

Abstract
Zinc (Zn) is an essential heavy metal that is incorporated into a number of human Zn metalloproteins. Zn plays important roles in nucleic acid metabolism, cell replication, and tissue repair and growth. Zn deficiency is associated with a range of pathological conditions, including impaired immunity, retarded growth, brain development disorders and delayed wound healing. Moreover, many reports have suggested that Zn is involved in cancer development and levels of Zn in serum and malignant tissues of patients with various types of cancer are abnormal. Zn may directly affect tumor cells by regulating gene expression profiles and/or cell viability, both of which are mediated in part by tumor‐induced changes in Zn transporter expression. On the other hand, Zn may indirectly influence tumor cells by affecting processes within the cancer microenvironment, including immune responses; the functions and/or activity levels of immune cells that attack tumor cells are influenced by the intracellular Zn concentrations within those cells. In both cases, Zn contributes to intracellular metal homeostasis and/or signal transduction in tumor and immune cells. In this review article, we will summarize the current understanding of the roles of Zn homeostasis and signaling primarily in immune cells, with a discussion of the contributions of these processes to oncogenesis. (Cancer Sci 2008; 99: 1515–1522)
The first discussion of the critical roles of zinc (Zn) in an organism was its reported requirement for the growth of Aspergillus niger.( 1 ) It has since been shown that Zn deficiency results in impaired growth, loss of hair, thickening and hyperkeratinization of the epidermis, and testicular atrophy in humans.( 2 ) Fujii suggested that Zn is involved in mitosis due to its presence in nucleoli during cell division.( 3 , 4 ) Prasad published a description of a syndrome that included iron‐deficiency anemia, hepatosplenomegaly hypogonadism, dwarfism and geophagia, speculating that Zn deficiency may cause growth retardation and hypogonadism.( 5 ) It has been estimated that more than 2 × 109 people have a nutritional deficiency for Zn in developing countries, which can result in growth retardation, immune dysfunction and cognitive impairment.( 6 ) These effects are reversible with Zn supplementation. Conditioned Zn deficiencies are also known to occur in many diseases and abnormal conditions, including malabsorption syndrome, chronic liver and renal diseases, sickle cell disease, excessive intake of alcohol, malignancies and other chronic debilitating conditions.( 6 )
Zinc is an essential trace element and a essential structural component of a great number of proteins, including intracellular signaling enzymes and transcription factors.( 7 , 8 ) In fact, Zn is required for the activity of more than 300 enzymes; as such, it participates in many enzymatic and metabolic functions in the body. More than 2000 transcription factors that work to regulate gene expression require Zn to maintain their structural integrity and bind to DNA.( 6 ) Thus, it is not surprising that the intracellular Zn concentration is tightly controlled by Zn transporters, Zn‐binding molecules and Zn sensors.( 9 , 10 , 11 , 12 , 13 , 14 ) Interestingly, a number of studies have found that the expression levels of Zn transporters in human tumors correlate with their malignancy, suggesting that alteration of intracellular Zn homeostasis can contribute to the severity of cancer.( 9 , 15 , 16 )
How do altered Zn levels affect the behaviors of the cells, which in turn result in localized and/or systemic abnormalities? Zn is critical for the activities of various enzymes that contribute to cellular signaling pathways as well as transcription factors. Zn is believed to maintain the structures of and/or serve as a cofactor in these proteins by binding tightly to various Zn‐binding motifs, including Zn finger (ZnF), ring finger, and LIM domains.( 7 , 8 ) Moreover, we have recently shown that, in addition to its structural role, Zn may also function as an intracellular signaling molecule.
Physiological and pathological roles of Zn homeostasis, transporters and metallothioneins
Phenotypic expression of the rare autosomal recessive disorder, acrodermatitis enteropathica (AE), was found to be due to defects in Zn metabolism.( 17 ) Mutations in ZIP4 were found to be causative of AE. ZIP4 encodes a member of the Zrt‐Irt‐like protein (ZIP)/SLC39 Zn transporter family that is expressed in intestinal organs, indicating that the intestinal absorption of Zn is a critical process.( 18 ) Various organs are affected by severe Zn deficiency, including the epidermal, gastrointestinal, central nervous, immune, skeletal and reproductive systems.( 19 ) Furthermore, the ZnT4 Zn transporter, which is expressed in breast epithelial cells, is responsible for the inherited Zn deficiency observed in lm mice, indicating that ZnT4‐mediated supplies of Zn in breast milk are critical for infant development.( 20 ) Moreover, studies of human breast‐fed infants have revealed a potential role for ZnT2 in the Zn content of milk.( 21 ) These findings indicate that transporter regulation of Zn homeostasis plays important cellular roles, disruption of which may induce disease states. The intracellular Zn concentration is tightly controlled by Zn importers (ZIP/SLC39),( 9 ) exporters (ZNT/SLC30),( 10 ) and binding proteins such as metallothioneins (Fig. 1).( 11 ) In addition, Zn‐sensing molecules, such as metal‐responsive element‐binding transcription factor‐1 (MTF‐1), regulate the expression of these molecules.( 12 )
Figure 1.
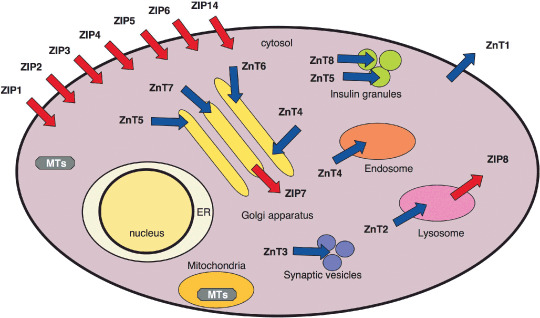
Subcellular localization of zinc (Zn) transporters. Subcellular localization and potential function of members of the ZIP and Zn transporter (ZnT) families are shown based on currently available information.( 92 , 98 , 99 , 100 , 101 , 102 , 103 , 104 ) Predicted directions of Zn mobilization are indicated with arrows. ER, endoplasmic reticulum; MT, metallothionein; MTF‐1, metal‐responsive element‐binding transcription factor‐1.
Fourteen members of the ZIP family, which were first discovered in Saccharomyces cerevisiae (Zrt proteins) and Arabidopsis thaliana (Irt proteins),( 15 , 16 ) have been reported in mammals, and some knockout mouse lines deficient for various ZIP proteins have been reported (Table 1). Mice lacking ZIP1, ZIP2 or ZIP3 show abnormal embryogenesis specifically under Zn‐limiting conditions. Homozygous ZIP4 knockout mouse embryos die during early development, whereas heterozygosity causes hypersensitivity to Zn deficiencies, as is observed in AE patients.( 18 , 22 ) ZIP6/Liv1 from zebrafish has been reported to control the epithelial–mesenchymal transition (EMT) following STAT3 activation,( 23 ) and the Drosophila gene fear of intimacy (foi), which is similar to mammalian ZIP6 and ZIP10, is essential for proper gonad formation, E‐cadherin expression and glial cell migration,( 24 , 25 , 26 ) suggesting that ZIP6/Liv1 and/or ZIP10 have important roles in cell migration. In fact, it has been suggested that ZIP10 is involved in the invasive behavior of breast cancer cells.( 27 ) We will return to this matter in the last section of this review. ZIP7 (KE4) was discovered during the characterization of the major histocompatibility complex (MHC) region on mouse chromosome 17,( 28 ) and mapped to the human leukocyte antigen (HLA) class II region on human chromosome 6.( 29 ) A Drosophila counterpart of ZIP7, Catsup, is reported to control melanin synthesis,( 30 ) whereas the plant homolog IAR1 regulates root elongation by controlling auxin conjugate sensitivity.( 31 )
Table 1.
Mutations of Zn transporters and methallothioneins
| Gene name | Protein name | Type | Phenotype | Drosophila melanogaster Caenorhabditis elegance | Reference |
|---|---|---|---|---|---|
| MT‐I MT‐II | MT‐I MT‐II | KO | Dysfunction of macrophage | Sugiura and Kuroda( 36 ) | |
| SLC39A1 | ZIP1 | KO | Abnormal morphogenesis of the embryo in zinc‐limiting condition | Dufner‐Beattie et al. ( 37 ) | |
| SLC39A2 | ZIP2 | KO | Abnormal morphogenesis of the embryo in zinc‐limiting condition | Peters et al. ( 38 ) | |
| SLC39A3 | ZIP3 | KO | Abnormal morphogenesis of the embryo and depletion of thymic pre‐T cells in zinc‐limiting condition | Dufner‐Beattie et al. ( 39 ) | |
| SLC39A4 | ZIP4 | AE | Die in utero during early development | Dufner‐Beattie et al. ( 40 ) | |
| hSLC39A4 | hZIP4 | Severe zinc deficiency | |||
| SLC39A6/10 | ZIP6/10 | – | FOI/defects in the formation of the embryoni ganad | Van Doren et al. ( 26 ) | |
| SLC39A7 | ZIP7(KE4) | – | Catsup/semi‐dominant lethal | Stathakis et al. ( 30 ) | |
| SLC30A1 | ZnT1 | KO | Embryonic lethal | Cdf‐1/died during larval development | Andrews et al. ( 41 ) Bruinsma( 22 ) |
| hSLC30A2 | hZnT2 | Self‐limiting AE | Produce zinc‐deficient milk in some women | Chowanadisai( 21 ) | |
| SLC30A3 | ZnT3 | KO | Prone to seizures elicited by kainic acid treatment | Cole et al. ( 42 ) | |
| SLC30A4 | ZnT4 | lm | Produce zinc‐deficient milk | Huang and Gitschier( 20 ) | |
| SLC30A5 | ZnT5 | KO | Poor growth; osteopenia; low bodyfat; muscle weakness; male‐specific cardiac death | Inoue et al. ( 43 ) |
AE, acrodermatitis enteropathica; KO, knockout; lm, lethal milk.
Cation diffusion facilitator (CDF) confers metal resistance to many eukaryotic cell types;( 32 ) 10 reported members of this protein family have been reported in mammals (named the Zn transporter [ZnT] family), some of which have been targeted in knockout mouse lines (Table 1). ZnT1 knockout mice are embryonic lethal. CDF1, a nematode ZnT1 ortholog, positively regulates Ras–Raf–MEK–ERK signal transduction by promoting Zn efflux and reducing the concentration of cytosolic Zn.( 33 ) ZnT3 knockout mice are prone to seize in response to kainic acid treatment. lm mice carry a nonsense mutation in the ZnT4 gene and produce Zn‐deficient milk. ZnT5 knockout mice show poor growth, osteopenia, low bodyfat, muscle weakness and male‐specific cardiac death. A point mutation in the human ZnT2 gene suggests that ZnT2 functions to enhance the Zn content of milk. A recent genome‐wide association study identified the region containing ZnT8 as a risk locus for type 2 diabetes.( 34 ) Interestingly, ZnT8 is expressed exclusively in pancreatic β cells.
Metallothioneins (MTs) – small cystein‐rich proteins that bind Zn as well as other metal ions – are thought to be responsible for regulating the intracellular Zn concentration and for nonessential heavy metal detoxification. When the concentration of intracellular free Zn reaches a threshold, activation of MTF‐1 induces the expression of MT, which then sequester the Zn ions.( 35 ) Therefore, MT serve as biochemical devices that control the concentration of free Zn by sequestering Zn and releasing it in response to other biochemical events, such as oxidative signaling. MT also participate in immune responses; macrophages from MT‐KO (MT‐I and MT‐II double knockout) mice show defects in phagocytosis and antigen presentation.( 36 )
Roles of Zn in immune responses
Because Zn deficiency is associated with many chronic diseases, experiments examining Zn homeostasis have employed mouse models to determine the impact of nutritional deficiency of a single element on immune function at the cellular and molecular levels.( 44 ) It is important to point out that chronic diseases, such as gastrointestinal disorders, renal disease, sickle cell anemia, cirrhosis, some cancers, cystic fibrosis, pancreatic insufficiency and autoimmune arthritis, have been shown to lead to suboptimal Zn status in humans.( 45 , 46 , 47 , 48 , 49 , 50 , 51 , 52 ) These disease states are associated with increased infections of prolonged duration, a clear indication of compromised immunity, which implies that Zn may contribute to immune cell homeostasis in vivo.( 47 , 50 ) Indeed, various studies using animal models of Zn deficiency have confirmed that decreased levels of Zn induce thymic atrophy, lymphopenia, and compromised cell‐ and antibody‐mediated immune responses. In this section, we will comment on the relationship between Zn and immunity, primarily through an examination of adaptive immunity including T cells and dendritic cells. Moreover, because immune responses against tumors may be affected by Zn deficiency, Zn may have indirect effects on tumorigenesis.
Zinc deficiency, which affects some patients with carcinomas, is reported to induce thymic atrophy. T cells, a critical antitumor population, develop in the thymus. Moreover, glucocorticoids, in particular corticosterone, are chronically elevated in Zn‐deficient mice and adrenalectomies or removal of these steroids prevent the thymus from atrophying under Zn‐deficient conditions.( 53 , 54 ) Additionally, Zn‐deficient diets cause substantial reductions in the number of CD4+CD8+ thymocytes, thymic cells known to show high rates of apoptosis in response to glucocorticoids.( 55 ) Thus, chronic overproduction of glucocorticoids in Zn‐deficient individuals may accelerate the rate of apoptosis in the thymus, which would reduce the number of peripheral T cells. On the other hand, mature CD4+ and CD8+ T cells are resistant to Zn deficiency and survive well in an otherwise atrophying thymus.( 55 ) Interestingly, in an experimental mouse model, Zn deficiency caused an imbalance between the peripheral functions of the T‐helper (Th)1 and Th2 cell populations; production of γ‐interferon (IFN‐γ) and interleukin (IL)‐2 (products of Th1 cells) decreased, whereas production of IL‐4, IL‐6 and IL‐10 (products of Th2 cells) were not affected.( 56 ) Of note, Th1 cells are known to play major roles in tumor suppression.
Another mechanism by which Zn deficiency contributes to impaired immunity, including peripheral lymphopenia and compromised immune responses, is altered gene expression profiles in various cell types including T cells. Indeed, mice with modest Zn deficiencies show changes in the expression levels of 1200 genes in their T cells.( 57 ) Altered gene expression may affect the survival and/or responses of T cells and dendritic cells, important cellular mediators of T‐cell homeostasis in vivo. In fact, culturing Th0 and Th1 cell lines in low‐Zn media followed by mitogenic stimulation leads to reduced expression of IL‐2 and IFN‐γ mRNA, which adversely affects the functional capacity of these cells.( 58 ) In addition to T cells, other types of immune cells show specific responses to Zn deficiency. HL‐60 cells, a human myeloid‐like precursor cell line, survive in cultures containing low concentrations of Zn.( 58 ) Interestingly, exposure of mouse dendritic cells to the bacterial endotoxin lipopolysaccharide (LPS), a toll‐like receptor (TLR) ligand, leads to a decrease in the intracellular free Zn concentration (Fig. 2). Moreover, artificially depleting the intracellular Zn using a Zn chelator triggers dendritic cell maturation. On the other hand, artificially elevating intracellular Zn levels suppresses the ability of dendritic cells to respond to LPS. In fact, Zn appears to suppress the surface expression of MHC class II molecules because Zn is required for the endocytosis of MHC class II molecules expressed on the plasma membrane, and Zn inhibits MHC class II vesicle trafficking to the plasma membrane from the perinuclear region. LPS affects the expression of a number of Zn import and export transporter molecules, resulting in a net increase in Zn transport out of cells. Importantly, overexpression of the Zn importer ZIP6, expression of which is reduced by LPS stimulation, suppresses dendritic cell maturation followed by inhibition of the stimulatory activities of CD4+ T cells. A similar effect of Zn has been observed in vivo; injections of LPS induce reduced intracellular free Zn levels and ZIP6 expression in dendritic cells and treatment with Zn‐depleting agents leads to increased dendritic cell maturation.( 59 ) These results clearly show that intracellular Zn homeostasis is critically involved in the maturation of dendritic cells, an important step for T‐cell activation (Fig. 3). In any case, intracellular Zn homeostasis in immune cells, which may be affected by tumor progression, plays a key role in responses of these cells, including those against tumors.
Figure 2.
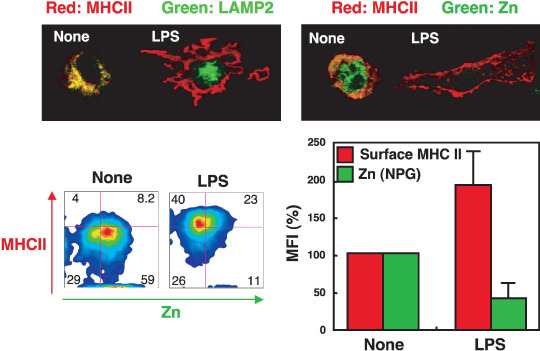
Decreased levels of intracellular free zinc (Zn) in dendritic cells are critical for lipopolysaccharide (LPS)‐mediated CD4+ T‐cell activation. LPS, a component of the Gram‐negative bacterial cell wall, transduces a signal that activates dendritic cells (DC). LPS binds to the TLR4 receptor on DC, resulting in MYD88‐ and TRIF‐mediated signaling. We showed that TRIF‐mediated signaling via TLR4 increased the level of Zn transporter (ZnT) family (Zn exporters) mRNA but decreased the mRNA expression of members of the Zip family (Zn importers), resulting in decreased levels of intracellular free Zn in DC. We also demonstrated that the reduction of the intracellular free Zn concentration in DC is critical for the surface expression of major histocompatibility complex (MHC) class II molecules, which are important for the activation of CD4+ T cells.
Figure 3.
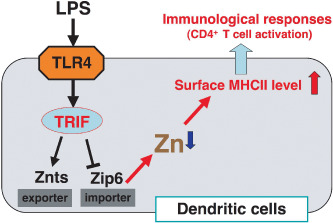
Summary of the roles of zinc (Zn) in dendritic cells. LPS, lipopolysaccharide; MHC, major histocompatibility complex.
Zinc signaling
As discussed above, Zn is known to be a critical structural constituent of a great number of proteins, including enzymes from cellular signaling pathways and transcription factors.( 7 , 8 ) In addition, Zn may also function as a signaling molecule. Microfluorescence imaging of Zn dynamics following presynaptic stimulation of hippocampal mossy fibers shows Zn release from terminal vesicles into the surrounding milieu.( 60 , 61 , 62 ) The Zn is then taken up into the cytoplasm of neighboring cells through gated Zn channels. Rapid Zn influx through Ca2+ permeable AMPA/kainate (Ca‐A/K) channels triggers the generation of reactive oxygen species, which are potently neurotoxic.( 63 ) As such, Zn functions similarly to neurotransmitters, which are stored in membrane‐enclosed synaptic vesicles and released by exocytosis to bind transmitter‐gated ion channels and activate postsynaptic cells.( 60 , 64 , 65 , 66 ) Using site‐directed mutagenesis, Hosie et al. identified a pair of Zn‐binding sites and completed the characterization of a third Zn‐binding site in the GABA receptor.( 67 ) Hirzel et al. used knockin mice carrying a D80A mutation (constructed in a Zn‐binding site) in the glycine receptor (GlyR) a1 subunit gene (Glra1) to show that Zn modulates neurotransmission.( 68 ) In the latter report, the authors showed that the hyperekplexia phenotype of the Glra1(D80A) mice was due to the loss of Zn‐mediated potentiation of a1 subunit‐containing GlyR. Therefore, synaptic Zn is essential for proper functioning of glycinergic neurotransmission in vivo. Zn is also known to inhibit NMDA receptor activity via two mechanisms: voltage‐dependent channel blockade and voltage‐independent reduction in the probability of channel opening.( 69 , 70 , 71 ) Huang et al. ( 72 ) recently reported that Zn‐mediated transactivation of TrkB potentiates hippocampal mossy fiber‐CA3 pyramid synapses.
Thus, it is likely that Zn acts as a neurotransmitter. The role of Zn as a neurotransmitter differs from the conventional concept of a secondary messenger in cells; neurotransmitters carry information between cells, whereas secondary messengers function intracellularly. Cyclic adenosine monophosphate (cAMP) was discovered in 1957 as the first intracellular secondary messenger. A limited number of secondary messenger species have now been identified, including Ca2+, cAMP, cyclic guanylic acid (cGMP), NO, lipid mediators, G‐proteins, protein kinases, protein phosphatases and nuclear receptors.( 73 ) We hypothesize that Zn acts as a secondary messenger as well. We have shown that the STAT3‐ZIP6 signaling cascade is critically involved in the EMT of zebrafish cells and is required for the nuclear localization of Snail, a ZnF‐containing repressor (Fig. 4).( 23 ) Moreover the nuclear localization of Snail is dependent on both its ZnF domain( 23 , 74 ) and the Zn transporter ZIP6, suggesting that Zn may act as an intracellular signaling molecule. It is also known that Zn regulates cyclic nucleotide signaling, a conventional secondary messenger system. Zn suppresses LPS‐induced tumor necrosis factor (TNF)‐α and IL‐1β release from monocytes. Moreover, this inhibitory effect is dependent on suppression of phosphodiesterase‐mediated hydrolyzation of cyclic nucleotides into 5′‐nucleotide monophosphate followed by an increase of the intracellular cGMP level. Because the NO donor S‐nitrosocystein (SNOC) also inhibits LPS‐induced TNF‐α and IL‐1β release, it is possible that increased levels of intracellular free Zn in response to SNOC play a role in this inhibition via augmentation of the cGMP level.( 75 ) Because cytokines can induce NO production to enhance the level of intracellular free Zn,( 76 ) Zn may act as a bridge between secondary signaling mediated by NO and cGMP. Additionally, Huberman's group showed in macrophage cell lines that the nuclear Zn concentration increases within 15 min following treatment with phorbol myristate acetate in a manner dependent on PKCβ.( 77 ) Interestingly, we have shown that TLR‐mediated signaling induces a decrease in the intracellular free Zn concentration in dendritic cells, and this decrease is required for dendritic cell activation and subsequent CD4+ T‐cell activation.( 59 ) LPS‐induced decreases in the concentration of intracellular free Zn are dependent on changes in the expression profiles of Zn transporters; expression of ZIP family members are downregulated, whereas those of the ZnT transporters are upregulated. The results show that extracellular stimuli affect intracellular free Zn concentrations and these changes are critically involved in the biological expression of extracellular stimuli. Taken together, these data support a role for Zn as an intracellular signaling molecule. We have also shown that extracellular stimulation of mast cells induces an increase in intracellular free Zn levels within minutes of the stimulation, a phenomenon that we have named the ‘Zn wave’.( 78 ) The Zn wave originates from the endoplasmic reticulum and/or the surrounding area. In mast cells, the Zn wave is dependent on Ca2+ influx and mitogen‐activated protein kinase activation (Fig. 5). Because extracellular Zn does not contribute to the Zn wave and the wave is induced several minutes after FcɛR1‐stimulation, Zn seems to be functioning as an intracellular secondary messenger under these conditions. This conclusion is supported by the following results: (i) extracellular stimuli, such those that cross‐link FcɛRI receptors, directly induce increases in the Zn wave; (ii) an intracellular compartment, possibly the endoplasmic reticulum compartment, serves as the source of Zn; and (iii) free Zn at a level similar to that observed in the Zn wave can affect intracellular signaling molecules, such as tyrosine phosphatases, and therefore may modulate the final output triggered by extracellular stimuli.
Figure 4.
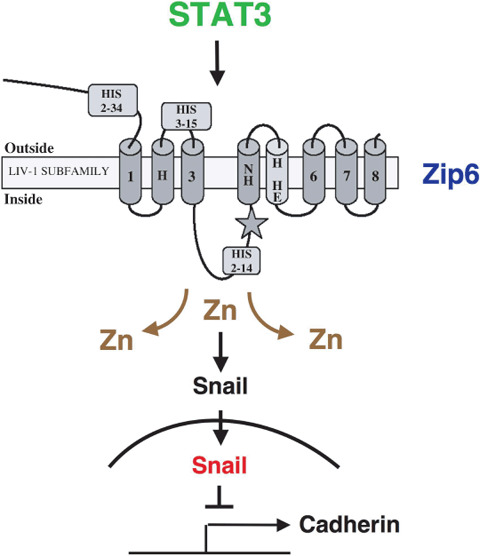
STAT3‐induced movement of organizer cells is dependent on ZIP6 expression. The STAT3–ZIP6 signaling cascade is critically involved in the epithelial–mesenchymal transition of the organizer cells, which is required for the nuclear localization of Snail, a Zn finger (ZnF)‐containing repressor of cadherin expression.
Figure 5.
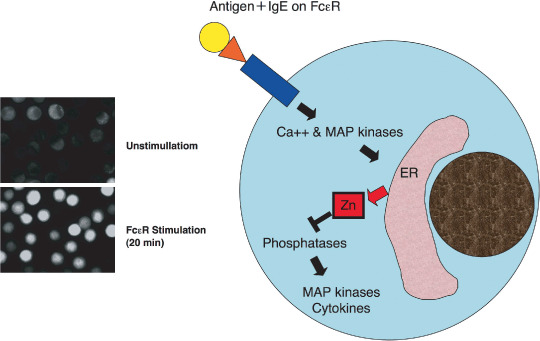
A zinc (Zn) wave is induced by FceRI‐mediated mast cell activation. FceRI stimulation via antigen plus immunoglobulin (Ig)E induces mast cell activation, including degranulation and cytokine production. Intracellular Zn levels, which can be detected using FluoZin‐3, increased within 20 min after FceRI stimulation. We have named this increase the ‘Zn wave’ and showed that these waves are dependent on Ca2+ influx and mitogen‐activated protein (MAP) kinase activation via FceRI stimulation. The Zn wave appears to originate in the endoplasmic reticulum (ER) and suppresses phosphatase activity, which is followed by MAP kinase activation and cytokine expression.
An important difference between the Zn wave in mast cells described above and other findings in zebrafish cells( 23 ) and dendritic cells( 59 ) is that the Zn wave is observed several minutes after stimulation, whereas the latter observations are critically dependent on transcriptional regulation of Zn transporters and are therefore detected several hours after stimulation. Thus, it appears that intracellular Zn signaling comprises an early component, such as the Zn wave, which is directly induced by extracellular stimuli, and a late component that is dependent on transcriptional regulation of transporter expression (Fig. 6). For the former, Zn acts as a conventional intracellular secondary messenger capable of transducing an extracellular stimulus into intracellular events. Collectively, these results support the idea that Zn is a secondary messenger/signaling ion that has the potential to influence many aspects of cellular signaling through its effects on a range of Zn‐binding proteins.
Figure 6.
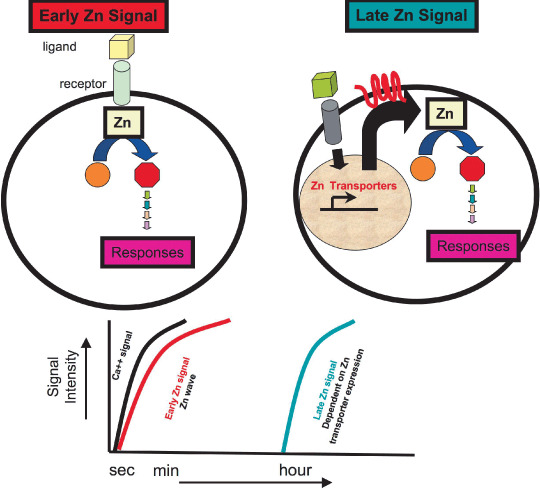
Early and late zinc (Zn) signaling. Intracellular Zn signaling can be classified as early signaling, such as the Zn wave, which is directly induced by extracellular stimuli, and late signaling, which is dependent on transcriptional regulation of Zn transporter expression.
Zinc and tumorigenesis
In this section, we discuss the cellular effects of altered intracellular Zn homeostasis. As described in the previous sections, the intracellular Zn concentration is tightly controlled by Zn transporters, Zn‐binding molecules and Zn‐sensing molecules.( 9 , 10 , 11 , 12 , 13 , 14 ) It has been pointed out, however, that the levels of Zn in sera and malignant tissues are abnormal in patients with various inflammatory diseases and tumors; serum Zn concentrations are lower in patients with such autoimmune diseases as rheumatoid arthritis.( 79 ) The relationship between tumor development and Zn levels, however, appears to be complicated. Indeed, Zn levels are reduced in patients suffering from carcinomas of the liver, gallbladder, digestive tract or prostate,( 80 , 81 , 82 ) whereas breast cancer patients show decreased and elevated Zn levels in sera and malignant tissues, respectively.( 82 , 83 , 84 )
For immune cells, Zn‐deficient conditions result in reduced natural killer cell‐mediated cytotoxic activity, antibody‐mediated responses, and host defense against pathogens and tumors,( 85 , 86 , 87 ) whereas excessive levels of Zn are cytotoxic; Zn induces apoptosis in lymphocytes, including T and B cells.( 88 , 89 ) Zn may have similar effects on tumor cells. Tumors need Zn to survive and grow, whereas excess Zn may induce tumor cell apoptosis, although the sensitivities of the different types of tumors are likely to vary. Indeed, some tumors are resistant to high Zn concentrations. In addition, Zn levels are affected by the microenvironment surrounding the cancerous tissue: (i) many cytokines and growth factors that are produced in these micrsoenvironments, including IL‐6, hepatocyte growth factor, epidermal growth factor and TNF‐α, directly or indirectly affect the expression profiles of various Zn transporters; (ii) mast cells, an important cell type in cancer microenvironments, contain high levels of Zn in granules that are released into the surround milieu; and (iii) oxidation/reduction reactions in these environments markedly influence the intracellular free Zn concentration. Moreover, it is likely that the activities of the diverse set of enzymes and transcription factors that require Zn to function, including matrix metalloproteases, are affected by the altered Zn concentrations in cancer microenvironments.
The relationships between the expression levels of Zn transporters, especially ZIP family members, and tumor malignancy have recently received a great deal of attention. There are at least 14 human ZIP transporters, which are believed to allow Zn influx into the cytosol.( 9 , 15 , 16 ) Some of these proteins are thought to be involved in cancer progression; ZIP1 is reported to be a suppressor of prostate cancer( 90 ) and some evidence suggests ZIP6 contributes to the metastasis of breast cancer to the lymph node( 91 , 92 ) although this issue is controversial.( 5 , 8 , 27 ) Interestingly, we showed that the STAT3‐ZIP6 signaling cascade is critically involved in the EMT of organizer cells and is required for the nuclear localization of Snail, a ZnF‐containing repressor.( 23 ) STAT3 is activated in the organizer of zebrafish to allow the movements of cells during gastrulation. The requirement for STAT3 is cell autonomous for the anterior migration of the organizer cells, and non‐cell autonomous for the convergence of neighboring cells.( 23 ) It is possible that ZIP6 acts as a downstream target of STAT3 to induce tumor cell migration followed by their metastasis. Moreover, we showed that ZIP6 is essential for the nuclear localization of the ZnF protein Snail, a master regulator of the EMT that suppresses the expression of cadherin.( 94 , 95 ) These results establish a molecular link between STAT3, ZIP6 and Snail during the EMT, which is not only important for embryonic development including the migration of the organizer cells but also for cancer metastasis.( 96 ) Indeed, the processes of embryonic development that rely on EMT for the generation of new tissue types have been reported to be co‐opted by tumors, resulting in uncontrolled proliferation and spatial expansion.( 7 , 81 ) We also demonstrated that the migratory activity of metastatic breast cancer cells was inhibited by knockdown of ZIP10 expression and Zn chelation. Importantly, analysis of clinical samples showed that breast cancers with lymph node metastases expressed significantly higher levels of the Zn transporter, ZIP10, than those without lymph node metastases.( 27 ) Furthermore, ZIP10 was recently shown to mediate Zn uptake and to act as a membrane transporter in vivo.( 97 ) Thus, altered intracellular Zn homeostasis due to changes in Zn transporter expression may be a key factor that determines tumor malignancy. Moreover, other types of proteins that regulate Zn levels, such as MT and MTF‐1, may contribute to tumor behavior. However, because systemic changes in Zn status will regulate the activities of immune cells, which in turn will affect tumors, serum Zn concentrations may not be a useful diagnostic marker clinically. Future studies should assess the possibility of using Zn levels to control tumor metastasis or tumor malignancy.
References
- 1. Raulin J. Chemical studies on vegetation. Annales des Sci Naturelles 1869; 11: 93–9. [Google Scholar]
- 2. Todd WR, Elvehjem CA, Hart EB. Zinc in the nutrition of the rat. Am J Physiol 1933; 107: 146–56. [Google Scholar]
- 3. Fujii T. Presence of zinc in nucleoli and its possible role in mitosis. Nature 1954; 174: 1108–9. [DOI] [PubMed] [Google Scholar]
- 4. Fujii T, Utida S, Mizuno T. Reaction of starfish spermatozoa to histidine and certain other substances considered in relation to zinc. Nature 1955; 176: 1068–9. 13272749 [Google Scholar]
- 5. Prasad AS, Halsted JA, Nadimi M. Syndrome of iron deficiency anemia, hepatosplenomegaly, hypogonadism, dwarfism and geophagia. Am J Med 1961; 31: 532–46. [DOI] [PubMed] [Google Scholar]
- 6. Brown H, Peerson JM, Allen LH, Rivera J. Effect of supplemental zinc on the growth and serum zinc concentrations of pre‐pubertal children: a meta‐analysis of randomized, controlled trials. Am J Clin Nutrition 2002; 75: 1062–71. [DOI] [PubMed] [Google Scholar]
- 7. Vallee BL, Auld DS. Cocatalytic zinc motifs in enzyme catalysis. Proc Natl Acad Sci USA 1993; 90: 2715–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Prasad AS. Zinc: an overview. Nutrition 1995; 11: 93–9. [PubMed] [Google Scholar]
- 9. Eide DJ. The SLC39 family of metal ion transporters. Pflugers Arch 2004; 447: 796–800. [DOI] [PubMed] [Google Scholar]
- 10. Palmiter RD, Huang L. Efflux and compartmentalization of zinc by members of the SLC30 family of solute carriers. Pflugers Arch 2004; 447: 744–51. [DOI] [PubMed] [Google Scholar]
- 11. Vallee BL. The function of metallothionein. Neurochem Int 1995; 27: 23–33. [DOI] [PubMed] [Google Scholar]
- 12. Andrews GK. Cellular zinc sensors: MTF‐1 regulation of gene expression. Biometals 2001; 14: 223–37. [DOI] [PubMed] [Google Scholar]
- 13. Palmiter RD. Protection against zinc toxicity by metallothionein and zinc transporter 1. Proc Natl Acad Sci USA 2004; 101: 4918–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Lichtlen P, Schaffner W. Putting its fingers on stressful situations: the heavy metal‐regulatory transcription factor MTF‐1. Bioessays 2001; 23: 1010–7. [DOI] [PubMed] [Google Scholar]
- 15. Zhao H, Eide D. The yeast ZRT1 gene encodes the zinc transporter protein of a high‐affinity uptake system induced by zinc limitation. Proc Natl Acad Sci USA 1996; 93: 2454–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Eide D, Broderius M, Fett J, Guerinot ML. A novel iron‐regulated metal transporter from plants identified by functional expression in yeast. Proc Natl Acad Sci USA 1996; 93: 5624–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Moynahan EJ. Letter: Acrodermatitis enteropathica: a lethal inherited human zinc‐deficiency disorder. Lancet 1974; 2: 399–400. [DOI] [PubMed] [Google Scholar]
- 18. Wang K, Zhou B, Kuo YM, Zemansky J, Gitschier J. A novel member of a zinc transporter family is defective in acrodermatitis enteropathica. Am J Hum Genet 2002; 71: 66–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Hambidge KM, Walravens PA. Disorders of mineral metabolism. Clin Gastroenterol 1982; 11: 87–117. [PubMed] [Google Scholar]
- 20. Huang L, Gitschier J. A novel gene involved in zinc transport is deficient in the lethal milk mouse. Nat Genet 1997; 17: 292–7. [DOI] [PubMed] [Google Scholar]
- 21. Chowanadisai W, Lonnerdal B, Kelleher SL. Identification of a mutation in SLC30A2 (ZnT‐2) in women with low milk zinc concentration that results in transient neonatal zinc deficiency. J Biol Chem 2006; 281: 39699–707. [DOI] [PubMed] [Google Scholar]
- 22. Küry S, Kharfi M, Kamoun R et al . Mutation spectrum of human SLC39A4 in a panel of patients with acrodermatitis enteropathica. Hum Mutat 2003; 22: 337–8. [DOI] [PubMed] [Google Scholar]
- 23. Yamashita S, Miyagi C, Fukada T, Kagara N, Che YS, Hirano T. Zinc transporter LIVI controls epithelial‐mesenchymal transition in zebrafish gastrula organizer. Nature 2004; 429: 298–302. [DOI] [PubMed] [Google Scholar]
- 24. Mathews WR, Ong D, Milutinovich AB, Van Doren M. Zinc transport activity of Fear of Intimacy is essential for proper gonad morphogenesis and DE‐cadherin expression. Development 2006; 133: 1143–53. [DOI] [PubMed] [Google Scholar]
- 25. Pielage J, Kippert A, Zhu M, Klämbt C. The Drosophila transmembrane protein Fear‐of‐intimacy controls glial cell migration. Dev Biol 2004; 275: 245–57. [DOI] [PubMed] [Google Scholar]
- 26. Van Doren M, Mathews WR, Samuels M, Moore LA, Broihier HT, Lehmann R. Fear of intimacy encodes a novel transmembrane protein required for gonad morphogenesis in Drosophila. Development 2003; 130: 2355–64. [DOI] [PubMed] [Google Scholar]
- 27. Kagara N, Tanaka N, Noguchi S, Hirano T. Zinc and its transporter ZIP10 are involved in invasive behavior of breast cancer cells. Cancer Sci 2007; 98: 692–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Lai JL, Preudhomme C, Zandecki M et al . Myelodysplastic syndromes and acute myeloid leukemia with 17p deletion. An entity characterized by specific dysgranulopoïesis and a high incidence of P53 mutations. Leukemia 1994; 9: 370–81. [PubMed] [Google Scholar]
- 29. Ando A, Kikuti YY, Shigenari A et al . cDNA cloning of the human homologues of the mouse Ke4 and Ke6 genes at the centromeric end of the human MHC region. Genomics 1996; 35: 600–2. [DOI] [PubMed] [Google Scholar]
- 30. Stathakis DG et al . The catecholamines up (Catsup) protein of Drosophila melanogaster functions as a negative regulator of tyrosine hydroxylase activity. Genetics 1999; 153: 361–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Lasswell J, Rogg LE, Nelson DC, Rongey C, Bartel B. Cloning and characterization of IAR1, a gene required for auxin conjugate sensitivity in Arabidopsis. Plant Cell 2000; 12: 2395–408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Palmiter RD, Findley SD. Cloning and functional characterization of a mammalian zinc transporter that confers resistance to zinc. Embo J 1995; 14: 639–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Bruinsma JJ, Jirakulaporn T, Muslin AJ, Kornfeld K. Zinc ions and cation diffusion facilitator proteins regulate Ras‐mediated signaling. Dev Cell 2002; 2: 567–78. [DOI] [PubMed] [Google Scholar]
- 34. Sladek R, Rocheleau G, Rung J et al . A genome‐wide association study identifies novel risk loci for type 2 diabetes. Nature 2007; 445: 881–5. [DOI] [PubMed] [Google Scholar]
- 35. Lichtlen P, Schaffner W. The ‘metal transcription factor’ MTF‐1: biological facts and medical implications. Swiss Med Wkly 2001; 131: 647–52. [DOI] [PubMed] [Google Scholar]
- 36. Sugiura T, Kuroda E, UY. Dysfunction of macrophages in metallothionein‐knock out mice. J UOEH 2004; 26: 193–205. [DOI] [PubMed] [Google Scholar]
- 37. Dufner‐Beattie J, Huang ZL, Geiser J, Xu W, Andrews GK. Mouse ZIP1 and ZIP3 genes together are essential for adaptation to dietary zinc deficiency during pregnancy. Genesis 2006; 44: 239–51. [DOI] [PubMed] [Google Scholar]
- 38. Peters JL, Dufner‐Beattie J, Xu W, Geiser J, Lahner B, Salt DE, Andrews GK. Targeting of the mouse Slc39a2 (Zip2) gene reveals highly cell‐specific patterns of expression, and unique functions in zinc, iron, and calcium homeostasis. Genesis 2007; 45: 339–52. [DOI] [PubMed] [Google Scholar]
- 39. Dufner‐Beattie J, Huang ZL, Geiser J, Xu W, Andrews GK. Generation and characterization of mice lacking the zinc uptake transporter ZIP3. Mol Cell Biol 2005; 25: 5607–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Dufner‐Beattie J, Weaver BP, Geiser J, Bilgen M, Larson M, Xu W, Andrews GK. The mouse acrodermatitis enteropathica gene Slc39a4 (Zip4) is essential for early development and heterozygosity causes hypersensitivity to zinc deficiency. Hum Mol Genet 2007; 16: 1391–9. [DOI] [PubMed] [Google Scholar]
- 41. Andrews GK, Wang H, Dey SK, Palmiter RD. Mouse zinc transporter 1 gene provides an essential function during early embryonic development. Genesis 2004; 40: 74–81. [DOI] [PubMed] [Google Scholar]
- 42. Cole TB, Robbins CA, Wenzel HJ, Schwartzkroin PA, Palmiter RD. Seizures and neuronal damage in mice lacking vesicular zinc. Epilepsy Res 2000; 39: 153–69. [DOI] [PubMed] [Google Scholar]
- 43. Inoue K, Matsuda K, Itoh M, Kawaguchi H, Tomoike H, Aoyagi T, Nagai R, Hori M, Nakamura Y, Tanaka T. Osteopenia and male‐specific sudden cardiac death in mice lacking a zinc transporter gene, Znt5. Hum Mol Genet 2002; 11: 1775–84. [DOI] [PubMed] [Google Scholar]
- 44. Hambidge M. Human zinc deficiency. J Nutr 2000; 130: 1344S–9S. [DOI] [PubMed] [Google Scholar]
- 45. Bhutta ZA, Bird SM, Black RE et al . Therapeutic effects of oral zinc in acute and persistent diarrhea in children in developing countries: pooled analysis of randomized controlled trials. Am J Clin Nutr 2000; 72: 1516–22. [DOI] [PubMed] [Google Scholar]
- 46. Dutta SK, Procaccino F, Aamodt R. Zinc metabolism in patients with exocrine pancreatic insufficiency. J Am Coll Nutr 1998; 17: 556–63. [DOI] [PubMed] [Google Scholar]
- 47. Fraker PJ, King LE, Laakko T, Vollmer TL. The dynamic link between the integrity of the immune system and zinc status. J Nutr 2000; 130: 1399S–406S. [DOI] [PubMed] [Google Scholar]
- 48. King LE, Osati‐Ashtiani F, Fraker PJ. Depletion of cells of the B lineage in the bone marrow of zinc‐deficient mice. Immunology 1995; 85: 69–73. [PMC free article] [PubMed] [Google Scholar]
- 49. Krebs NF, Sontag M, Accurso FJ, Hambidge KM. Low plasma zinc concentrations in young infants with cystic fibrosis. J Pediatr 1998; 133: 761–4. [DOI] [PubMed] [Google Scholar]
- 50. Prasad AS. Clinical and biochemical manifestation zinc deficiency in human subjects. J Pharmacol 1985; 16: 344–52. [PubMed] [Google Scholar]
- 51. Tapazoglou E, Prasad AS, Hill G, Brewer GJ, Kaplan J. Decreased natural killer cell activity in patients with zinc deficiency with sickle cell disease. J Laboratory Clin Med 1985; 105: 19–22. [PubMed] [Google Scholar]
- 52. Zemel BS, Kawchak DA, Fung EB, Ohene‐Frempong K, Stallings VA. Effect of zinc supplementation on growth and body composition in children with sickle cell disease. Am J Clin Nutr 2002; 75: 300–7. [DOI] [PubMed] [Google Scholar]
- 53. DePasquale‐Jardieu P, Fraker PJ. The role of corticosterone in the loss in immune function in the zinc‐deficient A/J mouse. J Nutr 1979; 109: 1847–55. [DOI] [PubMed] [Google Scholar]
- 54. DePasquale‐Jardieu P, Fraker PJ. Further characterization of the role of corticosterone in the loss of humoral immunity in zinc‐deficient A/J mice as determined by adrenalectomy. J Immunol 1980; 124: 2650–5. [PubMed] [Google Scholar]
- 55. King LE, Osati‐Ashtiani F, Fraker PJ. Apoptosis plays a distinct role in the loss of precursor lymphocytes during zinc deficiency in mice. J Nutr 2002; 132: 974–9. [DOI] [PubMed] [Google Scholar]
- 56. Prasad AS. Effects of zinc deficiency on Th1 and Th2 cytokine shifts. J Infect Dis 2000; 182 (Suppl 1): S62–8. [DOI] [PubMed] [Google Scholar]
- 57. Moore JB, Blanchard RK, McCormack WT, Cousins RJ. cDNA array analysis identifies thymic LCK as upregulated in moderate murine zinc deficiency before T‐lymphocyte population changes. J Nutr 2001; 131: 3189–96. [DOI] [PubMed] [Google Scholar]
- 58. Bao B, Prasad AS, Beck FW, Godmere M. Zinc modulates mRNA levels of cytokines. Am J Physiol Endocrinol Metab 2003; 285: E1095–102. [DOI] [PubMed] [Google Scholar]
- 59. Kitamura H, Morikawa H, Kamon H et al . Toll‐like receptor‐mediated regulation of zinc homeostasis influences dendritic cell function. Nat Immunol 2006; 7: 971–7. [DOI] [PubMed] [Google Scholar]
- 60. Li Y, Hough CJ, Suh SW, Sarvey JM, Frederickson CJ. Rapid translocation of Zn (2+) from presynaptic terminals into postsynaptic hippocampal neurons after physiological stimulation. J Neurophysiol 2001; 86: 2597–604. [DOI] [PubMed] [Google Scholar]
- 61. Qian J, Noebels JL. Visualization of transmitter release with zinc fluorescence detection at the mouse hippocampal mossy fibre synapse. J Physiol 2005; 566: 747–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Ueno S, Tsukamoto M, Hirano T et al . Mossy fiber Zn2+ spillover modulates heterosynaptic N‐methyl‐D‐aspartate receptor activity in hippocampal CA3 circuits. J Cell Biol 2002; 158: 215–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Weiss JH, Sensi SL. Ca2+–Zn2+ permeable AMPA or kainate receptors: possible key factors in selective neurodegeneration. Trends Neurosci 2000; 23: 365–71. [DOI] [PubMed] [Google Scholar]
- 64. Xie X, Smart TG. Modulation of long‐term potentiation in rat hippocampal pyramidal neurons by zinc. Pflugers Arch 1994; 427: 481–6. [DOI] [PubMed] [Google Scholar]
- 65. Hershfinkel M, Moran A, Grossman N, Sekler I. A zinc‐sensing receptor triggers the release of intracellular Ca2+ and regulates ion transport. Proc Natl Acad Sci USA 2001; 98: 11749–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Colvin RA, Fontaine CP, Laskowski M, Thomas D. Zn2+ transporters and Zn2+ homeostasis in neurons. Eur J Pharmacol 2003; 479: 171–85. [DOI] [PubMed] [Google Scholar]
- 67. Hosie AM, Dunne EL, Harvey RJ, Smart TG. Zinc‐mediated inhibition of GABA(A) receptors: discrete binding sites underlie subtype specificity. Nat Neurosci 2003; 6: 362–9. [DOI] [PubMed] [Google Scholar]
- 68. Hirzel K, Müller U, Latal AT et al . Hyperekplexia phenotype of glycine receptor alpha1 subunit mutant mice identifies Zn (2+) as an essential endogenous modulator of glycinergic neurotransmission. Neuron 2006; 52: 679–90. [DOI] [PubMed] [Google Scholar]
- 69. Mayer ML, Vyklicky L Jr. The action of zinc on synaptic transmission and neuronal excitability in cultures of mouse hippocampus. J Physiol 1989; 415: 351–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Christine CW, Choi DW. Effect of zinc on NMDA receptor‐mediated channel currents in cortical neurons. J Neurosci 1990; 10: 108–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Legendre P, Westbrook GL. The inhibition of single N‐methyl‐D‐aspartate‐activated channels by zinc ions on cultured rat neurones. J Physiol 1990; 429: 429–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Huang YZ, Pan E, Xiong ZQ, McNamara JO. Zinc‐mediated transactivation of TrkB potentiates the hippocampal mossy fiber‐CA3 pyramid synapse. Neuron 2008; 57: 546–58. [DOI] [PubMed] [Google Scholar]
- 73. Gomperts BD, Tatham PER, Kramer IM. Signal Transduction, xiv. San Diego, Calif.: Academic Press, 2002; 424. [Google Scholar]
- 74. Dominguez D, Montserrat‐Sentís B, Virgós‐Soler A et al . Phosphorylation regulates the subcellular location and activity of the snail transcriptional repressor. Mol Cell Biol 2003; 23: 5078–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Von Bulow V, Rink L, Haase H. Zinc‐mediated inhibition of cyclic nucleotide phosphodiesterase activity and expression suppresses TNF‐alpha and IL‐1 beta production in monocytes by elevation of guanosine 3′,5′‐cyclic monophosphate. J Immunol 2005; 175: 4697–705. [DOI] [PubMed] [Google Scholar]
- 76. Spahl DU, Berendji‐Grun D, Suschek CV, Kolb‐Bachofen V, Kroncke KD. Regulation of zinc homeostasis by inducible NO synthase‐derived, nuclear metallothionein translocation and intranuclear Zn2+ release. Proc Natl Acad Sci USA 2003; 100: 13952–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Glesne D, Vogt S, Maser J, Legnini D, Huberman E. Regulatory properties and cellular redistribution of zinc during macrophage differentiation of human leukemia cells. J Struct Biol 2006; 155: 2–11. [DOI] [PubMed] [Google Scholar]
- 78. Yamasaki S, Sakata‐Sogawa K, Hasegawa A et al . Zinc is a novel intracellular second messenger. J Cell Biol 2007; 177: 637–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Zoli A et al . Serum zinc and copper in active rheumatoid arthritis: correlation with interleukin 1 beta and tumour necrosis factor alpha. Clin Rheumatol 1998; 17: 378–82. [DOI] [PubMed] [Google Scholar]
- 80. Gupta SK, Singh SP, Shukla VK. Copper, zinc, and Cu/Zn ratio in carcinoma of the gallbladder. J Surg Oncol 2005; 91: 204–8. [DOI] [PubMed] [Google Scholar]
- 81. Costello LC, Franklin RB. The clinical relevance of the metabolism of prostate cancer; zinc and tumor suppression: connecting the dots. Mol Cancer 2006; 5: 17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Chakravarty PK, Ghosh A, Chowdhury JR. Zinc in human malignancies. Neoplasma 1986; 33: 85–90. [PubMed] [Google Scholar]
- 83. Margalioth EJ, Schenker JG, Chevion M. Copper and zinc levels in normal and malignant tissues. Cancer Sci 1983; 52: 868–72. [DOI] [PubMed] [Google Scholar]
- 84. Schwartz AE, Leddicotte GW, Fink RW, Friedman EW. Trace elements in normal and malignant human breast tissue. Surgery 1974; 76: 325–9. [PubMed] [Google Scholar]
- 85. Fernandes G, Nair M, Onoe K, Tanaka T, Floyd R, Good RA. Impairment of cell‐mediated immunity functions by dietary zinc deficiency in mice. Proc Natl Acad Sci USA 1979; 76: 457–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Keen CL, Gershwin ME. Zinc deficiency and immune function. Annu Rev Nutr 1990; 10: 415–31. [DOI] [PubMed] [Google Scholar]
- 87. Fraker PJ, Caruso R, Kierszenbaum F. Alteration of the immune and nutritional status of mice by synergy between zinc deficiency and infection with Trypanosoma cruzi . J Nutr 1982; 112: 1224–9. [DOI] [PubMed] [Google Scholar]
- 88. Telford WG, Fraker PJ. Preferential induction of apoptosis in mouse CD4+ CD8+ alpha beta TCRloCD3 epsilon lo thymocytes by zinc. J Cell Physiol 1995; 164: 259–70. [DOI] [PubMed] [Google Scholar]
- 89. Ibs KH, Rink L. Zinc‐altered immune function. J Nutr 2003; 133: 1452S–6S. [DOI] [PubMed] [Google Scholar]
- 90. Frederickson CJ, Koh JY, Bush AI. The neurobiology of zinc in health and disease. Nat Rev Neurosci 2005; 6: 449–62. [DOI] [PubMed] [Google Scholar]
- 91. Manning DL, Robertson JF, Ellis IO et al . Oestrogen‐regulated genes in breast cancer: association of pLIV1 with lymph node involvement. Eur J Cancer 1994; 30A: 675–8. [DOI] [PubMed] [Google Scholar]
- 92. Taylor KM, Nicholson RI. The LZT proteins; the LIV‐1 subfamily of zinc transporters. Biochim Biophys Acta 2003; 1611: 16–30. [DOI] [PubMed] [Google Scholar]
- 93. Kasper G et al . Expression levels of the putative zinc transporter LIV‐1 are associated with a better outcome of breast cancer patients. Int J Cancer 2005; 117: 961–73. [DOI] [PubMed] [Google Scholar]
- 94. Thiery JP. Epithelial‐mesenchymal transitions in tumour progression. Nat Rev Cancer 2002; 2: 442–54. [DOI] [PubMed] [Google Scholar]
- 95. Batlle E, Sancho E, Francí C et al . The transcription factor snail is a repressor of E‐cadherin gene expression in epithelial tumour cells. Nat Cell Biol 2000; 2: 84–9. [DOI] [PubMed] [Google Scholar]
- 96. Christiansen JJ, Rajasekaran AK. Reassessing epithelial to mesenchymal transition as a prerequisite for carcinoma invasion and metastasis. Cancer Res 2006; 66: 8319–26. [DOI] [PubMed] [Google Scholar]
- 97. Kaler P, Prasad R. Molecular cloning and functional characterization of novel zinc transporter, rZip10 (Slc39a10), involved in zinc uptake across rat renal brush border membrane. Am J Physiol Renal Physiol 2007; 292: F217–29. [DOI] [PubMed] [Google Scholar]
- 98. Begum NA, Kobayashi M, Moriwaki Y, Matsumoto M, Toyoshima K, Seya T. Mycobacterium bovis BCG cell wall and lipopolysaccharide induce a novel gene, BIGM103, encoding a 7‐TM protein: identification of a new protein family having Zn‐transporter and Zn‐metalloprotease signatures. Genomics 2002; 80: 630–45. [DOI] [PubMed] [Google Scholar]
- 99. Chimienti F, Devergnas S, Favier A, Seve M. Identification and cloning of a beta‐cell‐specific zinc transporter, ZnT‐8, localized into insulin secretory granules. Diabetes 2004; 53: 2330–7. [DOI] [PubMed] [Google Scholar]
- 100. Kambe T, Yamaguchi‐Iwai Y, Sasaki R, Nagao M. Overview of mammalian zinc transporters. Cell Mol Life Sci 2004; 61: 49–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Wang F, Kim BE, Dufner‐Beattie J, Petris MJ, Andrews G, Eide DJ. Acrodermatitis enteropathica mutations affect transport activity, localization and zinc‐responsive trafficking of the mouse ZIP4 zinc transporter. Hum Mol Genet 2004; 13: 563–71. [DOI] [PubMed] [Google Scholar]
- 102. Huang L, Kirschke CP, Zhang Y, Yu YY. The ZIP7 gene (Slc39a7) encodes a zinc transporter involved in zinc homeostasis of the Golgi apparatus. J Biol Chem 2005; 280: 15456–63. [DOI] [PubMed] [Google Scholar]
- 103. Kelleher SL, Lönnerdal B. Molecular regulation of milk trace mineral homeostasis. Mol Aspects Med 2005; 26: 328–39. [DOI] [PubMed] [Google Scholar]
- 104. Taylor KM, Hiscox S, Nicholson RI. Zinc transporter LIV‐1: a link between cellular development and cancer progression. Trends Endocrinol Metab 2004; 15: 461–3. [DOI] [PubMed] [Google Scholar]