

Abstract
Stomata are pores on plant aerial surfaces, each bordered by a pair of guard cells. They control gas exchange vital for plant survival. Understanding how guard cells respond to environmental signals such as atmospheric carbon dioxide (CO2) levels is not only insightful to fundamental biology but also relevant to real-world issues of crop productivity under global climate change. In the past decade, multiple important signaling elements for stomatal closure induced by elevated CO2 have been identified. Yet, there is no comprehensive understanding of high CO2-induced stomatal closure. In this work, we assemble a cellular signaling network underlying high CO2-induced stomatal closure by integrating evidence from a comprehensive literature analysis. We further construct a Boolean dynamic model of the network, which allows in silico simulation of the stomatal closure response to high CO2 in wild-type Arabidopsis thaliana plants and in cases of pharmacological or genetic manipulation of network nodes. Our model has a 91% accuracy in capturing known experimental observations. We perform network-based logical analysis and reveal a feedback core of the network, which dictates cellular decisions in closure response to high CO2. Based on these analyses, we predict and experimentally confirm that applying nitric oxide (NO) induces stomatal closure in ambient CO2 and causes hypersensitivity to elevated CO2. Moreover, we predict a negative regulatory relationship between NO and the protein phosphatase ABI2 and find experimentally that NO inhibits ABI2 phosphatase activity. The experimental validation of these model predictions demonstrates the effectiveness of network-based modeling and highlights the decision-making role of the feedback core of the network in signal transduction. We further explore the model’s potential in predicting targets of signaling elements not yet connected to the CO2 network. Our combination of network science, in silico model simulation, and experimental assays demonstrates an effective interdisciplinary approach to understanding system-level biology.
Stomata are pores on plant surfaces that can be opened or closed by pairs of guard cells in response to environmental signals. This study analyzes CO2-induced stomatal closure in Arabidopsis thaliana using network-based modelling and experimental validation, uncovering that feedback loops in a core signal transduction network regulate decision-making in guard cells.
Introduction
Stomata are microscopic pores on plant leaf surfaces that control gas exchange such as the uptake of CO2 and the release of water vapor and photosynthetically produced O2. Each stomate is bordered by 2 guard cells, whose shape change in response to the external and internal environment controls the stomate’s opening or closure. Because plants inevitably lose water via transpiration through the stomata, the size of the stomatal pores is tightly regulated to balance the competing needs of CO2 supply for photosynthesis and plant hydration. Stomata open in response to environmental signals such as light and low atmospheric CO2 levels and close in response to darkness, high CO2, drought, and low humidity [1]. As elucidated in detail for the stress hormone abscisic acid (ABA) [2–4], stimulus perception activates an intracellular signal transduction network, which leads to Ca2+ influx and anion efflux, with consequent membrane depolarization promoting K+ efflux. The outflow of ions increases cellular water potential, creating a driving force for water efflux through aquaporins, which finally causes guard cell deflation and stomatal closure. The extent to which the ABA signal transduction network is shared by other closure signals such as darkness and high CO2 has been studied and debated [2,5].
Understanding how stomata respond to high CO2 is a fundamental question for plant biology. Since stomata control gas exchange vital for plant water retention and photosynthesis, stomatal response to abiotic stress is an essential process for plant survival. Furthermore, understanding plant response to high CO2 also has imperative practical significance: over the past 60 years, CO2 concentration in the atmosphere has increased by 32% (NOAA at Mauna Loa [6]). The greenhouse effect induced by increased CO2 has increased global temperature, which exacerbates drought, compounding the stress. Plant stomatal responses to CO2 are therefore highly relevant to real-world issues of crop productivity under the adversity posed by climate change.
Current knowledge of high CO2-induced stomatal closure in guard cells
Stomatal movement in response to high CO2 concentration is a complex process in which numerous cellular elements have been implicated by prior studies. These elements interact with and regulate each other via chemical reactions, protein interactions, and physical processes, forming a complex cell signaling network that propagates the signal from CO2 to stomatal closure. Atmospheric CO2 enters guard cells through CO2 porins [7]. The CO2-binding carbonic anhydrase enzymes CA1, CA4, and CA7 [8–10] catalyze the reversible chemical reaction CO2 + H2O ↔ HCO3– + H+, and these enzymes are a demonstrated component of the CO2 signal transduction mechanism. HT1 (HIGH LEAF TEMPERATURE 1) is an important CO2 signaling element that inhibits the activation of the main anion channel SLAC1 (SLOW ANION CHANNEL-ASSOCIATED 1). Multiple mutants of HT1 have been shown to result in insensitivity to high CO2 [11–13]. Mitogen-activated protein kinases 4 and 12 (MPK4/MPK12) bind to HT1 and inhibit its kinase activity [13,14]. Bicarbonate enhances and stabilizes the binding of MPK4/MPK12 to HT1 [15,16], resulting in loss of HT1 activation of CBC1 kinase, which itself is an inhibitor of closure [15]. Arabidopsis RESISTANT TO HIGH CO2 (RHC1), a MATE-type transporter, may also link elevated CO2 concentration to repression of HT1, although this has been debated [17,18]. Disruption of the second messengers Ca2+cyt, ROS (reactive oxygen species), or NO (nitric oxide) impairs high CO2-induced closure [19–24], implicating them in CO2 signaling. Despite the recent identification of key elements and pathways in the high CO2 response, further elucidation is needed of how the known elements and pathways interact with each other to induce stomatal closure. A systems biology approach integrating existing evidence into a signaling network can identify gaps in knowledge and suggest potential ways to elucidate all the mechanisms that contribute to stomatal closure.
Prior modeling of guard cell signaling networks
In the past, our team has developed multiple networks and network-based models to characterize the guard cell signaling system. In 2006, Li and colleagues constructed a Boolean model of ABA-induced stomatal closure [25]. In 2014, Sun and colleagues developed a multilevel discrete dynamic model focusing on light-induced stomatal opening [26]. In 2017, Albert and colleagues made major updates to the ABA-induced stomatal closure model, including many more signaling elements discovered in recent years [3]. These network-based models revealed mechanistic insights and made novel predictions that were validated by follow-up experiments. Furthermore, we have developed novel theoretical concepts such as the concept of the stable motif, which forms the foundation of computational methods to determine the repertoire of long-time behaviors of biological systems and to identify nodes that can drive the system into a desired state [27–30].
Karanam and colleagues recently developed a graphical user interface to easily analyze Boolean models [31] and applied it to integrate a CO2 signaling pathway into our previous ABA-induced closure model [3], as an approach to describe CO2 signaling. While it is a good demonstration of their methodology, this preliminary model is not a rigorous representation of CO2 signaling in that: (1) it lacks a comprehensive literature review to identify elements that are differentially regulated in high CO2 signaling as compared to ABA signaling; and (2) it assumes that high CO2 is sufficient to directly activate water efflux through the membrane, as well as Ca2+ inflow, which together are sufficient to drive stomatal closure: these assumptions short-circuit all other CO2-related nodes, i.e., render the rest of their network superfluous. Because of the assumed direct connection to water efflux, this model would fail to recapitulate, for example, the experimental observations of impaired high CO2-induced closure for ht1 [12] and mpk4/mpk12 [14] mutants. A more rigorous network construction approach is required to produce a more accurate representation and simulation of high CO2-induced closure.
In this work, we compile and integrate an extensive body of literature evidence to assemble a guard cell signal transduction network that characterizes how high CO2 induces stomatal closure through the decrease in turgor and volume of guard cell pairs. To further capture the dynamics involved in the stomatal closure process, we develop a dynamic model of the signaling network. We rigorously evaluate the accuracy of the model against known experimental observations, obtaining a high (91%) accuracy. We perform analyses including network connectivity, model simulation, and stable motif analysis, which reveal a feedback core that dictates the cellular decisions in the CO2 response, and the roles of key nodes in the process. We then demonstrate the predictive power of our network model by predicting the outcomes of numerous perturbations (e.g., node knockouts) that can potentially influence stomatal response; such predictions are valuable for the prioritization of wet bench experiments. Finally, we experimentally test and validate 3 predictions related to the role of NO in stomatal closure. A flowchart of our work is shown below (Fig 1). Our modeling framework captures system-level biological processes and provides insights into their logic and key underlying mechanisms.
Fig 1. Flow chart of the process of network construction and analysis.

We construct a signaling network based on literature review, resulting in a network of 47 nodes and 95 edges. We then construct a Boolean dynamic model, which adds node states and regulatory logic to the network. We assess the accuracy of our network model by comparing its results with known experimental observations, finding that the model simulations successfully recapitulate 91% of experimental observations. We also perform multiple network-based analyses on the model. Finally, we experimentally validate selected model predictions.
Results
Construction and analysis of the signaling network of high CO2-induced stomatal closure
We conducted a comprehensive literature review, covering more than 160 publications relating to stomatal closure or to elements of guard cell CO2 signaling, and manually curated the interaction information from these publications (S1 Table), aiming to identify and integrate the mechanisms that lead to high CO2-induced rapid stomatal closure. For this purpose, guard cell development and long-term responses based on gene regulation are out of the scope of our model. Further, not all guard cell signaling elements are equally relevant to high CO2-induced closure. For example, cytosolic pH increases in response to ABA, but does not increase under high CO2 [9,32,33]. Another example is the kinase OST1 (Open Stomata1/SnRK2.6), known to phosphorylate SLAC1 as a key contributor to the ion flow that leads to stomatal closure induced by ABA. Although ost1 mutants show strongly impaired stomatal closure under both ABA and high CO2, recent studies showed that unlike ABA, high CO2 does not increase OST1 kinase activity in guard cells [34,35].
To focus on CO2 signaling, we divide the evidence into 5 categories based on relevance: (1) elements and their interactions that are validated to be involved in CO2 signaling; (2) generic interactions that can be assumed to be involved in CO2 signaling, e.g., chemical reactions whose reactants are reasonably expected to be present in the guard cell; (3) evidence that shows an element is not involved in CO2 signaling; (4) evidence that supports an element’s role in stomatal closure in response to another signal, but the element’s involvement in high CO2-induced closure has not been investigated; and (5) evidence that supports an element’s role in high CO2-induced closure but this element has no or insufficient connections to elements in categories (1) and (2). The resulting list of observations and their categorization is provided in S1 Table. We use category (1) and (2) evidence to construct the high CO2 signaling network and exclude category (3) and (4) evidence. Category (5) evidence is also necessarily excluded, as it is not possible to connect such nodes to the network. Such categorization is also helpful to wet bench investigators; e.g., category 4 and 5 elements would be of particular interest to investigate further in the future.
To represent signaling elements with different functions, we deploy a multi-node representation for the OST1, MPK, and SLAC1 proteins, separating the different (e.g., CO2-dependent and independent) mechanisms for their activation (see Fig 2C for an example, and see S2 Table and S1 Text for details). As the follow-up analyses require that each edge of the network points to a node, during network construction we redirected each evidence of influence on an edge to an equivalent influence on the target of the edge [36].
Fig 2.

(A) The high CO2 signaling network with 47 nodes and 95 edges. Yellow nodes represent validated elements in CO2 signaling, i.e., category (1); white nodes represent assumed elements based on generic evidence, i.e., category (2). (B) Acyclic graph representation of the network highlighting the in-component, the strongly connected-component and the out-component, forming a hierarchical structure of signal transduction. (C) Illustration of the multi-node representation of signaling elements that participate in multiple pathways and mechanisms, using MPKs as an example. MPKs are known to inhibit HT1 by a physical interaction induced by bicarbonate and independent of the MPKs’ kinase activity [15,16], and also are necessary in Ca2+-induced SLAC1 activation [37].
Biological description of the high CO2-induced stomatal closure network and its overlap with the ABA-induced closure network
The signaling network corresponding to high CO2-induced closure contains 47 nodes and 95 edges (Fig 2A). A summary table of all edges is presented in Table 1. The full list of nodes and edges, including the full node names, is provided in S2 Table. We mark nodes of the network (Fig 2A) in 3 colors, to reflect their specificity to high CO2-induced closure. Yellow nodes represent validated elements in CO2 signaling, i.e., category (1); white nodes represent assumed elements based on generic evidence, i.e., category (2). For example, the white nodes 8-nitro-cGMP, ADPRc, and cADPR represent metabolites known to be produced in response to NO (a validated CO2 element) [38] and are necessary to connect NO to the rest of the network. Although by default nodes in categories (3) to (5) are not included in the network, “OST1 activated” is a member of a node pair (it represents increased kinase activity of OST1), thus we include it and mark it in purple to indicate that it belongs to category (4): it is not involved in high CO2-induced closure but is important under ABA-induced closure.
Table 1. Edge list of the high CO2-induced closure network.
The edges represent interactions and regulatory effects. The start node (first column) indicates the source of the interaction and the end node (second column) indicates the target of the interaction. “Type” records the sign and type of the edge. Positive edge sign, “+”, indicates positive/activating regulation and “–” indicates negative/inhibitory regulation. For edge type, we use “D” for direct regulation, “I” for indirect regulation, and “A” for assumed regulation. In rare cases, we use “D/A” to represent an edge that is supported by a direct interaction but has assumed properties, e.g., assumed direction or sign. “Ref.” column records the reference that supports the edge. Edges representing known biophysical/biochemical processes, e.g., AnionEM → Depolarization, do not have a reference. The table follows alphabetic order of the start node.
| Start node | End node | Type | Ref. |
|---|---|---|---|
| 8-nitro-cGMP | ADPRc | +, A | [39–41] |
| ABI1 | OST1 minimum function | -, D | [42–44] |
| ABI1 | OST1 activated | -, D | [42–44] |
| ABI1 | SLAC1 generic | -, I | [45–47] |
| ABI1 | SLAC1 CO2 | -, I | [45–47] |
| ABI2 | OST1 minimum function | -, D | [44] |
| ABI2 | OST1 activated | -, D | [44] |
| ABI2 | GHR1 | -, D | [48] |
| ABI2 | SLAC1 generic | -, I | [45–47] |
| ABI2 | SLAC1 CO2 | -, I | [45–47] |
| ADPRc | cADPR | +, D | |
| AnionEM | Depolarization | +, D | |
| AnionEM | H2O Efflux | +, D | |
| Aquaporins | H2O Efflux | +, D | [7,49] |
| Ca2+ ATPase | Ca2+cyt | -, D | |
| CAs | HCO3- | +, D | [8–10] |
| CAs | RHC1 | +, D | [17] |
| Ca2+cyt | MPK activity | +, I | [13,18,37] |
| Ca2+cyt | Ca2+ ATPase | +, D | [25] |
| Ca2+cyt | CPKs | +, D | [46] |
| Ca2+cyt | QUAC1 | +, A | [50] |
| Ca2+cyt | H+ ATPase | -, D | [51] |
| Ca2+cyt | KEV | +, D | [52] |
| Ca2+cyt | Depolarization | +, D | |
| cADPR | CIS | +, I | [53] |
| CaIM | ABI1 | -, I | [54] |
| CaIM | ABI2 | -, I | [54] |
| CaIM | HAB1 | -, I | [54] |
| CaIM | Ca2+cyt | +, D | |
| CBC1/2 | SLAC1 CO2 | - | [55] |
| cGMP | 8-nitro-cGMP | +, D | |
| CIS | Ca2+cyt | +, D | |
| CO2 entry of cell | HCO3- | +, D | [8,9] |
| CO2 porins | CO2 entry of cell | +, D | [7] |
| CPKs | CPKs | +, D/A | [56] |
| CPKs | SLAC1 generic | +, D | [46,47,57] |
| Depolarization | KOUT | +, D | |
| GHR1 | CaIM | +, A | [48] |
| GHR1 | CPKs | +, D | [58] |
| GHR1 | SLAC1 generic | -, D | [48] |
| GHR1 | SLAC1 CO2 | -, D | [48] |
| GTP | cGMP | +, D | [59] |
| H+ ATPase | Depolarization | -, D | |
| H2O Efflux | Closure | +, D | |
| HAB1 | OST1 minimum function | -, D | [44] |
| HAB1 | OST1 activated | -, D | [44] |
| HCO3- | MPKs-HT1 complex | +, D | [13–16,18] |
| HCO3- | RHC1 | +, D | [17] |
| HCO3- | SLAC1 CO2 | +, A | [60] |
| High CO2 | CO2 entry of cell | +, D | |
| HT1 | CBC1/2 | +, D | [15] |
| HT1 | GHR1 | -, D | [13,17] |
| HT1 | OST1 minimum function | -, D | [13,17] |
| HT1 | OST1 activated | -, D | [13,17] |
| K+ efflux | Depolarization | -, D | |
| K+ efflux | H2O Efflux | +, D | |
| K+ efflux | K+ efflux | +, A | |
| KEV | Depolarization | +, D | |
| KEV | K+ efflux | +, D | |
| KOUT | K+ efflux | +, D | |
| MPKs | MPKs-HT1 complex | +, D | |
| MPKs | MPK activity | +, D | |
| MPKs-HT1 complex | HT1 | -, D | [13–16] |
| MPK activity | HT1 | -, A | [13,18,37] |
| MPK activity | MPK activity | +, D/A | [61] |
| MPK activity | SLAC1 generic | +, A | [37] |
| NAD+ | cADPR | +, D | |
| NADPH | NO | +, D | [62] |
| NADPH | ROS | +, D | |
| NIA1/2 | NO | +, D | [62] |
| Nitrite | NO | +, D | [62] |
| NO | 8-nitro-cGMP | +, I | [38] |
| NO | NOGC | +, D | [59] |
| NO | KOUT | -, I | [63] |
| NOGC | cGMP | +, D | [59] |
| OST1 activated | SLAC1 generic | +, D | [45] |
| OST1 minimum function | OST1 activated | +, D | |
| OST1 minimum function | RbohD/F | +, D | [64] |
| OST1 minimum function | SLAC1 generic | +, D | [45,65] |
| OST1 minimum function | SLAC1 CO2 | +, D | [45,65] |
| OST1 minimum function | QUAC1 | +, D | [66] |
| OST1 minimum function | Aquaporins | +, D | [49] |
| QUAC1 | AnionEM | +, D | [67] |
| RbohD/F | ROS | +, D | |
| RHC1 | HT1 | -, D | [17] |
| ROS | GHR1 | +, I | [48] |
| ROS | ABI1 | -, D | [68] |
| ROS | ABI2 | -, D | [69] |
| ROS | HAB1 | -, D | [70] |
| ROS | 8-nitro-cGMP | +, I | [38] |
| ROS | NIA1/2 | +, A | [71] |
| ROS | H+ ATPase | -, D | [72] |
| ROS | KOUT | -, I | [73] |
| SLAC1 CO2 | AnionEM | +, D | |
| SLAC1 nonCO2 | AnionEM | +, D |
The edges in the network are also marked to distinguish their properties: solid edges indicate a direct interaction/regulation, dashed edges indicate indirect regulation, and dotted edges indicate assumed regulation. An example of indirect regulation is the edge from ROS to the H+ ATPase, which is dashed. An example of an edge referring to assumed regulation is the edge from HCO3- to SLAC1 CO2, which is supported by a specific SLAC1 residue that is required for full CO2 signaling and is structurally predicted to interact with HCO3- [60], but this predicted physical interaction has not been experimentally verified. Like this example, all assumed edges in the model are well-supported. Further justification of the assumed edges can be found in S1 Text.
The high CO2-induced stomatal closure network is known to share nodes and pathways with the ABA-induced closure network, but it also has its unique signaling elements. Furthermore, through the literature review process, we also found multiple elements that participate in ABA-induced closure but are not yet evaluated in the context of high CO2-induced closure (also see S1 Table and S2 Text). We list in Table 2 the different categories for the combined set of the signaling elements (i.e., nodes) of this CO2 network and of our 2017 ABA network [3].
Table 2. Signaling elements that differ in the networks that lead to closure in response to ABA versus high CO2.
| Category | Nodes/signaling elements |
|---|---|
| CO2-specific signaling elements that are not involved in ABA-induced closure | CAs, HCO3-, RHC1, HT1, MPKs-HT1 inhibition |
| ABA signaling elements observed to be absent or not significant in high CO2-induced closure | Cytosolic pH increase, OST1 activation |
| ABA signaling elements* whose participation in high CO2 signaling has not been tested | AtRAC1, actin reorganization, GPA1, Malate, Microtubule depolymerization, PEPC, PP2CA, ROP11, S1P/PhytoS1P, SPHK1/2, SLAH3, TCTP; Phospholipases and molecules involved in reactions catalyzed by them: PtdIns(4,5)P2, PLC, DAG, InsP3, InsP6, PLDα, PLDδ, PA; Elements of the pathway leading to vacuolar acidification: PI3P5K, PtdIns(3,5)P2, V-PPase, V-ATPase, vacuolar acidification; |
| CO2 signaling elements with insufficient regulatory information to integrate into our CO2 network** | TPK1, KIN7, BIG1, AtABCB14, GCA2, RCARs |
The CO2 signaling network consists of 3 distinct modules
Of the 47 nodes in the network, 8 (including high CO2) are source nodes that are not regulated by any other node, and 1 node (Closure) is a sink node (output). The source nodes other than the signal “high CO2” mainly represent molecules needed as substrates for reactions incorporated in the network: CO2 porins, CAs, GTP, MPKs, NAD+, NADPH, Nitrite. Structural and connectivity analysis shows that the CO2 network is organized into 3 graph modules (called “connected components” in graph theoretical language), as shown in Fig 2B. First, 7 early CO2 signaling nodes form an in-component, i.e., these nodes are not regulated by any nodes from other components in the network. This module mainly represents how CO2 enters the guard cell and is converted to HCO3-. Second, a large strongly connected component (SCC) of 20 nodes forms the core of the signal transduction network. “Strongly connected” means that for any node pair A, B, there is at least 1 regulatory path from A to B and at least 1 path from B to A. The feedback loops underlying the strong connectivity allows complex dynamical behavior that ultimately dictates the homeostasis of the system. The 2 edges between the in-component and the SCC represent CO2 sensing mechanisms, e.g., the inhibitory effect of HCO3- on HT1 via the formation of the MPKs-HT1 complex or via RHC1. Third, 13 nodes, mainly representing ion transport mechanisms and the ion and water fluxes they mediate, as well as the output node “Closure,” form the out-component that ultimately implements stomatal closure. These nodes are mainly regulated by nodes of the SCC and do not regulate nodes in other connected components. These 3 modules determine a higher-level overall linear structure of signal propagation from the in-component (which includes the input CO2 signal) to the feedback-rich SCC, and finally to the out-component and closure (Fig 2B). The HCO3- activation of SLAC1 described in [60] forms a unique edge that connects the CO2 in-component directly to the out-component.
Construction of the dynamic model
A network of signaling elements reveals the interactions and regulatory relationships between them, yet this alone is not sufficient to capture the dynamic aspects of signal transduction. In the stomatal closure process, the biological system represented by the network evolves from a state corresponding to open stomata (in the absence of a closure signal) to a state corresponding to closed stomata after sufficient time has elapsed in the presence of a closure signal. To capture the dynamic state transition of the CO2 network, we perform dynamic modeling, where we describe each node of the network with a state variable that represents its abundance or activity. In this way, the propagation of the high CO2 signal is captured in the model by a series of node state changes propagating through the network, eventually leading to stomatal closure. We use a Boolean model, i.e., we describe each node with binary states: ON or 1 for higher-than-threshold abundance or activity; OFF or 0 for lower-than-threshold abundance and activity. Despite its simplicity, Boolean dynamics has proved capable of capturing complex biological behaviors [74], e.g., cell differentiation as a consequence of multi-stability [75,76], or sustained oscillations such as the cell cycle [77].
For each node, its state change is determined by the regulatory influences incident on it, which are captured by its Boolean regulatory function. The regulatory function uses the states of the node’s regulators in the network as inputs and combines these inputs using logical operators such that the function correctly reflects experimental observations. For example, the node “HCO3-” represents bicarbonate concentration in the guard cell. Its input nodes are “CO2 entry of cell” and “CAs” (the CO2-binding carbonic anhydrase enzymes CA1, CA4, and CA7). Since bicarbonate is produced from the reaction CO2 + H2O → HCO3– + H+, catalyzed by CAs, we know that both CO2 entry into the cell and CAs are required for the increase of intracellular bicarbonate concentration, thus the inputs are connected with a logical “AND” operator, and the regulatory function of node “HCO3-” is set as “ = CAs AND CO2 entry of cell.” The comprehensive list of regulatory functions and their justifications are provided in S1 Text. Each node’s regulatory function determines the future state of the node as determined by the current state of its regulators.
With the Boolean regulatory functions defined for each node, we can perform simulations of the signal transduction process over the entire CO2 network. We initialize the in silico stomata in a pre-stimulus, i.e., open state, with an ON state for closure-inhibiting nodes (namely ABI1, ABI2, CBC1/2, HAB1, HT1, and H+ ATPase) and other nodes OFF (see S1 Text for the complete initial condition). We deploy a discrete-time simulation, with stochastic random order asynchronous update, and simulate the network model for 30 timesteps (see Methods). Each simulated trajectory of the model will eventually converge into a stabilized behavior of the system as a whole, called an attractor. The most intuitive attractor is a steady state, where all nodes stabilize in a fixed state. Alternatively, a system can also have an oscillatory attractor, where a fraction of nodes indefinitely oscillates between 0 (OFF) and 1 (ON). A system can have multiple attractors (called multi-stability), which may be reachable from the same initial state if the system’s timing is stochastic. We obtain the consensus behavior of the system by performing 1,000 replicate simulations, i.e., 1,000 “in silico stomata” simulations starting in the same pre-stimulus state (see Methods), then calculating the percentage of node activation at each timestep as our metric, meaning the percentage of simulations in which the node (e.g., Closure) is in state 1 (ON). In this way, oscillations and multi-stability with different node states will display a percentage of node activation with a value between 0% and 100%. We demonstrate simulated high CO2-induced stomatal closure and lack of closure under ambient CO2 in Fig 3B and show a summary of node states in the corresponding attractors in Fig 3C.
Fig 3.

(A) Example demonstrating how a Boolean regulatory function is constructed from observations. (B) A demonstration of model simulation, capturing stomatal closure under high CO2, and lack of closure under ambient CO2. (C) Summary of node states in the attractor with closure ON under high CO2, and the attractor with closure OFF under ambient CO2, found from the model simulations. A green box represents the ON state, gray represents the OFF state, and X represents oscillation. If a prior experiment compared the states of a node under high and ambient CO2, we mark the corresponding cells with “e.” In all such cases, the model recapitulates the experimental result.
In the simulated wild-type system (Fig 3B), the percentage of closure in response to high CO2 increases over the timesteps from 0% to 100%. Despite the deliberate choice of stochasticity in the timing, all the simulations converge into the same attractor, in which the node Closure is ON. The row marked “high CO2” in Fig 3C shows the states of all nodes in this attractor, recapitulating the biological knowledge regarding the activity of nodes in stomata that closed in response to high CO2. We marked with “e” the node states that have been studied experimentally (see the entries in S1 Table in which Entity/Process A is High CO2 and which report measurements of Entity/Process B). For example, the node “OST1 activated” is OFF in this closure attractor, meaning that OST1 does not become activated under high CO2, consistent with recent publications [34,35]. The node Ca2+cyt oscillates between ON and OFF, capturing transient cytosolic Ca2+ concentration increases that have been observed during CO2 response [19,20]. Node states for which no prior experiments exist represent novel predictions. For example, the model indicates that high CO2 activates not only the CO2-specific “SLAC1 CO2” node but also the node “SLAC1 generic,” as all of these node’s regulators, including CPKs, are indirectly activated by high CO2. In the simulation curve corresponding to ambient CO2 (represented as the inactivity of the node “high CO2”), the percentage of closure remains 0% for the entire duration of the simulation (Fig 3B), consistent with the expectation that stomata are open in the absence of CO2 elevation. The row marked “ambient CO2” in Fig 3C shows the states of all nodes in the attractor under ambient CO2.
Validation of the network model
We assembled all experimental evidence related to high CO2-induced stomatal closure that was not used in model construction (see the entries with “V” for validation, in S1 Table), and checked if the model simulations of the relevant settings yield results consistent with the experimental evidence. An important concept that can be insufficiently appreciated by experimentalists is that all the pathway-level effects (e.g., the effect of a node manipulation on high CO2-induced stomatal closure) are emergent properties of the system that arise from multiple interactions; thus, the fact that the regulatory functions of each node are informed by experimental evidence is not sufficient to guarantee consistency with pathway-level observations.
The summary of validation and consistency is presented in Table 3. In a total of 47 settings, there are 43 cases of consistency between the experimental and simulation results and 4 inconsistent cases, yielding an accuracy of 43/47 = 91%. The high validation accuracy shows that our model correctly captures most known signaling behaviors in CO2-induced stomatal closure. Note that a few comparisons could be consistent by construction if the observed regulator node directly regulates the target node in the model. When assessing the validity of our model, we exclude this type of trivial consistency. A more detailed validation table including the trivially consistent cases, a description of each experimental observation, and explanations are provided in S3 Table.
Table 3. Comprehensive validation comparing simulations against known experimental observations.
In the 47 total simulations in the table, 43 are consistent and, 4 are inconsistent (marked with bold, underlined font), with accuracy 43/47 = 91%.
| Row # | Signal | Simulation setting | Experimental outcome | Ref | Simulated percentage of node |
|---|---|---|---|---|---|
| Consistent cases | |||||
| 1. | High CO2 | MPKs = OFF | No closure | [14,18] | Closure = 0% |
| 2. | High CO2 | MPK activity = OFF | Closure | [15,16] | Closure = 100% |
| 3. | High CO2 | MPK activity = ON | Closure | [16] | Closure = 100% |
| 4. | High CO2 | MPK activity = OFF, MPKs-HT1complex = OFF |
No or reduced closure | [16] | Closure = 0% |
| 5. | High CO2 | HT1 = OFF | Closure | [11,13] | Closure = 100% |
| 6. | High CO2 | HT1 = ON | No closure | [12,13] | Closure = 0% |
| 7. | High CO2 | RHC1 = OFF, HT1 = OFF | SLAC1 CO2 activation | [17] | SLAC1 CO2 = 100% |
| 8. | High CO2 | HT1 = OFF, OST1 minimum function = OFF | No or reduced Closure | [17] | Closure = 0% |
| 9. | High CO2 | CBC1/2 OFF | Closure | [15] | Closure = 100% |
| 10. | High CO2 | CAs = OFF | Reduced closure | [8] | Closure = 0% |
| 11. | High CO2 | CAs = OFF, HT1 = OFF | Closure | [8] | Closure = 100% |
| 12. | High CO2 | GHR1 = OFF | Reduced closure | [13,58] | Closure = 41.5% |
| 13. | High CO2 | HCO3- = OFF, high CO2 = ON | SLAC1 OFF | [9] | SLAC1 generic = 0%, SLAC1 CO2 = 0% |
| 14. | High CO2 | Ca2+cyt = OFF | No closure | [19–21] | Closure = 0% |
| 15. | High CO2 | ABI1 = ON | Reduced closure | [5,78] | Closure = 44.0% |
| 16. | High CO2 | ABI2 = ON | No closure | [5,78] | Closure = 39.9% |
| 17. | High CO2 | Default (high CO2 = ON) | Ca2+cyt > 0 | [19] | Ca2+cyt = 53.6% (oscillating) |
| 18. | High CO2 | ROS = OFF | Reduced closure | [22] | Closure = 0% |
| 19. | High CO2 | RbohD/F = OFF | Reduced closure | [22,79] | Closure = 0% |
| 20. | High CO2 | OST1 minimum function = OFF | No closure | [9] | Closure = 0% |
| 21. | High CO2 | Aquaporins = OFF | No closure | [7] | Closure = 0% |
| 22. | High CO2 | CO2 porins = OFF | No closure | [7] | Closure = 0% |
| 23. | High CO2 | Default (high CO2 = ON) | Anion efflux ON | [32] | AnionEM = 100% |
| 24. | High CO2 | Default (high CO2 = ON) | K+ efflux is ON | [32] | K+ efflux = 100% |
| 25. | High CO2 | QUAC1 = OFF | Slower closure | [67] | Closure reaches 100% more slowly than WT |
| 26. | High CO2 | CPKs = OFF (CPK 4,6,21,23) | Closure | [80] | Closure = 100% |
| 27. | High CO2 | Default (high CO2 = ON) | ROS is produced | [22,23] | ROS = 100% |
| 28. | High CO2 | GHR1 = OFF, HT1 = OFF | No or reduced Closure | [13] | Closure = 43.9% |
| 29. | High CO2 | SLAC1 CO2 = OFF, SLAC1 generic = OFF | No closure | [81,82] | Closure = 43.9% |
| 30. | High CO2 | SLAC1 CO2 = OFF, SLAC1 generic = OFF, QUAC1 = OFF | No closure | [82] | Closure = 0% |
| 31. | High CO2 | Default (high CO2 = ON) | H+-ATPase deactivation | [83] | H+-ATPase = 0% |
| 32. | Ca2+ | MPKs = OFF, high CO2 = OFF, CaIM = ON | Reduced AnionEM | [37] | AnionEM = 49.2% |
| 33. | Ca2+ | RHC1 = OFF, CaIM = ON, high CO2 = OFF | Closure | [17] | Closure = 100% |
| 34. | Ca2+ | CaIM = ON, high CO2 = OFF | Closure | [21] | Closure = 100% |
| 35. | Ca2+ | CPKs = OFF, CaIM = ON, high CO2 = OFF | Reduced closure | [21] | Closure = 41.1% |
| 36. | ROS | GHR1 = OFF, ROS = ON, high CO2 = OFF | No closure | [48] | Closure = 41.2% |
| 37. | ROS | ROS = ON, high CO2 = OFF | Closure | [62,79] | Closure = 100% |
| 38. | ROS | ROS = ON, RbohD/F = OFF | Closure | [79] | Closure = 100% |
| 39. | NO | NO = ON, high CO2 = OFF | Closure | [62,84] | Closure = 100% |
| 40. | 8-nitro-cGMP | 8-nitro-cGMP = ON, high CO2 = OFF | Closure | [38] | Closure = 100% |
| 41. | cADPR | cADPR = ON, high CO2 = OFF | Closure | [85] | Closure = 100% |
| 42. | HCO3 | HCO3- = ON, high CO2 = OFF | SLAC1 activation | [8] | SLAC1 CO2 = 100% |
| 43. | HCO3 | MPKs = OFF, high CO2 = OFF, HCO3- = ON | Impaired SLAC1 | [13,14,18] | SLAC1 generic = 0%, SLAC1 CO2 = 0% |
| Inconsistent cases | |||||
| 44. | High CO2 | CBC1/2 = OFF, HT1 = ON | Closure | [15] | Closure = 0% |
| 42. | High CO2 | NO = OFF | Reduced closure | [24] | Closure = 100% |
| 46. | High CO2 | R256 residue mutation interpreted as SLAC1 CO2 = OFF | Reduced closure | [60] | Closure reaches 100% more slowly than WT |
| 47. | Ca2+ | GHR1 = OFF, CaIM = ON, high CO2 = OFF | Closure | [48] | Closure = 40.3% |
Network-based analysis identifies the dynamic repertoire and decision-making mechanisms of the CO2 signaling system
In the previous section, we showed that our model can accurately reproduce existing experimental observations. Next, we explore the explanatory power of the model and its predictions about the biological system’s response in conditions for which experimental observations are not yet available.
We start by determining the full attractor repertoire of the model. The attractor repertoire of the model is important because attractors correspond to biological phenotypes, e.g., open or closed stomata are represented by 2 different attractors in the CO2 network model. Finding the attractor repertoire is the first step toward understanding how a biological system makes decisions on how to switch between different phenotypes. Understanding the decision-making behind attractor transitions of a biological system can reveal the key signal transduction mechanisms and suggest potential interventions to guide the system into a more desirable state. Finding attractors by simulations is difficult because the state space of a network model increases exponentially with its size, e.g., the CO2 network model with 47 nodes has a total of 247 states. An alternative and efficient way to find the attractors of a system is to utilize the network topology via stable motif analysis, developed by the Albert group [86]. Stable motifs are special feedback loops in a dynamic model that can self-sustain once their constituent nodes attain an associated state (see Methods). The important feature of stable motifs is that they are irreversible: after locking in they become independent of the rest of the system. This feature implies that the lock-in of a stable motif is like a decision of the system, and a series of stable motif lock-ins will lead to an attractor [28,30,86]. Therefore, finding all stable motifs and their lock-in sequence allows comprehensive identification of attractors. Stable motif-based attractor identification was shown to be more efficient [29] than methods based on mapping state transitions [87–89], algebraic methods [90], and methods based on network decomposition [91]. In addition, stable motif-based control outperforms other approaches in identifying interventions that can drive the system into a beneficial attractor [29].
Stable motifs identify the dynamic repertoire and decision-making of the system
Whether in ambient or high CO2, the unperturbed (wild type) system has a stable motif (see Fig 4A) that includes the activity of RbohD/F and ROS, the minimum functional level of OST1, and the inactivity of the 3 PP2C protein phosphatases (ABI1, ABI2, and HAB1). These elements form a positive feedback loop in which the enzyme activity of RBOH leads to sufficient ROS abundance, which inactivates all 3 PP2Cs, the inactivity of which is sufficient for activating the minimum functional level of OST1, which in turn activates RBOH. This positive feedback loop is self-sustaining, meaning that the corresponding active or inactive states can be maintained once established. We will refer to this stable motif as the Main Stable Motif (MSM). The lock-in of the MSM is observed in the closure ON attractor (the first row of Fig 3C). The opposite states of the nodes involved in the MSM, namely the inactivity of RbohD/F, ROS, the lack of OST1 function, and the activation of one of the PP2C protein phosphatases, when coupled with the inactivity of CaIM and GHR1, form 3 instances of a weaker type of stable motif, called a conditionally stable motif [92] (see Methods). Fig 4B indicates the superposition of the 3 conditionally stable motifs and shows that they can lock-in if HT1 is ON, which in the wild-type system only happens under ambient CO2. The lock-in of these conditionally stable motifs leads to the attractor with closure OFF (second row in Fig 3C), which is consistent with the CO2-independent high stomatal conductance of the ht1-3D and ht1-8D dominant mutants [12,13]. Importantly, the MSM and the 3 conditionally stable motifs are mutually exclusive. Since the lock-in of a stable motif is irreversible, the choice between them constitutes the decision-making process of the CO2 signaling network (Fig 4C). The system has 1 additional stable motif and 3 additional conditionally stable motifs that contribute in specific circumstances; these are described in section II of S2 Text.
Fig 4. Stable motifs of the network model.

(A) The MSM that leads to closure in the dynamic model. (B) Superposition of the 3 overlapping conditionally stable motifs that lead to attractors that represent open in silico stomata. Each conditionally stable motif contains a different PP2C protein family member, indicated by dashed, dash-dotted or dotted outline and edges, respectively. In the stable motifs, nodes whose name is preceded by ~ are inactive in the corresponding motif. The black circles indicate that the regulatory effects incident on a node form an AND gate; for example, ABI1 will activate only if ROS and CaIM are simultaneously inactive. The red edges starting from HT1 indicate that HT1 = ON is the condition necessary for the conditionally stable motifs. (C) Decision-making diagram of the CO2 signaling network. (D) Summary of the node states in the attractors of the system. Entries that contain a white box with * summarize multiple attractors in which the marked nodes take either on or off states. MSM, Main Stable Motif; ROS, reactive oxygen species.
Identifying the stable motifs allows comprehensive identification of all attractors of the system, using the algorithm we developed in [29]. We first confirm the unique Closure ON attractor in the presence of high CO2 (Fig 4D), where high CO2 leads to the locking-in of the MSM and precludes the activation of the 3 conditionally stable motifs. The MSM leads to the activation of anion flow through the SLAC1 channel and to K+ efflux, as well as to the activation of Aquaporins, which together satisfy all requirements to activate the node Closure. In the case of ambient CO2, stable motif analysis identifies 3 attractors. Two of them have the node Closure OFF, corresponding to the absence of stomatal closure observed under ambient CO2. These 2 attractors differ only in the state of CPKs (either ON or OFF). Interestingly, the third attractor under ambient CO2 has the node Closure ON and is highly similar to the closure ON attractor under high CO2—they differ only in the CO2 in-component and SLAC1 CO2 (see Fig 4D, row 2 versus row 1). Trajectories starting from the pre-stimulus initial state used in our simulation cannot reach this attractor under ambient CO2, consistent with biological reality and with the attractor found by simulation (Fig 3C). Yet, a different initial state, e.g., one in which HT1 is inactive, can lead to this attractor (see S1 Text). An intervention (e.g., sustained activation of a node) can also induce stomatal closure under ambient CO2, as we will present later.
Logic diagrams elucidate signal transduction mechanisms and the roles of driver nodes
To better understand how the system reaches its attractors (e.g., the closure ON attractor under high CO2), we introduce logic diagrams illustrating the signaling mechanisms (Fig 5). A logic diagram (see Methods) is an integration of the network and node states that yield a specific outcome (e.g., the ON state of the node Closure), constructed based on the framework of the “expanded network” developed by the Albert group [86,93,94]. Edges in the logic diagrams are logical implications (i.e., sufficient activations), and black dots represent AND gates. To make the logic diagrams easier to parse, we also applied network reduction techniques that do not change the dynamic behavior of the system [95], and we merged nodes that behave similarly or as a group (e.g., PP2Cs, see Methods).
Fig 5. Logic diagrams show the mechanisms that lead to various stomatal behaviors.

(A) Logic diagram of high CO2-induced stomatal closure. It also indicates that a closure attractor can be reached under ambient CO2 if the MSM is activated by an internal or external driver. Three such drivers (out of 17) are indicated by bold outlines. (B) Diagram of the mechanism for the lack of closure due to the activation of the conditionally stable motifs, e.g., under ambient CO2 or for ost1 knockout. (C) Logic diagram of the predicted mechanism of unstable closure that would arise with inactivity of the SLAC1 channels. Green nodes indicate the active state and gray means the inactive state; white nodes oscillate. MSM, Main Stable Motif.
As shown in Fig 5A, high CO2 activates the MSM (shown with bold font), which leads to ion and water efflux, and thus to an attractor with the node Closure ON. Fig 5B shows the mechanism for the lack of stomatal closure, e.g., under ambient CO2. When HT1 is ON, the conditionally stable motifs (bold-font nodes, with their dependence on HT1 = ON shown with red dotted edges) can lock in and cause the inactivity of the SLAC1 and QUAC1 channels, thus reaching a Closure OFF attractor. The knockout of OST1 also activates the conditionally stable motifs and leads to a similar Closure OFF attractor (see the third entry of Fig 4D). In addition to attractors with constant closure values (constant ON or OFF), the system also allows attractors with an unstable closure value that oscillates between ON and OFF indefinitely. Fig 5C shows the logic diagram of the predicted mechanism of unstable Closure state that could arise from the sustained inactivity of the SLAC1 channels (due, e.g., to knockout of GHR1, see the fourth entry of Fig 4D). The oscillation of Ca2+cyt leads to oscillating QUAC1 and K+ efflux, which in turn lead to an oscillating Closure state. Had SLAC1 been active, it would have sustained anion efflux, leading to sustained Closure ON.
In addition to finding attractors, stable motif analysis can also pinpoint key nodes called drivers, which determine the decisions in the system: once a driver node is sustained in a certain state, it can lock in a corresponding stable motif, forcing a decision in the system. Thus, external control of driver nodes can drive the system from one attractor to another [27,29]. For example, ROS (marked in bold outline) is an internal driver of the MSM, as it inhibits all 3 PP2C proteins, which then allows OST1 activity, which in turn activates RbohD/F, closing the cycle of self-sustained activity. Furthermore, CaIM (Ca2+ influx through the membrane) is an external driver of the MSM, as it inhibits the PP2Cs. When ROS or Ca2+ is externally provided, it will lock in the MSM, thus lead to a closure ON attractor, even in ambient CO2 (see Fig 4D). This explains the experimentally observed ROS or Ca2+-induced stomatal closure under ambient CO2 [21,79,96]. Moreover, ~HT1 (the OFF state of HT1) is also an external driver of the MSM, and leads to the same closure ON attractor as providing ROS or Ca2+ (see Fig 4D), which explains the low stomatal conductance observed in the ht1-2 kinase dead mutant regardless of the CO2 concentration [11].
Our model predicts how the perturbation of signaling elements affects high CO2 response
A model is most helpful in predicting unknown behaviors of a system. We first demonstrate the predictive power of our model by simulating the guard cell response to high CO2 under systematic single node perturbations. These simulations reflect how mutations or interventions could affect high CO2-induced closure. We simulate all single interventions that abolish a node’s expression or activity as maintaining the node in the state 0 (OFF) and denote it “KO”; we also simulate all single interventions that make a node constitutively active by maintaining the node in the state 1 (ON) and denote it “CA.” To quantify the stomatal closure response, we develop an improved approach from [3] combining 3 metrics: the final closure state, the cumulative percentage of closure (CPC), as well as the attractor repertoire of the system (see Methods). The CPC value represents the fraction of simulations in which Closure = 1, summed over 30 timesteps, thus it ranges from 0 to 30. As before [3,54], we observe and categorize 4 typical behaviors different than the wild type (WT) response: (1) we call the behavior in which the percentage of closure increases faster than in the WT system “hypersensitivity”; and (2) the behavior in which the percentage of closure increases more slowly than in the WT system “hyposensitivity”; (3) we refer to the behavior in which the percentage of closure converges to or oscillates around an intermediate value “reduced closure”; (4) and finally the category “no closure” is the lack of closure in simulated stomata. We present the summarized high CO2 closure response categories under systematic perturbations in Fig 6A. The complete perturbation results can be found in S4 Table. We illustrate simulated response curves of these categories in Fig 6B: hypersensitivity under external Ca2+ (CaIM CA), hyposensitivity under CaIM KO, reduced closure under H+-ATPase KO, and no closure under MPKs KO (mpk4/mpk12 mutant). These perturbation response categories offer a way to characterize each signaling element’s contribution to high CO2-induced closure, from weakest (nodes whose KO and CA are in the “Close to WT” and whose CA is in the “Hypersensitivity” category, such as ROS).
Fig 6. Systematic perturbation under high CO2 with illustrations.

(A) Response categorization for systematic perturbations of all nodes, both knockout (KO) and constitutive activation (CA) perturbations. (B) Simulation curves under high CO2 that exemplify response types: normal WT closure, hypersensitivity under external Ca2+ (Ca2+ influx across the membrane (CaIM) CA), hyposensitivity under CaIM KO, reduced CO2 sensitivity for H+-ATPase, or insensitivity to CO2 under MPKs KO. (C) Model simulation predicts 18 interventions that could induce stomatal closure under ambient CO2. Some of these interventions have been validated to induce closure under ambient CO2, and some interventions have supporting evidence in previous publications. (D) Logic diagram of the mechanism of closure in ambient CO2. The node “MSM” is a merged representation of the MSM. Bold outlines indicate 2 of the 18 interventions that can simultaneously activate the MSM and inactivate HT1 under ambient CO2. For example, NO leads to oscillations in Ca2+cyt, which lead to MPK activity, which inactivates HT1, which then activates the MSM and leads to closure. MSM, Main Stable Motif; NO, nitric oxide.
The 5 response categories are well-explained by our stable motif and network analysis
We find that in both hyper- and hypo-sensitivity cases, the final attractor of the system is almost the same as that of the wild-type system under high CO2; the final attractor only differs in the state of the perturbed node and up to a few other nodes regulated by it. As illustrated in the logic diagrams (Fig 5) earlier, the typical reason for hyper-sensitivity (i.e., faster closure) is the immediate accomplishment of a node state that would naturally happen only after a previous event; e.g., CaIM normally would only happen after GHR1 activation (Fig 5A), but it happens immediately with CaIM CA. The mechanism for slower closure is the elimination of one of 2 redundant signaling pathways: for example, CaIM KO will eliminate one pathway to increase the cytosolic Ca2+ level (a necessary condition of closure); the alternative pathway to activate Ca2+cyt is by CIS (Ca2+ influx from stores). The model prediction that perturbation of specific anion channels, e.g., QUAC1 KO or R256 residue KO (interpreted as SLAC1 CO2 = OFF) leads to hyposensitivity is due to the same type of mechanism. Reduced closure occurs when the state of the node Closure oscillates in all attractors, such as for GHR1 KO (see row 6 in Figs 4D and 5C). GHR1 KO leads to the deactivation of the SLAC1 anion channel, leaving QUAC1 as the only conduit of anion flow, and the activity of QUAC1 follows the Ca2+cyt oscillation, resulting in an attractor with oscillatory closure. The final category of behavior, “no closure,” is the lack of closure in all simulated stomata; the system converges into an attractor (or mix of attractors) with closure OFF. The mechanism that leads to this outcome is disruption of ion or water efflux, either through a mutation in a transporter (e.g., Aquaporins KO in Fig 4D), or indirectly through activation of the non-closure conditionally stable motifs (e.g., OST1 KO in Fig 4D; the mechanism is illustrated in Fig 5B).
Our model predicts interventions that yield closure even under ambient CO2
We performed systematic perturbation simulations of the model under ambient CO2 (i.e., with the node “high CO2” set to OFF) and found 18 interventions that can result in Closure = ON in the simulations (Fig 6C; comprehensive results are in S4 Table). Our stable motif analysis identifies that the reason for the effectiveness of these interventions is the activation of the MSM coupled with the inactivation of HT1 (see Fig 5A). HT1 KO itself is an example of such a stable-motif-activating intervention; this result agrees with the closed stomata or low stomatal conductance observed in the ht1-2 mutant [11,13]. Thus, the model predicts that constitutive activity of the nodes that lead (directly or indirectly) to the inhibition of HT1 can also drive the stable motifs and lead the system to closure in ambient CO2. Indeed, all the interventions listed in Fig 6C have this property. Fig 4D illustrates the Closure ON attractor that results from providing NO under ambient CO2 and Fig 6D illustrates the mechanism through which providing NO can lead to closure. In agreement with the model predictions, it was observed experimentally that provision of ROS [79] or CaIM (Ca2+ influx through the membrane, which is equivalent to providing external Ca2+) [21] results in closure under ambient CO2. The predicted closure-inducing effect of additional interventions is supported by experimental evidence, e.g., NO [62,84], cADPR [85], 8-nitro-cGMP [38] were shown to induce closure as a signaling element of the ABA-induced closure network.
Experimental validation of model predictions
Our model predicts that the activation of the MSM is an important part of the process of stomatal closure both in ambient and high CO2 (see Figs 5A and 6D). A follow-up conclusion from this prediction is that earlier activation of the MSM would lead to more rapid closure. One of the nodes whose constitutive activity is predicted to activate the MSM and lead to closure in ambient CO2 is NO. In our model, applying NO under high CO2 activates the MSM sooner. Hence, supplying NO is predicted to cause hypersensitivity of closure under high CO2 (i.e., faster high CO2-induced closure). To assess the validity of this prediction, we experimentally tested the effect of NO on high CO2 response by applying SNP (sodium nitroprusside dihydrate), an NO donor (Fig 7A, see Methodss for experimental assay description). To observe any acceleration of the closure process, we used short treatments (10 and 20 min). We found that under high (800 ppm) CO2, the stomatal aperture under SNP treatment (gray bars) is significantly lower than that without SNP treatment (blue bars, p = 2.8e-2 for 10 min and p = 3.4e-2 for 20 min, Student’s t test). Furthermore, the mean aperture corresponding to the 10 min combined treatment of 0.15 mM SNP and high CO2 is lower than the mean aperture corresponding to the 20 min high CO2 treatment (rightmost blue bar, p = 4.0e-2, Student’s t test), indicating that SNP treatment speeds up the high CO2-induced closure process, consistent with our model prediction. As further confirmation, we performed a similar experiment using the alternative NO donor SNAP (S-nitroso-N-acetylpenicillamine) and found similar results (see details in S2 Text section III).
Fig 7. Experimental validation of the predicted closure responses when applying NO (via the NO donor sodium nitroprusside dihydrate, SNP) in ambient or high CO2.
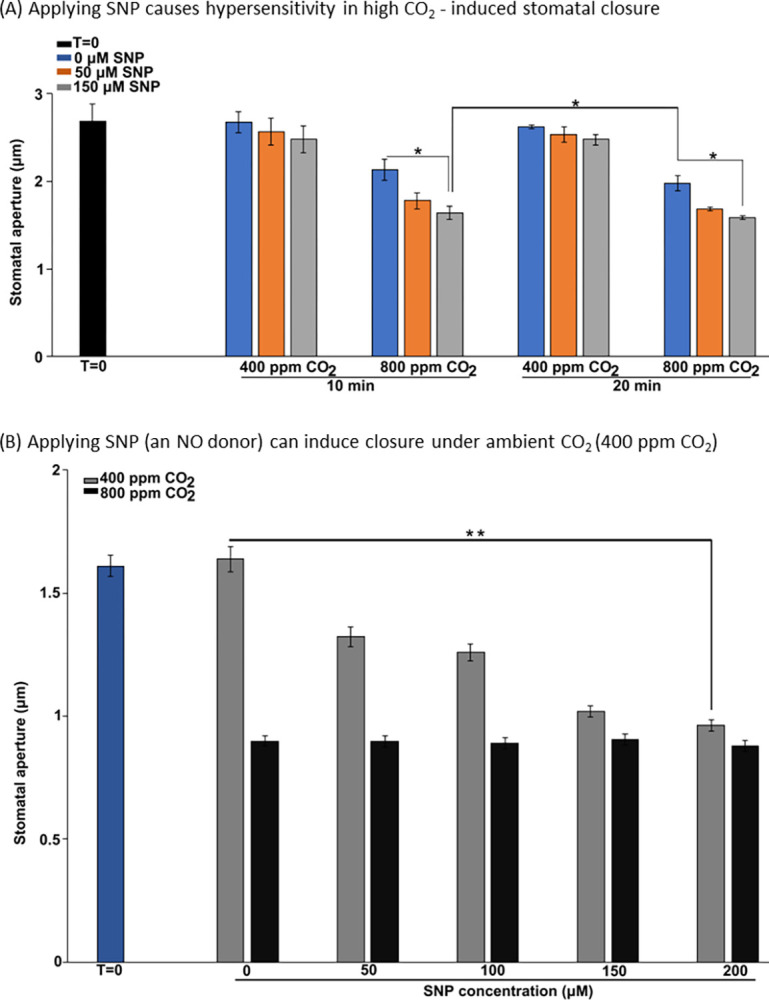
(A) As predicted by the model, applying SNP causes hypersensitivity in high CO2-induced stomatal closure: the combined treatment (orange and gray bars in the 800 ppm CO2 condition) leads to a lower mean aperture than high CO2 alone (blue bars), and a 10 min combined treatment leads to a lower mean aperture than a 20 min treatment of high CO2 alone. (B) Applying SNP (NO donor) under ambient (400 ppm) CO2 induces closure in a dose-dependent manner (gray bars; 400 ppm CO2), as predicted. The highest concentration of SNP yields a similar degree of closure as high (800 ppm) CO2 (black bars). The bars indicate the mean ± SE aperture from at least 3 independent experiments with at least 30 apertures measured per treatment. Significance of aperture differences is determined by Student’s t test: ** P < 0.01, * P < 0.05. The data underlying the graphs shown in the figure are provided in S1 Data.
Furthermore, we experimentally validate the prediction that NO can induce closure under ambient CO2. Our model predicts that under ambient CO2, NO can still activate the MSM, consequently inducing closure (Figs 4C and 6D). Previously, NO has been shown to induce closure as a signaling element of the ABA-induced closure network [62,84]. Yet, it has been unknown whether the degree of NO-induced closure (aperture reduction) is comparable to that of high CO2-induced closure. We found that applying the NO donor SNP under ambient CO2 can indeed induce stomatal closure (Fig 7B, gray bars), as predicted by our model. The reduction in aperture caused by 200 μM SNP is statistically significant (p = 1.6e-3, Student’s t test) and close to the reduction in aperture caused by CO2 of 800 ppm (black bars). This result confirms the model prediction that NO can induce closure in ambient CO2.
Taken together, these experiments not only validate our model predictions regarding the effects of NO, but also support the model-predicted importance of the MSM.
Stable motif analysis predicts a new regulatory relationship between NO and ABI2
Analysis of the inconsistencies between the model’s results and experimental observations can identify edges that are missing from the network model. The node NO also offers an example of this kind. For example, while our model recapitulates NO-induced closure in ambient CO2, it does not capture the observation that NO depletion leads to loss of stomatal closure under elevated CO2 in tomato [24]. We performed this experiment in Arabidopsis and observed that 200 μM of the NO scavenger cPTIO impairs high CO2-induced stomatal closure (Fig 8A). Under high CO2, the aperture size with cPTIO treatment is significantly different than that without cPTIO treatment (p = 1.9e-3, Student’s t test). This observation suggests that there is a not-yet-identified regulatory effect of NO that makes it necessary for a closure mechanism. This unidentified regulatory effect of NO can be predicted from our stable motif analysis. In our current model under high CO2 and NO KO, the MSM can still lock-in by the activation of its driver, the inactivity of HT1, thus leading to a closure ON attractor (Figs 8B and 4D). According to current knowledge, NO participates in the pathway that leads to calcium influx from intracellular stores into the cytosol (CIS) [97]. Disruption of this pathway eliminates one mechanism of increasing the cytosolic Ca2+ level, but it does not perturb the MSM. To be able to drive the system away from a closure ON attractor, NO KO should prevent the lock-in of the MSM that activates closure (Fig 8C). Thus, we hypothesize that NO regulates at least one of the nodes in the MSM, namely, it activates ROS, RbohD/F, or OST1 minimum function, or inhibits ABI1, ABI2, or HAB1. Indeed, we experimentally found that NO inhibits ABI2 phosphatase activity (Fig 8D and see Methods). This result suggests that the mechanism (or one of multiple contributing mechanisms) leading to impairment of stomatal closure under NO knockout could be the increase of ABI2 activity, which obstructs the minimal functional OST1 activity and precludes the lock-in of the MSM. This newly discovered inhibitory regulation likely contributes to NO-induced stomatal closure by the NO-induced decrease in the phosphatase activity of ABI2 making the activation of the MSM easier. The experimental confirmation of this stable-motif-based model prediction highlights the power of our model in predicting/prioritizing potential regulatory relationships and revealing the mechanisms of signal transduction.
Fig 8. Network-based analysis predicted a new regulatory edge between NO and PP2Cs, which was validated by experiments.

(A) The NO scavenger cPTIO impairs high CO2-induced stomatal closure. Under high (800 ppm) CO2, the aperture size with cPTIO treatment is significantly different than that without cPTIO treatment. (B) Logic diagram showing that high CO2 can activate closure despite NO KO. (C) Conceptual sketch of NO’s role in the network and the predicted edge. NO must regulate the MSM, otherwise loss of NO cannot impair high CO2-induced closure. (D) Experimental validation showing NO inhibits ABI2 phosphatase activity. The y axis shows the effect of increasing concentrations of the NO donor SNP on ABI2 activity, quantified by measuring the amount of phosphate released from a phosphopeptide substrate, as described in Methods. The bars indicate the means ± SE of 3 biological replicates. Significant differences between phosphatase activity of ABI2 alone and ABI2 treated with NO donor concentrations are indicated, as determined by Student’s t test: **P < 0.01, *P < 0.05. The data underlying the graphs shown in the figure are provided in S1 Data. MSM, Main Stable Motif; NO, nitric oxide.
Our model predicts which nodes of the high CO2 signaling network could be potential targets of currently unconnected nodes
There are signaling elements that are convincingly implicated in CO2 signaling, yet their connection to the signaling network remains unknown. During our literature review and data curation process, we placed such elements into the “insufficient connection” category. For example, big1 mutants are compromised in elevated CO2-induced stomatal closure and bicarbonate activation of S-type anion channel currents [98], yet BIG1’s connection to known guard cell signaling elements remains undiscovered. Connecting these elements to the existing CO2 network can improve understanding of guard cell CO2 signaling and could lead to testable predictions. Our model offers a way to identify and prioritize potential targets of these unconnected elements based on the effects of node perturbations (Fig 6). The idea is that, if a node perturbation yields a similar response in the model to the observed effect of the unconnected signaling element, the node is then a potential target of the signaling element. For example, the big1 mutant is known to have impaired CO2-induced closure. Our model simulations (Fig 6A) yield a comprehensive list of 24 single node perturbations that could impair closure. We predict that the big1 mutant regulates one of these nodes; any alternative single target of big1 would not cause loss of closure in the model. Moreover, one can utilize additional evidence to further narrow down the target candidates. For example, as big1 mutants are also compromised in bicarbonate activation of S-type anion channel currents, we can further filter the 24 candidate nodes to those that satisfy both conditions: (1) the node is downstream of bicarbonate; and (2) the node’s perturbation results in reduced SLAC1 activity (reflected by the node AnionEM, which incorporates the effect of both SLAC1 channel nodes). This filter narrows the list of potential targets of BIG1 to 9 nodes, namely, MPKs, ABI1, ABI2, GHR1, HT1, OST1 minimum function, RbohD/F, and ROS. One can design experiments to test if some of these nodes are indeed perturbed in big1 mutants. If a broader pool of candidate targets is needed, one could also consider the nodes whose perturbations causes hyposensitivity in high CO2-induced closure (5 nodes in the last row of Table 4). These nodes may be appropriate because it might be difficult to distinguish reduced sensitivity and hyposensitivity in experiments.
Table 4. Model-predicted potential regulation targets to impair high CO2-induced closure.
If a knockout of a signaling element impairs high CO2-induced closure, the model predicts the knockout will perturb one of these nodes. The first 3 rows are categorized based on the network location of the node, i.e., which component it is in. The last row is an extension where we hypothesize that perturbations causing hyposensitivity in the model could also be considered as having the potential to impair closure. Bold font indicates an example of 9 prioritized targets for BIG1, which are downstream of bicarbonate and whose perturbation can also cause reduced SLAC1 activity, as observed in the big1 mutant.
| Category | Potential regulation target to impair closure |
|---|---|
| High CO2 in-component or source node (6 nodes) | CAs, CO2 porins, CO2 entry of cell, HCO3-, MPKs, NADPH |
| Signal transduction SCC (10 nodes) | ABI1, ABI2, Ca2+cyt, Ca2+ATPase, GHR1, HT1, OST1 minimum function, RbohD/F, ROS |
| Ion Channel out-component (8 nodes) | H+ ATPase, Anion EM, Aquaporins, Depolarization, H2O Efflux, K+ efflux, KEV, KOUT |
| Nodes/perturbations causing hyposensitivity (5 nodes) | CaIM, CBC1/2, MPKs-HT1complex, QUAC1, SLAC1 CO2 |
By a similar logic, the “hypersensitivity” response category in Fig 6A allows target prediction for signaling elements whose perturbation (knockout or constitutive activation) promotes closure. For example, faster closure is observed in plants lacking the ABC transporter AtABCB14 [99]. Hence, our model predicts that the knockout of AtABCB14 causes a node perturbation that leads to hypersensitivity, as annotated in Fig 6A. Furthermore, one can predict/prioritize targets of a signaling element as those that can induce closure in ambient CO2, based on the node perturbations that can include closure in ambient CO2 (Fig 6C). To summarize, our model provides ways to prioritize potential targets to link previously unconnected nodes with the high CO2 signaling network. Importantly, we only used BIG1 and AtABCB14 as examples; the proposed prediction method is general.
Discussion
In this work, we constructed a signaling network that integrates current information regarding the signaling mechanisms underlying high CO2-induced stomatal closure in guard cells. We assembled the nodes and edges of the network model from a thorough literature review process, with rigorous definitions of which signaling elements are involved in CO2 signaling and which are not; we also deployed a multi-node presentation to capture multiple mechanisms of the same element. Finally, we conducted a comprehensive model performance evaluation, determining that our model indeed has high consistency with experimental observations. A previous model by Schroeder and colleagues of ABA and high CO2-induced closure, constructed as a case study of the Boolink graphical user interface [31], added to a model we originally created for ABA-induced stomatal closure [3] 6 nodes (CO2, CA1, CA4, MPK4/12, HT1, and CBC1/CBC2) and changed 5 functions (GHR1, CaIM, Ca2+c, Microtubule, and H2O efflux) without comprehensive evaluation of the model’s accuracy in recapitulating all experimental observations regarding high CO2-induced closure. In fact, 2 of the additions make the rest of the network superfluous: specifically, the function that controls CaIM has “OR CO2” in it, which means that high CO2 induces CaIM regardless of the rest of the regulators of CaIM; the function of H2O efflux also has “OR CO2” in it, which makes water efflux happen directly in response to CO2 regardless of the rest of the regulators of aquaporins. The combined effect of these 2 assumptions results in closure. In other words, the model of Karanam and colleagues short-circuits essentially the entire CO2 signaling network by making CO2 directly activate the very downstream part of the network. As a result, the model will not be able to capture the experimentally observed responses to many perturbations in the signaling network, e.g., the impaired high CO2-induced closure under slac1 [81], ht1-2 [12], and mpk4/mpk12 [14] mutants. By comparison, a rigorous model construction process allows our CO2 model to comprehensively reproduce experimental observations related to high CO2-induced closure.
Network methodologies reveal key decision-making motifs, pinpoint driver nodes, elucidate signal transduction mechanisms, predict new edges and responses
Our network-based modeling framework integrates piece-wise (node to node) biological knowledge to form a comprehensive view of the signal transduction network underlying high CO2-induced stomatal closure. This framework complements conventional biological approaches that often focus on single regulators or single pathways. Our analysis supports recent findings that HT1 is a key decision-making node of the CO2 signaling system [15,16] and offers additional insights regarding the ambient CO2 setting: closure induced by, e.g., exogenous ROS or Ca2+ in ambient CO2 must involve the inactivation of HT1 and the activation of a positive feedback loop (the MSM, formed by ROS, OST1, and the PP2Cs). Conversely, under conditions that activate HT1 the nodes of this positive feedback loop may also lock-in to their opposite states and lead to closure OFF.
We have also introduced the concept of logic diagrams to visually depict the logical implications of events in the closure process, highlighting key nodes and pathways (subgraphs) responsible for decision-making of the system. The logic diagrams explain not only how signal transduction leads to stomatal closure under high CO2, but also how closure is lost or restored. Leveraging these concepts, we explain the mechanisms behind the 5 types of simulated closure responses in the presence of perturbations.
Combining these network-based methodologies allows the generation of model predictions that lead to novel experimental findings. It is important to note that inconsistencies between the model and experimental results also yield insights. For example, we found that the model is incapable of reproducing the experimental observation that NO knockout impairs high CO2-induced closure; by understanding that this inconsistency results from the NO-independent lock-in of the MSM, we predicted that NO must regulate the MSM, and successfully validated this prediction experimentally, showing that NO inhibits ABI2, one of the PP2Cs in the MSM.
Unexplained mechanisms in CO2 signaling
There are signaling elements that are known to mediate closure in response to a certain signal, but are understudied in CO2 signaling. In our model construction process, when we designate each piece of knowledge regarding the regulation of a node by another into one of the 5 categories (see Tables 2 and S1), we place such elements into the “generic” or “insufficient evidence” categories. This way, our categorization of evidence can prioritize experiments to explore the roles of signaling elements in the context of CO2 signaling.
Past studies on the convergence of the signaling networks corresponding to ABA-induced stomatal closure and high CO2-induced stomatal closure yielded inconsistent results. This convergence was first suggested by Webb and Hetherington [5]. Xue and colleagues [9] found that the quadruple ABA receptor mutant pyr1;pyl1;pyl2;pyl4 showed slower closure but no other impairment under 800 ppm CO2. On the contrary, Chater and colleagues [22] and Dittrich and colleagues [100] found that quintuple mutants of ABA receptors from the PYR/PYL/RCAR family exhibit impaired high CO2-induced closure, and that ABA biosynthesis-deficient nced3;nced5 plants were impaired in high CO2-induced closure. Movahedi and colleagues [101] reported hypersensitivity to CO2 in cyp707a1 cyp707a3 ABA-hyper-accumulating mutants, supporting the convergence of the 2 pathways. By contrast, there is also evidence for ABA-independence of high CO2 signaling [2], e.g., OST1/SnRK2.6 protein kinase activity in guard cells is not activated by CO2 elevation [34,35], and cytosolic pH does not increase under high CO2 [9,32]. Moreover, CO2 regulation of PYR/PYL/RCAR signaling activity, as well as a mechanism by which PYR/PYL/RCAR receptors would affect high CO2-induced closure independently of changes in ABA concentration is not known. Considering all the above evidence, we placed RCARs in the “insufficient connection to CO2” category when constructing this model. Nevertheless, our model can be expanded to include a node that represents endogenous ABA if one assumes that it is necessary for OST1 minimum function but not sufficient to inhibit the PP2Cs and we now discuss this scenario in S2 Text section IV.
Our model construction and validation process included the examination of multiple different hypotheses regarding the role of certain nodes in high CO2 signaling. For example, the role of RHC1 in CO2 signaling is under debate: while Tian and colleagues showed experimental evidence that RHC1 is an essential CO2 signaling component [17], Tõldsepp and colleagues observed that a different rhc1 mutant shows no reduction of high CO2-induced closure. We tested both versions of the HT1 regulatory function, with and without RHC1, and found that the function without RHC1 has a higher accuracy in capturing experimental observations. Therefore, we adopt the HT1 function without RHC1, more consistent with [18]. Our interpretation is that RHC1’s regulatory effect on HT1 is weaker than that of MPKs, thus it is insufficient to include in a Boolean model.
Some CO2 signaling mechanisms remain unclear. For example, observations about guard cell cytosolic pH suggests undiscovered buffering capacity in the cytosol. Xue and colleagues and Brearley and colleagues [9,32] observed that cytosolic pH does not increase under high CO2-induced closure. On the other hand, after CO2 enters the guard cell, it undergoes the chemical reaction CO2 + H2O → HCO3– + H+ catalyzed by CAs, producing H+. So, there must be some buffering capacity or other mechanism to keep cytosolic pH constant. Understanding this mechanism may affect other components in the network, and its refinement may improve the model as well as our understanding of the high CO2 signaling mechanism.
Future discovery of new signaling elements and interactions that participate in high CO2-induced closure will call for updates of this model, including the addition of new nodes and/or new edges, the possible removal of assumed edges (shown as dotted lines in Fig 2), or changes to the regulatory functions of certain nodes. It is important to note that even when new signaling elements and interactions are incorporated into the CO2 model, certain structural and dynamic features of the model will be preserved. For example, the network will still have a strongly connected component (see Fig 1B) that includes at least the 20 nodes currently in it. The updates will be such that the model preserves the current model’s agreements with experimental results. The coupling of the structural and dynamical preservation means that the decision-making role of the MSM will also be maintained in the updated model, even if newly added edges will increase its size (for example, by NO joining it). It is likely that newly discovered interactions will support the regulatory relationships that drive the decisions of the system. For example, the inhibition of ABI2 by NO is an additional mechanism by which NO contributes to the activation of the MSM. Ultimately, a cycle between experimental and modeling approaches offers the most effective way to elucidate the full network of high CO2-induced stomatal closure.
Methods and data
Construction of the CO2-signaling network and dynamic model
As described in the main text, we extract regulatory relationships from the literature (S1 Table) and compile them into nodes and edges of the network (S2 Table). Then, we construct the regulatory functions (S1 Text) based on the regulatory relationships.
Simulation settings of the dynamical model
In simulation, we initialize the in silico stomata in a pre-stimulus open state, with closure-inhibiting nodes ON (namely ABI1, ABI2, HAB1, HT1, and H+ ATPase) and other nodes OFF (see S1 Text for complete initial condition). For example, Anion Efflux, a node that becomes activated during high CO2-induced closure, is initialized in the OFF state. We deploy a discrete-time simulation, with stochastic random order asynchronous update: at each discrete round of updates (or timestep), all nodes are updated in a randomly selected order. A different order is randomly generated in each timestep. The main advantage of random order asynchronous update is that it can capture consensus dynamics without implementation of exact timing, as the actual timing and durations of the processes happening in the system are typically unknown. The stochasticity of the update helps avoid spurious behaviors that depend on synchrony (i.e., which only appear under synchronous update, where all nodes are updated at the same time, which is biologically unrealistic). We perform this simulation for 30 timesteps, long enough for the system to reach its attractor (stable long-time behavior). In this way, we can simulate how a node’s state changes over time during the stomatal closure process.
We perform 1,000 replicate simulations, i.e., 1,000 “in silico stomata” simulations, then calculate the “percentage of node activation” at each timestep as our metric, meaning the percentage of simulations in which the node (e.g., Closure) is in state 1 (ON). For example, if 1,000 replicate simulations yield 510 cases where closure is ON at timestep 30, the simulated percentage of closure is 51%. In this way, oscillations and multistability with different node states will display a percentage of node activation value between 0% and 100%. A sample of our simulation code is provided on GitHub: https://github.com/jack802gan/CO2-model. We also recommend Cubewalker [102] as a more recent and more efficient simulation tool for Boolean dynamic models.
Stable motif and attractor analysis
Stable motifs are the smallest strongly connected components in a system that can sustain a unique steady state for the constituent nodes [28,86]. Stable motifs are identified in an expanded representation of the network (which includes the network’s regulatory functions as part of the network structure). A weaker version of a stable motif, called a conditionally stable motif [92], can sustain a unique steady state for its nodes as long as certain node(s) that regulate the motif are in a specific fixed state. The stable motifs and conditionally stable motifs of a Boolean network determine its attractors: one can uniquely associate sequences of stable motifs (stabilized in the order given by the sequence) to each attractor.
A driver (or driver set) of a stable motif is the node (or node combination) and its state that, when kept fixed, eventually leads to the locking in of the stable motif after sufficient updates. We used the python library pystablemotifs [29] to analyze the stable motifs, attractors, and their control methods. The library is available through the GitHub page https://github.com/jcrozum/pystablemotifs.
Validation against experimental observations
We use the experimental observations in S1 Table that were not used in model construction (a subset of the indirect regulations, marked as “V” for validation in S1 Table) as the ground truth for model validation. Because quantifying the (reduction of) stomatal aperture is difficult in both experiment and Boolean model, when comparing the experiment-based expected outcomes with the model simulation results, we employ a lenient criterion for consistency: any model-predicted defect (i.e., node percentage < 100%) is considered qualitatively consistent with either reduced response or no response. For example, if a simulation yields a closure percentage = 0.5, then it is considered consistent with both “reduced closure” and “no closure” in experimental observation. This type of consistency criterion was used in previous modeling as well [3]. We provide the comprehensive validation results in S3 Table. The supplementary file also includes the trivially consistent cases due to circular reasoning, denoted “true by construction.” For example, bicarbonate-induced activation of S-type anion channels was reduced in the dominant active abi1 mutants (ABI1 CA) [78], but because ABI1 is an input in both SLAC1 functions and ABI1 ON is sufficient to deactivate both SLAC1 nodes, this experimental observation is captured by model/rule construction, and calling it a validation would be circular reasoning. This type of validation is not counted in our assessment of model accuracy performance.
Definition of the response categories in CPC values, for simulated knockout or constitutive activity
We defined the CPC as the sum over 30 timesteps of the fraction of simulations that have the closure node in state 1 (ON), over 1,000 replicate simulations. Thus, CPC ranges from 0 (if the percentage of closure were 0% at every timestep) to 30 (if the percentage of closure were 100% at every timestep). The CPC metric helps capture time-sensitive behavior such as hypo-/hyper-sensitivity, reflected as a small CPC difference from wild-type CPC.
We define the 5 categories of responses based on the CPC value and attractor state: close to wild type, hypersensitivity, hyposensitivity, reduced closure, and no closure. We define the wild-type response range from the default high CO2-induced closure response. Since the simulation is stochastic (i.e., the CPC value can be different for each simulation), we determine the default CPC range by 600 batches of 1,000 replicate simulations under the default high-CO2 signal, and define its CPC range as the “close to wild type” CPC range (20.405 to 20.709). CPC values above this range are classified as hypersensitivity, meaning that the stomata reach the closure state significantly faster than wild type. CPC values below the wild type range with 100% percentage closure at the final timestep are classified as hyposensitivity. Reduced closure is the CPC value being much lower than wild type range, with the final percentage of closure being between 0 and 1. In all of the observed cases in this last category, the closure value is oscillating in the attractor. Lastly, no closure is defined as the CPC value being zero or very close to zero, with final percentage of closure being zero. A small positive CPC can occur here, because closure may temporarily turn ON during the response process, though it cannot be maintained. The comprehensive CPC simulation results are provided in S4 Table.
Logic diagrams of the CO2 network model
The logic diagram is an integration of the network and node states that yields a specific outcome (e.g., the ON state of the node Closure). It is an expansion of the concepts of elementary signaling mode [93] and logical domain of influence [94], both of which are subgraphs of the expanded network (also called logical hypergraph), which incorporates the Boolean functions of all the nodes in the network [86]. The logic diagram indicates the node states visually: nodes whose name is preceded by ~ and have gray background are inactive, nodes with green background are active, and nodes whose name are followed by “osc” and have a white background oscillate. A single edge in a logic diagram represents a sufficient regulatory relationship that can achieve the indicated state of the target node; multiple edges incident on the same black dot represents “AND” logic, indicating that all nodes are required to achieve the target node state. A logic diagram may condense fully linear pathways of nodes and edges in the original network, to simplify visualization. This type of condensation does not change the regulatory logic of the network model [95].
To help visual parsing in the CO2 network model, we also merged a few nodes of the original network. The node “PP2Cs” merges 3 members of the PP2C protein family, because these nodes have identical regulation. The technical definition of the merged node is PP2Cs = ABI1 or ABI2 or HAB1. The node “Ion flow” merges AnionEM and K+ efflux (i.e., Ion flow = AnionEM and K+ efflux). We also use a single node “Ca2+cyt osc” to represent the Ca2+cyt - Ca2+ ATPase negative feedback cycle, capturing the oscillating behavior of Ca2+cyt in the presence of a Ca2+ source (CaIM or CIS) and due to the inhibitory effect of the Ca2+ ATPase. We use hexagonal symbols for “PP2Cs” and “Ion efflux” as they represent a node set (Fig 5). Note the “Ion efflux” hexagon node needs multiple incident edges to activate, causing an exception of the edge definition.
The logic diagram can be read as a sequence of events and the conditions necessary for these events. For example, the sequence of events necessary for high CO2-induced closure described by Fig 5A is the following: high CO2 activates HCO3-, which inactivates HT1 via the formation of the MPKs-HT1 complex. HT1 being OFF then activates OST1 minimum function, which eventually activates the MSM. The simultaneous activation of the MSM and inactivation of HT1 (indicated by the black dot) activates GHR1, which in turn activates CPKs. GHR1 activates CaIM, which leads to cytosolic Ca2+ transients or oscillations. The MSM, GHR1, Ca2+cyt, HCO3-, as well as the inactivity of CBC1/2 contribute to the activation of Ion flows. The simultaneous activation of Ion flows and OST1-activated Aquaporin yield H2O efflux, which then leads to Closure. As another example, the logic diagram in Fig 6D shows that the activation of the MSM (e.g., by CaIM) leads to CIS, which then leads to cytosolic Ca2+ transients or oscillations, which activate MPK activity, which inhibits HT1, which then allows the activation of GHR1, then of the generic SLAC1 node, and finally the activation of the node Closure.
Plant materials and growth conditions
Arabidopsis thaliana Columbia (Col-0) accession plants were used as wild type. Col-0 were germinated on MS agar plates for 2 weeks and then grown in soil (1:1 mixture of Metro Mix 360: Sunshine Mix LC1 (Sun Gro Horticulture) in a growth chamber with an 8-h-light/16-h-dark cycle, 150 μmol photons m−2 s−1 light, and 22°C (day)/19°C (night)). Young fully expanded leaves from 4- to 5-week-old plants were used for stomatal aperture assay experiments.
Stomatal aperture assay
Stomatal apertures were assayed as described previously with slight modifications [103]. In brief, epidermal peels were hand-peeled from the abaxial surfaces of leaves and then were floated interior-side down on an opening solution (20 mM KCl, 10 mM MES-KOH (pH 6.15), 50 μM CaCl2) under white light (150 μmol m−2 sec−1) for 3 h to induce maximum opening of the stomata (initial aperture). After the 3 h preincubation period, epidermal peels were floated in a six-well plate (each well with a diameter of 35 mm and height of 17 mm) containing pre-equilibrated high (800 ppm) or ambient (400 ppm) CO2 opening solution (approximately 3 ml). Subsequently, the multiwell cell culture plate was placed in a petri dish chamber and sealed with parafilm. The high or ambient CO2 concentration was continuously maintained within the chamber using a Licor-6400 (flow rate of 500 μmol s−1) while the samples were further incubated under white light (150 μmol m−2 sec−1). After 3 h of incubation, the epidermal peels were imaged with a 40× objective using a light microscope (Nikon Diaphot 300) coupled to a digital camera (Nikon Coolpix 990). Stomatal apertures were measured in each image using ImageJ. To determine the effects of NO donor SNP (sodium nitroprusside dihydrate; Sigma, catalog number 71778), SNAP (S-Nitroso-N-acetyl-DL—penicillamine; Sigma, catalog number N3398), or NO scavenger (2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide; Sigma, catalog number C221) on CO2-induced stomatal closure, experiments were performed according to the method described by [104] with some modifications. Briefly, epidermal peels were incubated for 3 h in the opening buffer to induce stomatal opening. Subsequently, the epidermal peels were transferred to pre-equilibrated high (800 ppm) or ambient (400 ppm) CO2 opening solution (approximately 3 ml) added without or with NO donor or NO scavenger at the indicated concentrations. Then, the epidermal peels were treated under ambient CO2 (400 ppm CO2) or elevated CO2 (800 ppm CO2) in light for the indicated durations. A total of at least 30 stomatal apertures were measured per treatment for each individual experiment. The datapoint for each individual experiment is the mean aperture of these stomata. Data presented in Figs 7A, 7B, and 8A are the mean and standard error in the mean of the 3 data points coming from 3 independent experiments. The relevant data are provided in S1 Data.
Phosphopeptide-based phosphatase activity assay
To examine the effect of NO donor on ABI2 phosphatase activity, GST-tagged ABI2 was heterologously expressed in Escherichia coli and subsequently purified as described [105] with minor modifications. Briefly, pDEST15-ABI2 was constructed by transferring ABI2 cDNA from pCR8/GW/TOPO TA vector to the N-terminal GST fusion expression vector pDEST15 (Invitrogen) by Gateway LR recombination (Invitrogen). The cloned construct was verified by sequencing. The GST-ABI2 protein was expressed in E. coli BL21(DE3) pLysS Rosetta cells by induction with 0.5 mM IPTG for 16 h at 16°C. The cells were harvested by centrifugation (6,000 g, 8 min, 4°C) and stored at −80°C until use. All purification procedures were carried out at 4°C. The final cell pellet was resuspended in 10 ml of B-PER reagent (Pierce, catalog number 78243) supplemented with 1 mg/ml lysozyme (Sigma, catalog number L6876), 25 mg/ml DNAse and EDTA-free complete protease inhibitor (Thermo Fisher Scientific, catalog number A32955), and incubated for 60 min at 4°C with gentle shaking. The cell lysate was spun at 16,000g for 15 min at 4°C and the resulting supernatant was filtered using a 0.45 μm filter. Subsequently, the supernatant was loaded onto a 5 ml QIAGEN polypropylene column packed with GST glutathione agarose resin (Pierce, catalog number 16100) and the column-bound protein was eluted with equilibration buffer (25 mM Tris-HCl (pH 7.5), 150 mM NaCl) supplemented with 20 mM reduced L-glutathione (Sigma, catalog number G4251). The purity of the recombinant GST-ABI2 was estimated by analysis of aliquots separated on a 12% SDS polyacrylamide gel stained with Gel-Code Blue (Thermo), and the concentration was estimated based on known concentrations of Fraction V BSA (Pierce, catalog number 23209) run on the same gel.
The effect of NO donor on ABI2 phosphatase activity was assayed using the Ser/Thr phosphatase assay kit (Promega; catalog number V2460). The reaction was performed in a 50 μl volume containing 28 nM of purified recombinant GST-ABI2 protein in the presence or absence of the indicated concentration of NO donor SNP. After 30 min incubation at 30°C, the reaction was stopped by addition of 50 μl of molybdate dye/additive solution. The resulting mix was incubated for another 20 min at room temperature followed by absorbance measurements at 630 nm in a Synergy Neo2 multimode plate reader (Biotek). The amount of released phosphate was calculated based on a standard curve obtained using phosphate standard solutions of known concentration. The relevant data are provided in S1 Data.
Statistical testing
We used two-tailed Student’s t tests when comparing the stomatal aperture measurements, to evaluate if a significant change of aperture size or phosphatase activity is observed in an experiment. The statistical tests were performed in Python (Anaconda JupyterLab). Relevant data and an example of code are provided on GitHub: https://github.com/jack802gan/CO2-model. DOI: https://zenodo.org/doi/10.5281/zenodo.10665442
Supporting information
(DOCX)
(DOCX)
(XLSX)
(XLSX)
(DOCX)
(DOCX)
(XLSX)
Acknowledgments
The authors thank Dr. David Chakravorty, Dr. Yotam Zait, Dr. Ángel Ferrero-Serrano, and Dr. Jordan C. Rozum for helpful discussions.
Abbreviations:
- ABA
abscisic acid
- CPC
cumulative percentage of closure
- MSM
Main Stable Motif
- NO
nitric oxide
- ROS
reactive oxygen species
- SCC
strongly connected component
Data Availability
All relevant data are provided in the paper, its Supporting Information files, and the GitHub repository https://github.com/jack802gan/CO2-model. DOI: https://zenodo.org/doi/10.5281/zenodo.10665442.
Funding Statement
This work was supported by NSF grant MCB 1715826 to S.M.A. and R.A. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Assmann SM, Jegla T. Guard cell sensory systems: recent insights on stomatal responses to light, abscisic acid, and CO2. Curr Opin Plant Biol. 2016;33:157–167. Epub 20160809. doi: 10.1016/j.pbi.2016.07.003 [DOI] [PubMed] [Google Scholar]
- 2.Hsu PK, Dubeaux G, Takahashi Y, Schroeder JI. Signaling mechanisms in abscisic acid-mediated stomatal closure. Plant J. 2021;105(2):307–321. Epub 20201209. doi: 10.1111/tpj.15067 PubMed Central PMCID: PMC7902384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Albert R, Acharya BR, Jeon BW, Zañudo JGT, Zhu M, Osman K, et al. A new discrete dynamic model of ABA-induced stomatal closure predicts key feedback loops. PLoS Biol. 2017;15(9):e2003451. doi: 10.1371/journal.pbio.2003451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Munemasa S, Hauser F, Park J, Waadt R, Brandt B, Schroeder JI. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr Opin Plant Biol. 2015;28:154–162. doi: 10.1016/j.pbi.2015.10.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Webb AA, Hetherington AM. Convergence of the abscisic acid, CO2, and extracellular calcium signal transduction pathways in stomatal guard cells. Plant Physiol. 1997;114(4): 1557–1560. doi: 10.1104/pp.114.4.1557 PubMed Central PMCID: PMC158450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dr. Pieter Tans, NOAA/GML (gml.noaa.gov/ccgg/trends/) and Dr. Ralph Keeling, Scripps Institution of Oceanography (scrippsco2.ucsd.edu/).
- 7.Wang C, Hu H, Qin X, Zeise B, Xu D, Rappel WJ, et al. Reconstitution of CO2 Regulation of SLAC1 Anion Channel and Function of CO2-Permeable PIP2;1 Aquaporin as CARBONIC ANHYDRASE4 Interactor. Plant Cell. 2016;28(2):568–582. Epub 20160113. doi: 10.1105/tpc.15.00637 PubMed Central PMCID: PMC4790870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hu H, Boisson-Dernier A, Israelsson-Nordström M, Böhmer M, Xue S, Ries A, et al. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells. Nat Cell Biol. 2010;12(1):87–93. sup pp 1–18. Epub 20091213. doi: 10.1038/ncb2009 PubMed Central PMCID: PMC2906259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xue S, Hu H, Ries A, Merilo E, Kollist H, Schroeder JI. Central functions of bicarbonate in S-type anion channel activation and OST1 protein kinase in CO2 signal transduction in guard cell. EMBO J. 2011;30(8):1645–1658. Epub 20110318. doi: 10.1038/emboj.2011.68 PubMed Central PMCID: PMC3102275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sun P, Isner J-C, Coupel-Ledru A, Zhang Q, Pridgeon AJ, He Y, et al. Countering elevated CO2 induced Fe and Zn reduction in Arabidopsis seeds. New Phytol. 2022;235(5):1796–1806. doi: 10.1111/nph.18290 [DOI] [PubMed] [Google Scholar]
- 11.Hashimoto M, Negi J, Young J, Israelsson M, Schroeder JI, Iba K. Arabidopsis HT1 kinase controls stomatal movements in response to CO2. Nat Cell Biol. 2006;8(4):391–397. Epub 20060305. doi: 10.1038/ncb1387 [DOI] [PubMed] [Google Scholar]
- 12.Hashimoto-Sugimoto M, Negi J, Monda K, Higaki T, Isogai Y, Nakano T, et al. Dominant and recessive mutations in the Raf-like kinase HT1 gene completely disrupt stomatal responses to CO2 in Arabidopsis. J Exp Bot. 2016;67(11):3251–3261. Epub 20160331. doi: 10.1093/jxb/erw134 PubMed Central PMCID: PMC4892718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hõrak H, Sierla M, Tõldsepp K, Wang C, Wang YS, Nuhkat M, et al. A Dominant Mutation in the HT1 Kinase Uncovers Roles of MAP Kinases and GHR1 in CO2-Induced Stomatal Closure. Plant Cell. 2016;28(10):2493–2509. Epub 20160930. doi: 10.1105/tpc.16.00131 PubMed Central PMCID: PMC5134974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jakobson L, Vaahtera L, Tõldsepp K, Nuhkat M, Wang C, Wang YS, et al. Natural Variation in Arabidopsis Cvi-0 Accession Reveals an Important Role of MPK12 in Guard Cell CO2 Signaling. PLoS Biol. 2016;14(12):e2000322. Epub 20161206. doi: 10.1371/journal.pbio.2000322 PubMed Central PMCID: PMC5147794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Takahashi Y, Bosmans KC, Hsu P-K, Paul K, Seitz C, Yeh C-Y, et al. Stomatal CO2/bicarbonate sensor consists of two interacting protein kinases, Raf-like HT1 and non-kinase-activity requiring MPK12/MPK4. Sci Adv. 2022;8(49): doi: 10.1126/sciadv.abq6161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yeh C-Y, Wang Y-S, Takahashi Y, Kuusk K, Paul K, Arjus T, et al. MPK12 in stomatal CO2 signaling: function beyond its kinase activity. New Phytol. 2023;239(1). doi: 10.1111/nph.18913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tian W, Hou C, Ren Z, Pan Y, Jia J, Zhang H, et al. A molecular pathway for CO2 response in Arabidopsis guard cells. Nat Commun. 2015;6(1):6057. doi: 10.1038/ncomms7057 [DOI] [PubMed] [Google Scholar]
- 18.Tõldsepp K, Zhang J, Takahashi Y, Sindarovska Y, Hõrak H, Ceciliato PHO, et al. Mitogen-activated protein kinases MPK4 and MPK12 are key components mediating CO2-induced stomatal movements. Plant J. 2018;96(5):1018–1035. Epub 20181023. doi: 10.1111/tpj.14087 PubMed Central PMCID: PMC6261798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Webb AAR, McAinsh MR, Mansfield TA, Hetherington AM. Carbon dioxide induces increases in guard cell cytosolic free calcium. Plant J. 1996;9(3): 297–304. 10.1046/j.1365-313X.1996.09030297.x [DOI] [Google Scholar]
- 20.Young JJ, Mehta S, Israelsson M, Godoski J, Grill E, Schroeder JI. CO(2) signaling in guard cells: calcium sensitivity response modulation, a Ca(2+)-independent phase, and CO(2) insensitivity of the gca2 mutant. Proc Natl Acad Sci U S A. 2006;103(19):7506–7511. Epub 20060501. doi: 10.1073/pnas.0602225103 PubMed Central PMCID: PMC1464368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hubbard KE, Siegel RS, Valerio G, Brandt B, Schroeder JI. Abscisic acid and CO2 signalling via calcium sensitivity priming in guard cells, new CDPK mutant phenotypes and a method for improved resolution of stomatal stimulus-response analyses. Ann Bot. 2012;109(1):5–17. Epub 20111012. doi: 10.1093/aob/mcr252 PubMed Central PMCID: PMC3241576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chater C, Peng K, Movahedi M, Dunn JA, Walker HJ, Liang YK, et al. Elevated CO2-Induced Responses in Stomata Require ABA and ABA Signaling. Curr Biol. 2015;25(20):2709–2716. Epub 20151008. doi: 10.1016/j.cub.2015.09.013 PubMed Central PMCID: PMC4612465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kolla VA, Vavasseur A, Raghavendra AS. Hydrogen peroxide production is an early event during bicarbonate induced stomatal closure in abaxial epidermis of Arabidopsis. Planta. 2007;225(6):1421–1429. Epub 20061208. doi: 10.1007/s00425-006-0450-6 [DOI] [PubMed] [Google Scholar]
- 24.Shi K, Li X, Zhang H, Zhang G, Liu Y, Zhou Y, et al. Guard cell hydrogen peroxide and nitric oxide mediate elevated CO2 -induced stomatal movement in tomato. New Phytol. 2015;208(2):342–353. Epub 20150826. doi: 10.1111/nph.13621 [DOI] [PubMed] [Google Scholar]
- 25.Li S, Assmann SM, Albert R. Predicting essential components of signal transduction networks: a dynamic model of guard cell abscisic acid signaling. PLoS Biol. 2006;4(10):doi: 10.1371/journal.pbio.0040312 PubMed Central PMCID: PMC1564158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sun Z, Jin X, Albert R, Assmann SM. Multi-level Modeling of Light-Induced Stomatal Opening Offers New Insights into Its Regulation by Drought. PLoS Comput Biol. 2014;10(11):e1003930. doi: 10.1371/journal.pcbi.1003930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zañudo Jorge GT, Albert R. Cell Fate Reprogramming by Control of Intracellular Network Dynamics. PLoS Comput Biol. 2015;11(4):1–24. doi: 10.1371/journal.pcbi.1004193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rozum JC, Gómez Tejeda Zañudo J, Gan X, Deritei D, Albert R. Parity and time reversal elucidate both decision-making in empirical models and attractor scaling in critical Boolean networks. Sci Adv. 2021;7(29). Epub 20210716.doi: 10.1126/sciadv.abf8124 PubMed Central PMCID: PMC8284893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rozum JC, Deritei D, Park KH, Zañudo JGT, Albert R. pystablemotifs: Python library for attractor identification and control in Boolean networks. Bioinformatics. 2021. Epub 20211207. doi: 10.1093/bioinformatics/btab825 [DOI] [PubMed] [Google Scholar]
- 30.Gan X, Albert R. General method to find the attractors of discrete dynamic models of biological systems. Physical Review E. 2018;97(4). doi: 10.1103/PhysRevE.97.042308 [DOI] [PubMed] [Google Scholar]
- 31.Karanam A, He D, Hsu PK, Schulze S, Dubeaux G, Karmakar R, et al. Boolink: a graphical interface for open access Boolean network simulations and use in guard cell CO2 signaling. Plant Physiol. 2021;187(4):2311–2322. doi: 10.1093/plphys/kiab344 PubMed Central PMCID: PMC8644243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brearley J, Venis MA, Blatt MR. The effect of elevated CO2 concentrations on K+ and anion channels of Vicia faba L. guard cells. Planta. 1997;203(2):145–154. [Google Scholar]
- 33.Savchenko G, Wiese C, Neimanis S, Hedrich R, Heber U. pH regulation in apoplastic and cytoplasmic cell compartments of leaves. Planta. 2000;211(2):246–255. doi: 10.1007/s004250000280 [DOI] [PubMed] [Google Scholar]
- 34.Hsu PK, Takahashi Y, Munemasa S, Merilo E, Laanemets K, Waadt R, et al. Abscisic acid-independent stomatal CO2 signal transduction pathway and convergence of CO2 and ABA signaling downstream of OST1 kinase. Proc Natl Acad Sci U S A. 2018;115(42):E9971–E9980. Epub 20181003. doi: 10.1073/pnas.1809204115 PubMed Central PMCID: PMC6196521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Zhang L, Takahashi Y, Hsu PK, Kollist H, Merilo E, Krysan PJ, et al. FRET kinase sensor development reveals SnRK2/OST1 activation by ABA but not by MeJA and high CO. Elife. 2020:9. Epub 20200528. doi: 10.7554/eLife.56351 PubMed Central PMCID: PMC7289597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gan X, Albert R. Modeling biological information processing networks. In: Blagoev KB, Levine H, editors. Physics of Molecular and Cellular Processes. Graduate Texts in Physics. 1st ed. Springer International Publishing; 2022. [Google Scholar]
- 37.Jammes F, Song C, Shin D, Munemasa S, Takeda K, Gu D, et al. MAP kinases MPK9 and MPK12 are preferentially expressed in guard cells and positively regulate ROS-mediated ABA signaling. Proc Natl Acad Sci U S A. 2009;106(48):20520–20525. Epub 20091112. doi: 10.1073/pnas.0907205106 PubMed Central PMCID: PMC2776606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Joudoi T, Shichiri Y, Kamizono N, Akaike T, Sawa T, Yoshitake J, et al. Nitrated cyclic GMP modulates guard cell signaling in Arabidopsis. Plant Cell. 2013;25(2):558–571. Epub 2013/02/12. doi: 10.1105/tpc.112.105049 PubMed Central PMCID: PMC3608778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sethi JK, Empson RM, Galione A. Nicotinamide inhibits cyclic ADP-ribose-mediated calcium signalling in sea urchin eggs. Biochem J. 1996;319(Pt 2):613–617. PubMed Central PMCID: PMC1217811. doi: 10.1042/bj3190613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Walseth TF, Lee HC. Synthesis and characterization of antagonists of cyclic-ADP-ribose-induced Ca2+ release. Biochim Biophys Acta. 1993;1178(3):235–242. doi: 10.1016/0167-4889(93)90199-y [DOI] [PubMed] [Google Scholar]
- 41.Rakovic S, Cui Y, Iino S, Galione A, Ashamu GA, Potter BV, et al. An antagonist of cADP-ribose inhibits arrhythmogenic oscillations of intracellular Ca2+ in heart cells. J Biol Chem. 1999;274(25):17820–17827. doi: 10.1074/jbc.274.25.17820 [DOI] [PubMed] [Google Scholar]
- 42.Nishimura N, Sarkeshik A, Nito K, Park SY, Wang A, Carvalho PC, et al. PYR/PYL/RCAR family members are major in-vivo ABI1 protein phosphatase 2C-interacting proteins in Arabidopsis. Plant J. 2010;61(2):290–299. Epub 2009/10/31. doi: 10.1111/j.1365-313X.2009.04054.x PubMed Central PMCID: PMC2807913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Umezawa T, Sugiyama N, Mizoguchi M, Hayashi S, Myouga F, Yamaguchi-Shinozaki K, et al. Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proc Natl Acad Sci U S A. 2009;106(41):17588–17593. doi: 10.1073/pnas.0907095106 PubMed Central PMCID: PMC2754379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Vlad F, Rubio S, Rodrigues A, Sirichandra C, Belin C, Robert N, et al. Protein phosphatases 2C regulate the activation of the Snf1-related kinase OST1 by abscisic acid in Arabidopsis. Plant Cell. 2009;21(10):3170–3184. doi: 10.1105/tpc.109.069179 PubMed Central PMCID: PMC2782292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Geiger D, Scherzer S, Mumm P, Stange A, Marten I, Bauer H, et al. Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proc Natl Acad Sci U S A. 2009;106(50):21425–21430. Epub 2009/12/04. doi: 10.1073/pnas.0912021106 PubMed Central PMCID: PMC2795561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Scherzer S, Maierhofer T, Al-Rasheid KA, Geiger D, Hedrich R. Multiple calcium-dependent kinases modulate ABA-activated guard cell anion channels. Mol Plant. 2012;5(6):1409–1412. Epub 2012/08/31. doi: 10.1093/mp/sss084 [DOI] [PubMed] [Google Scholar]
- 47.Geiger D, Scherzer S, Mumm P, Marten I, Ache P, Matschi S, et al. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca2+ affinities. Proc Natl Acad Sci U S A. 2010;107(17):8023–8028. Epub 20100412. doi: 10.1073/pnas.0912030107 PubMed Central PMCID: PMC2867891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hua D, Wang C, He J, Liao H, Duan Y, Zhu Z, et al. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis. Plant Cell. 2012;24(6):2546–2561. Epub 20120622. doi: 10.1105/tpc.112.100107 PubMed Central PMCID: PMC3406912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Grondin A, Rodrigues O, Verdoucq L, Merlot S, Leonhardt N, Maurel C. Aquaporins Contribute to ABA-Triggered Stomatal Closure through OST1-Mediated Phosphorylation. Plant Cell. 2015;27(7):1945–1954. Epub 20150710. doi: 10.1105/tpc.15.00421 PubMed Central PMCID: PMC4531361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sasaki T, Mori IC, Furuichi T, Munemasa S, Toyooka K, Matsuoka K, et al. Closing plant stomata requires a homolog of an aluminum-activated malate transporter. Plant Cell Physiol. 2010;51(3):354–365. Epub 2010/02/16. doi: 10.1093/pcp/pcq016 PubMed Central PMCID: PMC2835873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kinoshita T, Nishimura M, Shimazaki K. Cytosolic Concentration of Ca2+ Regulates the Plasma Membrane H+-ATPase in Guard Cells of Fava Bean. Plant Cell. 1995;7(8):1333–1342. doi: 10.1105/tpc.7.8.1333 PubMed Central PMCID: PMC160955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Ward JM, Schroeder JI. Calcium-Activated K+ Channels and Calcium-Induced Calcium Release by Slow Vacuolar Ion Channels in Guard Cell Vacuoles Implicated in the Control of Stomatal Closure. Plant Cell. 1994;6(5):669–683. doi: 10.1105/tpc.6.5.669 PubMed Central PMCID: PMC160467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Guse AH. Cyclic ADP-ribose: a novel Ca2+-mobilising second messenger. Cell Signal. 1999;11(5):309–316. Epub 1999/06/22. doi: 10.1016/s0898-6568(99)00004-2 [DOI] [PubMed] [Google Scholar]
- 54.Maheshwari P, Du H, Sheen J, Assmann SM, Albert R. Model-driven discovery of calcium-related protein-phosphatase inhibition in plant guard cell signaling. PLoS Comput Biol. 2019;15(10):e1007429. Epub 20191028. doi: 10.1371/journal.pcbi.1007429 PubMed Central PMCID: PMC6837631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hayashi M, Sugimoto H, Takahashi H, Seki M, Shinozaki K, Sawasaki T, et al. Raf-like kinases CBC1 and CBC2 negatively regulate stomatal opening by negatively regulating plasma membrane H. Photochem Photobiol Sci. 2020;19(1):88–98. doi: 10.1039/c9pp00329k . [DOI] [PubMed] [Google Scholar]
- 56.Swatek KN, Wilson RS, Ahsan N, Tritz RL, Thelen JJ. Multisite phosphorylation of 14-3-3 proteins by calcium-dependent protein kinases. Biochem J. 2014;459(1):15–25. Epub 2014/01/21. doi: 10.1042/BJ20130035 PubMed Central PMCID: PMC4127189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Brandt B, Brodsky DE, Xue S, Negi J, Iba K, Kangasjarvi J, et al. Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proc Natl Acad Sci U S A. 2012;109(26):10593–10598. Epub 2012/06/13. doi: 10.1073/pnas.1116590109 PubMed Central PMCID: PMC3387046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sierla M, Hõrak H, Overmyer K, Waszczak C, Yarmolinsky D, Maierhofer T, et al. The Receptor-like Pseudokinase GHR1 Is Required for Stomatal Closure. Plant Cell. 2018;30(11):2813–2837. Epub 20181025. doi: 10.1105/tpc.18.00441 PubMed Central PMCID: PMC6305979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Mulaudzi T, Ludidi N, Ruzvidzo O, Morse M, Hendricks N, Iwuoha E, et al. Identification of a novel Arabidopsis thaliana nitric oxide-binding molecule with guanylate cyclase activity in vitro. FEBS Lett. 2011;585(17):2693–2697. Epub 2011/08/09. doi: 10.1016/j.febslet.2011.07.023 [DOI] [PubMed] [Google Scholar]
- 60.Zhang J, Wang N, Miao Y, Hauser F, McCammon JA, Rappel WJ, et al. Identification of SLAC1 anion channel residues required for CO2/bicarbonate sensing and regulation of stomatal movements. Proc Natl Acad Sci U S A. 2018;115(44):11129–11137. Epub 20181009. doi: 10.1073/pnas.1807624115 PubMed Central PMCID: PMC6217375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Nagy SK, Darula Z, Kallai BM, Bogre L, Banhegyi G, Medzihradszky KF, et al. Activation of AtMPK9 through autophosphorylation that makes it independent of the canonical MAPK cascades. Biochem J. 2015;467(1):167–175. Epub 2015/02/04. doi: 10.1042/bj20141176 [DOI] [PubMed] [Google Scholar]
- 62.Desikan R, Griffiths R, Hancock J, Neill S. A new role for an old enzyme: nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. Proc Natl Acad Sci U S A. 2002;99(25):16314–16318. Epub 20021121. doi: 10.1073/pnas.252461999 PubMed Central PMCID: PMC138608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sokolovski S, Blatt MR. Nitric oxide block of outward-rectifying K+ channels indicates direct control by protein nitrosylation in guard cells. Plant Physiol. 2004;136(4):4275–4284. doi: 10.1104/pp.104.050344 PubMed Central PMCID: PMC535857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sirichandra C, Gu D, Hu HC, Davanture M, Lee S, Djaoui M, et al. Phosphorylation of the Arabidopsis AtrbohF NADPH oxidase by OST1 protein kinase. FEBS Lett. 2009;583(18):2982–2986. Epub 20090829. doi: 10.1016/j.febslet.2009.08.033 [DOI] [PubMed] [Google Scholar]
- 65.Acharya BR, Jeon BW, Zhang W, Assmann SM. Open Stomata 1 (OST1) is limiting in abscisic acid responses of Arabidopsis guard cells. New Phytol. 2013;200(4):1049–1063. Epub 2013/09/17. doi: 10.1111/nph.12469 [DOI] [PubMed] [Google Scholar]
- 66.Imes D, Mumm P, Bohm J, Al-Rasheid KA, Marten I, Geiger D, et al. Open stomata 1 (OST1) kinase controls R-type anion channel QUAC1 in Arabidopsis guard cells. Plant J. 2013;74(3):372–382. Epub 2013/03/05. doi: 10.1111/tpj.12133 [DOI] [PubMed] [Google Scholar]
- 67.Meyer S, Mumm P, Imes D, Endler A, Weder B, Al-Rasheid KA, et al. AtALMT12 represents an R-type anion channel required for stomatal movement in Arabidopsis guard cells. Plant J. 2010;63(6):1054–1062. doi: 10.1111/j.1365-313X.2010.04302.x [DOI] [PubMed] [Google Scholar]
- 68.Meinhard M, Grill E. Hydrogen peroxide is a regulator of ABI1, a protein phosphatase 2C from Arabidopsis. FEBS Lett. 2001;508(3):443–446. doi: 10.1016/s0014-5793(01)03106-4 [DOI] [PubMed] [Google Scholar]
- 69.Meinhard M, Rodriguez PL, Grill E. The sensitivity of ABI2 to hydrogen peroxide links the abscisic acid-response regulator to redox signalling. Planta. 2002;214(5):775–782.Epub 20011121. doi: 10.1007/s00425-001-0675-3 [DOI] [PubMed] [Google Scholar]
- 70.Sridharamurthy M, Kovach A, Zhao Y, Zhu JK, Xu HE, Swaminathan K, et al. H2O2 inhibits ABA-signaling protein phosphatase HAB1. PloS ONE. 2014;9(12):e113643. Epub 20141202. doi: 10.1371/journal.pone.0113643 PubMed Central PMCID: PMC4252038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006;45(1):113–122. Epub 2005/12/22. doi: 10.1111/j.1365-313X.2005.02615.x [DOI] [PubMed] [Google Scholar]
- 72.Zhang X, Wang H, Takemiya A, Song CP, Kinoshita T, Shimazaki K. Inhibition of blue light-dependent H+ pumping by abscisic acid through hydrogen peroxide-induced dephosphorylation of the plasma membrane H+-ATPase in guard cell protoplasts. Plant Physiol. 2004;136(4):4150–4158. doi: 10.1104/pp.104.046573 PubMed Central PMCID: PMC535845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Kohler B, Hills A, Blatt MR. Control of guard cell ion channels by hydrogen peroxide and abscisic acid indicates their action through alternate signaling pathways. Plant Physiol. 2003;131(2):385–388. doi: 10.1104/pp.016014 PubMed Central PMCID: PMC1540280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang RS, Saadatpour A, Albert R. Boolean modeling in systems biology: an overview of methodology and applications. Phys Biol. 2012;9(5):14. doi: 10.1088/1478-3975/9/5/055001 WOS:000309514900003. [DOI] [PubMed] [Google Scholar]
- 75.Kauffman SA. METABOLIC STABILITY AND EPIGENESIS IN RANDOMLY CONSTRUCTED GENETIC NETS. J Theor Biol. 1969;22(3):437. doi: 10.1016/0022-5193(69)90015-0 WOS:A1969D012600005. [DOI] [PubMed] [Google Scholar]
- 76.Naldi A, Carneiro J, Chaouiya C, Thieffry D. Diversity and plasticity of Th cell types predicted from regulatory network modelling. PLoS Comput Biol. 2010;6(9):e1000912. Epub 20100902. doi: 10.1371/journal.pcbi.1000912 PubMed Central PMCID: PMC2932677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Deritei D, Rozum J, Ravasz Regan E, Albert R. A feedback loop of conditionally stable circuits drives the cell cycle from checkpoint to checkpoint. Sci Rep. 2019;9(1):16430. Epub 20191111. doi: 10.1038/s41598-019-52725-1 PubMed Central PMCID: PMC6848090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Merilo E, Laanemets K, Hu H, Xue S, Jakobson L, Tulva I, et al. PYR/RCAR receptors contribute to ozone-, reduced air humidity-, darkness-, and CO2-induced stomatal regulation. Plant Physiol. 2013;162(3):1652–1668. Epub 20130523. doi: 10.1104/pp.113.220608 PubMed Central PMCID: PMC3707544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kwak JM, Mori IC, Pei ZM, Leonhardt N, Torres MA, Dangl JL, et al. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003;22(11):2623–2633. doi: 10.1093/emboj/cdg277 PubMed Central PMCID: PMC156772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Schulze S, Dubeaux G, Ceciliato PHO, Munemasa S, Nuhkat M, Yarmolinsky D, et al. A Role for Calcium-Dependent Protein Kinases in Differential CO2- and ABA-Controlled Stomatal Closing and low CO2-induced Stomatal Opening in Arabidopsis. New Phytol. 2021;229(5):2765–2779. Epub 20201209. doi: 10.1111/nph.17079 PubMed Central PMCID: PMC7902375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Negi J, Matsuda O, Nagasawa T, Oba Y, Takahashi H, Kawai-Yamada M, et al. CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells. Nature. 2008;452(7186):483–486. Epub 20080227. doi: 10.1038/nature06720 [DOI] [PubMed] [Google Scholar]
- 82.Jalakas P, Nuhkat M, Vahisalu T, Merilo E, Brosché M, Kollist H. Combined action of guard cell plasma membrane rapid- and slow-type anion channels in stomatal regulation. Plant Physiol. 2021;187(4):2126–2133. doi: 10.1093/plphys/kiab202 PubMed Central PMCID: PMC8644578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ando E, Kollist H, Fukatsu K, Kinoshita T, Terashima I. Elevated CO2 induces rapid dephosphorylation of plasma membrane H+-ATPase in guard cells. New Phytol. 2022;236(6):2061–2074. doi: 10.1111/nph.18472 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Neill SJ, Desikan R, Clarke A, Hancock JT. Nitric Oxide Is a Novel Component of Abscisic Acid Signaling in Stomatal Guard Cells. Plant Physiol. 2002;128(1):13–16. doi: 10.1104/pp.010707 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Leckie CP, McAinsh MR, Allen GJ, Sanders D, Hetherington AM. Abscisic acid-induced stomatal closure mediated by cyclic ADP-ribose. Proc Natl Acad Sci U S A. 1998;95(26):15837–15842. Epub 1998/12/23. PubMed Central PMCID: PMC28131. doi: 10.1073/pnas.95.26.15837 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zañudo JG, Albert R. An effective network reduction approach to find the dynamical repertoire of discrete dynamic networks. Chaos. 2013;23(2):025111. doi: 10.1063/1.4809777 [DOI] [PubMed] [Google Scholar]
- 87.Garg A, Di Cara A, Xenarios I, Mendoza L, De Micheli G. Synchronous versus asynchronous modeling of gene regulatory networks. Bioinformatics (Oxford, England). 2008;24(17):1917–1925. doi: 10.1093/bioinformatics/btn336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Naldi A, Hernandez C, Abou-Jaoudé W, Monteiro PT, Chaouiya C, Thieffry D. Logical Modeling and Analysis of Cellular Regulatory Networks With GINsim 3.0. Front Physiol. 2018:9. doi: 10.3389/fphys.2018.00646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Veliz-Cuba A, Jarrah AS, Laubenbacher R. Polynomial algebra of discrete models in systems biology. Bioinformatics. 2010;26(13):1637–1643. doi: 10.1093/bioinformatics/btq240 [DOI] [PubMed] [Google Scholar]
- 90.Cheng D, Qi H, Li Z. Analysis and Control of Boolean Networks: A Semi-tensor Product Approach. Springer London; 2010. [Google Scholar]
- 91.Su C, Pang J. CABEAN: a software for the control of asynchronous Boolean networks. Bioinformatics. 2020;37(6):879–881. doi: 10.1093/bioinformatics/btaa752 [DOI] [PubMed] [Google Scholar]
- 92.Deritei D, Aird WC, Ercsey-Ravasz M, Regan ER. Principles of dynamical modularity in biological regulatory networks. Sci Rep. 2016;6:21957. Epub 20160316. doi: 10.1038/srep21957 PubMed Central PMCID: PMC4793241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Wang R-S, Albert R. Elementary signaling modes predict the essentiality of signal transduction network components. BMC Syst Biol. 2011;5(1):44. doi: 10.1186/1752-0509-5-44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Yang G, Gómez Tejeda Zañudo J, Albert R. Target Control in Logical Models Using the Domain of Influence of Nodes. Front Physiol. 2018:9. doi: 10.3389/fphys.2018.00454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Saadatpour A, Albert R, Reluga TC. A Reduction Method for Boolean Network Models Proven to Conserve Attractors. SIAM J Appl Dyn Syst. 2013;12(4):1997–2011. doi: 10.1137/13090537X [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Jeon BW, Acharya BR, Assmann SM. The Arabidopsis heterotrimeric G-protein β subunit, AGB1, is required for guard cell calcium sensing and calcium-induced calcium release. Plant J. 2019;99(2):231–244. 10.1111/tpj.14318 [DOI] [PubMed] [Google Scholar]
- 97.Gayatri G, Agurla S, Raghavendra A. Nitric oxide in guard cells as an important secondary messenger during stomatal closure. Front Plant Sci. 2013:4. doi: 10.3389/fpls.2013.00425 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.He J, Zhang R-X, Peng K, Tagliavia C, Li S, Xue S, et al. The BIG protein distinguishes the process of CO2 -induced stomatal closure from the inhibition of stomatal opening by CO2. New Phytol. 2018;218(1):232–241. Epub 2018/01/02. doi: 10.1111/nph.14957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Lee M, Choi Y, Burla B, Kim YY, Jeon B, Maeshima M, et al. The ABC transporter AtABCB14 is a malate importer and modulates stomatal response to CO2. Nat Cell Biol. 2008;10(10):1217–1223. Epub 20080907. doi: 10.1038/ncb1782 [DOI] [PubMed] [Google Scholar]
- 100.Dittrich M, Mueller HM, Bauer H, Peirats-Llobet M, Rodriguez PL, Geilfus CM, et al. The role of Arabidopsis ABA receptors from the PYR/PYL/RCAR family in stomatal acclimation and closure signal integration. Nat Plants. 2019;5(9):1002–1011. Epub 20190826. doi: 10.1038/s41477-019-0490-0 [DOI] [PubMed] [Google Scholar]
- 101.Movahedi M, Zoulias N, Casson SA, Sun P, Liang Y-K, Hetherington AM, et al. Stomatal responses to carbon dioxide and light require abscisic acid catabolism in Arabidopsis. Interface Focus. 2021;11(2):20200036. doi: 10.1098/rsfs.2020.0036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Park KH, Costa FX, Rocha LM, Albert R, Rozum JC. Robustness of biomolecular networks suggests functional modules far from the edge of chaos. bioRxiv. 2023:2023.06.30.547297. doi: 10.1101/2023.06.30.547297 [DOI] [Google Scholar]
- 103.Zhu M, Jeon BW, Geng S, Yu Y, Balmant K, Chen S, et al. Preparation of Epidermal Peels and Guard Cell Protoplasts for Cellular, Electrophysiological, and -Omics Assays of Guard Cell Function. In: Botella JR, Botella MA, editors. Plant Signal Transduction: Methods and Protocols. New York, NY: Springer New York; 2016. p. 89–121. [DOI] [PubMed] [Google Scholar]
- 104.Wang H, Xiao W, Niu Y, Chai R, Jin C, Zhang Y. Elevated Carbon Dioxide Induces Stomatal Closure of Arabidopsis thaliana (L.) Heynh. Through an Increased Production of Nitric Oxide. J Plant Growth Regul. 2015;34(2):372–380. doi: 10.1007/s00344-014-9473-6 [DOI] [Google Scholar]
- 105.Leung J, Merlot S, Giraudat J. The Arabidopsis ABSCISIC ACID-INSENSITIVE2 (ABI2) and ABI1 genes encode homologous protein phosphatases 2C involved in abscisic acid signal transduction. Plant Cell. 1997;9(5):759–771. doi: 10.1105/tpc.9.5.759 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(DOCX)
(XLSX)
(XLSX)
(DOCX)
(DOCX)
(XLSX)
Data Availability Statement
All relevant data are provided in the paper, its Supporting Information files, and the GitHub repository https://github.com/jack802gan/CO2-model. DOI: https://zenodo.org/doi/10.5281/zenodo.10665442.