

Abstract
Microscopy imaging has enabled us to establish the presence of fibrin(ogen) amyloid (fibrinaloid) microclots in a range of chronic, inflammatory diseases. Microclots may also be induced by a variety of purified substances, often at very low concentrations. These molecules include bacterial inflammagens, serum amyloid A, and the S1 spike protein of severe acute respiratory syndrome coronavirus 2. Here, we explore which of the properties of these microclots might be used to contribute to differential clinical diagnoses and prognoses of the various diseases with which they may be associated. Such properties include distributions in their size and number before and after the addition of exogenous thrombin, their spectral properties, the diameter of the fibers of which they are made, their resistance to proteolysis by various proteases, their cross-seeding ability, and the concentration dependence of their ability to bind small molecules including fluorogenic amyloid stains. Measuring these microclot parameters, together with microscopy imaging itself, along with methodologies like proteomics and imaging flow cytometry, as well as more conventional assays such as those for cytokines, might open up the possibility of a much finer use of these microclot properties in generative methods for a future where personalized medicine will be standard procedures in all clotting pathology disease diagnoses.
Keywords: microclots, platelet hyperactivation, inflammatory molecules, vascular complications
The processes of normal blood clotting are well established (for example 1 2 3 4 5 ). The terminal steps involve the self-assembly of fibrinogen molecules that have been cleaved by thrombin (which removes two “fibrinopeptides” 6 7 8 ) into long fibers with a diameter typically in the range of 50 to 100 nm. Some time ago, we established that blood frequently clotted into anomalous forms, referred to in earlier papers as “dense matted deposits” 9 10 11 12 , that were also rather resistant to fibrinolysis (proteolysis). 13 It was subsequently recognized 14 that this anomalous form was in fact amyloid in character 15 and could be stained with well-established, fluorogenic amyloid stains such as thioflavin T 15 16 17 18 19 20 21 22 or the oligothiophenes marketed as “Amytracker” dyes. 16 19 21 We used fluorescence microscopy imaging as our method of choice. To distinguish them from the fibrils in established amyloidosis 23 (commonly with diameters less than 15 nm 24 25 ) and to recognize that they consist mainly of fibrin aggregates (plus other trapped molecules) in the range of 2 to 200 μm and with individual fibers with diameters commonly in the range of 50 to 100 nm or more, we refer to them as fibrinaloid microclots. 17 Such fibrinaloid microclots or fibrin(ogen) aggregates, which “spontaneously” form in the circulation (presumably under the action of thrombin'), have recently been reported in the plasma of patients with type 2 diabetes mellitus (T2DM), 19 26 in those with acute coronavirus disease 2019 (COVID-19) 26 27 28 29 30 and in particular in those with persistent symptoms related to postacute sequelae of COVID (PASC), more commonly known 31 32 33 as long COVID. 17 34 35
In addition, platelets in the hematocrit and whole blood of participants with various inflammatory conditions are well known to be hyperactivated. 27 36 37 38 39 40 41 42 For these studies, we also used fluorescence microscopy imaging as our method of choice for platelet imaging. (After centrifuging whole blood, there are still platelets present in the hematocrit). These platelets might in fact be considered as those that were most fragile in vivo. These microclots and hyperactivated platelets have been implicated in the thrombotic and systemic inflammatory complications of various diseases. In addition, numerous well-known inflammatory molecules have been found trapped inside these insoluble microclots present in acute COVID-19 and long COVID. 17 35 These particular microclots (whose formation can be catalyzed by the severe acute respiratory syndrome coronavirus 2 [SARS-CoV-2] S1 spike protein) 34 are very resistant to digestion protocols. 13 15 35 However, microclots from T2DM (and the few that are always present in plasma samples of healthy participants) digest easily with standard protein digestion protocols. Important questions from these differential results are raised regarding the pathophysiological relevance of microclots and platelet hyperactivation. These findings warrant a discussion to determine if these pathologies are indeed a cause or a consequence of disease and if they are indeed a predictor of clinical thrombosis or contributors to disease pathologies noted in the patient. Further, if these microclots and activated platelets are indeed a predictor of clinical symptoms, including patient disease progression and, in particular, thrombotic endothelialitis, could they ultimately then also become a microscopy imaging monitoring tool to be used to assess the effectiveness and progress of treatment regimens? Finally, an arising question is whether these variables are just innocent bystanders, albeit present in all inflammatory conditions, or a significant element of the etiology of these diseases. In the latter case, one may anticipate significant relationships between the nature and extent of fibrinaloid microclot formation and the severity of the disease. In addition, variants such as omicron that are far less virulent (albeit more transmissible) are—as we have recently shown 43 —far less able to cause microclots. This implies strongly that the microclots are part of the disease pathway. If the latter is in fact occurring, then a naive view might be that all microclots are “the same” and that platelet hyperactivation is universally similar in all conditions. As with any quantitative determination (like microscopy imaging), this is almost certainly not true, and the purpose of the present commentary is to rehearse the evidence for the kinds of differences that are either known or may be anticipated. We also point out the usefulness of microscopy imaging, as such a method allows for a fast and reliable snapshot of clotting pathology, to study both microclot formation and platelet hyperactivation.
The General Pathway of Inflammation, Platelet Hyperactivation, and Microclot Formation in Chronic Disease
Fig. 1 shows the broad swathe of phenomena on which we focus, as they occur in response to an external stimulus 44 such as an infection. It recognizes that a chief set of pathways necessarily involves an external event—commonly an infection—that leads to inflammation (assessed via increases in the levels of inflammatory cytokines) and directly or indirectly to the formation of microclots and the hyperactivation of platelets. These phenomena can then feedback in a kind of autocatalytic cycle (in very unfavorable cases leading to a “cytokine storm” 45 46 ).
Fig. 1.
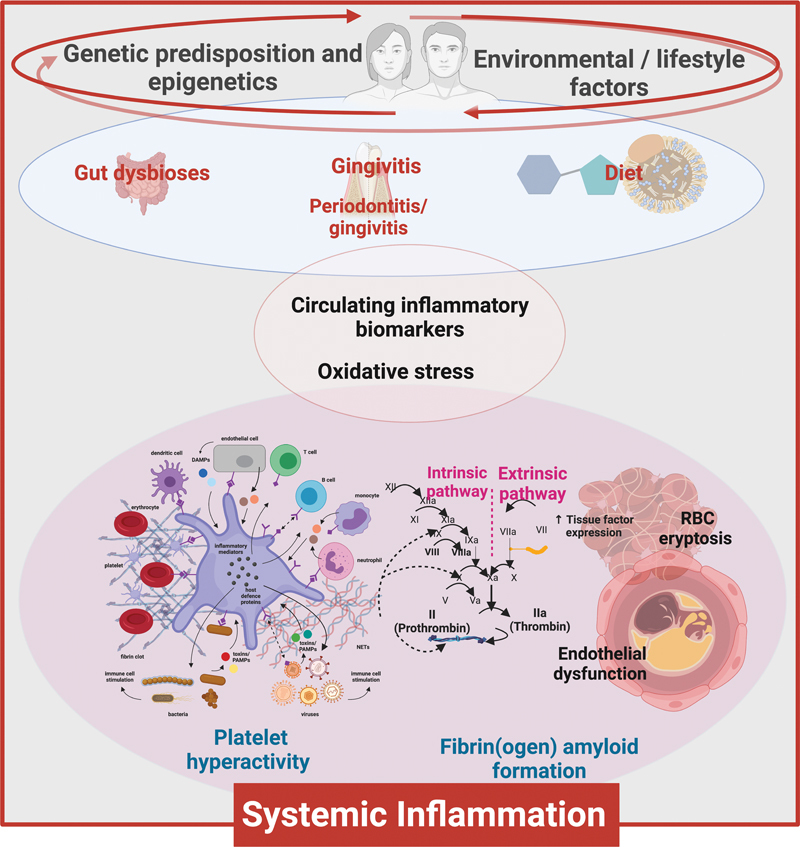
Visualizing various factors that influence disease to the understand the phenomena of the disease due to the response to an external stimulus 44 such as an infection. Figure created by authors using Biorender.com.
Variations in Patterns of Cytokines
The presence and concentration of molecules, 47 including pro- (and anti-) inflammatory molecules vary considerably between individuals, even in “health” 48 49 50 51 , not least because of variations in age 52 and in the gut microbiome. 53 Considerable variation similarly exists in those with the “same” disease, 54 such as an infection 55 (including by SARS-CoV-2 56 57 58 59 60 ), the patterns often correlating with severity and/or outcome. The same is true in myalgic encephalitis/chronic fatigue syndrome 61 and long COVID. 62 Consequently, the presence of a soup of inflammatory molecules in circulation that individually might bind to the numerous receptors on platelets, and have direct protein–protein interactions, may therefore be a crucial predictor of the detailed effects of any systemic inflammation. These interactions should be seen as central to the cause and effect of disease development and presentation of both diverse and overlapping symptoms. Clotting and platelet pathologies should therefore not simply be seen as a predictor of clinical thrombosis. Such assumption would be oversimplifying the pathophysiological value of both identifying and studying the underlying causes of the presence of the inflammatory molecules in circulation.
Differences in the Extent of Platelet Activation
There are many reasons why we might anticipate that platelet activation in chronic inflammatory diseases might have many origins, not least the plethora of receptors on platelets that can be activated individually by numerous circulating inflammatory molecules (see Fig. 2 and Table 1 for a basic list of the types of membrane receptors and their functions). Platelets are mostly seen as participating “only” in clot formation during wound healing, but they are actually most important as signaling entities in immuno-thrombosis. Signaling through different receptors can lead to different degrees of platelet activation; however, when a platelet is activated, it cannot return to its original state. Different diseases and even individuals with the same disease have a unique combination and concentration of inflammatory molecules in circulation. It would only need a few of these molecules to bind to any one or more of the various receptors on platelets, to cause platelet hyperactivation. The exact nature of the inflammatory molecule trigger may be different, but the end results will be an “outside-in” signaling activation that, in turn, results in “inside-out” platelet signaling and platelet hyperactivation. This is therefore a case where platelet activation and the concept of “cause and result” should be discussed with care. Nonetheless, microscopy imaging might be particularly useful in studying platelet hyperactivation, as the preparation methods we employ in our research allows for minimal handling of samples and short timeframes between obtaining the sample and doing the analysis. Fig. 3 shows examples of microscopy imaging of platelets from control and long COVID samples.
Fig. 2.
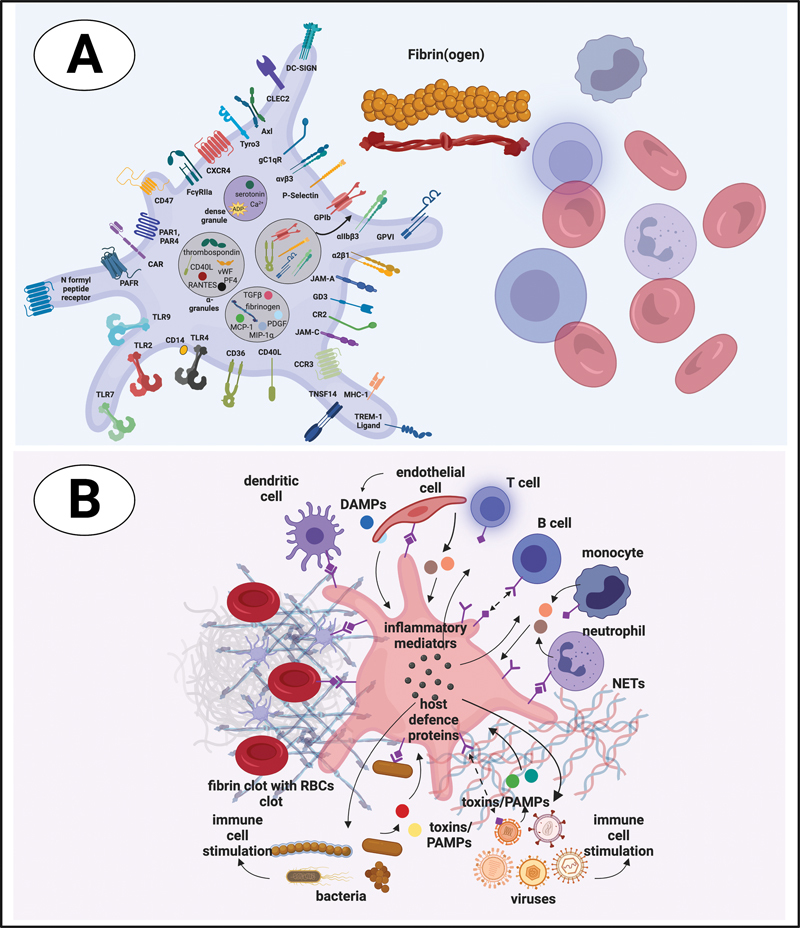
Platelet receptors and interactions with cells and proteins in circulation. Figure created by authors using Biorender.com.
Table 1. Receptor types found on the membranes of platelets and their various functions. For a discussion see 37 .
| Receptor type | Membrane receptor and function | References |
|---|---|---|
| Receptors associated with antigen presentation | • CD40 and CD40L (CD154, also known as CD40 ligand) - is a member of the tumor necrosis factor (TNF) family and CD40-mediated platelet activation is well known in the development of thrombosis, inflammation, and atherosclerosis. Human platelets carry preformed CD40L molecules, which rapidly appear on the platelet surface following stimulation by thrombin. CD40L is on the platelet surface for a short time and does participate in an immune reaction, although mainly by being released in soluble form and being a cytokine and not so much a receptor when the strict definition of a receptor is considered. • Major histocompatibility complex (MHC) class I - MHC (class I) is present on platelets, and platelets directly activate naive T cells in a platelet MHC class I-dependent manner. • Toll like receptors (TLRs)—TLR4 has a prominent functional impact on platelet activity, hemostasis, and thrombosis. • Fc receptor for IgG, [FcγRIIa or CD32])—human platelets express FcγRIIa, the low-affinity receptor for the constant fragment (Fc) of immunoglobulin (Ig) G that is also found on neutrophils, monocytes, and macrophages. Engagement by circulating ligands results in immune complexes that further triggers intracellular signaling events that lead to platelet activation and aggregation. An example of such a complex may be FcγRIIa/Integrin αIIbβ3 that might bind immunoglobulin. FcγRIIa may also be associated with GPIb-IX-V. • Complement receptors—platelets contain complement factors and bear complement receptors. |
123 124 125 126 127 128 129 130 131 132 |
| Activation and modulating platelet receptors | • CD63—this is a dense granule and lysosome membrane glycoprotein. After platelet activation and granule exocytosis, CD63 translocates to the platelet membrane, where it colocalizes with the αIIbβ3-CD9 complex and is incorporated in the cytoskeleton. This allows platelet interactions with other cells, for example, neutrophils. Both CD63 and CD9 are members of the tetraspanin superfamily of integral membrane proteins that functions as signaling complexes. • Glycoprotein VI[GPVI])—this receptor also signals through a immunoreceptor tyrosine-based activation motif, and is involved in platelet activation. It is a platelet-specific transmembrane type I receptor that non-covalently associates with the immunoreceptor tyrosine-based activation motif (ITAM) containing Fc receptor (FcR) γ-chain in the plasma membrane. It is the main collagen receptor. Activation leads to stable platelet adhesion and degranulation of platelet granules. • C-type-lectin-like receptor (CLEC2)—CLEC-2 is a C-type lectin-like type II transmembrane receptor and is involved in platelet aggregate stabilization. It uses a similar signaling pathway as the GPVI/FcRγ-(chain) complex but it involves tyrosine phosphorylation of only a single cytoplasmic YXXL motif. |
133 134 135 136 137 138 139 140 141 142 |
| Adhesion receptors | • GPIb-IX-V (sometimes also written as GPIb-V–IX or GP1b-IX)—This receptor can be classified as both an adhesion and a major signaling receptor, expressed on the surface of circulating platelets. It is composed of four subunits: GPIbα, GPIbβ, GPV, and GPIX. GPIbα and GPIbβ are linked by disulfide bridges, while the GPV and GPIX associate non-covalently with the complex. GPIbα subunit bears the binding site for von Willebrand factor (vWF), leukocyte integrin αMβ2, P-selectin, and the coagulation factors thrombin, and other factors like XI and XII, and Mac-1. GPIb-IX-V may also be associated with FcRγ and with FcγRIIa. • CD147—The extracellular matrix metalloproteinase (CD147) is a member of the immunoglobulin superfamily and is also known as EMMPRIN, localized to the open canalicular system (OCS), and potentially within the α-granules, as its stimulated expression coincided with CD62P release to the platelet surface. Its expression is upregulated in the coronary circulation in patients with stable coronary disease. Its expression is upregulated in response to platelet agonists, including ADP, collagen and thrombin. • P-selectin—It is an important adhesion receptor, and is an adhesion molecule (CD62P) component of the platelet membrane. After platelet activation, P-selectin is translocated from intracellular α granules to the external membrane, therefore α-degranulation results in increased surface expression of P-selectin. Shedding of P-selectin from the platelet membrane can also happen as released soluble ectodomain fragments that are also detectable in plasma. Plasma levels of soluble P-selectin (sP-selectin) are often used to demonstrate platelet activation. P-selectin mediates rolling of platelets and leukocytes on activated endothelial cells. Thrombin may also cause exposure of activated P-selectin. |
131 142 143 144 145 146 147 148 149 150 151 152 153 154 155 |
| Intergrins | • Integrin αVβ3—this is the vitronectin receptor. Vitronectin is a glycoprotein of the hemopexin family. It is also known to assemble fibronectin fibrils on platelets and mediates cell adhesion to the extracellular matrix. • Integrin αIIbβ3—this is a receptor for fibrinogen and von Willebrand Factor (VWF), and activation of this receptor induces platelet aggregation and may play an important role in platelet spreading. Upon platelet activation, inside-out signaling pathways increase the affinity of αIIbβ3 for fibrinogen and other ligands. Inside-out signaling therefore facilitates and initiates the conformational changes responsible for ligand binding. Ligand binding and integrin clustering follows, resulting outside-in signaling , which initiates and amplifies cellular events driving essential platelet processes such as spreading, thrombus consolidation, and clot retraction. • Integrin α2β1—this is a collagen receptor and collagen binds directly or indirectly to both α2β1 (and αIIbβ3, via VWF). • One of the main platelet activators is also thrombin that causes exposure of activated αIIbβ3, αvβ3, α2β1 (also P-selectin as mentioned in the previous section). |
137 155 156 157 158 159 160 161 |
| G-coupled receptors | • Protease-activated receptors (PAR1 and PAR4)—platelets contain both these receptors and PAR1 and PAR4 acts as a dual receptor system for responding to thrombin as ligand. Both PAR1 and PAR4 also signal via ADP. • ADP receptors, P2Y1 and P2Y12—they are involved in platelet activation and aggregation. • The thromboxane A2 (TXA2) receptor—it exists as two isoforms, TPα and TPβ, differing only in their C-terminal region. |
162 163 164 165 166 167 168 169 170 171 |
Fig. 3.
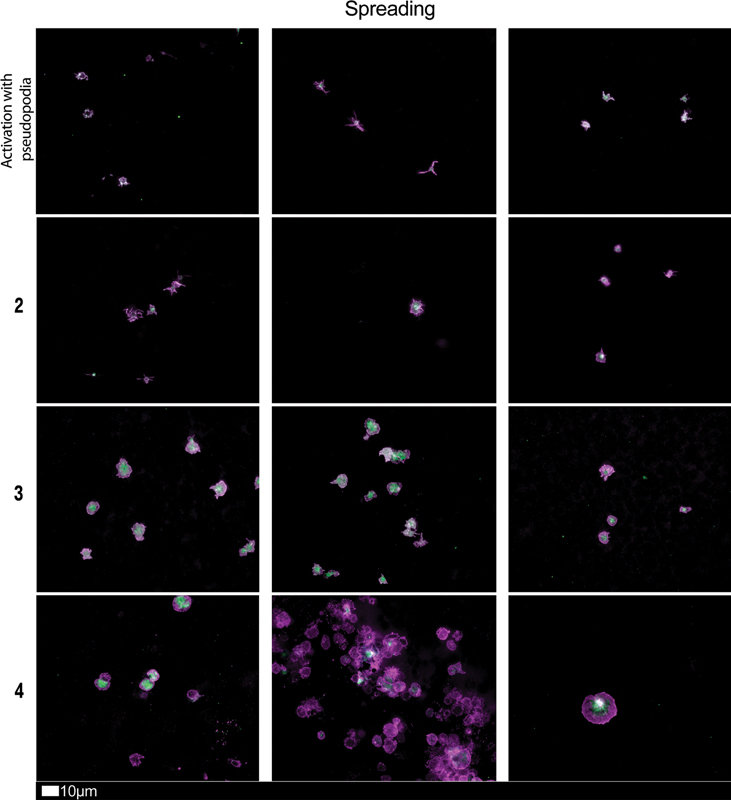
Fluorescence microscopy imaging examples of the different stages of platelet activation and spreading that was used to score platelet activation in long COVID patients. After centrifuging freshly collected samples, the hematocrit fraction of each sample was retained and incubated for 30 minutes at room temperature with the two fluorescent markers, CD62P (PE-conjugated) (platelet surface P-selectin) (IM1759U, Beckman Coulter, Brea, CA) and PAC-1 (FITC-conjugated) (340507, BD Biosciences, San Jose, CA). 10 mL of each exposed sample was placed on microscope slide and viewed using a 63x oil objective. Stage 1, with minimally activated platelets, seen as small round platelets with a few pseudopodia, seen as healthy/control platelets that progresses to Stage 4, with egg-shaped platelets, indicative of spreading and the beginning of clumping (with permission from the CC-BY publication 28 ).
Ten Things We know about the Fibrinaloid Microclots That Can Be Observed in Chronic, Inflammatory Diseases
We think it will be helpful to rehearse some of the known facts about fibrinaloid microclot formation in the form of a “ten things” 63 style ( Table 2 ); also please see Fig. 4 for examples of fluorescence microscopy imaging of microclots. Many proteins can adopt a more thermodynamically stable microstate with no change in the primary structure (sequence), in which the more stable contains an ordered β-sheet “amyloid” structure 15 (see Fig. 5 ). Normally, however, it is present in a less stable state that is kinetically more accessible during and following its synthesis. The more stable (labeled PrPSc) is separated from the initial state (PrPC) via a large energy barrier. This is true for amyloid proteins generally and is illustrated here for classical prion proteins. 17 64
Table 2. The “big picture” of microclots and ideas for future research related to microclots.
| Fibrinaloid microclots are widely present in all chronic, inflammatory diseases studied to date, and to a much greater extent than in ‘healthy’ controls. |
| These diseases include Alzheimer's, Parkinson's, rheumatoid arthritis, and infection with SARS-CoV-2 leading to acute or long COVID-19. |
| They can be stained with fluorogenic dyes such as thioflavin T or the Amytracker dyes. |
| They are commonly in the size range 2–200 μm. |
| They are comparatively resistant to the normal processes of fibrinolysis; some are even resistant to trypsin. |
| Their extent is often related to the severity of various diseases. |
| They can be induced in vitro (in both whole blood and platelet poor plasma) with a variety of substances, including bacterial lipopolysaccharide, lipoteichoic acid, 17-b-oestradiol, and SARS-CoV-2 S1 spike protein. |
| In some cases (especially SARS-Cov-2 infection) they are prevalent even without the addition of thrombin. |
| The diameter of the fibers can vary fairly considerably. |
| They can exhibit considerable structural (and even spectral) heterogeneity, reflecting the molecules that were bound to the fibrinogen before polymerization. |
Abbreviations: COVID-19, coronavirus disease 2019; SARS-Cov-2, severe acute respiratory syndrome coronavirus 2.
Fig. 4.
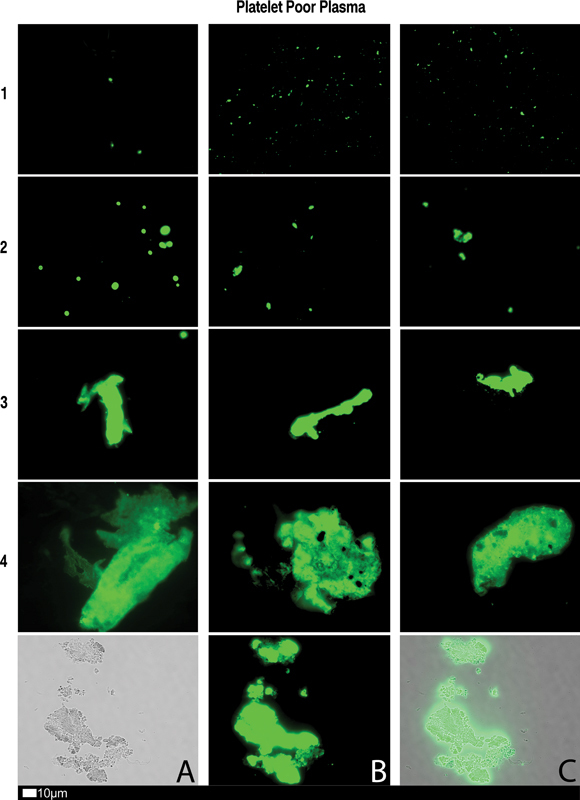
Fluorescence microscopy showing microclots in platelet poor plasma (PPP) with representative examples of the different stages of different stages of microclot formation. Stored or freshly prepared platelet poor plasma samples were exposed to Thioflavin T (ThT), a fluorogenic dye that binds to amyloid protein. 15 A final concentration of 0.005 mM was used (Sigma-Aldrich, St. Louis, MO). Plasma was exposed for 30 minutes (protected from light) at room temperature, whereafter 3 µL stained PPP was placed on a glass slide and covered with a coverslip. Stage 1 shows minimal microclot formation in healthy/control PPP which progresses to the presence of the severe microclotting Stage 4. Bottom row represents examples of stage 4 microclots using ( A ) bright-field microscopy, ( B ) fluorescence microscopy, and ( C ) an overlay of fluorescence and bright-field microscopy (with permission from the CC-BY publication 28 ).
Fig. 5.
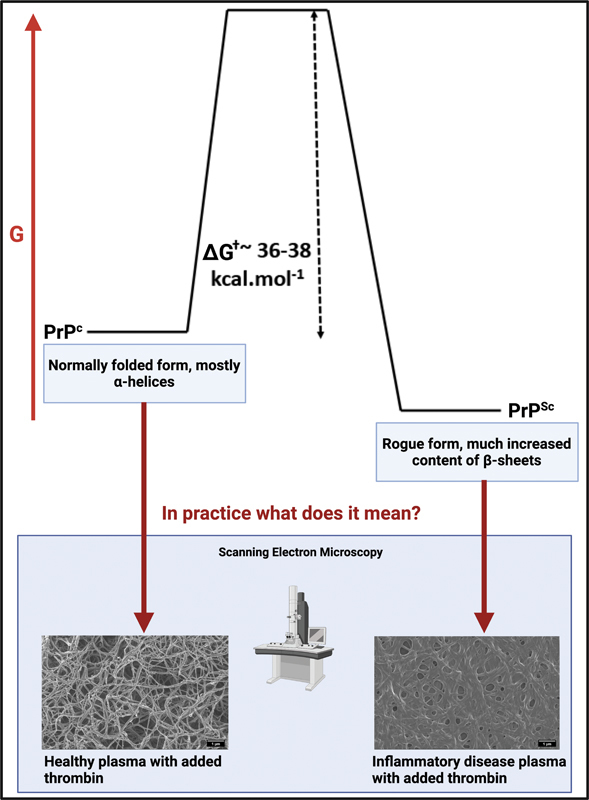
Protein–protein interactions may result in protein misfolding and have been shown to cause amyloidogenic changes to all kinds of proteins. This is illustrated in the upper part for the Prion protein PrP. The lower part shows electron micrographs of fibrin clots. Adapted in part from the CC-BY publications. 15 17 64 We also note that the spike protein is itself amyloidogenic. 172 Figure created by authors using Biorender.com.
Fig. 6 offers a comparative examination of microclot areas reanalyzing fluorescence microscopy data, previously published, 26 extending our prior analysis to include healthy participants, individuals with T2DM, and those currently experiencing acute COVID-19. Reexamining some of our previously collected micrograph data from previously published papers, we also provide evidence that microclots are visible in scanning electron microscopy (SEM) of whole blood samples. Fig. 7 showcases a selection of SEM micrographs of healthy whole blood and conditions where clotting pathologies are well-known. In Fig. 7A , a few plasma deposits (microclots) are discernible among or on the erythrocytes, while Fig. 7B to F show whole blood SEM micrographs of samples from an individual diagnosed with systemic lupus erythematosus ( Fig. 7B ), acute COVID-19 during the initial wave in 2020 ( Figs. 7C and D ), rheumatoid arthritis ( Fig. 7E ), and a sample from a patient diagnosed with Alzheimer's-type dementia ( Fig. 7F ). We only realized that these deposits were of significant importance during the last few years.
Fig. 6.
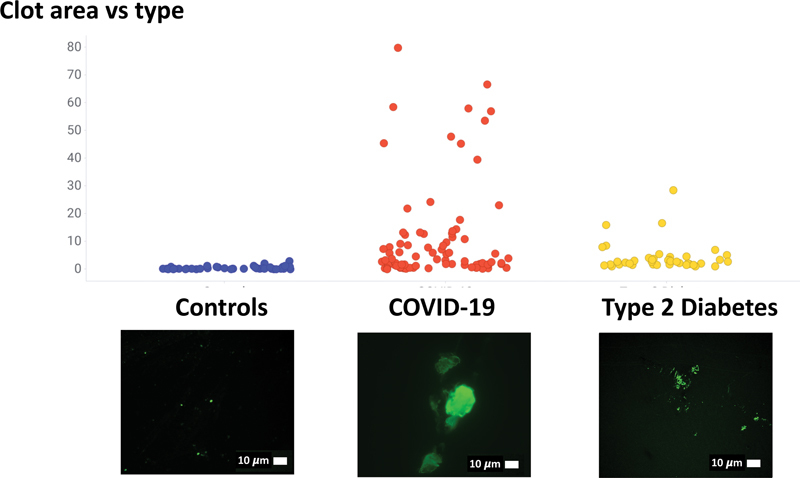
Comparison of microclot area in healthy participants, and participants with Type 2 diabetes and acute COVID-19. (Raw data reanalyzed, and available in 26 ).
Fig. 7.
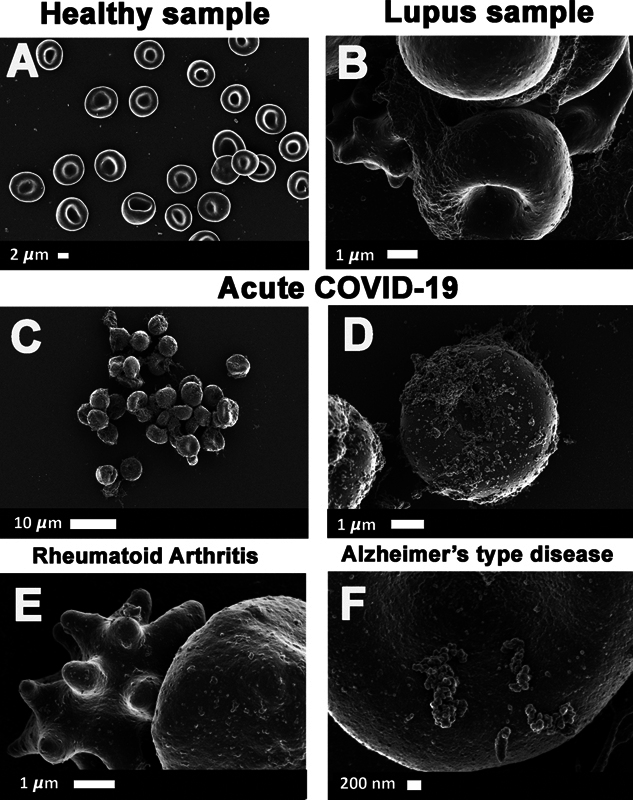
( A to F ) Whole blood scanning electron microscopy of micrograph data previously collected in published studies. ( A ) A few plasma deposits are discernible within the blood sample of a healthy participant 173 ; ( B ) systemic lupus erythematosus 174 ; ( C and D ) acute COVID-19 during the initial wave in 2020 30 ; ( E ) rheumatoid Arthritis 175 ; ( F ) Alzheimer's-type disease. 176
Variation in Microclot Properties
We remain relatively ignorant of the details of precisely what governs fibrin self-assembly following fibrinopeptide release (e.g., why it stops at a certain and nonconstant fiber diameter, even in health, 65 and why it can vary with disease 66 ). One thing we do know, however, is that microclots covary with (and in our view are thus largely responsible for) the severity of disease in both acute and long COVID 67 and in fact contribute to widespread thrombotic endothelialitis in long COVID. 62
What Kind of Molecules can Promote Fibrinaloid Formation?
Since the early discoveries that molecules such as oestrogens 68 69 could induce the formation during clotting of “dense matted deposits” (that we now refer to as fibrinaloid microclots, because of their amyloid character), we have also found that low concentrations of molecules such as bacterial lipopolysaccharide, 14 lipoteichoic acid, 21 serum amyloid A, 70 and SARS-CoV-2 S1 spike protein 34 can also do so. These dense matted deposits could be induced using both whole blood and platelet-poor plasma. It is moderately to highly unlikely that they are binding in the same locations as each other (the binding site of spike is known 71 ), and thus, the morphology of the clots they induce will be the same. In a similar vein, nonamyloid substances can also bind to and induce the aggregation of established amyloid proteins such as bacterial DNA promoting tau 72 or β-amyloid 73 aggregation.
Consequences of Microclot Formation
The fibrinaloid microclots that we discovered and that can be induced my miniscule amounts of molecules such as bacterial LPS or the SARS-CoV-2 spike protein are resistant to normal fibrinolysis for a least two reasons: (1) amyloid-type proteins are inherently more resistant to proteolysis 35 74 because of their crossed-β structure, 15 75 76 and (2) because they also entrap inhibitors of normal proteolysis such as α2-antiplasmin and plasminogen activator inhibitor 1 (PAI-1). 74 Consequently, they are able to block microcapillaries, leading to tissue ischemia and hypoxia, from which the great majority of symptoms can be seen to follow. 17 77 Importantly, our analysis provides a mechanistic link between external trigger events and the pathologies of present interest, as well as a candidate set of targets for treatment.
Anticipating Disease Diagnosis and Prognosis from Microclot Measurements, as Part of Disentangling Their Differences
To date, our microscopy imaging and analyses have mostly been semiquantitative, showing the existence of different microclots in various diseases. As with much of modern, postgenomic biology, 78 especially that influenced by “deep learning” 79 , it is time to move from a “discriminative” strategy. We suggest a “generative” strategy, in which we seek to solve the “inverse problem” (as in 80 ) by using the observables (microclot properties) to infer their “cause” (i.e., the diseases with which they are associated). The observables include the distribution of clot sizes and morphologies, 26 81 the diameter of individual fibers, 82 83 their ability to cross-seed other amyloidogenic proteins, 84 85 86 87 88 89 the spectral properties of the different stains when attached to fibers, 90 91 92 93 94 95 96 the susceptibility of the clots to proteolysis by different proteases, 35 97 and the extent to which they are naturally present in platelet-poor plasma versus being induced by the in vitro addition of thrombin. 17 34 98
The Power of Multivariate Data, and How the Manner of Fibrinogen Clotting Effects Dimensionality Reduction of the Various Plasma 'omes
Each property of an individual example in a system of interest can be seen as an element of a vector describing that system or as a dimension in multidimensional space. 99 100 101 When we have a series of properties of both a sample “as a whole” and indeed of individual objects therein, it becomes increasingly easy to discriminate them. 100 Care is needed, however, 102 since even comparatively small random variations in normal distributions can appear significant in individual dimensions when there are many to choose from, even in “unsupervised” methods (in which class membership, such as a particular disease, is not known 103 ). In favorable cases (e.g., 104 105 ), though, these are entirely sufficient. Supervised methods, in which a model is “trained” to predict an output set of properties (such as a particular disease) from “input” variables such as omics data, are even more prone to overtraining and other kinds of bias. 102 The solution here (e.g., 106 ) is to assess any predictions taken from such a mathematical model using separate “validation” examples whose class membership or other output properties are known but which are not used in the construction of the model.
Another modern trend is to recognize that the amount of unlabeled data available normally vastly exceeds that of labeled data and that such data can be used in the training of a supervised model; these methods are known as “semisupervised.” They have become preeminent in deep learning models 79 based on variational autoencoders 107 108 109 and transformers, 110 111 especially in natural language 112 113 and image processing. 114 115 116
In variational autoencoders, one essentially clusters input examples into a (much) lower dimensional space. This still allows considerable discrimination; however, even a normalized vector of just 20 in which an individual may be in the upper or lower half admits 2 20 (approximately 1 million) possibilities. 109 117 In the case of microclots, we consider that (distributions in) the many 1,000s of individual metabolites 118 119 120 and proteins 121 in serum or plasma can potentially each affect the size and shape of the microclots. This “harvesting” of all the molecules to which the fibrinogen is exposed, and which then determines how it polymerizes, effectively concentrates the vast numbers of metabolites, proteins, and even transcripts into a smaller number of dimensions; the microclots essentially act as a surrogate for the metabolome, proteome, and transcriptome present in the plasma at the time of clotting. Consequently, we are optimistic that an analysis of the detailed morphological and spectral properties of microclots will indeed serve to discriminate, possibly quite finely, individuals with different diseases.
Conclusion
In our previous work, we have focused more or less qualitatively on the presence of fibrinaloid microclots that we discovered using microscopy imaging while recognizing that some of their measurable properties differ in various conditions or diseases. Within the terminology of the deep learning agenda, 79 this is to be seen as a “discriminative” approach. The opposite strategy, amounting to the solution of the inverse problem, is referred to in general as a “generative” strategy. Here the aim, now clearly worthwhile, is to develop and exploit a more quantitative analysis in the prediction of diseased states, and the success of any treatment both in modifying the microclot properties and in curing or ameliorating the diseases. Various laboratories in the United States, United Kingdom, and Germany have successfully implemented microclot imaging and will also be publishing their results shortly. We have been using and described here, fluorescence microscopy imaging (and also briefly SEM methods) to visualize microclot presence and platelet hyperactivation. In addition, we are also developing imaging flow cytometry methods that may, in future, direct a meaningful translational outcome that could guide treatment options. We also suggest that automation of our microscopy imaging using software modalities like the MetaSystems platform ( https://metasystems-international.com/ ), might be particularly useful for unbiased quantitative analysis of microscopy imaging of both platelets and microclots. Other research teams are developing innovative strategies that integrate real-time one-dimensional-imaging fluorescence with deformability cytometry within a single instrument, denoted as RT-FDC. 122 RT-DC techniques are also currently being adapted to study and characterize the mechanical classification of microclots in whole blood, at rates of several hundred cells per second. We believe that the research agenda, using various novel methods, as set out here, will make the enterprise of unbiased quantitative analysis of microclots, worthwhile. We suggest that researchers direct increased attention toward a previously disregarded fraction of the blood sample. Microclots found in platelet-poor plasma could hold a significant diagnostic value for clotting-related issues in inflammatory diseases and other similar conditions, including postviral syndromes.
Funding Statement
Funding D.B.K. would like to thank the Novo Nordisk Foundation (grant NNF20CC0035580) and Balvi Research Foundation and funding. E.P. would like to thank the NRF of South Africa (grant number 142142), SA MRC (self-initiated research [SIR] grant), Balvi Research Foundation, and Polybio Research Foundation for funding. The content and findings reported and illustrated are the sole deduction, view, and responsibility of the researchers and do not reflect the official position and sentiments of the funders.
Conflict of Interest
E.P. is a director of Biocode Technologies, a Stellenbosch University start-up company. D.B.K. has no interests to declare.
References
- 1.Weisel J W. Fibrinogen and fibrin. Adv Protein Chem. 2005;70:247–299. doi: 10.1016/S0065-3233(05)70008-5. [DOI] [PubMed] [Google Scholar]
- 2.Adams R L, Bird R J. Review article: coagulation cascade and therapeutics update: relevance to nephrology. Part 1: overview of coagulation, thrombophilias and history of anticoagulants. Nephrology (Carlton) 2009;14(05):462–470. doi: 10.1111/j.1440-1797.2009.01128.x. [DOI] [PubMed] [Google Scholar]
- 3.Undas A, Ariëns R AS. Fibrin clot structure and function: a role in the pathophysiology of arterial and venous thromboembolic diseases. Arterioscler Thromb Vasc Biol. 2011;31(12):e88–e99. doi: 10.1161/ATVBAHA.111.230631. [DOI] [PubMed] [Google Scholar]
- 4.Wolberg A S. Thrombin generation and fibrin clot structure. Blood Rev. 2007;21(03):131–142. doi: 10.1016/j.blre.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 5.Palta S, Saroa R, Palta A. Overview of the coagulation system. Indian J Anaesth. 2014;58(05):515–523. doi: 10.4103/0019-5049.144643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mullin J L, Gorkun O V, Binnie C G, Lord S T. Recombinant fibrinogen studies reveal that thrombin specificity dictates order of fibrinopeptide release. J Biol Chem. 2000;275(33):25239–25246. doi: 10.1074/jbc.M004142200. [DOI] [PubMed] [Google Scholar]
- 7.Pechik I, Yakovlev S, Mosesson M W, Gilliland G L, Medved L. Structural basis for sequential cleavage of fibrinopeptides upon fibrin assembly. Biochemistry. 2006;45(11):3588–3597. doi: 10.1021/bi0525369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Riedel T, Suttnar J, Brynda E, Houska M, Medved L, Dyr J E. Fibrinopeptides A and B release in the process of surface fibrin formation. Blood. 2011;117(05):1700–1706. doi: 10.1182/blood-2010-08-300301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lipinski B, Pretorius E. Novel pathway of iron–induced blood coagulation: implications for diabetes mellitus and its complications. Pol Arch Med Wewn. 2012;122(03):115–122. [PubMed] [Google Scholar]
- 10.Pretorius E, Vermeulen N, Bester J, Lipinski B, Kell D B. A novel method for assessing the role of iron and its functional chelation in fibrin fibril formation: the use of scanning electron microscopy. Toxicol Mech Methods. 2013;23(05):352–359. doi: 10.3109/15376516.2012.762082. [DOI] [PubMed] [Google Scholar]
- 11.Pretorius E, Lipinski B. Differences in morphology of fibrin clots induced with thrombin and ferric ions and its pathophysiological consequences. Heart Lung Circ. 2013;22(06):447–449. doi: 10.1016/j.hlc.2012.10.010. [DOI] [PubMed] [Google Scholar]
- 12.Swanepoel A C, Visagie A, de Lange Z, Emmerson O, Nielsen V G, Pretorius E. The clinical relevance of altered fibrinogen packaging in the presence of 17β-estradiol and progesterone. Thromb Res. 2016;146:23–34. doi: 10.1016/j.thromres.2016.08.022. [DOI] [PubMed] [Google Scholar]
- 13.Kell D B, Pretorius E. The simultaneous occurrence of both hypercoagulability and hypofibrinolysis in blood and serum during systemic inflammation, and the roles of iron and fibrin(ogen) Integr Biol. 2015;7(01):24–52. doi: 10.1039/c4ib00173g. [DOI] [PubMed] [Google Scholar]
- 14.Pretorius E, Mbotwe S, Bester J, Robinson C J, Kell D B. Acute induction of anomalous and amyloidogenic blood clotting by molecular amplification of highly substoichiometric levels of bacterial lipopolysaccharide. J R Soc Interface. 2016;13(122):2.0160539E7. doi: 10.1098/rsif.2016.0539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kell D B, Pretorius E. Proteins behaving badly. Substoichiometric molecular control and amplification of the initiation and nature of amyloid fibril formation: lessons from and for blood clotting. Prog Biophys Mol Biol. 2017;123:16–41. doi: 10.1016/j.pbiomolbio.2016.08.006. [DOI] [PubMed] [Google Scholar]
- 16.de Waal G M, Engelbrecht L, Davis T, de Villiers W JS, Kell D B, Pretorius E. Correlative light-electron microscopy detects lipopolysaccharide and its association with fibrin fibres in Parkinson's disease, Alzheimer's disease and type 2 diabetes mellitus. Sci Rep. 2018;8(01):16798. doi: 10.1038/s41598-018-35009-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kell D B, Laubscher G J, Pretorius E. A central role for amyloid fibrin microclots in long COVID/PASC: origins and therapeutic implications. Biochem J. 2022;479(04):537–559. doi: 10.1042/BCJ20220016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Palhano F L, Lee J, Grimster N P, Kelly J W. Toward the molecular mechanism(s) by which EGCG treatment remodels mature amyloid fibrils. J Am Chem Soc. 2013;135(20):7503–7510. doi: 10.1021/ja3115696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pretorius E, Page M J, Engelbrecht L, Ellis G C, Kell D B. Substantial fibrin amyloidogenesis in type 2 diabetes assessed using amyloid-selective fluorescent stains. Cardiovasc Diabetol. 2017;16(01):141. doi: 10.1186/s12933-017-0624-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pretorius E, Page M J, Mbotwe S, Kell D B. Lipopolysaccharide-binding protein (LBP) can reverse the amyloid state of fibrin seen or induced in Parkinson's disease. PLoS One. 2018;13(03):e0192121. doi: 10.1371/journal.pone.0192121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pretorius E, Page M J, Hendricks L, Nkosi N B, Benson S R, Kell D B. Both lipopolysaccharide and lipoteichoic acids potently induce anomalous fibrin amyloid formation: assessment with novel Amytracker™ stains. J R Soc Interface. 2018;15(139):2.0170941E7. doi: 10.1098/rsif.2017.0941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pretorius E, Bester J, Page M J, Kell D B. The potential of LPS-binding protein to reverse amyloid formation in plasma fibrin of individuals with Alzheimer-type dementia. Front Aging Neurosci. 2018;10:257. doi: 10.3389/fnagi.2018.00257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Eisenberg D, Jucker M. The amyloid state of proteins in human diseases. Cell. 2012;148(06):1188–1203. doi: 10.1016/j.cell.2012.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Serpell L C, Sunde M, Benson M D, Tennent G A, Pepys M B, Fraser P E. The protofilament substructure of amyloid fibrils. J Mol Biol. 2000;300(05):1033–1039. doi: 10.1006/jmbi.2000.3908. [DOI] [PubMed] [Google Scholar]
- 25.Serpell L C. Alzheimer's amyloid fibrils: structure and assembly. Biochim Biophys Acta. 2000;1502(01):16–30. doi: 10.1016/s0925-4439(00)00029-6. [DOI] [PubMed] [Google Scholar]
- 26.Pretorius E, Venter C, Laubscher G J, Lourens P J, Steenkamp J, Kell D B. Prevalence of readily detected amyloid blood clots in 'unclotted' Type 2 Diabetes Mellitus and COVID-19 plasma: a preliminary report. Cardiovasc Diabetol. 2020;19(01):193. doi: 10.1186/s12933-020-01165-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Grobler C, Maphumulo S C, Grobbelaar L M et al. COVID-19: the rollercoaster of fibrin(Ogen), D-Dimer, Von Willebrand factor, P-selectin and their interactions with endothelial cells, platelets and erythrocytes. Int J Mol Sci. 2020;21(14):5168. doi: 10.3390/ijms21145168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Laubscher G J, Lourens P J, Venter C, Kell D B, Pretorius E. TEG ® , microclot and platelet mapping for guiding early management of severe COVID-19 coagulopathy . J Clin Med. 2021;10(22):5381. doi: 10.3390/jcm10225381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pretorius E, Vlok M, Venter Cet al. Persistent clotting protein pathology in Long COVID/ post-acute sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasminCardiovasc Diabetol 2021;20(172). Doi: org/10.1186/s12933-021-01359-7 [DOI] [PMC free article] [PubMed]
- 30.Venter C, Bezuidenhout J A, Laubscher G J et al. Erythrocyte, platelet, serum ferritin and P-selectin pathophysiology implicated in severe hypercoagulation and vascular complications in COVID-19. Int J Mol Sci. 2020;21(21):8234. doi: 10.3390/ijms21218234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Akbarialiabad H, Taghrir M H, Abdollahi A et al. Long COVID, a comprehensive systematic scoping review. Infection. 2021;49(06):1163–1186. doi: 10.1007/s15010-021-01666-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Davis H E, Assaf G S, McCorkell L et al. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. EClinicalMedicine. 2021;38:101019. doi: 10.1016/j.eclinm.2021.101019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Proal A D, VanElzakker M B. Long COVID or post-acute sequelae of COVID-19 (PASC): an overview of biological factors that may contribute to persistent symptoms. Front Microbiol. 2021;12:698169. doi: 10.3389/fmicb.2021.698169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Grobbelaar L M, Venter C, Vlok M et al. SARS-CoV-2 spike protein S1 induces fibrin(ogen) resistant to fibrinolysis: implications for microclot formation in COVID-19. Biosci Rep. 2021;41(08):BSR20210611. doi: 10.1042/BSR20210611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pretorius E, Vlok M, Venter C et al. Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc Diabetol. 2021;20(01):172. doi: 10.1186/s12933-021-01359-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Olumuyiwa-Akeredolu O O, Page M J, Soma P, Pretorius E. Platelets: emerging facilitators of cellular crosstalk in rheumatoid arthritis. Nat Rev Rheumatol. 2019;15(04):237–248. doi: 10.1038/s41584-019-0187-9. [DOI] [PubMed] [Google Scholar]
- 37.Pretorius E. Platelets as potent signaling entities in type 2 diabetes mellitus. Trends Endocrinol Metab. 2019;30(08):532–545. doi: 10.1016/j.tem.2019.05.003. [DOI] [PubMed] [Google Scholar]
- 38.Chu S G, Becker R C, Berger P B et al. Mean platelet volume as a predictor of cardiovascular risk: a systematic review and meta-analysis. J Thromb Haemost. 2010;8(01):148–156. doi: 10.1111/j.1538-7836.2009.03584.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Fintel D J. Oral antiplatelet therapy for atherothrombotic disease: overview of current and emerging treatment options. Vasc Health Risk Manag. 2012;8:77–89. doi: 10.2147/VHRM.S26030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Guo L, Rondina M T. The era of thromboinflammation: platelets are dynamic sensors and effector cells during infectious diseases. Front Immunol. 2019;10:2204. doi: 10.3389/fimmu.2019.02204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mancuso M E, Santagostino E. Platelets: much more than bricks in a breached wall. Br J Haematol. 2017;178(02):209–219. doi: 10.1111/bjh.14653. [DOI] [PubMed] [Google Scholar]
- 42.Ziegler M, Wang X, Peter K. Platelets in cardiac ischaemia/reperfusion injury: a promising therapeutic target. Cardiovasc Res. 2019;115(07):1178–1188. doi: 10.1093/cvr/cvz070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Grobbelaar L M, Kruger A, Venter C et al. Relative hypercoagulopathy of the SARS-CoV-2 beta and delta variants when compared to the less severe omicron variants is related to TEG parameters, the extent of fibrin amyloid microclots, and the severity of clinical illness. Semin Thromb Hemost. 2022;48(07):858–868. doi: 10.1055/s-0042-1756306. [DOI] [PubMed] [Google Scholar]
- 44.Kell D B, Pretorius E. No effects without causes: the iron dysregulation and dormant microbes hypothesis for chronic, inflammatory diseases. Biol Rev Camb Philos Soc. 2018;93(03):1518–1557. doi: 10.1111/brv.12407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kell D B, Pretorius E. To what extent are the terminal stages of sepsis, septic shock, SIRS, and multiple organ dysfunction syndrome actually driven by a toxic prion/amyloid form of fibrin? Semin Thromb Hemost. 2018;44:224–238. doi: 10.1055/s-0037-1604108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fajgenbaum D C, June C H. Cytokine storm. N Engl J Med. 2020;383(23):2255–2273. doi: 10.1056/NEJMra2026131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Williams R J. New York: John Wiley; 1956. Biochemical Individuality. [Google Scholar]
- 48.Kleiner G, Marcuzzi A, Zanin V, Monasta L, Zauli G. Cytokine levels in the serum of healthy subjects. Mediators Inflamm. 2013;2013:434010. doi: 10.1155/2013/434010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Li Y, Oosting M, Smeekens S P et al. A functional genomics approach to understand variation in cytokine production in humans. Cell. 2016;167(04):1099–1.11E17. doi: 10.1016/j.cell.2016.10.017. [DOI] [PubMed] [Google Scholar]
- 50.Wu D, Dinh T L, Bausk B P, Walt D R. Long-term measurements of human inflammatory cytokines reveal complex baseline variations between individuals. Am J Pathol. 2017;187(12):2620–2626. doi: 10.1016/j.ajpath.2017.08.007. [DOI] [PubMed] [Google Scholar]
- 51.Schirmer M, Kumar V, Netea M G, Xavier R J. The causes and consequences of variation in human cytokine production in health. Curr Opin Immunol. 2018;54:50–58. doi: 10.1016/j.coi.2018.05.012. [DOI] [PubMed] [Google Scholar]
- 52.Koelman L, Pivovarova-Ramich O, Pfeiffer A FH, Grune T, Aleksandrova K. Cytokines for evaluation of chronic inflammatory status in ageing research: reliability and phenotypic characterisation. Immun Ageing. 2019;16:11. doi: 10.1186/s12979-019-0151-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schirmer M, Smeekens S P, Vlamakis H et al. Linking the human gut microbiome to inflammatory cytokine production capacity. Cell. 2016;167(04):1125–1. doi: 10.1016/j.cell.2016.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Liu C, Chu D, Kalantar-Zadeh K, George J, Young H A, Liu G. Cytokines: from clinical significance to quantification. Adv Sci (Weinh) 2021;8(15):e2004433. doi: 10.1002/advs.202004433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Li Y, Oosting M, Deelen P et al. Inter-individual variability and genetic influences on cytokine responses to bacteria and fungi. Nat Med. 2016;22(08):952–960. doi: 10.1038/nm.4139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mudd P A, Crawford J C, Turner J S et al. Distinct inflammatory profiles distinguish COVID-19 from influenza with limited contributions from cytokine storm. Sci Adv. 2020;6(50):eabe3024. doi: 10.1126/sciadv.abe3024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Del Valle D M, Kim-Schulze S, Huang H H et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat Med. 2020;26(10):1636–1643. doi: 10.1038/s41591-020-1051-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Guo J, Wang S, Xia H et al. Cytokine signature associated with disease severity in COVID-19. Front Immunol. 2021;12:681516. doi: 10.3389/fimmu.2021.681516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wang S Y, Takahashi T, Pine A B et al. Challenges in interpreting cytokine data in COVID-19 affect patient care and management. PLoS Biol. 2021;19(08):e3001373. doi: 10.1371/journal.pbio.3001373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tjan L H, Furukawa K, Nagano T et al. Early differences in cytokine production by severity of coronavirus disease 2019. J Infect Dis. 2021;223(07):1145–1149. doi: 10.1093/infdis/jiab005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Montoya J G, Holmes T H, Anderson J N et al. Cytokine signature associated with disease severity in chronic fatigue syndrome patients. Proc Natl Acad Sci U S A. 2017;114(34):E7150–E7158. doi: 10.1073/pnas.1710519114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Turner S, Naidoo C A, Usher T J et al. Increased levels of inflammatory and endothelial biomarkers in blood of long COVID patients point to thrombotic endothelialitis. Semin Thromb Hemost. 2023;50(02):288–294. doi: 10.1055/s-0043-1769014. [DOI] [PubMed] [Google Scholar]
- 63.Kohane I S. HEALTH CARE POLICY. Ten things we have to do to achieve precision medicine. Science. 2015;349(6243):37–38. doi: 10.1126/science.aab1328. [DOI] [PubMed] [Google Scholar]
- 64.Kell D B, Pretorius E. Are fibrinaloid microclots a cause of autoimmunity in Long Covid and other post-infection diseases? Biochem J. 2023;480(15):1217–1240. doi: 10.1042/BCJ20230241. [DOI] [PubMed] [Google Scholar]
- 65.Bateman R M, Ellis C G, Suematsu M, Walley K R. S-nitrosoglutathione acts as a small molecule modulator of human fibrin clot architecture. PLoS One. 2012;7(08):e43660. doi: 10.1371/journal.pone.0043660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Bezuidenhout J, Venter C, Roberts Tet al. The atypical fibrin fibre network in rheumatoid arthritis and its relation to autoimmunity, inflammation and thrombosis bioRxiv 2020. May302020–5. [Google Scholar]
- 67.Turner S, Khan M A, Putrino D, Woodcock A, Kell D B, Pretorius E. Long COVID: pathophysiological factors and abnormalities of coagulation. Trends Endocrinol Metab. 2023;34(06):321–344. doi: 10.1016/j.tem.2023.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Pretorius E, Oberholzer H M, van der Spuy W J, Meiring J H. The changed ultrastructure of fibrin networks during use of oral contraception and hormone replacement. J Thromb Thrombolysis. 2010;30(04):502–506. doi: 10.1007/s11239-010-0502-4. [DOI] [PubMed] [Google Scholar]
- 69.Swanepoel A C, Lindeque B G, Swart P J, Abdool Z, Pretorius E. Estrogen causes ultrastructural changes of fibrin networks during the menstrual cycle: a qualitative investigation. Microsc Res Tech. 2014;77(08):594–601. doi: 10.1002/jemt.22378. [DOI] [PubMed] [Google Scholar]
- 70.Page M J, Thomson G JA, Nunes J M et al. Serum amyloid A binds to fibrin(ogen), promoting fibrin amyloid formation. Sci Rep. 2019;9(01):3102. doi: 10.1038/s41598-019-39056-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ryu J K, Sozmen E G, Dixit Ket al. SARS-CoV-2 spike protein induces abnormal inflammatory blood clots neutralized by fibrin immunotherapy bioRxiv 2021. Oct312021–10. [Google Scholar]
- 72.Tetz G, Pinho M, Pritzkow S, Mendez N, Soto C, Tetz V. Bacterial DNA promotes Tau aggregation. Sci Rep. 2020;10(01):2369. doi: 10.1038/s41598-020-59364-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tetz G, Tetz V. Bacterial extracellular DNA promotes β-amyloid aggregation. Microorganisms. 2021;9(06):1301. doi: 10.3390/microorganisms9061301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kruger A, Vlok M, Turner S et al. Proteomics of fibrin amyloid microclots in long COVID/post-acute sequelae of COVID-19 (PASC) shows many entrapped pro-inflammatory molecules that may also contribute to a failed fibrinolytic system. Cardiovasc Diabetol. 2022;21(01):190. doi: 10.1186/s12933-022-01623-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Nasica-Labouze J, Nguyen P H, Sterpone F et al. Amyloid β protein and Alzheimer's disease: when computer simulations complement experimental studies. Chem Rev. 2015;115(09):3518–3563. doi: 10.1021/cr500638n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Serpell L C, Benson M, Liepnieks J J, Fraser P E. Structural analyses of fibrinogen amyloid fibrils. Amyloid. 2007;14(03):199–203. doi: 10.1080/13506120701461111. [DOI] [PubMed] [Google Scholar]
- 77.Kell D B, Pretorius E. The potential role of ischaemia-reperfusion injury in chronic, relapsing diseases such as rheumatoid arthritis, long COVID, and ME/CFS: evidence, mechanisms, and therapeutic implications. Biochem J. 2022;479(16):1653–1708. doi: 10.1042/BCJ20220154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kell D B, Oliver S G. Here is the evidence, now what is the hypothesis? The complementary roles of inductive and hypothesis-driven science in the post-genomic era. BioEssays. 2004;26(01):99–105. doi: 10.1002/bies.10385. [DOI] [PubMed] [Google Scholar]
- 79.Kell D B, Samanta S, Swainston N. Deep learning and generative methods in cheminformatics and chemical biology: navigating small molecule space intelligently. Biochem J. 2020;477(23):4559–4580. doi: 10.1042/BCJ20200781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Shrivastava A D, Swainston N, Samanta S, Roberts I, Wright Muelas M, Kell D B. MassGenie: a transformer-based deep learning method for identifying small molecules from their mass spectra. Biomolecules. 2021;11(12):1793. doi: 10.3390/biom11121793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Meng F, Yoo J, Chung H S. Single-molecule fluorescence imaging and deep learning reveal highly heterogeneous aggregation of amyloid-β 42. Proc Natl Acad Sci U S A. 2022;119(12):e2116736119. doi: 10.1073/pnas.2116736119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Collet J P, Park D, Lesty C et al. Influence of fibrin network conformation and fibrin fiber diameter on fibrinolysis speed: dynamic and structural approaches by confocal microscopy. Arterioscler Thromb Vasc Biol. 2000;20(05):1354–1361. doi: 10.1161/01.atv.20.5.1354. [DOI] [PubMed] [Google Scholar]
- 83.Bezuidenhout J, Venter C, Roberts T et al. Detection of citrullinated fibrin in plasma clots of RA patients and its relation to altered structural clot properties, disease-related inflammation and prothrombotic tendency. Front Immunol. 2020;11:577523. doi: 10.3389/fimmu.2020.577523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Furukawa Y, Kaneko K, Nukina N. Tau protein assembles into isoform- and disulfide-dependent polymorphic fibrils with distinct structural properties. J Biol Chem. 2011;286(31):27236–27246. doi: 10.1074/jbc.M111.248963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Sarell C J, Stockley P G, Radford S E. Assessing the causes and consequences of co-polymerization in amyloid formation. Prion. 2013;7(05):359–368. doi: 10.4161/pri.26415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Hu R, Zhang M, Chen H, Jiang B, Zheng J. Cross-seeding interaction between β-amyloid and human islet amyloid polypeptide. ACS Chem Neurosci. 2015;6(10):1759–1768. doi: 10.1021/acschemneuro.5b00192. [DOI] [PubMed] [Google Scholar]
- 87.Morales R, Moreno-Gonzalez I, Soto C. Cross-seeding of misfolded proteins: implications for etiology and pathogenesis of protein misfolding diseases. PLoS Pathog. 2013;9(09):e1003537. doi: 10.1371/journal.ppat.1003537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ono K, Takahashi R, Ikeda T, Yamada M. Cross-seeding effects of amyloid β-protein and α-synuclein. J Neurochem. 2012;122(05):883–890. doi: 10.1111/j.1471-4159.2012.07847.x. [DOI] [PubMed] [Google Scholar]
- 89.Zhang M, Hu R, Chen H et al. Polymorphic cross-seeding amyloid assemblies of amyloid-β and human islet amyloid polypeptide. Phys Chem Phys. 2015;17(35):23245–23256. doi: 10.1039/c5cp03329b. [DOI] [PubMed] [Google Scholar]
- 90.Hammarström P, Lindgren M, Nilsson K PR. Weinheim: Wiley-VCH; 2013. Fluorescence spectroscopy as a tool to characterize amyloid oligomers and fibrils; pp. 211–243. [Google Scholar]
- 91.Rasmussen J, Mahler J, Beschorner N et al. Amyloid polymorphisms constitute distinct clouds of conformational variants in different etiological subtypes of Alzheimer's disease. Proc Natl Acad Sci U S A. 2017;114(49):13018–13023. doi: 10.1073/pnas.1713215114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Stepanchuk A A, Heyne B, Stys P K. Complex photophysical properties of K114 make for a versatile fluorescent probe for amyloid detection. ACS Chem Neurosci. 2021;12(07):1273–1280. doi: 10.1021/acschemneuro.1c00101. [DOI] [PubMed] [Google Scholar]
- 93.Stepanchuk A A, Barber P A, Lashley T, Joseph J T, Stys P K. Quantitative detection of grey and white matter amyloid pathology using a combination of K114 and CRANAD-3 fluorescence. Neurobiol Dis. 2021;161:105540. doi: 10.1016/j.nbd.2021.105540. [DOI] [PubMed] [Google Scholar]
- 94.Leite J P, Gales L. Fluorescence properties of the amyloid indicator dye thioflavin T in constrained environments. Dyes Pigments. 2019;160:64–70. [Google Scholar]
- 95.Sulatskaya A I, Turoverov K K, Kuznetsova I M.Spectral properties and factors determining high quantum yield of thioflavin T incorporated in amyloid fibrils Spectrosc Int J 201024(1–2):169–172. [Google Scholar]
- 96.Sulatskaya A I, Rychkov G N, Sulatsky M I et al. New evidence on a distinction between Aβ40 and Aβ42 amyloids: thioflavin t binding modes, clustering tendency, degradation resistance, and cross-seeding. Int J Mol Sci. 2022;23(10):5513. doi: 10.3390/ijms23105513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Stepanenko O V, Sulatsky M I, Mikhailova E V et al. Trypsin induced degradation of amyloid fibrils. Int J Mol Sci. 2021;22(09):4828. doi: 10.3390/ijms22094828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Pretorius E, Venter C, Laubscher G Jet al. Prevalence of symptoms, comorbidities, fibrin amyloid microclots and platelet pathology in individuals with Long COVID/ Post-Acute Sequelae of COVID-19 (PASC) Cardiovascular Diabetology 2022. Aug 6;2101148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Kell D B. Genotype-phenotype mapping: genes as computer programs. Trends Genet. 2002;18(11):555–559. doi: 10.1016/s0168-9525(02)02765-8. [DOI] [PubMed] [Google Scholar]
- 100.Hastie T, Tibshirani R, Friedman J. Berlin: Springer-Verlag; 2009. The Elements of Statistical Learning: Data Mining, Inference and Prediction, 2nd edition. [Google Scholar]
- 101.Davey H M, Davey C L. Multivariate data analysis methods for the interpretation of microbial flow cytometric data. Adv Biochem Eng Biotechnol. 2011;124:183–209. doi: 10.1007/10_2010_80. [DOI] [PubMed] [Google Scholar]
- 102.Broadhurst D, Kell D B. Statistical strategies for avoiding false discoveries in metabolomics and related experiments. Metabolomics. 2006;2(04):171–196. [Google Scholar]
- 103.Kell D B, King R D. On the optimization of classes for the assignment of unidentified reading frames in functional genomics programmes: the need for machine learning. Trends Biotechnol. 2000;18(03):93–98. doi: 10.1016/s0167-7799(99)01407-9. [DOI] [PubMed] [Google Scholar]
- 104.Day J P, Kell D B, Griffith G W. Differentiation of Phytophthora infestans sporangia from other airborne biological particles by flow cytometry. Appl Environ Microbiol. 2002;68(01):37–45. doi: 10.1128/AEM.68.1.37-45.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Kenny L C, Dunn W B, Ellis D I et al. Novel biomarkers for pre-eclampsia detected using metabolomics and machine learning. Metabolomics. 2005;1(03):227–234. [Google Scholar]
- 106.Screening for Pregnancy Endpoints Consortium . Kenny L C, Broadhurst D I, Dunn W et al. Robust early pregnancy prediction of later preeclampsia using metabolomic biomarkers. Hypertension. 2010;56(04):741–749. doi: 10.1161/HYPERTENSIONAHA.110.157297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kingma D P, Welling M.An Introduction to Variational AutoencodersArxiv2019 [Google Scholar]
- 108.Girin L, Leglaive S, Bie Xet al. Dynamical Variational Autoencoders: A Comprehensive ReviewArxiv2020 [Google Scholar]
- 109.Samanta S, O'Hagan S, Swainston N, Roberts T J, Kell D B. VAE-Sim: a novel molecular similarity measure based on a variational autoencoder. Molecules. 2020;25(15):3446. doi: 10.3390/molecules25153446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Vaswani A, Shazeer N, Parmar Net al. Attention Is All You NeedArxiv2017 [Google Scholar]
- 111.Lin T, Wang Y, Liu X, Qiu X.A Survey of TransformersArxiv2021 [Google Scholar]
- 112.Wolf T, Debut L, Sanh Vet al. Transformers: State-of-the-art Natural Language Processing.Proceedings of the 2020 EMNLP (Systems Demonstrations)201938–45.
- 113.Zhang H, Song H, Li S, Zhou M, Song D.A Survey of Controllable Text Generation using Transformer-based Pre-trained Language ModelsArxiv2022 [Google Scholar]
- 114.Liu Y, Zhang Y, Wang Yet al. A Survey of Visual TransformersArxiv2021 [DOI] [PubMed] [Google Scholar]
- 115.He K, Gan C, Li Zet al. Transformers in Medical Image Analysis: A ReviewArxiv2022 [Google Scholar]
- 116.Shamshad F, Khan S, Zamir S Wet al. Transformers in Medical Imaging: A SurveyArxiv2022 [DOI] [PubMed] [Google Scholar]
- 117.Shrivastava A D, Kell D B. FragNet, a contrastive learning-based transformer model for clustering, interpreting, visualising and navigating chemical space. Molecules. 2021;26(07):2065. doi: 10.3390/molecules26072065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Human Serum Metabolome (HUSERMET) Consortium . Dunn W B, Broadhurst D, Begley P et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat Protoc. 2011;6(07):1060–1083. doi: 10.1038/nprot.2011.335. [DOI] [PubMed] [Google Scholar]
- 119.Wright Muelas M, Roberts I, Mughal F, O'Hagan S, Day P J, Kell D B. An untargeted metabolomics strategy to measure differences in metabolite uptake and excretion by mammalian cell lines. Metabolomics. 2020;16(10):107. doi: 10.1007/s11306-020-01725-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Roberts I, Wright Muelas M, Taylor J M et al. Untargeted metabolomics of COVID-19 patient serum reveals potential prognostic markers of both severity and outcome. Metabolomics. 2021;18(01):6. doi: 10.1007/s11306-021-01859-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Anderson N L, Anderson N G. Proteome and proteomics: new technologies, new concepts, and new words. Electrophoresis. 1998;19(11):1853–1861. doi: 10.1002/elps.1150191103. [DOI] [PubMed] [Google Scholar]
- 122.Rosendahl P, Plak K, Jacobi A et al. Real-time fluorescence and deformability cytometry. Nat Methods. 2018;15(05):355–358. doi: 10.1038/nmeth.4639. [DOI] [PubMed] [Google Scholar]
- 123.Inwald D P, McDowall A, Peters M J, Callard R E, Klein N J. CD40 is constitutively expressed on platelets and provides a novel mechanism for platelet activation. Circ Res. 2003;92(09):1041–1048. doi: 10.1161/01.RES.0000070111.98158.6C. [DOI] [PubMed] [Google Scholar]
- 124.Dewitte A, Tanga A, Villeneuve J et al. New frontiers for platelet CD154. Exp Hematol Oncol. 2015;4:6. doi: 10.1186/s40164-015-0001-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Henn V, Steinbach S, Büchner K, Presek P, Kroczek R A. The inflammatory action of CD40 ligand (CD154) expressed on activated human platelets is temporally limited by coexpressed CD40. Blood. 2001;98(04):1047–1054. doi: 10.1182/blood.v98.4.1047. [DOI] [PubMed] [Google Scholar]
- 126.Chapman L M, Aggrey A A, Field D J et al. Platelets present antigen in the context of MHC class I. J Immunol. 2012;189(02):916–923. doi: 10.4049/jimmunol.1200580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Placke T, Örgel M, Schaller M et al. Platelet-derived MHC class I confers a pseudonormal phenotype to cancer cells that subverts the antitumor reactivity of natural killer immune cells. Cancer Res. 2012;72(02):440–448. doi: 10.1158/0008-5472.CAN-11-1872. [DOI] [PubMed] [Google Scholar]
- 128.Vallance T M, Zeuner M T, Williams H F, Widera D, Vaiyapuri S. Toll-like receptor 4 signalling and its impact on platelet function, thrombosis, and haemostasis. Mediators Inflamm. 2017;2017:9.605894E6. doi: 10.1155/2017/9605894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Qiao J, Al-Tamimi M, Baker R I, Andrews R K, Gardiner E E. The platelet Fc receptor, FcγRIIa. Immunol Rev. 2015;268(01):241–252. doi: 10.1111/imr.12370. [DOI] [PubMed] [Google Scholar]
- 130.Arman M, Krauel K. Human platelet IgG Fc receptor FcγRIIA in immunity and thrombosis. J Thromb Haemost. 2015;13(06):893–908. doi: 10.1111/jth.12905. [DOI] [PubMed] [Google Scholar]
- 131.Estevez B, Du X. New concepts and mechanisms of platelet activation signaling. Physiology (Bethesda) 2017;32(02):162–177. doi: 10.1152/physiol.00020.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Speth C, Rambach G, Würzner R et al. Complement and platelets: mutual interference in the immune network. Mol Immunol. 2015;67(01):108–118. doi: 10.1016/j.molimm.2015.03.244. [DOI] [PubMed] [Google Scholar]
- 133.Cha J K, Jeong M H, Jang J Y et al. Serial measurement of surface expressions of CD63, P-selectin and CD40 ligand on platelets in atherosclerotic ischemic stroke. A possible role of CD40 ligand on platelets in atherosclerotic ischemic stroke. Cerebrovasc Dis. 2003;16(04):376–382. doi: 10.1159/000072560. [DOI] [PubMed] [Google Scholar]
- 134.Israels S J, McMillan-Ward E M, Easton J, Robertson C, McNicol A. CD63 associates with the alphaIIb beta3 integrin-CD9 complex on the surface of activated platelets. Thromb Haemost. 2001;85(01):134–141. [PubMed] [Google Scholar]
- 135.Wichaiyo S, Lax S, Montague S J et al. Platelet glycoprotein VI and C-type lectin-like receptor 2 deficiency accelerates wound healing by impairing vascular integrity in mice. Haematologica. 2019;104(08):1648–1660. doi: 10.3324/haematol.2018.208363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Nieswandt B, Pleines I, Bender M. Platelet adhesion and activation mechanisms in arterial thrombosis and ischaemic stroke. J Thromb Haemost. 2011;9 01:92–104. doi: 10.1111/j.1538-7836.2011.04361.x. [DOI] [PubMed] [Google Scholar]
- 137.Nieswandt B, Watson S P. Platelet-collagen interaction: is GPVI the central receptor? Blood. 2003;102(02):449–461. doi: 10.1182/blood-2002-12-3882. [DOI] [PubMed] [Google Scholar]
- 138.Moroi M, Jung S M. Platelet glycoprotein VI: its structure and function. Thromb Res. 2004;114(04):221–233. doi: 10.1016/j.thromres.2004.06.046. [DOI] [PubMed] [Google Scholar]
- 139.Schönberger T, Ziegler M, Borst O et al. The dimeric platelet collagen receptor GPVI-Fc reduces platelet adhesion to activated endothelium and preserves myocardial function after transient ischemia in mice. Am J Physiol Cell Physiol. 2012;303(07):C757–C766. doi: 10.1152/ajpcell.00060.2012. [DOI] [PubMed] [Google Scholar]
- 140.Suzuki-Inoue K, Fuller G L, García A et al. A novel Syk-dependent mechanism of platelet activation by the C-type lectin receptor CLEC-2. Blood. 2006;107(02):542–549. doi: 10.1182/blood-2005-05-1994. [DOI] [PubMed] [Google Scholar]
- 141.Fuller G L, Williams J A, Tomlinson M G et al. The C-type lectin receptors CLEC-2 and Dectin-1, but not DC-SIGN, signal via a novel YXXL-dependent signaling cascade. J Biol Chem. 2007;282(17):12397–12409. doi: 10.1074/jbc.M609558200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Merten M, Thiagarajan P. P-selectin expression on platelets determines size and stability of platelet aggregates. Circulation. 2000;102(16):1931–1936. doi: 10.1161/01.cir.102.16.1931. [DOI] [PubMed] [Google Scholar]
- 143.Andrews R K, Gardiner E E, Shen Y, Whisstock J C, Berndt M C. Glycoprotein Ib-IX-V. Int J Biochem Cell Biol. 2003;35(08):1170–1174. doi: 10.1016/s1357-2725(02)00280-7. [DOI] [PubMed] [Google Scholar]
- 144.Gardiner E E, Karunakaran D, Shen Y, Arthur J F, Andrews R K, Berndt M C. Controlled shedding of platelet glycoprotein (GP)VI and GPIb-IX-V by ADAM family metalloproteinases. J Thromb Haemost. 2007;5(07):1530–1537. doi: 10.1111/j.1538-7836.2007.02590.x. [DOI] [PubMed] [Google Scholar]
- 145.Pennings G J, Kritharides L. CD147 in cardiovascular disease and thrombosis. Semin Thromb Hemost. 2014;40(07):747–755. doi: 10.1055/s-0034-1390001. [DOI] [PubMed] [Google Scholar]
- 146.Muramatsu T. Basigin (CD147), a multifunctional transmembrane glycoprotein with various binding partners. J Biochem. 2016;159(05):481–490. doi: 10.1093/jb/mvv127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Schmidt R, Bültmann A, Fischel S et al. Extracellular matrix metalloproteinase inducer (CD147) is a novel receptor on platelets, activates platelets, and augments nuclear factor kappaB-dependent inflammation in monocytes. Circ Res. 2008;102(03):302–309. doi: 10.1161/CIRCRESAHA.107.157990. [DOI] [PubMed] [Google Scholar]
- 148.Pennings G J, Yong A S, Kritharides L. Expression of EMMPRIN (CD147) on circulating platelets in vivo. J Thromb Haemost. 2010;8(03):472–481. doi: 10.1111/j.1538-7836.2009.03716.x. [DOI] [PubMed] [Google Scholar]
- 149.Canobbio I, Balduini C, Torti M. Signalling through the platelet glycoprotein Ib-V-IX complex. Cell Signal. 2004;16(12):1329–1344. doi: 10.1016/j.cellsig.2004.05.008. [DOI] [PubMed] [Google Scholar]
- 150.Yong A, Pennings G, Wong C et al. Intracoronary upregulation of platelet extracellular matrix metalloproteinase inducer (CD147) in coronary disease. Int J Cardiol. 2013;166(03):716–721. doi: 10.1016/j.ijcard.2011.11.093. [DOI] [PubMed] [Google Scholar]
- 151.Berger G, Hartwell D W, Wagner D D. P-Selectin and platelet clearance. Blood. 1998;92(11):4446–4452. [PubMed] [Google Scholar]
- 152.Blann A D. Soluble P-selectin: the next step. Thromb Res. 2014;133(01):3–4. doi: 10.1016/j.thromres.2013.10.030. [DOI] [PubMed] [Google Scholar]
- 153.Au A E, Josefsson E C. Regulation of platelet membrane protein shedding in health and disease. Platelets. 2017;28(04):342–353. doi: 10.1080/09537104.2016.1203401. [DOI] [PubMed] [Google Scholar]
- 154.Panzer S, Rosales S, Gisslinger H et al. Plasma levels of P-selectin are determined by platelet turn-over and the P-selectin Thr715Pro polymorphism. Thromb Res. 2008;121(04):573–579. doi: 10.1016/j.thromres.2007.05.025. [DOI] [PubMed] [Google Scholar]
- 155.Kim O V, Nevzorova T A, Mordakhanova E R et al. Fatal dysfunction and disintegration of thrombin-stimulated platelets. Haematologica. 2019;104(09):1866–1878. doi: 10.3324/haematol.2018.202309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Huynh K C, Stoldt V R, Scharf R E. Contribution of distinct platelet integrins to binding, unfolding, and assembly of fibronectin. Biol Chem. 2013;394(11):1485–1493. doi: 10.1515/hsz-2013-0182. [DOI] [PubMed] [Google Scholar]
- 157.Xu X R, Carrim N, Neves M A et al. Platelets and platelet adhesion molecules: novel mechanisms of thrombosis and anti-thrombotic therapies. Thromb J. 2016;14 01:29. doi: 10.1186/s12959-016-0100-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Gao W, Shi P, Chen X et al. Clathrin-mediated integrin αIIbβ3 trafficking controls platelet spreading. Platelets. 2018;29(06):610–621. doi: 10.1080/09537104.2017.1353682. [DOI] [PubMed] [Google Scholar]
- 159.Durrant T N, van den Bosch M T, Hers I. Integrin α IIb β 3 outside-in signaling . Blood. 2017;130(14):1607–1619. doi: 10.1182/blood-2017-03-773614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Coller B S.αIIbβ3: structure and function J Thromb Haemost 201513(Suppl 1, Suppl 1)S17–S25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Marjoram R J, Li Z, He L et al. α2β1 integrin, GPVI receptor, and common FcRγ chain on mouse platelets mediate distinct responses to collagen in models of thrombosis. PLoS One. 2014;9(11):e114035. doi: 10.1371/journal.pone.0114035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Sebastiano M, Momi S, Falcinelli E, Bury L, Hoylaerts M F, Gresele P. A novel mechanism regulating human platelet activation by MMP-2-mediated PAR1 biased signaling. Blood. 2017;129(07):883–895. doi: 10.1182/blood-2016-06-724245. [DOI] [PubMed] [Google Scholar]
- 163.Duvernay M T, Temple K J, Maeng J G et al. Contributions of protease-activated receptors PAR1 and PAR4 to thrombin-induced GPIIbIIIa activation in human platelets. Mol Pharmacol. 2017;91(01):39–47. doi: 10.1124/mol.116.106666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Kahn M L, Nakanishi-Matsui M, Shapiro M J, Ishihara H, Coughlin S R. Protease-activated receptors 1 and 4 mediate activation of human platelets by thrombin. J Clin Invest. 1999;103(06):879–887. doi: 10.1172/JCI6042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Shakhidzhanov S S, Shaturny V I, Panteleev M A, Sveshnikova A N. Modulation and pre-amplification of PAR1 signaling by ADP acting via the P2Y12 receptor during platelet subpopulation formation. Biochim Biophys Acta. 2015;1850(12):2518–2529. doi: 10.1016/j.bbagen.2015.09.013. [DOI] [PubMed] [Google Scholar]
- 166.Amann M, Ferenc M, Valina C M et al. Validation of a P2Y 12 -receptor specific whole blood platelet aggregation assay . Platelets. 2016;27(07):668–672. doi: 10.3109/09537104.2016.1153620. [DOI] [PubMed] [Google Scholar]
- 167.Cunningham M R, Nisar S P, Mundell S J. Molecular mechanisms of platelet P2Y(12) receptor regulation. Biochem Soc Trans. 2013;41(01):225–230. doi: 10.1042/BST20120295. [DOI] [PubMed] [Google Scholar]
- 168.Hensch N R, Karim Z A, Pineda J, Mercado N, Alshbool F Z, Khasawneh F T. P2Y 12 antibody inhibits platelet activity and protects against thrombogenesis . Biochem Biophys Res Commun. 2017;493(02):1069–1074. doi: 10.1016/j.bbrc.2017.09.083. [DOI] [PubMed] [Google Scholar]
- 169.Oestreich J H, Ferraris S P, Steinhubl S R, Akers W S. Pharmacodynamic interplay of the P2Y(1), P2Y(12), and TxA(2) pathways in platelets: the potential of triple antiplatelet therapy with P2Y(1) receptor antagonism. Thromb Res. 2013;131(02):e64–e70. doi: 10.1016/j.thromres.2012.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Chakraborty R, Bhullar R P, Dakshinamurti S, Hwa J, Chelikani P. Inverse agonism of SQ 29,548 and ramatroban on thromboxane A2 receptor. PLoS One. 2014;9(01):e85937. doi: 10.1371/journal.pone.0085937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Amano H, Ito Y, Eshima K et al. Thromboxane A2 induces blood flow recovery via platelet adhesion to ischaemic regions. Cardiovasc Res. 2015;107(04):509–521. doi: 10.1093/cvr/cvv139. [DOI] [PubMed] [Google Scholar]
- 172.Nyström S, Hammarström P. Amyloidogenesis of SARS-CoV-2 spike protein. J Am Chem Soc. 2022;144(20):8945–8950. doi: 10.1021/jacs.2c03925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Bester J, Pretorius E. Effects of IL-1β, IL-6 and IL-8 on erythrocytes, platelets and clot viscoelasticity. Sci Rep. 2016;6:32188. doi: 10.1038/srep32188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Pretorius E, du Plooy J, Soma P, Gasparyan A Y. An ultrastructural analysis of platelets, erythrocytes, white blood cells, and fibrin network in systemic lupus erythematosus. Rheumatol Int. 2014;34(07):1005–1009. doi: 10.1007/s00296-013-2817-x. [DOI] [PubMed] [Google Scholar]
- 175.Olumuyiwa-Akeredolu O O, Soma P, Buys A V, Debusho L K, Pretorius E. Characterizing pathology in erythrocytes using morphological and biophysical membrane properties: relation to impaired hemorheology and cardiovascular function in rheumatoid arthritis. Biochim Biophys Acta Biomembr. 2017;1859(12):2381–2391. doi: 10.1016/j.bbamem.2017.09.014. [DOI] [PubMed] [Google Scholar]
- 176.Bester J, Soma P, Kell D B, Pretorius E. Viscoelastic and ultrastructural characteristics of whole blood and plasma in Alzheimer-type dementia, and the possible role of bacterial lipopolysaccharides (LPS) Oncotarget. 2015;6(34):35284–35303. doi: 10.18632/oncotarget.6074. [DOI] [PMC free article] [PubMed] [Google Scholar]