

Abstract
Secreted frizzled-related proteins (SFRPs) are involved in the development of various types of cancer and function by suppressing the Wnt signaling pathway. To elucidate the clinical implications of SFRPs in uterine sarcoma, SFRP expression levels and their effects on uterine leiomyosarcoma cells were examined. Immunostaining for SFRP4 was performed on uterine smooth muscle, uterine fibroid and uterine leiomyosarcoma tissues. Additionally, the effects of SFRP4 administration on cell viability, migration and adhesion were evaluated in uterine leiomyosarcoma SKN cells using the WST-1 assay (Roche Diagnostics) and the CytoSelect™ 24-well Cell Migration Assay Kit and the CytoSelect™ 48-well Cell Adhesion Assay Kit. The expression levels of SFRP4 in uterine leiomyosarcoma tissues were lower than those in normal smooth muscle and uterine fibroid tissues. In addition, SFRP4 suppressed the viability and migration, and increased the adhesion ability of uterine leiomyosarcoma cells compared with in the control group. In conclusion, SFRP4 may suppress the viability and migration, and enhance the adhesion of sarcoma cells. These results suggested that SFRP4 could be considered as a novel therapeutic target for uterine sarcoma.
Keywords: secreted frizzled-related protein, uterine sarcoma, Wnt signaling pathway, migration, adhesion
Introduction
Surgery is the standard treatment for uterine sarcoma, as chemotherapy has a limited effect; therefore, novel treatment strategies are required. Given the low frequency of sarcoma in Japan compared with other malignant tumors, there is a delay in basic and clinical research, despite ongoing efforts to analyze various genetic abnormalities as potential factors involved in the development of sarcoma (1).
Our previous study revealed that resveratrol exhibits a concentration-dependent inhibitory effect on the proliferation of uterine sarcoma cells, suggesting its potential as a novel therapeutic agent for uterine sarcoma. Furthermore, our previous study demonstrated that the mechanism of resveratrol action was mediated by the Wnt signaling pathway (2). Additionally, resveratrol has been shown to exert anticancer effects on various types of carcinoma such as colon cancer and breast cancer, and it has been reported to be involved in the Wnt signaling pathway (3-5).
The present study focused on secreted frizzled-related proteins (SFRPs), which are proteins that affect the Wnt signaling pathway and are implicated in the development of various types of cancer (6,7). The inactivation of SFRPs could hypothesized to enhance the Wnt signaling pathway (6), promoting cancer growth; therefore, Wnt and SFRPs might be novel molecular targets for cancer treatment.
Benign uterine leiomyomas do not typically progress to uterine sarcoma, rendering treatment unnecessary as they tend to shrink after menopause (8). Nevertheless, considering cases where leiomyoma may have progressed to sarcoma, it is important to evaluate the progression of uterine fibroids to sarcoma. By elucidating the impact of SFRPs on sarcoma and the dynamics of the Wnt signaling pathway, SFRPs could emerge as a new therapeutic strategy for sarcoma.
Although there are few reports on the role of the Wnt signaling pathway in the proliferation and metastasis of uterine sarcoma cells (9,10), there are no reports, to the best of our knowledge, on treatment strategies for uterine sarcoma focusing on SFRPs. Therefore, the present study examined the differences in SFRP4 expression between normal uterine smooth muscle cells and uterine leiomyosarcoma. In addition, the inhibitory effect of SFRP4 on uterine sarcoma cells was assessed, as well as its effect on cell migration and adhesion.
Materials and methods
Cell line and culture
The SKN human uterine leiomyosarcoma cell line (European Collection of Authenticated Cell Cultures) was cultured in Dulbecco's Modified Eagle Medium/Nutrient Mixture F-12 (Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS, Thermo Fisher Scientific, Inc.), penicillin (100 U/ml) and streptomycin (100 µg/ml) in an incubator containing 5% CO2 at 37˚C.
Immunohistochemistry
SFRP4 was purchased from PeproTech, Inc. (cat. no. 120-50). SFRP4 cells were homogenized and filter-sterilized through a 0.2 µm filter, then frozen at -20˚C. Immunostaining of SFRP4 was performed on four cases of normal uterine smooth muscle, uterine fibroids and uterine leiomyosarcoma. The tissues were sectioned at a thickness of 5 µm. To block endogenous peroxidase and phosphatase activity, tissues were incubated with 3% hydrogen peroxide for 10 min at room temperature. Blocking was performed using 5% goat serum (Chemicon International; Thermo Fisher Scientific, Inc.) for 30 min at room temperature. Rabbit monoclonal antibodies against SFRP4 (dilution, 1:900; cat. no. 154167; Abcam) were added to tissues and incubated for 1 h at 37˚C. The tissues were incubated with the HRP-conjugated secondary antibody (1:1,000, cat. no. ab6721, Abcam) for 1 h at 37˚C. For chromogen detection, the Pierce DAB Substrate kit (cat. no. 34002; Thermo Fisher Scientific, Inc.) was used. As a positive control, immunohistochemical staining of SFRP4 in human colonic mucosa using SFRP4 polyclonal antibodies (1:400, cat. no. 15328-1-AP, Proteintech Group, Inc.) was performed. In the positive control tissues, the antigen-antibody reaction could be sufficiently visualized using the primary antibodies. Tissues were incubated for 1.5 h at 37˚C.
The intensity score (0-3) and proportion score (0-3) were used as evaluation methods (Table I). The human tissue samples used in the present study were obtained from patients who underwent surgery for uterine fibroids and uterine sarcoma at the Obstetrics and Gynecology Department of Tokushima University Hospital (Tokushima, Japan) between January 2000 and December 2020. There were a total of 19 patients, ranging in age from 46-69 years, with the mean age of 57 years. Normal uterine smooth muscle tissue was collected from 8 patients, aged 46-57 years, with a mean age of 52 years, who underwent surgery for uterine fibroids. The colon mucosal tissue used as a positive control was a portion of the sample from a patient who underwent surgery at Tokushima University Hospital that was scheduled to be discarded, but was kept in a way that could not identify the individual. There were no exclusion criteria for the present study. For using sample tissues, an opt-out method was used, where an information disclosure document was posted on the Tokushima University Hospital web page, thereby omitting the need to obtain consent from individual research subjects. Discontinuation/withdrawal from the study occurred when a research subject (or their representative consenter) expressed a refusal to continue participation, or when the research director or researcher deemed it appropriate to discontinue the research. The present study was approved by the Ethics Committee of Tokushima University Hospital (Tokushima, Japan; approval no. 4159).
Table I.
Intensity and proportion scores for normal uterine smooth muscle tissue, uterine fibroid tissue and uterine leiomyosarcoma tissue.
| Score | 0 | 1 | 2 | 3 |
|---|---|---|---|---|
| Intensity | Not stained at all | Slightly stained with strong to medium enlargement | Stained with low to medium enlargement | Stained well with weak enlargement |
| Proportion | <1% | 1-10% | 10-50% | >50% |
The intensity score (0-3) and proportion score (0-3) were used as evaluation methods. Intensity refers to degree of staining; depending on the degree of staining, stained cells are classified into one of four classes: Negative, weakly stained, moderately stained and strongly stained. Proportion refers to the percentage of stained cells. Depending on the percentage of stained cells, one of four classes (<1%, 1-10%, 10-50%, >50%) is assigned. A proportion value of 10% was scored as 2.
In order to confirm the expression of SFRP4 by immunostaining, in four cases of normal uterine smooth muscle and three cases of uterine leiomyosarcoma, the number of stained cells was measured using a BZ-800 Analyzer (fluorescence, Keyence Corporation), and the ratio of the area was measured (magnification, x20).
Cell treatment and viability assay
SKN uterine leiomyosarcoma cells were seeded on a 96-well microplate at a density of 5x102 cells/well and were incubated at 37˚C for 24 h prior to treatment. SFRP4 was added to cells at concentrations of 1.25 and 2.5 µg/ml based on the results of a previous study (11), and, after 48 h at 37˚C, cell viability was assessed at an absorbance of 450 nm using the WST-1 assay (Roche Diagnostics) according to the manufacturer's instructions.
Cell migration and adhesion assays
To assess changes in the migration of uterine sarcoma cells following SFRP4 administration, the CytoSelect™ 24-well Cell Migration Assay Kit (cat. CBA-100; Cell Biolabs, Inc.) was used. The CytoSelect Cell Migration Assay Kit contains polycarbonate membrane inserts (pore size, 8 µm) in a 24-well plate. The membrane serves as a barrier for differentiating migratory cells from non-migratory cells. Migratory cells can extend their protrusions towards chemoattractants, such as FBS (via actin cytoskeleton reorganization), and ultimately pass through the pores of the polycarbonate membrane (12). The migrating cells that passed through the membrane were collected and used for staining and semi-quantification.
Briefly, uterine leiomyosarcoma cells were seeded at a density of 5x102 cells/well in 100 µl culture in the CytoSelect 24-well Cell Migration Assay plate and divided into a group treated with SFRP4 at a concentration of 2.5 µg/ml at 37˚C for 24 h and a control group that received no treatment. WST-1 reagent (10%) was added to each well and the cells were incubated at 37˚C for 1 h. The samples were washed and the absorbance was measured at 450 nm using a microplate reader (Thermo Scientific Varioskan Flash Multimode reader; Thermo Fisher Scientific, Inc.).
Similarly, to assess changes in cell adhesion, uterine leiomyosarcoma cells were seeded at a density of 5x102 cells/well in a CytoSelect 48-well Cell Adhesion Assay plate (cat. no. CBA-070; Cell Biolabs, Inc.) and divided into a group treated with SFRP4 at a concentration of 2.5 µg/ml at 37˚C for 24 h and a control group that received no treatment. After adherence for 24 h to fibronectin, collagen I, collagen IV, laminin I, fibrinogen and BSA, the samples were washed twice with cold PBS and the absorbance was read at 450 nm using a microplate reader. BSA was used as a control and was included in the aforementioned kit.
Statistical analysis
Statistical analyses were performed on triplicate repeats using the Ekuseru-Toukei 2012 software (version 1.16; Social Survey Research Information Co., Ltd.). Data were expressed as the mean ± standard error of the mean. The statistical significance of the differences between the experimental and control groups was assessed using one-way ANOVA followed by Dunnett's test. Comparisons between two groups were evaluated using the unpaired Student's t-test. Immunohistochemistry scores were statistically analyzed using Kruskal-Wallis test followed by Dunn's test. P<0.05 was considered to indicate a statistically significant difference.
Results
The median intensities of SFRP4 expression in uterine smooth muscle, fibroids and leiomyosarcomas were 2.5, 3.0 and 1.0, respectively, and the median proportions were 3.0, 3.0 and 1.5, respectively. The expression levels of SFRP4 were significantly lower in uterine leiomyosarcoma tissues than those in normal smooth muscle and uterine fibroid tissues (P=0.01; Table II; Fig. 1).
Table II.
Expression of SFRP4 in different tissues.
| A, Expression of SFRP4 in normal uterine smooth muscle tissue | ||
|---|---|---|
| Tissue | Intensity | Proportion |
| Normal uterine smooth muscle 1 | 3 | 3 |
| Normal uterine smooth muscle 2 | 1 | 2 |
| Normal uterine smooth muscle 3 | 3 | 3 |
| Normal uterine smooth muscle 4 | 2 | 3 |
| B, Expression of SFRP4 in uterine fibroid tissue | ||
| Tissue | Intensity | Proportion |
| Uterine fibroid | 3 | 3 |
| Uterine fibroid 2 | 3 | 3 |
| Uterine fibroid 3 | 2 | 3 |
| Uterine fibroid 4 | 3 | 3 |
| C, Expression of SFRP4 in uterine leiomyosarcoma tissue | ||
| Tissue | Intensity | Proportion |
| Uterine leiomyosarcoma 1 | 1 | 1 |
| Uterine leiomyosarcoma 2 | 1 | 2 |
| Uterine leiomyosarcoma 3 | 1 | 1 |
| Uterine leiomyosarcoma 4 | 2 | 3 |
The expression of SFRP4 was assessed using immunostaining and evaluated based on the intensity and proportion scores. The median intensities and proportions of SFRP4 expression in uterine smooth muscle, fibroids and leiomyosarcomas were compared. SFRP4 expression in uterine leiomyosarcoma was significantly lower than that in normal uterine smooth muscle and uterine fibroids (n=4). P=0.01 vs. control. SFRP, secreted frizzled-related protein.
Figure 1.

Expression of secreted frizzled-related protein 4 in normal uterine smooth muscle, uterine fibroid and uterine leiomyosarcoma tissues was assessed through immunostaining and evaluated based on the intensity score and the proportion score (n=4). As a positive control, SFRP4 immunostaining was performed on human colonic mucosa previously collected at Tokushima University Hospital (Tokushima, Japan) Magnification, x20.
The number of stained cells was measured and the area ratio was calculated in normal uterine smooth muscle and uterine leiomyosarcoma. Normal smooth muscle showed an area ratio of 62.0%, whereas it was 23.7% in leiomyosarcoma, which was significantly lower than that in normal smooth muscle (P=0.03; Table III; Fig. 2).
Table III.
SFRP4 stained cell area ratio in normal uterine smooth muscle and uterine leiomyosarcoma tissues.
| Tissue | Total area, µm2 | Stained area, µm2 | Ratio, % |
|---|---|---|---|
| Normal uterine smooth muscle 1 | 579,907 | 329,900 | 57 |
| Normal uterine smooth muscle 2 | 288,917 | 235,857 | 82 |
| Normal uterine smooth muscle 3 | 1,062,726 | 569,027 | 54 |
| Normal uterine smooth muscle 4 | 634,039 | 350,073 | 55 |
| Uterine leiomyosarcoma 1 | 2,675,916 | 1,282,439 | 48 |
| Uterine leiomyosarcoma 2 | 2,328,849 | 2,273,09 | 10 |
| Uterine leiomyosarcoma 3 | 2,156,167 | 290,801 | 13 |
To confirm the expression of SFRP4 by immunostaining, the number of stained cells was measured in 4 cases of normal uterine smooth muscle and 3 cases of uterine leiomyosarcoma, and the area ratio was calculated. SFRP, secreted frizzled-related protein.
Figure 2.
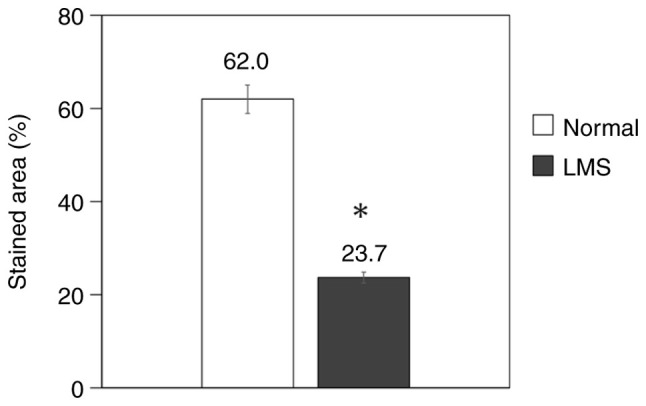
Ratio of SFRP4 expression. In four cases of normal uterine smooth muscle and three cases of uterine LMS, the number of cells stained was measured, and the ratio of the area was measured. SFRP4 expression in leiomyosarcoma was significantly lower than that in normal smooth muscle. *P=0.03 vs. normal uterine smooth muscle. SFRP, secreted frizzled-related protein; LMS, leiomyosarcoma.
SFRP4 was added to SKN uterine leiomyosarcoma cells at a concentration of 1.25 and 2.5 µg/ml and the absorbance was measured to examine the cell viability. The control group showed an absorbance value of 5.45, the SFRP4 1.25 µg/ml group showed an absorbance value of 5.28, whereas the SFRP4 2.5 µg/ml group showed an absorbance value of 4.95. Cell viability was significantly suppressed in response to a concentration of 2.5 µg/ml SFRP4 compared with that in the control group (P=0.001; Fig. 3).
Figure 3.
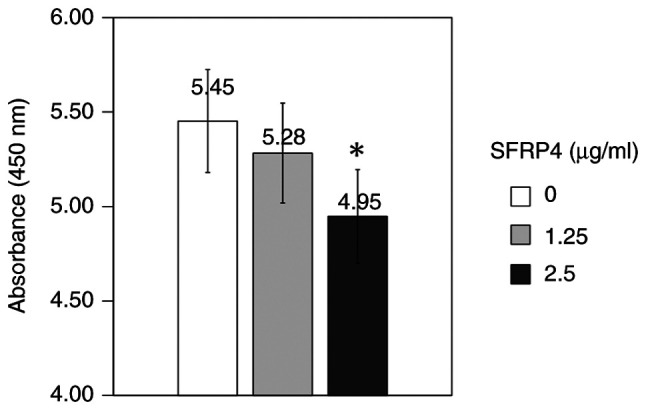
SFRP4 was used to treat uterine leiomyosarcoma SKN cells at concentrations of 1.25 and 2.5 µg/ml, and the absorbance was measured. Cell viability was significantly suppressed in response to SFRP4 at a concentration of 2.5 µg/ml (n=8). *P=0.001 vs. the control. SFRP, secreted frizzled-related protein.
SKN uterine leiomyosarcoma cells were cultured in the CytoSelect 24-well Cell Migration Assay plates, SFRP4 was added at a concentration of 2.5 µg/ml, and the absorbance was measured. The control group showed an absorbance value of 0.75, whereas the absorbance value was 0.55 in the SFRP4 group, showing a significant decrease in cell migration (P=0.007; Fig. 4).
Figure 4.
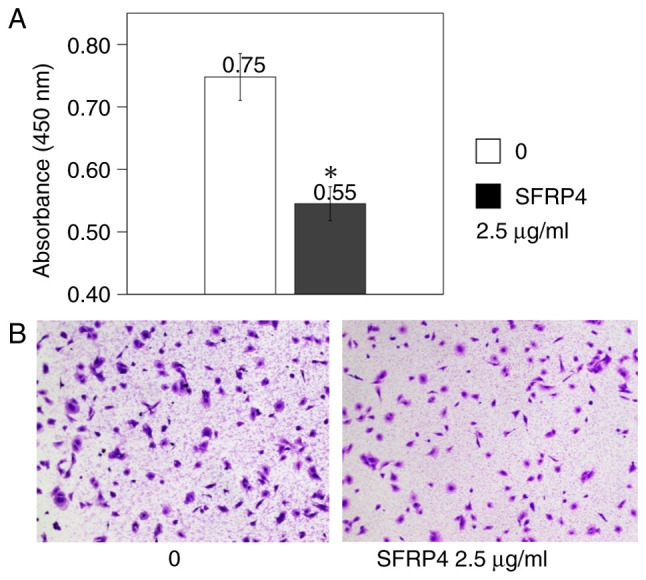
Uterine leiomyosarcoma SKN cells were cultured in the CytoSelect 24-well Cell Migration Assay plates, SFRP4 was added at a concentration of 2.5 µg/ml, and the absorbance was measured. The ‘0’ group was the SFRP4 untreated control group. (A) Leiomyosarcoma cells showed a significant decrease in migration (n=4). (B) A histological image comparing SFRP4-untreated cells and SFRP-treated (2.5 µg/ml) cells. A decrease in cell number was observed in the treated group compared with the untreated group. Magnification, x40. *P=0.007 vs. control. SFRP, secreted frizzled-related protein.
SKN uterine leiomyosarcoma cells were cultured in the CytoSelect 48-well Cell Adhesion Assay plates, SFRP4 was added at a concentration of 2.5 µg/ml, and the absorbance was measured. For each cell adhesion molecule (fibronectin, collagen-1, collagen-4, laminin fibrinogen and BSA), the values were 0.41, 0.30, 0.33, 0.32 and 0.40 in the control group and 1.16, 0.78, 0.61, 0.71 and 0.70 in the SFRP4 group, respectively. The adhesion ability of leiomyosarcoma cells to each cell adhesion molecule was significantly increased in response to SFRP4 compared with that in the control cells (P=0.0004, 0.002, 0.002, 0.001 and 0.007, respectively; Fig. 5).
Figure 5.

Uterine leiomyosarcoma SKN cells were cultured in CytoSelect 48-well Cell Adhesion Assay plates, SFRP4 was added at a concentration of 2.5 µg/ml, and the absorbance was measured. The ‘0’ group was the SFRP4 untreated control group. (A) Adhesion ability of the leiomyosarcoma cells was significantly increased to each cell adhesion molecule (n=4). (B) A histological image comparing SFRP4-untreated cells and SFRP-treated (2.5 µg/ml) cells. An increase in cell number was observed in the treated group compared with the untreated group. Magnification, x40. *P<0.01 vs. control. Col, collagen; Lam, laminin I; SFRP, secreted frizzled-related protein.
Discussion
Our previous study has shown that resveratrol suppresses the proliferation of uterine sarcoma cells via the Wnt signaling pathway (1). Therefore, the present study focused on SFRPs, which have been indicated to be involved in the Wnt signaling pathway, along with resveratrol. Wnt is an extracellular protein with a molecular weight of ~40,000 g/mol. When Wnt binds to Wnt receptor Frizzled in the cytoplasm, three types of intracellular signal transduction pathways are activated, namely the β-catenin pathway, the planar cell polarity pathway and the Ca2+ pathway (13).
The β-catenin pathway regulates cell proliferation and differentiation through gene expression by regulating the intracellular levels of β-catenin. Numerous genetic abnormalities within constituents of the β-catenin pathway have been identified in various types of human carcinoma, leading to abnormal accumulation of β-catenin in the cytoplasm and nucleus (13). Furthermore, abnormal accumulation of β-catenin is known to cause overexpression of cancer-related genes, such as c-myc, ultimately causing abnormal cell proliferation (13).
Fang et al (14) revealed that the Wnt signaling pathway is activated in osteosarcoma cells. Zou et al (3) reported that resveratrol suppresses the Wnt signaling pathway in osteosarcoma.
All members of the SFRP gene family are small proteins, ~30 kDa in size, belonging to five gene families, SFRP1-5, which are widely conserved across species. Comparison of amino acid sequence homology among families suggests that SFRP1, 2 and 5 and SFRP3 and 4 form separate subfamilies, and there are differences in their expression levels in each organ. SFRPs suppress the Wnt signaling pathway by directly inhibiting the binding of Wnt to its receptor, Frizzled, via extracellular secretion. The enhancement of Wnt signaling by inactivating SFRP may serve an important role in activation of the Wnt signaling pathway in cancer (15). Thus, Wnt and SFRP are considered novel molecular targets for cancer treatment.
Several studies have reported on the relationship between SFRP levels and several types of cancer. SFRP4 inhibits the Wnt signaling pathway in ovarian cancer (11). Furthermore, multiple SFRP genes are frequently inactivated in colorectal cancer (16). Decreased SFRP1 expression, and induction of apoptosis due to its overexpression, have been confirmed in cervical cancer (17). In addition, SFRP1 and SFRP2 suppress cell proliferation via the Wnt signaling pathway in cervical cancer (18). SFRP5 has been shown to inhibit cervical tumorigenesis by interfering with the Wnt pathway in vitro (19). In addition, the roles of the Wnt signaling pathway and SFRP4 in ovarian cancer have been reported (20).
To the best of our knowledge, there have been no reports on treatment strategies for sarcoma focusing on SFRPs. By elucidating the expression of SFRPs in uterine sarcoma and their effects on inhibitory action against sarcoma cell proliferation, migration and adhesion of sarcoma cells, SFRPs may be considered as a new therapeutic agent for uterine sarcoma.
In the present study, immunostaining revealed that uterine leiomyosarcoma cells expressed significantly less SFRP4 than normal uterine smooth muscle cells and uterine fibroids. The present study hypothesized that enhancement of Wnt signaling by the inactivation of SFRP may be involved in cancer growth, thus suggesting that SFRP inactivation may be involved in the growth of uterine sarcoma. In addition, cell viability was suppressed by the treatment of uterine sarcoma cells with SFRP4. Previous studies have reported that SFRP suppresses the Wnt signaling pathway, and it is possible that SFRP suppresses cell proliferation via this pathway.
In previous years, cell adhesion has been elucidated at the molecular level, and it has become clear that various cell adhesion molecules are involved in tumor metastasis through complex processes. Cell migration is a multistep process that serves an important role not only in tumor infiltration and metastasis, but also in wound healing, cell differentiation and embryonic development. Cell infiltration involves cell migration, wherein the cells migrate beyond the extracellular matrix and settle at new sites (21). Notably, attempts have been made to suppress tumor metastasis by controlling cell adhesion (22,23). In the present study, the treatment of uterine sarcoma cells with SFRP4 decreased their migratory ability and increased their adhesive ability; thus suggesting that SFRP4 may be involved in the infiltration and metastasis of uterine sarcoma cells.
In conclusion, the present study demonstrated that uterine leiomyosarcoma expresses less SFRP4 than normal uterine smooth muscle tissues, and that SFRP4 suppresses the proliferation of uterine leiomyosarcoma cells, reduces their migration and increases their adhesion.
These results should be interpreted with caution because of the small sample size of human samples used in the present study, and it is important to consider a number of limitations. A small sample size may skew statistical tests when identifying relationships and connections within the resulting data set. The role of SFRP4 could be better understood by increasing the number of samples. In addition, since SFRP4 was shown to inhibit the proliferation of SKN leiomyosarcoma cells, complementary experiments investigating apoptosis would provide evidence to support the results of the present study. Although we did not examine the Wnt signaling pathway in detail in this study, further analysis of the effects of SFRP on sarcoma, and changes in the factors involved in the Wnt signaling pathway may verify the use of SFRPs as a novel treatment option for uterine sarcoma.
Acknowledgements
For the present study, the laboratory and experimental equipment at the Support Center for Advanced Medical Sciences at the Tokushima University Graduate School of Biomedical Sciences (Tokushima, Japan) were used to perform cell culture and the absorbance measurements.
Funding Statement
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
ToK and AM analyzed and interpreted data regarding the effect of SFRP on uterine sarcoma. ToK and AM confirm the authenticity of all the raw data. ToK, AM, TN, AS, RA, HI, HN, AY, RK, YY, KY, TaK, MN and TI made substantial contributions to conception and design, or acquisition of data or analysis and interpretation of data. MN, KY and TI wrote and revised the manuscript. All authors have read and approved the final version of the manuscript.
Ethics approval and consent to participate
When using sample tissues in the present study, an opt-out method was used, where an information disclosure document was posted on the Tokushima University Hospital web page, thereby omitting the need to obtain consent from individual research subjects. The present study was approved by the Ethics Committee of Tokushima University Hospital (approval no. 4159).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Nakata E, Fujiwara T, Kunisada T, Ito T, Takihira S, Ozaki T. Immunotherapy for sarcomas. Jpn J Clin Oncol. 2021;51:523–537. doi: 10.1093/jjco/hyab005. [DOI] [PubMed] [Google Scholar]
- 2.Mineda A, Nishimura M, Kagawa T, Takiguchi E, Kawakita T, Abe A, Irahara M. Resveratrol suppresses proliferation and induces apotosis of uterine sarcoma cells by inhibiting the Wnt signaling pathway. Exp Ther Med. 2019;17:2242–2246. doi: 10.3892/etm.2019.7209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zou Y, Yang J, Jiang D. Resveratrol inhibits canonical Wnt signaling in human MG-63 osteosarcoma cells. Mol Med Rep. 2015;12:7221–7226. doi: 10.3892/mmr.2015.4338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Jang M, Cai L, Udeani GO, Slowing KV, Thomas CF, Beecher CW, Fong HH, Farnsworth NR, Kinghorn AD, Mehta RG, et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science. 1997;275:218–220. doi: 10.1126/science.275.5297.218. [DOI] [PubMed] [Google Scholar]
- 5.Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004;20:781–810. doi: 10.1146/annurev.cellbio.20.010403.113126. [DOI] [PubMed] [Google Scholar]
- 6.Lavergne E, Hendaoui I, Coulouarn C, Ribault C, Leseur J, Eliat PA, Mebarki S, Corlu A, Clément B, Musso O. Blocking Wnt signaling by SFRP-like molecules inhibits in vivo cell proliferation and tumor growth in cells carrying active β-catenin. Oncogene. 2011;30:423–433. doi: 10.1038/onc.2010.432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wu Q, Xu C, Zeng X, Zhang Z, Yang B, Rao Z. Tumor suppressor role of sFRP-4 in hepatocellular carcinoma via the Wnt/β-catenin signaling pathway. Mol Med Rep. 2021;23(336) doi: 10.3892/mmr.2021.11975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.De La Cruz MS, Buchanan EM. Uterine fibroids: Diagnosis and treatment. Am Fam Physician. 2017;95:100–107. [PubMed] [Google Scholar]
- 9.Pridgeon MG, Grohar PJ, Steensma MR, Williams BO. Wnt signaling in ewing sarcoma, osteosarcoma, and malignant peripheral nerve sheath tumors. Curr Osteoporos Rep. 2017;15:239–246. doi: 10.1007/s11914-017-0377-9. [DOI] [PubMed] [Google Scholar]
- 10.Singla A, Wang J, Yang R, Geller DS, Loeb DM, Hoang BH. Wnt signaling in osteosarcoma. Adv Exp Med Biol. 2020;1258:125–139. doi: 10.1007/978-3-030-43085-6_8. [DOI] [PubMed] [Google Scholar]
- 11.Ford CE, Jary E, Ma SSQ, Nixdorf S, Heinzelmann-Schwarz VA, Ward RL. The Wnt gatekeeper SFRP4 modulates EMT, cell migration and downstream Wnt signalling in serous ovarian cancer cells. PLoS One. 2013;8(e54362) doi: 10.1371/journal.pone.0054362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, Parsons JT, Horwitz AR. Cell migration: Integrating signals from front to back. Science. 2003;302:1704–1709. doi: 10.1126/science.1092053. [DOI] [PubMed] [Google Scholar]
- 13.Kikuchi A, Yamamoto H, Kishida S. Multiple of the interactions of Wnt proteins and their receptors. Cell Signal. 2007;19:659–671. doi: 10.1016/j.cellsig.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 14.Fang F, VanCleave A, Helmuth R, Torres H, Rickel K, Wollenzien H, Sun H, Zeng E, Zhao J, Tao J. Targeting the Wnt/β-catenin pathway in human osteosarcoma cells. Oncotarget. 2018;9:36780–36792. doi: 10.18632/oncotarget.26377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rattner A, Hsieh JC, Smallwood PM, Gilbert DJ, Copeland NG, Jenkins NA, Nathans J. A family of secreted proteins contains homology to the cysteine-rich ligand-binding domain of frizzled receptors. Proc Natl Acad Sci USA. 1997;94:2859–2863. doi: 10.1073/pnas.94.7.2859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Suzuki H, Watkins DN, Jair KW, Schuebel KE, Markowitz SD, Chen WD, Pretlow TP, Yang B, Akiyama Y, Van Engeland M, et al. Epigenetic inactivation of SFRP genes allows constitutive WNT signaling in colorectal cancer. Nat Genet. 2004;36:417–422. doi: 10.1038/ng1330. [DOI] [PubMed] [Google Scholar]
- 17.Ko J, Ryu KS, Lee YH, Na DS, Kim YS, Oh YM, Kim IS, Kim JW. Human secreted frizzled-related protein is down-regulated and induces apoptosis in human cervical cancer. Exp Cell Res. 2002;280:280–287. doi: 10.1006/excr.2002.5649. [DOI] [PubMed] [Google Scholar]
- 18.Chung MT, Lai HC, Sytwu HK, Yan MD, Shih YL, Chang CC, Yu MH, Liu HS, Chu DW, Lin YW. SFRP1 and SFRP2 suppress the transformation and invasion abilities of cervical cancer cells through Wnt signal pathway. Gynecol Oncol. 2009;112:646–653. doi: 10.1016/j.ygyno.2008.10.026. [DOI] [PubMed] [Google Scholar]
- 19.Lin YW, Chung MT, Lai HC, Yan MD, Shih YL, Chang CC, Yu MH. Methylation analysis of SFRP genes family in cervical adenocarcinoma. J Cancer Res Clin Oncol. 2009;135:1665–1674. doi: 10.1007/s00432-009-0613-5. [DOI] [PubMed] [Google Scholar]
- 20.Jacob F, Ukegjini K, Nixdorf S, Ford CE, Olivier J, Caduff R, Scurry JP, Guertler R, Hornung D, Mueller R, et al. Loss of secreted frizzled-related protein 4 correlates with an aggressive phenotype and predicts poor outcome in ovarian cancer patients. PLoS One. 2012;7(e31885) doi: 10.1371/journal.pone.0031885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kawauchi T. Cell adhesion and its endocytic regulation in cell migration during neural development and cancer metastasis. Int J Mol Sci. 2012;13:4564–4590. doi: 10.3390/ijms13044564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mei J, Jin LP, Ding D, Li MQ, Li DJ, Zhu XY. Inhibition of IDO1 suppresses cyclooxygenase-2 and matrix metalloproteinase-9 expression and decreases proliferation, adhesion and invasion of endometrial stromal cells. Mol Hum Reprod. 2012;18:467–476. doi: 10.1093/molehr/gas021. [DOI] [PubMed] [Google Scholar]
- 23.Tsukamoto H, Kondo S, Mukudai Y, Nagumo T, Yasuda A, Kurihara Y, Kamatani T, Shintani S. Evaluation of anticancer activities of benzo[c]phenanthridine alkaloid sanguinarine in oral squamous cell carcinoma cell line. Anticancer Res. 2011;31:2841–2846. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.