

Abstract
A monoclonal antibody (mAb) MY.1E12 was applied to detect MUC1 with sialylated glycans in a total of 55 formalin‐fixed, paraffin‐embedded surgical specimens of ovarian clear cell adenocarcinomas. A reverse correlation between the binding levels of this mAb and patient survival was demonstrated. To examine the role of MUC1 in ovarian clear cell carcinomas, two cDNA encoding MUC1 were transfected into ES‐2 ovarian clear cell carcinoma cells. By comparing these cells, the role of MUC1 in tumorigenicity, chemosensitivity and survival under anoikis conditions were assessed. The results indicate that MUC1 expressed on ovarian clear cell carcinoma cells is causally involved in the malignant behavior. (Cancer Sci 2007; 98: 1586–1591)
- Abbreviations: CDDP
cisplatin
- DMEM
Dulbecco's modified Eagle's medium
- EGF
epidermal growth factor
- FITC
fluoroscein‐5‐isothiocyanate
- Gal
galactose
- GalNAc
N‐acetylgalactosamine
- GlcNAc
N‐acetylglucosamine
- IC50
the concentration causing 50% growth inhibition
- mAb
monoclonal antibody
- PBS
phosphate‐buffered saline
- TX
paclitaxel
- WST
4‐[3‐(4‐iodophenyl)‐2‐(4‐nitrophenyl)‐2H‐5‐tetrazolio]‐1,3‐benzene disulfonate.
Ovarian epithelial cancer is a highly lethal neoplasm due to its metastatic capacity to the upper abdomen and its resistance to chemotherapy in its advanced stages.( 1 , 2 ) Advanced ovarian cancer is difficult to treat because it is impossible to completely resect diffuse tumors and to remove residual tumor cells by chemotherapy.( 1 , 2 ) Ovarian cancer has various histological types with different clinical features and prognosis,( 3 ) which are associated with various types of mucins and carbohydrate antigens.( 4 ) Clear cell adenocarcinoma of the ovary, whose incidence is increasing uniquely in Japan, exhibits several characteristics.( 4 ) Clear cell adenocarcinoma accounts for 15–20% of all cases of epithelial ovarian cancer.( 4 , 5 ) Up to 60% of patients with clear cell adenocarcinoma have stage I disease.( 6 ) Although patients with stage I ovarian cancer show a favorable prognosis, patients with clear cell adenocarcinoma are known to have a worse prognosis than those with other histological types of epithelial ovarian carcinomas.( 7 ) However, whether subpopulations of patients with clear cell carcinoma show better or worse prognosis is not known.
MUC1 is a mucin expressed by almost all epithelial cells and many carcinomas, and its expression and distribution are known to correlate with the progression of a variety of malignant diseases, as explored extensively in our and other laboratories.( 8 , 9 , 10 ) MUC1 is a transmembrane glycoprotein with a large extracellular domain that extends 200–500 nm above the cell membrane. The protein backbone of the extracellular domain consists mostly of 30–90 repeats of 20 amino acids that are highly similar.( 8 , 11 , 12 ) Proline residues hold the protein backbone straight, whereas serine and threonine residues, which are O‐glycosylated, make the structure very rigid.( 13 ) O‐Glycans of MUC1 are extended and modified in diverse ways depending on the cell type during their maturation and transport through the intercellular compartment.( 14 ) Despite many reports on the correlation of unique glycosylation status of MUC1 with disease progression, the role of MUC1 glycoforms in ovarian cancer biology is not well understood. Dong and colleagues reported that low expression of MUC1 in the apical membrane was associated with an early stage and a good outcome of invasive ovarian tumors, though their study included very few cases of clear cell tumors.( 15 ) In our previous studies using 17 ovarian carcinoma cell lines,( 4 ) cell lines from ovarian clear cell adenocarcinoma expressed MUC1 but no other mucins as far as tested at the mRNA and protein levels. In contrast, cell lines derived from other histological types expressed other mucins too. Therefore, we chose ovarian clear cell adenocarcinoma cells to address the biological and clinical significance of MUC1. In the present study, we used mAb MY.1E12,( 16 ) which was previously generated in our laboratory, to detect MUC1 with sialylated O‐glycans( 17 ) in formalin‐fixed, paraffin‐embedded surgical specimens of clear cell adenocarcinoma. A correlation between the level of expression of sialylated MUC1 and survival was demonstrated in ovarian clear cell adenocarcinoma.
In the present report, the significance of MUC1 expression with respect to chemosensitivity and sensitivity to apoptosis induced by loss of attachment status was also investigated by the use of established ovarian clear cell carcinoma cell lines. Chemosensitivity should represent another important aspect of diversity in the malignant behavior of ovarian clear cell carcinoma, and MUC1 overexpression on tumor cells was shown to confer resistance to anticancer drugs. Moreover, MUC1 overexpression was also shown to attenuate apoptosis induced under anoikis condition.
Materials and Methods
Specimens. In the 11 years between 1984 and 1995, 325 Japanese women with primary ovarian cancer were initially treated at the Department of Obstetrics and Gynecology of Keio University Hospital. Among them, 55 patients (aged from 39 to 76 years, mean 51.1 years) were diagnosed as having primary ovarian clear cell adenocarcinoma. All 55 patients received radical or cytoreductive surgery as the first step in their treatment. Between 1984 and 1989, our standard radical operation for patients with early cancer (stages I and II) was total abdominal hysterectomy, bilateral salpingo‐oophorectomy, and omentectomy. From 1990, pelvic and para‐aortic lymphadenectomy were added to the surgical procedures. For patients with advanced cancer (stages III and IV), cytoreductive surgery was carried out throughout the period from 1984 to 1995. After surgery, almost all patients received platinum‐containing chemotherapy, such as CDDP combined with doxorubicin and cyclophosphamide or CDDP combined with cyclophosphamide. Clinical data were collected from the clinical records at Keio University Hospital. Specimens of tumors were obtained during the initial operation from all 55 patients. The histopathological diagnosis of ovarian clear cell adenocarcinoma was verified by a single pathologist who examined hematoxylin–eosin‐stained, formalin‐fixed, and paraffin‐embedded materials.
Immunohistochemical staining. Surgical specimens fixed in formalin and embedded in paraffin were cut into 4‐µm sections. The sections were deparaffinized and treated with 0.3% hydrogen peroxide (Wako, Osaka, Japan) in methanol for 30 min to block endogenous peroxidase activity. The sections were rehydrated in PBS and incubated with 10% rabbit serum (Histofine SAB‐PO M kit; Nichirei, Tokyo, Japan) for 30 min at room temperature. Sections were incubated with mAb MY.1E12, purified from hybridoma cultures with protein G‐agarose,( 16 ) and diluted in PBS containing 3% bovine serum albumin overnight at 4°C. The sections were incubated with biotinylated goat antimouse immunoglobulin (Nichirei) for 30 min at room temperature and with a solution of horseradish peroxidase‐conjugated streptavidin (Nichirei) for 30 min at room temperature. After repeated washing, the sections were processed with an AEC (red) Substrate Kit (Zymed, South San Francisco, CA, USA) and stained with Mayer's hematoxylin (Wako).
The survival of each patient and the recurrence of the disease were recorded every month and analyzed by the Kaplan–Meier method. Differences in survival between subgroups of patients that were classified according to reactivity with mAb MY.1E12 (Table 1) were analyzed by the log‐rank test. A P‐value < 0.05 was considered statistically significant.
Table 1.
Classification of ovarian clear cell carcinoma patient according to the staining rate and intensity with monoclonal antibody MY.1E12
| Staining intensity | Positive rate | ||
|---|---|---|---|
| 0–5% | 5–50% | >50% | |
| 1+ | +/– | + | ++ |
| 2+ | + | ++ | +++ |
Staining with monoclonal antibody MY.1E12 was graded on the basis of the percentage of positively stained tumor cells as follows: –, no staining; +/–, less than 5% of tumor cells were stained; +, 5–50% of tumor cells were stained; ++, over 50% of tumor cells were stained. The staining intensity was graded as follows: –, negative; 1+, weak; 2+, strong. Final scoring (– to ++) was calculated as (staining intensity × positive rate), as indicated in the top row of the table.
Transfection with MUC1 cDNA. ES‐2 cells( 18 ) were purchased from the American Type Culture Collection (Rockville, MD, USA). ES‐2 is a human ovarian carcinoma cell line that demonstrated the lowest MUC1 level of expression among all of the clear cell adenocarcinoma cell lines examined.( 4 ) MUC1 cDNA encoding full‐length MUC1 protein containing 22 or 42 tandem repeats (which were kind gifts from Dr O. J. Finn [University of Pittsburgh School of Medicine, Pittsburgh, PA, USA] and Dr M. A. Hollingsworth [University of Nebraska Medical Centre, Omaha, NE, USA], respectively),( 19 , 20 ) were ligated into pCEP4 (Invitrogen, Carlsbad, CA, USA) vectors using the restriction enzyme BamHI and were stably transfected into ES‐2 cells by electroporation. After selection with hygromycin B (Calbiochem‐Novabiochem, San Diego, CA, USA), transfected cells were further selected by three passages through a cell sorter (Epics Elite; Beckman‐Coulter, Fullerton, CA, USA) using mAb MY.1E12. ES‐2 cells transfected with the cDNA encoding MUC1 protein containing 22 or 42 tandem repeats were designated as ES‐2/T‐22 cells and ES‐2/T‐42 cells, respectively. Finally, flow cytometric analysis was carried out using an Epics XL (Beckman‐Coulter) to determine the expression of sialylated MUC1 mucin. The indirect immunofluorescence method was used to stain cancer cells with mAb MY.1E12 (staining for 30 min on ice), followed by the addition of fluorescein‐conjugated goat antimouse affinity‐purified antibody (Cappel, West Chester, PA, USA) as the secondary antibody.
In vitro growth of transfected cells. Viable transfectants (0.5 × 104 cells) were cultured in 96‐well plates (Sumitomo Bakelite, Tokyo, Japan) for the times indicated. Then, the number of viable cells in each well was measured by a WST‐1 colorimetric assay (Roche Diagnostics, Mannheim, Germany) using a Model 550 Microplate Reader (Bio‐Rad, Hercules, CA, USA).
In vivo growth of transfected cells. To examine the in vivo growth of transfectants, viable cells (1 × 106 cells/100 µL PBS) were inoculated subcutaneously into the abdominal wall of 5‐week‐old KSN/nu female mice (Japan SLC, Hamamatsu, Japan). Tumor volumes were calculated as (1/2) × (longest diameter) × (shortest diameter)2 at the specified times. All mice were specific pathogen‐free and their handling was in accordance with the guidelines for the care and use Animal Committee of the Graduate School of Pharmaceutical Sciences, The University of Tokyo.
Anoikis condition assay. PolyHEMA plates were made by applying 2 mL of a 10‐mg/mL solution of polyhydroxyethylmethacrylate (Sigma, St Louis, MO, USA) in 50% ethanol to six‐well plates, and drying under the tissue culture hood. After repeating this procedure twice, extensive washing was carried out with PBS.( 21 , 22 ) Trypsinized ES‐2/mock cells, T‐22 cells and T‐42 cells (2 × 106) were then plated into six‐well plates with or without polyHEMA coating. After 6 and 12 h, cells were collected from the polyHEMA plates by pipetting and from the tissue culture plates by trypsinization. In control incubations in tissue culture plates, floating cells were combined with the attached cells before flow cytometric analysis to determine the spontaneous rate of apoptosis.
After the cells were washed with ice‐cold PBS and centrifuged, the supernatant was discarded and the pellet was resuspended in ice‐cold, diluted binding buffer (Immunotech, Marseille, France) at 106 cells/mL. After staining with propidium iodide and Annexin‐V–FITC (Immunotech) for 10 min on ice in the dark, the cells were analyzed by flow cytometry.
Chemosensitivity to CDDP and TX. ES‐2/mock cells, T‐22 cells and T‐42 cells were suspended in DMEM/F‐12 supplemented with 10% fetal bovine serum (Invitrogen) and seeded into the wells of 96‐well culture plates (Sumitomo Bakelite) at a density of 1 × 105/100 µL medium. After 3 h preincubation, new media (the above mentioned culture medium containing various concentrations of anticancer drugs) were added, and culture was continued for a further 24 h. The dose–response curve for each drug was determined by the WST‐1 assay, and the IC50 was calculated. CDDP and TX were provided by Nippon Kayaku Co. (Tokyo, Japan) and Bristol‐Myers Squibb Co. (Tokyo, Japan), respectively.
Statistical analysis. Statistical analysis was carried out using the unpaired Mann–Whitney U‐test or Fisher's exact probability test and a P‐value of less than 0.05 was considered to be significant.
Results
Monoclonal antibody MY.1E12 as a prognostic indicator of ovarian clear cell carcinomas. As shown in Table 1, staining with mAb MY.1E12 was graded on the basis of the percentage of positively stained tumor cells as follows: –, negative for staining; –/+, less than 5% of tumor cells were stained; +, 5–50% of tumor cells were stained; ++, over 50% of tumor cells were stained. We evaluated staining of the cytoplasm and of the apex on cells, and considered the sample to be positive for mAb MY.1E12 binding when at least one of these sites was positive. The staining intensity was graded as follows: –, negative; 1+, weak; 2+, strong.
Examples of mAb MY.1E12 binding, indicating the presence of MUC1 with sialoglycans in ovarian clear cell adenocarcinomas, are shown in Fig. 1a. Although clear cell carcinomas consist of two distinct tumor cell types, clear cell type and hobnail cell type, both were shown to be stained equally by this mAb. The immunohistochemical staining of mAb MY.1E12 revealed the distribution at the apical membrane, cytoplasm, vacuoles and extracellular secretions. The staining intensity was classified into two groups: a high‐score group (≥ ++) and a low‐score group (≤ +). Univariate (Kaplan–Meier) survival analysis showed that a low‐score (≤ +) group for mAb MY.1E12 reactivity represented patients with good prognosis (P < 0.05; Fig. 1b). The score according to immunohistochemical staining was also compared with clinical stage. As shown in Fig. 1c, the patients with a high score (≥ ++) for mAb MY.1E12 reactivity were associated with high clinical stages. Accordingly, mAb MY.1E12 reactivity was not considered as an independent prognostic factor for clear cell adenocarcinoma of the ovary.
Figure 1.
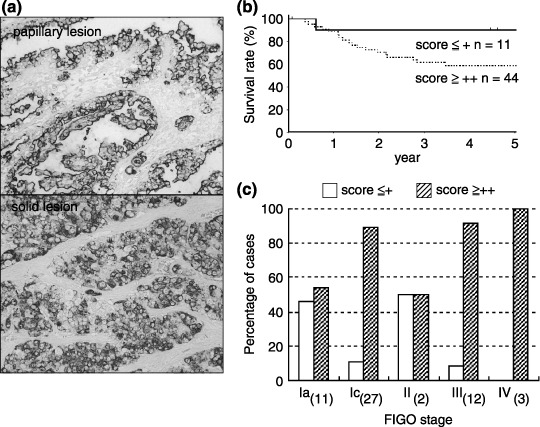
(a) Expression of MUC1 with sialoglycans detected by monoclonal antibody (mAb) MY.1E12 in an ovarian clear cell adenocarcinoma specimen. Both solid lesions and papillary lesions showed sialylated MUC1 expression on the apical membrane and the cytoplasm. (b) Relationship between sialylated MUC1 expression and survival of patients with clear cell adenocarcinoma. All patients were divided into two groups: low score (≤ +) and high score (≥ ++). (c) Frequency analysis of mAb MY.1E12 binding in clear cell adenocarcinoma of the ovary according to stage. The number of patients is given in parentheses.
Biological characteristics of ovarian carcinoma cells expressing MUC1. A clear cell carcinoma line ES‐2 was chosen as the recipient of MUC1 cDNA transfection. MUC1 cDNA with different numbers of tandem repeats were used. According to flow cytometric analysis with mAb MY.1E12, the expression of sialylated MUC1 was not detected on ES‐2/mock cells, whereas the levels were high on ES‐2/T‐42 and ES‐2/T‐22 cells. After three cycles of cell sorting and subcultures, profiles of mAb MY.1E12 binding were determined (Fig. 2). The fluorescence intensity of ES‐2/T‐42 cells was almost two‐fold that of ES‐2/T‐22 cells. Although other investigators often carry out cloning of transfectant cells, we chose to use cell sorting to avoid clonal artifacts. An advantage of this method is that the expression levels of all molecules, except the target molecule (i.e. MUC1 in this case), are supposed to be in a similar heterogeneous status as in the parental cells. We examined whether overexpression of MUC1 on ES‐2 cells influenced the expression of adhesion molecules. A difference was not found with regard to the expression of CD44, β1‐integrin, hyaluronan, heparan sulfate, sialyl Lewis X and sialic acidα2–3Galβ1–4GlcNAc recognized by Maackia amurensis leukoagglutinin between MUC1‐transfectant and mock transfectant cells (data not shown). The plating efficiency on a plastic tissue culture dish in a regular culture medium containing 10% fetal bovine serum did not show any difference between MUC1‐transfectant and mock‐transfectant cells (data not shown). These results demonstrate that the adhesive properties of ES‐2 cells were not affected by MUC1 expression. Also, the morphology of the transfectant cells observed under a phase‐contrast microscope was very similar (data not shown). The growth of MUC1 transfectant cells in monolayer cultures was examined by the WST‐1 assays. Because the plating efficiency of each cell line was not different, the same numbers of cells were plated. As shown in Fig. 3a, there was no difference in the growth rate in vitro.
Figure 2.
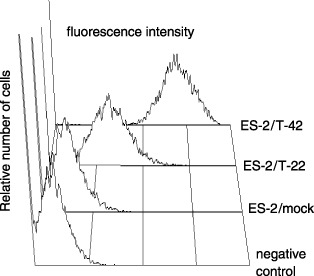
Fluorescence histograms of ES‐2/mock, T‐22 and T‐42 cells stained with monoclonal antibody (mAb) MY.1E12. The negative control was stained with a secondary antibody only. ES‐2/mock, T‐22 and T‐42 cells were prepared by cell sorting three times in an Epics Elite flow cytometer.
Figure 3.
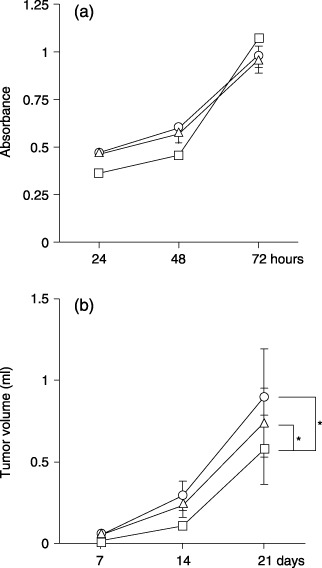
(a) Growth of ES‐2/mock, T‐22 and T‐42 cells in vitro in 96‐well culture plates. The number of viable cells in the wells was measured using the WST‐1 colorimetric assay. Squares, triangles and circles indicate ES‐2/mock cells, ES‐2/T‐22 cells and ES‐2/T‐42 cells, respectively. (b) Growth of ES‐2/mock cells, ES‐2/T‐22 cells and ES‐2/T‐42 cells in vivo. Viable cells (1 × 106 cells/100 µL phosphate‐buffered saline) were injected subcutaneously into the abdominal wall of KSN/nu female mice (5 weeks old). Tumor sizes were determined according to the calculated volumes (1/2) × (longest diameter) × (shortest diameter)2. Error bars indicate the standard deviations. Squares, triangles and circles show ES‐2/mock cells, ES‐2/T‐22 cells and ES‐2/T‐42 cells, respectively. Mann–Whitney U‐tests were used to assess the significance of the difference (n = 6, P < 0.05). Asterisks indicate P < 0.05.
We subsequently carried out xenografts in nude mice. Macroscopically, there was no substantial difference between tumors derived from MUC1‐transfectant and mock‐transfectant cells. All of the tumors were well encapsulated, and no evidence of increased infiltration into the surrounding tissues was observed. Examinations after hematoxylin–eosin staining of the tumor tissue sections did not reveal significant histological differences between MUC1 and mock transfectant cells (data not shown). As shown in Fig. 3b, there was a significant difference with regard to the tumor volume. Tumors from mock transfectant cells were significantly smaller than those from ES‐2/T‐42 or ES‐2/T‐22 cells (P < 0.05).
Cell viability under non‐adherent conditions was examined because cancer cells disseminated from a primary lesion into the peritoneal cavity should transiently survive anchorage‐independently. As shown in Fig. 4, the viability of ES‐2/T‐42 and ES‐2/T‐22 cells under anchorage‐independent conditions (anoikis assays) was significantly higher than ES‐2/mock cells at 6‐ and 12‐h time points, as judged by the cell number shown by the propidium iodide staining (dead cells), and by the Annexin‐V–FITC staining (apoptotic cells).
Figure 4.
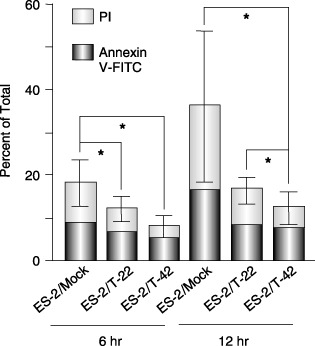
Results of the anoikis condition assays. Significant differences for the sums of propidium iodide (PI) and Annexin‐V–fluoroscein‐5‐isothiocyanate (FITC) intensities were assessed using the Mann–Whitney U‐test. Error bars indicate the standard deviation of sum of PI and Annexin‐V–FITC data. Asterisks indicate P < 0.05.
An important problem to be solved in the treatment of ovarian clear cell carcinoma is chemoresistance to anticancer drugs. The current standard chemotherapy regimen for ovarian cancer is a combination of CDDP and TX. We carried out a drug sensitivity test for CDDP and TX. The IC50 values were calculated at 24 h after the addition of these agents to cultures of MUC1‐transfectant cells. As shown in Table 2, ES‐2/mock cells showed a significantly higher sensitivity to both CDDP and TX than ES‐2/T‐22 cells or ES‐2/T‐42 cells (P < 0.05). ES‐2/T‐22 cells appeared to be more sensitive to these agents than ES‐2/T‐42 cells, but the difference was not statistically significant.
Table 2.
Chemosensitivity of cisplatin and paclitaxel
| IC50 (µg/mL) | ES‐2/mock | ES‐2/T‐22 | ES‐2/T‐42 |
|---|---|---|---|
| Cisplatin | 2.63 ± 0.55*1*2 | 7.74 ± 1.89*1 | 11.25 ± 3.43*2 |
| Paclitaxel | 5.95 ± 0.64*3 | 7.46 ± 2.08 | 9.76 ± 2.56*3 |
A dose–response curve for each drug was determined by WST‐1 assay, and the 50% growth‐inhibitory concentration (IC50) was calculated. Significance of the differences was assessed by Mann–Whitney U‐test. Asterisks (*1,*2,*3) show that the differences (*1 and *1,*2 and *2,*3 and *3) are statistically significant.
Discussion
Ovarian malignancies have a worse prognosis than other gynecologic malignancies and early diagnosis is difficult due to the anatomical location and lack of symptoms at an early stage. Einhorn and colleagues pointed out that the clinical stage and histological type are important determinants of prognosis and that these factors correlate with tumor spread.( 3 ) Dong and colleagues reported that a high expression of MUC1 in the apical membrane was associated with non‐mucinous ovarian tumors, whereas a low expression of MUC1 in the same lesion was associated with an early stage and a good outcome of invasive ovarian tumors.( 15 ) In our previous study at the mRNA, ovarian clear cell adenocarcinoma cell lines expressed MUC1 but no other mucins tested, whereas mucinous adenocarcinomas expressed MUC1 and other mucins. Interestingly, all cell lines derived from clear cell adenocarcinoma expressed MUC1 mRNA, and the glycoprotein product detected on the cell surface contained sialoglycans at a variety of levels, as differential expression was revealed by the binding of mAb MY.1E12.( 4 ) Thus, MUC1, particularly that with sialoglycans, might influence clinical features and biological behaviors of clear cell carcinomas. Clear cell carcinomas have uniquely been increasing in Japan, and the mechanism of progression and acquired chemoresistance is not well understood. In the present study, we found that the level of MUC1 with sialoglycans recognized by mAb MY.1E12 was inversely correlated with patient prognosis. However, sialylated MUC1 expression is associated with the clinical stage of ovarian cancer, and it therefore may not be considered as an independent prognostic factor in ovarian clear cell carcinomas. In ovarian carcinoma, the degree of histological differentiation is generally classified as grade 1 through to grade 3, determined according to the increase in the proportion of solid growth within the adenocarcinoma. But no classification system has been established for the differentiation of clear cell adenocarcinoma, and all of their tumors are classified as grade 3. According to our immunohistochemical results, the level of sialylated MUC1 expression in ovarian clear cell adenocarcinoma may be available as an index of malignancy, particularly in stage I disease. Further investigation including more samples is needed.
MUC1 on tumor cells has been shown to suppress homotypic cellular aggregation,( 23 ) to prevent cell–matrix adhesions, and to promote invasion in Matrigel.( 24 , 25 ) Moreover, MUC1 was reported to inhibit cytotoxic lymphocyte–target cell interactions in vitro.( 26 , 27 ) Furthermore, MUC1 was reported to induce apoptosis of lymphocytes.( 28 , 29 ) Glycoforms of MUC1 associated with these functions were not previously known. Therefore, ovarian clear cell carcinoma patients with high expression of sialylated MUC1 may be at an advanced stage and have a poor prognosis. To examine the role of MUC1 in the pathobiology of ovarian clear cell adenocarcinoma, we transfected full‐length cDNA encoding MUC1 with 22 and 42 tandem repeats into ES‐2 cells, which are derived from ovarian clear cell adenocarcinoma and express very low levels of MUC1.( 4 ) Although Wesseling and colleagues( 24 ) reported that transfection of MUC1 into a melanoma cell line induced loss of adhesion to the culture dish, we did not observe such a change after MUC1 transfection into the ES‐2 cell line. Moreover, morphological differences were not observed between mock‐ and MUC1‐transfectant cells. This discrepancy was probably dependent on the cellular origin that greatly influences the glycosylation of MUC1. A role of MUC1 in regulating tumor cell growth was previously reported in several cancer types,( 30 , 31 , 32 ) yet the results are contradictory among different reports. Our present results show the growth advantage of MUC1‐transfectant cells only in vivo. Recently, MUC1 was shown to be associated with all four EGF receptors, and that increased MUC1 expression potentiated EGF signaling through activation of the mitogenic MAP kinase pathways.( 33 ) Another study showed that MUC1 constitutively associates with the EGF receptor, and that EGF receptor‐mediated phosphorylation of MUC1 induces its binding to c‐Src tyrosine kinase and β‐catenin.( 34 ) However, it is still controversial whether these characteristics are associated with any of the MUC1 glycoforms or MUC1 on different cells with unique glycosylation. Effects of MUC1 on growth signaling cannot explain our result that MUC1‐transfectant cells grow faster in vivo, but not in vitro, than mock transfectant cells. In vivo growth of tumor cells does not necessarily correspond with in vitro growth because of hypoxia, attack by natural killer cells and macrophages, and transient loss of anchorage. It is known that MUC1 secreted or expressed on plasma membranes of tumor cells interferes with natural killer cell‐mediated lysis.( 35 ) As stated later, MUC1 renders resistance to apoptosis induced by anoikis. Therefore, it is likely that MUC1 overexpression in ovarian carcinoma cells provides an advantage to in vivo tumorigenicity and growth without directly influencing growth signaling. These possibilities should be further examined in the near future.
In advanced ovarian cancer, cancer cells are shed from the primary tumor into the peritoneal cavity, followed by attachment of cells to the peritoneal mesothelium that lines the bowel and abdominal wall. It is likely that specific adhesion molecules mediate this interaction between ovarian cancer cells and the peritoneal mesothelium. Among the surface membrane molecules that are capable of mediating cell adhesion, CD44H is a major receptor for hyaluronan that was reported to be partly responsible for promoting the adhesion of ovarian cancer cells to the peritoneum.( 36 , 37 ) When released from a primary tumor into the peritoneal cavity, cancer cells lose their anchorage and undergo apoptosis. Frisch and Francis first reported that apoptosis was induced by disruption of the interactions between epithelial cells and the extracellular matrix, and termed this phenomenon ‘anoikis’.( 22 ) To understand the mechanism of peritoneal dissemination, especially the events during the spread of cancer cells through the peritoneal cavity, the survival of floating cancer cells is critical. Our present in vitro model was considered to be suitable for studying such situations, and our results demonstrated that the expression of sialylated MUC1 led to resistance to apoptosis. Alteration of the environment (i.e. from anchorage to non‐adherent conditions) may induce apoptosis, whereas a high level of sialylated MUC1 may support tumor cell survival under such conditions.
Ovarian clear cell adenocarcinoma is one of the more intriguing epithelial ovarian cancers due to its distinct clinical features. Many gynecologists feel that clear cell adenocarcinoma of the ovary is less responsive to chemotherapy than the more common ovarian serous adenocarcinoma.( 5 , 38 , 39 , 40 , 41 ) The high recurrence rate of early disease may be due to chemoresistance. In support of this, cultured clear cell adenocarcinoma cell lines were found to exhibit resistance to CDDP.( 42 ) However, clear cell adenocarcinoma is regarded clinically as a high‐grade differentiated tumor and all patients with clear cell adenocarcinoma are recommended to have adjuvant chemotherapy. Investigation of the mechanism of chemoresistance in a subpopulation limited to ovarian clear cell adenocarcinoma may provide new strategies for chemotherapy. We demonstrated that MUC1 modified the cells to be more resistant against anticancer drugs than cells without MUC1. Although CDDP and TX have different mechanisms of cytotoxity,( 43 , 44 ) MUC1 expression induced resistance against both of them.
In conclusion, increased expression of MUC1 shown by the binding of mAb MY.1E12, specific for MUC1, with sialoglycans was associated with advanced stage of ovarian clear cell adenocarcinoma, and MUC1 containing sialoglycans expressed on clear cell adenocarcinoma cells influenced chemoresistance and apoptosis.
Acknowledgments
We thank Ms Miki Noji for her assistance in preparing this manuscript. This work was supported by grants‐in‐aid from the Ministry of Education, Science, Sports and Culture of Japan (11557180, 11672162 and 12307054), from the Research Association for Biotechnology, from the Program for Promotion of Fundamental Studies in Health Sciences of the Pharmaceutical and Medical Device Agency, from the Keio Gijuku Academic Development Fund, and by a Keio University Grant‐in‐Aid for Encouragement of Young Medical Scientists.
References
- 1. Cannistra SA. Cancer of the ovary. N Engl J Med 1993; 329: 1550–9. [DOI] [PubMed] [Google Scholar]
- 2. Omura GA, Brady MF, Homesley HD et al . Long‐term follow‐up and prognostic factor analysis in advanced ovarian carcinoma: the gynecologic oncology group experience. J Clin Oncol 1991; 9: 1138–50. [DOI] [PubMed] [Google Scholar]
- 3. Einhorn N, Nilsson B, Sjovall K. Factors influencing survival in carcinoma of the ovary. Study from a well‐defined Swedish population. Cancer 1985; 55: 2019–25. [DOI] [PubMed] [Google Scholar]
- 4. Tamada Y, Iida S, Aoki D, Nozawa S, Irimura T. Carbohydrate epitopes and mucins expressed by 17 human ovarian carcinoma cell lines. Oncol Res 1999; 11: 233–41. [PubMed] [Google Scholar]
- 5. Sugiyama T, Kamura T, Kigawa J et al . Clinical characteristics of clear cell carcinoma of the ovary: a distinct histologic type with poor prognosis and resistance to platinum‐based chemotherapy. Cancer 2000; 88: 2584–9. [PubMed] [Google Scholar]
- 6. Russell P. Surface epithelial–stromal tumor of the ovary. In: Kurman RJ, ed. Blaustein's Pathology of the Female Genital Tract, 4th edn. New York: Springer‐Verlag, 1995; 752–62. [Google Scholar]
- 7. O’Brien MER, Schofield JB, Tan S, Fryatt I, Fisher C, Wiltshaw E. Clear cell epithelial ovarian cancer (mesonephroid): bad prognosis only in early stages. Gynecol Oncol 1993; 49: 250–4. [DOI] [PubMed] [Google Scholar]
- 8. Hilkens J, Lightenberg MJL, Vos HL, Litvinov SV. Cell membrane‐associated mucins and their adhesion‐modulating property. Trends Biochem Sci 1992; 17: 359–63. [DOI] [PubMed] [Google Scholar]
- 9. Nakamori S, Ota DM, Cleary KR, Shirotani K, Irimura T. MUC1 mucin expression as a marker of progression and metastasis of human colorectal carcinoma. Gastroenterology 1994; 106: 353–61. [DOI] [PubMed] [Google Scholar]
- 10. Gendler SJ, Spicer AP. Epithelial mucin genes. Ann Rev Physiol 1995; 57: 607–34. [DOI] [PubMed] [Google Scholar]
- 11. Gendler SJ, Lancaster CA, Taylor‐Papadimitriou J et al . Molecular cloning and expression of human tumor‐associated polymorphic epithelial mucin. J Biol Chem 1990; 265: 15 286–93. [PubMed] [Google Scholar]
- 12. Lan MS, Batra SK, Qi W‐N, Metzgar RS, Hollingsworth MA. Cloning and sequencing of a human pancreatic tumor mucin cDNA. J Biol Chem 1990; 265: 15 294–9. [PubMed] [Google Scholar]
- 13. Jentoft N. Why are proteins O‐glycosylated? Trends Biochem Sci 1990; 15: 291–4. [DOI] [PubMed] [Google Scholar]
- 14. Litvinov SV, Hilkens J. The epithelial sialomucin, episialin, is sialylated during recycling. J Biol Chem 1993; 268: 21 364–7l. [PubMed] [Google Scholar]
- 15. Dong Y, Walsh MD, Cummings MC et al . Expression of MUC1 and MUC2 mucins in epithelial ovarian tumors. J Pathol 1997; 183: 311–17. [DOI] [PubMed] [Google Scholar]
- 16. Yamamoto M, Bhavanandan VP, Nakamori S, Irimura T. A novel monoclonal antibody specific for sialylated MUC1 mucin. Jpn J Cancer Res 1996; 87: 488–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Takeuchi H, Kato K, Denda‐Nagai K, Hanisch FG, Clausen H, Irimura T. The epitope recognized by the unique anti‐MUC1 monoclonal antibody MY.1E12 involves sialylα2‐3galactosylβ1‐3N‐acetylgalactosaminide linked to a distinct threonine residue in the MUC1 tandem repeat. J Immunol Meth 2002; 270: 199–209. [DOI] [PubMed] [Google Scholar]
- 18. Lau DHM, Lewis AD, Ehsan MN, Sikic BI. Multifactorial mechanisms associated with broad cross‐resistance of ovarian carcinoma cells selected by cyanomorpholino doxorubicin. Cancer Res 1991; 51: 5181–7. [PubMed] [Google Scholar]
- 19. Jerome KR, Bu D, Finn OJ. Expression of tumor‐associated epitopes on Epstein–Barr virus‐immortalized B‐cells and Burkitt's lymphomas transfected with epithelial mucin complementary DNA. Cancer Res 1992; 52: 5985–90. [PubMed] [Google Scholar]
- 20. Batra SK, Kern HF, Worlock AJ, Metzgar RS, Hollingsworth MA. Transfection of the human Muc1 mucin gene into a poorly differentiated human pancreatic tumor cell line, Panc1: integration, expression and ultrastructural changes. J Cell Sci 1991; 100: 841–9. [DOI] [PubMed] [Google Scholar]
- 21. Folkman J, Moscona A. The role of cell shape in growth control. Nature 1978; 273: 345–9. [DOI] [PubMed] [Google Scholar]
- 22. Frisch SM, Francis H. Disruption of epithelial cell–matrix interactions induces apoptosis. J Cell Biol 1994; 124: 619–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Ligtenberg MJ, Buijs F, Vos HL, Hilkens J. Suppression of cellular aggregation by high levels of episialin. Cancer Res 1992; 52: 2318–24. [PubMed] [Google Scholar]
- 24. Wesseling J, Van Der Valk SW, Vos HL, Sonnenberg A, Hilkens J. Episialin (MUC1) overexpression inhibits integrin‐mediated cell adhesion to extracellular matrix components. J Cell Biol 1995; 129: 255–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Wesseling J, Van Der Valk SW, Hilkens J. A mechanism for inhibition of E‐cadherin‐mediated cell–cell adhesion by the membrane‐associated mucin episialin/MUC1. Mol Biol Cell 1996; 7: 565–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Irimura T, Mclsaac AM, Carlson DA et al . Soluble factors in normal tissues that stimulate high‐molecular‐weight sialoglycoprotein production by human colon carcinoma cells. Cancer Res 1990; 50: 3331–8. [PubMed] [Google Scholar]
- 27. Van De Wiel‐van Kemenade E, Ligtenberg MJ, De Boer AJ et al . Episialin (MUC1) inhibits cytotoxic lymphocyte–target cell interaction. J Immunol 1993; 151: 767–76. [PubMed] [Google Scholar]
- 28. Gimmi CD, Morrison BW, Mainprice BA et al . Breast cancer‐associated antigen, DF3/MUC1, induces apoptosis of activated human T cells. Nat Med 1996; 2: 1367–70. [DOI] [PubMed] [Google Scholar]
- 29. Agrawal B, Klantz MJ, Reddish MA, Longenecker BM. Cancer‐associated MUCI mucin inhibits human T‐cell proliferation, which is reversible by IL‐2. Nat Med 1998; 4: 434–9. [DOI] [PubMed] [Google Scholar]
- 30. Makiguchi Y, Hinoda Y, Imai K. Effect of MUC1 mucin, an anti‐adhesion molecule, on tumor cell growth. Jpn J Cancer Res 1996; 87: 505–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Suwa T, Hinoda Y, Makiguchi Y et al . Increased invasiveness of MUC1 and cDNA‐transfected human gastric cancer MKN74 cells. Int J Cancer 1998; 76: 377–82. [DOI] [PubMed] [Google Scholar]
- 32. Satoh S, Hinoda Y, Hayashi T, Burdick MD, Imai K, Hollingsworth MA. Enhancement of metastatic properties of pancreatic cancer cells by MUC1 gene encoding an anti‐adhesion molecule. Int J Cancer 2000; 88: 507–18. [DOI] [PubMed] [Google Scholar]
- 33. Schroeder JA, Thompson MC, Gardner MM, Gendler SJ. Transgenic MUC1 interacts with epidermal growth factor receptor and correlates with mitogen‐activated protein kinase activation in the mouse mammary gland. J Biol Chem 2001; 276: 13 057–64. [DOI] [PubMed] [Google Scholar]
- 34. Li Y, Ren J, Yu W et al . The epidermal growth factor receptor regulates interaction of the human DF3/MUC1 carcinoma antigen with c‐Src and β‐catenin. J Biol Chem 2001; 276: 35 239–42. [DOI] [PubMed] [Google Scholar]
- 35. Zhang K, Sikut R, Hansson GC. A MUC1 mucin secreted from a colon carcinoma cell line inhibits target cell lysis by natural killer cells. Cell Immunol 1997; 176: 158–65. [DOI] [PubMed] [Google Scholar]
- 36. Cannistra SA, Kansas GS, Niloff J, DeFranzo B, Kim Y, Ottensmeier C. Binding of ovarian cancer cells to peritoneal mesothelium in vitro is partly mediated by CD44H. Cancer Res 1993; 53: 3830–8. [PubMed] [Google Scholar]
- 37. Cannistra SA, DeFranzo B, Niloff J, Ottensmeier C. Functional heterogeneity of CD44 molecules in ovarian cancer cell lines. Clin Cancer Res 1995; 1: 333–42. [PubMed] [Google Scholar]
- 38. Behbakht K, Randall TC, Benjamin I, Morgan MA, King S, Rubin SC. Clinical characteristics of clear cell carcinoma of the ovary. Gynecol Oncol 1998; 70: 255–8. [DOI] [PubMed] [Google Scholar]
- 39. Crozier MA, Copeland LJ, Silva EG, Gershenson DM, Stringer CA. Clear cell carcinoma of the ovary. A study of 59 cases. Gynecol Oncol 1989; 35: 199–203. [DOI] [PubMed] [Google Scholar]
- 40. Rubin SC, Wong GYC, Curtin JP, Barakat RR, Hakes TB, Hoskins WJ. Platinum‐based chemotherapy of high‐risk stage I epithelial ovarian cancer following comprehensive surgical staging. Obstet Gynecol 1993; 82: 143–7. [PubMed] [Google Scholar]
- 41. Bertelsen K. Tumor reduction surgery and long‐term survival in advanced ovarian cancer: a DACOVA study. Gynecol Oncol 1990; 38: 203–9. [DOI] [PubMed] [Google Scholar]
- 42. Gorai I, Nakazawa T, Miyagi E, Hirahara F, Nagashima Y, Minaguchi H. Establishment and characterization of two human ovarian clear cell adenocarcinoma lines from metastatic lesions with different properties. Gynecol Oncol 1995; 57: 33–46. [DOI] [PubMed] [Google Scholar]
- 43. Coukos G, Rubin SC. Chemotherapy resistance in ovarian cancer: new molecular perspectives. Obstet Gynecol 1998; 91: 783–92. [DOI] [PubMed] [Google Scholar]
- 44. St Croix B, Kerbel RS. Cell adhesion and drug resistance in cancer. Curr Opin Oncol 1997; 9: 549–56. [DOI] [PubMed] [Google Scholar]