

Abstract
Xylose is an abundant bioresource for obtaining diverse chemicals and added-value products. The production of xylose from green alternatives like enzymatic hydrolysis is an important step in a biorefinery context. This research evaluated the synergism among four classes of hydrolytic purified enzymes—endo-1,4-β-xylanase, α-l-arabinofuranosidase, β-xylosidase, and α-d-glucuronidase—over hydrolysis of glucuronoarabinoxylan (GAX) obtained from brewers’ spent grain (BSG) after alkaline extraction and ethanol precipitation. First, monosaccharides, uronic acids and glycosidic-linkages of alkaline extracted GAX fraction from BSG were characterized, after that different strategies based on the addition of one or two families of enzymes—endo-1,4-β-xylanase (GH10 and GH11) and α-l-arabinofuranosidase (GH43 and GH51)—cooperating with one β-xylosidase (GH43) and one α-d-glucuronidase (GH67) into enzymatic hydrolysis were assessed to obtain the best yield of xylose. The xylose release was monitored over time in the first 90 min and after a prolonged reaction up to 48 h of reaction. The highest yield of xylose was 63.6% (48 h, 40 ℃, pH 5.5), using a mixture of all enzymes devoid of α-l-arabinofuranosidase (GH43) family. These results highlight the importance of GH51 arabinofuranosidase debranching enzyme to allow a higher cleavage of the xylan backbone of GAX from BSG and their synergy with 2 endo-1,4-β-xylanase (GH10 and GH11), one β-xylosidase (GH43) and the inclusion of one α-d-glucuronidase (GH67) in the reaction system. Therefore, this study provides an environmentally friendly process to produce xylose from BSG through utilization of enzymes as catalysts.
Graphical Abstract

Keywords: Brewers’ spent grain, Glucuronoarabinoxylan, Arabinoxylan, Xylan saccharification, Synergism, Xylanolytic enzymes, Enzymatic hemicellulose hydrolysis
Introduction
In the context of biorefining, the utilization of hemicellulose as the second most abundant sugar component of lignocellulosic biomass is highly desired, hemicellulose can be transformed into oligosaccharides composed of xylose or in monomeric sugars, mainly xylose. Xylooligosaccharides (XOS) are considered prebiotic compounds presenting technical and health claims (Poletto et al. 2020; Swart et al. 2020). Xylose derived products including xylitol, ethanol, isobutanol, lactic acid or lipid and its derivatives by biological technologies (Long et al. 2022). Agricultural crop and industrial residues have been explored to obtain hemicellulose between them corncob, corn stover, wheat straw and the main byproduct of beer production known as brewers’ spent grain (BSG). BSG is a complex biomass composed of starch (2–13%), cellulose (13–21%), hemicellulose (19–42%) and lignin (12–16%), with a high protein content (19–30%) (Lynch et al. 2016; Rojas-Pérez et al. 2022).
Methods for converting plant material into monomeric sugars are required for their further use as feedstocks in the production of a range of value-added products, these procedures comprise pretreatment and enzymatic hydrolysis (EH) steps (Gonçalves et al. 2022). Pretreatment of lignocellulosic biomass is crucial prior to enzymatic hydrolysis. Various pretreatment options have been reported to fractionate, solubilize, hydrolyze, and separate cellulose, arabinoxylan (AX), and lignin components (Saha 2003). Specifically, AX can be extracted using water or hydrothermal pretreatment (Kaur et al. 2020), alkaline pretreatment (Pérez-Flores et al. 2019), ultrasound-assisted extraction (Reis et al. 2015) and microwave (Coelho et al. 2014). The AX of BSG are usually extracted by a well-established sequentially procedure using solutions of KOH with increasing concentration of 0.5 M, 1 M, and 4 M follow for ethanol precipitation (Mandalari et al. 2005; Vieira et al. 2014). Efficient hydrolysis of hemicellulose is a major challenge to the enzymatic conversion of lignocellulose (Liu et al. 2021). The conversion of AX into monosaccharides or derived compounds can be achieved by enzymatic or chemical catalysis, with the latter using metal oxides (Naidu et al. 2018). Although AX are readily hydrolyzed to monosaccharides by acid treatment, enzymatic hydrolysis is preferable for the industrial upgrading of AX, mainly because enzymatic hydrolysis permits specific, controlled modifications of the reaction that prevent the generation of undesirable byproducts and generally facilitate more environmentally friendly processes (Sørensen et al. 2007a, b; Meyer et al. 2009).
AX are characterized by a backbone of (β1 → 4)-linked d-xylopyranosyl residues partially substituted with single units of α-l-arabinofuranosyl at positions 2, 3, or both (Moreirinha et al. 2020). Phenolic acids, such as ferulic and p-coumaric acids, are esterified to arabinofuranosyl residues (Mandalari et al. 2005). Coelho et al. (2016) also identified other substituents in AX from BSG, such as uronic acid, methylated uronic acid, and an acetyl group, showing the presence of GAX in BSG. Due to the complex branching and heterogeneous composition of GAX, enzymatic degradation requires the synergistic action of depolymerizing and debranching enzymes. Depolymerization relies on endo-1,4-β-xylanases (EC 3.2.1.8) and β-xylosidases (EC 3.2.1.37). Endo-1,4-β-xylanases randomly attack the 1,4-β bonds within the xylan backbone to generate unsubstituted or branched xylo-oligosaccharides (XOS) and xylobiose, whereas β-xylosidases attack the non-reducing ends of short-chain xylo-oligosaccharides and xylobiose to release xylose (Sørensen et al. 2007a, b). Debranching enzymes mainly include α-l-arabinofuranosidases (EC 3.2.1.55), α-d-glucuronidases (EC 3.2.1.139), ferulic acid esterases (EC 3.1.1.73), and/or acetyl xylan esterases (EC 3.1.1.72).
Commercial cellulase–hemicellulase mixtures, enzyme extract from microorganisms, high purity enzymes or the combination between them had been probed in different hemicellulose fraction extracted from lignocellulosic biomass. Xiros et al. (2008) for example, evaluated a multi-enzymatic system (xylanase, endoglucanase, cellobiohydrolase, β-glucosidase, α-l-arabinofuranosidase, acetyl esterase, and feruloyl esterase) from N. crassa over BSG and alkali pre-treated BSG, found that enzymatic hydrolysis of alkali pre-treated BSG was increased about 50% compared with non-pretreated material and the released sugars (glucose, xylose, arabinose) from pre-treated BSG using the enzyme extract from N. crassa achieved a yield about 50% of total pentose content and about 60% of total glucose in the material. On the other hand, prolonged treatment (24 h with a 50:50 mixture of Celluclast® 1.5 L and Ultraflo® L at 50 °C, pH 5) in water-soluble wheat arabinoxylan achieve a xylose release of 62 wt% (Sørensen et al. 2005). Another work from Sørensen et al (2007a, b) found that 114.5% of xylose was released from water-soluble wheat AXs with a “minimal” enzyme 20:20:20:40 mixture of Abf II (α-l-arabinofuranosidase H. insolens GH43 family), Abf III (α-l-arabinofuranosidase M. giganteus GH51 family), Xyl III (endo-1,4-β-xylanase H. insolens GH10 family), and β-xyl (β-xylosidase T. reesei GH3 family), although the yield above 100% of the “theoretical maximum” presumably resulted from the degradation of arabinose and xylose during the acid hydrolysis. Mccleary et al. (2015) determined empirically an optimal enzyme mixture that contained β-xylanase (1300 U/mL), β-xylosidase (200 U/mL), B. adolescentis α-l-arabinofuranosidase (300 U/mL), U. maydis α-l-arabinofuranosidase (75 U/mL), and A. niger α-l-arabinofuranosidase (170 U/mL) as high purity enzymes over water-soluble wheat flour arabinoxylans (WAXs) and found a yield of 90% of l-arabinose and d-xylose release, although the addition of more enzymes did not increase the yield, suggesting the presence of other groups in the arabinoxylan, such as ferulic acid, which prevent complete hydrolysis of WAX to monosaccharides. Newly, Long et al. (2022) evaluated the hydrolysis of corncob arabinoxylans with low (CAX1) or high (CAX2) branching degrees reported that a new arabinofuranosidase EpABF62A of the GH62 family combined with a GH10 xylanase, a GH43 β-D-xylosidase and a GH67 α-glucuronidase released 75.0% or 64.5% xylose from CAX1 or CAX2, respectively.
The aim of this research was to evaluate different strategies based on the addition of one or two families of enzymes—endo-1,4-β-xylanase (GH10 and GH11) and α-l-arabinofuranosidase (GH43 and GH51)—cooperating with one β-xylosidase (GH43) and one α-d-glucuronidase (GH67)—over hydrolysis of GAX fraction (4 M KOH and ethanol precipitation) from BSG monitoring the time in the first 90 min and after a prolonged reaction up to 48 h of reaction, to obtain xylose as monomeric sugar.
Materials and methods
Extraction of GAX from BSG
A freeze-dried GAX from BSG were used as the raw material. This fraction was obtained from a supernatant solution by centrifugation from cellulosic residue after 4 step in a sequential alkaline extraction (4 M KOH + 5 mM Na2S2O5), subsequently acidified to pH 3 with citric acid to permit the precipitation of BSG proteins, the fraction soluble in citric acid, which contain the GAX were finally separated with ethanol precipitation, following the procedure described by Vieira et al. (2014).
Structural analysis of GAX extract from BSG
Monosaccharide composition analysis
Monosaccharides were released from cell wall polysaccharides by prehydrolysis with 0.2 mL of 72% H2SO4 (w/w) for 3 h at room temperature, followed by 2.5-h hydrolysis with 1 M of H2SO4 at 100 ℃ (Selvendran et al. 1979). Neutral sugars were analyzed to determine their alditol acetate content using gas chromatography flame ionization detection (GC-FID) (Blakeney et al. 1983; Harris et al. 1988). Duplicate hydrolysis was performed for all fractions.
Uronic acids
Uronic acids were quantified using the 3-phenyl phenol colorimetric method and the calibration curve for galacturonic acid (200 mg/mL) (Selvendran et al. 1979; Coimbra et al. 1996).
Glycosidic-linkage composition of the polysaccharide fraction
The glycosidic-linkage composition was determined using gas chromatography quadrupole mass spectrometry (GC–qMS) for the partially methylated alditol acetates, as described by Coelho et al. (2014) and Reis et al. (2015), using CH3I as a methylating reagent. Duplicate methylation was performed for all fractions.
Hydrolysis of GAX from BSG
Enzymes
Six pure enzymes were obtained from Megazyme International (Bray, County Wicklow, Ireland). Their families, substrates, and main characteristics are described in Table 1.
Table 1.
Summary of enzymes characteristics
| Enzyme | CAZy family | Microorganism | Substrate | T optima (℃) | pH optima |
|---|---|---|---|---|---|
|
endo-1,4-β-Xylanase (EC 3.2.1.8) |
GH11 | Neocallimastix patriciarum | Endo-hydrolysis of (1,4)-β-d-xylosidic linkages in xylans | 50 | 6.0 |
| GH10 | Cellvibrio japonicus | 60 | 5.0 | ||
|
α-l-Arabinofuranosidase (EC 3.2.1.55) |
GH43 | Bifidobacterium adolescentis | Highly specific hydrolysis of α-1,3-linked l-arabinofuranose residues from doubly substituted d-xylosyl or l-arabinosyl residues of arabinoxylans and branched arabinans, respectively | 50 | 6.0 |
| GH51 | Aspergillus niger | Hydrolysis of α-1,2- and α-1,3-linked l-arabinofuranose residues from arabinoxylans and branched arabinans. Hydrolyses α-1,5-linked arabino-oligosaccharides at a much lower rate | 40 | 4.0 | |
|
exo-1,4-β-d-Xylosidase (EC 3.2.1.37) |
GH43 | Selenomonas ruminantium | Hydrolysis of (1,4)-β-d-xylans and xylo-oligosaccharides to remove successive d-xylose residues from non-reducing termini | 50 | 5.0 |
|
α-d-Glucuronidase (EC 3.2.1.139) |
GH67 | Geobacillus stearothermophilus | Hydrolysis of the α-1,2 glycosidic bond between d-glucuronic acid or its ether 4-O-methyl-d-glucuronic acid from the terminal non-reducing d-xylose residues of xylo-oligosaccharides (aldo-uronic acids) and xylan | 70 | 7.0 |
Enzyme combinations
The reaction mixture was adjusted to a concentration of 1 g/L of GAX (4 M KOH) from BSG in 100 mM of sodium succinate buffer (pH 5.5) at temperature of 40 °C. A fixed dose of 0.2 mg of enzyme-protein/g potential GAX mixture was added, which contained N. patriciarum endo-1,4-β-xylanase GH11 (1300 U/mL), C. japonicus endo-1,4-β-xylanase GH10 (500 U/mL), B. adolescentis α-l-arabinofuranosidase GH43 (300 U/mL), A. niger α-l-arabinofuranosidase GH51 (170 U/mL), S. ruminantium β-d-xylosidase GH43 (200 U/mL), and G. stearothermophilus α-d-glucuronidase GH 67 (200 U/mL). For pH (5.5) and enzyme doses, the research adopted the values reported by Mccleary et al. (2015). The reaction temperature (40 °C) was established as a balance of the optimal temperatures for the six pure enzymes evaluated, considering both the optimal values reported in the enzyme datasheets and the references in the literature for this type of enzyme reaction (Xiros et al. 2011; Rasmussen et al. 2012; Mccleary et al. 2015). Table 2 shows the nomenclatures and definitions of the enzymes used in each test.
Table 2.
Definition of the addition of enzymes and nomenclature of the tests carried out to evaluate the synergistic effect
| Assay | Mixture enzyme | Enzymes present in the assay | Abbreviations |
|---|---|---|---|
| I | All | All enzymes | all |
| II | Without one enzyme | All enzymes devoid of endo-1,4-β-xylanase (GH11) | all-XGH11 |
| III | All enzymes devoid of endo-1,4-β-xylanase (GH10) | all-XGH10 | |
| IV | All enzymes devoid of α-l-arabinofuranosidase (GH43) | all-AGH43 | |
| V | All enzymes devoid of α-l-arabinofuranosidase (GH51) | all-AGH51 | |
| VI | Without two enzymes | All enzymes devoid of endo-1,4-β-xylanase (GH11) and α-l-arabinofuranosidase (GH43) | [all-(XGH11 and AGH43)] |
| VII | All enzymes devoid of endo-1,4-β-xylanase (GH11) and α-l-arabinofuranosidase (GH51) | [all-(XGH11 and AGH51)] | |
| VIII | All enzymes devoid of endo-1,4-β-xylanase (GH10) and α-l-arabinofuranosidase (GH43) | [all-(XGH10 and AGH43)] | |
| IX | All enzymes devoid of endo-1,4-β-xylanase (GH10) and α-l-arabinofuranosidase (GH51) | [all-(XGH10 and AGH51)] |
The xylose release was monitored to evaluate the effect of the enzymes with different families each one endo-1,4-β-xylanase and α-l-arabinofuranosidase. For this analysis, a progress curve was divided into two stages. The first stage covered the monitoring of the reaction from time 0 to 90 min of reaction, taking samples at 10, 20, 40, 60, and 90 min, and the second stage continued the reaction for up to 48 h, taking samples after 4, 18, 24, and 48 h. Samples were heated at 100 °C for 10 min to stop the enzymatic reaction. All trials were performed in duplicate, and the mean difference was statistically assessed using Tukey’s test. Release xylose was determined by sugar analysis derivatized as alditol acetates (2.3.3), at least in duplicate. The xylose yield was calculated using Eq. 1:
| 1 |
Sugar analysis after enzymatic hydrolysis
The released monosaccharides in supernatant enzymatic hydrolysis were analyzed as their alditol acetates by gas chromatography (Coimbra et al. 1996; Selvendran et al. 1979) using a FISONS 8340 chromatograph with a split injector (split ratio 1:60) and a FID detector. A DB-225 column (Agilent J and W, USA; 30 m × 0.25 mm × 0.15 Lm) was used. The injector and detector temperatures were 220 and 230 ℃, respectively. The oven temperature program started at 200–220 ℃ at a rate of 40 ℃ per min and was held at 220 ℃ for 15 min, then increased up to 230 °C with a rate of 20 ℃ per min and was held at 230 ℃ for 1 min. The flow rate of the carrier gas (H2) was set at 1 mL/min at 200 ℃. Using this technique, the xylose concentration is reported quantitatively, while the detection of arabinose was performed only qualitatively, reporting the presence or not of this sugar.
Results and discussion
Sugar and glycosidic-linkage composition of GAX extracted from BSG
GAX fraction contains 889 g/kg of carbohydrates, mainly neutral sugars, and uronic acids. Figure 1 shows the sugar composition of GAX extracted in 4 M of KOH from BSG on a free base of proteins and phenolic compounds, because these compounds were not characterized in this work.
Fig. 1.
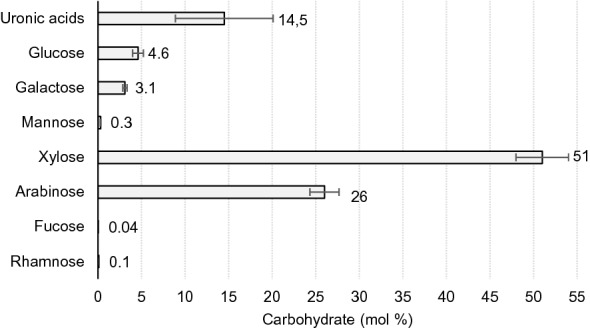
Carbohydrate composition of GAX fraction obtained by extraction of BSG with 4 M KOH
The carbohydrate was composed mainly of xylose (51% mol) and arabinose (26% mol). In this study, the quantification of uronic acids (14.5% mol) confirm the existence of glucuronoarabinoxylans in BSG, previously demonstrated qualitatively by Coelho et al. (2016) using electrospray ionization mass spectrometry (ESI-MS). The uronic acids amount found in this GAX were similar to those of steamed birchwood (11.5 mol %) (Kormelink and Voragen 1993). The GAX fraction of BSG also contains amounts of galactose (3.1% mol) and glucose (4.6% mol), this last probably due to the presence of residual starch (Reis et al. 2015) after the beer production. The remaining 11.1% (unknown) of the GAX surely corresponds to proteins, and phenolic acids. Proteins was already described to account 6–17% in AX extracted with 0.5 to 4 M KOH in BSG (Reis et al. 2015; Sajib et al. 2018). The presence of phenolic acids such p-coumaric and/or ferulic acid groups that could have remained esterified with arabinose in the branches of the main chain could also account with a small percentage to the extract; the values can range from 0.5 to 1.2 g/kg for ferulic acid and 0.1 g/kg for p-coumaric acid (Mandalari et al. 2005; Sajib et al. 2018).
According to part of the plant and the extraction method used, the AX fractions obtained can have different compositions, and consequently, their properties may differ (Biely et al. 2016; Bastos et al. 2018). Depending on the structural features of the xylan, different combinations of enzymes are required for maximum degradation (Kormelink and Voragen 1993). The structural features of GAX extracted from BSG were examined using methylation analysis (Fig. 2). Coelho et al. (2016) established the way to estimate the degree of polymerization (DP) of the AX is based on the principle that Xylp does not occur as AX branching residues. On this assumption, the DP is obtained by the calculation of the relative amount of total xylose divided by the amount of terminally linked xylose, for the alkaline fraction used in this work the GAX from BSG was mainly composed by a backbone of 15 residues. From the results of linkage analysis, it can be deduced that the general structure of the xylans from GAX alkaline extracted (4 M KOH) from BSG consists of a linear backbone of (1 → 4)-linked-d-xylopyranosyl units (Xylp) (42.3%). The degree of xylan main chain substitution by l-Araf and MeGlcA varies and depends heavily on the extraction conditions and part of the plant used (Biely et al. 2016). In this GAX fraction from BSG the branched regions of the xylan backbone had monosubstitutions at O-2 (16.1% mol), 2-Araf linkage was 3.2% indicates that α-1,2-linkage of 4-O-methyl-d-glucuronosyl units MeGlcA to the main chain Xylp is approximately the remaining 12.9%, highlighting the need to use an α-glucuronidase enzyme to achieve complete hydrolysis. Arabinose occupied the substitution in 3-Xylp position (3.1%). GAX also have disubstitutions at the 2 and 3 positions (6.1% mol). A lower degree of branching for arabinose was found, Ara:Xyl = 0.51, but higher for ratio of MeGlcA:Xyl = 3.57, showing an increase of this acid substitutions in the structure of the alkaline fraction extracted. Value similar was reported by Mandalari et al. (2005) for the fraction of BSG treated with cold water, 4 °C, 2 h (3.72) but lower than reported for the fraction treated with KOH 4.0 M, 2 h (13.95), although the fractionation protocol made by Vieira et al. (2014) was not as extensive as that reported by Mandalari et al. (2005).
Fig. 2.
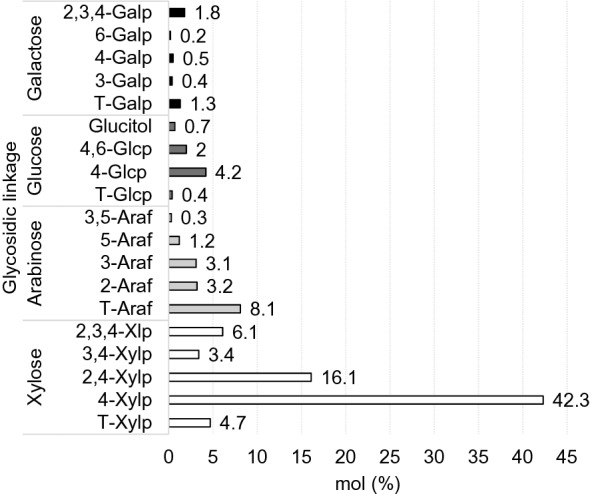
Glycoside linkage composition (mol %) of the GAX fraction obtained from BSG with 4 M KOH
Comparison of enzymatic synergistic interactions
Nine scenarios were used to evaluate xylose release by cooperation of six purified enzymes (Table 1). In the first assay, a mix of all enzymes (I) was assessed. In assays II–V, one family enzyme (endo-1,4-β-xylanase or α-l-arabinofuranosidase) was devoid, and for assays VI–IX, two enzymes of one family of endo-1,4-β-xylanase and one family of α-l-arabinofuranosidase were eliminated simultaneously according to the definition in Table 2. Figure 3A shows the release xylose where Test I (all enzymes) is compared with assays where one endo-1,4-β-xylanase (test II and III) or one α-l-arabinofuranosidase (test IV to V) family enzyme was omitted throughout 48 h of reaction. In general, the highest xylose release 0.36 g/L achieved by the test IV (all-AGH43) after 48 h of reaction, was equivalent to a hydrolysis yield of 63.6%, whereas in test I (all), the yield was 52.9%. In assays II (all-XGH11), III (all-XGH10), and V (all-AGH51), the yield was below 50%.
Fig. 3.

Xylose release from enzymatic hydrolysis. A Assays with all enzymes (I) versus assays without one enzyme (II to V). B Assays with all enzymes versus assays without two enzymes (VI to IX). Values represent the mean of two independent experiments, and error bars correspond to the standard deviations (SDs)
These results highlight the stronger synergistic effect of GH51 over GH43 arabinofuranosidase family as debranching enzyme to obtain xylose from GAX of BSG and showed the synergism with α-glucuronidase GH67 added in the multi-enzymatic system evaluated. α-l-Arabinofuranosidase from family GH51 only attacks the (1→2) or (1→3) bonds on singly substituted xylopyranosyls and hence directly provides unsubstituted xylopyranosyls while α-l-arabinofuranosidase from family GH43 only attacks the (1→3) linked arabinose on doubly substituted xylopyranosyls. After removing the (1→3) linked arabinose, the β-xylosidase still cannot work on the xylan backbone, because the α-(1→2) linked arabinose blocks the binding site (Rasmussen et al. 2012). Moreover α-d-glucuronidase attacks the α-1,2 glycosidic bond between d-glucuronic acid or its ether 4-O-methyl-d-glucuronic acid from the terminal non-reducing d-xylose residues of xylo-oligosaccharides (aldo-uronic acids) and xylan. In turn this enhances the probability of more unsubstituted xylopyranosyls at (or near) the non-reducing ends for β-xylosidase to attack. Rasmussen et al. (2012) also compared the xylose release but from soluble wheat arabinoxylan for 4 h, at this time they found similar behavior from the combined treatments of xylanase + β-xylosidase + α-l-arabinofuranosidase GH51 (AFAn) and the total combination of xylanase-β-xylosidase-α-l-arabinofuranosidase GH51 (AFAn) + α-l-arabinofuranosidase GH43 (AFBa). In this work a similar pattern was obtained up to 24 h between test I (all) and IV (all-AGH43), but after 24 h increased xylose release in the assay IV due to the presence of one α-d-glucuronidase of the family GH67. Long et al. (2022) also noted, that supplementation of the GH67 α-glucuronidase significantly enhanced (by 20–40%) the liberation of xylose from arabinoxylan of corncob during the enzyme hydrolysis (24 h), suggesting that MeGlcA substitution as an important limit factor of enzymatic saccharification remained in the substrate after the alkaline hydrogen peroxide pretreatment.
Figure 3A also shows the details of the first 90 min of the reaction in the left part of the figure. In these first 90 min of reaction, assays I (all), III (all-XGH10), IV (all-AGH43), and V (all-AGH51) showed a release of about 50% of the maximum xylose. Only assay II (all-XGH11) showed a release of 50% of the maximum xylose in less time (~ 10 min of reaction). The elimination of enzymatic activity related to endo-1,4-β-xylanases and α-l-arabinofuranosidases influenced the xylose yield over time. The results obtained showed that eliminating one of the endo-1,4-β-xylanases (GH10 or GH11 family) had different xylose release in the initial reaction rate (90 min). However, after 18 h and up to 48 h of reaction, assays II and III revealed no significant differences, showing the cooperation between these 2 kinds of xylanases. For the arabinose release presence of arabinose was evidenced at the first 90 min of the reaction for the assays I, II, III and IV, while for the assay V was not evident the arabinose presence corroborating the action of AGH51 not only to xylose release, but also in the arabinose release. Rasmussen et al. (2012) shows that the addition of xylanase and β-xylosidase each contributed to increase the arabinose release to different extents. However, neither xylanase nor β-xylosidase alone catalyzed the release of arabinose without the presence of arabinofuranosidase.
Figure 3B shows the xylose release in assays VI to IX, which omitted one endo-1,4-β-xylanase and one α-l-arabinofuranosidase simultaneously, compared with assay I (all enzymes present). The highest xylose release was 0.27 g/L achieved for assay I was equivalent to a hydrolysis yield of 52.9% compared with assays VI [all-(XGH11 and AGH43)] and VII [(all-(XGH11 and AGH51)], the yield for which was ~ 40%, whereas for assays VIII [all-(XGH10 and AGH43)] and IX [all-(XGH10 and AGH51)], the yield was ~ 30%. The results confirmed that the GH10 endo-1,4-β-xylanase family releases more xylose than the GH11 family; nevertheless, when they are at the same time this effect is not evident. The effect of eliminating GH43 or GH51 α-l-arabinofuranosidase families was less evident because the elimination of an endo-1,4-β-xylanase family was more important in the reaction. Rasmussen et al. (2012) inclusive indicated that the breaking down of the xylan backbone to smaller oligosaccharides, and thus exposing more (unsubstituted) non-reducing ends for β-xylosidase, is more important than removing the substituted arabinose to release more xylose from arabinoxylan.
The maximum yield (63.6%) reached in this work with the assay IV (endo-xylanase GH10 + endo-xylanase GH11 + α-L-arabinofuranosidase GH51 + α-glucuronidase GH67) is comparable to other arabinoxylan hydrolyzed enzymatically (Rasmussen et al. 2012; Long et al. 2022), but also show the necessity of supply the enzyme supplement with other accessory enzymes such as feruloyl esterase and α-l-galactosidase (Forssell et al. 2008; Mccleary et al. 2015; Biely et al. 2016; Long et al. 2022) and with other families of the α-glucuronidase such a GH115 to try increase the xylose release. Finally, due to the relevance of α-glucuronidase GH67 activity in the xylose release, it is important to probe other conditions of temperature and pH closest to the pH and temperature optima of this key enzyme.
Conclusion
This research studied the action of four debranching and depolymerizing hydrolytic purified enzymes on GAX from BSG, quantifying the xylose released in the process. An enzyme mixture of two endo-xylanases (from the GH10 and GH11 families), α-l-arabinofuranosidase GH51, β-xylosidase GH43, and α-d-glucuronidase GH67, showed the best cooperation, with 63.6% maximum xylose yield in 48 h (40 °C, pH 5.5). Although α-d-glucuronidase shows a key cooperation in the multi-enzymatic system evaluated, it is also recommended to use a feruloyl esterase and galactosidase to increase the yield of xylose production in futures studies.
Acknowledgements
LCRP greatly acknowledge the financial support provided by Universidad Nacional de Colombia DIB (Researches Division Campus Bogotá, project 35822). Thanks are also due to LAQV/REQUIMTE (UIDB/50006/2020, UIDP/50006/2020) through national founds and, where applicable, co-financed by the FEDER, within the PT2020 Partnership Agreement. EC thanks the research contract (CDL-CTTRI-88-ARH/2018–REF. 049-88-ARH/2018) funded by national funds (OE), through FCT, in the scope of the framework contract foreseen in the numbers 4, 5 and 6 of the article 23, of the Decree-Law 57/2016, of August 29, changed by Law 57/2017, of July 19.
Abbreviations
- Araf
L-Arabinofuranosyl residue
- AX
Arabinoxylan
- BSG
Brewers’ spent grain
- DP
Degree of polymerization
- EH
Enzymatic hydrolysis
- ESI-MSn
Electrospray ionization mass spectrometric detection
- GAX
Glucuronoarabinoxylan
- GC-FID
Gas chromatography flame ionization detection
- GC–qMS
Gas chromatography quadrupole mass spectrometry
- GlcA
D-Glucuronic acid
- MeGlcA
4-O-Methyl-d-glucuronic acid
- WAX
Wheat flour arabinoxylan
- XOS
Xylooligosaccharides
- Xyl
Xylose
- Xylp
D-Xylopyranosyl residue
Author contributions
LCRP designed and conducted all experiments, collected data, and prepared the manuscript, EC and MAMR reviewed the manuscript, PCNR and MAC supervised the research and revised the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by Ministry of Science, Technology, and Innovation (formerly known as COLCIENCIAS) for financial support (Grant 617-2014).
Availability of data and materials
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Bastos R, Coelho E, Coimbra MA. Arabinoxylans from cereal by-products: insights into structural features, recovery, and applications. In: Galanakis C, editor. Sustainable recovery and reutilization of cereal processing by-products. Amsterdam: Elsevier; 2018. [Google Scholar]
- Biely P, Singh S, Puchart V. Towards enzymatic breakdown of complex plant xylan structures: state of the art. Biotechnol Adv. 2016;34(7):1260–1274. doi: 10.1016/j.biotechadv.2016.09.001. [DOI] [PubMed] [Google Scholar]
- Blakeney AB, Harris PJ, Henry RJ, Stone BA. A simple and rapid preparation of alditol acetates for monosaccharide analysis. Carbohyd Res. 1983;113(2):291–299. doi: 10.1016/0008-6215(83)88244-5. [DOI] [Google Scholar]
- Coelho E, Rocha MAM, Saraiva JA, Coimbra MA. Microwave superheated water and dilute alkali extraction of brewers ’ spent grain arabinoxylans and arabinoxylo-oligosaccharides. Carbohyd Polym. 2014;99:415–422. doi: 10.1016/j.carbpol.2013.09.003. [DOI] [PubMed] [Google Scholar]
- Coelho E, Rocha MAM, Moreira ASP, Domingues MRM, Coimbra MA. Revisiting the structural features of arabinoxylans from brewers’ spent grain. Carbohyd Polym. 2016;139:167–176. doi: 10.1016/j.carbpol.2015.12.006. [DOI] [PubMed] [Google Scholar]
- Coimbra MA, Delgadillo I, Waldron KW, Selvendran RR (1996) Isolation and analysis of cell wall polymers from olive pulp. Anal Biochem 54(2):484–489
- Forssell P, Kontkanen H, Schols HA, Hinz S, Eijsink VGH, Treimo J, Robertson JA, Waldron KW, Faulds CB, Buchert J. Hydrolysis of brewers’ spent grain by carbohydrate degrading enzymes. J Inst Brew. 2008;114(4):306–314. doi: 10.1002/j.2050-0416.2008.tb00774.x. [DOI] [Google Scholar]
- Gonçalves MCP, Romanelli JP, Cansian ABM, Pucci EFQ, Guimarães JR, Tardioli PW, Saville BA. A review on the production and recovery of sugars from lignocellulosics for use in the synthesis of bioproducts. Ind Crops Prod. 2022;186:115213. doi: 10.1016/j.indcrop.2022.115213. [DOI] [Google Scholar]
- Harris PJ, Blakeney AB, Henry RJ, Stone BA. Gas-chromatographic determination of the monosaccharide composition of plant-cell wall preparations. J Assoc off Anal Chem. 1988;71(2):272–275. [Google Scholar]
- Kaur D, Singla G, Singh U, Krishania M. Efficient process engineering for extraction of hemicellulose from corn fiber and its characterization. Carbohydr Polym Technol Appl. 2020;1:100011. doi: 10.1016/j.carpta.2020.100011. [DOI] [Google Scholar]
- Kormelink FJM, Voragen AGJ. Degradation of different [(glucurono) arabino] xylans by a combination of purified xylan-degrading enzymes. Appl Microbiol Biotechnol. 1993;38(5):688–695. doi: 10.1007/BF00182811. [DOI] [Google Scholar]
- Liu J, Sun D, Zhu J, Liu C, Liu W. Carbohydrate-binding modules targeting branched polysaccharides: overcoming side-chain recalcitrance in a non-catalytic approach. Bioresour Bioprocess. 2021 doi: 10.1186/s40643-021-00381-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Long L, Sun L, Liu Z, Lin Q, Wang J, Ding S. Functional characterization of a GH62 family α-L-arabinofuranosidase from Eupenicillium parvum suitable for monosaccharification of corncob arabinoxylan in combination with key enzymes. Enzym Microb Technol. 2022;154:109965. doi: 10.1016/j.enzmictec.2021.109965. [DOI] [PubMed] [Google Scholar]
- Lynch KM, Steffen EJ, Arendt EK. Brewers’ spent grain: a review with an emphasis on food and health. J Inst Brew. 2016;122(4):553–568. doi: 10.1002/jib.363. [DOI] [Google Scholar]
- Mandalari G, Faulds CB, Sancho AI, Saija A, Bisignano G, Locurto R, Waldron KW. Fractionation and characterisation of arabinoxylans from brewers’ spent grain and wheat bran. J Cereal Sci. 2005;42(2):205–212. doi: 10.1016/j.jcs.2005.03.001. [DOI] [Google Scholar]
- Mccleary BV, Mckie VA, Draga A, Rooney E, Mangan D, Larkin J. Hydrolysis of wheat flour arabinoxylan, acid-debranched wheat flour arabinoxylan and arabino-xylo-oligosaccharides by b-xylanase, a-L-arabinofuranosidase and b-xylosidase. Carbohyd Res. 2015;407:79–96. doi: 10.1016/j.carres.2015.01.017. [DOI] [PubMed] [Google Scholar]
- Meyer AS, Rosgaard L, Sørensen HR. The minimal enzyme cocktail concept for biomass processing. J Cereal Sci. 2009;50(3):337–344. doi: 10.1016/j.jcs.2009.01.010. [DOI] [Google Scholar]
- Moreirinha C, Vilela C, Silva NHCS, Pinto RJB, Almeida A, Rocha MAM, Coelho E, Coimbra MA, Silvestre AJD, Freire CSR. Antioxidant and antimicrobial films based on brewers spent grain arabinoxylans, nanocellulose and feruloylated compounds for active packaging. Food Hydrocoll. 2020 doi: 10.1016/j.foodhyd.2020.105836. [DOI] [Google Scholar]
- Naidu DS, Hlangothi SP, John MJ. Bio-based products from xylan: a review. Carbohydr Polym. 2018;179:28–41. doi: 10.1016/j.carbpol.2017.09.064. [DOI] [PubMed] [Google Scholar]
- Pérez-Flores JG, Contreras-López E, Castañeda-Ovando A, Pérez-Moreno F, Aguilar-Arteaga K, Álvarez-Romero GA, Téllez-Jurado A. Physicochemical characterization of an arabinoxylan-rich fraction from brewers’ spent grain and its application as a release matrix for caffeine. Food Res Int. 2019;116:1020–1030. doi: 10.1016/j.foodres.2018.09.041. [DOI] [PubMed] [Google Scholar]
- Poletto P, Pereira GN, Monteiro CRM, Pereira MAF, Bordignon SE, de Oliveira D. Xylooligosaccharides: transforming the lignocellulosic biomasses into valuable 5-carbon sugar prebiotics. Process Biochem. 2020;91:352–363. doi: 10.1016/j.procbio.2020.01.005. [DOI] [Google Scholar]
- Rasmussen LE, Xu C, Sørensen JF, Nielsen MK, Meyer AS. Enzyme kinetics and identification of the rate-limiting step of enzymatic arabinoxylan degradation. Biochem Eng J. 2012;69:8–16. doi: 10.1016/j.bej.2012.08.004. [DOI] [Google Scholar]
- Reis SF, Coelho E, Coimbra MA, Abu-Ghannam N. Improved efficiency of brewer’s spent grain arabinoxylans by ultrasound-assisted extraction. Ultrason Sonochem. 2015;24:155–164. doi: 10.1016/j.ultsonch.2014.10.010. [DOI] [PubMed] [Google Scholar]
- Rojas-Pérez LC, Narváez-Rincón PC, Ballesteros I. Improving sugar extraction from brewers’ spent grain using sequential deproteinization and acid-catalyzed steam explosion in a biorefinery context. Biomass Bioenergy. 2022 doi: 10.1016/j.biombioe.2022.106389. [DOI] [Google Scholar]
- Saha BC. Hemicellulose bioconversion. J Ind Microbiol Biotechnol. 2003;30(5):279–291. doi: 10.1007/s10295-003-0049-x. [DOI] [PubMed] [Google Scholar]
- Sajib M, Falck P, Sardari RRR, Mathew S, Grey C, Karlsson EN, Adlercreutz P. Valorization of Brewer’s spent grain to prebiotic oligosaccharide: production, xylanase catalyzed hydrolysis, in-vitro evaluation with probiotic strains and in a batch human fecal fermentation model. J Biotechnol. 2018;268:61–70. doi: 10.1016/j.jbiotec.2018.01.005. [DOI] [PubMed] [Google Scholar]
- Selvendran RR, March JF, Ring SG. Determination of aldoses and uronic acid content of vegetable fiber. Anal Biochem. 1979;96(2):282–292. doi: 10.1016/0003-2697(79)90583-9. [DOI] [PubMed] [Google Scholar]
- Sørensen HR, Pedersen S, Viksø-Nielsen A, Meyer AS. Efficiencies of designed enzyme combinations in releasing arabinose and xylose from wheat arabinoxylan in an industrial ethanol fermentation residue. Enzyme Microb Technol. 2005;36(5–6):773–784. doi: 10.1016/j.enzmictec.2005.01.007. [DOI] [Google Scholar]
- Sørensen HR, Pedersen S, Jørgensen CT, Meyer AS. Enzymatic hydrolysis of wheat arabinoxylan by a recombinant "minimal” enzyme cocktail containing -xylosidase and novel endo -1, 4-b-xylanase and a-L-arabinofuranosidase activities. Biotechnol Prog. 2007;23:100–107. doi: 10.1021/bp0601701. [DOI] [PubMed] [Google Scholar]
- Sørensen HR, Pedersen S, Meyer AS. Synergistic enzyme mechanisms and effects of sequential enzyme additions on degradation of water insoluble wheat arabinoxylan. Enzyme Microb Technol. 2007;40(4):908–918. doi: 10.1016/j.enzmictec.2006.07.026. [DOI] [Google Scholar]
- Swart LJ, Bedzo OKK, van Rensburg E, Görgens JF. Pilot-scale xylooligosaccharide production through steam explosion of screw press–dried brewers’ spent grains. Biomass Convers Biorefinery. 2020 doi: 10.1007/s13399-020-01099-w. [DOI] [Google Scholar]
- Vieira E, Rocha MAM, Coelho E, Pinho O, Saraiva JA, Ferreira IMPLVO, Coimbra MA. Valuation of brewer’s spent grain using a fully recyclable integrated process for extraction of proteins and arabinoxylans. Ind Crops Prod. 2014;52:136–143. doi: 10.1016/j.indcrop.2013.10.012. [DOI] [Google Scholar]
- Xiros C, Topakas E, Katapodis P, Christakopoulos P. Hydrolysis and fermentation of brewer’s spent grain by Neurospora crassa. Biores Technol. 2008;99(13):5427–5435. doi: 10.1016/j.biortech.2007.11.010. [DOI] [PubMed] [Google Scholar]
- Xiros C, Katapodis P, Christakopoulos P. Factors affecting cellulose and hemicellulose hydrolysis of alkali treated brewers spent grain by Fusarium oxysporum enzyme extract. Biores Technol. 2011;102(2):1688–1696. doi: 10.1016/j.biortech.2010.09.108. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.