultaneously capture all the genetic material present in one sample and thus can show the taxonomic information across kingdoms and predict microbial functions. The first metagenomic study was performed by Jessica Bryant and colleagues in 2016 [73]. Following technical and analytical breakthroughs, multiomics, including metatranscriptomic, proteogenomic, and metabolomic approaches, have been used in recent studies to enable a comprehensive understanding of active members and acting functions of the plastisphere. These approaches can reveal the metabolic pathways active under different conditions and confirm the metagenomic functional predictions. Therefore, these methods have huge potential for analyzing the functional microorganisms in the plastispheres, such as potential microbes or enzymes for degrading plastics. For instance, Wright et al. used a combined proteogenomic and metabolomic approach to characterize the biodegradation and microbial community succession within the polyethylene terephthalate (PET) plastisphere [72]. Similarly, Wu et al. integrated metagenomic and metatranscriptomic technologies to evaluate the antibiotic resistomes in a laboratory‐incubated plastisphere, demonstrating that the antibiotic resistomes in the plastisphere were not only present but also actively expressed. Future multiomic approaches will considerably contribute to deciphering the characteristics of the plastispheres and their roles in microbial‐mediated biochemical transformations.
THE COMPOSITION AND ASSEMBLY OF THE PLASTISPHERE
The microbial community in the plastisphere has recently become an important research topic. Although diverse microorganisms, including bacteria, archaea, fungi, and other eukaryotes, can colonize microplastics, almost all current work is focused on bacterial communities. Community composition varies significantly across the soil, freshwater, and seawater plastisphere (Figure 3). Alpha‐, Beta‐, and Gamma‐Proteobacteria are typical enriched in freshwater plastisphere communities [66, 75, 76, 77]. The seawater plastisphere community mainly comprises bacteria belonging to the phylum Proteobacteria (Alpha‐ and Gamma‐) and to the Bacteroidetes [42, 78, 79, 80]. The vast majority of bacteria that colonize terrestrial plastics mainly belongs to the phyla Proteobacteria (Alpha‐) and Actinobacteria [27, 47, 81, 82, 83].
Figure 3.
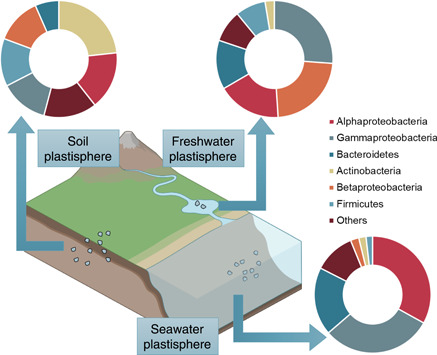
General structure of the bacterial communities in seawater, freshwater, and soil plastisphere (data based on REFs [17, 20, 22, 26, 27, 28, 29, 31, 32, 34, 40, 62, 63, 67, 74]).
The plastisphere community composition has been reported to significantly differ from that in the surrounding environment. Several studies even reported that certain microbial groups, i.e., members of Alpha‐Proteobacteria (e.g., Rhodospirillaceae), were consistently found on plastics [62, 81, 84, 85, 86]. However, increasing numbers of studies found that the bulk of microorganisms colonizing plastics are likely the same as those colonize other surfaces. The concept that the plastisphere exhibits a unique microbial community has yet to be conclusively demonstrated. It has been well established that the vast majority of planktonic microbes that populate the water column (e.g., Synechococcus and Prochlorococcus species) strongly diverge from those able to associate and form biofilms [87, 88, 89]. Thus, unsurprisingly, the marine and freshwater plastisphere would exhibit distinct microbial communities in comparison with the bulk water community. In fact, biofilms can be observed on any surface in aquatic environments, from artificial surfaces, such as bottles [74], ship hulls [90], oil platforms [91], and other man‐made items, to natural surfaces, such as animals, plants, zooplankton, micro/macro‐aggregates, transparent exopolymer particles, and rocks [92]. Comparing the plastisphere with other biofilms in aquatic environments would be more critical for understanding the core unique microbial community in the plastisphere. But until now, only several plastisphere studies in aquatic environments include comparable control materials. For example, glass, ceramic, shells, or wood were used in previous studies to compare the plastisphere with other biofilms formed on other materials [22, 45, 63, 93]. No microorganisms occur only on plastics, and the relative abundance profiles of the major bacterial groups on different materials (i.e., conventional microplastics and ceramics) are substantially similar [19]. For soil microorganisms, increasing studies demonstrate that biofilm is the predominant mode of life, whereby microbial species colonize various substrates and interfaces, including mineral surfaces [94], pore spaces [95], and plant roots [96]. Microplastics can also provide a surface for the growth and proliferation of soil microorganisms. The analogous sessile lifestyles may be the reason that the abundant bacterial groups are similar in the soil plastisphere and bulk soil.
The strongest differences in community structures between the plastisphere and the surrounding communities in both aquatic and terrestrial environments are commonly observed at the early stage of colonization [57]. For example, Ogonowski et al. [63] reported that the plastisphere on PE, PP, and PS microplastics was distinctly different from those on the nonplastic substrates (cellulose and glass beads) after 2 weeks of incubation in seawater under laboratory conditions. Alpha‐ (such as the Rhodobacteraceae) Gamma‐proteobacteria, and Flavobacteria are among the initial colonizers in the plastisphere and are typical fast‐growing opportunistic bacteria that can quickly respond to changes in the environment [97]. The surface properties of microplastics, including hydrophilicity, roughness, and electric charge, strongly affect the attachment of early microbial colonizers. Previous studies suggest that bacteria may preferentially colonize more hydrophilic surfaces, possibly due to the higher wettability and surface energies of these hydrophilic surfaces [61]. This could be why more intense biofilms are formed on more hydrophilic polymer surfaces [43]. Additionally, the high surface roughness of microplastics would supply more attachment points for microbes, and thus impact the microbial community composition at the early stage of the plastisphere [98]. Following the development of the plastisphere, the relative importance of substrates in shaping the microbial composition in the plastisphere declines. The environmental variables, such as temperature, salinity, pH, and nutrient content, influence the dynamic patterns of the microbial community in the plastisphere during the stages of development and maturation. For example, using a soil microcosm experiment with different microplastics under 15°C and 25°C, Sun et al. [27] reported that the temperature, rather than polymer type, significantly induced the differences between the plastisphere communities on PE and PLA microplastics. Wright et al. [99] estimated the global diversity of the plastisphere via a meta‐analysis and reported that environmental variables have the largest impact on microbial composition. Currently, there remains no clear conclusion about how environmental variables influence plastisphere microbial communities. The prioritized environmental factors are also unknown. Leached compounds from microplastics, that is, plastic additives and plastic oligomers, can partly impact the assembly of microbial communities in the plastisphere [43, 61]. Significantly distinct communities have been observed on biodegradable and conventional microplastics [82, 83, 100, 101]. A potential reason may be that biodegradable microplastics supply a nutrient‐rich environment compared with the conventional microplastics, thus recruiting copiotrophic microbes. Plastic additives may also influence the plastisphere communities as these artificial chemicals may promote or inhibit the biofilm growth. Overall, we may conclude that microplastic surface properties affect the stage of initial colonization, and that environmental factors and polymer inner characteristics play a critical role in the succession of the plastisphere.
Interesting is growing in understanding the ecological mechanisms that drive the formation and maturation of the plastisphere. The application and examination of ecological theories to plastisphere microbial communities could build insights into when and how this microbial community structure changes through plastisphere life. Furthermore, understanding how plastisphere communities assemble over time is essential to understand changes in community diversity, species, and trait abundance, interactions between community members, and ultimately, the functional genes in the plastisphere. Theoretically, ecologists have proposed that four ecological and evolutionary processes, namely selection, dispersal, drift, and diversification, drive all principles in community ecology [102]. Two distinct theoretical lines have been debated to examine and interpret the community assembly: the niche‐based theory and the neutral theory, which have been recently demonstrated to simultaneously drive community assembly [103, 104]. Several approaches have been used to obtain quantitative information on plastisphere community assembly, including quantifying assembly processes based on entire‐community null model analysis (QPEN) [105] and infer community assembly mechanisms by phylogenetic‐bin‐based null model analysis [106] as well as normalized stochasticity ratio (NST) [107] and the Sloan neutral model [108]. Most studies suggested that stochastic processes dominantly drive the microbial community assembly on microplastics [22, 26]. For example, Sun et al. [20] used the null model, NST, and the neutral model to estimate the assembly processes of bacterial communities in a field‐sampled aquatic plastisphere and found that the stochastic process of drift and dispersal limitation primarily contributed to the microbial community assembly. Studies on the soil plastisphere also indicated the dominant importance of neutral‐based processes on microbial assembly [24, 27, 100, 109]. However, Li et al. [29] reported that the niche‐based processes (deterministic) dominated the plastisphere community assembly as they observed low goodness‐of‐fit values for the neutral model of the plastisphere. These observations were ascribed to the heterogeneity of the microplastic samples. Many factors would influence the microbial assembly in the plastisphere. For instance, at the initial stage of colonization, the properties of the microplastic surface would impact the communities, and thus this selection will dominate the microbial assembly [100]. For the maturation stage of the plastisphere, the selection of microplastics may gradually decrease, and the importance of dispersal and drift would increase [100]. Parallelly, compounds that leach from microplastics may influence the composition of the plastisphere owing to microbial chemotaxis, which would further influence the assembly processes. Under the scenario that the plastisphere is sampled from large spatial areas, location‐specific relationships may be obtained, and probably, the dominant roles of selection. Simultaneously, particle relocation occurs during the transportation of plastics or microplastics at local and global scales. Such a phenomenon can increase the uncertainty when estimating the plastisphere characteristics. More work is needed to quantitatively determine the importance of stochastic and deterministic processes in the microbial community assembly in the plastisphere.
THE FUNCTIONS OF PLASTISPHERE
Whether or not the microorganisms in the plastisphere are degrading microplastics and contributing to the weight loss of plastics is an ongoing question. Several studies screened polymer‐degrading microorganisms from the plastisphere [110, 111]. For instance, Delacuvellerie et al. [110] compared the structures of bacterial communities from floating plastics, sediment‐associated plastics and sediments from the Mediterranean Sea and observed that hydrocarbon‐degrading bacteria such as Alcanivorax, Marinobacter and Arenibacter genera are enriched with plastics, implying that these bacteria were potentially involved in plastic degradation. Multiomics has also been used for evaluating the plastic degradation by plastisphere communities. Jessica Bryant and colleagues used metagenomic sequencing to observe that several putative xenobiotic biodegradation genes were more abundant on plastic particles. However, these data only allow for speculation as to whether the microorganisms residing on plastic particles are actually degrading plastics. Bhagwat et al. [112] also observed the upregulation of PET hydrolysis‐related enzyme genes in the plastisphere by using metagenomics, potentially suggesting plastic degradation. Through a combination of proteogenomic and metabolomic approach, Wright et al. characterized marine PET‐degrading enzymes and compared the degradable ability of plastisphere on different PET plastics during 6 weeks of incubation in the marine environment [72]. They reported that degradation depends on the recalcitrance of the substrate (i.e., crystallinity) and the accessibility of the substrate to the microbes. The study clearly demonstrates the potential for PET degradation existing in the marine plastisphere. However, the biodegradation of highly recalcitrant plastics (e.g., PE, PP, and PS) by the plastisphere has seldom been reported. For example, with an integrated metagenomics and metaproteomics approach, Oberbeckmann et al. observed polymer‐unspecific communities on PE and PS microplastics after 2 weeks of in situ incubation, but these plastics did not appear to be undergoing biodegradation [50]. Similarly, Delacuvellerie et al. [113] sampled PE and PP microplastics from the Mediterranean Sea, and used metagenomic and metaproteomic analysis to estimate the characteristics of the plastisphere. Despite the presence of hydrocarbon‐degrading bacteria was observed in the metagenomes, polymer degradation metabolism was not detected at the protein level. Therefore, whether plastispheres can degrade plastics seems to be closely related to the polymer types. PE, PP, and PS microplastics contain very stable backbones and are difficult to degrade, whereas PET, polyurethane, and polycarbonate are more susceptible to hydrolysis and to enzymes that catalyze the degradation. Further in‐depth exploration with integrated genomics is warranted to estimate plastic biodegradation by the plastisphere.
Plastisphere microorganisms are involved in functions related to elemental geochemical cycles. Pinnell and Turner observed the enrichment of adenylyl sulfate reductase and dissimilatory sulfite reductase genes in microorganisms on microplastics cultured at the sediment‐interface of a coastal lagoon, suggesting the stimulation of sulfate reduction in the plastisphere [48]. Bryant et al. [73] found genes with significantly higher abundance among plastic‐attached bacteria; these genes, such as nifH, suggest enrichment for nitrogen fixation in the plastisphere. Several studies provided substantive evidence that the plastisphere influences nitrogen cycling [114, 115, 116]. For instance, Su et al. [116] reported that plastisphere exhibits a higher denitrifying activity and N2O production than that in the surrounding bulk water, suggesting an overlooked N2O source. The plastisphere is more likely to recruit denitrifiers for colonization mainly because of the hypoxic conditions with the plastisphere and the denitrifiers utilize nitrate/nitrite as electron acceptors to sustain their metabolism [116]. Rahman et al. [117] performed a functional gene‐array analysis of microbial communities on PET and PLA microplastics and found that genes involved in carbon degradation and fixation, nitrogen fixation and denitrification, and sulfur reduction were more abundant. Although a majority of these studies could not precisely estimate the plastisphere‐induced changes in elemental flux, these results indicate that plastisphere can change the stability and function of the surrounding ecosystem. Considering the substantial plastic waste and microbial biomass, future research at a global scale is needed.
The presence of various potential pathogens has been reported from environmental plastic samples around the worldwide. For instance, Vibrio species are usually reported in the marine plastispheres from both temperate and tropical marine environments [68, 118, 119], and Pseudomonas (e.g., Pseudomonas monteilii, P. mendocina, P. angilliseptica, and P. syringae) is observed in freshwater and soil plastispheres [101, 120, 121]. A previous study identified that the ratios of potential pathogens/bacteria in the plastisphere were higher than those in the soil [28]. Furthermore, antibiotic resistance genes (ARGs) are frequently observed with the plastisphere in diverse environments [26, 35, 37, 56, 122]. For instance, Yang et al. [122] found that the abundance and diversity of ARGs in plastic microbiota were significantly greater than those in seawater microbiota in the North Pacific Gyre, which was the first report on ARGs in the plastisphere. Since this study, the roles of microplastics acting as reservoirs and refuges for ARGs are of increasing concern. As the bacteria in biofilms possess high diversity and metabolic complexity, the plastisphere is usually proposed to be a hotspot for horizontal gene transfer. Previous laboratory findings indicated that the frequency of plasmid transfer between plastic‐associated bacteria was higher than that in free‐living bacteria. The high level of gene changes would facilitate ARG propagation in the plastisphere. In addition, microorganisms embedded in the plastisphere are protected by extracellular polymeric substances and are more tolerant to the environmental stresses than the discrete cells. Thus, the acquisition of antibiotic tolerance by pathogens would be easier in the plastisphere, potentially presenting significant challenges for human health. However, reporting of microplastics as vectors for ARGs and potential pathogens should be taken with caution. Biofilms are often considered as natural reservoirs of ARGs and potential pathogens. Therefore, the higher durability, buoyancy, and transportability of microplastics compared with those of natural co‐occurring materials should be highlighted. These properties would induce the plastisphere to pose a higher risk because of the prolonged exposure time and distances. To date, studies still cannot answer the question of whether microplastics specifically recruit potential pathogens and ARGs. Several other vital questions, such as whether horizontal gene transfer of ARGs into pathogens is more frequent and easier on microplastics and whether pathogens could transfer from microplastics to organisms under natural conditions, must be addressed by future studies.
Compared with taxonomic diversity, the functional diversity of the plastisphere is considerably unexplored. Current functional predictions from sequencing methods revealed that diverse metabolic pathways exist in the plastisphere, potentially suggesting that the plastisphere community may participate in a large number of important ecological processes in the ecosystems. Multiple genes involved in xenobiotic degradation are enriched in the biofilm on microplastics and ARGs are frequently observed in plastisphere communities. However, importantly, few studies have shown the substantive evidence for the predicted functions. Multi‐omic technologies, coupled with verification experiments, are needed in future studies.
PERSPECTIVE
The contamination of aquatic and terrestrial ecosystems with plastics is a global threat in the Anthropocene and shows no sign of decreasing in the near future. Understanding the assembly processes and the associated environmental impacts of the plastisphere is vital to managing and predicting the risks posed by plastic pollution. This review systematically summarizes the experimental methods to explore the plastisphere, the community composition and assembly processes, and the associated ecological functions. However, continued assessment and elaborate experimentation are required to answer the open questions in plastisphere research.
Detailed information on microbial community succession during the early stages of the plastisphere is very limited. Biofilm communities on particle surfaces can be formed within several hours, however, few studies ventured into the formation and growth dynamics of early plastisphere communities. Traditional incubation and sequencing methods may not be suitable for tracking these processes. Microfluidic techniques can control structural and fluid behavior at the microscopic scale and can therefore simulate the heterogenous microenvironment between microplastics and the surrounding liquids. A combination of microfluidic devices and optical coherence tomography can provide valuable qualitative and quantitative information regarding the spatial structure of biofilms, which may be helpful for plastisphere studies.
The key factors influencing the plastisphere communities are unclear. Diverse environmental factors, including hydrographic conditions, soil textures, nutrient availability, temperature, salinity, pH, and microplastic characteristics, such as polymer types, surface hydrophilicity, potential charges, and surface morphologies, can affect plastisphere communities. There is a knowledge gap in the underlying mechanisms whereby these factors drive the communities.
Biological studies of the plastisphere have predominately focused on bacterial communities, whereas fungal and micro‐eukaryotic communities are poorly understood. All these microorganisms play vital roles in plastisphere ecological processes and create complex interactions between neighboring cells. They can exhibit cooperative behaviors to enhance stress resistance and nutrient uptake. In parallel, competition is always pervasive in the plastisphere because of the space and resource limitations present. The diversity and variation of microbes in the plastisphere ensure that these interactions are always manifold and dynamic, further shaping community structure and function. We still lack the knowledge of how microorganisms interact with each other in the plastisphere, how they respond to environmental and microplastic properties, and how they drive the microbial functions in the plastisphere.
Comprehensive methods, such as integrated genomics, metagenomics, metatranscriptomics, and metabolomics, are needed for an in‐depth understanding of the plastisphere and would facilitate the exploration of microbial diversity and functions to help reveal the molecular and ecological mechanisms and provide a holistic view of plastisphere. What we should notice is that these sequencing methods will unavoidably lead to errors. These errors may stem from the bioinformatic analysis, causing misalignment of short reads and mistakes in the genome assembly. Further studies should provide confirmatory evidence by culture‐dependent methods about the functional processes that occur within the plastisphere, such as biodegradation and pathogenicity.
AUTHOR CONTRIBUTIONS
Jie Wang and Linna Du contributed to the overall conceptualization and design. Yuanze Sun, Mochen Wu, and Jingxi Zang contributed to the writing and discussion of the main content of this manuscript. Muke Huang and Cheng Chen contributed to the figures and tables of this manuscript. All authors have read and approved the final manuscript.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflict of interest.
Supporting information
Supporting information.
Supporting information.
ACKNOWLEDGMENTS
This project was supported by the National Natural Science Foundation of China (No. 41907341) and the Beijing Innovation Consortium of Agriculture Research System (No. BAIC01‐2023‐27).
Sun, Yuanze , Wu Mochen, Zang Jingxi, Du Linna, Huang Muke, Chen Cheng, and Wang Jie. 2023. “Plastisphere Microbiome: Methodology, Diversity, and Functionality.” iMeta 2, e101. 10.1002/imt2.101
DATA AVAILABILITY STATEMENT
This manuscript does not generate any code or data. Supplementary materials (figures, tables, scripts, graphical abstract, slides, videos, Chinese translated version and update materials) may be found in the online DOI or iMeta Science http://www.imeta.science/.
REFERENCES
- 1. Andrady, Anthony L. , and Neal Mike A.. 2009. “Applications and Societal Benefits of Plastics.” Philosophical Transactions of the Royal Society B: Biological Sciences 364: 1977–84. 10.1098/rstb.2008.0304 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. “Plastics‐The Facts . 2021. An analysis of European latest plastics production, demand and waste data.”
- 3. Chamsa, Ali , Moon Hyunjin, Zheng Jiajia, Qiu Yang, Tabassum Tarnuma, Jang Jun Hee, Abu‐Omar Mahdi, Scott Susannah L., and Suh Sangwon. 2020. “Degradation Rates of Plastics in the Environment.” ACS Sustainable Chemistry & Engineering 8: 3494–511. 10.1021/acssuschemeng.9b06635 [DOI] [Google Scholar]
- 4. Bouwmeester, Hans , Hollman Peter C. H., and Peters Ruud J. B.. 2015. “Potential Health Impact of Environmentally Released Micro‐and Nanoplastics in the Human Food Production Chain: Experiences from Nanotoxicology.” Environmental Science & Technology 49: 8932–47. 10.1021/acs.est.5b01090 [DOI] [PubMed] [Google Scholar]
- 5. Foekema, Edwin M. , De Gruijter Corine, Mergia Mekuria T., van Franeker Jan Andries, Murk AlberTinka J., and Koelmans Albert A.. 2013. “Plastic in North Sea Fish.” Environmental Science & Technology 47: 8818–24. 10.1021/es400931b [DOI] [PubMed] [Google Scholar]
- 6. Thompson, Richard C. , Olsen Ylva, Mitchell Richard P., Davis Anthony, Rowland Steven J., John Anthony W. G., McGonigle Daniel, and Russell Andrea E.. 2004. “Lost At Sea: Where is All the Plastic?” Science 304: 838838. 10.1126/science.1094559 [DOI] [PubMed] [Google Scholar]
- 7. Hurley, Rachel , Woodward Jamie, and Rothwell James J.. 2018. “Microplastic Contamination of River Beds Significantly Reduced by Catchment‐Wide Flooding.” Nature Geoscience 11: 251–57. 10.1038/s41561-018-0080-1 [DOI] [Google Scholar]
- 8. Katija, Kakani , Choy C. Anela, Sherlock Rob E., Sherman Alana D., and Robison Bruce H.. 2017. “From the Surface To the Seafloor: How Giant Larvaceans Transport Microplastics Into the Deep Sea.” Science Advances 3: e1700715. 10.1126/sciadv.1700715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Evangeliou, N. , Grythe H., Klimont Z., Heyes C., Eckhardt S., Lopez‐Aparicio S., and Stohl A.. 2020. “Atmospheric Transport is a Major Pathway of Microplastics to Remote Regions.” Nature Communications 11(1): 3381. 10.1038/s41467-020-17201-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Nizzetto, Luca , Langaas Sindre, and Futter Martyn. 2016. “Pollution: Do Microplastics Spill on to Farm Soils?” Nature 537: 488488. 10.1038/537488b [DOI] [PubMed] [Google Scholar]
- 11. Allen, Steve , Allen Deonie, Phoenix Vernon R, Le Roux Gaël, Durántez Jiménez Pilar, Simonneau Anaëlle, Binet Stéphane, and Galop Didier. 2019. “Atmospheric Transport and Deposition of Microplastics in a Remote Mountain Catchment.” Nature Geoscience 12: 339–44. 10.1038/s41561-019-0335-5 [DOI] [Google Scholar]
- 12. de Souza Machado, Anderson Abel , Lau Chung Wai, Till Jennifer, Kloas Werner, Lehmann Anika, Becker Roland, and Rillig Matthias C.. 2018. “Impacts of Microplastics on the Soil Biophysical Environment.” Environmental Science & Technology 52: 9656–65. 10.1021/acs.est.8b02212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Xu, Shen , Ma Jie, Ji Rong, Pan Ke, and Miao Ai‐Jun. 2020. “Microplastics in Aquatic Environments: Occurrence, Accumulation, and Biological Effects.” Science of the Total Environment 703: 134699. 10.1016/j.scitotenv.2019.134699 [DOI] [PubMed] [Google Scholar]
- 14. Carbery, Maddison , O'Connor Wayne, and Palanisami Thavamani. 2018. “Trophic Transfer of Microplastics and Mixed Contaminants in the Marine Food Web and Implications for Human Health.” Environment International 115: 400–409. 10.1016/j.envint.2018.03.007 [DOI] [PubMed] [Google Scholar]
- 15. Koelmans, Albert A. , Redondo‐Hasselerharm Paula E., Nor Nur Hazimah Mohamed, de Ruijter Vera N., Mintenig Svenja M., and Kooi Merel. 2022. “Risk Assessment of Microplastic Particles.” Nature Reviews Materials 7: 138–52. 10.1038/s41578-021-00411-y [DOI] [Google Scholar]
- 16. Zettler, Erik R. , Mincer Tracy J., and Amaral‐Zettler Linda A.. 2013. “Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris.” Environmental Science & Technology 47: 7137–46. 10.1021/es401288x [DOI] [PubMed] [Google Scholar]
- 17. McCormick, Amanda , Hoellein Timothy J., Mason Sherri A., Schluep Joseph, and Kelly John J.. 2014. “Microplastic is an Abundant and Distinct Microbial Habitat in an Urban River.” Environmental Science & Technology 48: 11863–71. 10.1021/es503610r [DOI] [PubMed] [Google Scholar]
- 18. De Tender, C. , Schlundt C., Devriese LI, Mincer TJ, Zettler ER, and Amaral‐Zettler LA. 2017. “A Review of Microscopy and Comparative Molecular‐Based Methods to Characterize “Plastisphere” Communities.” Analytical Methods 9: 2132–43. 10.1039/C7AY00260B [DOI] [Google Scholar]
- 19. Amaral‐Zettler, Linda A. , Zettler Erik R., and Mincer Tracy J.. 2020. “Ecology of the Plastisphere.” Nature Reviews Microbiology 18: 139–51. 10.1038/s41579-019-0308-0 [DOI] [PubMed] [Google Scholar]
- 20. Sun, Yuanze , Zhang Mengjun, Duan Chongxue, Cao Na, Jia Weiqian, Zhao Zelong, Ding Changfeng, Huang Yi, and Wang Jie. 2021. “Contribution of Stochastic Processes to the Microbial Community Assembly on Field‐Collected Microplastics.” Environmental Microbiology 23: 6707–20. 10.1111/1462-2920.15713 [DOI] [PubMed] [Google Scholar]
- 21. Szabó, István , Al‐Omari Jafar, Szerdahelyi Gábor Soma, Farkas Milán, Al‐Omari Yazid, Szabó Péter Márton, Sebők Rózsa, et al. 2021. “In Situ Investigation of Plastic‐Associated Bacterial Communities in a Freshwater Lake of Hungary.” Water, Air, & Soil Pollution 232: 493. 10.1007/s11270-021-05445-0 [DOI] [Google Scholar]
- 22. Zhang, Sheng‐Jie , Zeng Yan‐Hua, Zhu Jian‐Ming, Cai Zhong‐Hua, and Zhou Jin. 2022. “The Structure and Assembly Mechanisms of Plastisphere Microbial Community in Natural Marine Environment.” Journal of Hazardous Materials 421: 126780. 10.1016/j.jhazmat.2021.126780 [DOI] [PubMed] [Google Scholar]
- 23. Nguyen, Hien Thi , Choi Woodan, Kim Eun‐Ju, and Cho Kyungjin. 2022. “Microbial Community Niches on Microplastics and Prioritized Environmental Factors Under Various Urban Riverine Conditions.” Science of the Total Environment 849: 157781. 10.1016/j.scitotenv.2022.157781 [DOI] [PubMed] [Google Scholar]
- 24. Luo, Gongwen , Jin Tuo, Zhang Huiru, Peng Jianwei, Zuo Ning, Huang Ying, Han Yongliang, et al. 2022. “Deciphering the Diversity and Functions of Plastisphere Bacterial Communities in Plastic‐Mulching Croplands of Subtropical China.” Journal of Hazardous Materials 422: 126865. 10.1016/j.jhazmat.2021.126865 [DOI] [PubMed] [Google Scholar]
- 25. Wallbank, Jessica A. , Lear Gavin, Kingsbury Joanne M., Weaver Louise, Doake Fraser, Smith Dawn A., Audrézet François, et al. 2022. “Into the Plastisphere, Where Only the Generalists Thrive: Early Insights in Plastisphere Microbial Community Succession.” Frontiers in Marine Science 626. 10.3389/fmars.2022.841142 [DOI] [Google Scholar]
- 26. Xu, Chenye , Lu Jiawei, Shen Chensi, Wang Jie, and Li Fang. 2022. “Deciphering the Mechanisms Shaping the Plastisphere Antibiotic Resistome on Riverine Microplastics.” Water Research 225: 119192. 10.1016/j.watres.2022.119192 [DOI] [PubMed] [Google Scholar]
- 27. Sun, Yuanze , Shi Jia, Wang Xiang, Ding Changfeng, and Wang Jie. 2022. “Deciphering the Mechanisms Shaping the Plastisphere Microbiota in Soil.” Msystems 7: e00352–22. 10.1128/msystems.00352-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Zhu, Dong , Ma Jun, Li Gang, Rillig Matthias C., and Zhu Yong‐Guan. 2022. “Soil Plastispheres as Hotspots of Antibiotic Resistance Genes and Potential Pathogens.” The ISME Journal 16: 521–32. 10.1038/s41396-021-01103-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Li, Changchao , Wang Lifei, Ji Shuping, Chang Mengjie, Wang Longfei, Gan Yandong, and Liu Jian. 2021. “The Ecology of the Plastisphere: Microbial Composition, Function, Assembly, and Network in the Freshwater and Seawater Ecosystems.” Water Research 202: 117428. 10.1016/j.watres.2021.117428 [DOI] [PubMed] [Google Scholar]
- 30. Erni‐Cassola, Gabriel , Wright Robyn J., Gibson Matthew I., and Christie‐Oleza Joseph A.. 2020. “Early Colonization of Weathered Polyethylene by Distinct Bacteria in Marine Coastal Seawater.” Microbial Ecology 79: 517–26. 10.1007/s00248-019-01424-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Hoellein, Timothy , Rojas Miguel, Pink Adam, Gasior Joseph, and Kelly John. 2014. “Anthropogenic Litter in Urban Freshwater Ecosystems: Distribution and Microbial Interactions.” PLoS ONE 9: e98485. 10.1371/journal.pone.0098485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Zhang, Mengjun , Zhao Yanran, Qin Xiao, Jia Weiqian, Chai Liwei, Huang Muke, Huang Yi. 2019. “Microplastics from Mulching Film is a Distinct Habitat for Bacteria in Farmland Soil.” Science of the Total Environment 688: 470–78. 10.1016/j.scitotenv.2019.06.108 [DOI] [PubMed] [Google Scholar]
- 33. Huang, Yi , Zhao Yanran, Wang Jie, Zhang Mengjun, Jia Weiqian, and Qin Xiao. 2019. “LDPE Microplastic Films Alter Microbial Community Composition and Enzymatic Activities in Soil.” Environmental Pollution 254: 112983. 10.1016/j.envpol.2019.112983 [DOI] [PubMed] [Google Scholar]
- 34. Puglisi, Edoardo , Romaniello Francesco, Galletti Serena, Boccaleri Enrico, Frache Alberto, and Cocconcelli Pier Sandro. 2019. “Selective Bacterial Colonization Processes on Polyethylene Waste Samples in an Abandoned Landfill Site.” Scientific Reports 9: 14138. 10.1038/s41598-019-50740-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Peng, Chu , Zhang Xiaofei, Zhang Xinyi, Liu Chunguang, Chen Zeyou, Sun Hongwen, and Wang Lei. 2022. “Bacterial Community Under the Influence of Microplastics in Indoor Environment and the Health Hazards Associated with Antibiotic Resistance Genes.” Environmental Science & Technology 56: 422–32. 10.1021/acs.est.1c04520 [DOI] [PubMed] [Google Scholar]
- 36. Di Pippo, Francesca , Venezia Cristina, Sighicelli Maria, Pietrelli Loris, Di Vito Stefania, Nuglio Simone, and Rossetti Simona. 2020. “Microplastic‐Associated Biofilms in Lentic Italian Ecosystems.” Water Research 187: 116429. 10.1016/j.watres.2020.116429 [DOI] [PubMed] [Google Scholar]
- 37. Di Pippo, Francesca , Crognale Simona, Levantesi Caterina, Vitanza Luca, Sighicelli Maria, Pietrelli Loris, Di Vito Stefania, Amalfitano Stefano, and Rossetti Simona. 2022. “Plastisphere in Lake Waters: Microbial Diversity, Biofilm Structure, and Potential Implications for Freshwater Ecosystems.” Environmental Pollution 310: 119876. 10.1016/j.envpol.2022.119876 [DOI] [PubMed] [Google Scholar]
- 38. Zhang, Xiaoyong , Xia Xiongjian, Dai Ming, Cen Jianwei, Zhou Lei, and Xie Jiefen. 2021. “Microplastic Pollution and its Relationship With the Bacterial Community in Coastal Sediments Near Guangdong Province, South China.” Science of the Total Environment 760: 144091. 10.1016/j.scitotenv.2020.144091 [DOI] [PubMed] [Google Scholar]
- 39. Baptista Neto, José Antonio , Gaylarde Christine, Beech Iwona, Bastos Alex Cardoso, da Silva Quaresma Valéria, and de Carvalho Diego Gomes. 2019. “Microplastics and Attached Microorganisms in Sediments of the Vitória Bay Estuarine System in SE Brazil.” Ocean & Coastal Management 169: 247–53. 10.1016/j.ocecoaman.2018.12.030 [DOI] [Google Scholar]
- 40. Wu, Nan , Zhang Ying, Zhao Ze, He Jiahui, Li Wenjie, Li Jiafu, Xu Wei'an, Ma Yongzheng, and Niu Zhiguang. 2020. “Colonization Characteristics of Bacterial Communities on Microplastics Compared with Ambient Environments (Water and Sediment) in Haihe Estuary.” Science of the Total Environment 708: 134876. 10.1016/j.scitotenv.2019.134876 [DOI] [PubMed] [Google Scholar]
- 41. Forero‐López, A. D. , Brugnoni L. I., Abasto B., Rimondino G. N., Lassalle V. L., Ardusso M. G., Nazzarro M. S., et al. 2022. “Plastisphere on Microplastics: In Situ Assays in an Estuarine Environment.” Journal of Hazardous Materials 440: 129737. 10.1016/j.jhazmat.2022.129737 [DOI] [PubMed] [Google Scholar]
- 42. Sun, Xuemei , Chen Bijuan, Xia Bin, Li Qiufen, Zhu Lin, Zhao Xinguo, Gao Yaping, and Qu Keming. 2020. “Impact of Mariculture‐Derived Microplastics on Bacterial Biofilm Formation and Their Potential Threat to Mariculture: A Case in Situ Study on the Sungo Bay, China.” Environmental Pollution 262: 114336. 10.1016/j.envpol.2020.114336 [DOI] [PubMed] [Google Scholar]
- 43. Rohrbach, Stephan , Gkoutselis Gerasimos, Hink Linda, Weig Alfons R., Obst Martin, Diekmann Astrid, Ho Adrian, Rambold Gerhard, and Horn Marcus A.. 2022. “Microplastic Polymer Properties as Deterministic Factors Driving Terrestrial Plastisphere Microbiome Assembly and Succession in the Field.” Environmental Microbiology. 10.1111/1462-2920.16234 [DOI] [PubMed] [Google Scholar]
- 44. Li, Huan‐Qin , Shen Ying‐Jia, Wang Wen‐Lei, Wang Hong‐Tao, Li Hu, and Su Jian‐Qiang. 2021. “Soil Ph Has a Stronger Effect than Arsenic Content on Shaping Plastisphere Bacterial Communities in Soil.” Environmental Pollution 287: 117339. 10.1016/j.envpol.2021.117339 [DOI] [PubMed] [Google Scholar]
- 45. Pinnell, Lee J. , and Turner Jeffrey W.. 2020. “Temporal Changes in Water Temperature and Salinity Drive the Formation of a Reversible Plastic‐Specific Microbial Community.” FEMS Microbiology Ecology 96. 10.1093/femsec/fiaa230 [DOI] [PubMed] [Google Scholar]
- 46. Chen, Xiao , LianLian Xiao‐ying, Wang Yi, Chen Sheng, Sun Yiran, Tao Guolin, Tan Qiaowen, and Feng Jingchun. 2023. “Impacts of Hydraulic Conditions on Microplastics Biofilm Development, Shear Stresses Distribution, and Microbial Community Structures in Drinking Water Distribution Pipes.” Journal of Environmental Management 325: 116510. 10.1016/j.jenvman.2022.116510 [DOI] [PubMed] [Google Scholar]
- 47. Yang, Yang , Li Teng, Liu Peng, Li Huixin, Hu Feng. 2022. “The Formation of Specific Bacterial Communities Contributes to the Enrichment of Antibiotic Resistance Genes in the Soil Plastisphere.” Journal of Hazardous Materials 436: 129247. 10.1016/j.jhazmat.2022.129247 [DOI] [PubMed] [Google Scholar]
- 48. Hidalgo‐Ruz, Valeria , Gutow Lars, Thompson Richard C., and Thiel Martin. 2012. “Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification.” Environmental Science & Technology 46: 3060–75. 10.1021/es2031505 [DOI] [PubMed] [Google Scholar]
- 49. Pinnell, Lee J. , and Turner Jeffrey W.. 2019. “Shotgun Metagenomics Reveals the Benthic Microbial Community Response to Plastic and Bioplastic in a Coastal Marine Environment.” Frontiers in microbiology 10: 1252. 10.3389/fmicb.2019.01252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Wen, Bin , Liu Jun‐Heng, Zhang Yuan, Zhang Hao‐Ran, Gao Jian‐Zhong, and Chen Zai‐Zhong. 2020. “Community Structure and Functional Diversity of the Plastisphere in Aquaculture Waters: Does Plastic Color Matter?” Science of the Total Environment 740: 140082. 10.1016/j.scitotenv.2020.140082 [DOI] [PubMed] [Google Scholar]
- 51. Oberbeckmann, Sonja , Bartosik Daniel, Huang Sixing, Werner Johannes, Hirschfeld Claudia, Wibberg Daniel, Heiden Stefan E., et al. 2021. “Genomic and Proteomic Profiles of Biofilms on Microplastics Are Decoupled from Artificial Surface Properties.” Environmental Microbiology 23: 3099–115. 10.1111/1462-2920.15531 [DOI] [PubMed] [Google Scholar]
- 52. Zhang, Xiaohan , Zhang Ying, Wu Nan, Li Wenjie, Song Xiaocui, Ma Yongzheng, and Niu Zhiguang. 2021. “Colonization Characteristics of Bacterial Communities on Plastic Debris: The Localization of Immigrant Bacterial Communities.” Water Research 193: 116883. 10.1016/j.watres.2021.116883 [DOI] [PubMed] [Google Scholar]
- 53. Zhang, Bin , Yang Xin, Liu Lingchen, Chen Liang, Teng Jia, Zhu Xiaopeng, Zhao Jianmin, and Wang Qing. 2021. “Spatial and Seasonal Variations in Biofilm Formation on Microplastics in Coastal Waters.” Science of the Total Environment 770: 145303. 10.1016/j.scitotenv.2021.145303 [DOI] [PubMed] [Google Scholar]
- 54. Li, Wenjie , Zhang Ying, Wu Nan, Zhao Ze, Xu Wei'an, Ma Yongzheng, and Niu Zhiguang. 2019. “Colonization Characteristics of Bacterial Communities on Plastic Debris Influenced by Environmental Factors and Polymer Types in the Haihe Estuary of Bohai Bay, China.” Environmental Science & Technology 53: 10763–73. 10.1021/acs.est.9b03659 [DOI] [PubMed] [Google Scholar]
- 55. Deng, Hui , Fu Qianqian, Zhang Yuqing, Li Dazhen, He Jianxiong, Feng Dan, Zhao Yuanyuan, Yu Huamei, and Ge Chengjun. 2022. “Bacterial Communities on Polyethylene Microplastics in Mangrove Ecosystems as a Function of Exposure Sites: Compositions and Ecological Functions.” Journal of Environmental Chemical Engineering 10: 107924. 10.1016/j.jece.2022.107924 [DOI] [Google Scholar]
- 56. Yang, Kai , Chen Qing‐Lin, Chen Mo‐Lian, Li Hong‐Zhe, Liao Hu, Pu Qiang, Zhu Yong‐Guan, and Cui Li. 2020. “Temporal Dynamics of Antibiotic Resistome in the Plastisphere during Microbial Colonization.” Environmental Science & Technology 54: 11322–32. 10.1021/acs.est.0c04292 [DOI] [PubMed] [Google Scholar]
- 57. Wright, Robyn J. , Gabriel Erni‐Cassola, Vinko Zadjelovic, Mira Latva, and Christie‐Oleza Joseph A.. 2020. “Marine Plastic Debris: A New Surface for Microbial Colonization.” Environmental Science & Technology 54: 11657–72. 10.1021/acs.est.0c02305 [DOI] [PubMed] [Google Scholar]
- 58. Zhang, Ying , Peng Yanwen, Peng Chu, Wang Ping, Lu Yuan, He Xiaosong, Wang Lei. 2021. “Comparison of Detection Methods of Microplastics in Landfill Mineralized Refuse and Selection of Degradation Degree Indexes.” Environmental Science & Technology 55: 13802–11. 10.1021/acs.est.1c02772 [DOI] [PubMed] [Google Scholar]
- 59. Zhao, Shiye , Zettler Erik R., Amaral‐Zettler Linda A., and Mincer Tracy J.. 2021. “Microbial Carrying Capacity and Carbon Biomass of Plastic Marine Debris.” The ISME Journal 15: 67–77. 10.1038/s41396-020-00756-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Schlundt, Cathleen , Mark Welch Jessica L., Knochel Anna M., Zettler Erik R., and Amaral‐Zettler Linda A.. 2020. “Spatial Structure in the “Plastisphere”: Molecular Resources for Imaging Microscopic Communities on Plastic Marine Debris.” Molecular ecology resources 20: 620–34. 10.1111/1755-0998.13119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Wang, Chengqian , Wang Liuwei, Ok Yong Sik, Tsang Daniel C. W., and Hou Deyi. 2022. “Soil Plastisphere: Exploration Methods, Influencing Factors, and Ecological Insights.” Journal of Hazardous Materials 430: 128503. 10.1016/j.jhazmat.2022.128503 [DOI] [PubMed] [Google Scholar]
- 62. Wang, Jie , Qin Xiao, Guo Jiabao, Jia Weiqian, Wang Qian, Zhang Mengjun, and Huang Yi. 2020. “Evidence of Selective Enrichment of Bacterial Assemblages and Antibiotic Resistant Genes by Microplastics in Urban Rivers.” Water Research 183: 116113. 10.1016/j.watres.2020.116113 [DOI] [PubMed] [Google Scholar]
- 63. Ogonowski, Martin , Motiei Asa, Ininbergs Karolina, Hell Eva, Gerdes Zandra, Udekwu Klas I., Bacsik Zoltan, and Gorokhova Elena. 2018. “Evidence for Selective Bacterial Community Structuring on Microplastics.” Environmental Microbiology 20: 2796–808. 10.1111/1462-2920.14120 [DOI] [PubMed] [Google Scholar]
- 64. Kettner, Marie Therese , Oberbeckmann Sonja, Labrenz Matthias, and Grossart Hans‐Peter. 2019. “The Eukaryotic Life on Microplastics in Brackish Ecosystems.” Frontiers in Microbiology 10: 538. 10.3389/fmicb.2019.00538 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Dudek, Kassandra L. , Cruz Bianca N., Polidoro Beth, and Neuer Susanne. 2020. “Microbial Colonization of Microplastics in the Caribbean Sea.” Limnology and Oceanography Letters 5: 5–17. 10.1002/lol2.10141 [DOI] [Google Scholar]
- 66. González‐Pleiter, Miguel , Velázquez David, Casero María Cristina, Tytgat Bjorn, Verleyen Elie, Leganés Francisco, Rosal Roberto, Quesada Antonio, and Fernández‐Piñas Francisca. 2021. “Microbial Colonizers of Microplastics in an Arctic Freshwater Lake.” Science of the Total Environment 795: 148640. 10.1016/j.scitotenv.2021.148640 [DOI] [PubMed] [Google Scholar]
- 67. De Tender, Caroline , Devriese Lisa I., Haegeman Annelies, Maes Sara, Vangeyte Jürgen, Cattrijsse André, Dawyndt Peter, and Ruttink Tom. 2017. “Temporal Dynamics of Bacterial and Fungal Colonization on Plastic Debris in the North Sea.” Environmental Science & Technology 51: 7350–60. 10.1021/acs.est.7b00697 [DOI] [PubMed] [Google Scholar]
- 68. Bowley, Jake , Baker‐Austin Craig, Porter Adam, Hartnell Rachel, and Lewis Ceri. 2021. “Oceanic Hitchhikers—Assessing Pathogen Risks from Marine Microplastic.” Trends in Microbiology 29: 107–16. 10.1016/j.tim.2020.06.011 [DOI] [PubMed] [Google Scholar]
- 69. Davidov, Keren , Iankelevich‐Kounio Evgenia, Yakovenko Iryna, Koucherov Yuri, Rubin‐Blum Maxim, Oren Matan. 2020. “Identification of Plastic‐Associated Species in the Mediterranean Sea Using DNA Metabarcoding With Nanopore MinION.” Scientific Reports 10: 17533. 10.1038/s41598-020-74180-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Qiang, Liyuan , Cheng Jinping, Mirzoyan Seda, Kerkhof Lee J., Häggblom Max M.. 2021. “Characterization of Microplastic‐Associated Biofilm Development along a Freshwater‐Estuarine Gradient.” Environmental Science & Technology 55: 16402–12. 10.1021/acs.est.1c04108 [DOI] [PubMed] [Google Scholar]
- 71. Wu, Xiaojian , Liu Zongbao, Li Meng, Bartlam Mark, and Wang Yingying. 2022. “Integrated Metagenomic and Metatranscriptomic Analysis Reveals Actively Expressed Antibiotic Resistomes in the Plastisphere.” Journal of Hazardous Materials 430: 128418. 10.1016/j.jhazmat.2022.128418 [DOI] [PubMed] [Google Scholar]
- 72. Wright, Robyn J. , Bosch Rafael, Langille Morgan G. I., Gibson Matthew I., and Christie‐Oleza Joseph A.. 2021. “A multi‐OMIC Characterisation of Biodegradation and Microbial Community Succession within the PET Plastisphere.” Microbiome 9: 141. 10.1186/s40168-021-01054-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Bryant, Jessica A. , Clemente Tara M., Viviani Donn A., Fong Allison A., Thomas Kimberley A., Kemp Paul, Karl David M., White Angelicque E., and DeLong Edward F.. 2016. “Diversity and Activity of Communities Inhabiting Plastic Debris in the North Pacific Gyre.” mSystems 1: e00024–00016. 10.1128/mSystems.00024-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Oberbeckmann, Sonja , Osborn A. Mark, and Duhaime Melissa B.. 2016. “Microbes on a Bottle: Substrate, Season and Geography Influence Community Composition of Microbes Colonizing Marine Plastic Debris.” PLoS ONE 11: e0159289. 10.1371/journal.pone.0159289 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Mughini‐Gras, Lapo , van der Plaats Rozemarijn Q. J., van der Wielen Paul W. J. J., Bauerlein Patrick S., and de Roda Husman Ana Maria. 2021. “Riverine Microplastic and Microbial Community Compositions: A Field Study in the Netherlands.” Water Research 192: 116852. 10.1016/j.watres.2021.116852 [DOI] [PubMed] [Google Scholar]
- 76. Miao, Lingzhan , Li Wanyi, Adyel Tanveer M., Yao Yu, Deng Ye, Wu Jun, Zhou Yongqiang, Yu Yue, and Hou Jun. 2023. “Spatio‐Temporal Succession of Microbial Communities in Plastisphere and their Potentials for Plastic Degradation in Freshwater Ecosystems.” Water Research 229: 119406. 10.1016/j.watres.2022.119406 [DOI] [PubMed] [Google Scholar]
- 77. Yang, Guiqin , Gong Mengting, Mai Lei, Zhuang Li, and Zeng Eddy Y.. 2021. “Diversity and Structure of Microbial Biofilms on Microplastics in Riverine Waters of the Pearl River Delta, China.” Chemosphere 272: 129870. 10.1016/j.chemosphere.2021.129870 [DOI] [PubMed] [Google Scholar]
- 78. Feng, Limin , He Lei, Jiang Shiqi, Chen Jinjun, Zhou Chunxia, Qian Zhong‐Ji, Hong Pengzhi, Sun Shengli, and Li Chengyong. 2020. “Investigating the Composition and Distribution of Microplastics Surface Biofilms in Coral Areas.” Chemosphere 252: 126565. 10.1016/j.chemosphere.2020.126565 [DOI] [PubMed] [Google Scholar]
- 79. Jiang, Peilin , Zhao Shiye, Zhu Lixin, and Li Daoji. 2018. “Microplastic‐Associated Bacterial Assemblages in the Intertidal Zone of the Yangtze Estuary.” Science of the Total Environment 624: 48–54. 10.1016/j.scitotenv.2017.12.105 [DOI] [PubMed] [Google Scholar]
- 80. Vaksmaa, Annika , Knittel Katrin, Abdala Asbun Alejandro, Goudriaan Maaike, Ellrott Andreas, Witte Harry J., Vollmer Ina, et al. 2021. “Microbial Communities on Plastic Polymers in the Mediterranean Sea.” Frontiers in Microbiology 12: 1021. 10.3389/fmicb.2021.673553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Ya, Haobo , Xing Yi, Zhang Tian, Lv Mingjie, and Jiang Bo. 2022. “LDPE Microplastics Affect Soil Microbial Community and form a Unique Plastisphere on Microplastics.” Applied Soil Ecology 180: 104623. 10.1016/j.apsoil.2022.104623 [DOI] [Google Scholar]
- 82. Li, Kang , Jia Weiqian, Xu Libo, Zhang Mengjun, and Huang Yi. 2023. “The Plastisphere of Biodegradable and Conventional Microplastics from Residues Exhibit Distinct Microbial Structure, Network and Function in Plastic‐Mulching Farmland.” Journal of Hazardous Materials 442: 130011. 10.1016/j.jhazmat.2022.130011 [DOI] [PubMed] [Google Scholar]
- 83. Zhang, Yu , Ma Jun, O'Connor Patrick, and Zhu Yong‐Guan. 2022. “Microbial Communities on Biodegradable Plastics Under Different Fertilization Practices in Farmland Soil Microcosms.” Science of the Total Environment 809: 152184. 10.1016/j.scitotenv.2021.152184 [DOI] [PubMed] [Google Scholar]
- 84. Li, Qihao , Tian Li, Cai Xunchao, Wang Yicheng, and Yanping Mao. 2022. “Plastisphere Showing Unique Microbiome and Resistome Different from Activated Sludge.” Science of the Total Environment 851: 158330. 10.1016/j.scitotenv.2022.158330 [DOI] [PubMed] [Google Scholar]
- 85. Zhang, Shuai , Liu Xingxiang, Qiu Pengxiang, Chen Bin, Xu Chenmin, Dong Weiliang, and Liu Tao. 2022. “Microplastics Can Selectively Enrich Intracellular and Extracellular Antibiotic Resistant Genes and Shape Different Microbial Communities in Aquatic Systems.” Science of the Total Environment 822: 153488. 10.1016/j.scitotenv.2022.153488 [DOI] [PubMed] [Google Scholar]
- 86. Agostini, Luana , Moreira Julio Cezar Fornazier, Bendia Amanda Gonçalves, Kmit Maria Carolina Pezzo, Waters Linda Gwen, Santana Marina Ferreira Mourão, Sumida Paulo Yukio Gomes, et al. 2021. “Deep‐Sea Plastisphere: Long‐Term Colonization by Plastic‐Associated Bacterial and Archaeal Communities in the Southwest Atlantic Ocean.” Science of the Total Environment 793: 148335. 10.1016/j.scitotenv.2021.148335 [DOI] [PubMed] [Google Scholar]
- 87. Bystrianský, Lukáš , Hujslová Martina, Hršelová Hana, Řezáčová Veronika, Němcová Lenka, Šimsová Jana, Gryndlerová Hana, et al. 2019. “Observations on Two Microbial Life Strategies in Soil: Planktonic and Biofilm‐Forming Microorganisms are Separable.” Soil Biology and Biochemistry 136: 107535. 10.1016/j.soilbio.2019.107535 [DOI] [Google Scholar]
- 88. Rickard, A. H. , McBain A. J., Ledder R. G., Handley P. S., and Gilbert P.. 2003. “Coaggregation Between Freshwater Bacteria within Biofilm and Planktonic Communities.” FEMS Microbiology Letters 220: 133–40. 10.1016/s0378-1097(03)00094-6 [DOI] [PubMed] [Google Scholar]
- 89. Booth, Sean C. , Workentine Matthew L., Wen Jing, Shaykhutdinov Rustem, Vogel Hans J., Ceri Howard, Turner Raymond J., and Weljie Aalim M.. 2011. “Differences in Metabolism between the Biofilm and Planktonic Response to Metal Stress.” Journal of Proteome Research 10: 3190–99. 10.1021/pr2002353 [DOI] [PubMed] [Google Scholar]
- 90. Salta, Maria , Wharton Julian A., Blache Yves, Stokes Keith R., and Briand Jean‐Francois. 2013. “Marine Biofilms on Artificial Surfaces: Structure and Dynamics.” Environmental Microbiology 15: 2879–93. 10.1111/1462-2920.12186 [DOI] [PubMed] [Google Scholar]
- 91. Neria‐González, Isabel , Wang En Tao, Ramírez Florina, Romero Juan M., and Hernández‐Rodríguez César. 2006. “Characterization of Bacterial Community Associated to Biofilms of Corroded Oil Pipelines from the Southeast of Mexico.” Anaerobe 12: 122–33. 10.1016/j.anaerobe.2006.02.001 [DOI] [PubMed] [Google Scholar]
- 92. Flemming, Hans‐Curt , and Wuertz Stefan. 2019. “Bacteria and Archaea on Earth and their Abundance In Biofilms.” Nature Reviews Microbiology 17: 247–60. 10.1038/s41579-019-0158-9 [DOI] [PubMed] [Google Scholar]
- 93. Miao, Lingzhan , Yu Yue, Adyel Tanveer M., Wang Chengqian, Liu Zhilin, Liu Songqi, Huang Liuyan, et al. 2021. “Distinct Microbial Metabolic Activities of Biofilms Colonizing Microplastics in Three Freshwater Ecosystems.” Journal of Hazardous Materials 403: 123577. 10.1016/j.jhazmat.2020.123577 [DOI] [PubMed] [Google Scholar]
- 94. Robert, Michel , and Chenu Claire. 2021. “Interactions Between Soil Minerals and Microorganisms.” Soil Biochemistry CRC Press: 307–404. [Google Scholar]
- 95. Gupta, Vadakattu V. S. R. , and Germida James J.. 2015. “Soil Aggregation: Influence on Microbial Biomass and Implications for Biological Processes.” Soil Biology and Biochemistry 80: A3–9. 10.1016/j.soilbio.2014.09.002 [DOI] [Google Scholar]
- 96. Rovira, A. D. 1965. “Interactions Between Plant Roots and Soil Microorganisms.” Annual Review of Microbiology 19: 241–66. 10.1146/annurev.mi.19.100165.001325 [DOI] [PubMed] [Google Scholar]
- 97. Qian, Pei‐Yuan , Cheng Aifang, Wang Ruojun, and Zhang Rui. 2022. “Marine Biofilms: Diversity, Interactions, and Biofouling.” Nature Reviews Microbiology 20: 671–84. 10.1038/s41579-022-00744-7 [DOI] [PubMed] [Google Scholar]
- 98. Ammar, Yasmine , Swailes David, Bridgens Ben, Chen Jinju. 2015. “Influence of Surface Roughness on the Initial Formation of Biofilm.” Surface and Coatings Technology 284: 410–16. 10.1016/j.surfcoat.2015.07.062 [DOI] [Google Scholar]
- 99. Wright, Robyn J. , Langille Morgan G. I., Walker Tony R.. 2020. “Food or Just a Free Ride? A Meta‐Analysis Reveals the Global Diversity of the Plastisphere.” The ISME Journal 15: 789–806. 10.1038/s41396-020-00814-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Ju, Zhicheng , Du Xiongfeng, Feng Kai, Li Shuzhen, Gu Songsong, Jin Decai, and Deng Ye. 2021. “The Succession of Bacterial Community Attached on Biodegradable Plastic Mulches During the Degradation in Soil.” Frontiers in microbiology 12. 10.3389/fmicb.2021.785737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Rüthi, Joel , Bölsterli Damian, Pardi‐Comensoli Lucrezia, Brunner Ivano, and Frey Beat. 2020. “The “Plastisphere” of Biodegradable Plastics is Characterized by Specific Microbial Taxa of Alpine and Arctic Soils.” Frontiers in Environmental Science 8: 562263. 10.3389/fenvs.2020.562263 [DOI] [Google Scholar]
- 102. Nemergut, Diana R. , Schmidt Steven K., Fukami Tadashi, O'Neill Sean P., Bilinski Teresa M., Stanish Lee F., Knelman Joseph E., et al. 2013. “Patterns and Processes of Microbial Community Assembly.” Microbiology and Molecular Biology Reviews 77: 342–56. 10.1128/MMBR.00051-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Zhou, Jizhong , and Ning Daliang. 2017. “Stochastic Community Assembly: Does it Matter in Microbial Ecology?” Microbiology and Molecular Biology Reviews 81: e00002–17. 10.1128/MMBR.00002-17 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Chave, J . 2004. “Neutral Theory and Community Ecology.” Ecology Letters 7: 241–53. 10.1111/j.1461-0248.2003.00566.x [DOI] [Google Scholar]
- 105. Stegen, James C. , Lin Xueju, Fredrickson Jim K., Chen Xingyuan, Kennedy David W., Murray Christopher J., Rockhold Mark L., and Konopka Allan. 2013. “Quantifying Community Assembly Processes and Identifying Features that Impose Them.” The ISME Journal 7: 2069–79. 10.1038/ismej.2013.93 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Ning, Daliang , Yuan Mengting, Wu Linwei, Zhang Ya, Guo Xue, Zhou Xishu, Yang Yunfeng, et al. 2020. “A Quantitative Framework Reveals Ecological Drivers of Grassland Microbial Community Assembly in Response to Warming.” Nature Communications 11(1): 4717. 10.1038/s41467-020-18560-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Ning, Daliang , Deng Ye, Tiedje James M, and Zhou Jizhong. 2019. “A General Framework for Quantitatively Assessing Ecological Stochasticity.” Proceedings of the National Academy of Sciences 116: 16892–98. 10.1073/pnas.1904623116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Sloan, William T. , Lunn Mary, Woodcock Stephen, Head Ian M., Nee Sean, and Curtis Thomas P.. 2006. “Quantifying the Roles of Immigration and Chance in Shaping Prokaryote Community Structure.” Environmental Microbiology 8: 732–40. 10.1111/j.1462-2920.2005.00956.x [DOI] [PubMed] [Google Scholar]
- 109. Li, Yongbin , Yang Rui, Guo Lifang, Gao Wenlong, Su Pingzhou, Xu Zhimin, Xiao Huan, et al. 2022. “The Composition, Biotic Network, and Assembly of Plastisphere Protistan Taxonomic and Functional Communities in Plastic‐Mulching Croplands.” Journal of Hazardous Materials 430: 128390. 10.1016/j.jhazmat.2022.128390 [DOI] [PubMed] [Google Scholar]
- 110. Delacuvellerie, Alice , Cyriaque Valentine, Gobert Sylvie, Benali Samira, Wattiez Ruddy. 2019. “The Plastisphere in Marine Ecosystem Hosts Potential Specific Microbial Degraders Including Alcanivorax Borkumensis as a Key Player for the Low‐Density Polyethylene Degradation.” Journal of Hazardous Materials 380: 120899. 10.1016/j.jhazmat.2019.120899 [DOI] [PubMed] [Google Scholar]
- 111. Joshi, Gajendra , Goswami Prasun, Verma Pankaj, Prakash Gopika, Simon Priya, Vinithkumar Nambali Valsalan, Dharani Gopal. 2022. “Unraveling the Plastic Degradation Potentials of the Plastisphere‐Associated Marine Bacterial Consortium as a Key Player for the Low‐Density Polyethylene Degradation.” Journal of Hazardous Materials 425: 128005. 10.1016/j.jhazmat.2021.128005 [DOI] [PubMed] [Google Scholar]
- 112. Bhagwat, Geetika , O'Connor Wayne, Grainge Ian, and Palanisami Thava. 2021. “Understanding the Fundamental Basis for Biofilm Formation on Plastic Surfaces: Role of Conditioning Films.” Frontiers in Microbiology 12: 687118. 10.3389/fmicb.2021.687118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Delacuvellerie, A. , Géron A., Gobert S., and Wattiez R.. 2022. “New Insights Into the Functioning and Structure of the PE and PP Plastispheres from the Mediterranean Sea.” Environmental Pollution 295: 118678. 10.1016/j.envpol.2021.118678 [DOI] [PubMed] [Google Scholar]
- 114. Li, Lu , Li Fangbai, Deng Min, Wu Chenxi, Zhao Xiaoli, Song Kang, and Wu Fengchang. 2022. “Microplastics Distribution Characteristics in Typical Inflow Rivers of Taihu Lake: Linking to Nitrous Oxide Emission and Microbial Analysis.” Water Research 225: 119117. 10.1016/j.watres.2022.119117 [DOI] [PubMed] [Google Scholar]
- 115. Huang, Jun‐Nan , Wen Bin, Miao Lin, Liu Xin, Li Zhong‐Jun, Ma Teng‐Fei, Xu Lei, Gao Jian‐Zhong, and Chen Zai‐Zhong. 2022. “Microplastics Drive Nitrification by Enriching Functional Microorganisms in Aquaculture Pond Waters.” Chemosphere 309: 136646. 10.1016/j.chemosphere.2022.136646 [DOI] [PubMed] [Google Scholar]
- 116. Su, Xiaoxuan , Yang Leyang, Yang Kai, Tang Yijia, Wen Teng, Wang Yingmu, Rillig Matthias C., et al. 2022. “Estuarine Plastisphere as an Overlooked Source of N2O Production.” Nature Communications 13: 3884. 10.1038/s41467-022-31584-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Rahman, Ishraq , Mujahid Aazani, Palombo Enzo A., and Müller Moritz. 2021. “A Functional Gene‐Array Analysis of Microbial Communities Settling on Microplastics in a Peat‐Draining Environment.” Marine Pollution Bulletin 166: 112226. 10.1016/j.marpolbul.2021.112226 [DOI] [PubMed] [Google Scholar]
- 118. Junaid, Muhammad , Siddiqui Junaid Ali, Sadaf Mamona, Liu Shulin, and Wang Jun. 2022. “Enrichment and Dissemination of Bacterial Pathogens by Microplastics in the Aquatic Environment.” Science of the Total Environment 830: 154720. 10.1016/j.scitotenv.2022.154720 [DOI] [PubMed] [Google Scholar]
- 119. Oberbeckmann, Sonja , and Labrenz Matthias. 2020. “Marine Microbial Assemblages on Microplastics: Diversity, Adaptation, and Role in Degradation.” Annual Review of Marine Science 12: 209–32. 10.1146/annurev-marine-010419-010633 [DOI] [PubMed] [Google Scholar]
- 120. MacLean, Joana , Mayanna Sathish, Benning Liane G., Horn Fabian, Bartholomäus Alexander, Wiesner Yosri, Wagner Dirk, and Liebner Susanne. 2021. “The Terrestrial Plastisphere: Diversity and Polymer‐Colonizing Potential of Plastic‐Associated Microbial Communities in Soil.” Microorganisms 9(9): 1876. 10.3390/microorganisms9091876 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Curren, Emily , and Leong Sandric Chee Yew. 2019. “Profiles of Bacterial Assemblages from Microplastics of Tropical Coastal Environments.” Science of the Total Environment 655: 313–20. 10.1016/j.scitotenv.2018.11.250 [DOI] [PubMed] [Google Scholar]
- 122. Yang, Yuyi , Liu Guihua, Song Wenjuan, Ye Chen, Lin Hui, Li Zan, Liu Wenzhi. 2019. “Plastics in the Marine Environment are Reservoirs for Antibiotic and Metal Resistance Genes.” Environment International 123: 79–86. 10.1016/j.envint.2018.11.061 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information.
Supporting information.
Data Availability Statement
This manuscript does not generate any code or data. Supplementary materials (figures, tables, scripts, graphical abstract, slides, videos, Chinese translated version and update materials) may be found in the online DOI or iMeta Science http://www.imeta.science/.