

Abstract
Non-perennial river segments — those that recurrently cease to flow or frequently dry — occur in all river networks and are globally more abundant than perennial (always flowing) segments. However, research and management have historically focused on perennial river segments. In this Review, we outline how non-perennial segments are integral parts of river networks. Repeated cycles of flowing, non-flowing and dry phases in non-perennial segments influence biodiversity and ecosystem dynamics at different spatial scales, from individual segments to entire river networks. Varying configurations of perennial and non-perennial segments govern physical, chemical and ecological responses to changes in the flow regimes of each river network, especially in response to human activities. The extent of non-perennial segments in river networks has increased owing to warming, changing hydrological patterns and human activities, and this increase is predicted to continue. Moreover, the dry phases of flow regimes are expected to be longer, drier and more frequent, albeit with high regional variability. These changes will likely impact biodiversity, potentially tipping some ecosystems to compromised stable states. Effective river-network management must recognize ecosystem services (such as flood risk management and groundwater recharge) provided by non-perennial segments and ensure their legislative and regulatory protection, which is often lacking.
Introduction
Rivers cover less than 2% of the surface of the Earth but contain approximately 13% of all described species and provide key ecosystem services such as provision of drinking water and food, regulation of climate and opportunities for recreation1,2. Despite their importance, rivers are among the most threatened ecosystem types in the world; one out of three riverine species in these biodiversity hotspots is threatened with extinction3. River science and management have historically focused on perennial segments, which flow year-round, but rivers are increasingly being conceptualized as spatially variable networks in which hydrological connections between perennial and non-perennial segments enable exchanges of water, materials and organisms that support network-scale biodiversity and ecosystem functioning4,5 (Fig. 1a).
Figure 1.

Non-perennial river segments: definition, abundance and flow regimes. a, An idealized river network, indicating the different types of non-perennial (ephemeral and intermittent) river segment and their linkages with nearby waters. b, A typical hydrological sequence of a non-perennial segment: flowing, non-flowing, dry and rewetting phases in the Calavon River, France. c, Global prevalence of non-perennial river networks. Panel b images courtesy of Bertrand Launay. Panel c reprinted with permission from ref. 4, Springer Nature Limited.
Non-perennial river segments (NPRs)4 recurrently experience flow cessation and lose most or all surface water (Fig. 1b). There is no global consensus in terminology6–9 owing to the high temporal and spatial variability of flows within and among NPRs10, but here they are classified generally as ‘ephemeral’, which only flow in response to rainfall, or ‘intermittent’, whose flows are longer and more predictable. When flowing (Fig. 1c), NPRs supply water, biota, energy, nutrients and other materials to connected waters, influencing their water quality, biodiversity and ecological integrity. These pulsed hydrological connections are often irregular in time and space yet generate dynamic transition zones between aquatic and terrestrial habitats that extend longitudinally down river channels, laterally onto floodplains and vertically into the underlying groundwaters11.
Every river network encompasses NPRs, particularly in the headwaters (Fig. 1a), but sometimes also in substantial lengths of the lower segments and often in braided sections and alluvial floodplains. NPRs naturally constitute more than half of the global river network length4 (Fig. 1c), a proportion that is predicted to rise in some regions because of climate change, land-use alteration and increased water abstraction12,13. Despite the ubiquity of NPRs and the ecological importance of the hydrological connectivity they provide, almost all policies and management practices for river networks are tailored for rivers that are perennial. The omission of NPRs from river management seriously risks undermining effective protection of the biodiversity and ecological integrity of entire river networks and their ecosystem services. For example, rubbish dumped into unmanaged headwater NPRs when surface water is absent will impact receiving perennial waters when flow resumes and carries contaminants downstream14. Scientists and managers need greater awareness of the importance of NPRs to biodiversity and ecosystem processes to protect entire river networks.
In this Review, we explore key insights from research on NPRs in the context of network-scale riverine connectivity. For brevity, we focus on aquatic responses to drying from segment to river network scale but acknowledge that interactions with terrestrial ecosystems are also crucial in NPRs, especially during dry phases. We conclude by exploring the drawbacks of excluding NPRs from river network management. River management strategies must treat river networks as arrays of co-occurring perennial segments and NPRs and must integrate NPRs into actions that maintain and, where possible, enhance network-scale biodiversity, ecosystem processes and ecosystem services.
Characterizing non-perennial segments
This section discusses NPR flow regimes, the types of connections between perennial segments and NPRs in river networks, their distribution within different networks and how such connections can alter fluxes of water, materials and organisms across river networks.
Typology and connections
Flow regimes.
The flow regime governs river geomorphology, water quality and ecology15 and, thus, is crucial to the understanding and management of river networks. In NPRs, the flow regime encompasses alternating flowing phases, non-flowing phases and, in many cases, dry phases (Fig. 1c). The frequency, duration and timing of each phase are powerful determinants of biodiversity, ecological integrity and ecosystem services, both at local scales within NPRs16,17 and at the river-network scale18,19.
Drying is a gradual process whose effects are separated by hydrological thresholds20. Early in the drying phase, while flow declines, surface water contracts and lateral aquatic habitats become disconnected. As drying progresses, riffles and other flowing surface habitats disappear, leaving the riverbed as disconnected pools (Fig. 1c). Eventually, the riverbed dries, although subsurface flow can continue in saturated sediments beneath the dry channel (the hyporheic zone)20–22 (Fig. 1c). This loss of surface water is governed by the type of surface– groundwater interactions occurring at the segment scale23–25. In losing segments, the water table of the underlying aquifer is generally deep and the hyporheic zone usually dries quickly21,25, whereas in gaining segments, upwelling groundwater can maintain a saturated hyporheic zone throughout the dry phase21,25.
During the rewetting phase, surface water returns to inundate dry channels, sometimes quickly as flash floods26,27 or slowly as a rise in the water table. Similar to all components of flow regimes, the characteristics of drying and rewetting transitional phases are driven by both natural climatic and geological factors and by human activities28–30 and vary greatly within and between river networks31,32. The consequences of such variability remain poorly understood but are likely to be an important determinant of the biodiversity and ecological integrity of a river network19,33.
Perennial and non-perennial segment connections.
Most NPRs are structurally connected to perennial segments by the spatial continuity of the river corridor (the channel, hyporheic zone and floodplain34) (Fig. 1a), reflecting their ubiquitous co-occurrence in river networks globally (Fig. 1b). This connectivity is well captured by the meta-ecosystem perspective that considers river networks as mutually dependent arrays of perennial river segments and NPRs that are connected to nearby aquatic and terrestrial ecosystems35. The connected terrestrial ecosystems range from riparian zones and uplands in the headwaters to floodplains downstream, and the connected aquatic ecosystems encompass nearby wetlands, lakes, subsurface groundwaters, reservoirs, estuaries and coastal waters (Fig. 1a).
The spatial arrangements of NPRs and perennial river segments and their connections are diverse, complex and dynamic (Fig. 2a,b). Most river networks have naturally non-perennial headwaters that span the interface between terrestrial and aquatic domains36. Flow in headwater segments is driven by a combination of surface runoff, groundwater inputs and/or meltwater and is often seasonal18,37. NPRs can also occur in the downstream parts of river networks (Fig. 2b). Typically, river networks that flow out onto porous alluvial plains and recharge the underlying groundwater have downstream NPRs, such as the Tagliamento River (Italy26) and the Albarine River (France38). Other causes of downstream NPRs include excessive evaporation such as in the Diamantina River (Australia39) and human activities such as damming and water abstraction cause artificial drying, for example, in the Colorado River (USA) and Yellow River (China40). In other river networks, the upper and lower segments are perennial but the mid-segments are non-perennial, such as in the Selwyn River (New Zealand41). In arid regions, whole river networks are often non-perennial, such as many rivers in northern and southwestern Africa42 and the Nordeste region in Brazil43. Most braided sections of rivers comprise NPRs, which can also be prevalent across large, alluvial floodplains26,44. The varying physical settings (such as channel shape and size, streambed permeability, groundwater influence and large wood deposits), flow regimes and catchment land-uses of these different network configurations drive physical, chemical and ecological responses to the connections between NPRs and perennial segments.
Figure 2.

The connections between non-perennial and perennial river segments. a, Examples of connections between perennial and non-perennial segments. b, River-network patterns of co-occurring non-perennial (dashed lines) and perennial (solid lines) segments. c, Functions affecting fluxes of water, materials and organisms through non-perennial segments (blue triangles) before entering downstream waters (grey triangles), as in ref. 45. Changes in arrow thickness reflect changes in the fluxes through the functional connections. Changes in arrow colour reflect conversion of material or energy form. Changes in arrow shape reflect delayed delivery of material or energy.
Functional connections among river segments.
The different functional roles played by the varying connections between perennial segments and NPRs in space and time can be classified by the general mechanisms by which stream segments influence fluxes to downstream waters45 (Fig. 2c). Fluxes of water, sediments, material and organisms from perennial to NPRs and vice versa are often altered, which can have contrasting effects on downstream river segments (Fig. 2c). These contrasting effects occur because the fluxes can cease, increase, decrease or be stored when they pass through NPRs before entering downstream waters45.
Connections between perennial segments and NPRs can act as sources of material, notably when the organic material that accumulates during dry phases is leached during rewetting, causing high nutrient fluxes to downstream waters46 (Fig. 2c). When NPRs dry, exchanges of water cease, both vertically between surface and groundwaters and longitudinally from upstream to downstream waters, transforming NPRs into sinks of material47 (Fig. 2c). As dry phases progress, terrestrial leaf litter from riparian zones is retained in NPRs and gradually accumulates on the streambed33 (Fig. 2c). Upon rewetting, this organic material is flushed downstream, sometimes en masse27, with NPRs functioning to delay its release and processing (Fig. 2c). Connections between NPRs and perennial river segments can also transform spatial patterns inchemical parameters such as dissolved oxygen when anoxic or hypoxic pulses of water from remnant pools or rewetting fronts are carried downstream to connected aquatic ecosystems by rewetting flows19,48 (Fig. 2c). The types and direction of processes that are altered are likely to reflect flow regime characteristics such as dry-phase duration and the longitudinal sequence of perennial segments and NPRs.
Global distribution and temporal trends
NPRs are prevalent on all continents, representing more than half of the global river network4 (Fig. 1b). For example, 94% of river lengths in the southwestern USA49 and more than 70% of river lengths in Australia are non-perennial50. NPRs typically dominate in arid, semi-arid and dry subhumid regions, which represent up to half of the land surfaces of the Earth40. These segments are also common across alpine, boreal, continental, Mediterranean, oceanic, polar and tropical regions16,51. Every river network on Earth includes NPRs, especially in their headwaters. Headwaters are estimated to represent more than 70% of the total river-network length and are typically prone to drying4,36.
The ecology of non-perennial segments
Drying in NPRs controls local biotic communities, ecosystem processes and ecosystem services. In addition, the spatial arrangement and type of connections between perennial segments and NPRs at the river-network scale shape the ecological integrity of river networks. This section discusses how drying influences the ecology of NPRs at the segment scale and how these effects propagate across the entire river network.
Non-perennial segment scale
Biotic groups respond locally to recurrent shifts between flowing, non-flowing and dry conditions in NPRs (Fig. 1c). Aquatic biodiversity declines steadily in response to non-perenniality as taxa lacking adaptations promoting resistance or resilience to drying are lost16,17. The extent of these declines is governed by hydrological parameters such as the duration of non-flowing and dry phases, with longer dry phases resulting in greater declines38,52,53. For example, a 10-day increase in the dry phase in the Albarine River, France, led to an additional loss of six invertebrate taxa from the benthic community and four invertebrate taxa from the hyporheic zone38. An increase in the duration of the non-flowing phase from 0 (perennial flow) to 78 days reduced the survival rate of Coho salmon (Oncorhynchus kisutch) populations from 59% to 11% in tributaries of the Russian River in California, USA52.
Concurrent increases in terrestrial species richness occur as colonizing species arrive from riparian and wider terrestrial habitats54. For example, 22 and 12 invertebrate taxa colonized the dry riverbeds of the Albarine River (France) and Oaky Creek (Australia), respectively, within 2 months of the onset of the dry phase54. Although aquatic species richness during the flowing phase at the site scale can be considerably lower in NPRs than in perennial segments55, NPRs contribution to regional biodiversity can exceed those of perennial segments because of the inherently high β-diversity (variability in community composition) in space and time and the presence of specialized species adapted to non-perennial conditions51,56.
When flow resumes, aquatic organisms recolonize from various refuges, including upstream pools, moist sediments and leaf litter57–59. Some aquatic organisms including invertebrates and diatoms have desiccation-tolerant forms that can survive in moist riverbed sediments60,61. Recovery rates and trajectories vary depending on connectivity to refuges and on dispersal abilities of species. Communities in NPRs connected to upstream perennial waters are soon replenished by colonists62, whereas community recovery in isolated NPRs can be slow and more stochastic58,63. Recovery can be modified by anthropogenic influences such as fragmentation by instream barriers that sever links between NPRs and sources of colonists in perennial segments64. Despite well-developed recovery mechanisms, differences persist between communities in perennial and non-perennial segments38,55. These differences can be particularly pronounced and long-lasting when unprecedented dry phases occur during drought events. By contrast, communities can recover rapidly, generally within a few weeks, after ‘normal’ seasonal dry phases63,65.
Aquatic species control fundamental ecosystem processes such as primary production and organic matter decomposition, and changes in aquatic biodiversity related to non-perenniality therefore alter ecosystem functioning. For example, desiccation-tolerant microorganisms in natural NPRs mediate biogeochemical cycling. When flow decreases, hypoxic conditions that develop in disconnected pools favour denitrification, reducing nitrate concentrations25. During dry phases, the microorganisms that make up biofilms coating the surfaces of sediment particles emit large quantities of CO2 (ref. 66) and upon rewetting, large CO2 pulses can occur from NPRs. Accounting for the global prevalence of NPRs, a single rewetting event contributes up to 10% of the daily carbon dioxide emissions from all perennial rivers and streams, particularly in temperate climates33. When water returns, ecosystem processes such as respiration67, nitrification and denitrification68 and decomposition of leaf litter69 quickly resume to previous levels. Recognizing the active contribution of NPRs to carbon cycling — during both wet and dry phases — could improve the accuracy of local-to-global-scale assessments70.
Owing to drying-driven decreases in the functional diversity of their aquatic communities, NPRs can collectively perform fewer ecosystem functions than perennial ones during flowing phases71. These declines can be mitigated by functional redundancy72 (multiple species sharing traits), making the drying-induced loss of individual species functionally inconsequential73. However, as losses accumulate, the risk of losing functionally unique species increases, potentially representing a tipping point that drives the ecosystem to an alternative state. For example, by eliminating pivotal species, especially predators, drying can alter the structure and functioning of food webs, potentially leading to partial food-web collapse74. Similarly, the loss of desiccation-sensitive microorganisms and invertebrate detritivores reduces decomposition rates of particulate organic matter that fuels food webs75, altering the quantity, quality and timing of energy sources transported downstream to perennial segments after flow resumes46,76.
River-network scale
River-network-scale responses to drying are unlikely to be simple additive effects of segment-scale responses but arise from complex, interacting effects of segment-scale drying with other drivers. For example, spatial and temporal patterns of drying vary among river networks32,77,78 (Fig. 2b). Longitudinal trends related to elevation and channel form are superimposed on idiosyncratic drying patterns, thwarting efforts to extrapolate segment-scale patterns or assume that ‘river continuum’ predictions are realistic18,19,41.
Complex responses to drying also result from the propagation of biogeochemical and ecological influences downstream by surface and subsurface flows. These transfers can slow, cease or be amplified by functional connections across hydrological phases, which is likely to lead to ‘hotspots’ and ‘hot moments’ of material processing along a river network25. For example, terrestrial plant litter accumulates in dry and non-flowing segments and this litter decomposes very slowly33 (Fig. 3). When flow resumes in these segments, it can trigger rapid microbial decomposition of the labile plant litter33 and flush the decomposing litter downstream27 to perennial segments, providing a delayed subsidy of a resource that may be limiting to downstream consumers79 (Fig. 3). However, the network-scale effects of non-perenniality largely depend on the spatial arrangement of NPRs within the river network (Fig. 3). Where NPRs are concentrated in the headwaters, particularly in deciduous forested areas36, the downstream effects of non-perenniality will include high seasonal inputs of unprocessed litter (Fig. 3) that can reduce downstream water quality80 or cause technical problems for dam intakes81. By contrast, where NPRs are in downstream segments of river networks, which are typically less dependent on terrestrial litter inputs from riparian zones, lower fluxes of unprocessed litter are expected in downstream river-network segments. Meanwhile, periods of disconnectivity retain leaf litter in the headwaters, which can thus become hotspots of carbon cycling (Fig. 3).
Figure 3.

Effects of non-perenniality on river-scale leaf litter decomposition and transport. Two theoretical river networks with contrasting spatial arrangement of non-perennial segments, upstream (panel a) and downstream (panel b). Leaf litter is poorly decomposed during dry phases in non-perennial segments. Instead, it accumulates and is then transported downstream en masse when flow resumes. As such, fluxes of decomposed and undecomposed litter vary substantially between the two river networks.
Synchrony can describe how local responses propagate at the river-network scale and can be enhanced by connectivity (upstream dispersal, advective transport) between segments or by a driver such as drying that simultaneously affects multiple segments82. By reducing longitudinal connectivity of flowing water, the effects of non-perenniality of river segments can reverberate throughout a river network and desynchronize, for example, diel dissolved oxygen fluctuations19 or the recovery of biological communities during flowing phases18. When flow resumes in NPRs, hydrological connectivity is restored, promoting network-scale synchronization of such fluctuations and processes19.
The dynamics of aggregate stream systems are typically less variable than their individual contributing segments83,84 because combining asynchronous contributions from many segments has a stabilizing effect (the ‘portfolio concept’83). However, widespread non-perenniality in river networks can synchronize dynamics across populations, increasing the risk of regional species extinctions85,86. Therefore, when aggregated in river networks, the variation in flow regimes between perennial segments and NPRs contributes to the persistence of regional biodiversity and, thus, stable ecosystem functioning and associated availability of ecosystem services84.
At the river-network scale, NPRs increase the β-diversity of aquatic communities because of the simultaneous coexistence of different successional stages at the river-network scale18,40,87 (Fig. 4). Depending on the spatial arrangement of NPRs and perennial segments within the river network, communities in NPRs can comprise a subset of the taxa inhabiting perennial segments, notably when NPRs are downstream of perennial segments acting as a source of colonizing organisms16 (Fig. 4). Alternatively, when non-perenniality is concentrated upstream, their biological communities can be more variable in space and time compared with their downstream perennial counterparts87,88 (Fig. 4). This variability is because recolonization from downstream perennial segments is limited by the unidirectional flow of water along river networks and by topographic barriers, particularly for weak aquatic dispersers, increasing the role of alternative sources of colonists in the landscape, such as perennial waterbodies and the underlying hyporheic zone.
Figure 4.

Non-perenniality impacts on biodiversity patterns at the rivernetwork scale. Predicted diversity patterns in two hypothetical river networks in which non-perennial sections are located upstream (panel a) and downstream (panel b). Circles represent communities hosting different species (shapes). For the two river networks, the magnitude of the effect of drying on biodiversity is the same (the same number of species disappears along a longitudinal gradient of drying). When non-perenniality is concentrated in the headwaters, the species-poor communities are composed of species not found in speciesrich communities in downstream perennial segments. Conversely, when non-perenniality occurs downstream, the species-poor communities of these segments are subsets of species-rich communities from upstream perennial segments. These network patterns emerge because recolonization is faster from upstream to downstream, following the directionality of river flow. Adapted with permission from ref. 10, Elsevier.
Including NPRs in the study of biodiversity dynamics within river networks demonstrates the importance of dispersal, a regional process that can dominate over the local process of species-sorting during rewetting phases18,89,90. The spatial extent of drying influences access to and from refuges in the network during dry phases and thus controls the dispersal of organisms and overall metacommunity dynamics18,91,92. However, the relative contribution of species dispersal in shaping riverine biodiversity patterns is highly context-dependent, as well as being taxon-specific and extends to nearby habitats such as riparian zones56, hyporheic zones and groundwaters93, and downstream ecosystems94.
The future of river networks
This section discusses the future changes predicted for NPR flow regimes, the likely responses by aquatic biota and how management strategies for entire river networks could be tailored to incorporate these changes. Currently, perennial river segments are potentially vulnerable to the lack of protection or restoration of NPRs elsewhere in the river network.
Widespread and increasing non-perennial flow
The spatial extent of NPRs in global river networks has increased12,95,96 owing to shifts in flow regimes (Fig. 5a,b). This increase is predicted to continue13,43,97, driven in part by climatic trends such as rising temperatures and associated increases in evaporation, changing precipitation patterns and the increasing occurrence of drought in many parts of the world98–100. For example, previously perennial rivers in Europe and China dried for the first time during the severe droughts that began in 2022 (refs. 101,102). In addition, intensifying use of water resources, including surface and groundwater abstraction, storage and diversion, are driving shifts from perennial to artificially non-perennial flow40,101. In KS, USA, for example, widespread irrigation from the High Plains aquifer since the mid-1900s has lowered the regional water table, sometimes by more than 50 m, shifting rivers and streams from naturally gaining and perennially flowing to artificially losing and non-perennial flow103,104.
Figure 5.

The future hydrological and biological fate of non-perennial rivers. a, Increasing or decreasing trends at the European scale (crosses indicate no trend) at the 10% significance level for the annual mean number of zero-flow days96. b, Mann–Kendall trends in annual no-flow days across the USA; red and blue indicate longer and shorter no-flow durations, respectively12. Unfilled circles indicate there was no significant trend. c, Relationships between annual no-flow duration (as a percentage) and the taxonomic richness (as the number of taxa) of aquatic communities. Labels mark (1) benthic invertebrates in the Albarine River, France38; (2) benthic invertebrates in the Selwyn River, New Zealand41; (3) hyporheic invertebrates in the Selwyn River187; (4) riparian plants in the San Pedro River, Arizona188; and (5) fish in the Selwyn River189. The lines are based on regression models published in the original studies. Panel a reprinted with permission from ref. 96, Taylor & Francis Group. Panel b is reprinted with permission from ref. 12, CC BY 4.0. Panel c is adapted with permission from ref. 16, Oxford University Press.
The predicted increase in the spatial extent of NPRs represents a network-scale decline in aquatic habitat availability and hydrological connectivity, which will alter riverine biodiversity, with consequences for ecosystem functioning and services. For example, a decrease in the availability and quality of wet refuges that support aquatic organisms during dry phases will synchronize biological responses to drying, reducing metacommunity resilience and local community recovery after flow resumes86.
Longer, drier and more frequent dry phases
Flow regime characteristics such as the frequency, duration, severity and timing of flowing, non-flowing and dry phases are changing, as is the rate of change during transitional drying and rewetting phases. Despite considerable regional variability in the evidence for such changes12,96 (Fig. 5a,b), there is high confidence that changes will intensify in the near future. These predicted changes include increases in dry-phase durations, frequency, severity99 and rates of onset of both dry phases and flow resumption105. Inherent within the predicted increase in dry-phase duration is the earlier onset and/or later cessation of dry phases in NPRs with seasonal flow regimes, as well as an increased co-occurrence of dry phases and extreme climatic events, particularly heatwaves106–108.
In the context of megadroughts109,110, dry phases could continue uninterrupted for years in NPRs that currently have seasonal flow regimes. Deglaciation and snow loss are expected to reduce summer flows, resulting in shifts to non-perennial flow111,112. Conversely, warmer winters with greater snowmelt and glacial melting could cause NPRs to become perennial at higher elevations and northern latitudes43.
Biological responses to future changes
Biological communities in NPRs are expected to respond to future changes in flow regime characteristics. An increase in dry-phase duration and severity (caused by a lack of precipitation, water abstraction and/or high temperatures and manifesting as reduced in-channel water availability) is likely to reduce the survival of desiccation-tolerant life stages of aquatic organisms within the riverbed sediments60. Similarly, an increase in dry-phase duration and frequency could eliminate desiccation-sensitive species without enabling colonization by tolerant equivalents72 (Fig. 5c). Faster wet-to-dry transitions could shorten the time between environmental cues that trigger insect metamorphosis and its completion, reducing the emergence of adults113. Rapid-onset rewetting phases that wash insects, crustaceans, amphibians or fish straight from refuges to downstream segments could reduce local community recovery rates114–116. Both earlier dry-phase onset and later dry-phase termination could reduce successful completion of aquatic stages of life cycles by riverine animals. For example, earlier onset of drying could reduce the proportion of aquatic juvenile insects that emerge as terrestrial adults in time to avoid desiccation113. Prolonging the dry phase could prevent egg-laying behaviours by species that oviposit on water117. Distances and connectivity to perennial refuges in the landscape may determine post-drying community composition86,118. In cases of shifts from perennial to non-perennial flow regimes, biological responses might be particularly dramatic, because species lack adaptations to drying. However, if NPRs are abundant in a river network, they could provide colonists adapted to the newly non-perennial conditions86,101.
In terms of ecosystem processes, biofilms generally recover within a few days upon flow resumption, from dormant forms and through drift119. Some ecosystem processes, such as primary production and ecosystem respiration, are therefore highly resilient to drying. As such, natural NPRs experiencing longer or more frequent dry phases might not be severely affected in the near future, although shifts towards greater reliance on external energy sources could occur120,121. Other processes could be more markedly affected by increasing drying, such as the decomposition of terrestrial leaf litter, because they are more dependent on macroorganisms101. Where once-perennial segments become non-perennial, biodiversity is predicted to respond strongly, with multiple cascading effects on ecosystem processes, although these effects will depend on the functional redundancy of a community and the types of organisms involved101,120,122. Finally, increasing non-perenniality is likely to occur in streams affected by multiple anthropogenic stressors, such as pollution and water abstraction, that interact to cause complex changes to ecosystem processes. For example, mesocosm experiments suggest that flow reductions and fine sediment pollution have synergistic effects on both algal biomass and thus primary production and on invertebrate abundance and thus leaf litter decomposition123.
In most cases, a reduction in aquatic biodiversity is likely to be the initial ecological effect of predicted future increases in drying. These taxonomic changes have functional consequences, altering ecosystem processes and associated ecosystem services. Although biological communities have proven resilient to drying, recovering within weeks to a few years even from rare dry phases in NPRs124,125, predicted future changes in riverine flow regimes have increasing potential to tip ecosystems to new, functionally compromised stable states. For example, decreases in flowing-phase duration and frequency could interact with concurrent stressors such as artificial enrichment by inorganic nutrients to shift aquatic vegetation communities from habitat-forming plants to filamentous algae. This change would alter basal food resources and habitat availability for invertebrates and fish, triggering trophic cascades that extend through food webs. In addition, plants act as ecosystem engineers that alter sediment dynamics, and therefore their loss could alter river shape126,127. By contrast, the ecological consequences of region-specific shifts towards perenniality could increase network-scale hydrological connectivity, promoting biotic dispersal and thus homogenizing communities90,128,129.
Managing NPRs in river networks
The effects of alternating flowing, non-flowing and dry phases on water quality, biodiversity, ecosystem processes and ecosystem services at the river-network scale mean that management expectations of natural NPRs must differ from those of perennial segments93,130,131. In particular, to be effective, management, conservation and restoration of river networks must explicitly recognize perennial segments and NPRs and their multifaceted connections. This section discusses the management implications of the presence of NPRs in river networks and of the connections between perennial segments and NPRs.
Management implications of non-perenniality
Understanding the connections between perennial segments and NPRs, and with other connected waters (lakes, reservoirs, wetlands, aquifers, estuaries and in coastal areas), is a crucial step towards integrated management of river networks. The processes mediated by these connections have major network-scale implications for biodiversity conservation, water quality management, mitigation of risks posed by floods and droughts and the provisioning of ecosystem services.
Dry-phase refuges located in NPRs are crucial to maintaining freshwater biodiversity at the river-network scale52,132. For example, disconnected pools maintained in intermittent streams in coastal Oregon133 and in tributaries of the Russian River52 during dry phases provide refuges that promote the survival of juvenile Coho salmon (Fig. 6a). In the Russian River tributaries, the mean cumulative survival of salmon in these pools reached 50% (ref. 52). The deterioration or loss of these habitats, together with the lack of access to them owing to fragmentation by anthropogenic barriers134, reduces fish populations, which, in turn, affects subsistence fishers52. Therefore, management strategies for biodiversity conservation of entire river networks should prioritize the identification and protection of these refuges135,136.
Figure 6.
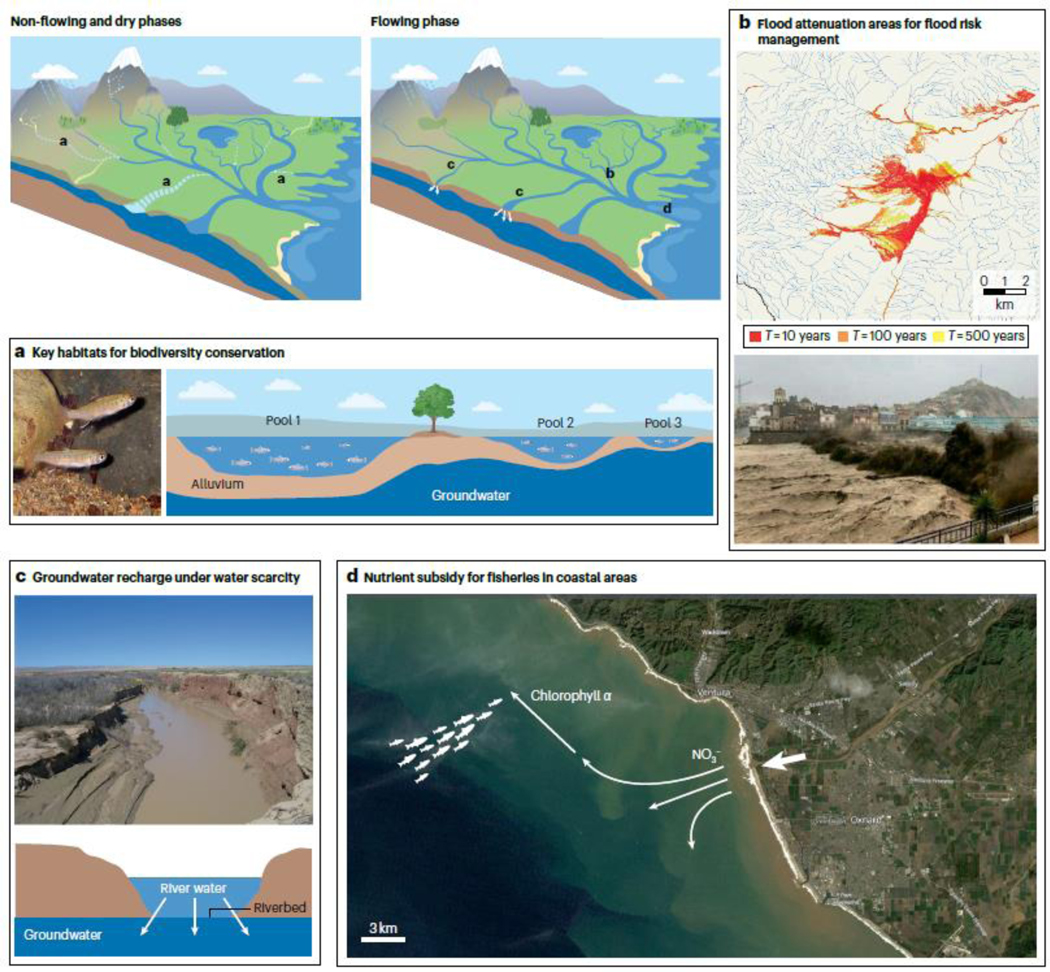
Examples of ecosystem processes and services occurring in non-perennial segments and management opportunities. a, Disconnected pools in a non-perennial river segment and Coho salmon juveniles in a non-perennial segment from the Russian River in California, USA. b, Rambla de Nogalte (southeastern Spain) during a flood in 2012, and an example of map showing inundation risks. c, The Río Puerco, a tributary of Río Grande, USA, during the flowing period and diagram showing groundwater recharge47. d, Satellite image of Santa Clara ephemeral river inputs to the Channel Islands in California, USA, during flooding in 1998. Nutrients and chlorophyll a were carried out from the river to the coastal waters during these floods139. The big white arrow shows the river mouth. Panel a schematic is adapted from ref. 190 CC BY 4.0. Panel a image courtesy of the Russian River Monitoring Program. Panel b image is reprinted with permission from ref. 190, CC BY 4.0. Panels c and d photo reprinted with permission from the USGS.
NPRs can attenuate floods and act as flood protection zones in the catchment (Fig. 6b). For example, the dry channel of the ephemeral river Rambla de Nogalte, southeastern Spain efficiently absorbs flash flood waters and sediments except where walls and embankments have been built137. Infiltration potential (the extent to which water can enter the sediments) in NPRs should be integrated into flood risk management at the river-network scale by identifying priority zones where this function is aximized. The high infiltration capacity of dry riverbeds in NPRs can also limit evaporative losses and facilitate groundwater recharge (Fig. 6c). For example, flooding after a rain event in the ephemeral Sand River, Kenya, recharged the groundwater level in only 1.5 h (ref. 138). Similarly, 49% of the monsoon flood volume from the ephemeral Río Puerco basin in New Mexico, USA recharged the aquifer and the rest entered a downstream reservoir47. The resultant availability of water can be a major benefit for people living and depending on these resources in arid and semi-arid regions.
Nutrients released from NPRs during flowing phases can subsidize downstream connected waters and support biodiversity and ecosystem functioning downstream, enhancing services provided by freshwater and marine fisheries (Fig. 6d). For example, the timing of a nutrient pulse from the ephemeral Santa Clara River (California, USA) to at least 20 km offshore during the 1998 floods was key to supporting marine productivity at a time when nutrient inputs from oceanic upwelling were less available139 (Fig. 6d).
Non-perenniality has major consequences for the provisioning, regulating and cultural ecosystem services made available by river networks. River drying prevents surface-water abstraction for uses such as crop irrigation and public water supply140. However, in many regions, a substantial fraction of public water supply comes from sources that include NPRs141 or their underlying groundwater sources142. Surface water loss is also likely to reduce fish populations that support subsistence fishers52. Regulating services such as water purification, flood mitigation and climate regulation are all compromised by drying143. For example, drying eliminates desiccation-sensitive microorganisms from biofilms and slows assimilation of inorganic nutrients (including anthropogenic pollutants) after flow resumes144. Drying also affects the cultural services provided by river networks by limiting water-associated activities such as boating while creating opportunities for new activities such as rambling140,145,146. These impacts on cultural services depend on how people perceive drying147,148, which greatly affects how river networks with extensive NPRs are likely to be managed.
Improving river-network management
Human activities alter flow regimes in NPRs, with major implications for functional connections and nearby perennial segments. However, compared with perennial segments, these NPRs are seldom as well protected by legislation and associated regulations from the impacts of human activities that might alter their flow101,149 (Fig. 7). Indeed, management practices have yet to be adapted to match new conceptual developments in river science4,5,101 that recognize the ecological importance of non-perenniality. Such adaptations in management practices would necessarily extend to other connected waters, such as floodplains, lakes and alluvial groundwaters, as well as the estuaries and near-shore coastal waters associated with river networks that drain to the sea.
Figure 7.

Examples of threats on non-perennial river segments. Rubbish in the dry riverbed of the Hodgsons Creek, Victoria (panel a) and in Madura gully, West Australia (panel b). A non-perennial segment of the Chitterne Brook flows through an intensively grazed cow pasture in England, UK (panel c). Sheep in the non-flowing segments of the Barranc del Carraixet, Spain (panel d). Sewage effluent turning the non-perennial segment of the Sant Miquel River artificially perennial in Spain (panel e). Gravel extractions from dry riverbeds in France (Albarine River) (panel f) and Bolivia ( Janq’u Qala) (panel g). A map showing the non-perennial river segments to be removed from protection by legislation in France (white; one of which is shown in panel j), on the basis of their non-perenniality (panel h); the perennial segment (yellow) is the only legally protected part of the river network. Sewage effluent generates a permanent pool in a non-perennial segment of the Albarine River, France (panel i). Non-perennial river segment that is no longer under protection in eastern France, le Ruisseau des Tendasses (panel j). Photos courtesy of T. Sykes (panel c) and H. Pella (panels j and h).
River-network management can be improved by recognizing and protecting the functional connections between perennial segments and NPRs. For example, if the legislative and regulatory protection of perennial segments in many countries were to be automatically extended to NPRs and their catchments, it would likely reduce the impacts of human activities that threaten the biodiversity and ecosystem functions of river networks and their connected ecosystems (Fig. 7). Some nations already have limited legislative protection for NPRs. In Australia, independent scientific assessment of the potential environmental impacts of large coal mining and coal seam gas developments on water-dependent biota and ecosystems in both perennial and non-perennial segments in the disturbance footprint is expressly required under legislation (the 2013 ‘water trigger’ amendment of the Environment Protection and Biodiversity Conservation Act 1999150).
Major obstacles need to be overcome to extend legislative protection for NPRs149,151. In Europe, the Water Framework Directive (WFD)152 largely omits NPRs from conservation, restoration and biomonitoring153. For example, the WFD only recognizes NPRs in one ‘river type’ in some countries in the Mediterranean region. Elsewhere, where NPRs are less prevalent but nonetheless diverse and extensive, such as in France154, the UK51 and the Czech Republic155, river typologies used to implement the WFD do not distinguish between perennial segments and NPRs. In some cases, such as France, there are attempts to remove NPRs from national legislation and regulations (Fig. 7h). In the USA, under implementation of the Clean Water Act, some NPRs are not included as Waters of the United States, potentially exposing them to impacts from activities such as dredging and waste dumping.
Another obstacle is the public perception of NPRs in river networks as less valuable than perennially flowing waters147,148,156. Consequently, NPRs are often overlooked in restoration and conservation plans148 and there is little appreciation of their ecosystem services143,146. Even in Australia, where the need for separate water quality guidelines for NPRs is accepted157, there is limited appreciation of the importance of connectivity between perennial segments and NPRs in influencing water quality. Scientists need to communicate the importance of ecosystem services provided by NPRs to the general public, river managers, politicians, policy makers and other stakeholders. Such evidence-informed actions could include producing fact sheets and policy briefs, using social media and conducting collaborative research projects with citizen scientists, river managers and stakeholders. For example, the open-source smartphone application DryRivers enables both citizen and professional scientists to map NPRs throughout Europe158 and has substantially increased public appreciation of the nature and extent of NPRs.
Logistically, there are serious limitations in fundamental hydrological data and hydrographic mapping for NPRs in river networks. Stream gauges that quantify flow and describe flow regime components of a stream segment are typically placed along larger perennial streams and fail to capture NPR flow regimes159. Most available maps are based on static, low-resolution surveys and cartography that omit many headwater NPRs36,160. Despite increasing efforts at various scales to statistically4,161–163 and mechanistically164,165 model the distribution and flow regimes of NPRs, such efforts are hampered by the scant stream gauge and groundwater-level data, which amplifies uncertainty over large areas4,163.
Further development and refinement of multiplatform remote sensing technology could be combined with modelling approaches that target stream gauging or field observations to reduce bias and fill gaps. Such approaches would enable the production of hydrographic maps that better reflect the dynamic connections between all segments in river networks. Remote-sensing platforms with synthetic aperture radar (SAR) are better able to capture surface water blocked by clouds, vegetation or shadows than multispectral platforms such as LandSat or CubeSat166,167. Future use of high spatial and temporal resolution SAR datasets (for example, from NASA-ISRO SAR) to map NPRs, in tandem with advances in data interpretation168, could support better integration of NPRs in distributed hydrological models169.
Another major obstacle to more effective management of connected perennial segments and NPRs in river networks is the limited availability of monitoring tools and approaches that perform equally well in both segment types. Most river management tools have been developed primarily or exclusively for perennial segments131,170,171 and typically perform poorly in NPRs. For example, biomonitoring indices used to indicate river health can rarely disentangle the effects of drying from those of stressors associated with human activities131,170,172,173. Functional traits173, metasystem approaches174, molecular tools175 and data on composition of terrestrial and semi-aquatic assemblages176,177 all have the potential to enhance assessment of NPR health. Developing common tools and approaches that are applicable in both perennial segments and NPRs could encourage river managers to include both types of segments in river-network biomonitoring.
Restoration and conservation of NPRs
Many NPRs are severely degraded by human activities and thus require restoration to recover lost biodiversity and ecological integrity. NPRs that are not degraded are seldom adequately protected yet many urgently need conservation to preserve their current values. However, ignorance of these values, especially in ephemeral NPRs178, and of the importance of NPRs to connected perennial segments has meant that efforts to restore or conserve NPRs are rare. It is likely that the same tools and approaches used for conserving and restoring perennial segments are equally applicable to NPRs. However, expectations of the outcomes, especially rates and trajectories of responses to restoration, need to consider inherent intermittence and resilience of these systems to different types of impacts (such as altered flow regimes versus altered water quality).
The few attempts at targeted restoration of NPRs have focused on riparian revegetation179. Of these attempts, even fewer have sought to evaluate restoration success or investigate pathways and mechanisms of ecological recovery. An experiment assessing ecosystem responses to reach-scale riparian replanting and livestock exclusion in three degraded NPRs in southeastern Australia found no differences in water quality, organic matter or aquatic invertebrate community composition between paired treatment and control sites after 6–8 years, ascribing the lack of response to a drought and pervasive effects of catchment-scale degradation180. The effectiveness of NPR restoration activities could also be enhanced by recovering natural flow regimes, reducing pollutant inputs, remediating degraded catchments, controlling invasive species and repairing damaged channels and streambeds. These multiple restoration activities must be implemented at appropriately broad spatial scales, and must prioritize recovery of lost functional connections between NPRs and perennial segments (such as through removal of instream barriers such as dams). As responses to restoration in NPRs are likely to be slower than in equivalent-sized perennial segments because of their inherent intermittence and often-arid or semi-arid climatic setting, expectations must be modified to reflect these key differences.
Similar to restoration, targeted conservation of NPRs at the river-network scale is rare, with most protected NPRs occurring in areas conserved for other reasons. For example, conservation actions targeting the endangered Coho salmon in the Russian River consisted of fish rescues during the non-flowing phase181. In NPRs in Texas, USA, local habitat restoration increased populations of the endangered desert pupfish (Cyprinidon bovinus)182. In Australia, standard methods have been developed to classify disconnected pools (riverine waterholes)183,184, informing actions taken to protect pools that act as refuges for biodiversity during drought58,184. However, connectivity is required among individual refuges to maintain metapopulation and metacommunity dynamics5, thus achieving effective network-scale conservation. In addition, effective network-scale conservation should seek to identify and protect key NPRs and their riparian zones. Systematic conservation planning tools such as Marxan185 are powerful approaches for identifying priority sites acting as refuges for fish across entire river networks135,136 and for evaluating the conservation value of NPRs considering both their aquatic and terrestrial species186.
We suggest that the most effective approaches for river-network management will be multifaceted, integrating targeted conservation and restoration strategies in an appropriate legislative context and explicitly acknowledging the importance of functional connections between NPRs and perennial segments. Although some biomonitoring approaches developed for perennial segments are effective in NPRs during flowing phases, biological indicators should include terrestrial communities to encompass dry phases and thus provide a more nuanced and comprehensive perspective of ecological responses to the management strategies176.
Summary and future perspectives
Repeated cycles of flowing, non-flowing and, in particular, dry phases govern the biodiversity and ecosystem processes of NPRs, in turn influencing ecosystem dynamics in connected perennial segments and downstream waters. NPR can function as sources, sinks and refuges for water, energy, materials and organisms and can delay and transform such ecosystem components, thus governing their fluxes across these connections. We contend that scientific recognition of the importance of these hydrological connections between perennial segments and NPRs in spatially variable river networks must be matched with a shift in river management. To facilitate such a shift, policy developments are needed to extend the legislation and regulations that protect perennial rivers to include NPRs.
There are five specific and actionable research domains within the next 3–5 years to further characterize how NPRs influence connected perennial segments and what this means for effective management at the river-network scale. First, researchers should identify how different functional connections affect fluxes of water, materials and organisms from NPRs to perennial segments at different locations within river networks (for example, upstream versus downstream segments). One approach is to use experimental manipulations to identify causal mechanisms. Such experiments should explore multiple fluxes concurrently owing to their likely interacting effects.
Second, the influence of physical and hydrological features on different types of fluxes must be characterized. For example, fluxes and their effects should be compared in small ephemeral NPRs and larger intermittent ones, as well as in single-thread and braided NPRs. Third, along these lines, the collective effects of different functional connections on ecosystem services provided by whole river networks must be evaluated and understood. For example, lagged connections could nullify or delay the influence of other types of connections upstream and alter the types, timing and location of ecosystem services contributed by NPRs. These three points largely relate to improving our knowledge of NPRs and their role in river networks, but understanding how human impacts and management actions alter NPRs is equally important.
Therefore, fourth, researchers must analyse how human activities modify functional connections in different river networks and, in particular, impact the provision of multiple ecosystem services. For example, the clearance — or restorative planting — of riparian vegetation along NPRs could change downstream functional connections and fluxes of organic matter. Finally, the scientific evidence provided by such research must be used to inform management actions as well as policy developments that enhance holistic legislative and regulatory protection for NPRs within river networks, to stop ongoing losses of biodiversity and ecological functions in river networks worldwide.
Sustaining the integrity of entire river networks and the quality of downstream waters, including rivers, lakes, reservoirs, groundwaters, estuaries and coastal areas, requires integrated management strategies that consider NPRs and their interconnections with perennial waters. Neglecting the important roles of NPRs compromises effective river management and could ultimately undermine actions taken to support the resilience of river networks as they adapt to global change.
Key Points.
Non-perennial segments comprise over half of the global river network. Ongoing climate change and human activities will further increase the occurrence of river drying.
Recurrent cycles of flowing, non-flowing and dry phases influence exchanges of water, energy, nutrients and organisms between non-perennial segments and connected perennial waters.
Physical, chemical and biological processes in non-perennial segments affect water quality and quantity, and ecological integrity in downstream receiving waters and entire river networks.
Historically, river science and management have focused on perennial river segments, neglecting the ubiquity and importance of non-perennial segments. This imbalance has often led to environmental problems such as poor water quality, loss of biodiversity and alteration of natural flow regimes at the river-network scale.
Sustaining the water quality and ecological integrity of entire river networks and associated downstream waters requires integrated management strategies that explicitly consider non-perennial segments and their connections with perennial ones.
Acknowledgements
T.D. and N.C. received support from the European Union’s Horizon 2020 Research and Innovation programme through the DRYvER project (Securing Biodiversity, Functional Integrity and Ecosystem Services in Drying River Networks, award number 869226). The views expressed in this paper are those of the authors and do not necessarily reflect the views or policies of the USEPA.
Glossary
- β-Diversity
Spatial and temporal variability in community composition
- Dry phases
In a non-perennial river segment, a period of time with no spatially continuous flowing or non-flowing surface water, although disconnected surface-water pools and subsurface water can be present
- Drying phase
In a non-perennial river segment, the transitional period between a flowing or non-flowing phase and a dry phase, during which most or all surface water is lost
- Ephemeral
A non-perennial flow regime in which water only flows in response to rainfall events, and flowing phases are thus unpredictable and typically short (hours to weeks)
- Flow cessation
The point in time at which surface water ceases to flow from upstream to downstream in a non-perennial segment
- Flowing phases
In a non-perennial river segment, the periods of time in which water flows from upstream to downstream
- Flow regime
The temporal variability in the quantity and timing of discharge
- Gaining segments
Stream segments in which flow increases owing to the upwelling of groundwater into the surface channel
- Intermittent
A non-perennial flow regime, often seasonal, that is typically characterized by long flowing phases (usually multiple months) and short dry phases
- Losing segments
Stream segments in which flow decreases owing to the infiltration of surface water into the streambed towards the groundwater
- Megadroughts
Droughts that exceed the duration of most droughts in the instrumental record
- Non-flowing phases
In a non-perennial river segment, the periods of time in which spatially continuous non-flowing (still or lentic) surface water is present
- Non-perennial segments
Stream segments in which surface water recurrently stops flowing. These segments lose all or most of their surface water
- Perennial segments
Stream segments in which surface water never stops flowing
- Rewetting phase
In a non-perennial river segment, the transitional period between a dry phase and a flowing or non-flowing phase
- Synchrony
The degree of concurrent change across spatially distinct segments or populations
Footnotes
Competing interests
The authors declare no competing interests.
References
- 1.Tonkin JD et al. Prepare river ecosystems for an uncertain future. Nature 570, 301–303 (2019). [DOI] [PubMed] [Google Scholar]
- 2.Tickner D. et al. Bending the curve of global freshwater biodiversity loss: an emergency recovery plan. BioScience 70, 330–342 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Reid AJ et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev 94, 849–873 (2019). [DOI] [PubMed] [Google Scholar]
- 4.Messager ML et al. Global prevalence of non-perennial rivers and streams. Nature 594, 391–397 (2021). [DOI] [PubMed] [Google Scholar]
- 5.Cid N. et al. From meta-system theory to the sustainable management of rivers in the Anthropocene. Front. Ecol. Environ 20, 49–57 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Uys MC & O’Keeffe JH Simple words and fuzzy zones: early directions for temporary river research in South Africa. Environ. Manage 21, 517–531 (1997). [DOI] [PubMed] [Google Scholar]
- 7.Williams DD The Biology of Temporary Waters (Oxford Univ. Press, 2006). [Google Scholar]
- 8.Gallart F. et al. A novel approach to analysing the regimes of temporary streams in relation to their controls on the composition and structure of aquatic biota. Hydrol. Earth Syst. Sci 16, 3165–3182 (2012). [Google Scholar]
- 9.Busch MH et al. What’s in a name? Patterns, trends, and suggestions for defining non-perennial rivers and streams. Water 12, 1980 (2020). [PMC free article] [PubMed] [Google Scholar]
- 10.Datry T. et al. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 377–403 (Academic Press, 2017). [Google Scholar]
- 11.Boulton AJ, Rolls RJ, Jaeger KL & Datry T. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 79–108 (Academic Press, 2017). [Google Scholar]
- 12.Zipper SC et al. Pervasive changes in stream intermittency across the United States. Environ. Res. Lett 16, 084033 (2021). [Google Scholar]
- 13.Zhang Y. et al. Future global streamflow declines are probably more severe than previously estimated. Nat. Water 1, 261–271 (2023). [Google Scholar]
- 14.Cuevas JG et al. Spatial distribution and pollution evaluation in dry riverbeds affected by mine tailings. Environ. Geochem. Health 10.1007/s10653-022-01469-5 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Poff NL et al. The natural flow regime. BioScience 47, 769–784 (1997). [Google Scholar]
- 16.Datry T, Larned ST & Tockner K. Intermittent rivers: a challenge for freshwater ecology. BioScience 64, 229–235 (2014). [Google Scholar]
- 17.Bogan MT et al. Resistance, resilience, and community recovery in intermittent rivers and ephemeral streams. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 349–376 (Academic Press, 2017). [Google Scholar]
- 18.Gauthier M. et al. Fragmentation promotes the role of dispersal in determining 10 intermittent headwater stream metacommunities. Freshw. Biol 65, 2169–2185 (2020). [Google Scholar]
- 19.Diamond JS et al. Light and hydrologic connectivity drive dissolved oxygen synchrony in stream networks. Limnol. Oceanogr 68, 322–335 (2023). [Google Scholar]
- 20.Boulton AJ Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshw. Biol 48, 1173–1185 (2003). [Google Scholar]
- 21.Stubbington R. The hyporheic zone as an invertebrate refuge: a review of variability in space, time, taxa and behaviour. Mar. Freshw. Res 63, 293–311 (2012). [Google Scholar]
- 22.DelVecchia AG et al. Reconceptualizing the hyporheic zone for nonperennial rivers and streams. Freshw. Sci 41, 167–182 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Malard F, Tockner K, Dole-Olivier M-J & Ward JV A landscape perspective of surface–subsurface hydrological exchanges in river corridors. Freshw. Biol 47, 621–640 (2002). [Google Scholar]
- 24.Boulton AJ, Datry T, Kasahara T, Mutz M. & Stanford JA Ecology and management of the hyporheic zone: stream–groundwater interactions of running waters and their floodplains. J. North. Am. Benthol. Soc 29, 26–40 (2010). [Google Scholar]
- 25.Gómez-Gener L. et al. Towards an improved understanding of biogeochemical processes across surface–groundwater interactions in intermittent rivers and ephemeral streams. Earth-Sci. Rev 220, 103724 (2021). [Google Scholar]
- 26.Arscott D, Tockner K, van der Nat D. & Ward J. Aquatic habitat dynamics along a braided alpine river ecosystem (Tagliamento River, Northeast Italy). Ecosystems 5, 0802–0814 (2002). [Google Scholar]
- 27.Corti R. & Datry T. Invertebrates and sestonic matter in an advancing wetted front travelling down a dry river bed (Albarine, France). Freshw. Sci 31, 1187–1201 (2012). [Google Scholar]
- 28.Costigan KH et al. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 51–78 (Academic Press, 2017). [Google Scholar]
- 29.Shanafield M, Bourke S, Zimmer M. & Costigan K. An overview of the hydrology of non-perennial rivers and streams. Wiley Interdiscip. Rev. Water 8, e1504 (2021). [Google Scholar]
- 30.Zipper S, Popescu I, Compare K, Zhang C. & Seybold EC Alternative stable states and hydrological regime shifts in a large intermittent river. Environ. Res. Lett 17, 074005 (2022). [Google Scholar]
- 31.Costigan KH, Jaeger KL, Goss CW, Fritz KM & Goebel PC Understanding controls on flow permanence in intermittent rivers to aid ecological research: integrating meteorology, geology and land cover. Ecohydrology 9, 1141–1153 (2016). [Google Scholar]
- 32.Hammond JC et al. Spatial patterns and drivers of non-perennial flow regimes in the contiguous United States. Geophys. Res. Lett 48, e2020GL090794 (2021). [Google Scholar]
- 33.Datry T. et al. A global analysis of terrestrial plant litter dynamics in non-perennial waterways. Nat. Geosci 11, 497–503 (2018). [Google Scholar]
- 34.Wohl E. Rivers in the critical zone. Dev. Earth Surf. Process 19, 267–293 (2015). [Google Scholar]
- 35.Gounand I, Harvey E, Little CJ & Altermatt F. Meta-ecosystems 2.0: rooting the theory into the field. Trends Ecol. Evol 33, 36–46 (2018). [DOI] [PubMed] [Google Scholar]
- 36.Benstead JP & Leigh DS An expanded role for river networks. Nat. Geosci 5, 678–679 (2012). [Google Scholar]
- 37.Pineda-Morante D. et al. Local hydrological conditions and spatial connectivity shape invertebrate communities after rewetting in temporary rivers. Hydrobiologia 849, 1511–1530 (2022). [Google Scholar]
- 38.Datry T. Benthic and hyporheic invertebrate assemblages along a flow intermittence gradient: effects of duration of dry events. Freshw. Biol 57, 563–574 (2012). [Google Scholar]
- 39.Costelloe JF, Grayson RB, Argent RM & McMahon TA Modelling the flow regime of an arid zone floodplain river, Diamantina River, Australia. Environ. Model. Softw 18, 693–703 (2003). [Google Scholar]
- 40.Larned ST, Datry T, Arscott DB & Tockner K. Emerging concepts in temporary-river ecology. Freshw. Biol 55, 717–738 (2010). [Google Scholar]
- 41.Arscott DB, Larned S, Scarsbrook MR & Lambert P. Aquatic invertebrate community structure along an intermittence gradient: Selwyn River, New Zealand. J. North. Am. Benthol. Soc 29, 530–545 (2010). [Google Scholar]
- 42.Grodek T. et al. The last millennium largest floods in the hyperarid Kuiseb River basin, Namib Desert. J. Quat. Sci 28, 258–270 (2013). [Google Scholar]
- 43.Döll P. & Schmied HM How is the impact of climate change on river flow regimes related to the impact on mean annual runoff? A global-scale analysis. Environ. Res. Lett 7, 014037 (2012). [Google Scholar]
- 44.Capderrey C, Datry T, Foulquier A, Claret C. & Malard F. Invertebrate distribution across nested geomorphic features in braided-river landscapes. Freshw. Sci 32, 1188–1204 (2013). [Google Scholar]
- 45.Leibowitz SG et al. Connectivity of streams and wetlands to downstream waters: an integrated systems framework. J. Am. Water Resour. Assoc 54, 298–322 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shumilova O. et al. Simulating rewetting events in intermittent rivers and ephemeral streams: a global analysis of leached nutrients and organic matter. Glob. Change Biol 25, 1591–1611 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Vivoni ER, Bowman RS, Wyckoff RL, Jakubowski RT & Richards KE Analysis of a monsoon flood event in an ephemeral tributary and its downstream hydrologic effects. Water Resour. Res 42, W03404 (2006). [Google Scholar]
- 48.Dahm CN, Candelaria-Ley RI, Reale CS, Reale JK & van Horn DJ Extreme water quality degradation following a catastrophic forest fire. Freshw. Biol 60, 2584–2599 (2015). [Google Scholar]
- 49.Levick LR et al. The Ecological and Hydrological Significance of Ephemeral and Intermittent Streams in the Arid and Semi-Arid American Southwest. Report No. EPA/600/R-08/134, ARS/233046 (US Environmental Protection Agency, Office of Research and Development, 2008). [Google Scholar]
- 50.Sheldon F. et al. Ecological roles and threats to aquatic refugia in arid landscapes: dryland river waterholes. Mar. Freshw. Res 61, 885–895 (2010). [Google Scholar]
- 51.Stubbington R, England J, Wood PJ & Sefton CEM Temporary streams in temperate zones: recognizing, monitoring and restoring transitional aquatic–terrestrial ecosystems. WIREs Water 4, e1223 (2017). [Google Scholar]
- 52.Vander Vorste R, Obedzinski M, Nossaman Pierce S, Carlson SM & Grantham TE Refuges and ecological traps: extreme drought threatens persistence of an endangered fish in intermittent streams. Glob. Change Biol 26, 3834–3845 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Arias-Real R, Gutiérrez-Cánovas C, Menéndez M, Granados V. & Muñoz I. Diversity mediates the responses of invertebrate density to duration and frequency of rivers’ annual drying regime. Oikos 130, 2148–2160 (2021). [Google Scholar]
- 54.Steward AL, Datry T. & Langhans SD The terrestrial and semi-aquatic invertebrates of intermittent rivers and ephemeral streams. Biol. Rev 97, 1408–1425 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Soria M, Leigh C, Datry T, Bini LM & Bonada N. Biodiversity in perennial and intermittent rivers: a meta-analysis. Oikos 126, 1078–1089 (2017). [Google Scholar]
- 56.Corti R. & Datry T. Terrestrial and aquatic invertebrates in the riverbed of an intermittent river: parallels and contrasts in community organisation. Freshw. Biol 61, 1308–1320 (2016). [Google Scholar]
- 57.Stubbington R. et al. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 217–243 (Academic Press, 2017). [Google Scholar]
- 58.Marshall JC et al. Go with the flow: the movement behaviour of fish from isolated waterhole refugia during connecting flow events in an intermittent dryland river. Freshw. Biol 61, 1242–1258 (2016). [Google Scholar]
- 59.Pařil P. et al. An unexpected source of invertebrate community recovery in intermittent streams from a humid continental climate. Freshw. Biol 64, 1971–1983 (2019). [Google Scholar]
- 60.Stubbington R. & Datry T. The macroinvertebrate seedbank promotes community persistence in temporary rivers across climate zones. Freshw. Biol 58, 1202–1220 (2013). [Google Scholar]
- 61.Barthès A. et al. Impact of drought on diatom communities and the consequences for the use of diatom index values in the River Maureillas (Pyrénées-Orientales, France). River Res. Appl 31, 993–1002 (2015). [Google Scholar]
- 62.Fournier RJ, de Mendoza G, Sarremejane R. & Ruhi A. Isolation controls reestablishment mechanisms and post-drying community structure in an intermittent stream. Ecology 104, e3911 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sarremejane R. et al. Stochastic processes and ecological connectivity drive stream invertebrate community responses to short-term drought. J. Anim. Ecol 90, 886–898 (2021). [DOI] [PubMed] [Google Scholar]
- 64.Gauthier M, Goff GL, Launay B, Douady CJ & Datry T. Dispersal limitation by structures is more important than intermittent drying effects for metacommunity dynamics in a highly fragmented river network. Freshw. Sci 40, 302–315 (2021). [Google Scholar]
- 65.Di Sabatino A, Coscieme L. & Cristiano G. No post-drought recovery of the macroinvertebrate community after five months upon rewetting of an irregularly intermittent Apennine River (Aterno River). Ecohydrol. Hydrobiol 10.1016/j.ecohyd.2022.11.005 (2022). [DOI] [Google Scholar]
- 66.von Schiller D. et al. Sediment respiration pulses in intermittent rivers and ephemeral streams. Glob. Biogeochem. Cycles 33, 1251–1263 (2019). [Google Scholar]
- 67.Placella S, Brodie E. & Firestone M. Rainfall-induced carbon dioxide pulses result from sequential resuscitation of phylogenetically clustered microbial groups. Proc. Natl Acad. Sci. USA 109, 10931–10936 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Arce MI, Sánchez-Montoya MM & Gómez R. Nitrogen processing following experimental sediment rewetting in isolated pools in an agricultural stream of a semiarid region. Ecol. Eng 77, 233–241 (2015). [Google Scholar]
- 69.del Campo R, Foulquier A, Singer G. & Datry T. in The Ecology of Plant Litter Decomposition in Stream Ecosystems (eds Swan CM, Boyero L. & Canhoto C) 73–100 (Springer, 2021). [Google Scholar]
- 70.Lauerwald R. et al. Inland water greenhouse gas budgets for RECCAP2: 1. State-of-the-art of global scale assessments. Glob. Biogeochem. Cycles 37, e2022GB007657 (2023). [Google Scholar]
- 71.B-Béres V. et al. Autumn drought drives functional diversity of benthic diatom assemblages of continental intermittent streams. Adv. Water Resour 126, 129–136 (2019). [Google Scholar]
- 72.Crabot J. et al. A global perspective on the functional responses of stream communities to flow intermittence. Ecography 44, 1511–1523 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Aspin TWH et al. Extreme drought pushes stream invertebrate communities over functional thresholds. Glob. Change Biol 25, 230–244 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ledger ME, Brown LE, Edwards FK, Milner AM & Woodward G. Drought alters the structure and functioning of complex food webs. Nat. Clim. Change 3, 223–227 (2013). [Google Scholar]
- 75.Foulquier A, Artigas J, Pesce S. & Datry T. Drying responses of microbial litter decomposition and associated fungal and bacterial communities are not affected by emersion frequency. Freshw. Sci 34, 1233–1244 (2015). [Google Scholar]
- 76.del Campo R, Corti R. & Singer G. Flow intermittence alters carbon processing in rivers through chemical diversification of leaf litter. Limnol. Oceanogr. Lett 6, 232–242 (2021). [Google Scholar]
- 77.Price AN, Jones CN, Hammond JC, Zimmer MA & Zipper SC The drying regimes of non-perennial rivers and streams. Geophys. Res. Lett 48, e2021GL093298 (2021). [Google Scholar]
- 78.Sauquet E, Beaufort A, Sarremejane R. & Thirel G. Predicting flow intermittence in France under climate change. Hydrol. Sci. J 66, 2046–2059 (2021). [Google Scholar]
- 79.Fritz KM, Pond GJ, Johnson BR & Barton CD Coarse particulate organic matter dynamics in ephemeral tributaries of a Central Appalachian stream network. Ecosphere 10, e02654 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hladyz S, Watkins SC, Whitworth KL & Baldwin DS Flows and hypoxic blackwater events in managed ephemeral river channels. J. Hydrol 401, 117–125 (2011). [Google Scholar]
- 81.Walczak N, Walczak Z. & Nieć J. Influence of debris on water intake gratings in small hydroelectric plants: an experimental study on hydraulic parameters. Energies 14, 3248 (2021). [Google Scholar]
- 82.Larsen S. et al. The geography of metapopulation synchrony in dendritic river networks. Ecol. Lett 24, 791–801 (2021). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Schindler DE, Armstrong JB & Reed TE The portfolio concept in ecology and evolution. Front. Ecol. Environ 13, 257–263 (2015). [Google Scholar]
- 84.Moore JW et al. Emergent stability in a large, free-flowing watershed. Ecology 96, 340–347 (2015). [DOI] [PubMed] [Google Scholar]
- 85.Ruetz CR, Trexler JC, Jordan F, Loftus WF & Perry SA Population dynamics of wetland fishes: spatio-temporal patterns synchronized by hydrological disturbance? J. Anim. Ecol 74, 322–332 (2005). [Google Scholar]
- 86.Sarremejane R. et al. Drought effects on invertebrate metapopulation dynamics and quasi-extinction risk in an intermittent river network. Glob. Change Biol 27, 4024–4039 (2021). [DOI] [PubMed] [Google Scholar]
- 87.Crabot J, Heino J, Launay B. & Datry T. Drying determines the temporal dynamics of stream invertebrate structural and functional beta diversity. Ecography 43, 620–635 (2020). [Google Scholar]
- 88.Sarremejane R, Mykrä H, Bonada N, Aroviita J. & Muotka T. Habitat connectivity and dispersal ability drive the assembly mechanisms of macroinvertebrate communities in river networks. Freshw. Biol 62, 1073–1082 (2017). [Google Scholar]
- 89.Datry T, Bonada N. & Heino J. Towards understanding the organisation of metacommunities in highly dynamic ecological systems. Oikos 125, 149–159 (2016). [Google Scholar]
- 90.Sarremejane R. et al. Local and regional drivers influence how aquatic community diversity, resistance and resilience vary in response to drying. Oikos 129, 1877–1890 (2020). [Google Scholar]
- 91.Cañedo-Argüelles M. et al. Dispersal strength determines meta-community structure in a dendritic riverine network. J. Biogeogr 42, 778–790 (2015). [Google Scholar]
- 92.Sarremejane R. et al. Do metacommunities vary through time? Intermittent rivers as model systems. J. Biogeogr 44, 2752–2763 (2017). [Google Scholar]
- 93.Stubbington R. et al. A comparison of biotic groups as dry-phase indicators of ecological quality in intermittent rivers and ephemeral streams. Ecol. Indic 97, 165–174 (2019). [Google Scholar]
- 94.Tonkin JD, Stoll S, Jähnig SC & Haase P. Contrasting metacommunity structure and beta diversity in an aquatic-floodplain system. Oikos 125, 686–697 (2016). [Google Scholar]
- 95.Ficklin DL, Abatzoglou JT, Robeson SM, Null SE & Knouft JH Natural and managed watersheds show similar responses to recent climate change. Proc. Natl Acad Sci. USA 115, 8553–8557 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Tramblay Y. et al. Trends in flow intermittence for European rivers. Hydrol. Sci. J 66, 37–49 (2021). [Google Scholar]
- 97.Ward AS, Wondzell SM, Schmadel NM & Herzog SP Climate change causes river network contraction and disconnection in the H.J. Andrews experimental forest, Oregon, USA. Front. Water 10.3389/frwa.2020.00007 (2020). [DOI] [Google Scholar]
- 98.Spinoni J, Naumann G, Carrao H, Barbosa P. & Vogt J. World drought frequency, duration, and severity for 1951–2010. Int. J. Climatol 34, 2792–2804 (2014). [Google Scholar]
- 99.Spinoni J, Vogt JV, Naumann G, Barbosa P. & Dosio A. Will drought events become more frequent and severe in Europe? Int. J. Climatol 38, 1718–1736 (2018). [Google Scholar]
- 100.Vicente-Serrano SM, Quiring SM, Peña-Gallardo M, Yuan S. & Domínguez-Castro F. A review of environmental droughts: increased risk under global warming? Earth-Sci. Rev 201, 102953 (2020). [Google Scholar]
- 101.Datry T. et al. Causes, responses, and implications of anthropogenic versus natural flow intermittence in river networks. BioScience 73, 9–22 (2023). [Google Scholar]
- 102.Toreti A. et al. Drought in Europe August 2022. Report no. JRC130493 (Publications Office of the European Union, 2022). [Google Scholar]
- 103.Kustu MD, Fan Y. & Robock A. Large-scale water cycle perturbation due to irrigation pumping in the US high plains: a synthesis of observed streamflow changes. J. Hydrol 390, 222–244 (2010). [Google Scholar]
- 104.Perkin JS et al. Groundwater declines are linked to changes in great plains stream fish assemblages. Proc. Natl. Acad. Sci. USA 114, 7373–7378 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Yuan X. et al. A global transition to flash droughts under climate change. Science 380, 187–191 (2023). [DOI] [PubMed] [Google Scholar]
- 106.Mazdiyasni O. & AghaKouchak A. Substantial increase in concurrent droughts and heatwaves in the United States. Proc. Natl Acad. Sci. USA 112, 11484–11489 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Sutanto SJ, Vitolo C, Di Napoli C, D’Andrea M. & Van Lanen HAJ Heatwaves, droughts, and fires: exploring compound and cascading dry hazards at the pan-European scale. Environ. Int 134, 105276 (2020). [DOI] [PubMed] [Google Scholar]
- 108.Tassone SJ et al. Increasing heatwave frequency in streams and rivers of the United States. Limnol. Oceanogr. Lett 8, 295–304 (2023). [Google Scholar]
- 109.Woodhouse CA & Overpeck JT 2000 Years of drought variability in the central United States. Bull. Am. Meteorol. Soc 79, 2693–2714 (1998). [Google Scholar]
- 110.Williams AP et al. Large contribution from anthropogenic warming to an emerging North American megadrought. Science 368, 314–318 (2020). [DOI] [PubMed] [Google Scholar]
- 111.Barnett TP, Adam JC & Lettenmaier DP Potential impacts of a warming climate on water availability in snow-dominated regions. Nature 438, 303–309 (2005). [DOI] [PubMed] [Google Scholar]
- 112.Milly PCD & Dunne KA Colorado River flow dwindles as warming-driven loss of reflective snow energizes evaporation. Science 367, 1252–1255 (2020). [DOI] [PubMed] [Google Scholar]
- 113.Drummond LR, McIntosh AR & Larned ST Invertebrate community dynamics and insect emergence in response to pool drying in a temporary river. Freshw. Biol 60, 1596–1612 (2015). [Google Scholar]
- 114.Stubbington R. et al. The response of perennial and temporary headwater stream invertebrate communities to hydrological extremes. Hydrobiologia 630, 299–312 (2009). [Google Scholar]
- 115.Jaeger KL, Olden JD & Pelland NA Climate change poised to threaten hydrologic connectivity and endemic fishes in dryland streams. Proc. Natl Acad. Sci. USA 111, 13894–13899 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Bogan MT, Leidy RA, Neuhaus L, Hernandez CJ & Carlson SM Biodiversity value of remnant pools in an intermittent stream during the great California drought. Aquat. Conserv. Mar. Freshw. Ecosyst 29, 976–989 (2019). [Google Scholar]
- 117.Hynes HBN The effect of drought on the fauna of a small mountain stream in Wales. Verhandlungen Int. Ver. Für Theor. Angew. Limnol 13, 826–833 (1958). [Google Scholar]
- 118.Bogan MT & Boersma KS Aerial dispersal of aquatic invertebrates along and away from arid-land streams. Freshw. Sci 31, 1131–1144 (2012). [Google Scholar]
- 119.Romaní A. & Sabater S. Metabolism recovery of a stromatolitic biofilm after drought in a Mediterranean stream. Arch. Hydrobiol 140, 261–271 (1997). [Google Scholar]
- 120.Acuña V, Casellas M, Corcoll N, Timoner X. & Sabater S. Increasing extent of periods of no flow in intermittent waterways promotes heterotrophy. Freshw. Biol 60, 1810–1823 (2015). [Google Scholar]
- 121.Ruffing CM et al. Prairie stream metabolism recovery varies based on antecedent hydrology across a stream network after a bank-full flood. Limnol. Oceanogr 67, 1986–1999 (2022). [Google Scholar]
- 122.Nyström M. Redundancy and response diversity of functional groups: implications for the resilience of coral reefs. Ambio 35, 30–35 (2006). [PubMed] [Google Scholar]
- 123.Matthaei CD, Piggott JJ & Townsend CR Multiple stressors in agricultural streams: interactions among sediment addition, nutrient enrichment and water abstraction. J. Appl. Ecol 47, 639–649 (2010). [Google Scholar]
- 124.Wood PJ & Petts GE The influence of drought on chalk stream macroinvertebrates. Hydrol. Process 13, 387–399 (1999). [Google Scholar]
- 125.Ledger ME & Hildrew AG Recolonization by the benthos of an acid stream following a drought. Arch. für Hydrobiol 152, 1–17 (2001). [Google Scholar]
- 126.Cotton JA, Wharton G, Bass JAB, Heppell CM & Wotton RS The effects of seasonal changes to in-stream vegetation cover on patterns of flow and accumulation of sediment. Geomorphology 77, 320–334 (2006). [Google Scholar]
- 127.Gurnell AM, van Oosterhout MP, de Vlieger B. & Goodson JM Reach-scale interactions between aquatic plants and physical habitat: river frome, dorset. River Res. Appl 22, 667–680 (2006). [Google Scholar]
- 128.Heino J. et al. Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshw. Biol 60, 845–869 (2015). [Google Scholar]
- 129.Rogosch JS & Olden JD Dynamic contributions of intermittent and perennial streams to fish beta diversity in dryland rivers. J. Biogeogr 46, 2311–2322 (2019). [Google Scholar]
- 130.Soria M. et al. Natural disturbances can produce misleading bioassessment results: identifying metrics to detect anthropogenic impacts in intermittent rivers. J. Appl. Ecol 57, 283–295 (2020). [Google Scholar]
- 131.Crabot J, Dolédec S, Forcellini M. & Datry T. Efficiency of invertebrate-based bioassessment for evaluating the ecological status of streams along a gradient of flow intermittence. Ecol. Indic 133, 108440 (2021). [Google Scholar]
- 132.Moidu H. et al. Ecological consequences of shifting habitat mosaics within and across years in an intermittent stream. Freshw. Biol 00, 1–15 (2023). [Google Scholar]
- 133.Wigington P Jr et al. Coho salmon dependence on intermittent streams. Front. Ecol. Environ 4, 513–518 (2006). [Google Scholar]
- 134.Archdeacon TP & Reale JK No quarter: lack of refuge during flow intermittency results in catastrophic mortality of an imperiled minnow. Freshw. Biol 65, 2108–2123 (2020). [Google Scholar]
- 135.Hermoso V, Ward DP & Kennard MJ Prioritizing refugia for freshwater biodiversity conservation in highly seasonal ecosystems. Divers. Distrib 19, 1031–1042 (2013). [Google Scholar]
- 136.Yu S, Rose PM, Bond NR, Bunn SE & Kennard MJ Identifying priority aquatic refuges to sustain freshwater biodiversity in intermittent streams in eastern Australia. Aquat. Conserv. Mar. Freshw. Ecosyst 32, 1584–1595 (2022). [Google Scholar]
- 137.Hooke JM Extreme sediment fluxes in a dryland flash flood. Sci. Rep 9, 1686 (2019). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Wekesa SS et al. Water flow behavior and storage potential of the semi-arid ephemeral river system in the Mara Basin of Kenya. Front. Environ. Sci 8, 95 (2020). [Google Scholar]
- 139.Warrick J, Washburn L, Brzezinski M. & Siegel D. Nutrient contributions to the Santa Barbara Channel, California, from the ephemeral Santa Clara River. Estuar. Coast. Shelf Sci 62, 559–574 (2005). [Google Scholar]
- 140.Koundouri P, Boulton A, Datry T. & Souliotis I. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 455–476 (Academic Press, 2017). [Google Scholar]
- 141.United States Environmental Protection Agency. Federal safe drinking water information system 4th quarter 2006 data. National Hydrography Dataset Plus at Medium Resolution https://www.epa.gov/cwa-404/surface-drinking-water-provided-intermittentephemeral-and-headwater-streams-state-maps (US EPA, 2023). [Google Scholar]
- 142.Allen DJ & Crane EJ The Chalk Aquifer of the Wessex Basin. Report no. RR/11/002 (British Geological Survey, 2019). [Google Scholar]
- 143.Datry T. et al. Flow intermittence and ecosystem services in rivers of the Anthropocene. J. Appl. Ecol 55, 353–364 (2018). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Timoner X, Acuña V, Von Schiller D. & Sabater S. Functional responses of stream biofilms to flow cessation, desiccation and rewetting. Freshw. Biol 57, 1565–1578 (2012). [Google Scholar]
- 145.Steward A, Von Schiller D, Tockner K, Marshall J. & Bunn S. When the river runs dry: human and ecological values of dry riverbeds. Front. Ecol. Environ 10, 202–209 (2012). [Google Scholar]
- 146.Stubbington R. et al. Ecosystem services of temporary streams differ between wet and dry phases in regions with contrasting climates and economies. People Nat. 2, 660–677 (2020). [Google Scholar]
- 147.Leigh C, Boersma KS, Galatowitsch ML, Milner VS & Stubbington R. Are all rivers equal? The role of education in attitudes towards temporary and perennial rivers. People Nat. 1, 181–190 (2019). [Google Scholar]
- 148.Cottet M, Robert A, Tronchère-Cottet H. & Datry T. ‘It’s dry, it has fewer charms!’: do perceptions and values of intermittent rivers interact with their management? Environ. Sci. Policy 139, 139–148 (2023). [Google Scholar]
- 149.Acuña V. et al. Why should we care about temporary waterways? Science 343, 1080–1081 (2014). [DOI] [PubMed] [Google Scholar]
- 150.Doody TM, Hancock PJ & Pritchard JL Information Guidelines Explanatory Note — Assessing Groundwater-Dependent Ecosystems. Report prepared for the Independent Expert Scientific Committee on Coal Seam Gas and Large Coal Mining Development through the Department of the Environment and Energy, Commonwealth of Australia https://www.iesc.gov.au/sites/default/files/2022-07/information-guidelinesexplanatory-note-assessing-groundwater-dependent-ecosystems.pdf (2019).
- 151.Sullivan SMP, Rains MC, Rodewald AD, Buzbee WW & Rosemond AD Distorting science, putting water at risk. Science 369, 766–768 (2020). [DOI] [PubMed] [Google Scholar]
- 152.Reyjol Y. et al. Assessing the ecological status in the context of the European water framework directive: where do we go now? Sci. Total Environ 497–498, 332–344 (2014). [DOI] [PubMed] [Google Scholar]
- 153.Kallis G. & Butler D. The EU water framework directive: measures and implications. Water Policy 3, 125–142 (2001). [Google Scholar]
- 154.Snelder TH et al. Regionalization of patterns of flow intermittence from gauging station records. Hydrol. Earth Syst. Sci 17, 2685–2699 (2013). [Google Scholar]
- 155.Crabot J, Polášek M, Launay B, Pařil P. & Datry T. Drying in newly intermittent rivers leads to higher variability of invertebrate communities. Freshw. Biol 66, 730–744 (2021). [Google Scholar]
- 156.Acuña V, Hunter M. & Ruhí A. Managing temporary streams and rivers as unique rather than second-class ecosystems. Biol. Conserv 211, 12–19 (2017). [Google Scholar]
- 157.Smith REW et al. Assessing and Managing Water Quality in Temporary Waters Technical Report. Australian and New Zealand Guidelines for Fresh and Marine Water Quality https://www.waterquality.gov.au/sites/default/files/documents/assessingand-managing-water-quality-in-temporary-waters.pdf (Australian and New Zealand Governments and Australian State and Territory Governments, 2020).
- 158.Truchy A. et al. Citizen scientists can help advance the science and management of intermittent rivers and ephemeral streams. BioScience 73, 513–521 (2023). [Google Scholar]
- 159.Krabbenhoft CA et al. Assessing placement bias of the global river gauge network. Nat. Sustain 5, 586–592 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Christensen JR et al. Headwater streams and inland wetlands: status and advancements of geospatial datasets and maps across the United States. Earth-Sci. Rev 235, 1–24 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Jaeger KL et al. Probability of Streamflow Permanence Model (PROSPER): a spatially continuous model of annual streamflow permanence throughout the Pacific Northwest. J. Hydrol. X 2, 100005 (2019). [Google Scholar]
- 162.Durighetto N, Vingiani F, Bertassello LE, Camporese M. & Botter G. Intraseasonal drainage network dynamics in a headwater catchment of the Italian Alps. Water Resour. Res 56, e2019WR025563 (2020). [Google Scholar]
- 163.Merritt AM, Lane B. & Hawkins CP Classification and prediction of natural streamflow regimes in arid regions of the USA. Water 13, 380 (2021). [Google Scholar]
- 164.Mahoney DT et al. Dynamics of streamflow permanence in a headwater network: insights from catchment-scale model simulations. J. Hydrol 620, 129422 (2023). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Ward A, Schmadel N. & Wondzell S. Simulation of dynamic expansion, contraction, and connectivity in a mountain stream network. Adv. Water Resour 114, 64–82 (2018). [Google Scholar]
- 166.Hou J, van Dijk A, Renzullo L, Vertessy R. & Mueller N. Hydromorphological attributes for all Australian river reaches derived from Landsat dynamic inundation remote sensing. Earth Syst. Sci. Data 11, 1003–1015 (2019). [Google Scholar]
- 167.Wang Z. & Vivoni ER Detecting streamflow in dryland rivers using CubeSats. Geophys. Res. Lett 49, e2022GL098729 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Stanislawski LV et al. Extensibility of U-Net neural network model for hydrographic feature extraction and implications for hydrologic modeling. Remote. Sens 13, 2368 (2021). [Google Scholar]
- 169.Gao S. et al. Mapping dynamic non-perennial stream networks using high-resolution distributed hydrologic simulation: a case study in the upper blue river basin. J. Hydrol 600, 126522 (2021). [Google Scholar]
- 170.Stubbington R. et al. Disentangling responses to natural stressor and human impact gradients in river ecosystems across Europe. J. Appl. Ecol 59, 537–548 (2022). [Google Scholar]
- 171.Datry T, Arscott DB & Sabater S. Recent perspectives on temporary river ecology. Aquat. Sci 73, 453–457 (2011). [Google Scholar]
- 172.Stubbington R. et al. Biomonitoring of intermittent rivers and ephemeral streams in Europe: current practice and priorities to enhance ecological status assessments. Sci. Total Environ 618, 1096–1113 (2018). [DOI] [PubMed] [Google Scholar]
- 173.Arias-Real R, Gutiérrez-Cánovas C, Menéndez M. & Muñoz I. Drying niches of aquatic macroinvertebrates identify potential biomonitoring indicators in intermittent and ephemeral streams. Ecol. Indic 142, 109263 (2022). [Google Scholar]
- 174.Cid N. et al. A metacommunity approach to improve biological assessments in highly dynamic freshwater ecosystems. BioScience 70, 427–438 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Blackman RC et al. Unlocking our understanding of intermittent rivers and ephemeral streams with genomic tools. Front. Ecol. Environ 19, 574–583 (2021). [Google Scholar]
- 176.Steward AL, Negus P, Marshall JC, Clifford SE & Dent C. Assessing the ecological health of rivers when they are dry. Ecol. Indic 85, 537–547 (2018). [Google Scholar]
- 177.Bunting G. et al. Aquatic and terrestrial invertebrate community responses to drying in chalk streams. Water Environ. J 35, 229–241 (2021). [Google Scholar]
- 178.Boulton AJ Conservation of ephemeral streams and their ecosystem services: what are we missing? Aquat. Conserv. Mar. Freshw. Ecosyst 24, 733–738 (2014). [Google Scholar]
- 179.Lake P, Bond N. & Reich P. in Intermittent Rivers and Ephemeral Streams: Ecology and Management 509–533 (Academic Press, 2017). [Google Scholar]
- 180.Reich P. et al. Aquatic invertebrate responses to riparian restoration and flow extremes in three degraded intermittent streams: an eight-year field experiment. Freshw. Biol 68, 325–339 (2023). [Google Scholar]
- 181.Beebe B, Bentley K, Buehrens T, Perry R. & Armstrong J. Evaluating fish rescue as a drought adaptation strategy using a life cycle modeling approach for imperiled Coho salmon. North. Am. J. Fish. Manag 10.1002/nafm.10532 (2021). [DOI] [Google Scholar]
- 182.Black AN et al. A review of the Leon springs pupfish (Cyprinodon bovinus) long-term conservation strategy and response to habitat restoration. Aquat. Conserv. Mar. Freshw. Ecosyst 26, 410–416 (2016). [Google Scholar]
- 183.Lobegeiger J. Refugial Waterholes Project. Research Highlights (State of Queensland, Department of Environment and Resource Management, 2010). [Google Scholar]
- 184.Department of Environment and Science. The Queensland Waterhole Classification Scheme. (Queensland Wetlands Program, Queensland Government, 2020). [Google Scholar]
- 185.Ball I, Possingham HP & Watts M. in Spatial Conservation Prioritisation: Quantitative Methods and Computational Tools 185–195 (Oxford Univ. Press, 2009). [Google Scholar]
- 186.Bruno D. et al. Ecological relevance of non-perennial rivers for the conservation of terrestrial and aquatic communities. Conserv. Biol 36, e13982 (2022). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Datry T, Larned ST & Scarsbrook MR Responses of hyporheic invertebrate assemblages to large-scale variation in flow permanence and surface–subsurface exchange. Freshw. Biol 52, 1452–1462 (2007). [Google Scholar]
- 188.Stromberg J, Bagstad K, Leenhouts J, Lite S. & Makings E. Effects of stream flow intermittency on riparian vegetation of a semiarid region river (San Pedro River, Arizona). River Res. Appl 21, 925–938 (2005). [Google Scholar]
- 189.Davey AJH & Kelly DJ Fish community responses to drying disturbances in an intermittent stream: a landscape perspective. Freshw. Biol 52, 1719–1733 (2007). [Google Scholar]
- 190.Bourke SA, Shanafield M, Hedley P, Chapman S. & Dogramaci S. A hydrological framework for persistent pools along non-perennial rivers. Hydrol. Earth Syst. Sci 27, 809–836 (2023). [Google Scholar]