

Abstract
Stem cells undergo partitioning through mitosis and separate into specific cells of each of the three embryonic germ layers: endoderm, mesoderm, and ectoderm. Pluripotency, reprogramming, and self-renewal are essential elements of embryonic stem cells (ESCs), and it is becoming evident that regulation of protein degradation mediated by the ubiquitin–proteasome system (UPS) is one of the key cellular mechanisms in ESCs. Although the framework of that mechanism may seem simple, it involves complicated proteolytic machinery. The UPS controls cell development, survival, differentiation, lineage commitment, migration, and homing processes. This review is centered on the connection between stem cell factors NANOG, OCT-3/4, SOX2, KLF4, C-MYC, LIN28, FAK, and telomerase and the UPS. Herein, we summarize recent findings and discuss potential UPS mechanisms involved in pluripotency, reprogramming, differentiation, and self-renewal. Interactions between the UPS and stem cell transcription factors can apply to various human diseases which can be treated by generating more efficient iPSCs. Such complexes may permit the design of novel therapeutics and the establishment of biomarkers that may be used in diagnosis and prognosis development. Therefore, the UPS is an important target for stem cell therapeutic product research.
Keywords: Deubiquitinating enzyme, Deubiquitination, E3 ligase, Post-translational modification, Ubiquitination
Introduction
Stem cells are undifferentiated cells that can undergo self-renewal. They undergo partitioning through mitosis and separate into specific cells of each of the three embryonic germ layers: endoderm, mesoderm, and ectoderm (Fig. 1) [1, 2]. Embryonic stem cells (ESCs) are not transformed, rather, they are pluripotent cells that are derived from the inner cell mass (ICM) of the mammalian blastocyst [3]. In 2006, four transcriptional factors, OCT-3/4, SOX2, KLF4, and C-MYC (referred to as Yamanaka factors), were identified. These four factors regulate pluripotency and self-renewal activity of stem cells and stimulate the formation of induced pluripotent stem cells (iPSCs) in mouse embryonic or adult fibroblasts [4]. The iPSCs are generated from mouse and human somatic cell types by enforcing the expression of transcription factors [5]. Post-translational modifications (PTMs) control the activity, interaction, subcellular localization, and stability of their target proteins. Ubiquitination is a type of PTMs mediated by ubiquitin ligases and deubiquitinating enzymes (DUBs), which regulate cellular functions and stability of proteins. Ubiquitination is a multi-step reaction involved in protein degradation via the 26S proteasome. The 26S proteasome is a large multi-catalytic/multi-subunit protease complex that is a part of the ubiquitin–proteasome system (UPS). The UPS is an essential system for extra-lysosomal cytosolic and nuclear protein degradation; additionally, it has various biological functions [6]. The 26S proteasome is involved in protein degradation and proteolytic cellular processes such as signal transduction, proliferation, differentiation, cell cycling, inflammation, gene transcription, development, senescence, antigen presentation, apoptosis, and stress responses. Ubiquitin is a highly conserved 76-amino acid polypeptide that has seven lysine residues: K6, K11, K27, K29, K33, K48, and K63. The K48-linked polyubiquitination chain mainly has an essential role in targeting proteins for 26S proteasomal degradation. On the other hand, the K63-linked polyubiquitination chain regulates intercellular signaling [4]. Ubiquitin is catalyzed in three enzymatic steps via ubiquitin-activating enzymes (E1), ubiquitin-conjugating enzymes (E2), and ubiquitin-protein ligases (E3). DUBs deconjugate ubiquitin from the substrate. Harmony between ubiquitination and deubiquitination controls stem cell destiny. Cross-regulation between E3 ligases and DUBs for stem cell transcription factors is important in regulation of stem cell function including pluripotency, differentiation, and self-renewal. E3 ligases ubiquitinate stemness-related proteins to regulate stem cell differentiation by attaching ubiquitin molecules to substrates, whereas DUBs maintain stemness-related proteins to prevent stem cell differentiation by removing ubiquitin molecules or the other way around [7]. Therefore, E3 ligases negatively regulate the roles of stem cell transcription factors, and DUBs positively regulate stem cell transcription factors or vice versa at the post-transcriptional level (Fig. 2). The UPS is involved in the pathogenesis of various human diseases, including cancer, genetic, autoimmune, neurodegenerative, and metabolic disorders. Therefore, the UPS is an important target for therapeutic product research [6].
Fig. 1.
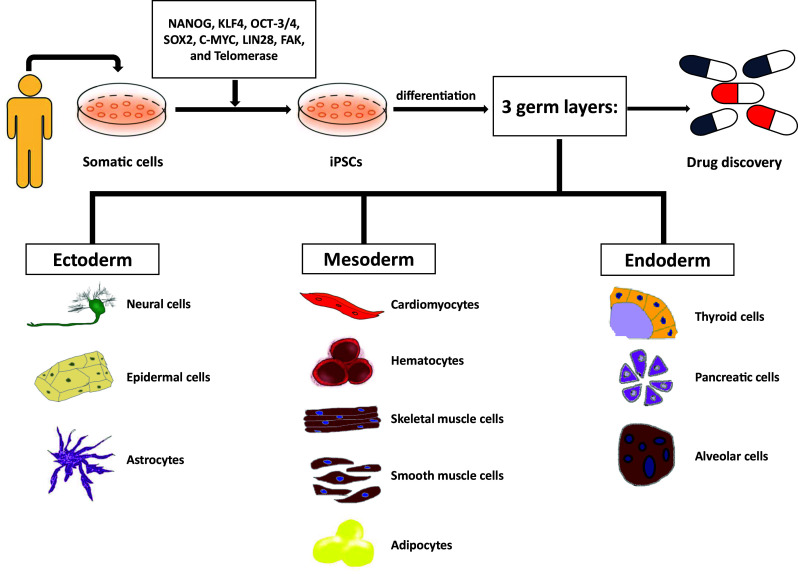
Reprogramming somatic cells into iPSCs. Differentiation process of stem cells from somatic cells to iPSCs is regulated by several kinds of reprogramming factors such as NANOG, KLF4, OCT-3/4, SOX2, C-MYC, LIN28, FAK, and telomerase
Fig. 2.

Interaction of the stem cell transcription factors with E3 ligases and DUBs. Various DUBs and E3 ligases regulate stem cell transcription factors including NANOG, KLF4, OCT-3/4, SOX2, C-MYC, LIN28, FAK, and telomerase. By deubiquitination, stem cell transcription factors regulate stem cell maintenance. Contrary to this, stem cell transcription factors regulate stem cell differentiation by ubiquitination
NANOG
The homeobox transcription factor NANOG has essential roles in the regulation of self-renewal and pluripotency of ESCs. Stabilization of NANOG is important for maintaining stemness of ESCs [8]. A study published in 2003 first showed NANOG to be a critical factor in the pluripotency of the ICM and ESCs. A NANOG-deficient ICM unsuccessfully created epiblasts, only delivering parietal endoderm-like cells [9]. NANOG has a relatively short half-life and is quickly degraded by the UPS. This means that during the developmental process, regulation of protein degradation determines cell fate. Another research group and we have demonstrated that the UPS regulates the expression level, stability, and activity of NANOG, and that the proline (P), glutamic acid (E), serine (S), threonine (T) (PEST) motif is important for the ubiquitination of the K48-type ubiquitin linkage [10]. F-box/WD repeat-containing protein 8 (FBXW8) is a widely known E3 ligase of NANOG that is promoted by activation of ERK1 phosphorylation. FBXW8 induces polyubiquitination and reduces NANOG protein stability resulting in differentiation of ESCs. The NANOG protein and phosphorylated ERKs locate in the nucleus and cytoplasm; phosphorylated ERKs translocate to the nucleus and accumulate during ESC differentiation. During ESC differentiation, ubiquitination of NANOG and phosphorylation of ERKs are induced. PD98059, an ERK inhibitor, decreases ubiquitination of NANOG [11]. USP21, a DUB for NANOG, was identified during yeast two-hybrid screening in our laboratory [4], as well as being identified by luciferase reporter assays [8] and DUB screening performed by two other groups [9]. USP21 deubiquitinates and stabilizes NANOG in ESCs, and this stabilization maintains the protein level of NANOG, controlling pluripotency. Our studies indicate that downregulation of USP21 results in degradation of NANOG and differentiation of ESCs. Recent study found out that downregulation of USP34 induced stemness of stem cell and increased the level of NANOG [12]. Thus, the interaction between the UPS and NANOG will have a synergistic effect on cell-based therapy.
OCT-3/4
Octamer-binding transcription factor 3/4 (OCT-3/4) was first reported 26 years ago and is a Pit–Oct–Unc (POU) transcription factor that regulates the fate of pluripotent ESCs and has an essential role in ESC reprogramming [13, 14]. OCT-3/4 has two specific domains, and it can form both heterodimers and homodimers. OCT-3/4 forms heterodimers with other transcription factors, and it forms homodimers within the DNA motif, depending on the one of several unique configurations of the octamer half-sites [15]. The POU domain is required for OCT-3/4 cytoplasmic localization [16]. OCT-3/4 can be replaced by orphan nuclear receptor Nr5a2 for the reprogramming function because Nr5a2 enhances the efficiency of reprogramming [17, 18]. When OCT-3/4 is overexpressed, stem cell differentiates into endoderm and mesoderm; however, when OCT-3/4 expression is downregulated, it undergoes differentiation into trophectoderm [19]. Ubiquitination of OCT-3/4 is directly regulated by the homologous to E6-AP carboxyl terminus (HECT)-type E3 ubiquitin ligase WWP2, which rapidly regulates protein degradation in ESCs with a short half-life. Significantly, OCT-3/4 expression is elevated with knockdown of WWP2 expression in differentiated ESCs. Single ubiquitin conjugation with OCT-3/4 inactivates transcriptional activity, while polyubiquitination, specifically the K63-linked polyubiquitination chain, decreases the OCT-3/4 level [20]. ITCH, an E3 ligase, belongs to the HECT family. It negatively regulates OCT-3/4 activity and maintains differentiated ESC self-renewal and pluripotency. Even though WWP2 and ITCH belong to the same family, they target differently to OCT-3/4. WWP2 mediates when OCT-3/4 is in full-length; on the other hand, ITCH targets the OCT-3/4 DNA binding domain [21]. RNF2 is a ring finger E3 ligase that interacts with OCT-3/4 and maintains stem cell pluripotency. This E3 ligase targets K6-, K27-, and K48-polyubiquitination chains. Recently, a study reported another E3 ligase, DPF2, which is a plant homeodomain (PHD) finger protein that interacts with OCT-3/4, and regulates the ubiquitination level and functions of it. PHD is one of the subgroups of RING-related E3 ligases and several studies have reported that the PHD finger protein has essential roles in ubiquitination and degradation of target proteins. DPF2 induces OCT-3/4 degradation by K48-polyubiquitination chain. Knockdown of DPF2 leads to induction of differentiation of human ESCs through an increase in OCT-3/4 level [22]. Several elements can affect the activity of stem cell transcription factors. First, the families belonging to E3 ligase may affect its activity. There are three E3 ligase subfamilies: HECT E3 ligases, the really interesting new gene (RING) finger-domain-containing E3 ligases, and the U box E3 ligases [21]. Second, targeting different regions in substrates by E3 ligases is also important. Even though WWP2 and ITCH belong to the same family, WWP2 targets the full-length of OCT-3/4, while ITCH targets a specific region of OCT-3/4. Third, cell types may also influence. For example, in undifferentiated stem cells, knockdown of WWP2 increases OCT-3/4 expression, while ITCH induces stem cell maintenance in differentiated stem cells. Lastly, substrate ubiquitination through 7 lysines may vary biological processes. K6-polyubiquitination has a cellular function for DNA repair [23], whereas K11- and K48-polyubiquitination target substrates for proteasomal degradation [24]. K27-polyubiquitination is related with mitophagy [25], K29- and K33-polyubiquitination are related with modification of kinases [26], and K63-polyubiquitination mediates intercellular signaling [27]. Through different lysine linkages, cellular functions of OCT-3/4 can be differently regulated. Therefore, several E3 ligases have different effects on OCT-3/4 activity because of their temporal and spatial expression. But, further investigation of their molecular mechanisms is required for understanding different effects on OCT-3/4. USP44 and USP34 are recently identified DUBs for OCT-3/4 and during ESC differentiation, they are downregulated. Absence of USP34 promoted the upregulation of OCT-3/4 to promote stemness [12]; on the other hand, absence of USP44 leads to the decrease of OCT-3/4 level in ESC differentiation [28]. However, their direct interaction is not yet studied. Moreover, OCT-3/4 is related to tumor progression, as overexpression of OCT-3/4 induces liver cancer cell resistance to chemotherapeutic drugs, while knockdown of OCT-3/4 expression reduces the cancer cell resistance level [22, 29–31]. OCT-3/4 has a crucial role in oncogenesis and may be an essential target for drug therapy of cancer.
SOX2
SRY box-containing gene 2 (SOX2) is a key component of iPSCs and neural stem cells (NSCs), and acts as a regulator in various phases of embryonic development and ESC differentiation [32]. SOX2 maintains NSC identity, therefore expression of this protein leads to cell cycle exit and aids in differentiation of NSCs [33]. This transcription factor has an essential role in re-establishing pluripotency in somatic cells by reprogramming them into iPSCs [34]. SOX2 contains the high mobility group (HMG) box, which interacts with the AT-rich motif to form an L-shaped binding surface. This L-shape surface leads to a substantial bend in DNA, which is involved in activation of transcription. The HMG domain has several regions associated with transactivation activity at the carboxy-terminus [17]. SOX2 overexpression induces differentiation into the neural lineage, and SOX2 can reprogram fibroblasts into multipotent NSC [32]. Suppression of ubiquitin-mediated protein degradation regulates the stabilization of SOX2 phosphorylation at Ser246, Ser249, Ser250, and Ser251. Phosphorylation of SOX2 enhances the self-renewal of mouse ESCs and stabilizes SOX2 by preventing protein degradation [35]. AKT1 functions as a kinase and can phosphorylate SOX2 at Thr118 for self-renewal of ESCs. mESC self-renewal ability is downregulated when AKT1 is inhibited, and caused the reduction of protein level of SOX2. Therefore, AKT1 positively regulates SOX2 transcriptional activity in mESC [36]. WWP2 is a C2-WW-HECT-type E3 ligase that interacts with K119 methylated SOX2 for the ubiquitination of SOX2. In addition, SET7 interacts with K119-monomethylated SOX2 to inhibit transcriptional activity of SOX2 and induces ubiquitination and degradation of SOX2. SOX2 enhances degradation by helping WWP2 to regulate SOX2 ubiquitination and proteasomal degradation [37]. More than 1200 genes within ESCs are responsible for SOX2-induced differentiation. Of these, OCT-3/4 and SOX2 act within autoregulatory positive feedback loops, which can reinforce the pluripotent state. Overexpression of OCT-3/4 is regulated by upregulation of SOX2 and vice versa, and this shows that functioning of SOX2 is essential in pluripotency of ESCs and activation of OCT-3/4 [38]. USP22 is a DUB of SOX2 which maintains stem cell pluripotency. This deubiquitinating enzyme enhanced the differentiation of ESCs into three germ layers [39]. During the reprogramming process in early phase stem cells, SOX2 induces differentiation into neural ectodermal, mesodermal, and trophectodermal cells, and it lowers mesendodermal gene expression [40]. USP7 [41], USP9X [42], USP15 [43], USP24 [43], USP25 [44], USP34 [12], USP37 [44], USP44 [14], and USP49 [45] are also DUBs of SOX2. The interactions between deubiquitinating enzymes and SOX2 give us new insight on stem cell regulation and stem cell fate specification.
KLF4
Krüppel-like factor 4 (KLF4), also called gut-enriched Krüppel-like factor (GKLF), belongs to the Krüppel-like factor (KLF) family, which controls numerous biological processes including proliferation, differentiation, development, and apoptosis [46]. This transcription factor contains transactivation, transrepression, and zinc finger domains [47]. Estrogen-related receptor beta (Esrrb) is an orphan nuclear receptor that is highly expressed in ESCs and has a role similar to that of KLF4 in the reprogramming of mouse embryonic fibroblasts (MEFs) [17, 18]. KLF4 is related to ubiquitination and is responsible for regulation of protein turnover in cells. Downregulation of KLF4 is controlled by serum stimulation. In MG132-treated cells, the level of KLF4 is increased and can be degraded by serum stimulation. Therefore, mESCs actively proliferate under a serum stimulation condition. To identify the specific site of ubiquitination, we created deletion constructs of KLF4. Essential lysines such as K32, K52, K232, and K252 on the N-terminal domain of KLF4 were identified in those constructs. These lysines are essential in proteolysis as well as in ubiquitination of the KLF4 protein; therefore, these results show that KLF4 undergoes proteasomal degradation [47]. In addition, KLF4 expression is downregulated by TGF-β-signaling, which is regulated by the UPS. Following proteasomal inhibitor MG132 treatment, proteasomal degradation of KLF4 increased. Cdh1/APC is an E3 ligase of KLF4 that has an important role in KLF4-related TFG-β-signaling. KLF4 is phosphorylated by ERK1 with β-TrCP1/TrCP2 E3 ligase activity, and this induces ubiquitination and protein degradation of KLF4 [32]. Krüppel-like factors are important in ESC self-renewal, and a low KLF4 level induces ESCs to undergo differentiation [46]. Recent studies have shown that KLF4 is important in remodeling cell fate via reprogramming of somatic cells to pluripotent cells.
C-MYC
C-MYC acts as a stem cell reprogramming inducer to maintain the pluripotency of cells [40]. This transcription factor regulates many cellular functions, cell division, cell growth, apoptosis, proliferation, tumorigenesis, and differentiation [48]. C-MYC does not act as a transcription factor of ESCs alone; rather, it functions with OCT-3/4, SOX2, or KLF4, to enhance the generation of partially reprogrammed ESCs [49–54]. In PTMs of C-MYC, ubiquitination and proteolysis are important in regulation of the stability or function of C-MYC. The half-life of C-MYC is approximately 20–30 min, which is less than that of other transcription factors. Proteasome inhibition using the proteasome inhibitors MG132 and lactacystin from the 26S proteasome induces C-MYC stability and differentiation [32]. C-MYC has several domains, and among them, MYC homology box I (MBI) and MBII are important in C-MYC proteolysis because these domains bind with a C-MYC ubiquitin ligase. C-MYC proteolysis and ubiquitination occur, especially, on the N-terminal part [47]. Another domain is the PEST motif, which has a crucial role in C-MYC degradation and proteolysis [32]. The degradation of C-MYC occurs in the nucleolus, but the site of its ubiquitination is not yet identified [55]. The first identified E3 ligase of C-MYC, S-phase kinase associated protein 2 (SKP2), is associated with the F-box protein, which interacts with several parts in C-MYC to induce ubiquitination [56]. FBXW7 is also an E3 ligase of C-MYC, and it promotes C-MYC ubiquitination through the phosphorylation of C-MYC on T58 with the help of glycogen synthase kinase 3 [32]. This prevents ESC differentiation, and it enhances cellular reprogramming and self-renewal through stabilization of C-MYC [57]. The other E3 ligases are SCFSkp2 and SCFFbw7, which rapidly regulate degradation of C-MYC [9]. Recent studies have identified DUBs of C-MYC, including USP22 [58], USP28 [59], USP36 [55], and USP37 [48]. USP22 induced by c-MYC may play a role in upregulation of SIRT1, which leads to the reduction of SIRT1 ubiquitination as well as increased stability in AML stem/progenitor cells [58]. USP28 was first reported as a DUB that does not interact directly with C-MYC, but, with the help of E3 ligase FBXW7, USP28 deubiquitinates C-MYC [59]. USP36 deubiquitinates and stabilizes C-MYC within the nucleus of the cell. When the level of USP36 is reduced, the levels of C-MYC and cell proliferation decrease [55, 60]. USP37 is a DUB that deubiquitinates C-MYC, and when the expression level of USP37 is increased, degradation of C-MYC is blocked. In contrast, a reduced USP37 level enhances C-MYC degradation [48].
LIN28
Abnormal cell lineage protein 28 (LIN28) is a transcription factor and is also referred to as RNA-binding protein LIN28. This transcription factor was first identified in Caenorhabditis elegans when screening for lineage-modifying genes [61]. The let-7 (Mirlet7) family of miRNAs is mediated by LIN28 at the transcriptional and post-transcriptional levels [62]. LIN28 binds with let-7 as a negative regulator and blocks processing of let-7 miRNA with high specificity in order to control ESC self-renewal and differentiation [63]. By using LIN28, OCT-3/4, SOX2, and NANOG, human fibroblasts have been reprogrammed into iPSCs successfully [64]. These transcription factors are so-called ‘Thomson factors’ and are different from Yamanaka factors. Yamanaka factors contain KLF4 and C-MYC, not LIN28 and NANOG [65]. When LIN28 expression is inhibited, it reduces the expression of OCT-3/4, which affects pluripotency and self-renewal of ESCs. LIN28 helps to regulate OCT-3/4 expression, revealing that when LIN28 is overexpressed, ESCs can be differentiated into the endoderm lineage [66]. The E3 ubiquitin ligases LIN41/TRIM71, LEP-2/MKRNS affect LIN28, but they do not regulate LIN28 directly. LIN41/TRIM71 decreases activity of let-7 causing negative regulation. During differentiation of ESCs, LIN41/TRIM71 and LIN28 levels are downregulated, while the level of let-7 is increased. Therefore, LIN41/TRIM71 is also a negative regulator of let-7 [67]. LIN41/TRIM71 negatively regulates LIN28 through let-7 by polyubiquitination [68]. LEP-2 is an allele of Makorin MKRN and regulation of LIN28 by LEP-2/MKRNS has not been fully elucidated, but this E3 ligase contain the RING domain, as in other E3 ligases, and tag LIN28 with ubiquitin to act as E3 ligases for proteasomal degradation [69]. The high level of LIN28 expression in the early stage of ESC development decreases upon differentiation of the cells [70]. In addition to the effects on humans, land plants can be also differentiated by Physcomitrella patens cold-shock domain protein 1 (PpCSP1), which is conserved as a portion of a domain of LIN28 in mammals. Recently, Li et al. revealed that without the 3′-untranslated region (3′-UTR) of PpCSP1, PpCSP1 has a similar role as mammalian LIN28 [71]. Another study has shown that overexpression of SOX2 induces proliferation of zebrafish Müller glial cells, and also directly regulates reprogramming factors of ascl1a (neurogenic) and LIN28 (reprogramming) [72].
FAK
Focal adhesion is a cellular process controlled by integrin, talin, vinculin, paxillin, Src, and focal adhesion kinase (FAK), essential proteins that transmit signals from the extracellular environment to the cell interior for application in actin remodeling and gene activation [73]. Human chromosome 8, mouse chromosome 15, chicken, and Xenopus all contain a 90% sequence of the FAK gene. FAK has a size of 125 kDa and belongs to non-receptor protein tyrosine kinase (PTK) or cytoplasmic tyrosine kinase families. FAK scaffolds proteins during organismal disease and development. This kinase was identified first as an integrin-regulated PTK. Integrin-dependent cell adhesion regulates the activity and phosphorylation of FAK including motility, spreading, proliferation, and survival. FAK contains three specific domains: the central kinase domain, the N-terminal FERM domain and the C-terminal focal adhesion targeting (FAT) sequence domain. In FAK, there are various sites for tyrosine phosphorylation and that process regulates FAK activity and interaction with SH2-domain containing proteins; for example, the autophosphorylation site of Tyr397, which is essential for FAK functions [74]. FAK also regulates a skeletal myogenesis process via the E2 enzyme UBE2H and the E3 ligase MG53 (mitsugumin 53)/TRIM72 (tripartite motif-containing 72). When FAK forms a complex with UBE2H and MG53/TRIM72, it undergoes ubiquitination followed by degradation. MG53/TRIM72, the first identified E3 ligase for FAK, is involved in skeletal myogenesis with FAK [73]. Ubiquitin C-terminal hydrolase-L1 (UCH-L1) is a DUB of FAK; moreover, it activates and stabilizes FAK during initial phases of adhesion [75]. Activated FAK can form a complex with Src family kinase (i.e., FAK/Src complex) and can initiate multiple downstream signaling pathways by phosphorylating other proteins for the regulation of different cellular functions. In angiogenesis, FAK has essential roles in embryonic development and cancer progression. The FAK expression level is increased in various human cancers. FAK functions in the nucleus to regulate p53 interaction with MDM2, and this regulation causes ubiquitination and degradation to occur. A decreased level of FAK enhances the expression level of p53, and the FERM domain of FAK binds with p53 and MDM2. In a kinase-independent manner, the FERM domain of FAK1 inactivates p53 that enhances MDM2-dependent p53 ubiquitination; as well, it induces cell proliferation and cell survival. Therefore, FAK might be a therapeutic target for treatment of cancer and cardiovascular diseases [74]. An analysis of expression patterns of mOCT-3/4 and mNANOG revealed that FAK1 protein expression was positively regulated by OCT-3/4 and NANOG. Forced ablation of mFAK1 inhibits foci formation with mOCT-3/4 and mNANOG. Both C-MYC and cyclin D1 are essential molecules in the cell cycle process. FAK1 regulates expression of C-MYC and cyclin D1 for cell proliferation, and FAK stimulates ERK1/2 pathways with these molecules. Therefore, knockdown of mFAK1 decreases cell proliferation by inhibiting ERK1/2 [76]. USP22 regulates FAK in cancer cell, therefore downregulation of USP22 may effectively work as a treatment for cancer [77], but in stem cell, studies have not been reported yet.
Telomerase
Telomeres are located at the end of eukaryotic chromosomes and consist of a repetitious hexameric DNA sequence (5′-TTAGGG-3′) [78]. Shortening of the telomere occurs when the cell undergoes differentiation, and telomerase, a polymerase of telomere, can reduce this reduction. Telomerase is a ribonucleoprotein and consists of human telomerase reverse transcriptase (hTERT), which is a catalytic protein subunit and is complementary to the hexameric repeat sequence (RNA template, hTR) [79]. Activity of telomerase exists in 90% of cancer cells, and when the telomerase activity decreases, it can induce cancer cell death. In somatic cells, telomerase is absent, but in stem cells, telomerase is activated so it can induce stem cell pluripotency [80]. PTMs of telomerase occur subsequent to hTERT binding with some proteins such as Hsp90. Hsp90 is a chaperone protein and binds to hTERT to promote telomerase activity. Geldanamycin (GA) is a chaperone inhibitor that blocks the interaction between HSP90 and telomerase. GA leads to the ubiquitination and proteasome-mediated degradation of hTERT; therefore, it decreases telomerase activity. Makorin 1 RING finger protein (MKRN1) is an E3 ligase of hTERT that can induce ubiquitin-mediated 26S proteasomal degradation of hTERT and regulate telomere homeostasis by decreasing telomerase activity [79]. The RING family is the largest class of ubiquitin E3 ligases to facilitate the transfer of ubiquitin to its substrate. Overexpression of MKRN1 induces degradation of hTERT and reduces telomerase activity, as well, it decreases telomere length even in the absence of GA [81]. Telomeric protein TPP1 recruits and regulates telomerase and this protein deubiquitinates by USP7 [82]. A future telomere research challenge will be the determination of telomere-related DUBs and their functional roles at the molecular level.
Conclusion
Many scientists have been investigating transcription factors, such as NANOG, OCT-3/4, SOX2, KLF4, C-MYC, LIN28, FAK, and telomerase that can regulate ESC self-renewal, reprogramming, and maintenance. OCT-3/4, SOX2, KLF4, and C-MYC are stem cell transcription factors that can induce pluripotency. Unlike SOX2, KLF4, and C-MYC, OCT-3/4 can be substituted by another factor, Nr5a2 [18]. OCT-3/4 blocks ESC differentiation into the trophoblast. In contrast, NANOG blocks differentiation into the primitive endoderm and germ layers. OCT-3/4 and NANOG are pluripotent marker proteins. Some studies have reported that NANOG is less stable than OCT-3/4 and degradation of OCT-3/4 occurs differently from that of NANOG [22]. Unlike other transcriptional factors, interaction between SOX2 with OCT-3/4 has a special function in maintaining pluripotency. When the OCT-3/4 activity is decreased, the level of SOX2 is also decreased [38]. These transcription factors are ubiquitinated by E3 ubiquitin ligases, and these E3 ligases have a positive effect on protein degradation. E3 ligases catalyze stemness-related proteins for ubiquitination, which leads to differentiation of stem cells. In contrast, when stemness-related proteins are deubiquitinated, stem cell differentiation is inhibited. Transcription factors have their own E3 ligases and DUBs (Table 1). DUBs exist in large multi-protein complexes that function in stem cell differentiation, pluripotency, and reprogramming [7]. Therefore, modulation of DUBs can improve cellular reprogramming efficiency during iPSC generation. Establishment of a fully reprogrammed iPSC will be useful in clinical applications. In the USA, ESC-derived cells have already been approved for use in some treatments [40]. Here, we defined various stem cell types related to the UPS (Table 2). When DUB enzymes and stem cell transcription factors form a complex, treatment approaches to various diseases such as cancer will become clearer. Such complexes may permit the design of novel therapeutics and the establishment of biomarkers that may be used in diagnosis and prognosis development.
Table 1.
Transcription factors related to E3 ligases and DUBs
| E3 ligases | DUBs | ||||
|---|---|---|---|---|---|
| NANOG | FBXW8 [12] | USP21 [4, 9, 10] | |||
| USP34 [13] | |||||
| OCT-3/4 | WWP2 [21] | DPF2 [22] | USP44 [24] | ||
| ITCH [23] | RNF2 [22] | USP34 [13] | |||
| SOX2 | WWP2 [33] | USP44 [15] | USP34 [13] | USP9X [38] | |
| USP49 [41] | USP37 [40] | ||||
| USP7 [37] | USP25 [40] | ||||
| USP15 [39] | USP24 [39] | ||||
| KLF4 | β-TrCP1/TrCP2 [28] | ||||
| C-MYC | FBXW7 [28, 55] | USP22 [54] | |||
| SKP2 [10, 52] | USP28 [55] | ||||
| USP36 [51, 56] | |||||
| USP37 [44] | |||||
| LIN28 | LIN41 [63] | LEP-2 [65] | |||
| TRIM71 [64] | MKRNS [65] | ||||
| FAK | MG53 [69] | UCH-L1 [71] | |||
| USP22 [73] | |||||
| Telomerase | MKRN1 [75, 77] | USP7 [78] | |||
Table 2.
Characteristics of various stem cells
| Characteristics | Stem cells | |||
|---|---|---|---|---|
| ESC | NSC | HSC | MSC | |
| Origins | ICM | Brain | Bone marrow | Fetal |
| Umbilical cord blood | Organs | |||
| 2 Fundamental properties | 1. Self-renewal | 1. Self-renewal | 1. Self-renewal | 1. Self-renewal |
| 2. Pluripotency | 2. Multipotent | 2. Differentiation | 2. Multipotent | |
| UPS | Regulates processes and molecular components | UCH-L1 promotes differentiation | Proteasome inhibitors regulate processes | Regulates Wnt/β-catenin, BMP signaling, and Notch pathway |
|
Transcription factors Regulated by UPS |
OCT-3/4 | NF-κB | NF-κB | |
| C-REL and RELA | SP7/OSTERIX | |||
| GATA-1 and GATA-2 | RUNX2 | |||
| E3 | WWP2 | MID | SMURF family | |
| UBR1 and UBR2 | ||||
| DDB1 | ||||
ESC, embryonic stem cell; NSC, neural stem cell; HSC, hematopoietic stem cell; MSC, mesenchymal stem cell; ICM, inner cell mass; UPS, ubiquitin-proteasome system; E3, ubiquitin ligase; UCH-L1, ubiquitin-c-terminal hydrolase-L1; BMP, bone morphology proteins; Oct-3/4, octamer-binding transcription factor-3/4; NF-κB, nuclear factor κB; GATA, GATA-binding factor; Runx2, runt-related transcription factor 2; Wwp2, WW domain-containing protein 2; MID1, mind bomb 1; UBR, ubiquitin protein ligase E3 component n-recognin; DDB1, DNA damage-binding protein 1; Smurf, smad ubiquitination regulatory factor
Acknowledgements
We would like to thank members of Baek laboratory for their critical comments on the manuscript.
Author contributions
JC: manuscript writing; KHB: manuscript writing, final approval of manuscript.
Compliance with ethical standards
Conflict of interest
The authors declare that they have no conflict of interest.
References
- 1.Shirazi R, Zarnani AH, Soleimani M, Nayernia K, Ragerdi Kashani I. Differentiation of bone marrow-derived stage-specific embryonic antigen 1 positive pluripotent stem cells into male germ cells. Microsc Res Tech. 2017;80(4):430–440. doi: 10.1002/jemt.22812. [DOI] [PubMed] [Google Scholar]
- 2.Zhang W, Sui Y, Ni J, Yang T. Insights into the Nanog gene: a propeller for stemness in primitive stem cells. Int J Biol Sci. 2016;12(11):1372–1381. doi: 10.7150/ijbs.16349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Smith AG. Embryo-derived stem cells: of mice and men. Annu Rev Cell Dev Biol. 2001;17:435–462. doi: 10.1146/annurev.cellbio.17.1.435. [DOI] [PubMed] [Google Scholar]
- 4.Kwon SK, Lee DH, Kim SY, Park JH, Choi J, Baek KH. Ubiquitin-specific protease 21 regulating the K48-linked polyubiquitination of NANOG. Biochem Biophys Res Commun. 2017;482(4):1443–1448. doi: 10.1016/j.bbrc.2016.12.055. [DOI] [PubMed] [Google Scholar]
- 5.Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- 6.Naujokat C, Saric T. Concise review: role and function of the ubiquitin-proteasome system in mammalian stem and progenitor cells. Stem Cells. 2007;25(10):2408–2418. doi: 10.1634/stemcells.2007-0255. [DOI] [PubMed] [Google Scholar]
- 7.Ramakrishna S, Kim KS, Baek KH. Posttranslational modifications of defined embryonic reprogramming transcription factors. Cell Reprogram. 2014;16(2):108–120. doi: 10.1089/cell.2013.0077. [DOI] [PubMed] [Google Scholar]
- 8.Jin J, Liu J, Chen C, Liu Z, Jiang C, Chu H, Pan W, Wang X, Zhang L, Li B, Jiang C, Ge X, Xie X, Wang P. The deubiquitinase USP21 maintains the stemness of mouse embryonic stem cells via stabilization of Nanog. Nat Commun. 2016;7:13594. doi: 10.1038/ncomms13594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Liu X, Yao Y, Ding H, Han C, Chen Y, Zhang Y, Wang C, Zhang X, Zhang Y, Zhai Y. USP21 deubiquitylates Nanog to regulate protein stability and stem cell pluripotency. Signal Transduct Target Ther. 2016;1:16024. doi: 10.1038/sigtrans.2016.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ramakrishna S, Suresh B, Lim KH, Cha BH, Lee SH, Kim KS, Baek KH. PEST motif sequence regulating human NANOG for proteasomal degradation. Stem Cells Dev. 2011;20(9):1511–1519. doi: 10.1089/scd.2010.0410. [DOI] [PubMed] [Google Scholar]
- 11.Kim SH, Kim MO, Cho YY, Yao K, Kim DJ, Jeong CH, Yu DH, Bae KB, Cho EJ, Jung SK, Lee MH, Chen H, Kim JY, Bode AM, Dong Z. ERK1 phosphorylates Nanog to regulate protein stability and stem cell self-renewal. Stem Cell Res. 2014;13(1):1–11. doi: 10.1016/j.scr.2014.04.001. [DOI] [PubMed] [Google Scholar]
- 12.Oh E, Kim JY, Sung D, Cho Y, Lee N, An H, Kim YJ, Cho TM, Seo JH. Inhibition of ubiquitin-specific protease 34 (USP34) induces epithelial-mesenchymal transition and promotes stemness in mammary epithelial cells. Cell Signal. 2017;36:230–239. doi: 10.1016/j.cellsig.2017.05.009. [DOI] [PubMed] [Google Scholar]
- 13.Jin W, Wang L, Zhu F, Tan W, Lin W, Chen D, Sun Q, Xia Z. Critical POU domain residues confer Oct4 uniqueness in somatic cell reprogramming. Sci Rep. 2016;6:20818. doi: 10.1038/srep20818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG, Gifford DK, Melton DA, Jaenisch R, Young RA. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005;122(6):947–956. doi: 10.1016/j.cell.2005.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Saxe JP, Tomilin A, Scholer HR, Plath K, Huang J. Post-translational regulation of Oct4 transcriptional activity. PLoS One. 2009;4(2):e4467. doi: 10.1371/journal.pone.0004467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Oka M, Moriyama T, Asally M, Kawakami K, Yoneda Y. Differential role for transcription factor Oct4 nucleocytoplasmic dynamics in somatic cell reprogramming and self-renewal of embryonic stem cells. J Biol Chem. 2013;288(21):15085–15097. doi: 10.1074/jbc.M112.448837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schmidt R, Plath K. The roles of the reprogramming factors Oct4, Sox2 and Klf4 in resetting the somatic cell epigenome during induced pluripotent stem cell generation. Genome Biol. 2012;13(10):251. doi: 10.1186/gb-2012-13-10-251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Heng JC, Feng B, Han J, Jiang J, Kraus P, Ng JH, Orlov YL, Huss M, Yang L, Lufkin T, Lim B, Ng HH. The nuclear receptor Nr5a2 can replace Oct4 in the reprogramming of murine somatic cells to pluripotent cells. Cell Stem Cell. 2010;6(2):167–174. doi: 10.1016/j.stem.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 19.Villodre ES, Kipper FC, Pereira MB, Lenz G. Roles of OCT4 in tumorigenesis, cancer therapy resistance and prognosis. Cancer Treat Rev. 2016;51:1–9. doi: 10.1016/j.ctrv.2016.10.003. [DOI] [PubMed] [Google Scholar]
- 20.Liao B, Jin Y. Wwp2 mediates Oct4 ubiquitination and its own auto-ubiquitination in a dosage-dependent manner. Cell Res. 2010;20(3):332–344. doi: 10.1038/cr.2009.136. [DOI] [PubMed] [Google Scholar]
- 21.Liao B, Zhong X, Xu H, Xiao F, Fang Z, Gu J, Chen Y, Zhao Y, Jin Y. Itch, an E3 ligase of Oct4, is required for embryonic stem cell self-renewal and pluripotency induction. J Cell Physiol. 2013;228(7):1443–1451. doi: 10.1002/jcp.24297. [DOI] [PubMed] [Google Scholar]
- 22.Liu C, Zhang D, Shen Y, Tao X, Liu L, Zhong Y, Fang S. DPF2 regulates OCT4 protein level and nuclear distribution. Biochim Biophys Acta. 1853;12:3279–3293. doi: 10.1016/j.bbamcr.2015.09.029. [DOI] [PubMed] [Google Scholar]
- 23.Akutsu M, Dikic I, Bremm A. Ubiquitin chain diversity at a glance. J Cell Sci. 2016;129(5):875–880. doi: 10.1242/jcs.183954. [DOI] [PubMed] [Google Scholar]
- 24.Grice GL, Nathan JA. The recognition of ubiquitinated proteins by the proteasome. Cell Mol Life Sci. 2016;73(18):3497–3506. doi: 10.1007/s00018-016-2255-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Birsa N, Norkett R, Wauer T, Mevissen TE, Wu HC, Foltynie T, Bhatia K, Hirst WD, Komander D, Plun-Favreau H, Kittler JT. Lysine 27 ubiquitination of the mitochondrial transport protein Miro is dependent on serine 65 of the Parkin ubiquitin ligase. J Biol Chem. 2014;289(21):14569–14582. doi: 10.1074/jbc.M114.563031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Davis ME, Gack MU. Ubiquitination in the antiviral immune response. Virology. 2015;479–480:52–65. doi: 10.1016/j.virol.2015.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wertz IE, Dixit VM. Regulation of death receptor signaling by the ubiquitin system. Cell Death Differ. 2010;17(1):14–24. doi: 10.1038/cdd.2009.168. [DOI] [PubMed] [Google Scholar]
- 28.Fuchs G, Shema E, Vesterman R, Kotler E, Wolchinsky Z, Wilder S, Golomb L, Pribluda A, Zhang F, Haj-Yahya M, Feldmesser E, Brik A, Yu X, Hanna J, Aberdam D, Domany E, Oren M. RNF20 and USP44 regulate stem cell differentiation by modulating H2B monoubiquitylation. Mol Cell. 2012;46(5):662–673. doi: 10.1016/j.molcel.2012.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tang YA, Chen CH, Sun HS, Cheng CP, Tseng VS, Hsu HS, Su WC, Lai WW, Wang YC. Global Oct4 target gene analysis reveals novel downstream PTEN and TNC genes required for drug-resistance and metastasis in lung cancer. Nucleic Acids Res. 2015;43(3):1593–1608. doi: 10.1093/nar/gkv024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wang YD, Cai N, Wu XL, Cao HZ, Xie LL, Zheng PS. OCT4 promotes tumorigenesis and inhibits apoptosis of cervical cancer cells by miR-125b/BAK1 pathway. Cell Death Dis. 2013;4:e760. doi: 10.1038/cddis.2013.272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lu Y, Zhu H, Shan H, Lu J, Chang X, Li X, Lu J, Fan X, Zhu S, Wang Y, Guo Q, Wang L, Huang Y, Zhu M, Wang Z. Knockdown of Oct4 and Nanog expression inhibits the stemness of pancreatic cancer cells. Cancer Lett. 2013;340(1):113–123. doi: 10.1016/j.canlet.2013.07.009. [DOI] [PubMed] [Google Scholar]
- 32.Suresh B, Lee J, Kim KS, Ramakrishna S. The importance of ubiquitination and deubiquitination in cellular reprogramming. Stem Cells Int. 2016;2016:6705927. doi: 10.1155/2016/6705927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ring KL, Tong LM, Balestra ME, Javier R, Andrews-Zwilling Y, Li G, Walker D, Zhang WR, Kreitzer AC, Huang Y. Direct reprogramming of mouse and human fibroblasts into multipotent neural stem cells with a single factor. Cell Stem Cell. 2012;11(1):100–109. doi: 10.1016/j.stem.2012.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nagano K, Itagaki C, Izumi T, Nunomura K, Soda Y, Tani K, Takahashi N, Takenawa T, Isobe T. Rb plays a role in survival of Abl-dependent human tumor cells as a downstream effector of Abl tyrosine kinase. Oncogene. 2006;25(4):493–502. doi: 10.1038/sj.onc.1208996. [DOI] [PubMed] [Google Scholar]
- 35.Ouyang J, Yu W, Liu J, Zhang N, Florens L, Chen J, Liu H, Washburn M, Pei D, Xie T. Cyclin-dependent kinase-mediated Sox2 phosphorylation enhances the ability of Sox2 to establish the pluripotent state. J Biol Chem. 2015;290(37):22782–22794. doi: 10.1074/jbc.M115.658195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jeong CH, Cho YY, Kim MO, Kim SH, Cho EJ, Lee SY, Jeon YJ, Lee KY, Yao K, Keum YS, Bode AM, Dong Z. Phosphorylation of Sox2 cooperates in reprogramming to pluripotent stem cells. Stem Cells. 2010;28(12):2141–2150. doi: 10.1002/stem.540. [DOI] [PubMed] [Google Scholar]
- 37.Fang L, Zhang L, Wei W, Jin X, Wang P, Tong Y, Li J, Du JX, Wong J. A methylation-phosphorylation switch determines Sox2 stability and function in ESC maintenance or differentiation. Mol Cell. 2014;55(4):537–551. doi: 10.1016/j.molcel.2014.06.018. [DOI] [PubMed] [Google Scholar]
- 38.Masui S, Nakatake Y, Toyooka Y, Shimosato D, Yagi R, Takahashi K, Okochi H, Okuda A, Matoba R, Sharov AA, Ko MS, Niwa H. Pluripotency governed by Sox2 via regulation of Oct3/4 expression in mouse embryonic stem cells. Nat Cell Biol. 2007;9(6):625–635. doi: 10.1038/ncb1589. [DOI] [PubMed] [Google Scholar]
- 39.Sussman RT, Stanek TJ, Esteso P, Gearhart JD, Knudsen KE, McMahon SB. The epigenetic modifier ubiquitin-specific protease 22 (USP22) regulates embryonic stem cell differentiation via transcriptional repression of sex-determining region Y-box 2 (SOX2) J Biol Chem. 2013;288(33):24234–24246. doi: 10.1074/jbc.M113.469783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Takahashi K, Yamanaka S. A decade of transcription factor-mediated reprogramming to pluripotency. Nat Rev Mol Cell Biol. 2016;17(3):183–193. doi: 10.1038/nrm.2016.8. [DOI] [PubMed] [Google Scholar]
- 41.Cox JL, Wilder PJ, Gilmore JM, Wuebben EL, Washburn MP, Rizzino A. The SOX2-interactome in brain cancer cells identifies the requirement of MSI2 and USP9X for the growth of brain tumor cells. PLoS One. 2013;8(5):e62857. doi: 10.1371/journal.pone.0062857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Oishi S, Premarathne S, Harvey TJ, Iyer S, Dixon C, Alexander S, Burne TH, Wood SA, Piper M. Usp9x-deficiency disrupts the morphological development of the postnatal hippocampal dentate gyrus. Sci Rep. 2016;6:25783. doi: 10.1038/srep25783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wuebben EL, Rizzino A. The dark side of SOX2: cancer-a comprehensive overview. Oncotarget. 2017;8(27):44917–44943. doi: 10.18632/oncotarget.16570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Qiu GZ, Sun W, Jin MZ, Lin J, Lu PG, Jin WL. The bad seed gardener: deubiquitinases in the cancer stem-cell signaling network and therapeutic resistance. Pharmacol Ther. 2017;172:127–138. doi: 10.1016/j.pharmthera.2016.12.003. [DOI] [PubMed] [Google Scholar]
- 45.Suresh B, Lee J, Kim H, Ramakrishna S. Regulation of pluripotency and differentiation by deubiquitinating enzymes. Cell Death Differ. 2016;23(8):1257–1264. doi: 10.1038/cdd.2016.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jiang J, Chan YS, Loh YH, Cai J, Tong GQ, Lim CA, Robson P, Zhong S, Ng HH. A core Klf circuitry regulates self-renewal of embryonic stem cells. Nat Cell Biol. 2008;10(3):353–360. doi: 10.1038/ncb1698. [DOI] [PubMed] [Google Scholar]
- 47.Lim KH, Kim SR, Ramakrishna S, Baek KH. Critical lysine residues of Klf4 required for protein stabilization and degradation. Biochem Biophys Res Commun. 2014;443(4):1206–1210. doi: 10.1016/j.bbrc.2013.12.121. [DOI] [PubMed] [Google Scholar]
- 48.Pan J, Deng Q, Jiang C, Wang X, Niu T, Li H, Chen T, Jin J, Pan W, Cai X, Yang X, Lu M, Xiao J, Wang P. USP37 directly deubiquitinates and stabilizes c-Myc in lung cancer. Oncogene. 2015;34(30):3957–3967. doi: 10.1038/onc.2014.327. [DOI] [PubMed] [Google Scholar]
- 49.Polo JM, Anderssen E, Walsh RM, Schwarz BA, Nefzger CM, Lim SM, Borkent M, Apostolou E, Alaei S, Cloutier J, Bar-Nur O, Cheloufi S, Stadtfeld M, Figueroa ME, Robinton D, Natesan S, Melnick A, Zhu J, Ramaswamy S, Hochedlinger K. A molecular roadmap of reprogramming somatic cells into iPS cells. Cell. 2012;151(7):1617–1632. doi: 10.1016/j.cell.2012.11.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Olariu V, Lovkvist C, Sneppen K. Nanog, Oct4 and Tet1 interplay in establishing pluripotency. Sci Rep. 2016;6:25438. doi: 10.1038/srep25438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Feng B, Jiang J, Kraus P, Ng JH, Heng JC, Chan YS, Yaw LP, Zhang W, Loh YH, Han J, Vega VB, Cacheux-Rataboul V, Lim B, Lufkin T, Ng HH. Reprogramming of fibroblasts into induced pluripotent stem cells with orphan nuclear receptor Esrrb. Nat Cell Biol. 2009;11(2):197–203. doi: 10.1038/ncb1827. [DOI] [PubMed] [Google Scholar]
- 52.Nakagawa M, Koyanagi M, Tanabe K, Takahashi K, Ichisaka T, Aoi T, Okita K, Mochiduki Y, Takizawa N, Yamanaka S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat Biotechnol. 2008;26(1):101–106. doi: 10.1038/nbt1374. [DOI] [PubMed] [Google Scholar]
- 53.Okita K, Ichisaka T, Yamanaka S. Generation of germline-competent induced pluripotent stem cells. Nature. 2007;448(7151):313–317. doi: 10.1038/nature05934. [DOI] [PubMed] [Google Scholar]
- 54.Carey BW, Markoulaki S, Hanna JH, Faddah DA, Buganim Y, Kim J, Ganz K, Steine EJ, Cassady JP, Creyghton MP, Welstead GG, Gao Q, Jaenisch R. Reprogramming factor stoichiometry influences the epigenetic state and biological properties of induced pluripotent stem cells. Cell Stem Cell. 2011;9(6):588–598. doi: 10.1016/j.stem.2011.11.003. [DOI] [PubMed] [Google Scholar]
- 55.Sun XX, He X, Yin L, Komada M, Sears RC, Dai MS. The nucleolar ubiquitin-specific protease USP36 deubiquitinates and stabilizes c-Myc. Proc Natl Acad Sci USA. 2015;112(12):3734–3739. doi: 10.1073/pnas.1411713112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kim SY, Herbst A, Tworkowski KA, Salghetti SE, Tansey WP. Skp2 regulates Myc protein stability and activity. Mol Cell. 2003;11(5):1177–1188. doi: 10.1016/S1097-2765(03)00173-4. [DOI] [PubMed] [Google Scholar]
- 57.Cartwright P, McLean C, Sheppard A, Rivett D, Jones K, Dalton S. LIF/STAT3 controls ES cell self-renewal and pluripotency by a Myc-dependent mechanism. Development. 2005;132(5):885–896. doi: 10.1242/dev.01670. [DOI] [PubMed] [Google Scholar]
- 58.Li L, Osdal T, Ho Y, Chun S, McDonald T, Agarwal P, Lin A, Chu S, Qi J, Li L, Hsieh YT, Dos Santos C, Yuan H, Ha TQ, Popa M, Hovland R, Bruserud O, Gjertsen BT, Kuo YH, Chen W, Lain S, McCormack E, Bhatia R. SIRT1 activation by a c-MYC oncogenic network promotes the maintenance and drug resistance of human FLT3-ITD acute myeloid leukemia stem cells. Cell Stem Cell. 2014;15(4):431–446. doi: 10.1016/j.stem.2014.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Diefenbacher ME, Chakraborty A, Blake SM, Mitter R, Popov N, Eilers M, Behrens A. Usp28 counteracts Fbw7 in intestinal homeostasis and cancer. Cancer Res. 2015;75(7):1181–1186. doi: 10.1158/0008-5472.CAN-14-1726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Welcker M, Orian A, Jin J, Grim JE, Harper JW, Eisenman RN, Clurman BE. The Fbw7 tumor suppressor regulates glycogen synthase kinase 3 phosphorylation-dependent c-Myc protein degradation. Proc Natl Acad Sci USA. 2004;101(24):9085–9090. doi: 10.1073/pnas.0402770101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ambros V, Horvitz HR. Heterochronic mutants of the nematode Caenorhabditis elegans. Science. 1984;226(4673):409–416. doi: 10.1126/science.6494891. [DOI] [PubMed] [Google Scholar]
- 62.Roush S, Slack FJ. The let-7 family of microRNAs. Trends Cell Biol. 2008;18(10):505–516. doi: 10.1016/j.tcb.2008.07.007. [DOI] [PubMed] [Google Scholar]
- 63.Balzer E, Heine C, Jiang Q, Lee VM, Moss EG. LIN28 alters cell fate succession and acts independently of the let-7 microRNA during neurogliogenesis in vitro. Development. 2010;137(6):891–900. doi: 10.1242/dev.042895. [DOI] [PubMed] [Google Scholar]
- 64.Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, Slukvin II, Thomson JA. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318(5858):1917–1920. doi: 10.1126/science.1151526. [DOI] [PubMed] [Google Scholar]
- 65.Kashyap V, Rezende NC, Scotland KB, Shaffer SM, Persson JL, Gudas LJ, Mongan NP. Regulation of stem cell pluripotency and differentiation involves a mutual regulatory circuit of the NANOG, OCT4, and SOX2 pluripotency transcription factors with polycomb repressive complexes and stem cell microRNAs. Stem Cells Dev. 2009;18(7):1093–1108. doi: 10.1089/scd.2009.0113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Qiu C, Ma Y, Wang J, Peng S, Huang Y. Lin28-mediated post-transcriptional regulation of Oct4 expression in human embryonic stem cells. Nucleic Acids Res. 2010;38(4):1240–1248. doi: 10.1093/nar/gkp1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Worringer KA, Rand TA, Hayashi Y, Sami S, Takahashi K, Tanabe K, Narita M, Srivastava D, Yamanaka S. The let-7/LIN-41 pathway regulates reprogramming to human induced pluripotent stem cells by controlling expression of prodifferentiation genes. Cell Stem Cell. 2014;14(1):40–52. doi: 10.1016/j.stem.2013.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Lee SH, Cho S, Kim MS, Choi K, Cho JY, Gwak HS, Kim YJ, Yoo H, Lee SH, Park JB, Kim JH. The ubiquitin ligase human TRIM71 regulates let-7 microRNA biogenesis via modulation of Lin28B protein. Biochim Biophys Acta. 2014;5:374–386. doi: 10.1016/j.bbagrm.2014.02.017. [DOI] [PubMed] [Google Scholar]
- 69.Herrera RA, Kiontke K, Fitch DH. Makorin ortholog LEP-2 regulates LIN-28 stability to promote the juvenile-to-adult transition in Caenorhabditis elegans. Development. 2016;143(5):799–809. doi: 10.1242/dev.132738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Moss EG, Tang L. Conservation of the heterochronic regulator Lin-28, its developmental expression and microRNA complementary sites. Dev Biol. 2003;258(2):432–442. doi: 10.1016/S0012-1606(03)00126-X. [DOI] [PubMed] [Google Scholar]
- 71.Li C, Sako Y, Imai A, Nishiyama T, Thompson K, Kubo M, Hiwatashi Y, Kabeya Y, Karlson D, Wu SH, Ishikawa M, Murata T, Benfey PN, Sato Y, Tamada Y, Hasebe M. A Lin28 homologue reprograms differentiated cells to stem cells in the moss Physcomitrella patens. Nat Commun. 2017;8:14242. doi: 10.1038/ncomms14242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Gorsuch RA, Lahne M, Yarka CE, Petravick ME, Li J, Hyde DR. Sox2 regulates Muller glia reprogramming and proliferation in the regenerating zebrafish retina via Lin28 and Ascl1a. Exp Eye Res. 2017;161:174–192. doi: 10.1016/j.exer.2017.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Nguyen N, Yi JS, Park H, Lee JS, Ko YG. Mitsugumin 53 (MG53) ligase ubiquitinates focal adhesion kinase during skeletal myogenesis. J Biol Chem. 2014;289(6):3209–3216. doi: 10.1074/jbc.M113.525154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhao X, Guan JL. Focal adhesion kinase and its signaling pathways in cell migration and angiogenesis. Adv Drug Deliv Rev. 2011;63(8):610–615. doi: 10.1016/j.addr.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Frisan T, Coppotelli G, Dryselius R, Masucci MG. Ubiquitin C-terminal hydrolase-L1 interacts with adhesion complexes and promotes cell migration, survival, and anchorage independent growth. FASEB J. 2012;26(12):5060–5070. doi: 10.1096/fj.12-211946. [DOI] [PubMed] [Google Scholar]
- 76.Ajjappala BS, Kim MS, Kim EY, Kim JH, Kang IC, Baek KH. Protein chip analysis of pluripotency-associated proteins in NIH3T3 fibroblast. Proteomics. 2009;9(16):3968–3978. doi: 10.1002/pmic.200800611. [DOI] [PubMed] [Google Scholar]
- 77.Ning Z, Wang A, Liang J, Xie Y, Liu J, Yan Q, Wang Z. USP22 promotes epithelial-mesenchymal transition via the FAK pathway in pancreatic cancer cells. Oncol Rep. 2014;32(4):1451–1458. doi: 10.3892/or.2014.3354. [DOI] [PubMed] [Google Scholar]
- 78.Greider CW. Telomere length regulation. Annu Rev Biochem. 1996;65:337–365. doi: 10.1146/annurev.bi.65.070196.002005. [DOI] [PubMed] [Google Scholar]
- 79.Kim JH, Park SM, Kang MR, Oh SY, Lee TH, Muller MT, Chung IK. Ubiquitin ligase MKRN1 modulates telomere length homeostasis through a proteolysis of hTERT. Genes Dev. 2005;19(7):776–781. doi: 10.1101/gad.1289405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Renaud S, Loukinov D, Bosman FT, Lobanenkov V, Benhattar J. CTCF binds the proximal exonic region of hTERT and inhibits its transcription. Nucleic Acids Res. 2005;33(21):6850–6860. doi: 10.1093/nar/gki989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Joazeiro CA, Weissman AM. RING finger proteins: mediators of ubiquitin ligase activity. Cell. 2000;102(5):549–552. doi: 10.1016/S0092-8674(00)00077-5. [DOI] [PubMed] [Google Scholar]
- 82.Zemp I, Lingner J. The shelterin component TPP1 is a binding partner and substrate for the deubiquitinating enzyme USP7. J Biol Chem. 2014;289(41):28595–28606. doi: 10.1074/jbc.M114.596056. [DOI] [PMC free article] [PubMed] [Google Scholar]