

Abstract
Plants undergo a major physiological change as they transition from vegetative growth to reproductive development. This transition is a result of responses to various endogenous and exogenous signals that later integrate to result in flowering. Five genetically defined pathways have been identified that control flowering. The vernalization pathway refers to the acceleration of flowering on exposure to a long period of cold. The photoperiod pathway refers to regulation of flowering in response to day length and quality of light perceived. The gibberellin pathway refers to the requirement of gibberellic acid for normal flowering patterns. The autonomous pathway refers to endogenous regulators that are independent of the photoperiod and gibberellin pathways. Most recently, an endogenous pathway that adds plant age to the control of flowering time has been described. The molecular mechanisms of these pathways have been studied extensively in Arabidopsis thaliana and several other flowering plants.
Keywords: Arabidopsis thaliana, Flowering time, Photoperiod, Vernalization, Gibberellic acid, Pathway integrators
Introduction
Land plants have evolved increasingly complex modes of reproduction. While today’s mosses and ferns still reproduce using motile spores/sperm, nonflowering (gymnosperms; with the notable exceptions of Ginkgo biloba and Cycadaceae) and flowering seed plants (angiosperms) do not. Instead, angiosperms have evolved specialized organs (the flower and components thereof) to further reproduction. The earliest fossil of a flowering plant, Archaefructus liaoningensis, dates back about 125 million years. Nowadays fruits of angiosperm flowers form a major source of the staple diet of people and livestock. Flowers are also appreciated for their aesthetic value, their fragrance and their medicinal properties. The formation of flowers is a prerequisite for successful sexual reproduction and as such, the correct timing of this event has adaptive value, in particular in non-self-fertile species in which flowering has to be synchronized between individuals. Even in self-fertile plants, the induction of flowering is tightly controlled by environmental and endogenous cues such as day length, temperature, and hormonal status. Because of their importance, plants and their flowers have attracted a lot of interest not only from breeders, but scientists as well.
Flowering time research in the pre-molecular biology era
Regulation of flowering time has been studied for more than 100 years [1]. These early studies established that plant flowering is regulated by factors that include day length (photoperiod). Subsequently, plants can be classified as long day (LD) plants that induce flowering when day length exceeds a certain threshold, short day (SD) plants that flower when days are short and nights are long, and day-neutral plants whose flowering is not dependent on the length of the day. This logically led to the question of how and where plants determine photoperiod.
Over the years, several hypotheses were put forward to explain how plants perceive photoperiod [1], but it was not until the 1930s that a more elaborate solution was suggested by Erwin Bünning. As a result of his investigations into “circadian oscillations,” Bünning proposed the existence of a “biological clock” that was entrained by the day-night cycle. Bünning further hypothesized that the 24 h day was divided into two phases, a light sensitive (photophile) and a dark sensitive (scotophile) phase, and that a circadian oscillator regulated the shift from one phase to the other. In this scenario, light behaved as an external signal because its presence during either phase would indicate to the plant if the day was long or short. The Bünning hypothesis was later expanded by Pittendrigh [2] into the “external coincidence” model. In contrast to earlier models, the external coincidence model depends on the presence of light at specific times during the 24-h cycle. Pittendrigh [3] later proposed an alternative mechanism, the “internal coincidence” model, in which two different circadian rhythms were entrained by dusk and dawn. As seasons progressed, one of these rhythms would shift phase relative to the other, resulting in (partial) overlap of the two oscillations, which would trigger downstream events, in this case the induction of flowering.
Around the same time that the works mentioned above established the basic principles that enable plants (or any other organism) to measure day length, others followed up on the question of where in the plant photoperiod is perceived. Knott [4], for example, exposed different parts of plants to light and found that the cue to flower required that the leaves, but not the shoot apex, be exposed to light. This suggested that under inductive photoperiod, plants produce in their leaves a flower-triggering substance that is transported to the shoot apex, an idea that was formalized in the “florigen hypothesis” by the Russian botanist Mikhail Chailakhyan. Subsequent experiments such as grafting leaves from flowering plants onto scions grown under noninductive photoperiod and exposing individual leaves to inductive day length soon confirmed the florigen hypothesis [1, 5, 6]. Interestingly, the velocity and pattern of movement of the florigen was found to match those of photosynthetic assimilates, indicating that the florigen might move through the phloem from the leaf to the apex [7, 8]. While the presence of a florigenic substance was confirmed in many experiments, its nature has been a matter of debate for a very long time.
Apart from day length, the quality of light also plays a significant role in the transition to flowering. Plants grown at high density or under a dense canopy experience a shift in the red to far-red ratio of the incoming light and respond by stem elongation and precocious induction of flowering, a process known as shade avoidance syndrome [9]. Apart from the red/far-red ratio, blue light is also known to regulate the transition to flowering. For example, it has been demonstrated that a day-neutral response can be induced in SD plants upon exposure to high intensities of blue or white lights [10]. While these experiments clearly show that various aspects of light (in particular day length, light quantity and quality) control the floral transition in many plant species, light is by no means the only environmental factor involved.
Other than the various aspects of light, temperature is probably the next most important external cue that affects flowering because plants need a conducive temperature to survive and propagate. In the context of flowering regulation, one can distinguish between the effects of the ambient growth temperature and those of a prolonged period of cold. Gassner [11] was among the first to describe the requirement of long periods of cold for flowering among different species of plants. He found a marked difference in cold requirements between biennials or winter annuals and spring plants or summer annuals. In 1928, the Russian scientist Lysenko coined the name “jarovization” to describe this response of plants. This was later translated into “vernalization.” The now accepted definition of vernalization as “the acquisition or acceleration of the ability to flower by a chilling treatment” was suggested in 1960 by Chourd. In general, summer annuals have a facultative vernalization response while the winter annuals have an obligate vernalization requirement and cannot flower without a prior cold treatment. The normal vernalization parameters range between 1 and 7°C for a period of 1–3 months, depending on the species. Furthermore, breaks of warm temperature were shown to disrupt the effect of vernalization in rye [12]. An interesting aspect of vernalization is that flowering does not necessarily commence immediately after plants experience normal growth temperatures. Instead, an extended period of time can pass before flowering is actually induced. However, once the vernalized state is achieved, it is mitotically stable. This is referred to as the “memory of winter” and is due, as we will discuss below, to the epigenetic silencing of certain vernalization responsive genes. The vernalized state is however not passed on from parent to progeny as silencing of these genes is reset during meiosis.
The role of temperature in plant development has been studied since the 18th century. Being sessile organisms, it is essential for plants to develop a mechanism to identify conducive temperatures for different life processes, including flowering. An early review of the effects of temperature on flowering was supplied by Wang [13]. A more recent review on how plants perceive temperature and differentiate between day-to-day fluctuations at a molecular level has been provided by Samach and Wigge [14].
In 1926, the Japanese scientist Kurosawa noticed that rice seedlings that were infected with the fungus Gibberella fujikuroi grew so tall that they were unable to stand upright. In 1938, gibberellic acid (GA), the chemical that caused this effect on the rice seedlings, was isolated. In 1952, Anton Lang applied GA to rosettes of Samolus parviflorus and Crepis tectorum and noticed that the plants responded by bolting and flowering. Subsequently, GA was often referred to as the flowering hormone. Lang however was able to distinguish between the florigen and GA and concluded that while GA was not the florigen, it somehow regulated the florigen [15, 16]. Conflicting results on the role of GA in flowering were observed in different species. While GA enhanced flowering in some plants, it suppressed flowering in others. Exogenous application of GA resulted in flowering in noninductive photoperiods in certain plants, but not in all cases investigated [17]. GA was also able to bypass the requirement for vernalization [18].
Besides GAs, carbohydrates have also been shown to play an important role in regulating the floral transition [19]. Sugars are produced through photosynthesis and play a vital role in plant development. The major plant sugar is sucrose, which has been shown to accumulate at the shoot apex just prior to transition to flowering. For example, S. alba plants grown in short days accumulated sugars in the apex upon increased irradiation [20].
Despite the success of these early works, it was not until the advent of modern plant genetics and molecular biology, particularly in Arabidopsis thaliana, that the mechanisms underlying the floral transition were better understood.
Flowering time mutants in Arabidopsis thaliana
It was Laibach [21] who proposed A. thaliana as a model plant for genetics. Its small genome size, the ease with which it could be crossed and cultivated, its short life cycle, and the large number of seeds produced made it an ideal model organism. Since then, A. thaliana has become the paradigm for understanding plant genetics and molecular biology, although several other plants species are also widely used for scientific research.
In general, no environmental conditions are known that completely prevent flowering of A. thaliana. Also, no A. thaliana mutants have been reported that, like the veg mutant in peas, fail to flower. However, genetic variation in the response to environmental cues clearly exists among natural accessions of A. thaliana [22–24]. Most accessions that are commonly used in the laboratory are summer annuals that do not require vernalization. However, winter annuals do exist, and genetic analyses have shown that natural alleles of two genes, FLOWERING LOCUS C and FRIGIDA, to a large extent account for the vernalization requirement of these accessions [25, 26]. With respect to photoperiod, flowering time in A. thaliana is dependent on the length of the day, with long days (16 h light) in general promoting floral transition compared to short days (8 h). However, A. thaliana will eventually flower even under SD and has hence been classified as a facultative LD plant.
Rédei [27] used X-ray irradiation to identify co, gi, and ld as loci that are involved in flowering. Later, Koornneef et al. [28] identified 11 loci that resulted in late flowering time when mutated in the Landsberg erecta (Ler) accession of A. thaliana. These loci included fd, fwa, fe, fpa, fy, fve, ft, fha, fca, and two of the loci (gi and co) that Rédei had previously identified. Most of the mutations were recessive, although co was intermediate and fwa was almost completely dominant. Flowering time of these mutants was assayed under different photoperiods and in response to vernalization. fca, fve, fy, and fpa were found to flower late under both SD and LD, but flowering could be accelerated by vernalization treatment. These genes define the core elements of what is now known as the autonomous pathway of flowering in A. thaliana. In contrast, mutations in gi, co, and fha delayed flowering specifically under LD suggesting that these genes are involved in a photoperiod-sensing pathway.
The last couple of years have seen tremendous progress in our understanding of the molecular regulation of flowering time. Numerous genes involved in this process have been identified, and we are beginning to understand how these genes integrate various endogenous and environmental cues to control the onset of flowering. Here we present a comprehensive overview of the current state of research on the different pathways that facilitate flowering and the different factors that regulate the transition from vegetative to reproductive growth.
Environmental control of flowering
As outlined above, flowering time is under the control of diverse environmental stimuli such as temperature and photoperiod. Photoperiod is perceived in the leaves from which the long distance signal dubbed the florigen is transmitted to the shoot apex to induce flowering. In the following sections, we will review the genetic and molecular mechanisms that allow plants to regulate flowering time in response to the environment.
Regulation of flowering by day length
The photoperiod pathway—or how to measure day length?
As one moves away from the equator, the length of the day varies significantly between summers and winters. Plants have developed the ability to sense this distinction and use it as an indicator to control the onset of flowering. The cascade of events responsible for measurement of day length and the subsequent initiation of flowering is referred to as the photoperiod pathway.
Light is perceived by plants at different wavelengths by specialized photoreceptors. Phototropins (blue), cryptochromes (blue), and phytochromes (red/far-red) are the three main classes of plant photoreceptors [29–31]. Several models have been proposed regarding how plants (or organisms in general) might measure day length (see above). Common to all of these hypotheses is that they require internal oscillators, i.e., genes regulated by the circadian clock, and environmental changes such as the day-night cycle to synchronize these rhythms. Interestingly, phytochromes and cryptochromes themselves have been shown to be regulated by the circadian clock, indicating the existence of a regulatory loop that modulates gating and resetting of the circadian clock [32].
Rédei [27] was the first to describe mutants that were insensitive to inductive day length. Among them was the constans (co) mutant. The CONSTANS (CO) gene encodes a putative zinc finger transcription factor [33], the temporal and spatial regulation of which turned out to be key to the photoperiod-dependent induction of flowering (Table 1) [34]. CO expression is under the control of the circadian clock, which causes a basic oscillation of CO expression with a phase of 24 h, and a maximum approximately 20 h after dawn under SD conditions [35]. This phasing of CO expression is further modified under LD by the activity of three other proteins: GIGANTEA (GI), FLAVIN-BINDING, KELCH REPEAT, F-BOX 1 (FKF1), and CYCLING DOF FACTOR1 (CDF1) [36–38].
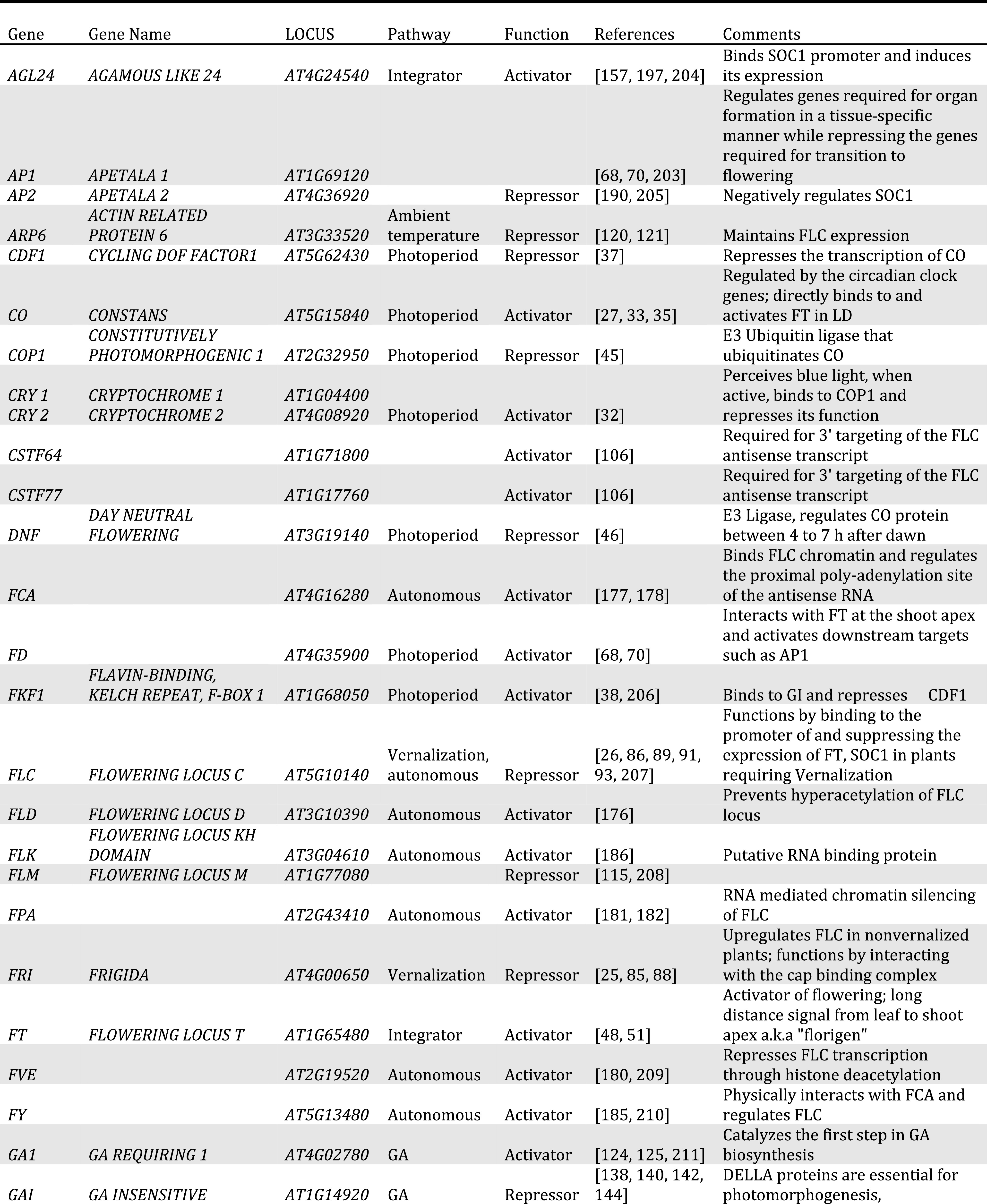
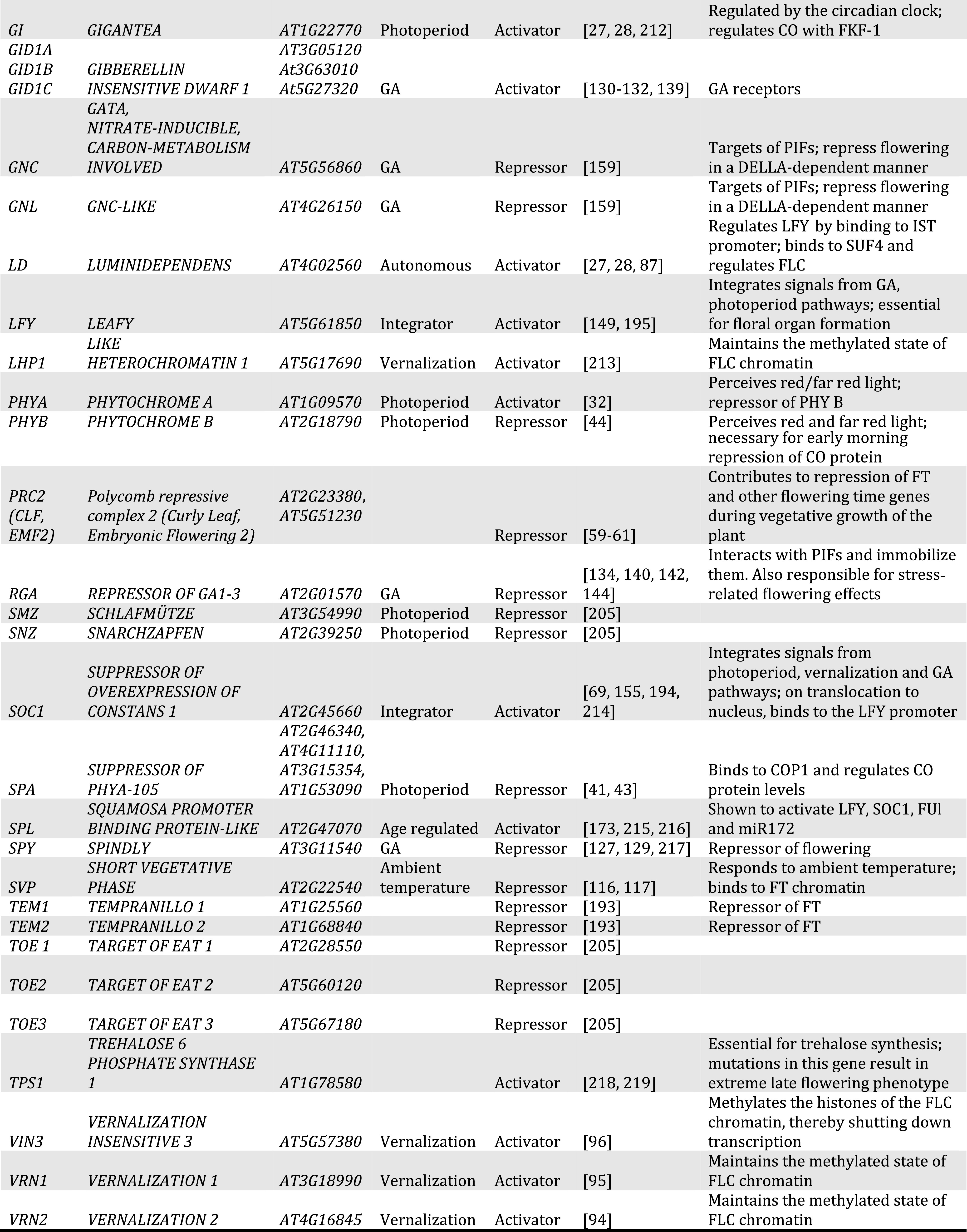
Table 1.
List of important flowering time regulators
Interestingly, these three genes are themselves regulated by the circadian clock. In long days, both the FKF1 and GI proteins follow the same phase with maximum levels being reached 13 h after dawn [38, 39]. In contrast, under SD conditions, GI peaks at 7 h after light onset, but FKF1 peaks 10 h after light onset [36].
Interaction assays in yeast showed that FKF1 physically interacts with GI [38]. Using truncated FKF1 protein constructs, the regions of interaction were narrowed down to the LOV (light, oxygen, or voltage) domain of FKF1 and the N terminus of GI. Interestingly, FKF1 protein binds GI only in the presence of blue light, which it perceives through its flavin-binding domain. As a result of this, FKF1-GI complexes are formed much more efficiently during long days when there is sufficient exposure of the FKF1 protein to blue light and FKF1 and GI proteins peak at the same time, unlike under short days, where the proteins are in different phase and the light, which is required for FKF1-GI complex formation, is lacking [38] (Fig. 1).
Fig. 1.

a, b Regulation of CONSTANS at a transcriptional and protein level. a In short days, FKF1 and GI proteins peak at different times and hence are not able to efficiently repress CDF1, a transcriptional inhibitor of CO. CO protein levels are very low to start with in SD as indicated by the graph. PHYB plays a vital role in maintaining this low level of CO in the early hours of the day. Another protein, DNF, is important for maintaining low levels of CO between 4 and 7 h after dawn. Active CRY protein represses COP1, a ubiquitin ligase that marks CO for degradation. In the dark, the inactive CRY is no longer able to repress COP1 resulting in almost no CO protein being present. b In long days, both FKF1 and GI peak at approximately 13 h after dawn, resulting in active repression of CDF1, and thereby, CO transcription. The protein levels are regulated by PHYB in the early morning hours, while active CRY and PHYA repress PHYB during the rest of the day. Active CRY protein also binds to and inhibits transport of COP1 into the nucleus, hence preventing it from efficiently ubiquitinating the CO protein. Genes are represented in green, and proteins in orange. Dull colors represent inactive genes/proteins, while bold colors indicate active genes/proteins. Dashed box shows weak complex formation, and the grey box shows efficient complex formation. The clock is a 24 h clock. The graph represents expression of CO protein through the day (SD/LD), with the day length represented on the x-axis
FKF1 and GI do not regulate CO expression directly but through interactions of FKF1-GI with CDFs [36, 37]. The CDFs are a family of transcription factors that play an important role in maintenance of CO mRNA levels. The quadruple cdf mutant accumulates CO mRNA both during the day and night and flowers early both in short and long days. CDF1 has been shown to directly bind to the CO regulatory regions and act as a repressor of CO transcription [37]. Chromatin immunoprecipitation (ChIP) using tagged versions of the GI protein also showed enrichment of 17 different amplicons distributed throughout the CO promoter [38]. In addition, ChIP using tagged versions of FKF1 showed that this protein binds to similar regions on the CO promoter as GI and CDF1 [38]. Finally, analysis of the abundance of the three proteins showed that CDF1 peaks first, followed by GI, and then finally FKF1 peaks in the afternoon in long days [37, 38]. Together these studies suggest that CDF1 protein first binds to the CO promoter in the morning. As soon as there is sufficient GI, the CDF1-GI complex is formed that represses CO transcription. Once FKF1 protein peaks, it interacts with the CDF1-GI complex and targets CDF1 for degradation through its F-Box domain to finally activate transcription of the CO gene (Fig. 1) [38]. While CDF1 and CDF2 are both targets of the FKF1-GI ubiquitination pathway, it is unknown whether the other members of this family follow the same mode of degradation [36]. Taken together, the activity of FKF1/GI/CDFs results in a second peak of CO expression towards the end of the subjective LD at approximately 16 h after dawn (Fig. 1).
CO, however, is not only regulated at a transcriptional level, but also at the level of its protein stability and accumulation. Central to the posttranslational regulation of CO are CONSTITUTIVELY PHOTOMORPHOGENIC (COP1) and members of the SUPPRESSOR OF PHYA-105 (SPA) protein family (Fig. 1). COP1 functions as an E3 ubiquitin ligase and has been shown to act downstream of the cryptochrome signalling but upstream of CO. The flowering phenotype of the cop1 co double mutant resembled the co single mutant in both long and short days, placing CO genetically downstream of COP1. Similarly, expression of FLOWERING LOCUS T (FT), a major target of CO (see below), was upregulated in the cop1 single mutants in both short and long days but not in cop1 co double mutants, suggesting that COP1 acts as a negative regulator of CO function, possibly by directing CO for degradation by the 26S proteasome-dependent pathway. This was later shown to be the case, when Liu et al. [40] reported that CO-GST was ubiquitinated specifically by COP1. Furthermore, constitutive overexpression of the CO protein fused to luciferase in cop1 mutants resulted in a drastic increase in luciferase signal when compared to wild type, providing evidence that degradation of CO by COP1 also occurred in vivo. Finally, yeast-2-hybrid analysis and in vitro protein interaction studies also verified that COP1 interacted with CO. The interaction domain was further narrowed down to the WD repeat domain of the COP1 protein. These interactions were also confirmed in vivo by fusing COP1 and CO to the yellow and cyan fluorescence proteins and observing their co-localization in nuclear bodies [40].
Besides COP1, SPA proteins have also been shown to regulate CO [41, 42]. In A. thaliana, the SPA protein family consists of four members that have a WD domain similar to COP1. The spa1 mutant flowered early in short days but was indistinguishable from wild type in long days. The early flowering phenotype of the spa1 mutant was completely suppressed by mutations in co. The other three spa single mutants did not show any difference in flowering in short or long days. The spa1 spa3 spa4 triple mutant, however, flowered earlier than the spa1 single mutant in short days but was only slightly earlier than wild type in long days. This indicates that the SPA3 and SPA4 proteins act redundantly with SPA1 to de-repress flowering specifically in SD conditions [42]. While CO mRNA levels were found to be unaltered in the spa triple mutants, CO protein levels were strongly elevated in the triple mutants when compared to wild type, suggesting that SPA proteins were regulating the CO protein posttranslationally [42]. In agreement with this hypothesis, co-immunoprecipitation studies established that all four SPA proteins indeed interacted with CO through its CCT domain. Further, the SPA1, SPA3, and SPA4 proteins were shown to physically interact with the coiled coil domain of COP1 [41, 43]. These results suggest that SPA proteins enable degradation of the CO protein by the COP1-mediated ubiquitination [42].
Analysis of CO protein accumulation was also performed under different light conditions using CO:GFP fusions. GFP fluorescence was detectable in plants grown under white, blue and far red but not in plants that had been exposed to red or were kept in the dark. This indicated that the accumulation of the CO protein was influenced by a photoreceptor [44]. Subsequently, phyB mutants were shown to exhibit increased levels of CO in the red light and early morning hours, indicating that PHYB plays a major role in regulation of CO in the early hours of the day (Fig. 1) [44, 45].
Another interesting protein that has been shown to repress CO independently of GI/FKF1/CDF is DAY NEUTRAL FLOWERING (DNF) [46]. In dnf mutants, the circadian rhythm of CO is disturbed, resulting in precocious expression of CO and early flowering, in photoperiods as short as 6 h. The molecular mechanism by which DNF regulates CO expression is currently unknown. However, DNF encodes a functional membrane-bound E3 ligase, suggesting that DNF targets a repressor of CO for degradation by the proteasome pathway.
In the end, the complex regulation of CO enables the plant to discriminate SD, where CO protein is not being stably produced, from LD, where CO protein accumulates towards the end of the day. The mechanisms involved turn out to be a mix of both the internal and external coincidence mechanisms originally proposed by Pittendrigh [2, 3]. The former is implemented in the synchronized expression of GI and FKF1, which ensures a boosted CO expression by timed degradation of the CDFs specifically under LD. The latter is enacted in the regulation of FKF1 and COP1/SPA activity through light, which leads to the accumulation of CO protein specifically towards the end of a long day. An important aspect of this is that regulation of CO happens in the leaves and not at the shoot apex where flowers will eventually be formed [34].
The photoperiod pathway—or what good is knowing day length anyway?
For flowering to occur, the information that a plant experiences in the inductive photoperiod needs to be transferred from the leaves to the apex. The question arose as to whether CO itself might constitute a long distance signal (florigen). However, expression of CO mRNA from various tissue-specific promoters suggested that CO regulates production of a systemic flower-promoting signal in the leaves, but does not act as a florigen [34, 47].
Instead, several lines of evidence now indicate that a protein called FLOWERING LOCUS T (FT) is contributing to the floral induction by acting as a long distance signal between leaves and the shoot meristem. FT was simultaneously cloned by two independent groups using an activation tagging approach [48, 49] and a large chromosomal deletion mutant caused by a T-DNA insertion [50, 51]. The FT gene encodes a protein with similarities to Raf kinase inhibitory protein (RKIP) and phosphatidylethanolamine binding protein (PEBP). These proteins are known to inhibit Raf, and thereby result in signal transduction through the MAP kinase pathway. However, since FT lacks certain key residues conserved in all PEBP and RKIP proteins [52], the molecular function of FT is not entirely clear. Analysis of FT expression revealed not only that its expression is much higher in long days, but also that it follows a circadian pattern, peaking in the evening [35, 53]. Promoter GUS constructs showed that the FT gene is transcribed in the phloem companion cells, where CO is also present [54]. Temporal and spatial expression of FT in the vasculature is controlled by a complex orchestration of activating and repressive inputs. The latter include proteins that regulate chromatin structure [55] and thus accessibility of FT locus for transcription factor binding. Several studies have demonstrated that trimethylation of lysine 27 in the amino terminus of histone H3 (H3K27me3) provides an assembly platform for repressive complexes. In this context it is interesting to note that recent genome-wide surveys indicate that all flowering time genes but CO are H3K27me3 targets [56–58]. H3K27 trimethylation is carried out by the polycomb repressive complex 2 (PRC2) and mutants in a number of PRC2 genes [i.e. CURLY LEAF (CLF), EMBRYONIC FLOWER 2, etc.] flower early [59–61]. In these mutants, early flowering was shown to be at least in part due to ectopic expression of FT, suggesting that PRC2 complexes play a major role in repressing FT during vegetative growth. Chromatin-immunoprecipitation experiments revealed that CLF in fact bound FT chromatin, establishing a direct link between PRC2 and FT repression [62]. While PRC2 components can be identified rather easily in plants, proteins homologous to PRC1 are more elusive. However, it has been suggested that LIKE HETEROCHROMATIN PROTEIN1 (LHP1) might act as a PRC1-like corepressor [63]. lhp1 mutants flower somewhat earlier than wild type and, similar to mutants in PRC2 components, this early flowering has been attributed to increased FT expression. Furthermore, LHP1 is directly associated with the FT locus [64], indicating that, like PRC2, LHP1 (PRC1) contributes to FT repression.
FT mRNA is not readily detected in short days, but mRNA levels rise rapidly in the leaves upon transfer from short to long days and are detectable even after exposure to a single long day [37, 65, 66].
Several lines of evidence place FT genetically downstream of CO. In the phloem of SUC2::CO plants, FT mRNA abundance was increased and ft mutations strongly suppressed the early flowering of SUC2::CO [34]. Overexpression of CO in ft-10 plants did not rescue the late flowering phenotype, but FT, when expressed from the SUC2 promoter in co mutants, was able to completely rescue the late flowering phenotype [67]. Further, pFT::GUS was shown to be expressed in a CO-dependent manner [54]. In addition, microarray analysis of plants shifted from short days to long days showed CO-dependent upregulation of FT [68]. Finally, treatment of 35S::CO:GR plants with dexamethasone and cycloheximide resulted in an increase in FT mRNA within 1 h of induction [51, 66, 69]. Corbesier et al. [65] later demonstrated that treating a single leaf from co mutant plants carrying a pCO::GR:CO rescue construct with dexamethasone was sufficient to induce FT mRNA and subsequently flowering. Taken together, these data clearly indicate that FT is a primary target of CO in leaves.
Interestingly, there is strong evidence that FT is not acting in leaves but might promote flowering at the shoot meristem. In particular the finding that FT can interact with the meristem-specific bZIP transcription factor FD immediately suggested that FT might play an important role in conveying the information to initiate flowering from the leaves to the apex [68, 70]. However, it should be noted that formation of the FT-FD protein complex at the shoot meristem has yet to be demonstrated. Several scenarios have been suggested as to how FT might be expressed in leaves but act at the shoot apex to regulate flowering. First, one can conceive an indirect mechanism in which FT triggers expression of an unknown factor X in leaves. X would then move to the shoot apex where it would activate expression of FT, which would be free to interact with FD. Alternatively, FT could directly move either as mRNA or as protein from the leaves to the shoot apex (Fig. 3). Several lines of evidence suggest that the latter is the case in A. thaliana.
Fig. 3.

Integration of flowering time pathways. Light is perceived in the leaves, where it is perceived by photoreceptors such as PHYA, PHYB, CRY1, and CRY2 and regulates expression of genes such as GI, FKF1, and CDF1, all of which have direct or indirect effects on CO expression. CO is a transcriptional activator of FT. miR172 is regulated both by the circadian clock as well as SPLs, which are in turn regulated by miR156. miR172 targets the AP2 family of transcription factors, which play an important role in transcriptional repression of FT in the leaf. The different autonomous pathway genes regulate FLC, a suppressor of FT and SOC1. Another major environmental factor that affects FLC is temperature. FRI activates FLC, while the histone modification proteins VIN3 and VRN1/2 repress it, thereby promoting flowering. Ambient temperatures affect expression of yet another transcriptional repressor of FT, SVP. As the florigen, FT protein moves from the leaf to the apex, where, with the bZIP transcription factor FD, it activates AP1 and SOC1. In the GA pathway, GA regulates levels of the DELLA proteins, which in turn repress miRNA159, a repressor of MYB. MYBs positively control LFY levels in the meristem. Thus the signals from different pathways integrate at LFY, FT, and/or SOC1. SOC1 and AGL24 regulate each other and act together to activate LFY transcription. TFL1 and LFY repress each other. SOC1 activates FUL, which is also a target of the SPL proteins. Activation of SPLs by miR156 forms a novel pathway for regulation of flowering called the aging pathway. SPL proteins upregulate LFY, AP1, FUL, and SOC1. Hence, the different integrators directly or indirectly activate AP1, which marks the beginning of floral organ formation. All genes are represented in green, microRNAs in red, and proteins in orange
For example, an artificial microRNA against FT driven by the 35S and SUC2 promoters delayed flowering, but no change in flowering time was observed when amiRNA-FT was expressed at the apex using the FD promoter. This indicates that FT mRNA was required in the phloem companion cells to induce flowering, but not at the apical meristem [71]. Similarly, Jäger and Wigge [72] could show that trapping FT protein in the phloem companion cells by fusion with a strong nuclear localization signal prevented FT from inducing flowering. Because in this experiment the FT mRNA would be free to move, this finding also indicated that it was the FT protein rather than the mRNA that was acting as a long distance signal in flowering control. In addition, expression of a translational fusion of FT with three molecules of yellow fluorescence protein (YFP) from the SUC2 promoter did not induce flowering. As the FT-3xYFP protein was shown to promote flowering when expressed from a constitutive promoter [71], this also suggested that FT functions by direct movement rather than a relay mechanism. Because in this particular experiment FT had been separated from YFP by a tobacco etch virus (TEV) cleavage sequence, it was possible to release the mature FT protein from the FT-3xYFP precursor by in vivo cleavage using TEV protease expressed from the SUC2 promoter. Release of FT protein resulted in very early flowering, demonstrating that FT protein in the phloem companion cells was sufficient to induce flowering. In agreement with this, Corbesier et al. [65] demonstrated by fluorescence microscopy that a GFP:FT fusion protein was exported from the vasculature to the base of the meristem. Finally, expression of a synthetic FT gene with synonymous mutations in every possible triplet (synFT) was shown to promote flowering just as well as wild type FT [73]. Grafting experiments confirmed that synFT was fully functional. As the primary sequence and the predicted folding of the synFT mRNA are quite different from those of wild type FT, this finding supported the idea that FT mRNA was not acting as a florigen. In agreement with this, synFT-T7 mRNA was not detected in the shoot apical region of ft-1 stock plants grafted on 35S::synFT-T7 scions. In summary, all of these data strongly suggested that FT protein rather than the mRNA is acting as a long distance signal in A. thaliana.
However, more recently Li and colleagues [74] reported that a cis element within the first 102 bases of the FT ORF was sufficient to facilitate movement of a heterologous protein throughout the plant. While movement of nontranslatable FT mRNA alone was not sufficient to induce flowering, indicating that FT protein was needed, these latest results suggest that FT mRNA might contribute to the induction of flowering after all.
The CO-FT module is conserved in other plant species
Although FT has been established as a florigen in A. thaliana, the function of its homologs is less well studied, especially in SD and day-neutral plants. In rice (an SD plant), Heading date 1 (Hd1) and Hd3a have been identified as orthologs of CO and FT, respectively [75, 76]. Both genes were identified by QTL mapping as the key activators of flowering in rice. In addition to Hd3a, rice encodes another close homolog, Rice Flowering Locus T 1 (Rft1). Both genes show diurnal expression, peak at the transition to flowering, and RNAi experiments suggest that both Hd3a and Rft1 are essential for flowering in rice [77].
While the presence of CO and FT orthologs in rice suggest a certain degree of evolutionary conservation of the pathways that control flowering in LD (Arabidopsis) and SD plants (rice), there are also clear differences. For example, in rice, Hd3a was regulated independently of Hd1 by the B-type regulator Early Heading Date 1 (Ehd1) [78]. Induction of another protein, Grain number, plant height, heading date 7 (Ghd7), by photoperiod could repress Ehd1 and establish an acute and ecologically relevant threshold of the day length required for photoperiodic flowering [79]. In contrast to rice, two-component signalling cascades do not appear to play a major role in the regulation of flowering in A. thaliana. Regardless of the details of its regulation, Hd3a is eventually produced in leaves and is thought to travel to the shoot meristem in rice, similar to FT in A. thaliana [80]. Thus, the mode of FT/Hd3a function appears to be quite similar in A. thaliana and rice, demonstrating that the core function of the CO-FT module is conserved in these two only distantly related species.
Homologs of FT were also identified in the SD plant Pharbitis nil [81]. PnFT 1 and PnFT 2 are genes that are closely related to A. thaliana FT. Their regulation was, however, markedly different from what had been observed for FT in A. thaliana. While both PnFT genes were regulated by the circadian clock, their mRNAs accumulated in the night and peaked in the early morning hours of SD reared plants, but these mRNAs were undetectable in plants reared under LD conditions. This suggests that regulation of PnFT1 occurs by a clock output protein that is active during the dark period [81] and as such functions in a different mode from the CO-FT module in the LD plant A. thaliana. Despite these differences, PnFT 1 and PnFT 2 were able to rescue the A. thaliana co mutant phenotype when expressed from the 35S promoter, indicating that these two genes were true orthologs of the A. thaliana FT gene [81]. Flowering in day-neutral tomato plants was shown to be brought about by SFT (SINGLE-FLOWER TRUSS), an ortholog of FT. In tomato, sft mutant plants are late-flowering, and grafting experiments showed that SFT was able to complement all the defects of the sft mutant [82]. In addition, 35S::SFT was able to induce flowering in the SD flowering Maryland Mammoth tobacco plants under LD conditions [82, 83], which indicated that SFT could act as a flower-promoting factor in a different species.
These results, obtained from diverse species, suggest that the basic mode of action of the CO-FT module is conserved. However, certain variations of a common theme have evolved in plants that utilize different strategies for photoperiod-regulated flowering.
Regulation of flowering by temperature
Effects of vernalization on flowering
Besides light and photoperiod, temperature is a major determinant of flowering time. Temperature affects flowering in two ways: first, many plants require a prolonged period of cold (vernalization) to induce flowering the following spring, and second, the ambient temperatures a plant experiences throughout its vegetative growth have a marked effect on the timing of flowering; these mechanisms explain the wide range of flowering time responses in natural accessions of A. thaliana [84]. Some are rapid cyclers and flower early, while most late flowering accessions follow a winter-annual life style and require vernalization before they can flower.
Analyses of the genetic differences between rapid cycling and winter-annual varieties of A. thaliana revealed that the dominant locus FRIGIDA (FRI) played a major role in conferring a vernalization requirement to natural accessions of A. thaliana [85]. FRI function is compromised in many rapid-cycling accessions, and FRI is the major determinant of this life history variation [25]. The FRI locus of the winter annuals can be considered the ancestral state. This locus was evolutionarily modified by several rounds of deletions and mutations, which resulted in the summer-annual loss of function phenotype [25]. Further studies revealed that another gene, FLOWERING LOCUS C (FLC), and FRI are both required for vernalization to occur [26, 86, 87]. FRI functions by upregulating the expression of FLC, which is a potent floral repressor [88]. The mechanism by which FRI regulates expression of FLC is still not fully understood, although it was recently shown that FRI protein interacts with the cap binding complex (CBC) through its two coiled coil domains, and that this interaction is essential for FRI function [88].
FLC’s mode of action is better characterized than that of FRI. FLC encodes a MADS box protein that acts to directly repress certain flowering time genes [86, 89, 90]. FLC was shown to block the transcriptional activation of SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1) and FT by directly interacting with CArG boxes on their respective chromatin [91, 92]. This binding reduced the effect of photoperiodic activation of these genes. Since the FLC null allele was able to completely suppress the late-flowering phenotype of FRI, it was concluded that FRI mediates vernalization via FLC [93]. Together, FRI and FLC are responsible for the winter-annual life history; loss of either of the two genes usually results in early flowering and loss of the vernalization requirement. Interestingly, loss of FRI and/or FLC have occurred multiple times, indicating that summer-annual life histories have evolved independently in different accessions of A. thaliana [84].
High levels of FLC expression appear to be responsible for the winter-annual behavior of FRI/FLC positive accessions. To better understand how expression of FLC is regulated in response to vernalization, a genetic screen was performed to identify plants that flowered late even after exposure to long periods of cold. Two important regulators of FLC, VERNALIZATION1 (VRN1) and VERNALIZATION2 (VRN2), were identified from the screen [94, 95]. These studies demonstrated that FLC is epigenetically silenced in response to vernalization. Interestingly, initial silencing of FLC was completely normal in both the vrn1 and vrn2 mutants, but FLC levels increased after plants were returned to higher temperatures, indicating that these genes are required for maintenance, rather than initiation, of the FLC silencing. VRN1 and VRN2 were found to encode a DNA binding protein and a nuclear localized zinc finger protein with similarity to the polycomb group protein Su(Z)12 from Drosophila; this further supported the hypothesis that VRN1 and VRN2 maintaining the epigenetic repression of FLC.
Sung and Amasino [96] identified yet another protein, VERNALIZATION INSENSITIVE 3 (VIN3), that is essential for FLC regulation and that helped to explain the basic principle of FLC silencing. In contrast to vrn1 and vrn2, which fail to maintain FLC silencing, repression of FLC does not occur in vin3 plants. This indicates that VIN3 is required for the initial repression of FLC during cold exposure (Fig. 2). VIN3 encodes a PHD finger protein and is transiently induced by cold temperatures [96, 97]. Lesions in VRN2 locus were shown to affect the structure of FLC chromatin [94], indicating that VRN2 may play a role in FLC chromatin remodelling during silencing. Additionally, VIN3 was shown to interact with members of the PRC2 [98, 99], which is responsible for trimethylation of lysine 27 of histone H3 (H3K27me3), a typical sign of gene silencing [60, 100]. This particular methylation mark increases at the transcription start site of FLC in response to vernalization [98, 101]. This results in the recruitment of VRN1, VRN2, and LHP1, which together maintain the repressed state of FLC. Thus VIN3 is induced in response to vernalization and establishes the initial silencing of FLC. VRN1 and VRN2 are then required to maintain FLC in a silenced state (Fig. 2).
Fig. 2.

Regulation of FLC. In plants requiring vernalization, FLC chromatin is acetylated in a nonvernalized state, resulting in active transcription. The first step to negate the effects of FLC is the transcriptional repression of its RNA by COOLAIR, the antisense transcript of FLC during early exposure to cold. Another noncoding RNA called COLDAIR is transcribed from the first intron of FLC and also plays a major role in downregulating FLC transcript levels. Upon initiation of vernalization (late cold), VIN3 methylates lysine residues of histone H3. This vernalized state is maintained by VRN1 and VRN2 upon vernalization, even after the temperatures become warmer. The autonomous pathway regulators FLD and FVE also function by controlling methylation of lysine residues of histone H3. The RNA binding elements Cst64 and Cst77 and the autonomous pathway regulators FPA, FCA, and FY all regulate FLC transcript levels. Levels of FLC RNA (black) are plotted against different stages of cold and compared to levels of COOLAIR RNA (red), COLDAIR RNA (green), and VIN3 protein (orange)
In addition to PRC proteins, noncoding RNAs are emerging as a new family of regulators of gene expression. The production of antisense FLC RNA called COOLAIR RNA (cold induced long antisense intragenic RNA) was recently shown to be the first response to cold treatment. The transcription of COOLAIR RNA was able to repress sense strand transcription before VIN3 exhibited any effects [102]. More recently, Heo and Sung [103] have identified another noncoding RNA from the sense strand of the first intron of FLC that is distinct from COOLAIR and has been named COLD ASSISTED INTRONIC NON CODING RNA (COLDAIR). COLDAIR is temporally correlated with flowering time; its transcript levels were shown to increase within the first 10 days of vernalization. COLDAIR is also mechanistically associated with the flowering time pathway. FLC contains a cryptic COLDAIR promoter, which is activated when FLC is repressed. COLDAIR was further shown to be necessary for recruitment of CLF to FLC. Although CLF is a component of the PRC2 complex, COLDAIR’s role in maintaining PRC2 association with FLC after vernalization is unclear [103]. Despite this final caveat, noncoding RNAs clearly play an important role in the regulation of FLC expression. A detailed review of noncoding RNAs and their function in chromatin regulation has recently been published [104].
RNA binding proteins and epigenetic regulators have also been shown to play important roles in FLC RNA regulation [105]. For example, the FLC repressor FCA was suppressed by mutants that mapped to the loci CstF64 and CftF77 [106]. CstF64 and CstF77 are two 3′ RNA processing factors that were shown to be essential for 3′ targeting of the antisense transcripts of FLC [106].
Despite diverse efforts, clear orthologs of FLC have been identified only within the Brassicaceae [107–109]. Within this family, however, FLC orthologs such as Arabis alpina PERPETUAL FLOWERING 1 (PEP1) have acquired additional functions. PEP1 appears to be important not only for the induction of flowering in response to vernalization but also for the establishment of a perennial life history [110]. Similar to FLC in A. thaliana, PEP1 is initially repressed in response to vernalization, and flowering commences. However, only the main shoot and those axillary shoots that had developed before vernalization become reproductive by the end of the vernalization treatment. In contrast, PEP1 was active in axillary shoots that were initiated during or after the vernalization and remained vegetative. Chromatin methylation studies showed that H3K27Me3 increased upon vernalization but was not maintained at the PEP1 locus when A. alpina plants were returned to warm temperatures, which allowed for perennial growth of A. alpina [110].
Although vernalization has been studied in a variety of species, no clear orthologs of FLC have been identified outside the Brassicaceae, suggesting that vernalization in diverse taxa is the result of evolutionary convergence. In wheat, for example, vernalization was found to regulate the expression of ZCCT1 (and ZCCT2), a protein that harbors a CCT domain related the one found in CO-like proteins [111]. Repression of ZCCT1 in response to vernalization was gradual and stable suggesting that, even though it belongs to a different family of transcription factors, it might have a function analogous to FLC in A. thaliana.
Regulation of flowering by ambient temperature
Flowering is also affected by the ambient temperatures a plant experiences throughout its vegetative development [14]. The flowering response to ambient temperatures is diverse among species, and this diversity extends to different accessions of A. thaliana. Higher temperatures promote flowering. This was demonstrated in A. thaliana by growing natural accessions under SD conditions at elevated ambient temperatures (25 or 27°C) [112]. Under these conditions, many accessions flowered as early under SD as they normally would under 23°C LD. Thus, in many accessions, higher temperatures can serve as a substitute inductive LD. Several flowering time mutants show temperature dependence. phyB mutants were shown to flower earlier at 23°C but not at 16°C [113]. Similarly, cry2 mutants show an exaggerated delay in flowering at 16°C compared to 23°C [114]. Also, accessions with nonfunctional fri and flc alleles responded strongly to higher temperatures and flowered much earlier at 27°C than at 23°C. In contrast, FRI/FLC accessions showed a much weaker response to elevated temperatures, indicating that FLC plays a role in suppressing thermal induction [112]. Further analysis revealed the existence of natural accessions that were unresponsive to thermal induction despite having nonfunctional fri/flc alleles. In the case of Nd-1, the causal gene could be mapped to a deletion at the FLOWERING LOCUS M (FLM) locus [115]. FLM is a MADS-box protein that shows strong sequence similarities with FLC. Interestingly, the effect of the mutation in the Nd-1 accession was apparent in 23°C but was masked at 27°C. This masking persisted with or without a functional FLM allele at 27°C SD, suggesting that FLM also participates in the temperature pathway [112]. Microarray analyses revealed that genes associated with alternative splice site selection were specifically affected by thermal induction. Temperature-dependent alternative splicing of FLM [112] also suggests that splicing is an important regulator of flowering.
Another major regulator of flowering in response to ambient temperatures is SHORT VEGETATIVE PHASE (SVP). SVP, a MADS box protein, binds to the CArG motifs on the FT and SOC1 promoters and acts as a floral repressor (Fig. 3) [116, 117]. SVP acts downstream of the autonomous pathway mutants FCA and FVE, which are known to play a role in ambient temperature sensing in A. thaliana [114]. Genetic interactions between SVP and FLC indicated that SVP did not regulate FLC. The proteins, however, were shown to co-immunoprecipitate indicating that they may act in concert. Western blotting and co-immunoprecipitation studies performed on 5-day-old seedlings revealed that SVP and FLC are mutually dependent and exhibit similar temporal and spatial expression. ChIP analysis of FLC and SVP showed common binding sites in both the flowering integrators FT and SOC1. SVP may therefore regulate these genes in an FLC-dependent manner [118].
While genetic and molecular analyses have identified several genes that are involved in regulating flowering in response to ambient temperature, the mechanism by which plants detect differences in temperature remains unknown. Only recently, microarray analyses of plants induced to flower by temperature and photoperiod showed expression of HEAT SHOCK PROTEIN 70 (HSP70) to be highly correlated with gradual increases in temperature [112, 119]. Based on this finding, a genetic screen was designed to identify factors involved in temperature perception. HSP70::LUC plants were mutagenized and screened for increased LUC expression when plants were shifted from 12 to 27°C for 3 h [119]. This resulted in the identification of the ACTIN RELATED PROTEIN 6 (ARP6) as a component in temperature-mediated flowering [119]. ARP6 is a nuclear protein that acts to repress flowering by maintaining FLC expression [120, 121]. arp6 mutants phenocopied warm-grown plants and show a constitutive warm temperature response. ARP6 is part of the SWR1 chromatin remodelling complex and functions by introducing histone H2A.Z rather than H2A into nucleosomes. H2A.Z nucleosomes appear to wrap DNA more tightly than their H2A counterparts. The tight wrapping of DNA by H2A.Z nucleosomes can be overcome by higher temperatures, thereby providing a possible mechanism for temperature-dependent gene regulation [119].
Plant-derived flowering time regulators
Gibberellic acid pathway: a hormonal control of flowering
In 1935, Teijiro Yabuta observed that rice seedlings infected with the fungus G. fujikuroi grew so quickly that they tipped over. It was later discovered that gibberellins (gibberellic acids or GAs) produced by the fungus were regulating growth in the host plants. Their role in normal plant development was, however, studied only much later. Since then, numerous GAs have been isolated from plants, but not all of them are biologically active. The active GAs include GA1, GA3, GA4, and GA7 (numbered based on the order of their discovery [122]). A large-scale X-ray and EMS mutagenesis screen was performed in A. thaliana by Koornneef and Van der Veen [123] to identify mutants with an impaired GA pathway. They isolated 37 mutants that showed poor germination and improper floral organ formation, but no striking flowering time phenotype was observed under LD conditions. These mutants could be completely rescued by the exogenous application of GA, indicating that they were most likely affected in GA biosynthesis. The first committed step of GA biosynthesis requires the GA1 gene, which encodes an ent-kaurene synthase [124]. ga1 loss of function mutants of A. thaliana displayed a mild flowering time phenotype compared to mutants affected in the photoperiod pathway [124–126]. Interestingly, flowering of ga1–3 loss of function mutants was almost normal under LD, but they never flowered in SD unless supplemented with exogenous GA. These results were interpreted as evidence that GA was required for floral transition in SD, but not in LD. Another genetic screen for mutants that were resistant to the inhibitor of GA biosynthesis, paclobutrazol [127], identified SPINDLY (SPY) as a negative regulator of GA signalling. SPY encodes an O-linked N-acetylglucosamine transferase. Recent work in rice indicates that SPY regulates the GA pathway by regulating the DELLA proteins (see below) [128]. ga1 spy4 double mutants and wild type plants flowered similarly, indicating that the spy4 mutation was able to overcome the late flowering phenotype of ga1 [129].
Bioactive GAs are perceived by plants through a cytoplasmic/nuclear localized receptor, GIBBERELLIC INSENSITIVE DWARF 1 (GID), which was originally identified in rice [130]. A. thaliana has three functionally redundant copies of the GID1 receptor [131, 132]. Interestingly, the A. thaliana gid1 triple mutant was described to be either moderately late flowering [131] or extremely late flowering (or not flowering at all) [132], even in LD. These findings indicated that contrary to previous results [126], GA signalling contributed to promoting flowering under LD and that its role in regulating flowering time in A. thaliana was not limited to SD. The finding that the ga1–3 mutant accumulates detectable levels of bioactive GAs [133, 134] provides a simple explanation for the observed difference in the severity of phenotypes between ga1–3 and the gid1 triple mutant.
To further define the role of GAs in LD-induced flowering, the ga1–3 mutation was introduced into the co mutant background by genetic crosses. The ga1–3 co double mutant flowered later than both parents, indicating that deficiency in GA biosynthesis has an additive effect on the late-flowering phenotype of co mutants in LD [33]. The relationship between FT and GA was studied by measuring the levels of FT mRNA in ga1–3 mutants upon a shift from SD to LD. It was observed that FT mRNA levels increased 15-fold upon application of GA, indicating that ga1–3 plants required GA in addition to an inductive photoperiod to produce FT mRNA [135]. Finally, application of the GA-biosynthesis inhibitor paclobutrazol to wild type plants resulted in delayed flowering in LD. Supplementing the inhibitor with exogenous GA completely restored proper flowering [135]. Taken together, these results indicate that GAs regulate the expression of FT and function in parallel with CO in LD to promote flowering.
GID1 regulates GA signal transduction through interaction with members of the DELLA protein family. The DELLA proteins are named after a conserved protein motif starting with the amino acids D, E, L, L, and A. They belong to the GRAS family of transcriptional regulators that function as repressors of plant growth and development [136, 137]. An important role for this protein family was suggested by the finding that a deletion in the DELLA domain resulted in a semi-dwarf phenotype that resembled the GA-deficient mutant that could not be rescued by GA supplementation [138]. It was shown recently that the GA-bound form of GID1 induced a conformational change upon binding to the N-terminal region of DELLA proteins [139]. Because the conformational change promotes ubiquitination by an E3 ubiquitin ligase, the DELLA proteins become susceptible to degradation via the 26S proteasome pathway. DELLA proteins have been shown to be important integrators of GA signalling and play a significant role in many aspects of plant development, in particular photomorphogenesis [140–142]. DELLA proteins have been shown to immobilize the PHYTOCHROME INTERACTING FACTOR (PIF) proteins by directly interacting with them [143, 144]. Interestingly, Oda et al. [145] showed that suppression of PIF3 by antisense RNA induced CO and FT, resulting in early flowering in LD. Since PIFs are regulated by light and GA via the DELLA proteins, they represent a point of convergence of light and GA pathways.
Other important points of convergence between the GA, the photoperiod and the vernalization pathways are the floral integrators LEAFY (LFY) and SOC1 (Fig. 3). Application of GA was shown to increase LFY promoter activity in SD [146]. In SD, LFY transcription is absent in ga1–3 plants, as evidenced by a β-glucuronidase reporter fused to the LFY promoter (LFY::GUS). A more sensitive analysis demonstrated that LFY mRNA was reduced 10-fold in the ga1–3 mutant plants when compared to wild type, indicating that both the endogenous LFY promoter and the LFY::GUS construct were less active in the ga1–3 background. In addition, analysis of the GUS activity in the spy mutant indicated an increase in LFY promoter activity especially in SD [147]. These findings lead to the conclusion that GA regulates the LFY promoter and that at least part of the flower-stimulating activity of GAs is due to an activation of LFY expression by GAs [147]. Different GAs were tested for an effect on the LFY promoter. GA4 followed by GA3 showed maximum effect in activation of the LFY promoter. GA4 was found to be the predominant bioactive GA in A. thaliana, and GA4 levels strongly increased at the shoot apex during transition to flowering [148]. Taken together these results indicate that LFY constitutes an important point of integration of signals from the photoperiod and GA pathways.
A more detailed analysis of the LFY promoter identified a cis regulatory sequence that was required for LFY expression in response to GA treatment. This regulatory sequence conforms to the consensus binding site for MYB transcription factors [149]. Interestingly, MYB transcription factors have previously been implicated in GA signalling in other plant species. In particular, GAMYBs, a family of R2R3 type MYB transcription factors, have been shown to play an important role during germination in cereals. In A. thaliana, AtMYB33, a potential homolog of GAMYBs, was found to be expressed in the shoot apex as a response to endogenous GAs or application of exogenous GAs [150]. In addition, MYB33 protein was shown by EMSA studies to bind to the predicted GA-responsive element in the LFY promoter. Analysis of the Lolium temulentum homolog of GAMYB, LtGAMYB, showed that the protein is expressed in the shoot apex during floral transition. Furthermore, its levels increased in synchrony with GAs at the apex, indicating that GAs may regulate the floral transition in L. temulentum via LtGAMYB [133, 151]. Interestingly, MYB33 and its closest paralogs, MYB65 and MYB101, are predicted targets of the microRNA159 (miR159). Regulation of MYB33, MYB65, and MYB101 by miR159 has recently been shown to play a major role in regulating the spatial expression of these genes [152, 153]. miR159 was also shown to be downregulated by the DELLA proteins, indicating that GA mediates flowering in response to miR159 by repressing DELLA proteins [154].
Another critical gene for promoting flowering in response to GA signalling is SOC1. Moon et al. [155] demonstrated that SOC1 expression is nearly undetectable in ga1–3 mutants in SD. The exogenous application of GA resulted in an increase in SOC1 expression indicating that GA plays an important role in the regulation of SOC1. These authors further demonstrated that overexpression of SOC1 was able to overcome the flowering defects of ga1–3 in SD. Furthermore, Achard et al. [156] showed that the plant stress hormone ethylene delayed flowering by repressing LFY and SOC1 in a DELLA-dependent signalling pathway.
The exogenous application of GA also resulted in an increased transcript level for AGAMOUS LIKE 24 (AGL24) [157]. This response was shown to be SOC1 dependent, as the soc1–2 mutation completely prevented the upregulation of AGL24 levels upon GA application [158]. Additionally, AGL24 was shown to directly bind the SOC1 promoter. Upon treatment of wild type plants with GA, both SOC1 and AGL24 transcript levels increased compared to untreated plants. The soc1–2 agl24-1 double mutants did not flower in short days without the application of GA. Taken together, SOC1 and AGL24 may regulate each other in a GA-dependent manner to regulate flowering especially in SD [158]. Finally, SVP, a repressor of flowering and a negative regulator of SOC1, was also shown to be regulated by GAs. SVP levels decreased in GA-treated wild type plants, while ga1–3 mutants showed consistently higher levels of SVP than their wild type counterparts. It can be concluded that GA regulates SOC1 expression at several levels by promoting expression of SOC1-inducing genes (such as AGL24) and at the same time downregulating floral repressors such as SVP [118].
More recently, two GATA-like transcription factors, GNC (GATA, NITRATE-INDUCIBLE, CARBON-METABOLISM INVOLVED) and GNL (GNC-LIKE), have been shown to participate in GA signalling, and gnc and gnl single and double mutants flowered earlier than wild type in LD. These two genes were shown by ChIP to be direct targets of the PIF transcription factors and thus are regulated by GA in a DELLA-dependent manner. gnc and gnl were able to partially suppress the flowering defects of the ga1 mutants indicating that GNC and GNL function to repress flowering in a GA-dependent manner [159].
Other endogenous factors promoting flowering
In addition to GA, several other plant-derived signals have been shown to affect flowering. Sugars, which are the major products of photosynthesis, are essential for regulation of several metabolic and developmental processes such as germination, flowering, senescence, and stress response. Sucrose in particular has been shown to promote flowering in various plant species. For example, the accumulation of sucrose at the apex during the transition to flowering was noticed in pineapple [160] and Rudbeckia [161]. In some species such as S. alba, apical exudates actually showed a diurnal fluctuation in sucrose levels in displaced SD and LD [162]. However, sucrose levels did not increase during floral transition in all the species investigated. For example, sucrose levels did not change at the shoot apex during the transition to flowering in L. temulentum, suggesting that the contribution of sucrose to flowering might be species-specific [163].
In A. thaliana, Ohto et al. [164] observed that very high concentrations of sucrose had a negative impact on flowering time. A marked increase in the number of rosette and cauline leaves was observed when plants were grown on 5% sucrose. Results were similar for different hexoses such as glucose and fructose. A delay of flowering at high concentrations of sucrose was also observed in late flowering mutants such as ld, co, fca, gi, and fha [164]. However, at lower concentrations (1%), sucrose had the opposite effect and induced early flowering in these mutants [165]. Sucrose at a concentration of 1% also accelerated flowering for some late-flowering accessions [165], indicating that the regulation of flowering is strongly dependent on sucrose homeostasis but is not mediated through a specific flowering time pathway. In a QTL study performed on a recombinant inbred line population derived from a cross between Landsberg erecta and Kondara accessions, it was later found that flowering-time QTLs colocalized with carbohydrate and starch QTLs at the bottom of chromosomes 1, 2, and 3 [166]. In addition, mutations that affect sugar and starch accumulation in leaves and at the shoot apex often also cause changes in the timing of the floral transition. In order to assess whether the accumulation of sucrose at the apex is due to starch mobilization from the leaf during the floral transition, the starchless phosphoglucomutase (pgm) and the starch excess 1 (sex1) mutant were analyzed [167, 168]. Corbesier et al. [169] showed that starch mobilization was essential for the increase of sucrose at the apex. Upon induction of flowering by exposure to LD, there was an increase in the carbohydrates exported from the leaf [169].
Another sugar that is shown to have a marked effect on the transition to flowering is trehalose [170]. Trehalose is synthesized from UDP-glucose and glucose-6-phosphate via an intermediate, trehalose-6-phosphate (T6P). In most plants, trehalose can only be found in micromolar concentrations and its function is not entirely clear. However, T6P appears to be essential for normal plant development because loss of function mutations in the TREHALOSE-6-PHOSPHATE SYNTHASE1 (TPS1) gene have been shown to be embryo lethal [171]. Homozygous tps1 individuals could, however, be obtained by expressing TPS1 under the control of a chemically inducible promoter (GVG::TPS1) during embryogenesis [172]. These homozygous tps1-2 GVG::TPS1 plants grew more slowly than wild type, but the most obvious phenotype was an extreme delay in flowering. Thus, T6P is important for the regulation of flowering.
How the information about the carbohydrate status is integrated into flowering time regulation is poorly understood. However, it was recently shown that the levels of a microRNA that is known to regulate flowering, miR156, decreased with increasing age of the plant. Interestingly, regulation of flowering by miR156 appears to define a novel flowering time pathway that acts independently of the photoperiod, vernalization, and GA pathways [173]. Since the aging process affects both miR156 abundance and carbohydrate metabolism, it is tempting to speculate that miR156 and its targets, the SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) genes, function as a read-out of carbohydrate status.
Autonomous pathway of flowering regulation
Autonomous pathway mutants are characterized by delayed flowering irrespective of day length. The autonomous pathway genes include LUMINIDEPENDENS (LD), FCA, FY, FPA, FLOWERING LOCUS D (FLD), FVE, FLK, and REF6 [105, 174]. Koornneef et al. [175] created 42 crosses between different late flowering mutants. All genes in the autonomous pathway act by repressing FLC expression, and the late flowering observed in the mutants can largely be explained by elevated FLC levels. Thus, genes in the autonomous pathway normally promote flowering indirectly by repressing the floral repressor FLC (Fig. 2). As the common lab strains Col-0 and Ler have mutations in the FLC gene that lead to a low level of FLC transcripts, overexpression of upstream autonomous pathway genes usually has no or only mild effects on flowering [175].
The proteins encoded by the genes in the autonomous pathway generally fall into two broad functional categories: general chromatin remodelling and maintenance factors and proteins that affect RNA processing. One example of the former is the FLD protein, which shares similarities with proteins found in mammalian histone deacetylase complexes. FLD was shown to regulate FLC by preventing hyperacetylation of the locus, thereby acting as a repressor of FLC transcription [176]. fld-3 mutants showed hyperacetylation of histone H4 and a twofold increase in histone H3K4 dimethylation [176, 177]. Genetic analyses indicated that FLD and FCA function in the same genetic pathway, with FCA being epistatic to FLD with respect to flowering time [177]. FCA contains two RNA recognition motifs (RRM) and a WW domain suggesting a role in posttranscriptional RNA modifications. FCA mRNA was shown to be subject to alternative splicing, and different transcripts were expressed at different levels in different tissues [178]. Interestingly, alternative splicing of FCA seems to require functional FCA. The WW domain was shown to be important for FCA autoregulation [179]. FCA was shown to be associated with the FLC coding region at exon 6 and intron 6 where it regulates the proximal polyadenylation site of the antisense RNA [177]. Another FLC repressor, FVE, is a homolog of the mammalian putative retinoblastoma-associated proteins RbAp46 and RbAp48 and acts through participation in a histone deacetylation complex [180] (Fig. 2). Interestingly, the loss of FCA function was found to be additive with mutations in FVE.
The FPA gene encodes a protein with three RRM. FPA is expressed throughout the plant’s life, in particular in newly formed tissues and meristems [181]. FPA and FCA were shown to act in a partially redundant fashion to control RNA-mediated chromatin silencing of FLC [182, 183]. Apart from its participation in chromatin silencing, FPA has also been implicated in alternative cleavage and polyadenylation of RNAs [183].
In contrast to FPA, FY, an RNA 3′ end-processing factor, has been shown to directly interact with FCA [184]. Interaction is facilitated through the WW domain of FCA and two proline-rich (PPLPP) motifs in the C-terminus of FY. FCA/FY interaction is not only required for downregulation of FLC, but apparently also plays an important role in the autoregulation of FCA expression [184]. FCA/FY interaction is required for the selection of the promoter-proximal polyadenylation site selection in the FCA pre-mRNA. There exists some natural variation at FY, and a mutation in the second PPLPP motif of FY in Bla-6 was recently shown to contribute to the relative insensitivity of this accession to the flower-promoting effects of a reduced red light to far-red light (R/FR) ratio [185].
FLK also encodes a putative RNA binding protein. As is common for autonomous pathway mutants, flk flowered late under both LD and SD [186]. The delayed flowering of flk was most likely caused by activation of FLC expression, which in turn resulted in the downregulation of FT and SOC1. The late-flowering phenotype could be suppressed by vernalization and application of exogenous gibberellins [186].
Finally, LD was identified in several genetic screens for late flowering mutants [27, 28]. The gene was eventually cloned by Lee et al. [87] and was shown to encode a protein with similarities to transcriptional regulators and to contain two bipartite nuclear localization domains and a glutamate-rich region. The late flowering phenotype of ld mutants was completely suppressed by vernalization. The LD protein was found to localize to the nucleus and regulate the LFY promoter [187]. The LD protein also binds to SUPPRESSOR OF FRIGIDA 4 (SUF4) preventing its action on the FLC locus [188].
Given that the proteins in the autonomous pathway function by regulating chromatin modification and/or RNA processing, it is not surprising that mutants in some of these genes are not only late flowering but display additional phenotypes. For example, Bäurle et al. [182] showed that the fca fpa double mutant was defective in female gametophytic development and early embryonic development.
Integrators of flowering: the crosstalk between pathways
The induction of flowering is a central event in the life cycle of plants. When timed correctly, it helps ensure reproductive success and therefore has adaptive value. Because of its importance, flowering is under the control of a complex genetic circuitry that integrates environmental and endogenous signals, such as photoperiod, temperature, and hormonal status. Genetic and molecular analyses over the last years have identified numerous genes that participate in the regulation of flowering. However, it has also become clear that the genetically defined pathways that regulate flowering are not strictly separated. Instead there is increasing evidence for extensive crosstalk between the pathways. In the end, however, the various signalling pathways regulate the expression of a relatively small number of common targets, which have been referred to as central floral pathway integrators or “integrator genes” [189].
One such integrator gene is FT. At the most basal level, expression of FT in leaves is controlled by a number of proteins involved in chromatin remodelling. Next are a number of transcription factors that regulate FT expression in a more gene-specific manner. The role of CO as a positive regulator of FT has been discussed above. However, CO is by no means the only factor to regulate FT expression. In particular, several repressors of FT have lately been identified. These include two AP2-domain-containing transcription factors of the RAV family [TEMPRANILLO1 (TEM1), TEM2] and six genes of the euAP2 linage [APETALA2, SCHLAFMÜTZE (SMZ), SNARCHZAPFEN, TARGET OF EAT1 (TOE1), TOE2, TOE3] [190] that are targets of miR172 [191, 192]. Overexpression of any of these genes delayed flowering, indicating that the encoded proteins function as floral repressors. In addition, binding of TEM1 and SMZ to regulatory regions of FT has been demonstrated by ChIP, indicating that these factors directly repress FT [191, 193]. Besides these AP2-like transcription factors, FT is also a direct target of FLC [89, 91].
As outlined above, FLC is repressed in response to vernalization, indicating cross-talk between the photoperiod and vernalization pathways (Fig. 3) [89, 91]. The FLC transcript can be detected in the hypocotyl and cotyledons of young seedlings where FLC might contribute to preventing precocious activation of FT. FLC can also be detected at the shoot apex where it most likely acts on targets different from FT, as the latter is not normally expressed at the meristem.
A likely target of FLC at the shoot meristem (and possibly also in leaves) is SOC1. SOC1 was initially cloned as a suppressor of CO overexpression but was later shown to also be regulated by GA signalling [155]. More recently, Wang et al. [173] demonstrated that SOC1 mRNA levels also increase in an miR156/SPL-dependent manner. As the latter have been linked to the regulation of flowering dependent on plant age, SOC1 apparently integrates signals not only from three but (at least) four different pathways.
SOC1’s function as a positive regulator of flowering at the shoot apex is tightly associated with the activity of yet another MADS box protein, AGL24. Similar to what has been described for SOC1, expression of AGL24 is positively regulated by vernalization. However, in contrast to SOC1, which is a direct target of FLC, induction of AGL24 was found to be independent of FLC [204]. The situation is further complicated by the fact that SOC1 and AGL24 directly regulate one another’s expression [158]. In addition, heterodimerization of the two proteins seems to be a prerequisite for SOC1 translocation into the nucleus and binding to the LFY promoter [194].
LFY was first recognized for its function in flower meristem development, and lfy mutants showed homeotic transformations with leaf-like structures replacing the floral organs [195]. In agreement with its function in flower development, LFY RNA is most strongly expressed in floral meristems [146, 147, 195]. However, later analyses indicated that LFY mRNA was also detectable in young leaf primordia [146], suggesting that LFY might also have a function during vegetative development, and subsequently it was demonstrated that overexpression of LFY resulted in an early flowering phenotype [146, 196]. Several regulating pathways converge on the LFY promoter. As mentioned above, LFY is a direct target of SOC1 [194]. In addition, expression of LFY is also controlled by GA [148]. LFY was recently shown to be a target of the miR156-regulated SPLs as well [66]. Taken together these findings indicate that LFY is regulated by multiple input signals and has a dual function in flowering time and flower meristem identity regulation.
LFY is by no means the only protein that, in addition to its role in regulating the floral transition, also functions in flower development. For example, overexpression of AGL24 resulted in, besides early flowering, the formation of ectopic floral organs [197]. This indicated that flowering time and flower development pathways are not always clearly separated but are in part controlled by the same factors. Furthermore, higher-order mutant combinations of agl24, soc1, and svp displayed clear homeotic transformation [198].
Overall, recent studies indicate that activation of flower development genes by the floral integrators is rather direct and apparently does not require a long signalling cascade. For example, FD has been reported to bind directly to the promoter of the A-class gene, APETALA1 [68]. In addition, expression of class B and C homeotic genes was shown to be regulated by SVP, SOC1, and AGL24, through direct control of the LFY coregulator SEP3 [198]. Repression of SEP3 by SOC1 and AGL24 was further shown to be essential to prevent precocious organ formation in flowers [198]. The exact order of events that control flower meristem formation and flower differentiation has recently been reviewed [199–201].
The flower development genes not only activate organ formation, but they also turn off the flowering time genes in the emerging flower primordia, thus ensuring a sharp transition to flower formation. This has been demonstrated nicely in recent studies that used chromatin-immunoprecipitation followed by hybridization to tiling arrays or high throughput sequencing to identify targets of a number of homeotic genes on a genome-wide scale. A good example is AP1, which was shown to induce SEP3 while simultaneously downregulating AGL24 and SVP during early flower development. Ultimately this results in the formation of AP1/SEP3 heterodimers in the emerging flower bud, which then activate additional floral homeotic genes required for floral organ formation [202]. At the same time, however, AP1 also binds to and represses FD and FDP in the emerging flower from stage two onward [203]. Similarly, SEP3 was shown to bind to and repress SOC1 and SVP, while activating expression of a large number of floral homeotic genes. Finally, the A-class protein AP2 was also shown to bind to and negatively regulate SOC1, among other genes [192].
Conclusions and outlook
Correct timing of the floral transition is crucial to ensure reproductive success. The floral transition is thus regulated by an intricate network of genetically defined but nevertheless interacting pathways that perceive and respond to a variety of endogenous and environmental stimuli. The last few years have seen great advances in our understanding of the molecular mechanisms that control the transition from vegetative growth to reproductive development. Dozens of genes involved in flowering time regulation have been identified and characterized in detail. However, despite these recent advances, many questions remain open.
For example, it is now widely accepted that the FT proteins constitute a florigenic signal that relays the information to initiate flowering in response to inductive photoperiod from the leaves to the shoot apex. However, it is completely unclear how FT is loaded to the phloem sieve elements and how unloading at the shoot meristem is accomplished. Is this purely passive or do checkpoints exist that regulate FT movement, for example at the plasmodesmata? Also, the molecular function of FT is not fully understood. The interaction of FT with the transcription factor FD suggests a role of FT in regulating transcription; however, evidence to this end is sparse.
Another field that will likely attract a lot of attention in the future will be the analyses of the processes that control the temporal and spatial order of events at the shoot meristem that eventually lead to the formation of flowers. The genome-wide identification of targets of only a limited number of transcription factors involved in flowering time regulation and flower development has already greatly advanced our understanding of these processes. At the same time these experiments clearly demonstrate that the transcriptional processes that govern flowering are more complex than previously thought. Also, as long as information on the changes that occur during the floral transition at the level of the chromatin landscape is lacking, our understanding of the floral transition will remain incomplete. In the end it will be necessary to obtain tissue-specific quantitative information on the various factors and parameters that control flowering, as only this type of information will put us in a position to model the regulation of flowering and predict the effects a changing environment will have on this ecologically and economically important trait.
Acknowledgments
The authors would like to thank Dr. Yasushi Kobayashi, Dr. Sureshkumar Balasubramanian, Dr. Jia Wei Wang, Dr. Lisa Smith, Dr. Dan Koenig, Dr. Beth Rowan, Dr. George Wang and Vinicius Galvao for comments on the manuscript and the Max-Planck Gesellschaft for funding.
Abbreviations
- GA
Gibberellic acid
- SD
Short day
- LD
Long day
References
- 1.Kobayashi Y, Weigel D. Move on up, it’s time for change—mobile signals controlling photoperiod-dependent flowering. Genes Dev. 2007;21:2371–2384. doi: 10.1101/gad.1589007. [DOI] [PubMed] [Google Scholar]
- 2.Pittendrigh CS. Circadian rhythms and the circadian organization of living systems. Cold Spring Harb Symp Quant Biol. 1960;25:159–184. doi: 10.1101/sqb.1960.025.01.015. [DOI] [PubMed] [Google Scholar]
- 3.Pittendrigh C. Circadian surfaces and the diversity of possible roles of circadian organization in photoperiodic induction. Proc Natl Acad Sci USA. 1972;69:2734–2737. doi: 10.1073/pnas.69.9.2734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Knott J. Effect of a localized photoperiod on spinach. Proc Soc Hortic Sci. 1934;31:152–154. [Google Scholar]
- 5.Evans LT. Flower induction and the florigen concept. Annu Rev Plant Physiol. 1971;22:365–394. [Google Scholar]
- 6.Turck F, Fornara F, Coupland G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu Rev Plant Biol. 2008;59:573–594. doi: 10.1146/annurev.arplant.59.032607.092755. [DOI] [PubMed] [Google Scholar]
- 7.King RW, Evans LT, Wardlaw IF. Translocation of the floral stimulus in Pharbitis nil in relation to that of assimilates. Z Pflanzenphysiol. 1968;59:377–388. [Google Scholar]
- 8.King RW, Zeevaart JA. Floral stimulus movement in perilla and flower inhibition caused by noninduced leaves. Plant Physiol. 1973;51(4):727–738. doi: 10.1104/pp.51.4.727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Smith H, Whitelam GC. The shade avoidance syndrome: multiple responses mediated by multiple phytochromes. Plant Cell Environ. 1997;20(6):840–844. [Google Scholar]
- 10.Tsuchiya T, Ishiguri Y. Role of the quality of light on the photoperiodic flowering response in four latitudinal ecotypes of Chenopodium rubrum . Plant Cell Physiol. 1981;22:525–532. [Google Scholar]
- 11.Gassner G. Beiträge zur physiologischen Charakteristik sommer- und winterannueller Gewächse, insbesondere der Getreidepflanzen. Z Bot. 1918;10:417–480. [Google Scholar]
- 12.Purvis ON, Gregory FG. The reversibility by high temperature of the vernalised condition in Petkus winter rye. Ann Bot. 1952;16:1–21. [Google Scholar]
- 13.Wang JY. A critique of the heat unit approach to plant-response studies. Ecology. 1960;41(4):785–790. [Google Scholar]
- 14.Samach A, Wigge P. Ambient temperature perception in plants. Curr Opin Plant Biol. 2005;8:483–486. doi: 10.1016/j.pbi.2005.07.011. [DOI] [PubMed] [Google Scholar]
- 15.Lang A. The effect of gibberellin upon flower formation. Proc Natl Acad Sci USA. 1957;43:709–717. doi: 10.1073/pnas.43.8.709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lang A. Gibberellin-like substances in photoinduced and vegetative Hyoscyamus plants. Planta. 1960;54:498–504. [Google Scholar]
- 17.Langridge J. Effect of day-length and gibberellic acid on the flowering of Arabidopsis . Nature. 1957;180:36–37. [Google Scholar]
- 18.Chandler J, Dean C. Factors influencing the vernalization response and flowering time of late flowering mutants of Arabidopsis thaliana (L.) Heynh. J Exp Bot. 1994;45:1279–1288. [Google Scholar]
- 19.Bernier G, Havelange A, Houssa C, Petitjean A, Lejeune P. Physiological signals that induce flowering. Plant Cell. 1993;5:1147–1155. doi: 10.1105/tpc.5.10.1147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bodson M, Outlaw WH. Elevation in the sucrose content of the shoot apical meristem of Sinapis alba at floral evocation. Plant Physiol. 1985;79(2):420–424. doi: 10.1104/pp.79.2.420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Laibach F. Arabidopsis thaliana (L.) Heynh. als Objekt für genetische und entwicklungsphysiologische Untersuchungen. Bot Arch. 1943;44:439–455. [Google Scholar]
- 22.Koornneef M, Alonso-Blanco C, Vreugdenhil D. Naturally occurring genetic variation in Arabidopsis thaliana . Annu Rev Plant Biol. 2004;55:141–172. doi: 10.1146/annurev.arplant.55.031903.141605. [DOI] [PubMed] [Google Scholar]
- 23.Lempe J, Balasubramanian S, Sureshkumar S, Singh A, Schmid M, Weigel D. Diversity of flowering responses in wild Arabidopsis thaliana strains. PLoS Genet. 2005;1:109–118. doi: 10.1371/journal.pgen.0010006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sung S, Amasino RM. Vernalization and epigenetics: how plants remember winter. Curr Opin Plant Biol. 2004;7(1):4–10. doi: 10.1016/j.pbi.2003.11.010. [DOI] [PubMed] [Google Scholar]
- 25.Johanson U, West J, Lister C, Michaels S, Amasino R, Dean C. Molecular analysis of FRIGIDA, a major determinant of natural variation in Arabidopsis flowering time. Science. 2000;290:344–347. doi: 10.1126/science.290.5490.344. [DOI] [PubMed] [Google Scholar]
- 26.Michaels S, Amasino R. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell. 1999;11:949–956. doi: 10.1105/tpc.11.5.949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rédei GP. Supervital mutants of Arabidopsis . Genetics. 1962;47:443–460. doi: 10.1093/genetics/47.4.443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Koornneef M, Hanhart C, van der Veen J. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana . Mol Gen Genet. 1991;229:57–66. doi: 10.1007/BF00264213. [DOI] [PubMed] [Google Scholar]
- 29.Lariguet P, Dunand C. Plant photoreceptors: phylogenetic overview. J Mol Evol. 2005;61:559–569. doi: 10.1007/s00239-004-0294-2. [DOI] [PubMed] [Google Scholar]
- 30.Li Q, Yang H. Cryptochrome signaling in plants. Photochem Photobiol. 2007;83:94–101. doi: 10.1562/2006-02-28-IR-826. [DOI] [PubMed] [Google Scholar]
- 31.Quail P, Boylan M, Parks B, Short T, Xu Y, Wagner D. Phytochromes: photosensory perception and signal transduction. Science. 1995;268:675–680. doi: 10.1126/science.7732376. [DOI] [PubMed] [Google Scholar]
- 32.Tóth R, Kevei E, Hall A, Millar A, Nagy F, Kozma-Bognar L. Circadian clock-regulated expression of phytochrome and cryptochrome genes in Arabidopsis . Plant Physiol. 2001;127:1607–1616. doi: 10.1104/pp.010467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Putterill J, Robson F, Lee K, Simon R, Coupland G. The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell. 1995;80:847–857. doi: 10.1016/0092-8674(95)90288-0. [DOI] [PubMed] [Google Scholar]
- 34.An H, Roussot C, Suárez-López P, Corbesier L, Vincent C, Píniro M, Hepworth S, Mouradov A, Justin S, Turnbull C, Coupland G. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis . Development. 2004;131:3615–3626. doi: 10.1242/dev.01231. [DOI] [PubMed] [Google Scholar]
- 35.Suárez-López P, Wheatley K, Robson F, Onouchi H, Valverde F, Coupland G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis . Nature. 2001;410:1116–1120. doi: 10.1038/35074138. [DOI] [PubMed] [Google Scholar]
- 36.Fornara F, Panigrahi K, Gissot L, Sauerbrunn N, Rühl M, Jarillo J, Coupland G. Arabidopsis DOF transcription factors act redundantly to reduce CONSTANS expression and are essential for a photoperiodic flowering response. Dev Cell. 2009;17:75–86. doi: 10.1016/j.devcel.2009.06.015. [DOI] [PubMed] [Google Scholar]
- 37.Imaizumi T, Schultz T, Harmon F, Ho L, Kay S. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis . Science. 2005;309:293–297. doi: 10.1126/science.1110586. [DOI] [PubMed] [Google Scholar]
- 38.Sawa M, Nusinow D, Kay S, Imaizumi T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis . Science. 2007;318:261–265. doi: 10.1126/science.1146994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Imaizumi T, Tran H, Swartz T, Briggs W, Kay S. FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis . Nature. 2003;426:302–306. doi: 10.1038/nature02090. [DOI] [PubMed] [Google Scholar]
- 40.Liu L, Zhang Y, Li Q, Sang Y, Mao J, Lian H, Wang L, Yang H. COP1-mediated ubiquitination of CONSTANS is implicated in cryptochrome regulation of flowering in Arabidopsis . Plant Cell. 2008;20:292–306. doi: 10.1105/tpc.107.057281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hoecker U, Quail P. The phytochrome A-specific signaling intermediate SPA1 interacts directly with COP1, a constitutive repressor of light signaling in Arabidopsis . J Biol Chem. 2001;276:38173–38178. doi: 10.1074/jbc.M103140200. [DOI] [PubMed] [Google Scholar]
- 42.Laubinger S, Marchal V, Le Gourrierec J, Gentilhomme J, Wenkel S, Adrian J, Jang S, Kulajta C, Braun H, Coupland G, Hoecker U. Arabidopsis SPA proteins regulate photoperiodic flowering and interact with the floral inducer CONSTANS to regulate its stability. Development. 2006;133:3213–3222. doi: 10.1242/dev.02481. [DOI] [PubMed] [Google Scholar]
- 43.Laubinger S, Hoecker U. The SPA1-like proteins SPA3 and SPA4 repress photomorphogenesis in the light. Plant J. 2003;35:373–385. doi: 10.1046/j.1365-313x.2003.01813.x. [DOI] [PubMed] [Google Scholar]
- 44.Valverde F, Mouradov A, Soppe W, Ravenscroft D, Samach A, Coupland G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science. 2004;303:1003–1006. doi: 10.1126/science.1091761. [DOI] [PubMed] [Google Scholar]
- 45.Jang S, Marchal V, Panigrahi K, Wenkel S, Soppe W, Deng X, Valverde F, Coupland G. Arabidopsis COP1 shapes the temporal pattern of CO accumulation conferring a photoperiodic flowering response. EMBO J. 2008;27:1277–1288. doi: 10.1038/emboj.2008.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Morris K, Thornber S, Codrai L, Richardson C, Craig A, Sadanandom A, Thomas B, Jackson S. DAY NEUTRAL FLOWERING represses CONSTANS to prevent Arabidopsis flowering early in short days. Plant Cell. 2010;22(4):1118–1128. doi: 10.1105/tpc.109.066605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ayre BG, Turgeon R. Graft transmission of a floral stimulant derived from CONSTANS. Plant Physiol. 2004;135(4):2271–2278. doi: 10.1104/pp.104.040592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kardailsky I, Shukla V, Ahn J, Dagenais N, Christensen S, Nguyen J, Chory J, Harrison M, Weigel D. Activation tagging of the floral inducer FT. Science. 1999;286:1962–1965. doi: 10.1126/science.286.5446.1962. [DOI] [PubMed] [Google Scholar]
- 49.Weigel D, Ahn JH, Blazquez MA, Borevitz JO, Christensen SK, Fankhauser C, Ferrandiz C, Kardailsky I, Malancharuvil EJ, Neff MM, Nguyen JT, Sato S, Wang ZY, Xia Y, Dixon RA, Harrison MJ, Lamb CJ, Yanofsky MF, Chory J. Activation tagging in Arabidopsis . Plant Physiol. 2000;122(4):1003–1013. doi: 10.1104/pp.122.4.1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kaya H, Sato S, Tabata S, Kobayashi Y, Iwabuchi M, Araki T. hosoba toge toge, a syndrome caused by a large chromosomal deletion associated with a T-DNA insertion in Arabidopsis . Plant Cell Physiol. 2000;41(9):1055–1066. doi: 10.1093/pcp/pcd032. [DOI] [PubMed] [Google Scholar]
- 51.Kobayashi Y, Kaya H, Goto K, Iwabuchi M, Araki T. A pair of related genes with antagonistic roles in mediating flowering signals. Science. 1999;286:1960–1962. doi: 10.1126/science.286.5446.1960. [DOI] [PubMed] [Google Scholar]
- 52.Ahn J, Miller D, Winter V, Banfield M, Lee J, Yoo S, Henz S, Brady R, Weigel D. A divergent external loop confers antagonistic activity on floral regulators FT and TFL1. EMBO J. 2006;25:605–614. doi: 10.1038/sj.emboj.7600950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Harmer S, Hogenesch J, Straume M, Chang H, Han B, Zhu T, Wang X, Kreps J, Kay S. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science. 2000;290:2110–2113. doi: 10.1126/science.290.5499.2110. [DOI] [PubMed] [Google Scholar]
- 54.Takada S, Goto K. Terminal flower2, an Arabidopsis homolog of heterochromatin protein1, counteracts the activation of flowering locus T by constans in the vascular tissues of leaves to regulate flowering time. Plant Cell. 2003;15(12):2856–2865. doi: 10.1105/tpc.016345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Farrona S, Coupland G, Turck F. The impact of chromatin regulation on the floral transition. Semin Cell Dev Biol. 2008;19(6):560–573. doi: 10.1016/j.semcdb.2008.07.015. [DOI] [PubMed] [Google Scholar]
- 56.Exner V, Aichinger E, Shu H, Wildhaber T, Alfarano P, Caflisch A, Gruissem W, Kohler C, Hennig L. The chromodomain of LIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development. PLoS One. 2009;4(4):e5335. doi: 10.1371/journal.pone.0005335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Turck F, Roudier F, Farrona S, Martin-Magniette ML, Guillaume E, Buisine N, Gagnot S, Martienssen RA, Coupland G, Colot V. Arabidopsis TFL2/LHP1 specifically associates with genes marked by trimethylation of histone H3 lysine 27. PLoS Genet. 2007;3(6):e86. doi: 10.1371/journal.pgen.0030086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhang X, Clarenz O, Cokus S, Bernatavichute YV, Pellegrini M, Goodrich J, Jacobsen SE. Whole-genome analysis of histone H3 lysine 27 trimethylation in Arabidopsis . PLoS Biol. 2007;5(5):e129. doi: 10.1371/journal.pbio.0050129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Köhler C, Hennig L, Bouveret R, Gheyselinck J, Grossniklaus U, Gruissem W. Arabidopsis MSI1 is a component of the MEA/FIE Polycomb group complex and required for seed development. EMBO J. 2003;22(18):4804–4814. doi: 10.1093/emboj/cdg444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Schubert D, Primavesi L, Bishopp A, Roberts G, Doonan J, Jenuwein T, Goodrich J. Silencing by plant Polycomb-group genes requires dispersed trimethylation of histone H3 at lysine 27. EMBO J. 2006;25(19):4638–4649. doi: 10.1038/sj.emboj.7601311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yoshida N, Yanai Y, Chen L, Kato Y, Hiratsuka J, Miwa T, Sung ZR, Takahashi S. EMBRYONIC FLOWER2, a novel polycomb group protein homolog, mediates shoot development and flowering in Arabidopsis . Plant Cell. 2001;13(11):2471–2481. doi: 10.1105/tpc.010227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Jiang D, Wang Y, He Y. Repression of FLOWERING LOCUS C and FLOWERING LOCUS T by the Arabidopsis Polycomb repressive complex 2 components. PLoS One. 2008;3(10):e3404. doi: 10.1371/journal.pone.0003404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hennig L, Derkacheva M. Diversity of Polycomb group complexes in plants: same rules, different players? Trends Genet. 2009;25(9):414–423. doi: 10.1016/j.tig.2009.07.002. [DOI] [PubMed] [Google Scholar]
- 64.Adrian J, Farrona S, Reimer J, Albani M, Coupland G, Turck F. cis-Regulatory elements and chromatin state coordinately control temporal and spatial expression of FLOWERING LOCUS T in Arabidopsis . Plant Cell. 2010;22(5):1425–1440. doi: 10.1105/tpc.110.074682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Corbesier L, Vincent C, Jang S, Fornara F, Fan Q, Searle I, Giakountis A, Farrona S, Gissot L, Turnbull C, Coupland G. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis . Science. 2007;316:1030–1033. doi: 10.1126/science.1141752. [DOI] [PubMed] [Google Scholar]
- 66.Yamaguchi A, Kobayashi Y, Goto K, Abe M, Araki T. TWIN SISTER OF FT (TSF) acts as a floral pathway integrator redundantly with FT. Plant Cell Physiol. 2005;46(8):1175–1189. doi: 10.1093/pcp/pci151. [DOI] [PubMed] [Google Scholar]
- 67.Yoo SK, Chung KS, Kim J, Lee JH, Hong SM, Yoo SJ, Yoo SY, Lee JS, Ahn JH. CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis . Plant Physiol. 2005;139(2):770–778. doi: 10.1104/pp.105.066928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wigge P, Kim M, Jaeger K, Busch W, Schmid M, Lohmann J, Weigel D. Integration of spatial and temporal information during floral induction in Arabidopsis . Science. 2005;309:1056–1059. doi: 10.1126/science.1114358. [DOI] [PubMed] [Google Scholar]
- 69.Samach A, Onouchi H, Gold S, Ditta G, Schwarz-Sommer Z, Yanofsky M, Coupland G. Distinct roles of CONSTANS target genes in reproductive development of Arabidopsis . Science. 2000;288:1613–1616. doi: 10.1126/science.288.5471.1613. [DOI] [PubMed] [Google Scholar]
- 70.Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science. 2005;309:1052–1056. doi: 10.1126/science.1115983. [DOI] [PubMed] [Google Scholar]
- 71.Mathieu J, Warthmann N, Küttner F, Schmid M. Export of FT protein from phloem companion cells is sufficient for floral induction in Arabidopsis . Curr Biol. 2007;17:1055–1060. doi: 10.1016/j.cub.2007.05.009. [DOI] [PubMed] [Google Scholar]
- 72.Jäger K, Wigge P. FT protein acts as a long-range signal in Arabidopsis . Curr Biol. 2007;17:1050–1054. doi: 10.1016/j.cub.2007.05.008. [DOI] [PubMed] [Google Scholar]
- 73.Notaguchi M, Abe M, Kimura T, Daimon Y, Kobayashi T, Yamaguchi A, Tomita Y, Dohi K, Mori M, Araki T. Long-distance, graft-transmissible action of Arabidopsis FLOWERING LOCUS T protein to promote flowering. Plant Cell Physiol. 2008;49:1645–1658. doi: 10.1093/pcp/pcn154. [DOI] [PubMed] [Google Scholar]
- 74.Li C, Zhang K, Zeng X, Jackson S, Zhou Y, Hong Y. A cis element within flowering locus T mRNA determines its mobility and facilitates trafficking of heterologous viral RNA. J Virol. 2009;83:3540–3548. doi: 10.1128/JVI.02346-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, Yano M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002;43(10):1096–1105. doi: 10.1093/pcp/pcf156. [DOI] [PubMed] [Google Scholar]
- 76.Yano M, Katayose Y, Ashikari M, Yamanouchi U, Monna L, Fuse T, Baba T, Yamamoto K, Umehara Y, Nagamura Y, Sasaki T. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell. 2000;12(12):2473–2484. doi: 10.1105/tpc.12.12.2473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Komiya R, Ikegami A, Tamaki S, Yokoi S, Shimamoto K. Hd3a and RFT1 are essential for flowering in rice. Development. 2008;135:767–774. doi: 10.1242/dev.008631. [DOI] [PubMed] [Google Scholar]
- 78.Doi K, Izawa T, Fuse T, Yamanouchi U, Kubo T, Shimatani Z, Yano M, Yoshimura A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004;18(8):926–936. doi: 10.1101/gad.1189604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Itoh H, Nonoue Y, Yano M, Izawa T. A pair of floral regulators sets critical day length for Hd3a florigen expression in rice. Nat Genet. 2010;42(7):635–638. doi: 10.1038/ng.606. [DOI] [PubMed] [Google Scholar]
- 80.Tamaki S, Matsuo S, Wong HL, Yokoi S, Shimamoto K. Hd3a protein is a mobile flowering signal in rice. Science. 2007;316(5827):1033–1036. doi: 10.1126/science.1141753. [DOI] [PubMed] [Google Scholar]
- 81.Hayama R, Agashe B, Luley E, King R, Coupland G. A circadian rhythm set by dusk determines the expression of FT homologs and the short-day photoperiodic flowering response in Pharbitis . Plant Cell. 2007;19:2988–3000. doi: 10.1105/tpc.107.052480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Lifschitz E, Eviatar T, Rozman A, Shalit A, Goldshmidt A, Amsellem Z, Alvarez J, Eshed Y. The tomato FT ortholog triggers systemic signals that regulate growth and flowering and substitute for diverse environmental stimuli. Proc Natl Acad Sci USA. 2006;103:6398–6403. doi: 10.1073/pnas.0601620103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lifschitz E, Eshed Y. Universal florigenic signals triggered by FT homologues regulate growth and flowering cycles in perennial day-neutral tomato. J Exp Bot. 2006;57:3405–3414. doi: 10.1093/jxb/erl106. [DOI] [PubMed] [Google Scholar]
- 84.Alonso-Blanco C, Koornneef M. Naturally occurring variation in Arabidopsis: an underexploited resource for plant genetics. Trends Plant Sci. 2000;5(1):22–29. doi: 10.1016/s1360-1385(99)01510-1. [DOI] [PubMed] [Google Scholar]
- 85.Napp-Zinn K. Vernalization, environmental and genetic regulation. In: Atherton JG, editor. Manipulation of flowering. London: Butterworths; 1987. pp. 123–132. [Google Scholar]
- 86.Koornneef M, Blankestijn-de Vries H, Hanhart C, Soppe W, Peeters T. The phenotype of some late-flowering mutants is enhanced by a locus on chromosome 5 that is not effective in the Landsberg erecta wild-type. Plant J. 1994;6:911–919. [Google Scholar]
- 87.Lee I, Aukerman M, Gore S, Lohman K, Michaels S, Weaver L, John M, Feldmann K, Amasino R. Isolation of LUMINIDEPENDENS: a gene involved in the control of flowering time in Arabidopsis . Plant Cell. 1994;6:75–83. doi: 10.1105/tpc.6.1.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Geraldo N, Bäurle I, Kidou S, Hu X, Dean C. FRIGIDA delays flowering in Arabidopsis via a cotranscriptional mechanism involving direct interaction with the nuclear cap-binding complex. Plant Physiol. 2009;150:1611–1618. doi: 10.1104/pp.109.137448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Searle I, He Y, Turck F, Vincent C, Fornara F, Krober S, Amasino RA, Coupland G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis . Genes Dev. 2006;20(7):898–912. doi: 10.1101/gad.373506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Sheldon CC, Burn JE, Perez PP, Metzger J, Edwards JA, Peacock WJ, Dennis ES. The FLF MADS box gene: a repressor of flowering in Arabidopsis regulated by vernalization and methylation. Plant Cell. 1999;11(3):445–458. doi: 10.1105/tpc.11.3.445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Helliwell C, Wood C, Robertson M, Peacock WJ, Dennis E. The Arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex. Plant J. 2006;46:183–192. doi: 10.1111/j.1365-313X.2006.02686.x. [DOI] [PubMed] [Google Scholar]
- 92.Hepworth SR, Valverde F, Ravenscroft D, Mouradov A, Coupland G. Antagonistic regulation of flowering-time gene SOC1 by CONSTANS and FLC via separate promoter motifs. EMBO J. 2002;21(16):4327–4337. doi: 10.1093/emboj/cdf432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Michaels S, Amasino R. Loss of FLOWERING LOCUS C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous pathway mutations but not responsiveness to vernalization. Plant Cell. 2001;13:935–941. doi: 10.1105/tpc.13.4.935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Gendall A, Levy Y, Wilson A, Dean C. The VERNALIZATION 2 gene mediates the epigenetic regulation of vernalization in Arabidopsis . Cell. 2001;107:525–535. doi: 10.1016/s0092-8674(01)00573-6. [DOI] [PubMed] [Google Scholar]
- 95.Levy Y, Mesnage S, Mylne J, Gendall A, Dean C. Multiple roles of Arabidopsis VRN1 in vernalization and flowering time control. Science. 2002;297:243–246. doi: 10.1126/science.1072147. [DOI] [PubMed] [Google Scholar]
- 96.Sung S, Amasino R. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature. 2004;427:159–164. doi: 10.1038/nature02195. [DOI] [PubMed] [Google Scholar]
- 97.Bond D, Dennis E, Pogson B, Finnegan E. Histone acetylation, VERNALIZATION INSENSITIVE 3, FLOWERING LOCUS C, and the vernalization response. Mol Plant. 2009;2:724–737. doi: 10.1093/mp/ssp021. [DOI] [PubMed] [Google Scholar]
- 98.De Lucia F, Crevillen P, Jones AM, Greb T, Dean C. A PHD-polycomb repressive complex 2 triggers the epigenetic silencing of FLC during vernalization. Proc Natl Acad Sci USA. 2008;105(44):16831–16836. doi: 10.1073/pnas.0808687105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wood CC, Robertson M, Tanner G, Peacock WJ, Dennis ES, Helliwell CA. The Arabidopsis thaliana vernalization response requires a polycomb-like protein complex that also includes VERNALIZATION INSENSITIVE 3. Proc Natl Acad Sci USA. 2006;103(39):14631–14636. doi: 10.1073/pnas.0606385103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Cao R, Zhang Y. The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr Opin Genet Dev. 2004;14(2):155–164. doi: 10.1016/j.gde.2004.02.001. [DOI] [PubMed] [Google Scholar]
- 101.Finnegan EJ, Dennis ES. Vernalization-induced trimethylation of histone H3 lysine 27 at FLC is not maintained in mitotically quiescent cells. Curr Biol. 2007;17(22):1978–1983. doi: 10.1016/j.cub.2007.10.026. [DOI] [PubMed] [Google Scholar]
- 102.Swiezewski S, Liu F, Magusin A, Dean C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature. 2009;462:799–802. doi: 10.1038/nature08618. [DOI] [PubMed] [Google Scholar]
- 103.Heo JB, Sung S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science. 2011;331(6013):76–79. doi: 10.1126/science.1197349. [DOI] [PubMed] [Google Scholar]
- 104.De Lucia F, Dean C (2010) Long non-coding RNAs and chromatin regulation. Curr Opin Plant Biol. doi:10.1016/j.pbi.2010.11.006 [DOI] [PubMed]
- 105.Simpson G. The autonomous pathway: epigenetic and post-transcriptional gene regulation in the control of Arabidopsis flowering time. Curr Opin Plant Biol. 2004;7:570–574. doi: 10.1016/j.pbi.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 106.Liu F, Marquardt S, Lister C, Swiezewski S, Dean C. Targeted 3′ processing of antisense transcripts triggers Arabidopsis FLC chromatin silencing. Science. 2010;327:94–97. doi: 10.1126/science.1180278. [DOI] [PubMed] [Google Scholar]
- 107.Hecht V, Foucher F, Ferrandiz C, Macknight R, Navarro C, Morin J, Vardy ME, Ellis N, Beltran JP, Rameau C, Weller JL. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 2005;137(4):1420–1434. doi: 10.1104/pp.104.057018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Schläppi M, Patel M. Biennialism and vernalization-promoted flowering in Hyoscyamus niger: a comparison with Arabidopsis . Flower Newsl. 2001;31:25–32. [Google Scholar]
- 109.Tadege M, Sheldon CC, Helliwell CA, Upadhyaya NM, Dennis ES, Peacock WJ. Reciprocal control of flowering time by OsSOC1 in transgenic Arabidopsis and by FLC in transgenic rice. Plant Biotechnol J. 2003;1(5):361–369. doi: 10.1046/j.1467-7652.2003.00034.x. [DOI] [PubMed] [Google Scholar]
- 110.Wang R, Farrona S, Vincent C, Joecker A, Schoof H, Turck F, Alonso-Blanco C, Coupland G, Albani MC. PEP1 regulates perennial flowering in Arabis alpina . Nature. 2009;459(7245):423–427. doi: 10.1038/nature07988. [DOI] [PubMed] [Google Scholar]
- 111.Yan L, Loukoianov A, Blechl A, Tranquilli G, Ramakrishna W, SanMiguel P, Bennetzen JL, Echenique V, Dubcovsky J. The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science. 2004;303(5664):1640–1644. doi: 10.1126/science.1094305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Balasubramanian S, Sureshkumar S, Lempe J, Weigel D. Potent induction of Arabidopsis thaliana flowering by elevated growth temperature. PLoS Genet. 2006;2:e106. doi: 10.1371/journal.pgen.0020106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Halliday KJ, Salter MG, Thingnaes E, Whitelam GC. Phytochrome control of flowering is temperature sensitive and correlates with expression of the floral integrator FT. Plant J. 2003;33(5):875–885. doi: 10.1046/j.1365-313x.2003.01674.x. [DOI] [PubMed] [Google Scholar]
- 114.Blázquez MA, Ahn JH, Weigel D. A thermosensory pathway controlling flowering time in Arabidopsis thaliana . Nat Genet. 2003;33(2):168–171. doi: 10.1038/ng1085. [DOI] [PubMed] [Google Scholar]
- 115.Werner JD, Borevitz JO, Uhlenhaut NH, Ecker JR, Chory J, Weigel D. FRIGIDA-independent variation in flowering time of natural Arabidopsis thaliana accessions. Genetics. 2005;170(3):1197–1207. doi: 10.1534/genetics.104.036533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Hartmann U, Höhmann S, Nettesheim K, Wisman E, Saedler H, Huijser P. Molecular cloning of SVP: a negative regulator of the floral transition in Arabidopsis . Plant J. 2000;21:351–360. doi: 10.1046/j.1365-313x.2000.00682.x. [DOI] [PubMed] [Google Scholar]
- 117.Lee J, Yoo S, Park S, Hwang I, Lee J, Ahn J. Role of SVP in the control of flowering time by ambient temperature in Arabidopsis . Genes Dev. 2007;21:397–402. doi: 10.1101/gad.1518407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Li D, Liu C, Shen L, Wu Y, Chen H, Robertson M, Helliwell C, Ito T, Meyerowitz E, Yu H. A repressor complex governs the integration of flowering signals in Arabidopsis . Dev Cell. 2008;15:110–120. doi: 10.1016/j.devcel.2008.05.002. [DOI] [PubMed] [Google Scholar]
- 119.Kumar S, Wigge P. H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis . Cell. 2010;140:136–147. doi: 10.1016/j.cell.2009.11.006. [DOI] [PubMed] [Google Scholar]
- 120.Choi K, Kim S, Kim SY, Kim M, Hyun Y, Lee H, Choe S, Kim SG, Michaels S, Lee I. SUPPRESSOR OF FRIGIDA3 encodes a nuclear ACTIN-RELATED PROTEIN6 required for floral repression in Arabidopsis . Plant Cell. 2005;17(10):2647–2660. doi: 10.1105/tpc.105.035485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Deal RB, Kandasamy MK, McKinney EC, Meagher RB. The nuclear actin-related protein ARP6 is a pleiotropic developmental regulator required for the maintenance of FLOWERING LOCUS C expression and repression of flowering in Arabidopsis . Plant Cell. 2005;17(10):2633–2646. doi: 10.1105/tpc.105.035196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hedden P, Phillips AL. Gibberellin metabolism: new insights revealed by the genes. Trends Plant Sci. 2000;5(12):523–530. doi: 10.1016/s1360-1385(00)01790-8. [DOI] [PubMed] [Google Scholar]
- 123.Koornneef M, van der Veen J. Induction and analysis of gibberellin sensitive mutants in Arabidopsis thaliana (L.) Heynh. Theor Appl Genet. 1980;58(6):257–263. doi: 10.1007/BF00265176. [DOI] [PubMed] [Google Scholar]
- 124.Sun T, Goodman H, Ausubel F. Cloning the Arabidopsis GA1 locus by genomic subtraction. Plant Cell. 1992;4:119–128. doi: 10.1105/tpc.4.2.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Koornneef M, Vaneden J, Hanhart CJ, Dejongh AMM. Genetic fine-structure of the Ga-1 locus in the higher-plant Arabidopsis thaliana (L.) Heynh. Genet Res. 1983;41(1):57. [Google Scholar]
- 126.Wilson R, Heckman J, Somerville C. Gibberellin is required for flowering in Arabidopsis thaliana under short days. Plant Physiol. 1992;100:403–408. doi: 10.1104/pp.100.1.403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Jacobsen SE, Olszewski NE. Mutations at the SPINDLY locus of Arabidopsis alter gibberellin signal transduction. Plant Cell. 1993;5(8):887–896. doi: 10.1105/tpc.5.8.887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Shimada A, Ueguchi-Tanaka M, Sakamoto T, Fujioka S, Takatsuto S, Yoshida S, Sazuka T, Ashikari M, Matsuoka M. The rice SPINDLY gene functions as a negative regulator of gibberellin signaling by controlling the suppressive function of the DELLA protein, SLR1, and modulating brassinosteroid synthesis. Plant J. 2006;48(3):390–402. doi: 10.1111/j.1365-313X.2006.02875.x. [DOI] [PubMed] [Google Scholar]
- 129.Swain S, Tseng T, Olszewski N. Altered expression of SPINDLY affects gibberellin response and plant development. Plant Physiol. 2001;126:1174–1185. doi: 10.1104/pp.126.3.1174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ueguchi-Tanaka M, Ashikari M, Nakajima M, Itoh H, Katoh E, Kobayashi M, Chow T, Hsing Y, Kitano H, Yamaguchi I, Matsuoka M. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin. Nature. 2005;437:693–698. doi: 10.1038/nature04028. [DOI] [PubMed] [Google Scholar]
- 131.Griffiths J, Murase K, Rieu I, Zentella R, Zhang Z, Powers S, Gong F, Phillips A, Hedden P, Sun T, Thomas S. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis . Plant Cell. 2006;18:3399–3414. doi: 10.1105/tpc.106.047415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Willige B, Ghosh S, Nill C, Zourelidou M, Dohmann E, Maier A, Schwechheimer C. The DELLA domain of GA INSENSITIVE mediates the interaction with the GA INSENSITIVE DWARF1A gibberellin receptor of Arabidopsis . Plant Cell. 2007;19:1209–1220. doi: 10.1105/tpc.107.051441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.King RW, Moritz T, Evans LT, Junttila O, Herlt AJ. Long-day induction of flowering in Lolium temulentum involves sequential increases in specific gibberellins at the shoot apex. Plant Physiol. 2001;127(2):624–632. [PMC free article] [PubMed] [Google Scholar]
- 134.Silverstone A, Ciampaglio C, Sun T. The Arabidopsis RGA gene encodes a transcriptional regulator repressing the gibberellin signal transduction pathway. Plant Cell. 1998;10:155–169. doi: 10.1105/tpc.10.2.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Hisamatsu T, King R. The nature of floral signals in Arabidopsis. II. Roles for FLOWERING LOCUS T (FT) and gibberellin. J Exp Bot. 2008;59:3821–3829. doi: 10.1093/jxb/ern232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Harberd NP, Belfield E, Yasumura Y. The angiosperm gibberellin-GID1-DELLA growth regulatory mechanism: how an “inhibitor of an inhibitor” enables flexible response to fluctuating environments. Plant Cell. 2009;21(5):1328–1339. doi: 10.1105/tpc.109.066969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Sun TP. Gibberellin-GID1-DELLA: a pivotal regulatory module for plant growth and development. Plant Physiol. 2010;154(2):567–570. doi: 10.1104/pp.110.161554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Koornneef M, Elgersma A, Hanhart CJ, Vanloenenmartinet EP, Vanrijn L, Zeevaart JAD. A gibberellin insensitive mutant of Arabidopsis thaliana . Physiol Plant. 1985;65(1):33–39. [Google Scholar]
- 139.Murase K, Hirano Y, Sun T, Hakoshima T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature. 2008;456:459–463. doi: 10.1038/nature07519. [DOI] [PubMed] [Google Scholar]
- 140.Achard P, Liao L, Jiang C, Desnos T, Bartlett J, Fu X, Harberd N. DELLAs contribute to plant photomorphogenesis. Plant Physiol. 2007;143:1163–1172. doi: 10.1104/pp.106.092254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Alvey L, Harberd N. DELLA proteins: integrators of multiple plant growth regulatory inputs? Physiol Plant. 2005;123(2):153–160. [Google Scholar]
- 142.Fu X, Harberd NP. Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature. 2003;421(6924):740–743. doi: 10.1038/nature01387. [DOI] [PubMed] [Google Scholar]
- 143.de Lucas M, Daviere JM, Rodriguez-Falcon M, Pontin M, Iglesias-Pedraz JM, Lorrain S, Fankhauser C, Blazquez MA, Titarenko E, Prat S. A molecular framework for light and gibberellin control of cell elongation. Nature. 2008;451(7177):480–484. doi: 10.1038/nature06520. [DOI] [PubMed] [Google Scholar]
- 144.Schwechheimer C, Willige BC. Shedding light on gibberellic acid signalling. Curr Opin Plant Biol. 2009;12(1):57–62. doi: 10.1016/j.pbi.2008.09.004. [DOI] [PubMed] [Google Scholar]
- 145.Oda A, Fujiwara S, Kamada H, Coupland G, Mizoguchi T. Antisense suppression of the Arabidopsis PIF3 gene does not affect circadian rhythms but causes early flowering and increases FT expression. FEBS Lett. 2004;557(1–3):259–264. doi: 10.1016/s0014-5793(03)01470-4. [DOI] [PubMed] [Google Scholar]
- 146.Blázquez MA, Soowal LN, Lee I, Weigel D. LEAFY expression and flower initiation in Arabidopsis . Development. 1997;124(19):3835–3844. doi: 10.1242/dev.124.19.3835. [DOI] [PubMed] [Google Scholar]
- 147.Blázquez M, Green R, Nilsson O, Sussman M, Weigel D. Gibberellins promote flowering of Arabidopsis by activating the LEAFY promoter. Plant Cell. 1998;10:791–800. doi: 10.1105/tpc.10.5.791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Eriksson S, Böhlenius H, Moritz T, Nilsson O. GA4 is the active gibberellin in the regulation of LEAFY transcription and Arabidopsis floral initiation. Plant Cell. 2006;18:2172–2181. doi: 10.1105/tpc.106.042317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Blázquez M, Weigel D. Integration of floral inductive signals in Arabidopsis . Nature. 2000;404:889–892. doi: 10.1038/35009125. [DOI] [PubMed] [Google Scholar]
- 150.Gocal G, Sheldon C, Gubler F, Moritz T, Bagnall D, MacMillan C, Li S, Parish R, Dennis E, Weigel D, King R. GAMYB-like genes, flowering, and gibberellin signaling in Arabidopsis . Plant Physiol. 2001;127:1682–1693. [PMC free article] [PubMed] [Google Scholar]
- 151.Gocal G, Poole A, Gubler F, Watts R, Blundell C, King R. Long-day up-regulation of a GAMYB gene during Lolium temulentum inflorescence formation. Plant Physiol. 1999;119:1271–1278. doi: 10.1104/pp.119.4.1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Palatnik JF, Allen E, Wu X, Schommer C, Schwab R, Carrington JC, Weigel D. Control of leaf morphogenesis by microRNAs. Nature. 2003;425(6955):257–263. doi: 10.1038/nature01958. [DOI] [PubMed] [Google Scholar]
- 153.Park W, Li J, Song R, Messing J, Chen X. CARPEL FACTORY, a Dicer homolog, and HEN1, a novel protein, act in microRNA metabolism in Arabidopsis thaliana . Curr Biol. 2002;12(17):1484–1495. doi: 10.1016/s0960-9822(02)01017-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Achard P, Herr A, Baulcombe D, Harberd N. Modulation of floral development by a gibberellin-regulated microRNA. Development. 2004;131:3357–3365. doi: 10.1242/dev.01206. [DOI] [PubMed] [Google Scholar]
- 155.Moon J, Suh S, Lee H, Choi K, Hong C, Paek N, Kim S, Lee I. The SOC1 MADS-box gene integrates vernalization and gibberellin signals for flowering in Arabidopsis . Plant J. 2003;35:613–623. doi: 10.1046/j.1365-313x.2003.01833.x. [DOI] [PubMed] [Google Scholar]
- 156.Achard P, Baghour M, Chapple A, Hedden P, Van Der Straeten D, Genschik P, Moritz T, Harberd N. The plant stress hormone ethylene controls floral transition via DELLA-dependent regulation of floral meristem-identity genes. Proc Natl Acad Sci USA. 2007;104:6484–6489. doi: 10.1073/pnas.0610717104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Yu H, Xu Y, Tan E, Kumar P. AGAMOUS-LIKE 24, a dosage-dependent mediator of the flowering signals. Proc Natl Acad Sci USA. 2002;99:16336–16341. doi: 10.1073/pnas.212624599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Liu C, Chen H, Er H, Soo H, Kumar P, Han J, Liou Y, Yu H. Direct interaction of AGL24 and SOC1 integrates flowering signals in Arabidopsis . Development. 2008;135:1481–1491. doi: 10.1242/dev.020255. [DOI] [PubMed] [Google Scholar]
- 159.Richter R, Behringer C, Müller I, Schwechheimer C. The GATA-type transcription factors GNC and GNL/CGA1 repress gibberellin signaling downstream from DELLA proteins and PHYTOCHROME-INTERACTING FACTORS. Genes Dev. 2010;24:2093–2104. doi: 10.1101/gad.594910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Madhusudanan KN, Nandakumar S. Carbohydrate changes in shoot tip and subtending leaves during ontogenetic development of pineapple. Z Pflanzenphysiol. 1983;110(5):429–438. [Google Scholar]
- 161.Komarova EN, Milyaeva EL. Changes in content and distribution of starch in stem apices of bicolored coneflower during the period of flowering evocation. Sov Plant Physiol. 1991;38(1):46–51. [Google Scholar]
- 162.Lejeune P, Bernier G, Requier MC, Kinet JM. Sucrose increase during floral induction in the phloem sap collected at the apical part of the shoot of the long-day plant Sinapis alba L. Planta. 1993;190(1):71–74. [Google Scholar]
- 163.King RW, Evans LT. Shoot apex sugars in relation to long-day induction of flowering in Lolium temulentum L. Aust J Plant Physiol. 1991;18(2):121–135. [PMC free article] [PubMed] [Google Scholar]
- 164.Ohto M, Onai K, Furukawa Y, Aoki E, Araki T, Nakamura K. Effects of sugar on vegetative development and floral transition in Arabidopsis . Plant Physiol. 2001;127:252–261. doi: 10.1104/pp.127.1.252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Roldán M, Gómez-Mena C, Ruiz-García L, Salinas J, Martínez-Zapater J. Sucrose availability on the aerial part of the plant promotes morphogenesis and flowering of Arabidopsis in the dark. Plant J. 1999;20:581–590. doi: 10.1046/j.1365-313x.1999.00632.x. [DOI] [PubMed] [Google Scholar]
- 166.El-Lithy M, Reymond M, Stich B, Koornneef M, Vreugdenhil D. Relation among plant growth, carbohydrates and flowering time in the Arabidopsis Landsberg erecta × Kondara recombinant inbred line population. Plant Cell Environ. 2010;33(8):1369–1382. doi: 10.1111/j.1365-3040.2010.02155.x. [DOI] [PubMed] [Google Scholar]
- 167.Caspar T, Huber SC, Somerville C. Alterations in growth, photosynthesis, and respiration in a starchless mutant of Arabidopsis thaliana (L.) deficient in chloroplast phosphoglucomutase activity. Plant Physiol. 1985;79(1):11–17. doi: 10.1104/pp.79.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Caspar T, Lin TP, Kakefuda G, Benbow L, Preiss J, Somerville C. Mutants of Arabidopsis with altered regulation of starch degradation. Plant Physiol. 1991;95(4):1181–1188. doi: 10.1104/pp.95.4.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Corbesier L, Lejeune P, Bernier G. The role of carbohydrates in the induction of flowering in Arabidopsis thaliana: comparison between the wild type and a starchless mutant. Planta. 1998;206:131–137. doi: 10.1007/s004250050383. [DOI] [PubMed] [Google Scholar]
- 170.Paul MJ, Primavesi LF, Jhurreea D, Zhang Y. Trehalose metabolism and signaling. Annu Rev Plant Biol. 2008;59:417–441. doi: 10.1146/annurev.arplant.59.032607.092945. [DOI] [PubMed] [Google Scholar]
- 171.Eastmond PJ, van Dijken AJ, Spielman M, Kerr A, Tissier AF, Dickinson HG, Jones JD, Smeekens SC, Graham IA. Trehalose-6-phosphate synthase 1, which catalyses the first step in trehalose synthesis, is essential for Arabidopsis embryo maturation. Plant J. 2002;29(2):225–235. doi: 10.1046/j.1365-313x.2002.01220.x. [DOI] [PubMed] [Google Scholar]
- 172.van Dijken AJ, Schluepmann H, Smeekens SC. Arabidopsis trehalose-6-phosphate synthase 1 is essential for normal vegetative growth and transition to flowering. Plant Physiol. 2004;135(2):969–977. doi: 10.1104/pp.104.039743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Wang J, Czech B, Weigel D. miR156-regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana . Cell. 2009;138:738–749. doi: 10.1016/j.cell.2009.06.014. [DOI] [PubMed] [Google Scholar]
- 174.Noh B, Lee SH, Kim HJ, Yi G, Shin EA, Lee M, Jung KJ, Doyle MR, Amasino RM, Noh YS. Divergent roles of a pair of homologous jumonji/zinc-finger-class transcription factor proteins in the regulation of Arabidopsis flowering time. Plant Cell. 2004;16(10):2601–2613. doi: 10.1105/tpc.104.025353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Koornneef M, Alonso-Blanco C, Blankestijn-de Vries H, Hanhart CJ, Peeters AJ. Genetic interactions among late-flowering mutants of Arabidopsis . Genetics. 1998;148(2):885–892. doi: 10.1093/genetics/148.2.885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.He Y, Michaels S, Amasino R. Regulation of flowering time by histone acetylation in Arabidopsis . Science. 2003;302:1751–1754. doi: 10.1126/science.1091109. [DOI] [PubMed] [Google Scholar]
- 177.Liu F, Quesada V, Crevillén P, Bäurle I, Swiezewski S, Dean C. The Arabidopsis RNA-binding protein FCA requires a lysine-specific demethylase 1 homolog to downregulate FLC. Mol Cell. 2007;28:398–407. doi: 10.1016/j.molcel.2007.10.018. [DOI] [PubMed] [Google Scholar]
- 178.Macknight R, Bancroft I, Page T, Lister C, Schmidt R, Love K, Westphal L, Murphy G, Sherson S, Cobbett C, Dean C. FCA, a gene controlling flowering time in Arabidopsis, encodes a protein containing RNA-binding domains. Cell. 1997;89:737–745. doi: 10.1016/s0092-8674(00)80256-1. [DOI] [PubMed] [Google Scholar]
- 179.Quesada V, Macknight R, Dean C, Simpson G. Autoregulation of FCA pre-mRNA processing controls Arabidopsis flowering time. EMBO J. 2003;22:3142–3152. doi: 10.1093/emboj/cdg305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Ausín I, Alonso-Blanco C, Jarillo J, Ruiz-García L, Martínez-Zapater J. Regulation of flowering time by FVE, a retinoblastoma-associated protein. Nat Genet. 2004;36:162–166. doi: 10.1038/ng1295. [DOI] [PubMed] [Google Scholar]
- 181.Schomburg F, Patton D, Meinke D, Amasino R. FPA, a gene involved in floral induction in Arabidopsis, encodes a protein containing RNA-recognition motifs. Plant Cell. 2001;13:1427–1436. doi: 10.1105/tpc.13.6.1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Bäurle I, Smith L, Baulcombe D, Dean C. Widespread role for the flowering-time regulators FCA and FPA in RNA-mediated chromatin silencing. Science. 2007;318:109–112. doi: 10.1126/science.1146565. [DOI] [PubMed] [Google Scholar]
- 183.Hornyik C, Terzi LC, Simpson GG. The spen family protein FPA controls alternative cleavage and polyadenylation of RNA. Dev Cell. 2010;18(2):203–213. doi: 10.1016/j.devcel.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 184.Simpson GG, Dijkwel PP, Quesada V, Henderson I, Dean C. FY is an RNA 3′ end-processing factor that interacts with FCA to control the Arabidopsis floral transition. Cell. 2003;113(6):777–787. doi: 10.1016/s0092-8674(03)00425-2. [DOI] [PubMed] [Google Scholar]
- 185.Adams S, Allen T, Whitelam GC. Interaction between the light quality and flowering time pathways in Arabidopsis . Plant J. 2009;60(2):257–267. doi: 10.1111/j.1365-313X.2009.03962.x. [DOI] [PubMed] [Google Scholar]
- 186.Lim MH, Kim J, Kim YS, Chung KS, Seo YH, Lee I, Hong CB, Kim HJ, Park CM. A new Arabidopsis gene, FLK, encodes an RNA binding protein with K homology motifs and regulates flowering time via FLOWERING LOCUS C. Plant Cell. 2004;16(3):731–740. doi: 10.1105/tpc.019331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Aukerman M, Lee I, Weigel D, Amasino R. The Arabidopsis flowering-time gene LUMINIDEPENDENS is expressed primarily in regions of cell proliferation and encodes a nuclear protein that regulates LEAFY expression. Plant J. 1999;18:195–203. doi: 10.1046/j.1365-313x.1999.00442.x. [DOI] [PubMed] [Google Scholar]
- 188.Kim S, Choi K, Park C, Hwang HJ, Lee I. SUPPRESSOR OF FRIGIDA4, encoding a C2H2-type zinc finger protein, represses flowering by transcriptional activation of Arabidopsis FLOWERING LOCUS C. Plant Cell. 2006;18(11):2985–2998. doi: 10.1105/tpc.106.045179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Simpson G, Dean C. Arabidopsis, the Rosetta stone of flowering time? Science. 2002;296:285–289. doi: 10.1126/science.296.5566.285. [DOI] [PubMed] [Google Scholar]
- 190.Kim S, Soltis PS, Wall K, Soltis DE. Phylogeny and domain evolution in the APETALA2-like gene family. Mol Biol Evol. 2006;23(1):107–120. doi: 10.1093/molbev/msj014. [DOI] [PubMed] [Google Scholar]
- 191.Mathieu J, Yant LJ, Murdter F, Küttner F, Schmid M. Repression of flowering by the miR172 target SMZ. PLoS Biol. 2009;7(7):e1000148. doi: 10.1371/journal.pbio.1000148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Yant L, Mathieu J, Dinh T, Ott F, Lanz C, Wollmann H, Chen X, Schmid M. Orchestration of the floral transition and floral development in Arabidopsis by the bifunctional transcription factor APETALA2. Plant Cell. 2010;22:2156–2170. doi: 10.1105/tpc.110.075606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Castillejo C, Pelaz S. The balance between CONSTANS and TEMPRANILLO activities determines FT expression to trigger flowering. Curr Biol. 2008;18(17):1338–1343. doi: 10.1016/j.cub.2008.07.075. [DOI] [PubMed] [Google Scholar]
- 194.Lee J, Oh M, Park H, Lee I. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates leafy. Plant J. 2008;55(5):832–843. doi: 10.1111/j.1365-313X.2008.03552.x. [DOI] [PubMed] [Google Scholar]
- 195.Weigel D, Alvarez J, Smyth DR, Yanofsky MF, Meyerowitz EM. LEAFY controls floral meristem identity in Arabidopsis . Cell. 1992;69(5):843–859. doi: 10.1016/0092-8674(92)90295-n. [DOI] [PubMed] [Google Scholar]
- 196.Weigel D, Nilsson O. A developmental switch sufficient for flower initiation in diverse plants. Nature. 1995;377(6549):495–500. doi: 10.1038/377495a0. [DOI] [PubMed] [Google Scholar]
- 197.Yu H, Ito T, Wellmer F, Meyerowitz E. Repression of AGAMOUS-LIKE 24 is a crucial step in promoting flower development. Nat Genet. 2004;36:157–161. doi: 10.1038/ng1286. [DOI] [PubMed] [Google Scholar]
- 198.Liu C, Xi W, Shen L, Tan C, Yu H. Regulation of floral patterning by flowering time genes. Dev Cell. 2009;16(5):711–722. doi: 10.1016/j.devcel.2009.03.011. [DOI] [PubMed] [Google Scholar]
- 199.Wagner D. Flower morphogenesis: timing is key. Dev Cell. 2009;16(5):621–622. doi: 10.1016/j.devcel.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 200.Liu Z, Mara C. Regulatory mechanisms for floral homeotic gene expression. Semin Cell Dev Biol. 2010;21:80–86. doi: 10.1016/j.semcdb.2009.11.012. [DOI] [PubMed] [Google Scholar]
- 201.Moyroud E, Kusters E, Monniaux M, Koes R, Parcy F. LEAFY blossoms. Trends Plant Sci. 2010;15(6):346–352. doi: 10.1016/j.tplants.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 202.Gregis V, Sessa A, Colombo L, Kater MM. AGAMOUS-LIKE24 and SHORT VEGETATIVE PHASE determine floral meristem identity in Arabidopsis . Plant J. 2008;56(6):891–902. doi: 10.1111/j.1365-313X.2008.03648.x. [DOI] [PubMed] [Google Scholar]
- 203.Kaufmann K, Wellmer F, Muiño J, Ferrier T, Wuest S, Kumar V, Serrano-Mislata A, Madueño F, Krajewski P, Meyerowitz E, Angenent G, Riechmann J. Orchestration of floral initiation by APETALA1. Science. 2010;328:85–89. doi: 10.1126/science.1185244. [DOI] [PubMed] [Google Scholar]
- 204.Michaels SD, Ditta G, Gustafson-Brown C, Pelaz S, Yanofsky M, Amasino RM. AGL24 acts as a promoter of flowering in Arabidopsis and is positively regulated by vernalization. Plant J. 2003;33(5):867–874. doi: 10.1046/j.1365-313x.2003.01671.x. [DOI] [PubMed] [Google Scholar]
- 205.Yant L, Mathieu J, Schmid M. Just say no: floral repressors help Arabidopsis bide the time. Curr Opin Plant Biol. 2009;12(5):580–586. doi: 10.1016/j.pbi.2009.07.006. [DOI] [PubMed] [Google Scholar]
- 206.Nelson DC, Lasswell J, Rogg LE, Cohen MA, Bartel B. FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis . Cell. 2000;101(3):331–340. doi: 10.1016/s0092-8674(00)80842-9. [DOI] [PubMed] [Google Scholar]
- 207.Lee I, Michaels SD, Masshardt AS RMA. The late-flowering phenotype of FRIGIDA and LUMINIDEPENDENS is suppressed in the Landsberg erecta strain of Arabidopsis . Plant J. 1994;6:903–909. [Google Scholar]
- 208.Scortecci KC, Michaels SD, Amasino RM. Identification of a MADS-box gene, FLOWERING LOCUS M, that represses flowering. Plant J. 2001;26(2):229–236. doi: 10.1046/j.1365-313x.2001.01024.x. [DOI] [PubMed] [Google Scholar]
- 209.Kim HJ, Hyun Y, Park JY, Park MJ, Park MK, Kim MD, Lee MH, Moon J, Lee I, Kim J. A genetic link between cold responses and flowering time through FVE in Arabidopsis thaliana . Nat Genet. 2004;36(2):167–171. doi: 10.1038/ng1298. [DOI] [PubMed] [Google Scholar]
- 210.Henderson IR, Liu F, Drea S, Simpson GG, Dean C. An allelic series reveals essential roles for FY in plant development in addition to flowering-time control. Development. 2005;132(16):3597–3607. doi: 10.1242/dev.01924. [DOI] [PubMed] [Google Scholar]
- 211.Koornneef M, Rolff E, Spruit CJP. Genetic-control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L.) Heynh. Z Pflanzenphysiol. 1980;100(2):147–160. [Google Scholar]
- 212.Rubio V, Deng XW. Plant science. Standing on the shoulders of GIGANTEA. Science. 2007;318(5848):206–207. doi: 10.1126/science.1150213. [DOI] [PubMed] [Google Scholar]
- 213.Gaudin V, Libault M, Pouteau S, Juul T, Zhao G, Lefebvre D, Grandjean O. Mutations in LIKE HETEROCHROMATIN PROTEIN 1 affect flowering time and plant architecture in Arabidopsis . Development. 2001;128(23):4847–4858. doi: 10.1242/dev.128.23.4847. [DOI] [PubMed] [Google Scholar]
- 214.Lee J, Lee I. Regulation and function of SOC1, a flowering pathway integrator. J Exp Bot. 2010;61:2247–2254. doi: 10.1093/jxb/erq098. [DOI] [PubMed] [Google Scholar]
- 215.Wu G, Park MY, Conway SR, Wang JW, Weigel D, Poethig RS. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis . Cell. 2009;138(4):750–759. doi: 10.1016/j.cell.2009.06.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Yamaguchi A, Wu MF, Yang L, Wu G, Poethig RS, Wagner D. The microRNA-regulated SBP-Box transcription factor SPL3 is a direct upstream activator of LEAFY, FRUITFULL, and APETALA1. Dev Cell. 2009;17(2):268–278. doi: 10.1016/j.devcel.2009.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Tseng T, Swain S, Olszewski N. Ectopic expression of the tetratricopeptide repeat domain of SPINDLY causes defects in gibberellin response. Plant Physiol. 2001;126:1250–1258. doi: 10.1104/pp.126.3.1250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Blázquez M, Santos E, Flores C, Martínez-Zapater J, Salinas J, Gancedo C. Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding trehalose-6-phosphate synthase. Plant J. 1998;13:685–689. doi: 10.1046/j.1365-313x.1998.00063.x. [DOI] [PubMed] [Google Scholar]
- 219.Vogel G, Aeschbacher R, Müller J, Boller T, Wiemken A. Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: identification by functional complementation of the yeast tps2 mutant. Plant J. 1998;13:673–683. doi: 10.1046/j.1365-313x.1998.00064.x. [DOI] [PubMed] [Google Scholar]