

Significance
In humans, ~600 enzyme complexes can carry out protein ubiquitylation, and the most abundant class of these are the cullin 3-RING-ligase complexes (CRL3s). CRL3s are multiprotein complexes built around a BTB/cullin 3 core, and the incorporation of different BTB proteins into this scaffold results in distinct architectures that ubiquitylate a wide range of substrates. In most cases, it is not known how the complexes are tuned to their substrates. We show that the BTB protein KCTD5 is the central organizer in a CRL3KCTD5 complex and that the architecture and internal dynamics of KCTD5 are essential for positioning a Gβγ substrate protein near an activated ubiquitin for the transfer reaction. This explains how KCTD5 targets Gβγ for proteasomal degradation and regulates cellular activities.
Keywords: ubiquitin ligase, BTB proteins, G proteins, cryo electron microscopy
Abstract
Heterotrimeric G proteins can be regulated by posttranslational modifications, including ubiquitylation. KCTD5, a pentameric substrate receptor protein consisting of an N-terminal BTB domain and a C-terminal domain, engages CUL3 to form the central scaffold of a cullin-RING E3 ligase complex (CRL3KCTD5) that ubiquitylates Gβγ and reduces Gβγ protein levels in cells. The cryo-EM structure of a 5:5:5 KCTD5/CUL3NTD/Gβ1γ2 assembly reveals a highly dynamic complex with rotations of over 60° between the KCTD5BTB/CUL3NTD and KCTD5CTD/Gβγ moieties of the structure. CRL3KCTD5 engages the E3 ligase ARIH1 to ubiquitylate Gβγ in an E3-E3 superassembly, and extension of the structure to include full-length CUL3 with RBX1 and an ARIH1~ubiquitin conjugate reveals that some conformational states position the ARIH1~ubiquitin thioester bond to within 10 Å of lysine-23 of Gβ and likely represent priming complexes. Most previously described CRL/substrate structures have consisted of monovalent complexes and have involved flexible peptide substrates. The structure of the KCTD5/CUL3NTD/Gβγ complex shows that the oligomerization of a substrate receptor can generate a polyvalent E3 ligase complex and that the internal dynamics of the substrate receptor can position a structured target for ubiquitylation in a CRL3 complex.
Protein ubiquitylation plays a central role in eukaryotic biology and is often used to regulate intracellular protein levels through proteasomal degradation. The selection of targets for modification is largely determined by the E3 ubiquitin ligases, and the family of cullin-RING E3 ligases (CRLs) catalyze ubiquitin transfer by positioning lysine residues from substrates next to activated E2~ubiquitin or RING-between-RING (RBR) E3~ubiquitin conjugates (1, 2). These multicomponent modular assemblies use a cullin arm to bridge substrate-binding receptors (SRs) to the RBX1/2 RING component of the ubiquitylation machinery. The cullin 3-RING-ligase complex (CRL3) family, defined by the inclusion of cullin 3 (CUL3), can directly engage over 100 different SRs that typically combine a BTB CUL3-binding domain and a substrate-binding domain in a single polypeptide (3). Notably, BTB domains can self-associate into stable dimers, pentamers, and oligomers (4–9) and thus drive the multimerization of CRL3 complexes. While multivalency is not a unique feature of the CRL3s (10), most structures of CRL complexes reported to date have involved truncated components that result in monovalent complexes (11, 12).
Proteomic studies have identified KCTD5, KCTD2, and KCTD17, a family of homologous pentameric BTB CRL3 substrate receptors, as interactors of some Gβγ heterodimers (Gβ1γ2, Gβ1γ7, Gβ2γ7, and Gβ2γ2) (13–17), and functional studies have shown that these receptors can promote the ubiquitylation of G protein subunits via CRL3KCTD complexes (17–20). KCTD5 has roles in neurodevelopment and is linked to sleep disorders (21, 22), and members of the wider family of 25 human KCTD proteins have been associated with neurological disorders, obesity, and cancer through a variety of mechanisms (23–26). G proteins, consisting of Gαβγ heterotrimers and, once activated by GPCRs, the dissociated Gα monomer and Gβγ heterodimers, activate downstream cellular effectors. Gβγ subunits are involved in a wide range of signaling events in many different subcellular locations (27–29).
CRLs ubiquitylate a wide variety of substrates, ranging from intrinsically disordered proteins anchored via short degron motifs to ordered targets (for example, refs. 30–33). In the case of ordered substrates such as Gβγ, little is known about how specific lysine residues are selected for modification. Here, we show that a CRL3KCTD5 complex directly ubiquitylates Gβ1γ2 in a reaction that depends on the RBR E3 ligase ARIH1 (34) and that KCTD5 modulates Gβγ levels in cells in a proteosome-dependent manner. Cryo-EM structures of KCTD5 in complex with Gβ1γ2 and the N-terminal domain of CUL3 (CUL3NTD) show that KCTD5 forms a central pentameric scaffold that simultaneously engages five Gβγ substrates via its C-terminal domain (KCTD5CTD) and five CUL3NTD subunits via its N-terminal BTB domain (KCTD5BTB). Internal flexibility is a common feature of E3 ligases, and rotational and translational dynamics between the KCTD5CTD/Gβγ and KCTD5BTB/CUL3 moieties of the 5:5:5 complex generate multiple poses of Gβγ relative to CUL3. We used the experimental structure to model the full reaction complex and show that the internal dynamics of KCTD5 generate conformers in which lysine-23 of Gβ1 is positioned near an activated ARIH1~ubiquitin held by the neddylated CUL3/RBX1 domain and likely represent the priming complex.
Results
KCTD5 Assembles a KCTD5/CUL3NTD/Gβγ Complex.
We expressed and purified KCTD5, CUL3NTD, and Gβ1γ2 and used tagged KCTD5 to show that CUL3NTD and Gβγ bind individually and in combination with KCTD5 (Fig. 1A). Next, we used biolayer interferometry (BLI) with anchored KCTD5 for more quantitative analyses of the interactions. CUL3NTD binds with a 30 nM dissociation constant, consistent with previous results (6, 8, 35), while Gβγ binds with micromolar affinity (Fig. 1B and SI Appendix, Fig. S1 and Table S1). The presence of CUL3NTD had little effect on the binding affinity of Gβγ for KCTD5, suggesting little or no cooperativity between the binding of the two partners. Notably, while CUL3NTD and Gβγ had similar association rates, the dissociation rate for Gβγ was approximately 60-fold higher than for CUL3NTD and largely accounts for the differences in the affinities between the components. In size-exclusion chromatography (SEC) experiments, KCTD5 alone elutes as a single peak consistent with the expected homopentamer seen in crystal structures (36). Additions of CUL3NTD to KCTD5 resulted in the formation of a larger species as detected by SEC, with a maximum shift at a 1:1 molar ratio (SI Appendix, Fig. S2). In contrast, the addition of Gβγ did not produce a complex with KCTD5 that was detectable by SEC, even at a twofold molar excess. We attribute this result to the rapid exchange kinetics between KCTD5 and Gβγ on the time scale of the SEC experiments (~24 min from injection to elution). However, a recent publication showed that KCTD5 and Gβγ could form a stable complex by SEC (20). Overall, these experiments show that both CUL3 and Gβγ bind directly and independently to KCTD5, but that Gβγ interacts more weakly and has a shorter residence time, at least under the tested conditions.
Fig. 1.

KCTD5 is a CRL3 substrate receptor for Gβγ. (A) CUL3NTD and Gβγ bind directly and independently to ALFA-tagged KCTD5 (ALFA-K5). Coomassie-stained SDS-PAGE. ALFA-resin fractions were eluted with SDS-sample buffer after washing the resin to remove unbound components. (B) BLI of sensor-anchored KCTD5 with CUL3NTD (Top) and Gβγ (Bottom). (C–E) anti-Gβ1 western blots. (C) CUL3Nedd8/Rbx1/UBE2L3/ARIH1 ubiquitylates Gβ in a KCTD5-dependent manner (all reactions 2 min, 37 °C). (D) CRL3KCTD5/ARIH1/UBE2L3 catalyzes the ubiquitination of a single site on Gβ and has moderate ubiquitin-K48 chain extension activity. K5: KCTD5; K0: ubiquitin with all seven native lysines replaced with arginines; K48R ubiquitin with a single K48R substitution, K48 only: ubiquitin with six lysine to arginine substitutions, preserving the native lysine at position 48. (E) CRL3KCTD5 uses UBE2L3/ARIH1 but not UBE2D2 for the attachment of the initial ubiquitylation of Gβ. All experiments were repeated at least three times, and representative data are shown.
A Reconstituted CRL3KCTD5 Complex Ubiquitylates Gβγ.
We assembled a defined ubiquitylation system to show that CRL3KCTD5 can directly ubiquitylate Gβ subunits, consistent with previous cell-based experiments (17). The reaction requires the E1 protein UBE1, the E2 protein UBE2L3, neddylated CUL3, the RBR E3 ligase ARIH1, and KCTD5 (Fig. 1 C–E). Thus, CRL3KCTD5 uses an E3-E3 superassembly with ARIH1 for the substrate priming reaction, as previously shown for several CRL1 complexes (31, 34). The widely used E2 enzyme UBE2D2, which can also directly ubiquitin some substrates (37, 38), was not active under our reaction conditions (Fig. 1E). We were also able to detect the ubiquitylation of Gγ subunits, but the reaction had a higher background, so we focused primarily on Gβ ubiquitylation as a readout for the reaction (SI Appendix, Fig. S3). The small amount of Gβ with more than one ubiquitin was due to a limited amount of ubiquitin K48 chain formation with UBE2L3/ARIH1, and reactions with a ubiquitin with all seven native lysines replaced with arginines (K0 ubiquitin), and a K48R ubiquitin mutant shows that there is a single ubiquitylation site on Gβ (Fig. 1D).
Structure of the KCTD5/CUL3NTD/Gβγ Complex.
KCTD5 has two main folded regions: a BTB domain from residues 45 to 150 and a CTD from 156 to 209. KCTD5 crystallized under two different conditions revealed that the BTB and CTD domains form independent homopentamers with roughly aligned symmetry axes and that these can adopt conformational states that differ primarily by a rotation of ~30° between the N-terminal BTB and C-terminal CTD moieties (36). Molecular dynamics simulations have shown even greater degrees of flexibility between the two domains (20, 39). We determined the structure of a complex containing full-length KCTD5, CUL3NTD, and Gβγ by cryoelectron microscopy. Complexes were prepared in situ by directly mixing stock solutions of the three proteins prior to spotting onto cryogrids and plunge freezing. Initial map construction converged to volumes for either KCTD5BTB/CUL3NTD or KCTD5CTD/Gβγ, and maps with density for full complexes could only be obtained by dividing the particles images into subsets, indicative of large-scale internal motions (Fig. 2 and SI Appendix, Figs. S4 and S5). Maps generated from the particle image set in which either the bottom KCTD5BTB/CUL3NTD region or the top KCTD5CTD/Gβγ region were subtracted (“top only” and “bottom only” maps, respectively) showed improved detail with resolutions of 2.9 Å and 3.5 Å resolution, respectively. Despite minor improvements upon imposing C5 symmetry during map refinements, all final map calculations were carried out without the imposition of symmetry, and we used these subtracted maps to build models for KCTD5CTD/Gβγ and KCTD5BTB/CUL3NTD components of the complex. We also obtained maps for multiple conformational states of the entire complex via the subdivided particle image sets (SI Appendix, Fig. S5). A recent report of the structure of a two-component KCTD5/Gβγ complex required a head-to-tail fusion of KCTD5 with Gγ to stabilize the assembly for cryo-EM (20). In addition, C5 symmetry was imposed in the refinements. Our structures of the three-component KCTD5/CUL3NTD/Gβγ complex were obtained without artificial cross-links or imposed symmetry and reveal large-scale internal motions and pronounced deviations from C5 symmetry. These features have important consequences for the mechanism of substrate ubiquitylation and are described in the sections below.
Fig. 2.

Structure of the pentameric KCTD5/CUL3NTD/Gβγ complex. (A) Cryo-EM map of the KCTD5CTD/Gβγ “top” part of the complex. Densities for KCTD5 are colored different shades of red. Gβ is blue and Gγ is purple. (B) Cryo-EM map of the KCTD5BTB/CUL3NTD “bottom” part of the complex. CUL3NTD is green. (C) Side view of the model fit to a map of the intact complex showing only one Gβγ and CUL3NTD chain for clarity. (D) Contact interfaces in the complex as per panel C in an “open book” format with buried surfaces in gray. The primary and distal interfaces from adjacent subunits in “c” are indicated with dark/light shades, respectively. (E) Schematic of the interfaces with key residues indicated. L209* is KCTD5 with a stop codon introduced at residue position 209. (F) Ubiquitylation activity of KCTD5 mutants. (G) Ubiquitylation activity of Gβ mutants. F and G are anti-Gβ western blots.
KCTD5CTD/Gβγ Subcomplex.
In the top half of the complex, each of five Gβγ units in the pentameric complex interacts with three adjacent chains of the KCTD5 CTD in two contact patches (Fig. 2 A–D and SI Appendix, Fig. S6 and Movie S1). Five Gβγ units bind edge-on and project radially from the central KCTD5CTD pentamer. All KCTD5CTD contacts involve Gβ only and there are no interactions with Gγ. There are no Gβγ interactions with KCTD5BTB. Remarkably, the two contact patches on Gβ are similar to those engaged by Gα subunits (40) and GRK2 (41) (SI Appendix, Fig. S7) despite the lack of homology between the binding partners. As a result, Gα, GRK2, and KCTD5 binding are likely to be mutually exclusive, suggesting that KCTD5 interacts with free Gβγ. Remarkably, KCTD12, a pentameric KCTD protein that does not interact with CUL3, uses a C-terminal H-domain to engage 5 Gβγ units using an unrelated architecture involving different structural elements to regulate GABAB receptors (42). The H1 fold is not compatible with the pentamers formed by the KCTD5 CTD. In the major contact between KCTD5 CTD and Gβγ (patch “a” in Fig. 2 C and D), the outermost strand of Gβ blade 1 comprising residues N88-R96 nestles into a shallow groove formed by β1, α1, and β3 from a KCTD5CTD subunit. This surface also includes contacts between L55 and D76 from Gβ blade 1 to residues T169 and S173 in the KCTD5CTD helix from the adjacent subunit. These contacts bury a similar surface on Gβ as the N-terminal helix of Gα in G protein heterotrimers (40). This interaction surface has a small nonpolar center (KCTD5 L161, W179, and Gβ A92, P94) flanked by basic residues K78, K89, and R96 on Gβ and acidic residues E167 and D177 on KCTD5.
The second contact patch (“b” in Fig. 3 C and D) involves residues from the KCTD5 C-terminal tail that follows the BTB CTD, residues 210 to 234. In the crystal structures of KCTD5, no density was observed for this region, suggesting that these residues in KCTD5 are disordered in the uncomplexed protein (36). The KCTD5 tail is not included in the recent cryo-EM structure of KCTD5 linked to Gβγ (20). In our structure of the complex, the KCTD5 tail snakes under the long Gβ loop 125 to 134 and culminates in an α-helix from residues 222 to 234, which we call the KCTD5 switch helix. The connection from KCTD 214 to 221 is in weak density, but the Gβ loop 125 to 134, which is normally flexible in free Gβγ, makes multiple interchain contacts and is well ordered. Overall, the structure of the KCTD5 C-terminal tail is consistent with the AlphaFold prediction that shows highly exposed residues from 212 to 221 followed by isolated terminal α-helix (https://alphafold.ebi.ac.uk/entry/Q9NXV2). The basic KCTD5 switch helix is wedged between adjacent Gβγ units and interacts mainly with the “hot spot” acidic patch of the first Gβ, notably residues D228 and D246, but also forms minor contacts with blade 2 of the adjacent Gβ unit. The acidic Gβ hotspot has been observed to bind to a wide range of helical peptides (43), and the KCTD5 switch helix adds to this repertoire.
Fig. 3.
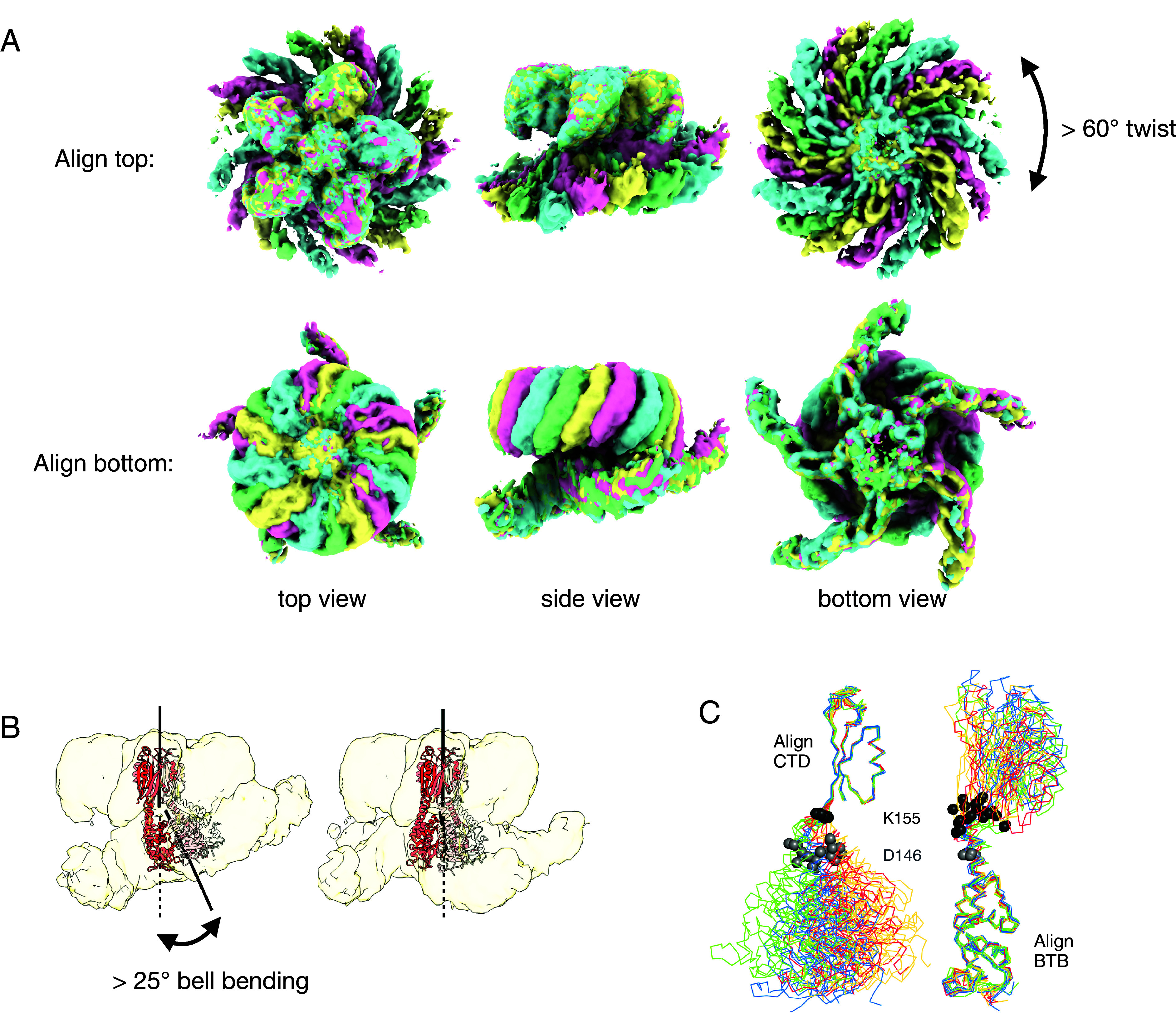
Structural variability in the complex. (A) Alignment of the consensus cryo-EM maps for states A, B, C, and D (green, cyan, pink, and yellow, respectively). Note that the two pseudo-C5 axes between the top and bottom of the complex are not colinear in these consensus maps. (B) Multi-body refinement reveals additional “bell-bending” variability between the two moieties for each state. (C) Structural alignments between the KCTD5 models built into the consensus cryo-EM maps show that the conformational variability in the complex is largely limited to a ~10 residue region between the BTB and CTD domains. Twenty models corresponding to the five chains of states A, B, C, and D are shown, colored according to the maps in A. The Cα atoms of KCTD5 residues 146 and 155 are shown in gray and black spheres, respectively. Bell-binding generates additional variability, but the changes remain localized to the same linker region.
We used structure-guided mutagenesis to test residue substitutions at the KCTD/Gβγ interfaces (Fig. 2 E–G and SI Appendix, Figs. S8 and S9 and Table S1). The main contact patch is required for the KCTD5/Gβγ association, as mutations L161R in KCTD5 and K78E, K89E, or A92D in Gβ disrupt binding without affecting the fold of either protein or CUL3NTD association. Accordingly, these mutations also fully disrupt the ubiquitylation activity of the complex. In contrast, deletion of the KCTD5 tail via mutation L209* reduces the Gβγ binding affinity by approximately 10-fold. Basic substitutions in Gβ at positions D228 or D246 or KCTD5 mutant L209* reduce but do not fully disrupt activity, demonstrating that the KCTD5 tail is of secondary importance in stabilizing the complex.
Consensus refinement of CTD/Gβγ with no imposed symmetry restrains reveals deviations from C5 symmetry in this part of the complex. The angles between the adjacent Gβγ subunits range between 70° and 74° with an average of 72° as expected for fivefold symmetry. Notably, the sandwiched KCTD5 C-terminal helix density is weaker when the Gβγ arms are closer together and stronger when they are farther apart, suggesting that a fully engaged KCTD5 helix pushes the Gβγ units apart from the ideal 72° spacing. In other words, it appears that the pentameric complex cannot accommodate 5 fully engaged terminal helices at the same time, indicating a degree of frustration in the complex. A 3DFlex analysis (44) of the dynamics in the complex shows that the outer edges of the Gβγ units, which include the N-terminal helices from Gβ and Gγ, are more dynamic than the central core near the pseudo-C5 axis (Movie S1).
KCTD5BTB/CUL3 Subcomplex.
In the bottom part of the complex, the CUL3NTD interacts with two adjacent KCTD5BTB domains (contact patch “c” in Fig. 2 C and D), consistent with earlier modeling studies (6, 8) and a recently determined KCTD7 structure (20). All reported BTB/CUL3, SKP1/CUL1, ELOC/CUL2, and ELOC/CUL5 structures involve a common primary interface and a second smaller distal interface that varies widely between substrate adaptor/cullin pairs (4, 6, 45). A major component of the primary interface is the α2/v-loop/β3 region from the common BTB/SKP1/ELOC fold (6, 45) (SI Appendix, Fig. S10), which consists of residues F69-R94 in KCTD5. These elements contact H2, H4, and H5 in the cullin-repeat 1 (CR1) domain of their cognate cullins and include residues that are displaced by CAND1 to promote SR exchange (46, 47). In the dimeric KLHL and SPOP BTB families, the v-loop includes the critical φ-X-E motif (4, 48), but this motif is not found in the v-loops of the KCTD family (6), even though members of both families bind to CUL3 with nanomolar affinity. In the adjacent distal surface, the short KCTD5 helix α1 (R59 to R64) on the adjacent BTB subunit fits into a shallow groove between CUL3 helices H1 and H2. In an apparent example of convergent evolution, the distal interface engages nonhomologous elements from the three-box region of the BACK domain from the same chain in KLHL/SPOP, or from the F-box or BC-Box in SKP1 and ELOC, respectively (SI Appendix, Fig. S10). As expected, mutation at KCTD5 position F128 in the BTB domain disrupted the interaction with CUL3NTD (6) but not with Gβγ, and abolished the ubiquitylation activity of the complex (Fig. 2 E and F and SI Appendix, Figs. S8 and S9 and Table S1).
Pentameric KCTDBTB rings often deviate from C5 symmetry, and “ring opening” has been observed in KCTD BTB domains (6–8). This is recapitulated in the bottom-only consensus cryo-EM map in which there are no contacts between one pair of adjacent BTB domains (Fig. 2B). A 3DFlex (44) analysis of the KCTD5BTB/CUL3NTD subcomplex reveals structural states ranging from a closed BTB pentamer to an open ring (Movie S1). One consequence of these dynamics is that the CUL3NTD chain at the open ring position engages KCTD5 via the primary interface only and does not make the distal contact (SI Appendix, Fig. S11). Notably, the map density for this CUL3 chain is weaker than for the other four chains, suggesting partial occupancy for this subunit due to reduced affinity.
Structure and Dynamics in the Intact KCTD5/CUL3/Gβγ Complex.
We were able to obtain maps for the entire complex only by subdividing the cleaned particle image set into subgroups; these maps differed in the relative positions of the top and bottom of the complex. Larger numbers of subgroups (with smaller numbers of particle images in each) lead to finer differences between the maps, suggesting continuous motions in the complex rather than discrete heterogeneity. We tested different methods to analyze heterogeneity in this dataset (44, 49, 50) with full and subdivided particle image sets. We found that multi-body refinement (49) on four subsets was best suited to the current problem and yielded clear improvements for both moieties of the structures. We first generated four ~4 Å maps from consensus refinements on particle image subsets A, B, C, and D; these had unambiguous density for the intact complexes but still had significant smearing in the KCTD5BTB/CUL3NTD regions, and we combined the top and bottom focused maps to improve these densities (SI Appendix, Table S2). For each of the four substates, we generated 10 composite multi-body maps for a total of 40 maps (Fig. 3 and SI Appendix, Fig. S5). In principle, map density at the interfaces between bodies is ill-defined, but we were able to unambiguously determine the connectivity between the BTB and CTD domains despite weaker density in these regions (SI Appendix, Fig. S12).
The resulting models differ mainly by on-axis rotations of over 60° (“twist”) between the two pseudo-C5 moieties and by rotations of up to 25° about an axis perpendicular to the pseudo-C5 axes (bell-bending) (Movie S2). The consensus maps A, B, C, and D mostly capture the twisting component of the ensemble of states, while the multi-body component maps mostly reveal bell-bending in each state. There are also on-axis translations of up to 5 Å. There are few contacts between the two halves of the complex, and CUL3NTD and Gβγ come into direct contact only at higher bell-bending angles.
Essentially all of the structural variability in the complex can be localized to a 10-residue connection between the BTB and CTD domains of KCTD5 (Fig. 3C), and this is recapitulated in an analysis of the crystal structures of uncomplexed KCTD5 (36) (SI Appendix, Fig. S13). The hinge region begins at residue D146 in the last α-helix of the BTB domain and ends at K155 prior to the first β-strand of the CTD. The flexibility in the hinge, which accounts for twist, bell-bending, and translation, appears to be evenly distributed over the 10 residues. KCTD5 homologs have low sequence conservation in this region, apart from a composition of mostly hydrophilic amino acids (SI Appendix, Fig. S13).
CRL3KCTD5 Positions Gβγ for Ubiquitylation.
We generated models of intact CRL3KCTD5 complexes by extending CUL3 to include structures of the ubiquitylation machinery assembled at the C-terminal half of the cullin. Modeling was based on the CUL1/RBX1/ARIH1(Rcat)~ubiquitin components of the TS2 state of a CRL1FBXW7 structure (31). In the model, all five CUL3 arms arc toward the CTD of the KCTD5 pentamer in a suprafacial arrangement (11) while allowing just enough space for the inclusion of Gβγ. The arms are positioned such that all five ubiquitin C termini face the central pseudo-C5 axis of the pentamer and do not produce unfavorable clashes over the ensemble of conformations (Fig. 4 and Movie S2). The twist and bell-bending motions between the two moieties of the complex generate a “cloud” of ARIH1(Rcat)~ubiquitin reactive sites that populate a solid toroid that surrounds the Gβγ surface (Fig. 5 B and C). Some of the structural states result in close approaches between the activated ubiquitin C termini and the Gβγ units, notably surface-exposed lysines K23 on Gβ and K29, K32, and K46 on Gγ (Fig. 4B and SI Appendix, Fig. S14). Of these, Gβ K23 is the most conserved across the human isoforms (SI Appendix, Fig. S15). Although only a fraction of the conformational states are poised for ubiquitin transfer to Gβ K23, this appears to be sufficient for the reaction, as CRL3KCTD5 ubiquitylates Gβ exclusively at K23 (Fig. 4D and SI Appendix, Fig. S16). It is important to note that the ARIH1(Rcat)~Ub transferase module is highly dynamic relative to the cullin N-terminal domain (31), and these additional degrees of freedom would further expand the volume of the reactive cloud.
Fig. 4.

Model of an active CRL3KCTD5 complex. (A) Side view of conformational state B of the KCTD5/CUL3NTD/Gβγ complex with one CUL3 arm modeled to include the C-terminal catalytic machinery based on the TS2 state of the CRL1FBXW7 complex (PDB ID 7B5M). Rbx1 is in gray, AriH1 (Ariadne and Rcat domains) is in white, and ubiquitin is in yellow. Only one CUL3 arm and one Gβγ unit are shown on the KCTD5 pentamer for clarity. The conformational state shown here places the NZ atom of Gβ K23 within 9 Å of the ubiquitin G75 CA atom with an unobstructed path to the thioester bond of the ubiquitin/AriH1~ubiquitin conjugate. The structure of the complex is consistent with membrane-anchored Gβγ. (B and C) Surface representation of one conformer of the extended complex. The small yellow circles indicate the positions of the ubiquitin G75 Cα in the ensemble of the conformations of the complex based on multi-body maps for states A–D. (D) Ubiquitylation of Gβ WT and point mutants (anti-Gβ western blot). (E) Kinetics of the ubiquitylation reaction for KCTD5 WT (black) and linker deletion mutants Δ2 (green), Δ4 (red), and Δ7 (blue). Error bars are SD with n = 3.
Fig. 5.

Reducing KCTD5 levels alters Gβγ expression and ubiquitination patterns. (A) The level of Gβγ is altered with KCTD5 knockdown or overexpression. BiFC was measured between the split YFP fusion proteins Gβ1-NYFP(1–158) and Gγ2-CYFP(159–238) in the presence or absence of overexpressed HA-KCTD5 or 100 nM siRNA. Fluorescence intensities are presented as mean ± SEM for four different experiments. Data were analyzed by two-way ANOVA followed by Tukey’s multiple comparisons test where **P < 0.01. (B) Pan-Gβ1-4 expression measured by western blot in parental and KCTD5 KO cell lines stably expressing either the WT, the C-terminal truncated (1 to 208*), F128A, or L161R KCTD5 mutants. For the sake of comparison, both parental and KCTD5 KO cell lines expressed a non-KCTD5 control protein (Venus, a YFP). Bars represent the average values from five independent experiments expressed as relative values normalized to expression of β-tubulin. Error bars are SEM. Statistical analysis was done by one-way ANOVA followed by a Dunnett post hoc test, *P < 0.05, **P < 0.01, and ns means nonsignificant. (C) KCTD5 levels alter ubiquitylation of Gβγ and their interacting proteins. Western blot analysis from parental and KCTD5 KO lines expressing FLAG-ubiquitin, HA-TAP-Gβ1, and HA-Gγ2 that were treated or not with 10 μM MG-132 for 8 h, lysed, and subjected to streptavidin purification. Blots are representative of three independent experiments.
In order to test the effect of KCTD5 interdomain flexibility on the priming reaction, we measured the ubiquitylation rates of a series of KCTD5 linker deletion mutants (Fig. 4E and SI Appendix, Fig. S17). We used K0 ubiquitin for these studies to monitor the priming reaction. Deletions of 2 and 4 residues (Δ2, Δ4) had no measurable effect of the reaction rates, while a seven-residue deletion (Δ7) resulted in a significantly slower reaction. While we expect a shortened linker to reduce the internal flexibility of the complex, it is difficult to predict the degree of this effect because the unwinding of BTB helix α6 (residues S147 to S150) is seen in some structural states (Fig. 3 and SI Appendix, Figs. S6 and S13). Thus, helix unwinding might compensate for the shortening of the linker in the Δ2 and Δ4 deletions. Alternatively, these results might indicate that there are specific interactions between the CUL3 catalytic machinery and Gβγ, leading to the selection of orientations between activated ubiquitin and the substrate, and productive orientations are compatible with Δ2 and Δ4 but not Δ7. The participation of substrate in the priming reaction has been recently described in a CRL2FEM1C-Sil1-UBE2R2~ubiquitin complex (51) but has not yet been described in a CRL3 system. These interpretations are not exclusive, and we plan to address these issues in future work.
KCTD5 Regulates Gβγ Levels in Cells.
Validating the KCTD5/Gβγ interaction from ref. 14, we showed that KCTD5 could be pulled down with TAP-Gβ1 and not with TAP-Smad2 (SI Appendix, Fig. S18). Pulling down KTCD5-FLAG, we also show that the coimmunoprecipitation was bidirectional. We next used bimolecular fluorescence complementation between Gβ and Gγ subunits and examined the effects of knocking down KCTD5 in HEK 293 cells. Knocking down KCTD5 with siRNA increased the levels of Gβγ pairs (Fig. 5A) and were reduced by overexpressing KCTD5. Next, to confirm effects on native Gβ levels, we used western blotting with an antibody that recognizes Gβ1-4 in parental cells and in a line of HEK 293 cells where we knocked out KCTD5 using CRISPR/Cas9. In KCTD5 KO cells, the levels of Gβ are significantly increased compared to parental cells (see column 1 vs. 2 in Fig. 5B). Restoring expression of KCTD5 WT lowered the levels of detected Gβ, but KCTD5 mutants L209*, F128A and L161R did not (Fig. 5B). These mutants disrupt Gβγ binding, although the deletion of the KCTD5 C-terminal tail in L209* retained some biochemical activity (Fig. 2 and SI Appendix, Figs. S1, S8, and S9 and Table S1). Next, we examined the ubiquitylation of Gβ using TAP tagging. Gβ was pulled down in parental cells, with and without treatment with the proteasome inhibitor MG-132 (Fig. 5 C, Left). MG-132 treatment increases Gβ ubiquitylation, demonstrating the involvement of the 26S proteasome in regulating Gβ levels. In contrast, very little ubiquitylation is seen in the KCTD5 KO cells and levels of Gβ are increased but insensitive to MG-132 treatment (Fig. 5 C, Right), concordant with the role for KCTD5 as a substrate receptor for Gβ regulation by CUL3-mediated proteasomal degradation. Gβ1 mutations K23A and K23R reduced the amount of ubiquitylated Gβγ in HEK 293 cells (SI Appendix, Fig. S19), consistent with K23 as a major target residue for ubiquitin attachment.
Discussion
The CRL framework depends on the internal dynamics between multiple structural blocks to adapt to a variety of substrates. For example, there are large reorganizations of the UBE2D~ubiquitin or ARIH1(Rcat)~ubiquitin transferase modules relative to the substrate scaffolding module over the catalytic cycle (30, 31). At the substrate end, intrinsically disordered degrons or phosphodegrons can occupy an extended, fuzzy volume (52), and ubiquitylation can occur only when the volumes sampled by the activated ubiquitin thioester and substrate nucleophiles intersect (53). This is a necessary condition, but additional factors, such as nucleophile positioning, are important. With ordered substrates, the requirements for close approach and proper orientation are likely more difficult to achieve and can be expected to be case-specific. The CRL3KCTD5 system described here includes large-scale internal motions in the KCTD5 substrate receptor within a pseudo-C5 pentamer. We propose that the partial decoupling of the KCTD5CTD/Gβγ block from the KCTD5BTB/CUL3NTD block allows the rigid Gβγ to approach and interact with the ubiquitin transferase module to position the target lysines in the priming reaction (Fig. 6). The architecture of the complex is remarkably tuned to the substrate—the shape and position of Gβγ on KCTD5CTD, the geometry of the KCTD5BTB/CUL3 interface, the arc of the cullin, and the position of the ARIH1~Ub on the modeled transferase module—all contribute to the favorable placement of the active groups with little evidence of steric clashes.
Fig. 6.

Model for the role of CRL dynamics in positioning an activated ubiquitin C terminus near to a target substrate. S: substrate, SBD: substrate-binding domain, UCE: ubiquitin-carrying enzyme. See text for details.
The CRL3KCTD reaction complex we describe here is compatible with membrane-bound Gβγ (Fig. 4), leading to a model where the recruitment of pentameric KCTD5 nucleates a cluster of Gβγ and CUL3 complexes at inner membrane surfaces. KCTD5-driven ubiquitylation can modulate Gβγ levels in cells (Fig. 5 and refs. 18–20), and it will be important to determine how this activity is regulated. Possible mechanisms include posttranslational modifications (although none are apparently required for substrate binding), component concentrations, and subcellular localizations. The existence of substoichiometric CRL3KCTD5 complexes with incomplete occupancy would lead to unproductive states, and activity may depend on threshold effects due to the concentrations and availability of limiting factors. The fast substrate exchange kinetics and slow CUL3 dissociation rates we describe here have been observed in other CRL complexes (54). Component availability and exchange rates, both spontaneous and catalyzed, are widely used strategies in the regulation of CRL1 networks (46), and we expect these effects to be at work in CRL3KCTD networks as well.
The KCTD family of proteins are involved in a wide range of biological processes and participate in a variety of cellular signaling pathways (23–25). Members of clades E (KCTD2/5/17) and F (KCTD8/12/16) use distinct binding modes to engage Gβγ subunits and down-regulate GPCR signaling through CUL3-dependent and CUL3-independent mechanisms, respectively (SI Appendix, Fig. S7 and refs. 16–19). Published studies of phenotypic effects of KCTD proteins demonstrate a remarkable plasticity which likely results from different levels or subtypes of KCTD proteins present in the different cell models used. The regulation of the levels or activity of distinct populations of Gβγ subunits via KCTD proteins provides a rationale for the pleiotropic effects in distinct cells, as Gβγ subunits are central players in the organization and regulation of cellular signaling (27, 28, 55).
Materials and Methods
Protein Expression and Purification.
Expression plasmids and mutants for KCTD5, Gβ1, Gγ2, CUL3, and RBX1 were made by gene synthesis (Genscript) unless noted otherwise. Human KCTD5 was expressed as thioredoxin-6His-(TEV)-AviALFA-(3C)-KCTD5 or 6xHis-SUMO-(TEV)-AviALFA-(3C)-KCTD5 from pET32 vectors in Escherichia coli (the bracketed terms indicate cleavage sites for TEV and 3C proteases, respectively). CUL3NTD (residues 1 to 381), UBA1, UBE2L3, and ARIH191–557 were expressed as N-terminal His-tagged fusions from pMCSG7, pET28, pET28aLIC, and pProExHTb vectors in E. coli, respectively. E. coli–expressed proteins were purified by Ni-NTA affinity chromatography, and tags were removed by TEV, 3C, or thrombin digestion. Proteins were further purified by size-exclusion chromatography into buffer A (25 mM HEPES pH 7.5, 100 mM NaCl, 0.1 mM TCEP, and 5% glycerol) prior to concentration and flash freezing in liquid nitrogen.
Gβ1γ2 and CUL3/RBX1 were prepared by coexpression in HEK 293 cells. Gβ1 was cloned into a piggyBac vector (PB) (56) without any tags. Full-length Gγ2(C68S), a prenylation-deficient variant, was inserted into a PB vector downstream of GFP(3C). Plasmids for Gβ1 and GFP-Gγ2 were mixed with PBase and PB-RB vectors (56) and transfected into HEK 293 GnT1−/− cells. Stable cell lines were selected for resistance to puromycin and blasticidin and scaled up in suspension cultures using FreeStyle 293 media (ThermoFisher). Expression was induced with doxycycline and cells were harvested after 48 to 72 h. GFP-tagged proteins were purified from cell lysates on GFP affinity columns and eluted with 3C protease. Proteins were further purified by size-exclusion chromatography into 25 mM HEPES pH 7.5, 100 mM NaCl, 0.1 mM TCEP, and 5% glycerol prior to concentration and flash freezing in liquid nitrogen. All Gβγ proteins in this study were in the Gβ1γ2(C68S) background. Proteins were further purified by size-exclusion chromatography into buffer A prior to concentration and flash freezing. GFP(3C)-CUL3 and untagged RBX1 were coexpressed in HEK 293 cells and purified as described for Gβγ, except that the protein was neddylated on-resin with NAE1/UBA3, UBE2M, and NEDD8 (R&D Systems) prior to 3C elution.
Size-Exclusion Chromatography.
Analytical size-exclusion chromatography was carried out on a Waters HPLC with an ENrichTM SEC 650 10 × 300 column (Bio-Rad) in 20 mM HEPES pH 8.0, 150 mM NaCl, and 0.1 mM TCEP at a flow rate of 1.0 mL/min.
Copurification Binding Assay.
ALFA-tagged KCTD5 was added to anti-ALFA nanobody resin in buffer A and washed three times. CUL3NTD and/or Gβγ were added and incubated at 4 °C for 15 min, followed by a second wash and elution into SDS sample buffer. Samples were analyzed by SDS/PAGE and visualized with Coomassie staining.
In Vitro Ubiquitylation Assay.
We adapted a pulse–chase ubiquitylation assay (34) to measure CRL3KCTD5 activity. Ubiquitin-charged UBE2L3 or UBE2D2 (E2~Ub) were generated with 0.3 µM mUBA1, 10 µM UBE2L3 or UBE2D2, and 10 µM ubiquitin in 25 mM HEPES pH 7.5, 150 mM NaCl, 5 mM MgCl2, and 1 mM ATP at 37 °C for 15 min and quenched with 10 U/µL apyrase. Reactions were initiated by mixing equal volumes of E2~Ub with neddylated CUL3/RBX1, KCTD5, Gβ1γ2, and ARIH1 for UBE2L3~Ub. Reactions were stopped with SDS-PAGE sample buffer and analyzed by western blotting with anti-Gγ2 (7-RE20, Santa Cruz sc-134344) and anti-Gβ1 (Sigma-Aldrich sab2701168). %Gβ ubiquitylation was calculated as 100*a/(a+b), where a is the integrated intensity of the Gβ~Ub band, and b is the integrated intensity of the Gβ band.
Biolayer Interferometry.
An Octet RED384 system (Sartorius) instrument was used for biolayer interferometry. Thioredoxin-fusion KCTD5 was anchored onto anti-Penta-His (HIS1K) biosensors and dipped into wells containing CUL3NTD or Gβγ in 20 mM HEPES pH 8, 150 mM NaCl, 1 mM TCEP, and 0.05% Tween-20. A different sensor was used for each CUL3NTD concentration because of the slow off-rate for this analyte. Instrument software was used to process and fit the data to a 1:1 kinetic model and a steady-state model.
Cryoelectron Microscopy and Model Building.
Cryo-EM samples were prepared by incubating a mixture of 12 µM KCTD5, 18 µM CUL3NTD, and 18 µM Gβγ in 25 mM HEPES pH 8.0, 150 mM NaCl, and 0.1 mM TCEP at 4 °C for 30 min. The solution was applied to glow-discharged homemade nanofabricated holey gold grids (57) in a Vitrobot Mark IV and vitrified by plunging into liquid ethane. High-resolution data were collected on a Titan Krios G3 electron microscope (Thermo Fisher Scientific) operated at 300 kV equipped with a prototype Falcon 4 direct detector device camera. Movies were collected at a nominal magnification of 75,000×, corresponding to a calibrated pixel size of 1.03 Å, and a total exposure of ∼49.3 e−/Å2 over 40 frames. Data from untilted movies and 40° tilted movies were processed with cryoSPARC (58). Masks of the initial bottom and top moieties were used in particle subtraction to generate top-only and bottom-only particle image datasets. Ab initio maps were calculated from the subtracted particle sets and used for nonuniform refinement (59). Final maps were sharpened with DeepEMhancer (60). Models for KCTD5 (3DRX), CUL3NTD (4EOZ), and Gβγ (1GP2) were docked into the maps and rebuilt and refined with COOT (61) and Phenix (62) to give final coordinates for KCTD5CTD/Gβγ and KCTD5BTB/CUL3NTD.
The unsubtracted particle image set was subdivided into sets A, B, C, and D with cryoSPARC heterogeneous refinement and imported to RELION (63) for de novo map generation and initial 3D refinement. We improved these maps in two ways. In the first method, we used RELION multi-body refinement (49, 64) with the four RELION consensus maps and masks for the top and bottom bodies. In each of the four cases, the second most abundant component reflected bell-bending and was selected for further analysis. Ten multi-body composite maps for each state were used to build a total of 40 models with Namdinator (65) and further refined with Phenix. The forty multi-body maps and models have been deposited at Zenodo. In the second method, we used phenix.combine_focused_maps with the four RELION consensus maps as templates and the top-only and bottom-only focused maps. The four maps and models from this method have been deposited to the EMDB and PDB (SI Appendix, Table S2). Figures were prepared with ChimeraX (66) and Pymol (67). Software used in this project was curated by SBgrid (68).
Further details regarding the methods used in this study are available in SI Appendix.
Supplementary Material
Appendix 01 (PDF)
Cryo-EM maps of the dynamics in the top and bottom parts of the complex as revealed by 3DFlex analysis.
Dynamics in the CRL3KCTD5/Gβ1γ2 complex, including the extended model that includes full-length CUL3 (green), RBX1 (grey), ARIH1 (white) and ubiquitin (yellow). The C-terminus of ubiquitin is shown as a yellow sphere.
Acknowledgments
This research was funded by Canadian Institutes of Health Research (CIHR) grant PJT-191900 to G.G.P. D.M.N. was supported by a CIHR Canada Graduate Scholarship - Masters’s Award. We thank Frank Sicheri, Samuel Lunenfeld Institute, for the expression vector for ARIH1 and the Structural Genomics Consortium for expression vectors for UBE1 and UBE2L3. T.E.H. is supported by funding from the Heart and Stroke Society of Canada and PJT-159687 from CIHR. T.E.H. holds the Canadian Pacific Chair in Biotechnology. We thank Tiffany Villleneuve and Fenneke KleinJan for instrument support, Dr. Richard Wargachuk for construction of KCTD5-FLAG, and Dr. Tamara Ouspenskaia for construction of TAP-Smad2. Titan Krios cryo-EM data were collected at the Toronto High-Resolution High-Throughput Cryo-EM facility supported by the Canada Foundation for Innovation and Ontario Research Fund.
Author contributions
D.M.N., D.H.R., D.D., A.X.J., N.N., D.Y., A.Z., D.A.K., M.U.Q.M., J.L.R., T.E.H., and G.G.P. designed research; D.M.N., D.H.R., D.D., D.P., R.R., A.X.J., N.N., D.Y., A.Z., D.A.K., M.U.Q.M., N.C.P., A.F.A.K., S.B., J.L.R., and G.G.P. performed research; D.M.N., D.H.R., D.D., M.T.M.-J., J.L.R., and G.G.P. contributed new reagents/analytic tools; D.M.N., D.H.R., D.D., T.E.H., and G.G.P. analyzed data; and D.M.N., D.D., T.E.H., and G.G.P. wrote the paper.
Competing interests
The authors declare no competing interest.
Footnotes
This article is a PNAS Direct Submission. J.D.G. is a guest editor invited by the Editorial Board.
Data, Materials, and Software Availability
The cryo-EM maps and associated atomic models of top and bottom complex and conformational states A, B, C, and D have been deposited in the Electron Microscopy Data Bank with accession codes EMD-41994 (69), EMD-41995 (70), EMD-41996 (71), EMD-41997 (72), EMD-41998 (73), EMD-41999 (74), EMD-42000 (75), EMD-42001 (76), EMD-42002 (77), EMD-42003 (78), EMD-42004 (79), EMD-42005 (80), EMD-42006 (81), EMD-42007 (82), EMD-42008 (83), EMD-42009 (84), EMD-42010 (85), EMD-42011 (86) and the Protein Data Bank with accession codes 8U7Z (87), 8U80 (88), 8U81 (89), 8U82 (90), 8U83 (91), and 8U84 (92). The 40 maps and models from the multi-body analysis are available from Zenodo at https://zenodo.org/record/8341597 (93). Microscope movies and particle image sets are available from EMPIAR at https://www.ebi.ac.uk/empiar/EMPIAR-11734 (94).
Supporting Information
References
- 1.Rusnac D.-V., Zheng N., Structural biology of CRL ubiquitin ligases. Adv. Exp. Med. Biol. 1217, 9–31 (2020). [DOI] [PubMed] [Google Scholar]
- 2.Harper J. W., Schulman B. A., Cullin-RING ubiquitin ligase regulatory circuits: A quarter century beyond the F-box hypothesis. Annu. Rev. Biochem. 90, 403–429 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang P., Song J., Ye D., CRL3s: The BTB-CUL3-RING E3 ubiquitin ligases. Adv. Exp. Med. Biol. 1217, 211–223 (2020). [DOI] [PubMed] [Google Scholar]
- 4.Errington W. J., et al. , Adaptor protein self-assembly drives the control of a cullin-RING ubiquitin ligase. Structure 20, 1141–1153 (2012). [DOI] [PubMed] [Google Scholar]
- 5.Canning P., et al. , Structural basis for Cul3 protein assembly with the BTB-Kelch family of E3 ubiquitin ligases. J. Biol. Chem. 288, 7803–7814 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ji A. X., et al. , Structural insights into KCTD protein assembly and Cullin3 recognition. J. Mol. Biol. 428, 92–107 (2016). [DOI] [PubMed] [Google Scholar]
- 7.Smaldone G., et al. , The BTB domains of the potassium channel tetramerization domain proteins prevalently assume pentameric states. FEBS Lett. 590, 1663–1671 (2016). [DOI] [PubMed] [Google Scholar]
- 8.Pinkas D. M., et al. , Structural complexity in the KCTD family of Cullin3-dependent E3 ubiquitin ligases. Biochem. J. 474, 3747–3761 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cuneo M. J., O’Flynn B. G., Lo Y.-H., Sabri N., Mittag T., Higher-order SPOP assembly reveals a basis for cancer mutant dysregulation. Mol. Cell 83, 731–745.e4 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Toma-Fukai S., Shimizu T., Structural diversity of ubiquitin E3 ligase. Molecules 26, 6682 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tang X., et al. , Suprafacial orientation of the SCFCdc4 dimer accommodates multiple geometries for substrate ubiquitination. Cell 129, 1165–1176 (2007). [DOI] [PubMed] [Google Scholar]
- 12.Hao B., Oehlmann S., Sowa M. E., Harper J. W., Pavletich N. P., Structure of a Fbw7-Skp1-cyclin E complex: Multisite-phosphorylated substrate recognition by SCF ubiquitin ligases. Mol. Cell 26, 131–143 (2007). [DOI] [PubMed] [Google Scholar]
- 13.Ahmed S. M., Daulat A. M., Meunier A., Angers S., G protein betagamma subunits regulate cell adhesion through Rap1a and its effector Radil. J. Biol. Chem. 285, 6538–6551 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Campden R., et al. , Tandem affinity purification to identify cytosolic and nuclear gβγ-interacting proteins. Methods Mol. Biol. Clifton NJ 1234, 161–184 (2015). [DOI] [PubMed] [Google Scholar]
- 15.Zha Z., et al. , A non-canonical function of Gβ as a subunit of E3 ligase in targeting GRK2 ubiquitylation. Mol. Cell 58, 794–803 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brockmann M., et al. , Genetic wiring maps of single-cell protein states reveal an off-switch for GPCR signalling. Nature 546, 307–311 (2017). [DOI] [PubMed] [Google Scholar]
- 17.Young B. D., Sha J., Vashisht A. A., Wohlschlegel J. A., Human multisubunit E3 ubiquitin ligase required for heterotrimeric G-protein β-subunit ubiquitination and downstream signaling. J. Proteome Res. 20, 4318–4330 (2021). [DOI] [PubMed] [Google Scholar]
- 18.Muntean B. S., et al. , Members of the KCTD family are major regulators of cAMP signaling. Proc. Natl. Acad. Sci. U. S. A. 119, e2119237119 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sloan D. C., Cryan C. E., Muntean B. S., Multiple potassium channel tetramerization domain (KCTD) family members interact with Gβγ, with effects on cAMP signaling. J. Biol. Chem. 299, 102924 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jiang W., Wang W., Kong Y., Zheng S., Structural basis for the ubiquitination of G protein βγ subunits by KCTD5/Cullin3 E3 ligase. Sci. Adv. 9, eadg8369 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Li Q., et al. , Conserved properties of Drosophila insomniac link sleep regulation and synaptic function. PLoS Genet. 13, e1006815 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Barfield R., et al. , Epigenome-wide association analysis of daytime sleepiness in the multi-ethnic study of atherosclerosis reveals African-American-specific associations. Sleep 42, zsz101 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu Z., Xiang Y., Sun G., The KCTD family of proteins: Structure, function, disease relevance. Cell Biosci. 3, 45 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Skoblov M., et al. , Protein partners of KCTD proteins provide insights about their functional roles in cell differentiation and vertebrate development. BioEssays News Rev. Mol. Cell. Dev. Biol. 35, 586–596 (2013). [DOI] [PubMed] [Google Scholar]
- 25.Teng X., et al. , KCTD: A new gene family involved in neurodevelopmental and neuropsychiatric disorders. CNS Neurosci. Ther. 25, 887–902 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Angrisani A., Di Fiore A., De Smaele E., Moretti M., The emerging role of the KCTD proteins in cancer. Cell Commun. Signal. 19, 56 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tennakoon M., et al. , Subtype-dependent regulation of Gβγ signalling. Cell. Signal. 82, 109947 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Khan S. M., et al. , The expanding roles of Gβγ subunits in G protein-coupled receptor signaling and drug action. Pharmacol. Rev. 65, 545–577 (2013). [DOI] [PubMed] [Google Scholar]
- 29.Khan S. M., Sung J. Y., Hébert T. E., Gβγ subunits—Different spaces, different faces. Pharmacol. Res. 111, 434–441 (2016). [DOI] [PubMed] [Google Scholar]
- 30.Baek K., et al. , NEDD8 nucleates a multivalent cullin-RING-UBE2D ubiquitin ligation assembly. Nature 578, 461–466 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Horn-Ghetko D., et al. , Ubiquitin ligation to F-box protein targets by SCF-RBR E3–E3 super-assembly. Nature 590, 671–676 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mena E. L., et al. , Structural basis for dimerization quality control. Nature 586, 452–456 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jenkyn-Bedford M., et al. , A conserved mechanism for regulating replisome disassembly in eukaryotes. Nature 600, 743–747 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Scott D. C., et al. , Two distinct types of E3 ligases work in unison to regulate substrate ubiquitylation. Cell 166, 1198–1214.e24 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Balasco N., et al. , Molecular recognition of Cullin3 by KCTDs: Insights from experimental and computational investigations. Biochim. Biophys. Acta 1844, 1289–1298 (2014). [DOI] [PubMed] [Google Scholar]
- 36.Dementieva I. S., et al. , Pentameric assembly of potassium channel tetramerization domain-containing protein 5. J. Mol. Biol. 387, 175–191 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wu K., Kovacev J., Pan Z.-Q., Priming and extending: A UbcH5/Cdc34 E2 handoff mechanism for polyubiquitination on a SCF substrate. Mol. Cell 37, 784–796 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hill S., et al. , Robust cullin-RING ligase function is established by a multiplicity of poly-ubiquitylation pathways. eLife 8, e51163 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Barone D., Balasco N., Vitagliano L., KCTD5 is endowed with large, functionally relevant, interdomain motions. J. Biomol. Struct. Dyn. 34, 1725–1735 (2016). [DOI] [PubMed] [Google Scholar]
- 40.Wall M. A., et al. , The structure of the G protein heterotrimer Gi alpha 1 beta 1 gamma 2. Cell 83, 1047–1058 (1995). [DOI] [PubMed] [Google Scholar]
- 41.Lodowski D. T., Pitcher J. A., Capel W. D., Lefkowitz R. J., Tesmer J. J. G., Keeping G proteins at bay: A complex between G protein-coupled receptor kinase 2 and Gbetagamma. Science 300, 1256–1262 (2003). [DOI] [PubMed] [Google Scholar]
- 42.Zheng S., Abreu N., Levitz J., Kruse A. C., Structural basis for KCTD-mediated rapid desensitization of GABAB signalling. Nature 567, 127–131 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Smrcka A. V., Fisher I., G-protein βγ subunits as multi-functional scaffolds and transducers in G-protein-coupled receptor signaling. Cell. Mol. Life Sci. 76, 4447–4459 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Punjani A., Fleet D. J., 3DFlex: Determining structure and motion of flexible proteins from cryo-EM. Nat. Methods 20, 860–870 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stogios P. J., Downs G. S., Jauhal J. J. S., Nandra S. K., Privé G. G., Sequence and structural analysis of BTB domain proteins. Genome Biol. 6, R82 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Baek K., et al. , Systemwide disassembly and assembly of SCF ubiquitin ligase complexes. Cell 186, 1895–1911 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Shaaban M., et al. , Structural and mechanistic insights into the CAND1-mediated SCF substrate receptor exchange. Mol. Cell 83, 2332–2346.e8 (2023). [DOI] [PubMed] [Google Scholar]
- 48.Ji A. X., Privé G. G., Crystal structure of KLHL3 in complex with Cullin3. PLoS ONE 8, e60445 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Nakane T., Kimanius D., Lindahl E., Scheres S. H., Characterisation of molecular motions in cryo-EM single-particle data by multi-body refinement in RELION. eLife 7, e36861 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhong E. D., Bepler T., Berger B., Davis J. H., CryoDRGN: Reconstruction of heterogeneous cryo-EM structures using neural networks. Nat. Methods 18, 176–185 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Li J., et al. , Cullin-RING ligases employ geometrically optimized catalytic partners for substrate targeting. Mol. Cell 84, 1304–185 (2024). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mittag T., et al. , Dynamic equilibrium engagement of a polyvalent ligand with a single-site receptor. Proc. Natl. Acad. Sci. U.S.A. 105, 17772–17777 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Baek K., Scott D. C., Schulman B. A., NEDD8 and ubiquitin ligation by cullin-RING E3 ligases. Curr. Opin. Struct. Biol. 67, 101–109 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pierce N. W., et al. , Cand1 promotes assembly of new SCF complexes through dynamic exchange of F box proteins. Cell 153, 206–215 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Martins-Cannavino K., Hébert T. E., Gβγ signaling from an eponymous past to a specific future. Cell Syst. 12, 289–290 (2021). [DOI] [PubMed] [Google Scholar]
- 56.Li Z., Michael I. P., Zhou D., Nagy A., Rini J. M., Simple piggyBac transposon-based mammalian cell expression system for inducible protein production. Proc. Natl. Acad. Sci. U.S.A. 110, 5004–5009 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Marr C. R., Benlekbir S., Rubinstein J. L., Fabrication of carbon films with ~500nm holes for cryo-EM with a direct detector device. J. Struct. Biol. 185, 42–47 (2014). [DOI] [PubMed] [Google Scholar]
- 58.Punjani A., Rubinstein J. L., Fleet D. J., Brubaker M. A., cryoSPARC: Algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017). [DOI] [PubMed] [Google Scholar]
- 59.Punjani A., Zhang H., Fleet D. J., Non-uniform refinement: Adaptive regularization improves single-particle cryo-EM reconstruction. Nat. Methods 17, 1214–1221 (2020). [DOI] [PubMed] [Google Scholar]
- 60.Sanchez-Garcia R., et al. , DeepEMhancer: A deep learning solution for cryo-EM volume post-processing. Commun. Biol. 4, 874 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Emsley P., Lohkamp B., Scott W. G., Cowtan K., Features and development of Coot. Acta Crystallogr. D, Biol. Crystallogr. 66, 486–501 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Adams P. D., et al. , PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D, Biol. Crystallogr. 66, 213–221 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kimanius D., Dong L., Sharov G., Nakane T., Scheres S. H. W., New tools for automated cryo-EM single-particle analysis in RELION-4.0. Biochem. J. 478, 4169–4185 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nakane T., Scheres S. H. W., Multi-body refinement of Cryo-EM images in RELION. Methods Mol. Biol. Clifton NJ 2215, 145–160 (2021). [DOI] [PubMed] [Google Scholar]
- 65.Kidmose R. T., et al. , Namdinator—Automatic molecular dynamics flexible fitting of structural models into cryo-EM and crystallography experimental maps. IUCrJ 6, 526–531 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pettersen E. F., et al. , UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004). [DOI] [PubMed] [Google Scholar]
- 67.DeLano W. L., The PyMOL Molecular Graphics System (Schrödinger, LLC, 2000). [Google Scholar]
- 68.Morin A., et al. , Collaboration gets the most out of software. eLife 2, e01456 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: Local Refinement of KCTD5(CTD)/Gbeta1gamma2. EMD-41994. https://www.ebi.ac.uk/emdb/EMD-41994. Deposited 15 September 2023.
- 70.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: Local Refinement of KCTD5(BTB)/Cullin3(NTD). EMD-41995. https://www.ebi.ac.uk/emdb/EMD-41995. Deposited 15 September 2023.
- 71.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State A: Composite Map from RELION Multi-body Refinement. EMD-41996. https://www.ebi.ac.uk/emdb/EMD-41996. Deposited 15 September 2023.
- 72.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State A: Reference Map for Composite Map EMD-41996. EMD-41997. https://www.ebi.ac.uk/emdb/EMD-41997. Deposited 15 September 2023.
- 73.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State A Body 1: RELION Multi-body Refinement of KCTD5(CTD)/Gbeta1gamma2. EMD-41998. https://www.ebi.ac.uk/emdb/EMD-41998. Deposited 15 September 2023.
- 74.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State A Body 2: RELION Multi-body Refinemnent of KCTD5(BTB)/Cullin3(NTD). EMD-41999. https://www.ebi.ac.uk/emdb/EMD-41999. Deposited 15 September 2023.
- 75.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State B: Composite Map from RELION Multi-body Refinement. EMD-42000. https://www.ebi.ac.uk/emdb/EMD-42000. Deposited 15 September 2023.
- 76.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State B: Reference Map for Composite Map EMD-42000. EMD-42001. https://www.ebi.ac.uk/emdb/EMD-42001. Deposited 15 September 2023.
- 77.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State B Body 1: RELION Multi-body Refinement of KCTD5(CTD)/Gbeta1gamma2. EMD-42002. https://www.ebi.ac.uk/emdb/EMD-42002. Deposited 15 September 2023.
- 78.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State B Body 2: RELION Multi-body Refinement of KCTD5(BTB)/Cullin3(NTD). EMD-42003. https://www.ebi.ac.uk/emdb/EMD-42003. Deposited 15 September 2023.
- 79.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State C: Composite Map from RELION Multi-body Refinement. EMD-42004. https://www.ebi.ac.uk/emdb/EMD-42004. Deposited 15 September 2023.
- 80.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State C: Reference Map for Composite Map EMD-42004. EMD-42005. https://www.ebi.ac.uk/emdb/EMD-42005. Deposited 15 September 2023.
- 81.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State C Body 1: RELION Multi-body Refinement of KCTD5(CTD)/Gbeta1gamma2. EMD-42006. https://www.ebi.ac.uk/emdb/EMD-42006. Deposited 15 September 2023.
- 82.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State C Body 2: RELION Multi-body Refinement of KCTD5(BTB)/Cullin3(NTD). EMD-42007. https://www.ebi.ac.uk/emdb/EMD-42007. Deposited 15 September 2023.
- 83.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State D: Composite Map from RELION Multi-body Refinement. EMD-42008. https://www.ebi.ac.uk/emdb/EMD-42008. Deposited 15 September 2023.
- 84.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State D: Reference Map for Composite Map EMD-42008. EMD-42009. https://www.ebi.ac.uk/emdb/EMD-42009. Deposited 15 September 2023.
- 85.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State D Body 1: RELION Multi-body Refinement of KCTD5(CTD)/Gbeta1gamma2. EMD-42010. https://www.ebi.ac.uk/emdb/EMD-42010. Deposited 15 September 2023.
- 86.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex State D Body 2: RELION Multi-body Refinemnent of KCTD5(BTB)/Cullin3(NTD). EMD-42011. https://www.ebi.ac.uk/emdb/EMD-42011. Deposited 15 September 2023.
- 87.Kuntz D. A., Nguyen D. M., Narayanan N., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: Local Refinment of KCTD5(CTD)/Gbeta1gamma2. PDB ID: 8U7Z. https://www.rcsb.org/structure/8U7Z. Deposited 15 September 2023.
- 88.Kuntz D. A., Nguyen D. M., Narayanan N., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: Local Refinment of KCTD5(BTB)/Cullin3(NTD). PDB ID: 8U80. https://www.rcsb.org/structure/8U80. Deposited 15 September 2023.
- 89.Kuntz D. A., Nguyen D. M., Narayanan N., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: State A From Composite RELION Multi-body Refinement Map. PDB ID: 8U81. https://www.rcsb.org/structure/8U81. Deposited 15 September 2023.
- 90.Kuntz D. A., Nguyen D. M., Narayanan N., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: State B From Composite RELION Multi-body Refinement Map. PDB ID: 8U82. https://www.rcsb.org/structure/8U82. Deposited 15 September 2023.
- 91.Kuntz D. A., Nguyen D. M., Narayanan N., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: State C From Composite RELION Multi-body Refinement Map. PDB ID: 8U83. https://www.rcsb.org/structure/8U83. Deposited 15 September 2023.
- 92.Kuntz D. A., Nguyen D. M., Narayanan N., Privé G. G., KCTD5/Cullin3/Gbeta1gamma2 Complex: State D From Composite RELION Multi-body Refinement Map. PDB ID: 8U84. https://www.rcsb.org/structure/8U84. Deposited 15 September 2023.
- 93.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., Multi-body cryo-em maps and models of a pentameric KCTD5/Cullin3/Gβγ E3 ubiquitin ligase complex. Zenodo. https://zenodo.org/records/8341597. Deposited 13 September 2023.
- 94.Nguyen D. M., Narayanan N., Kuntz D. A., Privé G. G., Structure and dynamics of a pentameric KCTD5/Cullin3/GBeta1Gamma2 E3 ubiquitin ligase complex. EMPIAR. https://www.ebi.ac.uk/empiar/EMPIAR-11734/. Deposited 20 September 2023.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix 01 (PDF)
Cryo-EM maps of the dynamics in the top and bottom parts of the complex as revealed by 3DFlex analysis.
Dynamics in the CRL3KCTD5/Gβ1γ2 complex, including the extended model that includes full-length CUL3 (green), RBX1 (grey), ARIH1 (white) and ubiquitin (yellow). The C-terminus of ubiquitin is shown as a yellow sphere.
Data Availability Statement
The cryo-EM maps and associated atomic models of top and bottom complex and conformational states A, B, C, and D have been deposited in the Electron Microscopy Data Bank with accession codes EMD-41994 (69), EMD-41995 (70), EMD-41996 (71), EMD-41997 (72), EMD-41998 (73), EMD-41999 (74), EMD-42000 (75), EMD-42001 (76), EMD-42002 (77), EMD-42003 (78), EMD-42004 (79), EMD-42005 (80), EMD-42006 (81), EMD-42007 (82), EMD-42008 (83), EMD-42009 (84), EMD-42010 (85), EMD-42011 (86) and the Protein Data Bank with accession codes 8U7Z (87), 8U80 (88), 8U81 (89), 8U82 (90), 8U83 (91), and 8U84 (92). The 40 maps and models from the multi-body analysis are available from Zenodo at https://zenodo.org/record/8341597 (93). Microscope movies and particle image sets are available from EMPIAR at https://www.ebi.ac.uk/empiar/EMPIAR-11734 (94).