

Abstract
Loneliness has broad public health importance, especially in older adulthood, and there is some evidence suggesting it is associated with several personality disorders (PDs). The etiology of these PD-loneliness associations, however, has rarely been studied, especially in the context of the maladaptive traits of the DSM-5 alternative model of personality disorder (AMPD). To address these limitations, we estimated phenotypic, genetic, and unique environmental associations between loneliness and maladaptive personality traits in a sample of older adults from the Minnesota Twin Registry (n = 1,356, Mage = 70.4). Loneliness was moderately to strongly associated with each of the AMPD domains of negative affect, detachment, antagonism, disinhibition, and psychoticism (r = .22–.58), with evidence of both genetic (rg = .45–.75) and unique environmental (re = .10–.48) influences explaining the associations to varying degrees. We argue that loneliness may be an underappreciated concomitant of personality pathology, with PD traits perhaps underlying its development. Indeed, these findings suggest that loneliness may be a manifestation of the genetic and environmental forces that also lead to pathological personality variation.
Keywords: loneliness, personality, personality inventory for DSM-5, alternative model for personality disorder, biometric model
Public Health Relevance of Loneliness in Older Adulthood
Loneliness is a subjective emotional experience, commonly defined as the distressing feeling accompanying the perception that one’s social needs are not being met by one’s relationships (Hawkley & Cacioppo, 2010). Loneliness is associated with numerous adverse physical and mental health outcomes, including internalizing psychopathology (Beutel et al., 2017), cognitive decline (Kuiper et al., 2015), and cardiovascular disease (Valtorta et al., 2016). In fact, the impact of loneliness is comparable in magnitude to other well-established risk factors for mortality like substance abuse, obesity, and low physical activity (Holt-Lunstad et al., 2015).
These associations are of additional importance in older adult populations because these individuals often face predisposing factors for loneliness like living alone, loss of family and friends, sensory impairments, and chronic illness. In a systematic review of the risk factors in older adults, prediction of longitudinal change in loneliness was found across studies for (a) not being partnered and partner loss, (b) limited social networks, (c) low levels of social activity, (d) poor self-perceived health, and (e) depressed mood and an increase in depression (Dahlberg et al., 2022). Along those lines, the National Academies of Sciences (NAS) has recently emphasized the importance of studying the basic mechanisms of loneliness in older adults, recommending “increased funding of basic research as a key to achieving the goal of developing a more robust evidence base on effective prevention, assessment, and intervention” (National Academies of Sciences, 2020, p. 61).
Evidence-based efforts to reduce loneliness are in their early stages, though becoming increasingly common (Hickin et al., 2021; Lim et al., 2020). The increased study of loneliness as a primary intervention target reflects the high frequency with which it is encountered by community clinicians as cross-diagnostic presenting complaint (Pérez-Rojas et al., 2017) and surveys of clinicians indicating that patient’s presenting complaints tend to shape treatment planning more strongly than do diagnostic categories (First et al., 2018). Despite this clear public health relevance and importance as cross-diagnostic intervention target, loneliness has been infrequently studied in relation to personality pathology.
Loneliness in Personality Pathology
Preliminary evidence suggests that loneliness is commonly experienced across a range of personality pathologies. Based on a meta-synthesis of qualitative studies exploring loneliness among people with PD diagnoses or traits, Ikhtabi et al. (2022) concluded that “experiences of loneliness [are] a central part of the difficulties faced by this group of people” (p. 11), and argue that, given loneliness’s salience, interventions for PD should focus on psychological and social factors that help people connect with others.
Perhaps, the most studied at this intersection is relations between loneliness and borderline PD. Patients diagnosed with borderline PD report feeling considerably lonelier than controls (Hauschild et al., 2018; Liebke et al., 2017). Twin studies allow for inference on the degree to which genetic and environmental factors are involved in the covariance of two phenotypes. For instance, in a large community sample of twins, several borderline personality features were correlated with loneliness (r = .51 for a composite score) and those correlations were explained by both genetic (rg = .64) and unique environmental (re = .40) influences, suggesting common etiological factors (Schermer et al., 2020). Similarly, in an adolescent twin sample, Skaug et al. (2022) demonstrated high genetic overlap between borderline traits and loneliness, with overlap in the unique environment also accounting for a statistically significant but smaller portion of the covariance. Data on associations between loneliness and other PD syndromes is sparser, with limited evidence suggesting cross-sectional links with schizoid (Levi-Belz et al., 2014), avoidant (Hayakawa et al., 2018), and, of smaller magnitude, antisocial (Özdel et al., 2015) PDs. Some dimensional measures of traditional PD syndromes have been related to loneliness as well, such as paranoid beliefs (Alsuhibani et al., 2022), schizotypy (Benson & Park, 2019), histrionic traits (Berryman et al., 2018), vulnerable narcissism (Kealy et al., 2022), social avoidance (Yuan et al., 2022), and dependent personality features (Overholser, 1996). Of the traditional syndromes or trait measures of those syndromes, only obsessive-compulsive PD has not been demonstrated to be associated with higher levels of loneliness (for a review: Reinhard et al., 2022).
The flaws of traditional, categorical PD diagnostic approaches have been well documented (e.g., Tyrer et al., 2007), making it especially important to study relations between loneliness and dimensional measures of personality, like those in modern PD taxonomies. The DSM-5 alternative model for personality disorder (AMPD) defines PD as clinically significant difficulty in self and interpersonal personality functioning (criterion A) as expressed in high levels of at least one maladaptive personality trait (criterion B; American Psychiatric Association [APA], 2013). The pathological personality traits of criterion B are arranged hierarchically with 25 constituent trait facets indexed across five overarching domains, typically measured with the Personality Inventory for DSM-5 (PID-5; Krueger et al., 2012).
The AMPD traits have theoretical (Krueger et al., 2012) and empirically demonstrated links to the Five Factor Model (FFM) of normal-range personality, with strong associations between negative affect and neuroticism, detachment and low extraversion, antagonism and low agreeableness, disinhibition and low conscientiousness, and more modest associations between psychoticism and openness (for a review: Barchi-Ferreira Bel & Osório, 2020). The FFM has been a useful framework for understanding individual differences in loneliness, with meta-analytic evidence suggesting strong cross-sectional associations with neuroticism and low extraversion, as well as moderate associations with low conscientiousness and low agreeableness (Buecker et al., 2020). The limited longitudinal evidence suggests that the influence of neuroticism on residual change in loneliness is stronger than that of loneliness on residual change in neuroticism, though statistically significant relations have been observed in both directions, while the impact of other traits has been modest (Abdellaoui et al., 2019; Freilich, Mann, & Krueger, 2022; Mund & Neyer, 2016). Loneliness appears to have strong genetic correlations (i.e., |rg| > .50) with both neuroticism and, negatively, with extraversion, as well as moderate, negative genetic correlations (rg < −.20) with agreeableness and conscientiousness, and small to moderate unique environmental correlations (.10 < |re| < .50) with each of these four traits (Abdellaoui et al., 2019; Freilich, Mann, South, & Krueger, 2022; Schermer & Martin, 2019). Though not necessarily indicative of causality, longitudinal prediction of residual change in loneliness and significant unique environmental correlations is consistent with broad personality traits, especially neuroticism and low extraversion, being involved in the development of loneliness.
Given these links between the FFM and AMPD and the evidence consistent with the FFM traits being involved in the development of loneliness, it stands to reason that maladaptive personality might also be etiologically involved, perhaps with a larger effect. To our knowledge, the relations between AMPD dimensions and loneliness have been reported in only two previous studies. Roche et al. (2019) related the PID-5 domains to daily single-item loneliness self-reports in a diary design. They found that negative affect (r = .47) and detachment (r = .29) were significantly associated with mean loneliness levels, but antagonism (r = .11), disinhibition (r = .06), and psychoticism (r = .09) were not. Further, they found that negative affect, but not detachment, uniquely amplified associations between daily increases in loneliness and cognitive distortions, concluding that negative affect may be implicated in maintaining loneliness, making it a theoretically compelling target of psychotherapy. Next, in a cross-sectional study with an adolescent sample, Romero and Alonso (2019) found that a three-item loneliness measure was significantly associated with negative affect (r = .20), detachment (r = .21), and psychoticism (r = .30), but not antagonism (r = .06) or disinhibition (r = .11).
In sum, a wide range of personality pathology dimensions appear to involve loneliness. This suggests that loneliness may be a central feature across PD, with an etiology possibly involving PD traits. “Pathological conscientiousness” may be an exception, where loneliness has not been clearly linked to measures of either maladaptively high (i.e., obsessive-compulsive PD) or low (i.e., PID-5 disinhibition) constraint. Evidence is limited for several key domains, notably those of the AMPD, with results regarding disagreeableness (i.e., PID-5 antagonism and antisocial PD diagnoses) and psychoticism appearing to be mixed.
The Present Study
There is tentative but limited evidence suggesting loneliness may be a salient feature across many to most domains of personality pathology. Despite the broad public health importance of loneliness, as well as specific relevance to healthy aging and psychotherapy, these associations have rarely been studied in the context of the AMPD. Further, limited work has explored why personality pathology and loneliness are linked, with evidence from the normal-range personality literature suggesting broad traits may be etiologically involved. Finally, no study has examined loneliness and personality pathology in an older adult sample, a population that does demonstrate PD pathology (Oltmanns & Balsis, 2011) and new onset of PDs in particular (Dupree et al., 2023). Therefore, we sought to examine associations between the AMPD domains and loneliness using a large older adult twin sample.
Having hypothesized significant phenotypic correlations between loneliness and the AMPD domains and facets, we planned a priori to model the degree to which genetic and unique environmental influences explained the associations. We hypothesized that loneliness would have strong associations with negative affect and detachment and moderate associations with antagonism and psychoticism. Further, we hypothesized that the genetic and unique environmental influences between loneliness and those of the AMPD domains would be significantly correlated and that a medium to large amount of the genetic variance in loneliness would overlap with that of the AMPD domains, collectively. We also aimed to extend this work in an exploratory manner to the narrow personality facets of the AMPD to understand associations between loneliness and a wide range of specific traits relevant to personality pathology. These hypotheses are consistent with loneliness resulting from many of the same genetic and environmental forces that impact personality pathology.
Method
Participants
The sample includes adults from the Minnesota Twin Registry (MTR), a birth-record-based registry of twins born in the state of Minnesota (Krueger & Johnson, 2002; Lykken et al., 1990). The recent data collection effort took place from 2017 to 2022 and involved twins born between 1936 and 1959. The current wave also involved eight twin pairs from the Carolina African American Twin Study of Aging (Whitfield, 2013). The age of participants spanned 62–85 years at the time of data collection (M = 70.4; 62% female). Full sample demographics (n = 1,377) are reported in Table S1 in the online supplemental material. Notably, there was limited racial diversity (98% White, which for the MTR sample is reflective of the Minnesota state population for the birth years sampled), but considerable diversity in terms of education (40% without a college degree, 43% with undergraduate degree, and 17% with graduate degree) and marital status (74% married or living common law, 10% widowed, 11% divorced or separated, and 5% single).
Because of the public health significance of loneliness in older individuals, the sample being comprised of aging adults, many of whom are retired (60%), is an important consideration. Participants were recruited in tandem with their twins, so only 21 twins were part of a “broken pair” (i.e., their cotwin is not present in the data). Thus, the final analytic sample consisted of 1,356 twin persons, constituting 678 twin pairs and 21 persons whose cotwin did not participate. Of the 678 twin pairs, 420 pairs (262 female and 158 male) were monozygotic (MZ) and 258 (160 female and 98 male) were dizygotic (DZ). All twin pairs were of the same biological sex.
Measures
UCLA Loneliness Scale Version 3
The UCLA Loneliness Scale is a 20-item self-report questionnaire designed to index an individual’s subjective feelings of loneliness (Russell, 1996). Items include, for example, “How often do you feel that you lack companionship?” and “How often do you feel that there is no one you can turn to?,” rated on a 4-point Likert scale (1 = never; 2 = rarely; 3 = sometimes; 4 = often). Of the 20 items, nine are reverse scored to index loneliness (e.g., “How often do you feel part of a group of friends?”). The 20-item scale demonstrated high internal consistency in the sample (α = .93, ωT = .94). We tested four confirmatory factor analytic models of the scale’s structure and concluded that a unidimensional loneliness factor was optimal after accounting for method effects that correspond to the valence of each item (i.e., negative vs. positive valence, the latter requiring reverse scoring to index loneliness). These analyses are discussed in supplemental materials and reported in Table S2 in the online supplemental material.
PID-5 Short Form
The PID-5 is a self-report measure assessing the 25 maladaptive personality trait facets and five superordinate domains of criterion B of the AMPD (Krueger et al., 2012). Items are rated on a 4-point Likert scale (0 = very or often false; 1 = sometimes or somewhat false; 2 = sometimes or somewhat true; 3 = very or often true). Originally developed as a 220-item measure, item-response theory-based analyses were used to adapt the 100-item shortened form (which retained similar reliability and criterion validity properties) that was used in this study (Maples et al., 2015). Four items are averaged to index each of the facets, and three characteristic facets are averaged to index each of the domains. For example, the grandiosity, deceitfulness, and manipulativeness facets are measured by four items and then averaged to index the antagonism domain. Some PID-5 facets are “interstitial,” meaning they are located between domains (e.g., depressivity is considered primarily a facet of detachment, but, in factor analyses, has moderately large loadings onto negative affect as well). Facets that have been demonstrated to contribute primarily to a given domain (i.e., are less interstitial; have smaller cross-loadings) are used in scoring. The five domains each had moderate to high internal consistency (.80 ≤ α ≤ .87, .83 ≤ ωT ≤ .90). Most of the facets displayed adequate internal consistency (α and ωT ≥ .55 for 24 facets; α and ωT ≥ .70 for 17 facets). Irresponsibility did not, however (α = .38, ωT = .42), nor did any three-item subset of its four items, so analyses should be interpreted cautiously.1
Data Preparation
After participants submitted their scores via Qualtrics or on a paper questionnaire (and subsequently entered into Qualtrics), data were screened by hand for careless responding by the investigators using intuitive approaches (e.g., implausible response consistency, inconsistency, and/or endorsement of an impossible response option to an attention item). After anomalous responses were flagged, research assistants conducted a second assessment shortly after where participants either corrected or confirmed their entries. Following this second assessment, all participant responses were deemed sufficiently valid and were used in the analyses. In addition, prior to conducting other analyses, data were inspected for meeting the statistical assumption of normality. Visual inspection of the scale histograms and calculation of skewness (i.e., skew coefficient g > 0.75) suggested that each of the PID-5 domains and 23 of the 25 PID-5 facets had a positive skew (all except restricted affectivity and submissiveness).2 The loneliness measure was not skewed. Biometric models that decompose variance into additive genetic (A), dominant genetic (D), common environmnetal (C), and unique environmnetal (E) variance components (i.e., ACE and ADE models) may have decreased accuracy for non-normally distributed variables (Arbet et al., 2020), so a rank-based Blom transformation was applied to skewed variables (Blom, 1958; Wright et al., 2017). As recommended for biometric analyses, all variables were then regressed on sex, the linear and quadratic effects of age, and the age–sex interaction. Regressing out these effects is recommended to remove similarity between twins that is due to being the same age and sex, and thus serves to avoid overestimating twin intraclass correlations (ICCs; McGue & Bouchard, 1984). The standardized residuals of these regressions were then carried forward to the analyses.
Statistical Analysis
First, we calculated bivariate and semipartial phenotypic correlations between the pathological personality traits and loneliness. Then, for each construct, we calculated MZ and DZ twin ICCs to compare the similarity across twins for a given phenotype. The greater similarity among MZ twin pairs than DZ twin pairs suggests genetic influences on the phenotype. Next, we estimated univariate ACE or ADE and AE variance decompositions. When the DZ ICC was less than half of the MZ ICC, an ADE rather than ACE model was fit. Personality traits (both maladaptive and normative) and loneliness often show little to no common environmental components when examining broad, average variance estimates (Boomsma et al., 2006; Wright et al., 2017), suggesting that a more parsimonious model that excludes a common environmental component (C) might fit the data well (i.e., an AE model). Thus, for each phenotype either an ACE and AE or ADE and AE model was fit. We used the Akaike information criteria (AIC) to compare the ACE or ADE model to the simpler AE model for each phenotype; the model with the lowest AIC was determined to best fit the observed data. In estimating these models, the direct symmetric approach was used, which estimates model variance components rather than model paths because it is less likely to produce biased estimates (Verhulst et al., 2019).
After modeling the univariate decompositions, we then estimated bivariate ACE, ADE, and AE “Cholesky decompositions” of the covariance between loneliness and the personality traits (Loehlin, 1996). The Cholesky model can be transformed to estimate various parameters. Because we were interested in the overlap between influences on personality pathology and loneliness, we calculated genetic and environmental correlations (i.e., correlations between the latent variance components), and the degree to which each phenotypic correlation was attributable genetic and environmental factors (e.g., proportion of phenotypic covariance attributable to common additive genetic effects). Therefore, for each pair (i.e., given personality trait and loneliness), the best fitting of these models was interpreted as a “correlated factors” model to estimate genetic and environmental correlations as well as a “common and specific factors” model to estimate the percentage of phenotypic covariance mediated by genetic and environmental factors (Loehlin, 1996). Finally, we fit a multivariate Cholesky decomposition involving each of the AMPD domains (modeled first ordinally) and loneliness (modeled last). Having hypothesized that loneliness largely results from similar forces as personality pathology, this model allowed us to estimate the proportion of genetic and environmental variance in loneliness that was attributable to the combined effects of the genetic and environmental variance components of the five measured personality domains.
Transparency and Openness
Our sample size results from the complete, recent MTR wave of data collection (n = 1,377), without data exclusions apart from participants without a twin in the dataset (n = 21) and missingness (2 ≤ n ≤ 20). All analysis code and model results are available at https://osf.io/y6k94/ (Freilich, 2023). Biometric models were estimated in R using the OpenMx package (Boker et al., 2022), modifying sample scripts by Dr. Hermine Maes (https://hermine-maes.squarespace.com/). This study’s design and its analysis were not preregistered. Study procedures were approved by the Institutional Review Board of the University of Minnesota.
Results
Phenotypic Associations
Phenotypic correlations between loneliness and the maladaptive personality trait domains are reported in Table 1. Correlations are provided both before and after transformation and regression procedures. The personality domains each displayed moderate to strong correlations with loneliness (.22 ≤ r ≤ .58), with associations being strongest with detachment (r = .58) and negative affect (r = .46). Phenotypic associations between loneliness and the narrow personality trait facets were of a similar magnitude (.13 ≤ r ≤ .57) and are reported in Table 3.
Table 1.
Correlations Among Loneliness and the AMPD Domains
| Variable | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
|
| ||||||
| Age | −0.04 | −0.02 | 0.05 | −0.07 | −0.04 | −0.04 |
| Sex (1 = male, 2 = female) | 0.00 | 0.01 | 0.00 | −0.24 | −0.04 | −0.14 |
| 1. Loneliness | 0.94 | 0.46 | 0.59 | 0.21 | 0.39 | 0.34 |
| 2. Negative affect | 0.46 | 0.89 | 0.46 | 0.41 | 0.61 | 0.47 |
| 3. Detachment | 0.58 | 0.48 | 0.90 | 0.27 | 0.45 | 0.38 |
| 4. Antagonism | 0.22 | 0.46 | 0.32 | 0.83 | 0.45 | 0.45 |
| 5. Disinhibition | 0.39 | 0.62 | 0.47 | 0.47 | 0.84 | 0.54 |
| 6. Psychoticism | 0.37 | 0.49 | 0.42 | 0.45 | 0.54 | 0.84 |
Note. Correlations are reported above the diagonal before transformation procedures and regressions that control for age and sex and below the diagonal after such procedures. McDonald’s omega total reliability estimate is reported along the diagonal. n ≥ 1,360 pairwise, inclusive of participants without a twin in the dataset. Later tables include only the approximately 669–677 (n = 1,338–1,354) twin pairs. AMPD = alternative model for personality disorder.
Table 3.
Bivariate Relations Between Personality Pathology and Loneliness
| Variable | Phenotypic correlation (% mediated through A) | Semipartial correlation (rs) | Genetic correlation [95% CI] | Unique environmental correlation [95% CI] |
|---|---|---|---|---|
|
| ||||
| Negative affect | 0.46* (48%) | .15* | 0.60 [0.46–0.72] | 0.38 [0.30–0.45] |
| Anxiousness | 0.41* (49%) | .11* | 0.52 [0.38–0.65] | 0.35 [0.27–0.42] |
| Emotional lability | 0.39* (50%) | .07 | 0.61 [0.44–0.78] | 0.29 [0.21–0.37] |
| Hostility | 0.41* (49%) | .14* | 0.59 [0.43–0.73] | 0.32 [0.24–0.40] |
| Perseveration | 0.40* (62%) | .07 | 0.70 [0.56–0.84] | 0.24 [0.15–0.32] |
| Restricted affectivity | 0.28* (57%) | .15* | 0.46 [0.30–0.63] | 0.19 [0.11–0.28] |
| Separation anxiety | 0.34* (45%) | .07* | 0.53 [0.34–0.66] | 0.27 [0.19–0.35] |
| Submissiveness | 0.25* (64%) | −.01 | 0.43 [0.27–0.59] | 0.15 [0.06–0.23] |
| Detachment | 0.58* (48%) | .38* | 0.75 [0.64–0.84] | 0.48 [0.41–0.55] |
| Anhedonia | 0.57* (45%) | .17* | 0.73 [0.60–0.83] | 0.48 [0.41–0.55] |
| Depressivity | 0.49* (43%) | .11* | 0.76 [0.59–0.92] | 0.40 [0.32–0.47] |
| Intimacy avoidance | 0.39* (43%) | .08* | 0.53 [0.35–0.69] | 0.33 [0.24–0.40] |
| Suspiciousness | 0.42* (55%) | .10* | 0.73 [0.57–0.88] | 0.28 [0.19–0.36] |
| Withdrawal | 0.53* (54%) | .19* | 0.73 [0.62–0.79] | 0.40 [0.32–0.47] |
| Antagonism | 0.22* (71%) | −.07 | 0.45 [0.27–0.62] | 0.10 [0.01–0.19] |
| Attention seeking | 0.14* (93%) | .03 | 0.38 [0.20–0.56] | 0.02 [−0.07–0.10] |
| Callousness | 0.25* (37%) | .17* | 0.35 [0.09–0.48] | 0.22 [0.13–0.30] |
| Deceitfulness | 0.24* (72%) | .15* | 0.62 [0.41–0.85] | 0.10 [0.01–0.10] |
| Grandiosity | 0.20* (62%) | .06 | 0.39 [0.19–0.59] | 0.11 [0.02–0.20] |
| Manipulativeness | 0.13* (77%) | −.04 | 0.31 [0.11–0.51] | 0.05 [−0.04–0.13] |
| Disinhibition | 0.39* (63%) | .03 | 0.64 [0.51–0.78] | 0.24 [0.15–0.32] |
| Distractibility | 0.38* (58%) | .23* | 0.59 [0.44–0.73] | 0.18 [0.26–0.34] |
| Impulsivity | 0.24* (69%) | .02 | 0.51 [0.33–0.65] | 0.11 [0.02–0.20] |
| Irresponsibility | 0.29* (75%) | .12* | 0.74 [−0.95–0.99] | 0.10 [0.01–0.19] |
| Rigid perfectionism | 0.23* (65%) | .10* | 0.40 [0.24–0.56] | 0.14 [0.05–0.22] |
| Risk taking | 0.13* (66%) | −.02 | 0.27 [0.07–0.48] | 0.07 [−0.02–0.15] |
| Psychoticism | 0.37* (47%) | .08* | 0.55 [0.38–0.71] | 0.30 [0.22–0.38] |
| Eccentricity | 0.38* (51%) | .29* | 0.58 [0.42–0.74] | 0.29 [0.20–0.37] |
| Perceptual dysregulation | 0.23* (29%) | .06 | 0.22 [−0.86–0.36] | 0.24 [0.15–0.32] |
| Unusual beliefs and experiences | 0.23* (46%) | .03 | 0.48 [0.21–0.71] | 0.17 [0.09–0.25] |
Note. Percentage of phenotypic covariance explained by shared additive genetic influences (A) reported alongside phenotypic correlation. In semipartials, covariation was removed with other traits in the same scale (e.g., semipartial with detachment removes covariation with other four PID-5 domains. Semipartial with anhedonia removes covariation with other four detachment facets). Correlations are reported after transformation procedures and after regressions that control for age and sex. Maximum likelihood 95% confidence intervals in brackets; those which do not contain zero are deemed to be significant (bolded). CI = confidence intervals; PID-5 = Personality Inventory for DSM-5.
p < .01.
Semipartial correlations were calculated to estimate the association between each of the domains, net of the other four domains, with loneliness. Semipartial correlations may suggest the degree to which a given domain uniquely predicts loneliness. Semipartial correlations with loneliness are reported in Table 3 and were statistically significant (at p < .01) for detachment (r = .38), negative affect (r = .15), and psychoticism (r = .08), but not for either antagonism or disinhibition. Similarly, we estimated semipartial correlations within each domain (e.g., association between loneliness and anxiousness, net of the other facets of negative affect). This was done to infer the degree to which facets of a given domain uniquely predict loneliness, accounting for the other facets in that domain (Table 3).
Criterion-Predictor Overlap
Some of the items from the loneliness scale could be interpreted as overlapping with negative affect or detachment items. For instance, the loneliness item “How often do you feel that there is no one you can turn to?” could theoretically assess alienation. In order to assess criterion-predictor overlap, we calculated correlations between each of the loneliness items and each of the facets of detachment and negative affect. Detachment results are reported in Table S3 in the online supplemental material, and negative affect results are reported in Table S4 in the online supplemental material.
Detachment and its facets had consistent correlations across the 20 loneliness items (detachment r’s .29–.46; anhedonia r’s .19–.45; withdrawal r’s .27–.51; depressivity r’s .15–.41; suspiciousness r’s .12–.32; intimacy avoidance r’s .16–.33). Overall, the item “How often do you feel close to people?”3 was most associated with the detachment facets (average r = .38), while the item “How often do you feel shy?” was least associated (average r = .22). Similarly, negative affect and its facets had relatively consistent correlations across the 20 loneliness items (negative affect r’s .16–.40; anxiousness r’s .19–.35; hostility r’s .16–.33; perseveration r’s .17–.32; emotional lability r’s .11–.33; separation anxiety r’s .09–.34; restricted affectivity r’s .14–.28; submissiveness r’s .09–.25). The loneliness item “How often do you feel isolated from others?” was most associated with the negative affect facets (average r = .30), while “How often do you feel shy?” was least associated (average r = .16). Because no individual loneliness item stood out as especially overlapping with any particular facet, we proceeded with a loneliness measurement model that utilized all 20 items.
Univariate Biometric Analyses
For loneliness and each of the domains and facets, the MZ ICC (.16 ≤ r ≤ .42) was greater than the DZ ICC (−.08 ≤ r ≤ .24), suggesting genetic influences on the phenotypes. Given this pattern of ICCs, either an ACE or ADE model (depending on if MZ ICC > 2 × DZ ICC) and a reduced AE model were fit and compared based on AIC. ICCs, model results, and fit statistics are reported in Table 2. In 28 of the 31 phenotypes, the reduced AE model had a lower AIC, suggesting the genetic dominance (D) and shared environmental (C) variance components were not meaningfully greater than zero and could be dropped from the model. Heritability estimates ranged from .14 to .41, with unique environment influences accounting for the remaining variance (.59 ≤ E ≤ .86).
Table 2.
Univariate Parameter Estimates and Fit Statistics for Loneliness and the AMPD Traits
| Variable | MZ pairs | DZ pairs | MZ ICC | DZ ICC | A [95% Cl] | D [95% Cl] | C [95% Cl] | E [95% Cl] | AIC |
|---|---|---|---|---|---|---|---|---|---|
|
| |||||||||
| Loneliness | 419 | 258 | 0.37 | 0.16 | 0.24 [−0.22–0.69] | 0.14 [−0.32–0.63] | — | 0.62 [0.54–0.71] | 3.778.2 |
| 0.38 [0.28–0.47] | — | — | 0.62 [0.55–0.71] | 3,776.5 | |||||
| Negative affect | 415 | 254 | 0.36 | 0.22 | 0.27 [0.00–0.56] | — | 0.08 [−0.16–0.32] | 0.64 [0.56–0.73] | 3.727.4 |
| 0.37 [0.28–0.46] | — | — | 0.63 [0.56–0.71] | 3,725.9 | |||||
| Anxiousness | 415 | 254 | 0.41 | 0.24 | 0.28 [0.00–0.57] | — | 0.12 [−0.14–0.36] | 0.60 [0.53–0.68] | 3.713.0 |
| 0.41 [0.32–0.49] | — | — | 0.59 [0.52–0.67] | 3,711.9 | |||||
| Emotional lability | 415 | 254 | 0.26 | 0.18 | 0.20 [−0.09–0.49] | — | 0.07 [−0.18–0.31] | 0.73 [0.64–0.83] | 3.760.0 |
| 0.28 [0.19–0.37] | — | — | 0.72 [0.64–0.81] | 3,758.3 | |||||
| Hostility | 415 | 254 | 0.34 | 0.10 | 0.09 [−0.43–0.60] | 0.24 [−0.28–0.79] | — | 0.67 [0.59–0.76] | 3.754.2 |
| 0.32 [0.23–0.41] | — | — | 0.68 [0.60–0.77] | 3,753.0 | |||||
| Perseveration | 415 | 254 | 0.34 | 0.20 | 0.31 [0.04–0.61] | — | 0.04 [−0.21–0.27] | 0.65 [0.57–0.75] | 3.744.1 |
| 0.35 [0.26–0.45] | — | — | 0.65 [0.57–0.73] | 3,742.2 | |||||
| Restricted affectivity | 415 | 254 | 0.33 | 0.14 | 0.21 [−0.28–0.68] | 0.13 [−0.36–0.64] | — | 0.66 [0.58–0.76] | 3.753.1 |
| 0.33 [0.24–0.42] | — | — | 0.67 [0.59–0.76] | 3,751.4 | |||||
| Separation anxiety | 415 | 254 | 0.22 | 0.12 | 0.17 [−0.13–0.49] | — | 0.04 [−0.23–0.30] | 0.79 [0.69–0.89] | 3.772.7 |
| 0.22 [0.13–0.30] | — | — | 0.78 [0.69–0.88] | 3,770.8 | |||||
| Submissiveness | 415 | 254 | 0.38 | 0.16 | 0.23 [−0.26–0.69] | 0.16 [−0.32–0.67] | — | 0.62 [0.54–0.71] | 3.735.2 |
| 0.37 [0.28–0.47] | — | — | 0.63 [0.55–0.71] | 3,733.6 | |||||
| Detachment | 415 | 254 | 0.38 | 0.21 | 0.31 [0.04–0.61] | — | 0.06 [−0.20–0.30] | 0.63 [0.55–0.71] | 3.728.7 |
| 0.38 [0.29–0.47] | — | — | 0.62 [0.55–0.70] | 3,726.9 | |||||
| Anhedonia | 415 | 254 | 0.35 | 0.14 | 0.29 [−0.24–0.79] | 0.04 [−0.47–0.59] | — | 0.67 [0.58–0.75] | 3.744.8 |
| 0.33 [0.24–0.42] | — | — | 0.67 [0.59–0.75] | 3,742.8 | |||||
| Depressivity | 415 | 254 | 0.21 | 0.14 | 0.14 [−0.16–0.44] | — | 0.07 [−0.20–0.32] | 0.79 [0.69–0.89] | 3.762.3 |
| 0.21 [0.12–0.30] | — | — | 0.79 [0.69–0.87] | 3,760.5 | |||||
| Intimacy avoidance | 415 | 254 | 0.25 | 0.17 | 0.15 [−0.14–0.45] | — | 0.10 [−0.16–0.35] | 0.75 [0.66–0.85] | 3.764.8 |
| 0.26 [0.17–0.35] | — | — | 0.74 [0.65–0.83] | 3,763.4 | |||||
| Suspiciousness | 415 | 254 | 0.24 | 0.14 | 0.25 [−0.21–0.71] | 0.00 [−0.48–0.50] | — | 0.75 [0.65–0.85] | 3.772.3 |
| 0.25 [0.16–0.35] | — | — | 0.75 [0.66–0.84] | 3,770.3 | |||||
| Withdrawal | 415 | 254 | 0.42 | 0.18 | 0.28 [−0.19–0.74] | 0.14 [−0.32–0.64] | — | 0.57 [0.50–0.66] | 3.717.0 |
| 0.42 [0.33–0.52] | — | — | 0.58 [0.51–0.66] | 3,715.3 | |||||
| Antagonism | 415 | 254 | 0.35 | 0.09 | 0.00 [ 0.49–0.46] | 0.36 [ 0.13–0.87] | — | 0.65 [0.56–0.73] | 3,733.6 |
| 0.33 [0.24–0.43] | — | — | 0.67 [0.58–0.75] | 3.733.7 | |||||
| Attention seeking | 415 | 254 | 0.36 | 0.11 | 0.14 [−0.38–0.64] | 0.21 [−0.31–0.75] | — | 0.65 [0.57–0.74] | 3.739.7 |
| 0.34 [0.25–0.43] | — | — | 0.66 [0.58–0.74] | 3,738.3 | |||||
| Callousnessa | 415 | 254 | 0.23 | −0.08 | −0.57 [−1.08–0.06] | — | 0.79 [0.26–1.34] | 0.77 [0.68–0.87] | 3.772.6 |
| 0.18 [0.09–0.27] | — | — | 0.82 [0.72–0.92] | 3,779.2 | |||||
| Deceitfulness | 414 | 254 | 0.23 | 0.04 | −0.09 [−0.59–0.39] | 0.32 [−0.19–0.85] | — | 0.77 [0.67–0.87] | 3.771.0 |
| 0.21 [0.12–0.30] | — | — | 0.79 [0.69–0.89] | 3,770.5 | |||||
| Grandiositya | 415 | 254 | 0.29 | 0.03 | −0.19 [−0.68–0.28] | — | 0.49 [−0.01–1.00] | 0.61 [0.70–0.80] | 3.761.0 |
| 0.26 [0.17–0.36] | — | — | 0.74 [0.64–0.83] | 3,762.7 | |||||
| Manipulativeness | 415 | 254 | 0.31 | 0.06 | −0.05 [−0.55–0.43] | 0.35 [−0.15–0.88] | — | 0.70 [0.61–0.79] | 3.750.0 |
| 0.28 [0.19–0.37] | — | — | 0.72 [0.63–0.80] | 3,749.8 | |||||
| Disinhibition | 415 | 254 | 0.40 | 0.23 | 0.36 [0.10–0.64] | — | 0.04 [−0.20–0.27] | 0.59 [0.52–0.68] | 3.713.5 |
| 0.41 [0.32–0.50] | — | — | 0.59 [0.52–0.67] | 3,711.7 | |||||
| Distractibility | 415 | 254 | 0.39 | 0.20 | 0.36 [0.08–0.66] | — | 0.02 [−0.24–0.26] | 0.62 [0.54–0.70] | 3.729.1 |
| 0.38 [0.30–0.48] | — | — | 0.62 [0.54–0.70] | 3,727.2 | |||||
| Impulsivity | 415 | 254 | 0.26 | 0.17 | 0.21 [−0.08–0.50] | — | 0.06 [−0.18–0.30] | 0.73 [0.64–0.83] | 3.761.4 |
| 0.28 [0.19–0.37] | — | — | 0.72 [0.64–0.81] | 3,759.7 | |||||
| Irresponsibility | 415 | 254 | 0.22 | 0.06 | −0.01 [−0.47–0.44] | 0.25 [−0.23–0.75] | — | 0.76 [0.66–0.87] | 3.781.2 |
| 0.22 [0.13–0.32] | — | — | 0.78 [0.69–0.89] | 3,780.2 | |||||
| Rigid perfectionism | 415 | 254 | 0.42 | 0.12 | 0.08 [−0.43–0.56] | 0.34 [−0.15–0.87] | — | 0.59 [0.51–0.67] | 3.723.7 |
| 0.40 [0.31–0.49] | — | — | 0.60 [0.53–0.68] | 3,723.5 | |||||
| Risk taking | 415 | 254 | 0.28 | 0.07 | −0.01 [−0.52–0.48] | 0.29 [−0.23–0.82] | — | 0.72 [0.63–0.81] | 3.754.9 |
| 0.26 [0.17–0.35] | — | — | 0.74 [0.64–0.82] | 3,754.1 | |||||
| Psychoticism | 415 | 254 | 0.28 | 0.14 | 0.26 [−0.23–0.74] | 0.02 [−0.47–0.54] | — | 0.72 [0.63–0.82] | 3.765.7 |
| 0.28 [0.19–0.37] | — | — | 0.72 [0.64–0.81] | 3,763.7 | |||||
| Eccentricity | 415 | 254 | 0.28 | 0.17 | 0.29 [0.01–0.59] | — | 0.00 [−0.24–0.23] | 0.70 [0.61–0.80] | 3.763.5 |
| 0.30 [0.21–0.40] | — | — | 0.70 [0.62–0.80] | 3,761.5 | |||||
| Perceptual dysregulation | 415 | 254 | 0.24 | 0.10 | 0.14 [−0.34–0.61] | 0.11 [−0.38–0.62] | — | 0.75 [0.65–0.85] | 3.767.5 |
| 0.24 [0.15–0.34] | — | — | 0.76 [0.66–0.85] | 3,765.7 | |||||
| Unusual beliefs and experiences | 415 | 254 | 0.16 | 0.03 | −0.01 [−0.56–0.54] | — | 0.15 [−0.41–0.73] | 0.85 [0.75–0.96] | 3.791.8 |
| 0.14 [0.05–0.25] | — | — | 0.86 [0.76–0.97] | 3,790.0 | |||||
Note. AMPD = alternative model for personality disorder; MZ pairs = number of monozygotic twin pairs; DZ pairs = number of dizygotic twin pairs; MZ ICC = monozygotic intraclass correlation, or the correlation on a given trait across MZ twin pairs; DZ ICC = dizygotic intraclass correlation; AIC = Akaike information criteria; A = additive genetic; D = dominant genetic; C = common environment; E = unique environment; CI = confidence intervals. Maximum likelihood 95% confidence intervals are displayed in brackets. Full ACE or ADE models are in the first row for each variable, followed by AE models. The bolded row is considered the best-fitting model, indicated by the lowest AIC.
AE model interpreted (and bolded) despite higher AIC because ADE model provided nonsensical parameters (i.e., negative variance components), perhaps due to lower than expected DZ ICCs. Models are estimated using direct symmetric approach, which does not place lower bound of 0 on variance components estimates to avoid upward bias in estimates, at the potential risk of decreasing interpretability (Verhulst et al., 2019).
Bivariate and Multivariate Biometric Analyses
In each of the bivariate decompositions, the reduced AE model had a lower AIC than the ACE or ADE model, suggesting it had the superior fit to the data. Therefore, we proceeded with interpreting correlated factors AE models. These models allow for the calculation of additive genetic (rg) and unique environmental (re) correlations, which estimate the degree to which genetic and unique environmental components of the PID-5 traits covaried with those of loneliness. Each of the domains and many of the facets had genetic and unique environmental correlations that were significantly greater than zero, with associations being strongest for detachment and negative affect and their underlying facets. For instance, the latent additive genetic components of negative affect and of loneliness were estimated to have a correlation of rg = .60, while the latent unique environmental components had a correlation of re = .38. Each of these correlations and their 95% confidence intervals is reported in Table 3.
In addition, we interpreted common and specific factors AE models, which estimate the proportion of phenotypic covariance attributable to additive genetic and unique environmental influences. For instance, negative affect and loneliness had a phenotypic correlation of r = .46, and an estimated 48% of that covariance was mediated through additive genetic factors, while the remaining 52% was attributable to shared factors in the unique environment. For each personality trait, we report the percentage of covariance with loneliness attributable to additive genetic (A) components in Table 3 (with the remaining percentage attributable to the unique environment, given that only AE models were interpreted). Based on these percentages, we partitioned phenotypic correlations between each trait and in loneliness into A and E components to display in Figure 1. For instance, because 48% of the r = .46 correlation between negative affect and loneliness was mediated through additive genetic factors, the figure displays the .46 total as an “A component” of .22 (.46 × 48%) and an “E component” of .24 (.46 × 52%).
Figure 1.
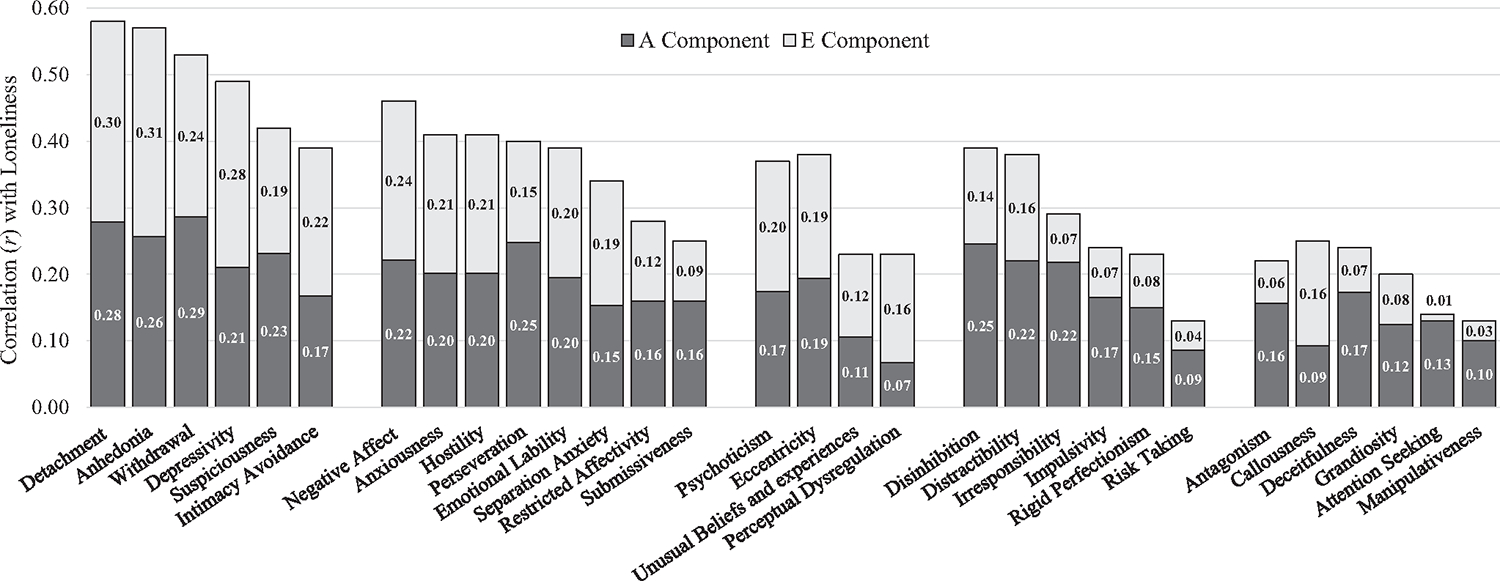
Phenotypic Correlations of AMPD Traits With Loneliness Partitioned Into Additive Genetic and Unique Environmental Components
Note. AMPD = alternative model for personality disorder.
We interpreted a reduced AE multivariate model because it had superior fit indices over an ACE or ADE model. Loneliness was the final entry in the model, so the path from the latent A variance component of loneliness to the phenotype loneliness (labeled a66 because it travels from the sixth additive genetic variance component in the model to the sixth phenotype) estimates the remaining additive genetic influences on loneliness not accounted for by the additive genetic influences of each of the previous phenotypes in the model (the five AMPD domains). The value of the a66 path was .3678, suggesting that 36.7% (.36782 [a66 path squared]/.3683[heritability of loneliness]) of the additive genetic influences on loneliness are not attributable to the AMPD domains. The e66 path similarly estimates the remaining unique environmental variance in loneliness. e66 had a value of .6647, suggesting that 70.3% of the unique environmental variance in loneliness is not shared with the AMPD domains. Full results are in Tables S5 and S6 in the online supplemental material.
Discussion
Summary of Findings
In the current sample of aging twins, loneliness was significantly associated with each of the pathological personality traits of the AMPD. Consistent with prior evidence, loneliness and each of those traits were moderately heritable, with unique rather than shared environmental influences accounting for the phenotypic variance not influenced by genetic factors (Boomsma et al., 2006; Wright et al., 2017). Further, the covariance between the traits and loneliness was partially due to shared additive genetic and unique environmental influences, suggesting some common etiological factors, though a large portion of the (especially environmental) variance was unique to loneliness. These estimates varied both within and across personality domains in interesting ways; for instance, there was moderate genetic and environmental overlap between loneliness and both detachment and negative affect, while genetic overlap with antagonism was more modest and environmental overlap was near zero.
Domain-Specific Results
Detachment
Among the broad domains, phenotypic (r = .58), semipartial (rs = .38), genetic (rg = .75), and unique environmental (re = .48) correlations with loneliness were strongest for detachment. The phenotypic associations were stronger than in previous college (r = .29; Roche et al., 2019) and adolescent (r = .21; Romero & Alonso, 2019) samples, perhaps suggesting detachment has a stronger influence on loneliness later in life, though methodological differences (e.g., daily diary assessment, briefer measures) may also contribute to the discrepancy. Perhaps as individuals age, work less, and have less family around them, their propensity toward detachment is more likely to be expressed, which in turn may lead to loneliness. Detachment has conceptual links with low extraversion and avoidant PD, both of which have had similar phenotypic associations with loneliness in previous samples (Buecker et al., 2020; Hayakawa et al., 2018). Each facet of detachment was strongly correlated with loneliness (r = .39–.57) and remained significant in the semipartial associations (rs = .08–.19) that remove covariation with the other four facets.
Large genetic (rg = .53–.76) and moderate environmental (re = .28–.48) associations were observed for each of the facets. Loneliness appears to be involved across a wide range of pathological processes relevant to detachment; associations were strongest for anhedonia, withdrawal, and depressivity, rather than intimacy avoidance and suspiciousness, but differences were modest. Genetic and unique environmental factors appear to each contribute greatly to the covariance (i.e., 43%–55% of phenotypic correlations mediated through additive genetic factors across facets4), suggesting both are relevant to the etiology. Overall, similar mechanisms appear to be involved in a person’s tendency to experience a wide range of detachment processes (e.g., social withdrawal, suspiciousness, emotional blunting) and their tendency to feel lonely.
Negative Affect
Phenotypic (r = .46), semipartial (rs = .15), genetic (rg = .60), and unique environmental (re = .38) correlations with loneliness were similarly strong for negative affect. Phenotypic associations were similar to those observed in a previous college sample (r = .47; Roche et al., 2019) and larger than those from an adolescent sample (r = .20; Romero & Alonso, 2019). Evidence is consistent with previous links between loneliness and neuroticism (Buecker et al., 2020), borderline PD (Hauschild et al., 2018), histrionic traits (Berryman et al., 2018), dependency (Overholser, 1996), and vulnerable narcissism (Kealy et al., 2022). A possibly notable difference from normal-range personality is that where neuroticism has had larger semipartial and unique environmental correlations with loneliness than low extraversion (Schermer & Martin, 2019), its conceptual partner, negative affect, had weaker associations than detachment (extraversion’s conceptual partner). Like those of detachment, each of the negative affect’s facets was significantly associated with loneliness phenotypically (r = .25–.41), genetically (rg = .43–.70), and unique environmentally (re = .15–.35), with relatively small differences across facets. Shared genetic and environmental factors contributed to the covariance to a similar degree (i.e., 45%–64% of phenotypic correlations mediated through additive genetic factors). Taken together, this similarly suggests that many of the genetic and unique environmental factors that influence a person’s tendency to experience a wide range of distinctive negative emotions (e.g., anxiousness, hostility, and emotional lability) also influence a person’s tendency to feel lonely.
Psychoticism
Psychoticism was similarly associated with loneliness phenotypically (r = .38), genetically (rg = .55), and environmentally (re = .30) and was the only other domain significantly associated net of the others (rs = .08). Phenotypic associations were comparable to that of an adolescent sample (r = .30; Romero & Alonso, 2019), larger than that of a college sample (r = .09; Roche et al., 2019), and consistent with prior evidence of strong links with schizotypy (Benson & Park, 2019) and paranoid beliefs (Alsuhibani et al., 2022). Among the facets, only eccentricity (r = .38; rs = .29) uniquely predicted loneliness net of the others (perceptual dysregulation r = .23; rs = .06; unusual beliefs and experiences r = .23; rs = .03). Unique environmental factors accounted for the covariance with loneliness to a similar degree for each of the facets (re = .17–.29), but the genetic correlation was considerably larger for eccentricity than perceptual dysregulation (rg = .58 compared to rg = .22). In total, this may suggest that links between loneliness and psychotic experiences are more heterogenous than with detachment and negative affect. Likely due to a larger shared genetic liability, eccentric beliefs and behaviors may be more relevant to the development of loneliness than other components of psychoticism.
Disinhibition
Prior evidence has not linked disinhibition specifically to loneliness (e.g., r = .06, Roche et al., 2019; r = .11; Romero & Alonso, 2019), though both borderline (Hauschild et al., 2018) and antisocial PD (Özdel et al., 2015)—which contain impulsivity in their diagnostic criteria—and low conscientiousness (Buecker et al., 2020) have been associated. In the current sample, disinhibition was moderately associated with loneliness (r = .39) with genetic (rg = .64) and unique environmental (re = .24) factors contributing to the covariance, though the association was not significant net of the other AMPD domains (rs = .03). Distractibility was most strongly associated facet in phenotypic (r = .38), semipartial (rs = .23), and unique environmental (re = .18) correlations. Interestingly, rigid perfectionism, a negative indicator of disinhibition, was positively associated with loneliness as well, perhaps suggesting that over-constraint may also be socially maladaptive. Overall, associations with the facets of disinhibition were more modest than those of other domains and likely due in larger part to overlapping genetic (rg = .27–.74; 58%–75% of covariance) rather than environmental (re = .07–.18; 25%–42%) factors.
Antagonism
Among the PID-5 domains, phenotypic (r = .22), semipartial phenotypic (rs = −.07), genetic (rg = .45), and unique environmental (re = .19) correlations with loneliness were weakest for antagonism. This is relatively consistent with the evidence of modest associations with antisocial PD (Özdel et al., 2015), antagonism in a college (r = .11; Roche et al., 2019) and adolescent (r = .06; Romero & Alonso, 2019) sample, and low agreeableness (Buecker et al., 2020). Associations with agreeableness also tend to not be statistically significant when accounting for correlated personality domains (Freilich, Mann, South, & Krueger, 2022; Schermer & Martin, 2019). Among the facets, associations with callousness (r = .25; rs = .17) and deceitfulness (r = .24; rs = .15) were strongest, perhaps suggesting that direct interpersonal harms have a greater impact on developing loneliness than disturbances related to self or identity (e.g., grandiosity and attention seeking). Apart from callousness (re = .22), these associations likely result in larger part from a shared genetic liability (62%–93% of phenotypic covariance attributable to A), rather than being linked environmentally (re− = .02–.11). Overall, associations across antagonism facets were relatively modest, and perhaps an artifact of covariation with other domains of pathological personality resulting from shared genetic influences.
Conclusions
Taken together, this evidence suggests that many if not most domains of personality pathology involve and perhaps underlie the development of loneliness to varying degrees. Given its centrality across diverse pathologies and its adverse health consequences, intervention on loneliness is becoming increasingly emphasized; further inquiry into its etiology and maintenance will likely benefit such interventions. Evidence from the normal-range personality literature suggests that broad tendencies to experience positive and negative emotions (i.e., extraversion and neuroticism) predict residual change in loneliness (Abdellaoui et al., 2019; Mund & Neyer, 2016) and that the associations are environmentally mediated (Freilich, Mann, South, & Krueger, 2022; Schermer & Martin, 2019). We hypothesized that biometric results would extend to (and perhaps be eclipsed in magnitude by) the maladaptive variants of the FFM. Results were quite similar; we observed moderate to strong phenotypic and genetic correlations and weak to moderate environmental correlations across each of the broad domains, with detachment and negative affect having the largest associations. Further, those associations were similar in magnitude to those of low extraversion and neuroticism observed elsewhere (Freilich, Mann, South, & Krueger, 2022; Schermer & Martin, 2019). This suggests that shared etiological factors (beyond genetic confounding) are involved in the development of personality traits and loneliness, and previous research suggests directionality (i.e., traits underlying loneliness development). Conclusions about causality and directionality cannot be made from such cross-sectional data, though it does appear that loneliness is a salient feature of personality pathology and PD in older adulthood.
The construct of loneliness is usually measured with stand-alone scales, meant to index a subjective feeling or emotional experience, and considered a sign or symptom of various psychopathologies (e.g., depression, borderline PD). Traditional self-report scales assess broad tendencies to feel connected and close to others (e.g., I [“never,” “rarely,” “sometimes,”, “often”] feel that I lack companionship; Russell, 1996).5 As such, interindividual differences in loneliness are largely “trait-like,” that is, though there are changes through the lifespan, the rank-order stability is quite high (similar to personality traits; Mund, Freuding et al., 2020; Mund, Lüdtke et al., 2020). Put differently, some individuals tend to experience more loneliness than others, through changing situations and environments, just as some people tend to be more neurotic than others. This is not to suggest the constructs are redundant (indeed, a large portion variance in loneliness could not be accounted for by the personality pathology domains), though traditional loneliness inventories may be well conceived as measuring the trait-like tendency to experience a high degree of negative emotion and a low degree of positive emotion in interpersonal relationships where high neuroticism and low extraversion represent those propensities broadly across contexts.
Efforts are being made in daily diary or intensively longitudinal study designs to measure the contextual and more acute or rapidly varying aspects of loneliness (e.g., Kuczynski et al., 2022). However, as traditionally measured, loneliness likely results from many of the same genetic forces that also lead to pathological personality variation. Therefore, loneliness may be conceived as a facet of personality pathology, similar to a trait like depressivity, which the PID-5 measures as a subordinate facet of the broader detachment and negative affect domains. Regardless of whether it is conceived of as a separate symptom or an inherent facet of personality pathology, loneliness appears to be concomitant with many PD traits, and those traits offer a useful organizing framework for understanding individual differences.
Limitations
The reliance on self-report scales is a clear limitation of the present work. There is likely some degree of shared method variance contributing to the observed associations. One source of shared method variance is evaluative consistency bias, or the tendency for respondents to be consistent in rating themselves as having or not having positive qualities (Anusic et al., 2009). If there was a large degree of evaluative consistency bias among respondents, we would conclude that some portion of the observed associations was artifactual rather than meaningful. Future studies would benefit from multimethod assessment, including the use of multiple informants and clinical interviews. In addition, though we evaluated the degree to which any individual loneliness items overlapped with negative affect or detachment facets, criterion-predictor overlap could similarly bias the analyses. Alternative questionnaires and means of assessments (e.g., daily diary) may provide a measure of loneliness that is less overlapping with personality.
The cross-sectional nature of the data creates limitations on inference about temporal sequencing. A unique environmental correlation suggests the degree to which co-occurrence of two phenotypes is due to factors in the environment that make identical twins differ (accounting for genetic and shared environmental similarity). Thus, these correlations are consistent with the conclusion that (a) pathological personality traits could underlie the development of loneliness, (b) loneliness could underlie the development of pathological personality traits, or (c) a third unmeasured environmental variable could underlie changes in both. We argue that this pattern of results is consistent with personality underlying the development of loneliness, given the nature of the constructs and longitudinal evidence from the normal-range personality literature.
Future work would also benefit from measuring Criterion A to study the role of loneliness in both aspects of the AMPD. This would also allow calculation of composite scores of traditional PD syndromes (e.g., specific combinations of Criterion A scores and PID-5 facets to index borderline PD) which may provide additional insight on the role of loneliness in PD, beyond what traits offer. Finally, the data were derived from a community sample, and, as result, relatively low mean levels of personality pathology were observed. Though the PID-5 was designed as a continuous measure of maladaptive personality traits (and, indeed, some evidence suggests the instrument is invariant across clinical and community samples, e.g., Bach et al., 2018; Somma et al., 2019), it is unclear if associations would generalize across PD severity.
Supplementary Material
Acknowledgments
This study was supported by a grant from the National Institute on Aging (R01-AG053217). Colin D. Freilich was supported by the National Institute on Drug Abuse (T32DA050560).
Footnotes
The authors have no conflict of interests to declare.
All analysis code and model results are available at https://osf.io/y6k94/.
This study’s design and its analysis were not preregistered.
Colin D. Freilich served as the lead for formal analysis and writing–original draft and contributed equally to methodology. Matt McGue served as the lead for writing–review and editing and served in a supporting role for conceptualization, funding acquisition, methodology, and validation. Susan C. South served in a supporting role for methodology and validation. Glenn I. Roisman served as the lead for funding acquisition and served in a supporting role for conceptualization, validation, and writing–review and editing. Robert F. Krueger served as the lead for funding acquisition, project administration, resources, software, and supervision and contributed equally to validation. Colin D. Freilich and Robert F. Krueger contributed equally to conceptualization. Susan C. South and Robert F. Krueger contributed to writing–review and editing equally.
Supplemental materials: https://doi.org/10.1037/per0000635.supp
This is likely due to limited variability in the item-level responses, but note, however, that later results involving irresponsibility are seemingly logical (e.g., moderate positive correlation with loneliness, moderate heritability).
Because this is a community sample, it is expected for participants, on average, to endorse relatively low levels of personality pathology.
Note, this item is reverse scored to index loneliness.
Though genetic correlations were greater than environmental correlations, a similar amount of the covariance was mediated through genetic and environmental factors because a greater portion of the phenotypic variances were attributable to environmental factors (i.e., heritabilities were 0.21–0.42 for detachment facets and loneliness).
The oft-used UCLA (Russell, 1996) and De Jong Giervand (De Jong Gierveld & Van Tilburg, 2010) loneliness scales notably both index general frequency (e.g., “rarely” and “often”), in contrast measures that assess symptomatology during a specific timepoint (e.g., in the past week).
References
- Abdellaoui A, Chen H-Y, Willemsen G, Ehli EA, Davies GE, Verweij KJH, Nivard MG, de Geus EJC, Boomsma DI, & Cacioppo JT (2019). Associations between loneliness and personality are mostly driven by a genetic association with neuroticism. Journal of Personality, 87(2), 386–397. 10.1111/jopy.12397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alsuhibani A, Shevlin M, Freeman D, Sheaves B, Bentall RP, & Jonason PK (2022). Why conspiracy theorists are not always paranoid: Conspiracy theories and paranoia form separate factors with distinct psychological predictors. PLoS ONE, 17(4), Article e0259053. 10.1371/journal.pone.0259053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- American Psychiatric Association. (2013). Diagnostic and statistical manual of mental disorders DSM-5 fifth edition. 10.1176/appi.books.9780890425596 [DOI] [Google Scholar]
- Anusic I, Schimmack U, Pinkus RT, & Lockwood P (2009). The nature and structure of correlations among Big Five ratings: The halo-alpha-beta model. Journal of Personality and Social Psychology, 97(6), 1142–1156. 10.1037/a0017159 [DOI] [PubMed] [Google Scholar]
- Arbet J, McGue M, & Basu S (2020). A robust and unified framework for estimating heritability in twin studies using generalized estimating equations. Statistics in Medicine, 39(27), 3897–3913. 10.1002/sim.8564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bach B, Sellbom M, & Simonsen E (2018). Personality inventory for DSM-5 (PID-5) in clinical versus nonclinical individuals: Generalizability of psychometric features. Assessment, 25(7), 815–825. 10.1177/1073191117709070 [DOI] [PubMed] [Google Scholar]
- Barchi-Ferreira Bel AM, & Osório FL (2020). The personality inventory for DSM-5: Psychometric evidence of validity and reliability-updates. Harvard Review of Psychiatry, 28(4), 225–237. 10.1097/HRP.0000000000000261 [DOI] [PubMed] [Google Scholar]
- Benson TL, & Park S (2019). Increased plasticity of bodily self-experience in individuals who may carry latent liability for schizophrenia. Schizophrenia Research, 207, 58–62. 10.1016/j.schres.2018.05.004 [DOI] [PubMed] [Google Scholar]
- Berryman C, Ferguson CJ, & Negy C (2018). Social media use and mental health among young adults. Psychiatric Quarterly, 89(2), 307–314. 10.1007/s11126-017-9535-6 [DOI] [PubMed] [Google Scholar]
- Beutel ME, Klein EM, Brähler E, Reiner I, Jünger C, Michal M, Wiltink J, Wild PS, Münzel T, Lackner KJ, & Tibubos AN (2017). Loneliness in the general population: Prevalence, determinants and relations to mental health. BMC Psychiatry, 17(1), Article 97. 10.1186/s12888-017-1262-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blom G (1958). Statistical estimates and transformed beta-variables (1st ed.). John Wiley and Sons. [Google Scholar]
- Boker SM, Neale MC, Maes HH, Wilde MJ, Spiegel M, Brick TR, Estabrook R, Bates TC, Mehta P, von Oertzen T, Gore RJ, Hunter MD, Hackett DC, Karch J, Brandmaier AM, Pritikin JN, Zahery M, Kirkpatrick RM, Wang Y, … Niesen J (2022). OpenMx: Extended structural equation modelling (Version 2.20.7). https://cran.rproject.org/web/packages/OpenMx/citation.html [Google Scholar]
- Boomsma DI, Cacioppo JT, Slagboom PE, & Posthuma D (2006). Genetic linkage and association analysis for loneliness in Dutch twin and sibling pairs points to a region on chromosome 12q23–24. Behavior Genetics, 36(1), 137–146. 10.1007/s10519-005-9005-z [DOI] [PubMed] [Google Scholar]
- Buecker S, Maes M, Denissen JJA, & Luhmann M (2020). Loneliness and the big five personality traits: A meta-analysis. European Journal of Personality, 34(1), 8–28. 10.1002/per.2229 [DOI] [Google Scholar]
- Dahlberg L, McKee KJ, Frank A, & Naseer M (2022). A systematic review of longitudinal risk factors for loneliness in older adults. Aging and Mental Health, 26(2), 225–249. 10.1080/13607863.2021.1876638 [DOI] [PubMed] [Google Scholar]
- De Jong Gierveld J, & Van Tilburg T (2010). The De Jong Gierveld short scales for emotional and social loneliness: Tested on data from 7 countries in the UN generations and gender surveys. European Journal of Ageing, 7(2), 121–130. 10.1007/s10433-010-0144-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupree J, South SC, & Oltmanns TF (2023). Late onset personality disorders in mid-life and older adults. Journal of Personality Disorders, 37(3), 304–316. 10.1521/pedi.2023.37.3.304 [DOI] [PubMed] [Google Scholar]
- First MB, Rebello TJ, Keeley JW, Bhargava R, Dai Y, Kulygina M, Matsumoto C, Robles R, Stona A-C, & Reed GM (2018). Do mental health professionals use diagnostic classifications the way we think they do? A global survey. World Psychiatry, 17(2), 187–195. 10.1002/wps.20525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freilich CD (2023, February 1). Personality pathology and loneliness. OSF https://osf.io/y6k94/ [Google Scholar]
- Freilich CD, Mann FD, & Krueger RF (2022). Comparing associations between personality and loneliness at midlife across three cultural groups. Journal of Personality, 91(3), 653–666. 10.1111/jopy.12765 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freilich CD, Mann FD, South SC, & Krueger RF (2022). Comparing phenotypic, genetic, and environmental associations between personality and loneliness. Journal of Research in Personality, 101, Article 104314. 10.1016/j.jrp.2022.104314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauschild S, Winter D, Thome J, Liebke L, Schmahl C, Bohus M, & Lis S (2018). Behavioural mimicry and loneliness in borderline personality disorder. Comprehensive Psychiatry, 82, 30–36. 10.1016/j.comppsych.2018.01.005 [DOI] [PubMed] [Google Scholar]
- Hawkley LC, & Cacioppo JT (2010). Loneliness matters: A theoretical and empirical review of consequences and mechanisms. Annals of Behavioral Medicine, 40(2), 218–227. 10.1007/s12160-010-9210-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayakawa K, Kato TA, Watabe M, Teo AR, Horikawa H, Kuwano N, Shimokawa N, Sato-Kasai M, Kubo H, Ohgidani M, Sagata N, & Kanba S (2018). Blood biomarkers of hikikomori, a severe social withdrawal syndrome. Scientific Reports, 8(1), Article 2884. 10.1038/s41598-018-21260-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hickin N, Käll A, Shafran R, Sutcliffe S, Manzotti G, & Langan D (2021). The effectiveness of psychological interventions for loneliness: A systematic review and meta-analysis. Clinical Psychology Review, 88, Article 102066. 10.1016/j.cpr.2021.102066 [DOI] [PubMed] [Google Scholar]
- Holt-Lunstad J, Smith TB, Baker M, Harris T, & Stephenson D (2015). Loneliness and social isolation as risk factors for mortality: A meta-analytic review. Perspectives on Psychological Science, 10(2), 227–237. 10.1177/1745691614568352 [DOI] [PubMed] [Google Scholar]
- Ikhtabi S, Pitman A, Toh G, Birken M, Pearce E, & Johnson S (2022). The experience of loneliness among people with a “personality disorder” diagnosis or traits: A qualitative meta-synthesis. BMC Psychiatry, 22(1), Article 130. 10.1186/s12888-022-03767-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kealy D, Woolgar S, & Hewitt JMA (2022). Investigating pathological narcissism and loneliness, and the link with life satisfaction. Scandinavian Journal of Psychology, 63(1), 32–38. 10.1111/sjop.12773 [DOI] [PubMed] [Google Scholar]
- Krueger RF, Derringer J, Markon KE, Watson D, & Skodol AE (2012). Initial construction of a maladaptive personality trait model and inventory for DSM-5. Psychological Medicine, 42(9), 1879–1890. 10.1017/S0033291711002674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krueger RF, & Johnson W (2002). The Minnesota Twin Registry: Current status and future directions. Twin Research, 5(5), 488–92. 10.1375/136905202320906336 [DOI] [PubMed] [Google Scholar]
- Kuczynski AM, Halvorson MA, Slater LR, & Kanter JW (2022). The effect of social interaction quantity and quality on depressed mood and loneliness: A daily diary study. Journal of Social and Personal Relationships, 39(3), 734–756. 10.1177/02654075211045717 [DOI] [Google Scholar]
- Kuiper JS, Zuidersma M, Oude Voshaar RC, Zuidema SU, van den Heuvel ER, Stolk RP, & Smidt N (2015). Social relationships and risk of dementia: A systematic review and meta-analysis of longitudinal cohort studies. Ageing Research Reviews, 22, 39–57. 10.1016/j.arr.2015.04.006 [DOI] [PubMed] [Google Scholar]
- Levi-Belz Y, Gvion Y, Horesh N, Fischel T, Treves I, Or E, Stein-Reisner O, Weiser M, David HS, & Apter A (2014). Mental pain, communication difficulties, and medically serious suicide attempts: A case-control study. Archives of Suicide Research, 18(1), 74–87. 10.1080/13811118.2013.809041 [DOI] [PubMed] [Google Scholar]
- Liebke L, Bungert M, Thome J, Hauschild S, Gescher DM, Schmahl C, Bohus M, & Lis S (2017). Loneliness, social networks, and social functioning in borderline personality disorder. Personality Disorders: Theory, Research, and Treatment, 8(4), 349–356. 10.1037/per0000208 [DOI] [PubMed] [Google Scholar]
- Lim MH, Penn DL, Thomas N, & Gleeson JFM (2020). Is loneliness a feasible treatment target in psychosis? Social Psychiatry and Psychiatric Epidemiology, 55(7), 901–906. 10.1007/s00127-019-01731-9 [DOI] [PubMed] [Google Scholar]
- Loehlin JC (1996). The Cholesky approach: A cautionary note. Behavior Genetics, 26(1), 65–69. 10.1007/BF02361160 [DOI] [Google Scholar]
- Lykken DT, Bouchard TJ, McGue M, & Tellegen A (1990). The Minnesota Twin Family Registry: Some initial findings. Acta Geneticae Medicae et Gemellologiae: Twin Research, 39(1), 35–70. 10.1017/S0001566000005572 [DOI] [PubMed] [Google Scholar]
- Maples JL, Carter NT, Few LR, Crego C, Gore WL, Samuel DB, Williamson RL, Lynam DR, Widiger TA, Markon KE, Krueger RF, & Miller JD (2015). Testing whether the DSM-5 personality disorder trait model can be measured with a reduced set of items: An item response theory investigation of the Personality Inventory for DSM-5. Psychological Assessment, 27(4), 1195–1210. 10.1037/pas0000120 [DOI] [PubMed] [Google Scholar]
- McGue M, & Bouchard TJ (1984). Adjustment of twin data for the effects of age and sex. Behavior Genetics, 14(4), 325–343. 10.1007/BF01080045 [DOI] [PubMed] [Google Scholar]
- Mund M, Freuding MM, Möbius K, Horn N, & Neyer FJ (2020). The stability and change of loneliness across the life span: A meta-analysis of longitudinal studies. Personality and Social Psychology Review, 24(1), 24–52. 10.1177/1088868319850738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mund M, Lüdtke O, & Neyer FJ (2020). Owner of a lonely heart: The stability of loneliness across the life span. Journal of Personality and Social Psychology, 119(2), 497–516. 10.1037/pspp0000262 [DOI] [PubMed] [Google Scholar]
- Mund M, & Neyer FJ (2016). The winding paths of the lonesome cowboy: Evidence for mutual influences between personality, subjective health, and loneliness. Journal of Personality, 84(5), 646–657. 10.1111/jopy.12188 [DOI] [PubMed] [Google Scholar]
- National Academies of Sciences. (2020). Social isolation and loneliness in older adults: Opportunities for the health care system. National Academies Press (US). 10.17226/25663 [DOI] [PubMed] [Google Scholar]
- Oltmanns TF, & Balsis S (2011). Personality disorders in later life: Questions about the measurement, course, and impact of disorders. Annual Review of Clinical Psychology, 7(1), 321–349. 10.1146/annurev-clinpsy-090310-120435 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Overholser JC (1996). The dependent personality and interpersonal problems. The Journal of Nervous and Mental Disease, 184(1), 8–16. 10.1097/00005053-199601000-00003 [DOI] [PubMed] [Google Scholar]
- Özdel K, Türkçapar MH, Guriz SO, Hamamci Z, Duy B, Taymur I, & Riskind JH (2015). Early maladaptive schemas and core beliefs in antisocial personality disorder. International Journal of Cognitive Therapy, 8(4), 306–317. 10.1521/ijct.2015.8.4.306 [DOI] [Google Scholar]
- Pérez-Rojas AE, Lockard AJ, Bartholomew TT, Janis RA, Carney DM, Xiao H, Youn SJ, Scofield BE, Locke BD, Castonguay LG, & Hayes JA (2017). Presenting concerns in counseling centers: The view from clinicians on the ground. Psychological Services, 14(4), 416–427. 10.1037/ser0000122 [DOI] [PubMed] [Google Scholar]
- Reinhard MA, Nenov-Matt T, & Padberg F (2022). Loneliness in personality disorders. Current Psychiatry Reports, 24(11), 603–612. 10.1007/s11920-022-01368-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roche MJ, Pincus AL, & Cole PE (2019). Linking dimensions and dynamics in psychopathology research: An example using DSM-5 instruments. Journal of Research in Personality, 82, Article 103852. 10.1016/j.jrp.2019.103852 [DOI] [Google Scholar]
- Romero E, & Alonso C (2019). Maladaptive personality traits in adolescence: Behavioural, emotional and motivational correlates of the PID-5-BF scales. Psicothema, 31(3), 263–270. 10.7334/psicothema2019.86 [DOI] [PubMed] [Google Scholar]
- Russell DW (1996). UCLA Loneliness Scale (Version 3): Reliability, validity, and factor structure. Journal of Personality Assessment, 66(1), 20–40. 10.1207/s15327752jpa6601_2 [DOI] [PubMed] [Google Scholar]
- Schermer JA, Colodro-Conde L, Grasby KL, Hickie IB, Burns J, Ligthart L, Willemsen G, Trull TJ, Martin NG, & Boomsma DI (2020). Genetic and environmental causes of individual differences in borderline personality disorder features and loneliness are partially shared. Twin Research and Human Genetics, 23(4), 214–220. 10.1017/thg.2020.62 [DOI] [PubMed] [Google Scholar]
- Schermer JA, & Martin NG (2019). A behavior genetic analysis of personality and loneliness. Journal of Research in Personality, 78, 133–137. 10.1016/j.jrp.2018.11.011 [DOI] [Google Scholar]
- Skaug E, Czajkowski NO, Waaktaar T, & Torgersen S (2022). The role of sense of coherence and loneliness in borderline personality disorder traits: A longitudinal twin study. Borderline Personality Disorder and Emotion Dysregulation, 9(1), Article 19. 10.1186/s40479-022-00190-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somma A, Krueger RF, Markon KE, Borroni S, & Fossati A (2019). Item response theory analyses, factor structure, and external correlates of the Italian translation of the personality inventory for DSM-5 Short Form in community-dwelling adults and clinical adults. Assessment, 26(5), 839–852. 10.1177/1073191118781006 [DOI] [PubMed] [Google Scholar]
- Tyrer P, Coombs N, Ibrahimi F, Mathilakath A, Bajaj P, Ranger M, Rao B, & Din R (2007). Critical developments in the assessment of personality disorder. The British Journal of Psychiatry, 190(S49), s51–s59. 10.1192/bjp.190.5.s51 [DOI] [PubMed] [Google Scholar]
- Valtorta NK, Kanaan M, Gilbody S, Ronzi S, & Hanratty B (2016). Loneliness and social isolation as risk factors for coronary heart disease and stroke: Systematic review and meta-analysis of longitudinal observational studies. Heart, 102(13), 1009–1016. 10.1136/heartjnl-2015-308790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verhulst B, Prom-Wormley E, Keller M, Medland S, & Neale MC (2019). Type I error rates and parameter bias in multivariate behavioral genetic models. Behavior Genetics, 49(1), 99–111. 10.1007/s10519-018-9942-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitfield KE (2013). A registry of adult African American twins: The Carolina African American Twin Study of Aging. Twin Research and Human Genetics, 16(1), 476–480. 10.1017/thg.2012.79 [DOI] [PubMed] [Google Scholar]
- Wright ZE, Pahlen S, & Krueger RF (2017). Genetic and environmental influences on Diagnostic and Statistical Manual of Mental Disorders-Fifth Edition (DSM-5) maladaptive personality traits and their connections with normative personality traits. Journal of Abnormal Psychology, 126(4), 416–428. 10.1037/abn0000260 [DOI] [PubMed] [Google Scholar]
- Yuan Y, Jiang S, Wen X, Han Z, Wu D, Wang X, Ye T, Hu Y, Jeong J, & Xiang M (2022). The chain-mediation pathway of social avoidance to depression in college students is regulated by self-esteem. Frontiers in Psychology, 13, Article 802161. 10.3389/fpsyg.2022.802161 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.