

Abstract
Several cat breeds are defined by morphological variation of the tail. The Japanese Bobtail is a breed that has been accepted for registration only within the past 50 years; however, the congenital kinked tail variants defining this breed were documented in the Far East centuries ago and the cats are considered ‘good luck’ in several Asian cultures. The recent discovery of the mutation for the tailless Manx phenotype has demonstrated that the Japanese Bobtail does not have a causative mutation in the same gene (T-Box). Here, a simple segregation analysis of cats bred from a pedigreed Japanese Bobtail demonstrated a simple autosomal dominant mode of inheritance with variable expression of the tail length and kink placement. Unexpectedly, radiological examinations of the entire vertebral column of kink-tailed cats indicated variation from the normal vertebral feline formula (C7, T13, L7, S3, Cd20–24), including cats with mostly one reduction of thoracic vertebrae (C7, T12, L7, S3), and an average of 15.8 caudal vertebrae. A few cats had variation in the number of cervical vertebrae. Several transitional vertebrae and anomalous ribs were noted. One cat had a bifid vertebra in the tail. Most cats had hemivertebrae that were usually included in the tail kink, one of which was demonstrated by gross pathology and histopathology. The abnormal vertebral formula or the placement of the kink in the tail did not coincide with morbidity or mortality.
Short Communication
Axial skeletal development is the foundation of vertebrates. The domestic cat can be an important model for spinal cord and tail development when considering the different breeds that have no tail (anury) or shortened tails (brachyury).1–3 The bobtail of the Japanese Bobtail breed is suggested to be inherited as an incomplete dominant single gene trait; however, the tail variation has not been scientifically documented. We examine the inheritance of the bobtail trait in the Japanese Bobtail, demonstrating mode of inheritance and variation in the vertebral formulae of cats with kinked tailed phenotypes.
One female Japanese Bobtail cat was donated for breeding trials (Figure 1). Housing and husbandry was approved by the institutional animal care and use committee (protocol 16691). The female Japanese Bobtail was mated to a Persian and Bengal – breeds with normal, straight tails but different body morphologies – producing 10 kittens (Table 1; Figure 2). The parentage of breedings was confirmed by DNA typing, as previously described. 4 Tail phenotypes were determined by visual inspection and palpation. All first-generation offspring (F1) had variable tail abnormalities. The donated cat was suggested to be homozygous for the tail trait by the breeder, which was supported by the breeding data as all 10 F1 cats had a tail abnormality (Table 1; Figure 2). Three F1 and one second-generation offspring were mated with straight-tailed cats that did not have Japanese Bobtail progenitors, producing 37 kittens (Table 1; Figure 2). Two kittens were stillborn males and unavailable for tail evaluation owing to consumption by the queen. Fifteen of 35 offspring had kinked or shortened tails (43%), indicating no significant deviation from an autosomal dominant mode of inheritance (χ2 = 0.714, 1 df; P = 0.40). The breeding of the male F1 produced two normal-tailed females and six normal-tailed males, excluding X- or Y-linked inheritance. No sex bias was observed (χ2 = 0.191, 1 df; P = 0.66). Litter sizes were variable, ranging from one to seven kittens (average is four per litter), thus reduced litter sizes were not suggested. However, no bobtail–bobtail crosses were conducted so that the potential of in utero loss of homozygous bobtailed kittens was not evaluated. However, in pedigreed cats, the bobtail trait is considered to be fixed and homozygous. Breeders normally breed bobtailed to bobtailed cats. The breedings support an autosomal dominant mode of inheritance with variable expression.
Figure 1.

The Japanese Bobtail breed. Show-quality tails of the Japanese Bobtail must be no longer than 3 inches. Most cats are expected to be homozygous for the bobtail mutation. The high white ‘Mi-Ke’ coloration (left) and black and white (right) are highly popular. (Images courtesy of Richard Katris – Chanan Photography)
Table 1.
Simple segregation analysis of the bobtail kinked tail trait in cats
| Breeding* | Sex | Bobtail | Non-bobtail | Unknown | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | Male | Female | Kittens | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ | ♂ | ♀ |
| NN × AA | 6678 | 11662 | 6 | 4 | 2 | 0 | 0 | 0 | 0 | 4 | 2 |
| NN × AA | 4445 | 11662 | 4 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 4 |
| Total F1 kittens | 4 | 6 | 0 | 0 | 0 | 0 | 4 | 6 | |||
| NN × Aa | 11896 | 12173 | 6 | 0 | 1 | 4 | 1 | 0 | 0 | 4 | 2 |
| NN × Aa | 13380 | 12173 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 |
| NN × Aa | 10456 | 13230 | 11 (2 kittens) | 4 | 2 | 2 | 2 | 1 | 0 | 7 | 4 |
| NN × Aa | 13380 | 13230 | 4 | 1 | 0 | 0 | 2 | 0 | 0 | 1 | 2 |
| NN × Aa | 11894 | 16527 | 3 | 2 | 0 | 0 | 0 | 1 | 0 | 3 | 0 |
| Aa × NN | 13227 | 15240 | 7 (2 litters) | 0 | 4 | 2 | 1 | 0 | 0 | 2 | 5 |
| Aa × NN | 13227 | 13225 | 6 | 0 | 1 | 4 | 1 | 0 | 0 | 4 | 2 |
| Total second-generation kittens | 7 | 8 | 12 | 8 | 2 | 0 | 21 | 16 | |||
| Total kittens | 11 | 14 | 12 | 8 | 2 | 0 | 25 | 22 | |||
Breeding genotypes (sire × queen): NN is homozygous normal tail (wild-type), AA is homozygous bobtail, Aa is heterozygous bobtail. χ2 = 0.714 (P = 0.40) for bobtail vs non-bobtail of the second-generation offspring. χ2 = 0.676 (P = 0.411) for male vs female of the second-generation offspring. χ2 = 0.191 (P = 0.66) for male vs female for all offspring. Heterozygous matings (Aa) were not available for evaluation
Figure 2.

Pedigree of cats segregating for the Japanese Bobtail tail phenotype. Squares are males, circles are females, diamonds are unknown. Cats with filled symbols have the kinked tail trait from the Japanese Bobtail; open symbols indicate a normal, straight (wild-type) tail. Arrow indicates the proband homozygous pedigreed Japanese Bobtail. Small symbols are stillborn kittens with unknown tail type; symbols with a slash indicate dead cats. Cats with a lower left filled circle were radiographed. Cat with the ‘H’ (13229) was submitted for pathology of the tail
Right lateral and ventrodorsal radiographs of the pelvis and tail were obtained from 24 cats, including three offspring with normal tails, and reviewed by a board-certified radiologist (REP) (Table 2). Twenty-one of 24 cats also underwent right lateral radiograph of the cervical, thoracic and lumbar spine (Table 2). All 17 kinked-tailed cats for which the entire spine was radiographed had abnormal vertebral formulae. Seven cats had block vertebrae and one cat had a bifid vertebra at Cd7.
Table 2.
Radiographic vertebral determination of kinked-tail phenotype of Japanese Bobtail cats
| Cd vertebrae in kink | Transitional vertebrae |
Cd block vertebrae | Cd hemivertebrae | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ID | C | T | L | S | Cd | C–T | S–Cd (disk space) | |||
| 16626 | 7 | 13 | 7 | 3 | 20 | Not applicable | None | S2–S3 | None | None |
| 17804 | 7 | 13 | 7 | 3 | 22 | Not applicable | None | S2–S3 | None | None |
| 17800 | 7 | 13 | 7 | 3 | 21 | Not applicable | None | None | None | None |
| 13230 | 7 | 12 | 7 | 3 | 15 | 6, 10 | Rib: C7 | S2–S3 | 11–12, 14–15 | 6, 10 |
| 13227 | 7 | 12 | 7 | 3 | 18 | 9, 14, 16–18 | None | None | 16–18 | 9, 14 |
| 14055 | 7 | 12 | 7 | 3 | 18 | 14–16 | None | None | None | 9, 14 |
| 12173 | 7 | 12 | 7 | 3 | 12 | 6–8, 10–12 | Rib: C7, L1 | None | 6–8, 10–12 | None |
| 13232 | 7 | 12 | 7 | 3 | 16 | 7, 11, 13, 15 | Rib: C6, C7 | None | None | 7, 13 |
| 13231 | 7 | 12 | 7 | 3 | 15 | 9, 11 | None | None | 13–15 | 9, 11 |
| 16625 | 7 | 12 | 7 | 3 | 20 | 19–20 | Ribs:C7, sacralized L7 | None | None | None |
| 16628 | 7 | 12 | 7 | 3 | 11 | 7–8, 10–11 | None | None | 7–8 | None |
| 16528* | 7 | 12 | 7 | 3 | 13 | 7, 9, 11–13 | None | S2–S3 | 7–8 | 7, 9, 11, 13 |
| 17801 | 7 | 12 | 7 | 3 | 12 | 5–8 | None | S2–S3 | 6–7 | 8 |
| 17796 | 7 | 12 | 7 | 3 | 18 | 6–8, 16 | None | S1–S2 | None | 6, 8, 16 |
| 17799 | 7 | 12 | 7 | 3 | 19 | 5–7, 10–13, 15, 17 | None | S2–S3 | None | 5–7, 10–13, 15, 17 |
| 17803 | 7 | 12 | 7 | 3 | † | † | None | None | None | † |
| 16627 | 7 | 12 | 7 | 3 | 15 | 9, 15 | Rib: C7 | S2–S3 | None | 9 |
| 17802 | 6 | 13 | 7 | 3 | 21 | 15 | None | None | None | 15 |
| 17805 | 6 | 13 | 7 | 3 | 14 | 4–6, 9 | None | S2–S3 | None | 4, 6, 9 |
| 16527 | 6 | 13 | 6 | 3 | 19 | 13, 17–19 | None | None | None | 13, 17, 18 |
| 12175 | 6 | 3 | 12 | 6–12 | None | S2–S3 | None | None | ||
| 12172 | 3 | 17 | 11–17 | None | None | None | 4, 9 | |||
| 12174 | 3 | 13 | 13–15 | None | S1–S2 | None | 13, 15 | |||
Bifid vertebra at Cd7
Tail very short and superimposed over pelvis with lots of anomalous vertebrae; vertebrae could not be accurately counted
Fourteen of 17 (82.4%) kinked-tailed cats had the vertebral formula C7, T12, L7, S3, Cd11–20 (Figures 3 and 4). Eight cats (47%) had hemivertebra involving the caudal vertebrae. The number of hemivertebrae ranged from two to nine, and the location varied. Seven cats (50%) had block vertebra all involving the caudal vertebrae. Five of 14 cats (36%) had transitional cervicothoracic regions (ribs on C7). One had a transitional thoracolumbar junction (rib on L1). One had a transitional lumbosacral junction (sacralization of L7). Eight of 14 (57%) had a transitional sacrocaudal region (visible disk space between S2 and S3).
Figure 3.

Kinked-tail phenotypic variation in cats with the Japanese Bobtail trait. Lateral and ventral radiographic views and tail photographs of the same cats demonstrating variation of the kinked-tail phenotype in individuals derived from a pedigreed Japanese Bobtail cat. (a), (d) and (i) represent the same cat; (b), (e) and (j) represent the same cat; (e), (g) and (k) represent the same cat; and (d), (h) and (l) represent the same cat
Two of 17 (12%) kinked-tailed cats had the vertebral formula C6, T13, L7, S3, Cd14–21. Both had hemivertebrae (1–3) involving the caudal vertebrae. Neither cat had a block vertebra. Neither cat had transitional segments at the cervicothoracic, thoracolumbar or lumbosacral junctions. One had a transitional sacrocaudal region (visible disk space between S2 and S3).
One kinked-tailed cat had the vertebral formula of C6, T13, L6, S3, Cd19 (Figure 4). This cat had three hemivertebrae involving Cd13, Cd17 and Cd18, but no block vertebrae and no transitional segments.
Figure 4.
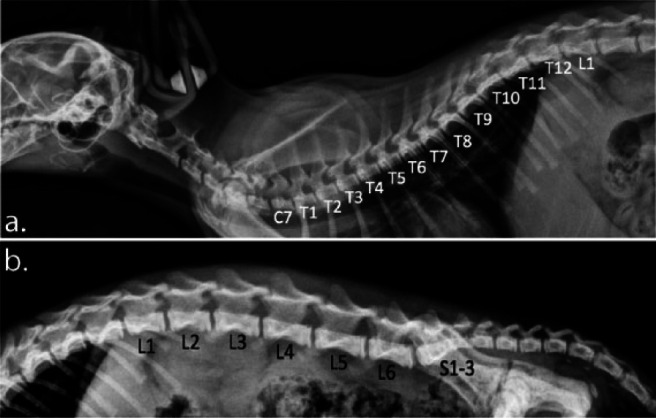
Lateral radiographs of the cervical and thoracic spine of cats with kinked tails from the Japanese Bobtail trait. (a) Unlike a normal, wild-type-tailed cat that has 13 thoracic vertebrae, this cat has only 12 thoracic vertebrae (T1–12). This cat has seven cervical vertebrae and seven lumbar vertebrae (not all included in this study) like a tailed cat. (b) This cat has only six lumbar vertebrae (L1–6). The sacrum is normal consisting of three fused segments (S1–3)
Three of 20 kinked-tailed cats underwent radiography of the pelvis and tail only. The visible vertebral formula for these cats was S3, Cd12–18. Two had hemivertebra (two each) involving the caudal vertebrae. There were no block vertebrae. None had transitional segments at the lumbosacral junctions but all three had a transitional sacrocaudal region (visible disk space between S1 and S2 [n = 1] or S2 and S3 [n = 2]).
Three cats with straight tails underwent radiography of the entire spine. All cats had a normal vertebral formula (C7, T13, L7, S3 and Cd20–24). 5 No block vertebrae, hemivertebra or bifid vertebra in any portion of the vertebral column, and no transitional segments in the cervicothoracic, thoracolumbar or lumbosacral regions, were observed. Two straight-tailed cats had a transitional sacrocaudal junction (a radiographically visible disk space between S2 and S3).
One F1 cat died of unrelated causes and was submitted to necropsy for histopathology of the kinked tail (Figure 5). The affected regions of tail were immersed in 10% neutral buffered formalin for fixation followed by demineralization in a 15% formic acid solution. These regions were longitudinally sectioned, processed routinely and embedded in paraffin. Tissue sections (4 μm thick) were stained with hematoxylin and eosin. The clinically described scoliosis of the tail was due to three coccygeal hemivertebrae. Affected vertebrae were shortened and misshapen but were otherwise normal. The affected vertebrae and adjacent soft tissues lacked histologic evidence of degeneration or other abnormalities, consistent with a congenital abnormality.
Figure 5.

Gross pathology and histopathology of a cat tail with the Japanese Bobtail kinked tail phenotype (cat 13229). (a) Photograph of the tail, caudal abdomen and pelvic limbs. Abnormal lateral curvature of the tail is present. (b) Longitudinal section of formalin-fixed and demineralized tail. Three vertebral bodies, including one small, wedge-shaped hemivertebra, are evident. A single misshapen intervertebral disk space bridges the three vertebrae; ruler = 1 cm. (c) Subgross histologic section from the same region. Note the normal lamellar bone of the vertebral bodies and the intact, but misshapen intervertebral disk. Haematoxylin and eosin stain; bar = 1 cm
Historical paintings and wooden carvings from the Far East, such as Tokyo’s Gotokuji Temple, constructed in 1697, depict cats with shortened, curled tails. The bobtailed cats are thought to have first arrived in Japan from China or Korea, arriving with the Buddhist monks in 600–700 AD. Although commonly known for centuries, the bobtailed Japanese cat has developed into a defined breed in the USA within the past 50 years. The Cat Fanciers’ Association accepted the breed with provisional status in 1971 and for championship competition in 1976. 6 Although the breed remains rare, several registries throughout the world have since accepted the Japanese Bobtail.
The Japanese Bobtail is a medium-build cat with a well-muscled, slender body. The moderately dolichocephalic head is distinct from the other Oriental breeds (Figure 1) and the three-color, high white spotting pattern (‘Mi-Ke’) is popular (Figure 1, left). The tail is supposedly unique to each individual. For competition, the tail must be clearly visible and is composed of one or more curves, angles or kinks, or any combination thereof. The Japanese Bobtail is considered a natural breed and has allowable outcrosses with cats from Japan depending on the specific cat registry regulations. 6 Previous studies have used microsatellites (short tandem repeats) and single nucleotide polymorphisms to demarcate genetically breeds and their relationships to random-bred populations.7–9 Despite the Japanese Bobtail’s suggested origins of the Far East, these studies indicate that the origins cannot be clearly identified, suggesting the breed may represent a hybridization of Eastern and Western cats.
Domestic cats have several well-established breeds defined by single gene structural mutations, such as taillessness of the Manx breed.10–13 The inheritance and associated health concerns of taillessness are well documented, making the breed controversial.14–16 Several genetic mutations in T-Box are identified for taillessness in Manx cats and other species, 17 including dogs and mice.18,19 The Japanese, American and Kurilian Bobtails examined in the Manx study did not have T-Box mutations, suggesting that an additional gene(s) exists in cats that controls tail and vertebrae formation. In addition, many tailless Manx do not have health complications, suggesting genetic modifiers of the T-box mutations that could be positively selected within the population. The locus symbol, BTADJ, is suggested to represent this autosomal dominant kinked tail mutation of the Japanese Bobtail. Other breeds may share variants at this locus, have different variants or have other causative genes.
The Tailless locus of Manx cats is associated with many health detriments in cats, such as incontinence, lameness and spina bifida with myelomeningocele. Radiography was used in a subset of cats to define the abnormalities of the vertebral column non-invasively, documenting the extent of the variable expression of the phenotype. Almost all cats with kinked tails had reduced numbers of caudal vertebrae, ranging from 11–21, and averaging 16. The reduction in caudal vertebrae was less pronounced than the Manx, where caudal vertebral numbers range from 0–23. 20 All kinked-tailed cats had several caudal vertebrae involved in the kink and 12/20 (60%) cats had at least one hemivertebra included in the kinks. The tail kinks were almost all remote to the sacrum, thereby reducing the likelihood of concurrent neurologic deficits.
Vertebral anomalies remote to the sacrum have been well described in cats and are frequently identified incidentally during radiographic examination for unrelated reasons. 21 Transitional vertebrae at the cervicothoracic or thoracolumbar junction are not associated with clinical signs nor are reduced or increased numbers of vertebrae at any location in the spine. 22 However, the identification of anomalous vertebral numbers or transitional segments is clinically important when planning for surgical intervention in the spine as surgical site selection is often based on the location of the last rib.
Unexpectedly, besides abnormalities in the caudal vertebra, all kinked-tailed cats had abnormal skeletal vertebral formulae with reduced vertebral numbers. Moreover, a large number of transitional vertebrae were identified in the bobtailed cats described. When compared with previously published data in a random assortment of tailed cats,24 the number of kinked-tailed cats in this study with reduced vertebral numbers in the cervical (18% vs 0%) and thoracic region (64% vs 1%) was quite high. Similarly, the number of cats with transitional segments at the cervicothoracic (36% vs 1%) and sacrocaudal junctions (45% vs 10%) was also high, implying that the kinked tail is associated with variations in the vertebral formula and vertebral transitionality. Although clearly a single gene, the variation in presentation is substantial and warrants the consideration of genetic modifiers. Determination of the causative gene will shed light on the evolution and development of the basic body plan of all vertebrates. Ancedotal comments from Pixie-Bob breeders have also noted cats with missing thoracic verterbrae, suggesting some Pixie-Bob may be of Japanese Bobtail origin or have a DNA variant in the same gene. Bobtail breeders report that pedigreed bobtails, which are likely homozygous for the mutation, also have a wide and unpredictable range in tail presentation. A comparison of the range in the tail presentation between homozygous and heterozygous cats will be possible once the mutation is identified. From a clinical standpoint, knowing the number of vertebrae and location of ribs is important when planning surgical approaches in animals with neurologic dysfunction. However, transitional vertebrae and reduced cervical and thoracic vertebral numbers were not associated with significant morbidity in this group of cats.
Conclusions
The bobbed tail of the Japanese Bobtail breed has autosomal dominant inheritance with variable expression. Besides a reduced number of caudal vertebrae, the trait is associated with caudal hemivertebrae, abnormal vertebral formulae and the presence of transitional vertebrae. Cats were generally adopted or maintained in the colony, and no health concerns have been reported in cats, which are, to date, all aged 5 years or younger.
Acknowledgments
The efforts of radiography technicians Jennifer Lazzari, Michele Santoro and Linda Savely were invaluable. We thank the Japanese Bobtail owners and breeders who so generously contributed time to this study, especially Jennifer Reding and Nikki Crandall-Seibert for their assistance with cats, additional radiographs and commentary on the breeds. We appreciate the support of Erica Creighton with the pedigree and image construction. We appreciate the donation of cat food by Royal Canin USA, Inc (St Charles, MO, USA).
Footnotes
Supplementary material: Radiographs from this project are available from the corresponding author upon request.
The authors do not have any potential conflicts of interest to declare.
Funding: This work was supported, in part, by funding from the National Center for Research Resources R24 RR016094 and is currently supported by the Office of Research Infrastructure Programs/OD R24OD010928, the Winn Feline Foundation (W10-014 and W10-015), and the George and Phyllis Miller Feline Health Fund, Center for Companion Animal Health, School of Veterinary Medicine, University of California, Davis (2008-36-F).
Accepted: 22 July 2014
References
- 1. Kitchen H, Murray R, Cockrell B. Animal model for human disease. Spina bifida, sacral dysgenesis and myelocele. Animal model: Manx cats. Am J Pathol 1972; 68: 203. [PMC free article] [PubMed] [Google Scholar]
- 2. Green ST, Green FA. The Manx cat: an animal model for neural tube defects. Mater Med Pol 1987; 19: 219–221. [PubMed] [Google Scholar]
- 3. Woodside J, Dail W, McGuire E, et al. The Manx cat as an animal model for neurogenic vesical dysfunction associated with myelodysplasia: a preliminary report. J Urol 1982; 127: 180–183. [DOI] [PubMed] [Google Scholar]
- 4. Lipinski M, Amigues Y, Blasi M, et al. An international parentage and identification panel for the domestic cat (Felis catus). Anim Genet 2007; 38: 371–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Thrall DE. Textbook of veterinary diagnostic radiology. 5th ed. St Louis, MO: Saunders Elsevier, 2012, p 684. [Google Scholar]
- 6. The Cat Fanciers’ Association. Japanese Bobtail. http://www.cfainc.org/Breeds/BreedsCJ/JapaneseBobtail.aspx (accessed 27 February 2014).
- 7. Kurushima JD, Lipinski MJ, Gandolfi B, et al. Variation of cats under domestication: genetic assignment of domestic cats to breeds and worldwide random-bred populations. Anim Genet 2013; 44: 311–324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lipinski MJ, Froenicke L, Baysac KC, et al. The ascent of cat breeds: genetic evaluations of breeds and worldwide random-bred populations. Genomics 2008; 91: 12–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Menotti-Raymond M, David VA, Pflueger SM, et al. Patterns of molecular genetic variation among cat breeds. Genomics 2008; 91: 1–11. [DOI] [PubMed] [Google Scholar]
- 10. Todd N. The Manx factor in domestic cats: a possible genetic basis for expressivity of taillessness and other associated anomalies. J Hered 1964; 55: 225. [PubMed] [Google Scholar]
- 11. Todd NB. The inheritance of taillessness in Manx cats. J Hered 1961; 52: 228–232. [Google Scholar]
- 12. Howell JM, Siegel PB. Morphological effects of the Manx factor in cats. J Hered 1966; 57: 100–104. [DOI] [PubMed] [Google Scholar]
- 13. Howell JM, Siegel PB. Phenotypic variability of taillessness in Manx cats. J Hered 1963; 54: 167–169. [DOI] [PubMed] [Google Scholar]
- 14. James CC, Lassman LP, Tomlinson BE. Congenital anomalies of the lower spine and spinal cord in Manx cats. J Pathol 1969; 97: 269–276. [DOI] [PubMed] [Google Scholar]
- 15. Martin A. A congenital defect in the spinal cord of the Manx cat. Vet Path Online 1971; 8: 232–238. [DOI] [PubMed] [Google Scholar]
- 16. Tomlinson B. Abnormalities of the lower spine and spinal cord in Manx cats. J Clin Pathol 1971; 24: 480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Buckingham KJ, McMillin MJ, Brassil MM, et al. Multiple mutant T alleles cause haploinsufficiency of Brachyury and short tails in Manx cats. Mamm Genome 2013; 24: 400–408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hytönen MK, Grall A, Hédan B, et al. Ancestral T-box mutation is present in many, but not all, short-tailed dog breeds. J Hered 2009; 100: 236–240. [DOI] [PubMed] [Google Scholar]
- 19. Kavka AI, Green JB. Tales of tails: Brachyury and the T-box genes. Biochim Biophys Acta 1997; 1333: F73–F84. [DOI] [PubMed] [Google Scholar]
- 20. Deforest ME, Basrur PK. Malformations and the Manx syndrome in cats. Can Vet J 1979; 20: 304–314. [PMC free article] [PubMed] [Google Scholar]
- 21. Newitt A, German AJ, Barr FJ. Congenital abnormalities of the feline vertebral column. Vet Radiol Ultrasound 2008; 49: 35–41. [DOI] [PubMed] [Google Scholar]
- 22. Morgan JP. Congenital anomalies of the vertebral column of the dog: a study of the incidence and significance based on a radiographic and morphologic study. J Am Vet Radiol Soc 1968; 9: 21–29. [Google Scholar]