

Abstract
Background
γ-Hexachlorocyclohexane (γ-HCH), an organochlorine insecticide of anthropogenic origin, is a persistent organic pollutant (POP) that causes environmental pollution concerns worldwide. Although many γ-HCH-degrading bacterial strains are available, inoculating them directly into γ-HCH-contaminated soil is ineffective because of the low survival rate of the exogenous bacteria. Another strategy for the bioremediation of γ-HCH involves the use of transgenic plants expressing bacterial enzyme for γ-HCH degradation through phytoremediation.
Results
We generated transgenic Arabidopsis thaliana expressing γ-HCH dehydrochlroninase LinA from bacterium Sphingobium japonicum strain UT26. Among the transgenic Arabidopsis T2 lines, we obtained one line (A5) that expressed and accumulated LinA well. The A5-derived T3 plants showed higher tolerance to γ-HCH than the non-transformant control plants, indicating that γ-HCH is toxic for Arabidopsis thaliana and that this effect is relieved by LinA expression. The crude extract of the A5 plants showed γ-HCH degradation activity, and metabolites of γ-HCH produced by the LinA reaction were detected in the assay solution, indicating that the A5 plants accumulated the active LinA protein. In some A5 lines, the whole plant absorbed and degraded more than 99% of γ-HCH (10 ppm) in the liquid medium within 36 h.
Conclusion
The transgenic Arabidopsis expressing active LinA absorbed and degraded γ-HCH in the liquid medium, indicating the high potential of LinA-expressing transgenic plants for the phytoremediation of environmental γ-HCH. This study marks a crucial step toward the practical use of transgenic plants for the phytoremediation of POPs.
Supplementary Information
The online version contains supplementary material available at 10.1186/s12896-024-00867-0.
Keywords: Transgenic plants, Phytoremediation, Organochlorine pesticides, POPs, γ-HCH dehydrochlorinase, Arabidopsis thaliana
Background
γ-Hexachlorocyclohexane (γ-HCH), an organochlorine insecticide of anthropogenic origin, was used worldwide in the 1940s. Its release into the environment caused serious environmental problems due to its toxicity and long-term persistence in upland soils [1–3]. Although many countries have prohibited its use, γ-HCH remains in environments around the world [1], and γ-HCH and its byproducts, α-HCH and β-HCH, were listed as persistent organic pollutants (POPs) at the Stockholm Convention [3]. Thus, there is a global need for HCH removal and remediation actions.
Many bacterial strains, notably those from Sphingomonadaceae family, have been extensively studied at the molecular level for their ability to degrade γ-HCH and its isomers under aerobic conditions [2, 4, 5]. However, it was strongly suggested that bacterial activities in natural environments differ significantly from those under laboratory conditions [6–8]. We are now encountering problems with the practical use of bacteria in complex natural environments (e.g., bioremediation and biological control of plant diseases) since bacteria do not always perform the functions they exhibited under laboratory conditions [6, 9, 10]. Indeed, direct inoculation of bacteria into the soil can affect the viability of inoculated organisms due to factors such as temperature, humidity, pH, organic matter content, and biological predators and competitors [11].
Phytoremediation is another strategy to remove organic and inorganic contaminants from the environment [12]. The low cost of phytoremediation makes it suitable for the remediation of large areas of contaminated land. However, two problems present obstacles to the practical use of phytoremediation in soils contaminated with hydrophobic and recalcitrant compounds like γ-HCH: plants are generally unable to absorb large amounts of hydrophobic compounds, and they lack enzymes that degrade recalcitrant compounds such as γ-HCH.
Cucurbita species can take up a large number of POPs from soil [13–16]. It was reported that major latex-like proteins (MLPs) in their xylem sap are related to the efficient translocation of hydrophobic contaminants [17]. Therefore, we reasoned that generating transgenic Cucurbita species expressing γ-HCH-degrading enzymes might show promise as a new strategy for phytoremediation of γ-HCH. Our previous study produced transgenic hairy root cultures of Cucurbita moschata expressing bacterial γ-HCH dehydrochlorinase LinA [18]. In that study, we synthesized codon-optimized linA (relinA) (Accession No. LC006108), and further modified it to remove unwanted restriction enzyme recognition sites and codons that plant cells rarely use. The final relinA sequence comprised 74 (16%) modified base pairs (bp) that did not change the predicted amino acid sequence of LinA [18]. In addition, it was necessary to fuse an endoplasmic reticulum targeting signal peptide to LinA for stable accumulation in hairy roots. The resultant LinA-expressed transgenic hairy root cultures successfully degraded γ-HCH, indicating that LinA has strong potential for phytoremediation of environmental γ-HCH [18].
LinA is a unique dehydrochlorinase with no significant homologous sequence [19, 20]. LinA was originally identified as an enzyme that catalyzes the first two steps of dehydrochlorination from γ-HCH to 1,3,4,6-tetrachloro-1,4-cyclohexadiene (1,4-TCDN) via γ-pentachlorocyclohexene (γ-PCCH) (Fig. 1) in Sphingobium japonicum strain UT26 [19, 20]. This strain is one of the best characterized of the bacterial strains degrading γ-HCH and has been the focus of extensive analyses of the degradation pathway of γ-HCH [4]. 1,4-TCDN is hypothesized to undergo a nonenzymatic conversion to 1,2,4-trichlorobenzene (1,2,4-TCB) due to the inherent instability of its diene-type structure, leading to its transformation into the more stable aromatic ring structure of 1,2,4-TCB (Fig. 1) [21].
Fig. 1.
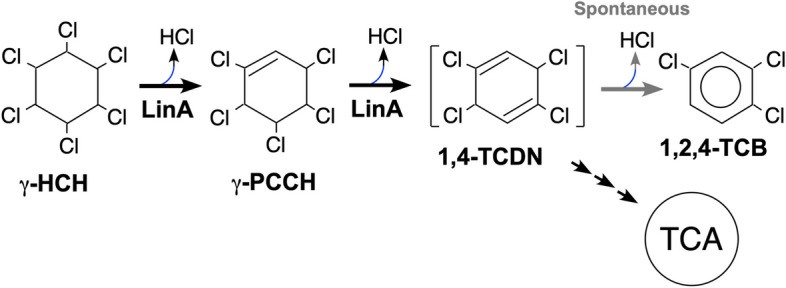
Reactions catalyzed by γ-HCH dehydrochlorinase LinA. LinA catalyzes the first two steps of the γ-HCH degradation in γ-HCH-degrading bacteria Sphingobium japonicum strain UT26 [19, 20]. LinA converts γ-HCH to 1,3,4,6-tetrachloro-1,4-cyclohexadiene (1,4-TCDN) via γ-pentachlorocyclohexene (γ-PCCH). 1,4-TCDN is further metabolized to TCA in UT26 [4]. 1,4-TCDN is hypothesized to undergo a nonenzymatic conversion to 1,2,4-trichlorobenzene (1,2,4-TCB) due to the inherent instability of its diene-type structure, leading to its transformation into the more stable aromatic ring structure of 1,2,4-TCB [21]
In addition to γ-HCH and γ-PCCH, LinA also catalyzes the dehydrochlorination of α-, δ-, and ε-HCHs and the corresponding PCCHs [20, 22, 23]. Dehydrochlorination by LinA occurs stereoselectively by acting specifically on a trans and diaxial pair of hydrogen and chlorine atoms [21, 24]. The overall three-dimensional structure of LinA predicted by computer modelling was verified by site-directed mutagenesis, and its D25, H73, and R129 residues were shown to form catalytically critical residues of LinA and to be essential for its activity [25]. Later, the crystal structure of LinA was experimentally solved, and it supported the proposed 1,2-anti dehydrochlorination of γ-HCH by LinA [26]. LinA did not dehydrochlorinate the other chlorinated compounds tested [20], indicating its narrow substrate specificity. Hexabromocyclododecanes were the only reported substrates of LinA besides HCH-related compounds [27–29]. In addition, we recently demonstrated that LinA could also convert another notorious synthetic organochlorine insecticide, 1,1,1-trichloro-2,2-bis(4-chlorophenyl)-ethane (DDT), which is also listed as a POP, to 1,1-dichloro-2,2-bis(4-chlorophenyl)-ethylene (DDE) [30], indicating the high potential of LinA for the bioremediation of POPs. LinA catalyzes the reaction without any cofactor, and this feature is an advantage for the heterologous expression of LinA [20].
In the previous study, we produced transgenic hairy root cultures of C. moschata expressing LinA [18]. For practical application, however, it is crucial to generate the whole transgenic plant expressing LinA. Thus, in this study we used the Arabidopsis thaliana plant, which grows fast and is easily modified genetically, to assess the possibility of using LinA-expressing transgenic plants for the phytoremediation of γ-HCH.
Results
Production of transgenic Arabidopsis into which the relinA gene was introduced
In the previous study, we generated hairy root cultures of C. moschata that express LinA fused to the ER-targeting signal peptide from the ascorbate oxidase of Cucumis sativus [31], named AOs hereafter [18]. Using pAOs::relinA, the same vector we used in the previous study, we transformed A. thaliana. PCR analysis confirmed the relinA gene's introduction, and six independent T2 lines (A1, A2, A4, A5, A6, and A8) of transgenic A. thaliana were obtained (Fig. 2A). We conducted subculture for the selected six T2 lines and retained 2 to 4 individual T3 plants that grew well for each line. We then conducted RT-qPCR analysis of 10 individual T3 lines for expression of relinA at the transcriptional level (Fig. 2B). Four A5-derived T3 lines (A5-1, A5-2, A5-3, and A5-4) showed apparent relinA expression at the transcriptional level. Of these, A5-3 expressed relinA at the highest level.
Fig. 2.

Production and expression analysis of transgenic A. thaliana. A PCR analysis of the genomic DNA of T2 Arabidopsis lines transformed with pAOs::relinA (lane 1 to 9 indicates T2 transgenic lines, A1 to A9, respectively). Theoretically, 1,089 bp DNA is amplified by the primer set for the relinA gene. M, 1-kb ladder of DNA size markers; WT, genomic DNA of wide type Arabidopsis; E, genomic DNA of Arabidopsis line transformed with control vector pIG-sGFP [32]; V, plasmid DNA of pAOs::relinA (positive control). The full uncropped gel images for this figure is provided as Figure S1. B RT-qPCR analysis of relinA gene transcription in T3 transgenic Arabidopsis lines. The relative expression level of relinA to that of the endogenous reference gene ACTIN2 is shown. Error bars indicate the mean ± standard deviation (SD), n = 3 (biological replicates)
Selection of transgenic Arabidopsis plants expressing LinA
Western blot analysis of LinA protein detected an approximately 18-kDa protein in four A5-derived plants (Fig. 3A), but not in wild type Arabidopsis plant (WT) or transgenic Arabidopsis E11-1 line transformed by pIG-sGFP [32]. The protein's molecular mass was larger than the 16.5-kDa molecular mass of native LinA expressed in bacterial strain UT26 [20]. In the previous study, LinA expressed in transgenic hairy root cultures of C. moschata was modified with high mannose-type N-linked oligosaccharide(s) [18]. N-glycosidase treatment using Endo H or PNGase F sifted the molecular mass to that of the native LinA expressed in bacterial cells (Fig. 3B). In contrast, the O-glycosidase treatment had no detectable effect (Fig. 3B), indicating that LinA in Arabidopsis was also modified with high mannose-type N-linked oligosaccharide(s). In addition, another band for a ca. 17-kDa protein was detected in this experiment and remained after N-glycosidase treatment (Fig. 3B), suggesting an additional unknown modification of LinA in Arabidopsis. A significantly higher amount of LinA was detected in the roots than in the leaves (Fig. 3C), indicating that LinA is accumulated more in the roots than in the leaves.
Fig. 3.

Analysis of LinA expressed in transgenic Arabidopsis. A Western blot analysis of protein extracts of T3 transgenic Arabidopsis lines transformed with pAOs::relinA (A8-1, A6-1, A5-4, A5-3, A5-2, A5-1, and A1-1) and with control empty vector (E11-1), wild type Arabidopsis (WT), and bacterial strain UT26 cells (UT26) was performed using an anti-LinA antibody. Actin was used as a loading control. Black and white triangles indicate native LinA expressed in UT26 cells and LinA expressed in transgenic Arabidopsis, respectively. B Analysis of the glycosylation of LinA expressed in transgenic Arabidopsis line. Protein extract of A5-3 line was treated with ( +) or without (-) glycopeptidases, Endo H (E), PNGase F (P), and O-glycosidase (O). Western blot analysis was performed using an anti-LinA antibody. Actin was used as a loading control. Black and white triangles indicate native LinA expressed in UT26 cells and N-glycosylated LinA expressed in transgenic Arabidopsis, respectively. Gray triangle indicates ca. 17-kDa protein expressed in transgenic Arabidopsis with an unknown modification. C Western blot analysis of protein extracts from root (R) and leaf (L) of transgenic Arabidopsis was performed using an anti-LinA antibody. Actin was used as a loading control. The full uncropped blot images for these figures are provided as Figure S2
Tolerance of transgenic Arabidopsis expressing LinA to γ-HCH
To estimate the effect of γ-HCH, the WT, E11-1, and A5-derived lines (A5-1 to A5-4) were cultivated on MS medium containing γ-HCH at three different concentrations (10, 20, and 50 ppm) (Fig. 4). No growth difference was observed in the 10 ppm γ-HCH condition (Fig. 4A). At 20 ppm, however, apparent growth defects were observed in WT and E11-1 but not in the A5-derived lines (Fig. 4A). A5-derived lines grew even under 50 ppm γ-HCH, although growth defects were observed to some extent (Fig. 4A). To quantitatively estimate the effects of γ-HCH on the plants, the fresh weights of plants grown on medium containing γ-HCH were measured (Fig. 4B). This result confirmed that Arabidopsis plants are sensitive to γ-HCH but becomes more tolerant to it by the expression of LinA.
Fig. 4.

γ-HCH tolerance of transgenic Arabidopsis expressing LinA. Effects of γ-HCH on T3 transgenic Arabidopsis lines transformed with pAOs::relinA (A5-1 to A5-4) and with control empty vector (E11-1), and wild type Arabidopsis (WT) were estimated. A The seedlings of plants were transferred to MS medium containing γ-HCH at three different concentrations (10, 20, and 50 ppm) (0 day), and cultivated for 14 days. B The wet weight of plants cultivated on the medium containing γ-HCH for 14 days was measured. Error bars indicate the mean ± standard deviation (SD), n = 3 (biological replicates)
Degradation of γ-HCH by crude extracts of Arabidopsis expressing LinA
We prepared the crude extracts of WT, E11-1, and A5-derived lines, incubated each with 10 ppm of γ-HCH, and analyzed by gas chromatography with an electron capture detector (GC-ECD). Apparent decreases in γ-HCH were observed in the A5-derived lines but not in WT and E11-1 (Fig. 5A). The activity of A5-4 was slightly weaker than those of other A5- derived lines. Furthermore, the increase and decrease in γ-PCCH during the reaction and the appearance of 1,2,4-TCB were observed only in A5-devived lines. Representative chromatograms for WT and A5-3 are shown in Fig. 5B. These results indicated that LinA proteins expressed in A5-derived lines had γ-HCH dehydrochlorinase activity.
Fig. 5.

Degradation of γ-HCH by crude extracts of Arabidopsis expressing LinA. A Crude extracts of T3 transgenic Arabidopsis lines transformed with pAOs::relinA (A5-1 to A5-4) and with control empty vector (E11-1), and wild type Arabidopsis (WT) were added to the reaction buffer containing 10 ppm of γ-HCH, and the residual amounts of γ-HCH were measured by GC-ECD. Error bars indicate the mean ± standard deviation (SD), n = 3 (biological replicates). B Representative chromatograms of GC-ECD analysis for γ-HCH degradation by crude extracts of WT (right) and A5-3 line (left) plants. Chromatograms immediately after adding crude extract into the reaction buffer (0 h) and atter incubation for 12 and 24 h are shown. Dieldrin was added as the internal standard. Note that commercially available γ-HCH chemical contains γ-PCCH, and thus γ-PCCH was also detected in a negative control sample without crude extract
Degradation of γ-HCH in liquid medium by the whole Arabidopsis plants expressing LinA
To assess the whole plant γ-HCH degradation activity of the A5-derived lines, the WT, E11-1, and A5-derived lines grown on solid medium were incubated in a liquid medium containing 10 ppm of γ-HCH. When whole plants of the A5-derived lines (A5-1, A5-2, and A5-3) were incubated in the medium, most of the γ-HCH in the medium disappeared after 36 h of incubation, unlike the case when whole plants of the WT and E11-1 lines or no plant were incubated in the medium (Fig. 6A). When whole plant of the A5-4 line was incubated in the medium, most of the γ-HCH in the medium disappeared after 36 h incubation, but a small amount of γ-HCH remained (Fig. 6A). In addition, far smaller amounts of γ-HCH were detected in the A5-derived line bodies after incubation in the medium for 36 h than in the WT and E11-1 line bodies (Fig. 6B). We estimated the total amount of γ-HCH in the assay system (Table 1). More than 99% of γ-HCH in the assay system disappeared when the A5-1, A5-2, and A5-3 lines were incubated in the medium. Despite a slightly lower reduction rate compared to the others, the A5-4 line caused more than 92% of γ-HCH in the assay system to disappear. These results indicated that the A5-derived lines can absorb and degrade γ-HCH in liquid medium.
Fig. 6.
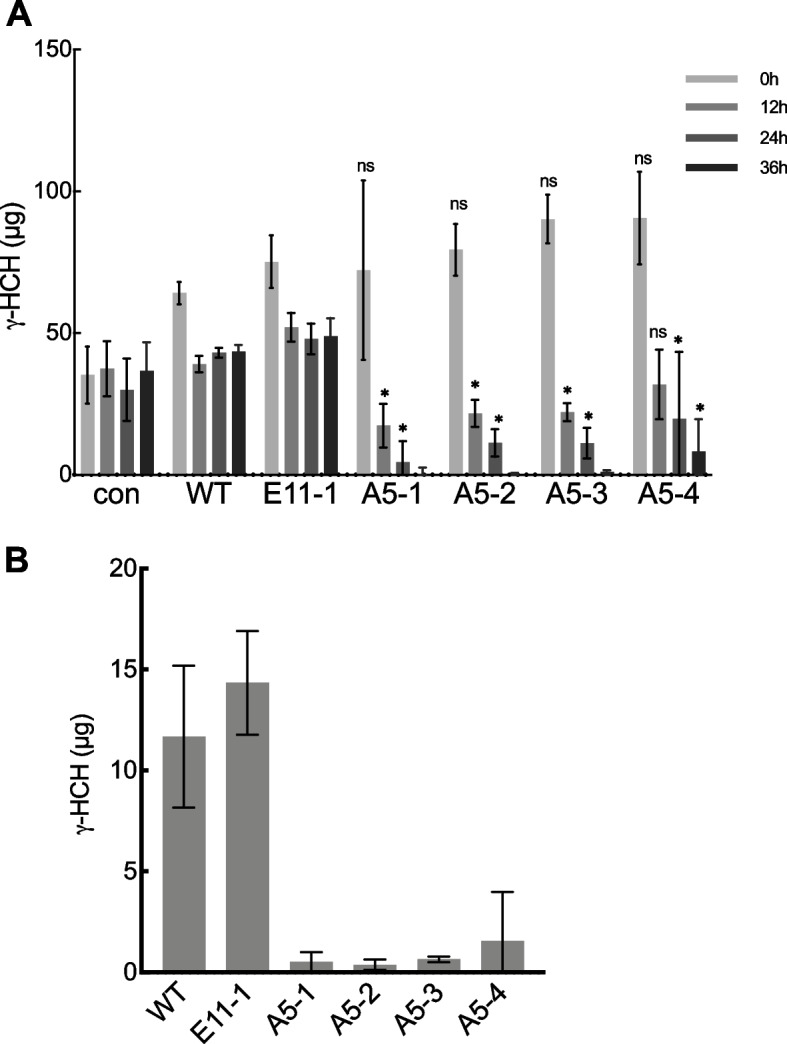
Degradation of γ-HCH in liquid medium by whole Arabidopsis expressing LinA. A Whole plants of T3 transgenic Arabidopsis lines transformed with pAOs::relinA (A5-1 to A5-4) and with control empty vector (E11-1), and wild type Arabidopsis (WT) were incubated in a liquid medium containing ten ppm of γ-HCH for 36 h. The concentration of γ-HCH in the medium was measured by GC-ECD, and total amount of γ-HCH in the medium was estimated. Error bars indicate the mean ± standard deviation (SD), n = 3 (biological replicates). Asterisks indicate significant differences from the WT detected by follow-up Two Factor ANOVA (*, P < 0.05). con, control without plant. B Total amount of γ-HCH in each plant body after incubation for 36 h was estimated. Error bars indicate the mean ± standard deviation (SD), n = 3 (biological replicates)
Table 1.
Change of total amount of γ -HCH in the assay system
| WT | E11-1 | A5-1 | A5-2 | A5-3 | A5-4 | ||
|---|---|---|---|---|---|---|---|
| Amounts of γ -HCH (µg) | |||||||
| Before incubation | Medium | 64.2 ± 1,2 | 75.2 ± 6.6 | 72.2 ± 29 | 79.4 ± 6.3 | 90.2 ± 5.8 | 90.6 ± 14 |
| After 36 h incubation |
Medium + Plant body |
55.2 ± 4 | 63.4 ± 6.1 | 0.5 ± 0.42 | 0.4 ± 0.2 | 0.7 ± 0.1 | 7.1 ± 5.9 |
| Reduction of γ -HCH(%) | 14 | 15.7 | 99.3 | 99.5 | 99.3 | 92.2 | |
Values are the mean ± standard deviation (SD), n = 3 (biological replicates)
Discussion
In the present study, we generated the transgenic Arabidopsis into which the bacterial gene encoding γ-HCH dehydrochlorinase LinA was introduced. One of the T2 generation lines (A5) expressed and accumulated LinA protein well, and the A5-derived T3 lines degraded γ-HCH rapidly in liquid medium. This study is the first report on whole transgenic plants expressing active LinA. The transgenic Nicotiana tabacum plant expressing human cytochrome P4502E1 was reported to be capable of degrading γ-HCH [33]. However, the γ-HCH degradation activity of the transgenic N. tabacum decreased quickly. One of the reasons for this may be that P4502E1 requires a cofactor, NADPH, for its reaction. It is noteworthy that LinA did not require any cofactor for its reaction [20]. Thus, transgenic plants expressing LinA are more suitable for the sustainable removal of γ-HCH from the environment.
LinA converts γ-HCH into 1,2,4-TCB. Because of the high volatility of 1,2,4-TCB, it is plausible that 1,2,4-TCB evaporates during the incubation of γ-HCH with transgenic plants expressing LinA. Indeed, we did not detect 1,2,4-TCB in the γ-HCH degradation experiment in liquid medium. Since 1,2,4-TCB is toxic to plants [34, 35], it is advantageous that 1,2,4-TCB does not remain in the plant for continuous γ-HCH degradation. Moreover, the toxicity of 1,2,4-TCB for rats is approximately 10 times lower than that of γ-HCH [36, 37]. Therefore, the conversion of γ-HCH to 1,2,4-TCB at least reduces environmental toxicity. However, the possibility remains that 1,4-TCDN and 1,2,4-TCB are converted to other substances in plant cells by unknown enzymatic or nonenzymatic reaction(s). Further research on the metabolic pathway of γ-HCH in plant cells and the toxicological properties of the metabolites would be required for practical use of transgenic plants expressing LinA. LinA cannot degrade β-HCH [20, 22, 23], but LinB degrades β-HCH. Especially, a variant of LinB (LinBMI) from Sphingobium sp. strain MI1205 degrades β-HCH more effectively compared with LinBUT from strain UT26 [38]. Co-expressing other lin genes, such as linB and linC [4], may be more efficient for the complete detoxification of an HCH-contaminated site.
Although we obtained six T2 lines into which relinA was introduced (Fig. 2A), only one plant line (A5) expressed relinA well at both transcriptional (Fig. 2B) and translational (Fig. 3A) levels. The genome positions into which the relinA gene was integrated and the copy number of the introduced relinA gene may be important for the stable expression of LinA in plant cells. Even among the T3 A5-derived lines, A5-4 showed weaker LinA activity than the other three A5-derived lines (Figs. 4, 5 and 6). Although the reason remains unclear, selecting a stable line that expresses LinA well among many transgenic plant lines is crucial for the practical phytoremediation.
LinA was modified by glycosylation with high mannose-type N-linked oligosaccharide(s) in the hairy roots culture of C. moschata [18]. LinA expressed in A. thaliana also underwent the same glycosylation, although LinA expressed in Arabidopsis may have undergone additional modification(s) (Fig. 3B). The glycosylation probably does not significantly affect LinA activity since the predicted glycosylation residue N137 is not an amino acid residue critical to the LinA reaction [18]. Indeed, LinA expressed in Arabidopsis showed γ-HCH dehydrochlorinase activity without glycosidase treatment. Glycosylation may be important for the stable accumulation of LinA in plant cells.
WT Arabidopsis and the E11-1 line were sensitive to γ-HCH at more than 20 ppm of γ-HCH on the solid medium (Fig. 4). Since the A5-derived lines expressing active LinA showed tolerance to γ-HCH, γ-HCH itself seems toxic towards the Arabidopsis plant. The phytotoxic effects of γ-HCH on other plants have also been reported [33, 39]. γ-HCH causes nerve excitation symptoms by blocking an insect's central nervous system GABA (γ-aminobutyric acid) receptor [40]. However, the effects of γ-HCH on plants varied depending on the species [39], and the toxic mechanism of γ-HCH in plants remains unknown.
In a preliminary experiment, we assessed the ability of the A5-derived lines to degrade γ-HCH in soil. Although no apparent reduction of γ-HCH in soil was observed, the amounts of γ-HCH in plant bodies of the A5-derived lines were lesser than that of WT plants after cultivation in γ-HCH-polluted soil, suggesting that γ-HCH absorbed from soil was degraded in the plant bodies of the A5-derived lines (data not shown). LinA expressed in the transgenic Arabidopsis plant accumulated more in the roots than in the leaves (Fig. 3C). Although the reason for this remains unclear, this feature benefits soil's phytoremediation of γ-HCH. In any case, absorption of γ-HCH from soil is crucial for the practical use of transgenic plants. Therefore, generating whole transgenic Cucurbita species expressing LinA is a reasonable strategy for phytoremediation of γ-HCH in soil, as suggested previously [18].
Conclusion
The successful establishment of whole transgenic Arabidopsis plants expressing and accumulating active LinA is a crucial step toward the practical use of transgenic plants in the phytoremediation of γ-HCH. We demonstrated that these plants absorbed and degraded γ-HCH in a liquid medium. This study confirms the possibility of a new strategy for the use of transgenic plants for the degradation of γ-HCH in the environment. We believe that the present results will play a vital role for the practical application of transgenic plants to remediate POPs.
Methods
Plant materials and growth conditions
Seeds of Arabidopsis thaliana (Columbia ecotype) were surface-sterilized with 20% NaClO for 15 min, followed by washing five times with sterile water. The sterilized Arabidopsis seeds were put on Murashige and Skoog (MS) medium with 0.8% agar in Petri dish. After incubation at 4°C for 72 h in darkness, the Petri dish was transferred into a growth incubator for germination and development (22°C, 16 h light/8 h dark). After incubation for 7 days, the seedlings were transplanted into soil and grew in a growth chamber (22°C, 16 h light/8 h dark).
Construction of transgenic Arabidopsis plants
The vector pAOs::relinA [18] was introduced into Agrobacterium tumefaciens strain EHA105 by electroporation [41]. Using the resultant strain, A. thaliana plants were transformed by floral dip method [42]. Seeds of T0 transgenic plants were screened on MS medium containing 50 mg/L of kanamycin and carbenicillin 100 mg/L, after which the T1 plant seedlings were transferred to soil. Finally, the homozygous genotypes of T2 transgenic plants were obtained from self-fertilization, and homozygous lines were identified in the T3 generation via segregation analysis. One of the T3 transgenic Arabidopsis lines (E11-1) transformed by pIG-sGFP [32] was used as a negative control (empty) line.
DNA extraction from Arabidopsis and PCR analysis
Genomic DNA of Arabidopsis was extracted from the leaves of 14- to18-day-old plants as described previously [43]. PCR analysis was performed using a primer set (AOs::relinA, 5′- CCTAGAAGCTAATTCCCGATCTAG-3′ and 5′- AAGGCCATCGTTGAAG-3′) as follow: 94 °C for 2 min, 30 cycles at 94 °C for 30 s, 52 °C for 30 s, and at 68 °C for 150 s, and a final extension step at 68 °C for 5 min. PCR products were analyzed by electrophoresis using 1.0% agarose gel.
RNA extraction from Arabidopsis and RT-qPCR analysis
Total RNA of Arabidopsis was extracted from 14-day-old plants by RNeasy Plant Mini Kit (Qiagen) and treated with DNase I (Takara) at 37 °C for 1 h to eliminate contaminated genomic DNA. The total RNA was reverse transcribed using the SuperScript II First-Strand Sythesis System (Thermo Fisher Scientific) with an Oligo dot Primer. Real-time DNA amplification using primer sets (relinA, 5′-CGCTGACAAAGTGAACGGTA-3′ and 5′-TAGTTCGTGCATGCATTCCT-3′; ACTIN2, 5′-GCACCACCTGAAAGGAAGTACA-3′ and 5′-CGATTCCTGGACCTGCCTCATC-3) was monitored using Bio-Rad CFX Maestro software (Bio-Rad Laboratories). The expression level of the relinA gene was normalized to that of the endogenous reference gene ACTIN2.
Preparation of crude extract from Arabidopsis and bacterial cells
Leaves and roots of 14- to 18-day-old Arabidopsis were ground using a disposable pestle (Bio-Bik), and their total soluble proteins were extracted with an extraction buffer containing 50 mM HEPES–KOH (pH7.5), 100 mM NaCl, 2.5 mM EDTA, 1 mM PMSF, 20 µM Leupeptin, and 1 mM DTT. The homogenate was then centrifuged at 12,000 rpm at 4 °C for 2 min to remove debris. Total proteins were extracted from bacterial strain UT26 as follows: UT26 cells grown at 30 °C in 1/3LB (3.3 g of Bacto tryptone, 1.7 g of yeast extracts, and 5 g of sodium chloride per liter) were resuspended in 50 mM potassium phosphate buffer (pH 7.5), and disrupted by sonication (Branson Sonifier 250A). After centrifugation, the supernatant was used as crude extract. Protein concentration was estimated by Bradford method with bovine γ-globulin as a standard.
Western blot analysis
Protein samples were separated using SDS-PAGE with 15% gel and transferred to PVDF membranes (Trans-Blot Turbo Trans Pack, Bio-Rad Laboratories) using Trans-Blot Turbo System (Bio-Rad Laboratories). Membranes were blocked for 30 min in 5%(w/v) nonfat dry milk in TBS-T containing 25 mM Tris–HCl (pH7.4), 0.15 M NaCl, and 0.05%(v/v) Tween-20. The membranes were incubated with an anti-LinA antibody [44] (1: 2,000 dilution) or an anti-Actin (Plant) antibody (1:3,000 dilution, Sigma-Aldrich) in Solution I for primary antibody (Can Get Signal™ Immunoreaction Enhancer Solution, Toyobo) at 4 °C overnight, followed by incubation with anti-rabbit IgG HRP or anti-mouse IgG HPR (Thermo Fisher Scientific) in Solution II for secondary antibody (Can Get Signal™ Immunoreaction Enhancer Solution, Toyobo) at room temperature for 2 h. Antigen–antibody complexes were detected using an Chemi-Lumi Ultra (Nacalai Tesque).
LinA glycosylation analysis
The total soluble proteins were deglycosylated by treatment with Endo H, PNGase F and O-glycosidase (New England BioLabs) under denaturing conditions according to the manufacturer’s instructions. Western blot analysis was performed for the treaded samples.
γ-HCH tolerance assay using whole plants
Seeds of Arabidopsis were germinated on MS medium with 0.8% agar containing 50 mg/L of kanamycin and grew in an incubator for germination and development (22 °C, 16 h light/8 h dark). After cultivation for 14 days, the whole plants were transplanted to MS medium with 0.8% agar containing 10, 20 and 50 ppm of γ-HCH, and incubated for development (22 °C, 16 h light/8 h dark). Fresh weights of the whole plants after 14 days cultivation were measured.
γ-HCH degradation assay using crude extracts
Leaves and roots of 14- to 18-day-old Arabidopsis plants were immersed in liquid nitrogen and ground to fine powder using a mortar and pestle. The fine powder was mixed with extraction buffer containing 50 mM potassium phosphate buffer (pH 7.5) and centrifuged at 6,200 rpm at 4 °C for 10 min. The supernatant was used as the crude extract. In a 1 mL glass reaction vial with a mininat valve (Osaka Chemical, Osaka, Japan), 200 µL of the crude extract was mixed with 800 µL glycine–NaOH buffer (pH 8.6). After 1 µL of 10 mg/mL γ-HCH was added to the solution, the mixture was incubated at 30 °C. At appropriate time points, 100 µL was sampled and then mixed with an equivalent volume of ethyl acetate containing 2 ppm of dieldrin as the internal standard. After centrifugation, the ethyl acetate layer was recovered and analyzed by GC-ECD.
GC-ECD analysis
GC-ECD analysis was conducted using a GC-17A gas chromatograph (Shimadzu, Kyoto, Japan) and a Rtx-1 capillary column (30 m × 0.25 mm × 0.25 µm; Restek, Bellafonte, PA, USA). The column temperature was increased from 160 °C to 280 °C at a rate of 40 °C/min, and the gas flow rate was 30 mL/min. γ-HCH was purchased from Tokyo Chemical Industry Co. (Tokyo, Japan).
γ-HCH degradation assay using whole plants
Seeds of Arabidopsis plants were germinated in MS medium with 0.8% agar containing 50 mg/L of kanamycin and grew in a incubator for germination and development (22 °C, 16 h light/8 h dark). After cultivation for 14 days, the whole plants were transferred to 10 mL of MS liquid medium containing 10 ppm of γ-HCH (100 µg of γ-HCH in the assay system), and incubated at 22 °C on a rotary shaker (100 rpm) for 36 h. At appropriate time points, 100 µL of the medium was sampled and then mixed with an equivalent volume of ethyl acetate containing 2 ppm of dieldrin as the internal standard. The ethyl acetate layer was recovered after centrifugation and analyzed by GC-ECD. After incubation for 36 h, whole plant was immersed in liquid nitrogen and ground to fine powder using a mortar and pestle. The fine powder was mixed with extraction buffer containing 50 mM potassium phosphate buffer (pH 7.5) and then mixed with an equivalent volume of ethyl acetate containing 2 ppm of dieldrin as the internal standard. After centrifugation, the ethyl acetate layer was recovered and analyzed by GC-ECD.
Supplementary Information
Acknowledgments
Disclosure statement
No potential conflict of interest was reported by the authors.
Authors’ contributions
Wenhao Deng, Yoshihiko Nanasato, Yutaka Tabei, Masao Watanabe and Yuji Nagata conceived the study; Wenhao Deng, Yoshinobu Takada and Kouhei Kishida performed the experiments; Wenhao Deng, Yoshinobu Takada, Leonardo Stari and Yuji Nagata analyzed the data and wrote the manuscript with help of Kouhei Kishida, Yoshiyuki Ohtsubo, Yoshihiko Nanasato, Yutaka Tabei and Masao Watanabe, and all authors have read and approved the final manuscript.
Funding
This work was supported by Institute for Fermentation, Osaka (IFO) (Grant ID: K-2016–004 to Yuji Nagata), Grants-in-Aid for Scientific Research (B) from the Japan Society for the Promotion of Science (Grant ID: 19H02865 and 22H02233 to Yuji Nagata), and the Ministry of Agriculture, Forestry and Fisheries of Japan (Genomics for Agricultural Innovation, GMB0002 to Yoshihiko Nanasato, Yutaka Tabei, and Yuji Nagata). This work was supported in part by MEXT KAKENHI (Grant Numbers JP22H05172 and JP22H05179 to Masao Watanabe), JSPS KAKENHI (Grant Numbers JP21H02162 and JP23K18058 to Masao Watanabe; JP19K05963 and JP22K05581 to Yoshinobu Takada).
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Declarations
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Phillips TM, Seech AG, Lee H, Trevors JT. Biodegradation of hexachlorocyclohexane (HCH) by microorganisms. Biodegradation. 2005;16:363–392. doi: 10.1007/s10532-004-2413-6. [DOI] [PubMed] [Google Scholar]
- 2.Lal R, Pandey G, Sharma P, Kumari K, Malhotra S, Pandey R, Raina V, Kohler HP, Holliger C, Jackson C, Oakeshott J. Biochemistry of microbial degradation of hexachlorocyclohexane and prospects for bioremediation. Microbiol Mol Biol Rev. 2010;74:58–80. doi: 10.1128/MMBR.00029-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vijgen J, Abhilash PC, Li YF, Lal R, Forter M, Torres J, Singh N, Yunus M, Tian C, Schaffer A, Weber R. Hexachlorocyclohexane (HCH) as new Stockholm Convention POPs-a global perspective on the management of Lindane and its waste isomers. Environ Sci Pollut Res Int. 2011;18:152–162. doi: 10.1007/s11356-010-0417-9. [DOI] [PubMed] [Google Scholar]
- 4.Nagata Y, Endo R, Ito M, Ohtsubo Y, Tsuda M. Aerobic degradation of lindane (gamma-hexachlorocyclohexane) in bacteria and its biochemical and molecular basis. Appl Microbiol Biotechnol. 2007;76:741–752. doi: 10.1007/s00253-007-1066-x. [DOI] [PubMed] [Google Scholar]
- 5.Tabata M, Ohhata S, Nikawadori Y, Kishida K, Sato T, Kawasumi T, Kato H, Ohtsubo Y, Tsuda M, Nagata Y. Comparison of the complete genome sequences of four γ -hexachlorocyclohexane-degrading bacterial strains: insights into the evolution of bacteria able to degrade a recalcitrant man-made pesticide. DNA Res. 2016;23:581–599. doi: 10.1093/dnares/dsw041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.van Veen J, van Overbeek L, van Elsas J. Fate and activity of microorganisms introduced into soil. Mirobiol Mol Biol Rev. 1997;61:121–135. doi: 10.1128/mmbr.61.2.121-135.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rediers H, Rainey PB, Vanderleyden J, De Mot R. Unraveling the secret lives of bacteria: use of in vivo expression technology and differential fluorescence induction promoter traps as tools for exploring niche-specific gene expression. Microbiol Mol Biol Rev. 2005;69:217–261. doi: 10.1128/MMBR.69.2.217-261.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nagata Y, Senbongi J, Ishibashi Y, Sudo R, Miyakoshi M, Ohtsubo Y, Tsuda M. Identification of Burkholderia multivorans ATCC 17616 genetic determinants for fitness in soil by using signature-tagged mutagenesis. Microbiology. 2014;160:883–891. doi: 10.1099/mic.0.077057-0. [DOI] [PubMed] [Google Scholar]
- 9.Ramos JL, Marques S, van Dillewijn P, Espinosa-Urgel M, Segura A, Duque E, Krell T, Ramos-Gonzalez MI, Bursakov S, Roca A, Solano J, Fernadez M, Luis Niqui J, Pizarro-Tobias P, Wittich RM. Laboratory research aimed at closing the gaps in microbial bioremediation. Trends Biotechnol. 2011;29:641–647. doi: 10.1016/j.tibtech.2011.06.007. [DOI] [PubMed] [Google Scholar]
- 10.Tyagi M, da Fonseca MM, de Carvalho CC. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation. 2011;22:231–241. doi: 10.1007/s10532-010-9394-4. [DOI] [PubMed] [Google Scholar]
- 11.Mrozik A, Piotrowska-Seget Z. Bioaugmentation as a strategy for cleaning up of soils contaminated with aromatic compounds. Microbiol Res. 2010;165:363–375. doi: 10.1016/j.micres.2009.08.001. [DOI] [PubMed] [Google Scholar]
- 12.Pilon-Smits E. Phytoremediation. Annu Rev Plant Biol. 2005;56:15–39. doi: 10.1146/annurev.arplant.56.032604.144214. [DOI] [PubMed] [Google Scholar]
- 13.White JC. Plant-facilitated mobilization and translocation of weathered 2,2-bis(p-chlorophenyl)-1,1-dichloroethylene (p, p′-DDE) from an agricultural soil. Environ Toxicol Chem. 2001;20:2047–2052. doi: 10.1897/1551-5028(2001)020<2047:pfmato>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 14.Mattina MI, Isleyen M, Eitzer BD, Iannucci-Berger W, White JC. Uptake by cucurbitaceae of soil-Bome contaminants depends upon plant genotype and pollutant properties. Environ Sci Technol. 2006;40:1814–1821. doi: 10.1021/es051572s. [DOI] [PubMed] [Google Scholar]
- 15.Otani T, Seike N, Sakata Y. Differential uptake of dieldrin and endrin from soil by several plant families and genera. Soil Sci Plant Nutr. 2007;53:86–94. doi: 10.1111/j.1747-0765.2007.00102.x. [DOI] [Google Scholar]
- 16.Namiki S, Otani T, Seike N. Fate and plant uptake of persistent organic pollutants in soil. Soil Sci Plant Nutr. 2013;59:669–679. doi: 10.1080/00380768.2013.813833. [DOI] [Google Scholar]
- 17.Inui H, Sawada M, Goto J, Yamazaki K, Kodama N, Tsuruta H, Eun H. A major latex-like protein is a key factor in crop contamination by persistent organic pollutants. Plant Physiol. 2013;161:2128–2135. doi: 10.1104/pp.112.213645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Nanasato Y, Namiki S, Oshima M, Moriuchi R, Konagaya K, Seike N, Otani T, Nagata Y, Tsuda M, Tabei Y. Biodegradation of γ -hexachlorocyclohexane by transgenic hairy root cultures of Cucurbita moschata that accumulate recombinant bacterial LinA. Plant Cell Rep. 2016;35:1963–1974. doi: 10.1007/s00299-016-2011-1. [DOI] [PubMed] [Google Scholar]
- 19.Imai R, Nagata Y, Fukuda M, Takagi M, Yano K. Molecular cloning of a Pseudomonas paucimobilis gene encoding a 17-kilodalton polypeptide that eliminates HCl molecules from γ -hexachlorocyclohexane. J Bacteriol. 1991;173:6811–6819. doi: 10.1128/jb.173.21.6811-6819.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nagata Y, Hatta T, Imai R, Kimbara K, Fukuda M, Yano K, Takagi M. Purification and characterization of γ-hexachlorocyclohexane (γ-HCH) dehydrochlorinase (LinA) from Pseudomonas paucimobilis. Biosci Biotech Biochem. 1993;57:1582–3. [DOI] [PubMed]
- 21.Nagasawa S, Kikuchi R, Nagata Y, Takagi M, Matsuo M. Stereochemical analysis of γ-HCH degradation by Pseudomonaspaucimobilis UT26. Chemosphere. 1993;26:1187–201.
- 22.Wu J, Hong Q, Sun Y, Hong Y, Yan Q, Li S. Analysis of the role of LinA and LinB in biodegradation of γ -hexachlorocyclohexane. Environ Microbiol. 2007;9:2331–2340. doi: 10.1111/j.1462-2920.2007.01350.x. [DOI] [PubMed] [Google Scholar]
- 23.Bala K, Geueke B, Miska ME, Rentsch D, Poiger T, Dadhwal M, Lal R, Holliger C, Kohler HP. Enzymatic conversion of ε-hexachlorocyclohexane and a heptachlorocyclohexane isomer, two neglected components of technical hexachlorocyclohexane. Environ Sci Technol. 2012;46:4051–4058. doi: 10.1021/es204143x. [DOI] [PubMed] [Google Scholar]
- 24.Trantirek L, Hynkova K, Nagata Y, Murzin A, Ansorgova A, Sklenar V, Damborsky J. Reaction mechanism and stereochemistry of γ -hexachlorocyclohexane dehydrochlorinase LinA. J Biol Chem. 2001;276:7734–7740. doi: 10.1074/jbc.M007452200. [DOI] [PubMed] [Google Scholar]
- 25.Nagata Y, Mori K, Takagi M, Murzin AG, Damborsky J. Identification of protein fold and catalytic residues of γ-hexachlorocyclohexane dehydrochlorinase LinA. Proteins. 2001;45:471–477. doi: 10.1002/prot.10007. [DOI] [PubMed] [Google Scholar]
- 26.Okai M, Kubota K, Fukuda M, Nagata Y, Nagata K, Tanokura M. Crystal structure of γ -hexachlorocyclohexane Dehydrochlorinase LinA from Sphingobium japonicum UT26. J Mol Biol. 2010;403:260–269. doi: 10.1016/j.jmb.2010.08.043. [DOI] [PubMed] [Google Scholar]
- 27.Heeb NV, Wyss SA, Geueke B, Fleischmann T, Kohler HE, Lienemann P. LinA2, a HCH-converting bacterial enzyme that dehydrohalogenates HBCDs. Chemosphere. 2014;107:194–202. doi: 10.1016/j.chemosphere.2013.12.035. [DOI] [PubMed] [Google Scholar]
- 28.Heeb NV, Wyss SA, Geueke B, Fleischmann T, Kohler HE, Bernd Schweizer W, Moor H, Lienemann P. Stereochemistry of enzymatic transformations of (+)β- and (-)β-HBCD with LinA2-α-HCH-degrading bacterial enzyme of Sphingobium indicum B90A. Chemosphere. 2015;122:70–78. doi: 10.1016/j.chemosphere.2014.11.008. [DOI] [PubMed] [Google Scholar]
- 29.Heeb NV, Hubeli J, Fleischmann T, Lienemann P, Nayyar N, Lal R, Kohler HE. Transformation of ε-HBCD with the Sphingobium Indicum enzymes LinA1, LinA2 and LinATM, a triple mutant of LinA2. Chemosphere. 2021;267:129217. doi: 10.1016/j.chemosphere.2020.129217. [DOI] [PubMed] [Google Scholar]
- 30.Yusuf Habibullah KO, Ito R, Stari L, Kishida K, Ohtsubo Y, Masai E, Fukuda M, Miyauchi K, Nagata Y. Degradation of DDT by γ -hexachlorocyclohexane dehydrochlorinase LinA. Biosci Biotechnol Biochem. 2023;88:123–130. doi: 10.1093/bbb/zbad141. [DOI] [PubMed] [Google Scholar]
- 31.Ohkawa J, Okada N, Shinmyo A, Takano M. Primary structure of cucumber (Cucumis sativus) ascorbate oxidase deduced from cDNA sequence: homology with blue copper proteins and tissue-specific expression. Proc Natl Acad Sci U S A. 1989;86:1239–1243. doi: 10.1073/pnas.86.4.1239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nanasato Y, Konagaya K, Okuzaki A, Tsuda M, Tabei Y. Agrobacterium-mediated transformation of kabocha squash (Cucurbita moschata Duch) induced by wounding with aluminum borate whiskers. Plant Cell Rep. 2011;30:1455–64. [DOI] [PMC free article] [PubMed]
- 33.Singh S, Sherkhane PD, Kale SP, Eapen S. Expression of a human cytochrome P4502E1 in Nicotiana tabacum enhances tolerance and remediation of γ -hexachlorocyclohexane. N Biotechnol. 2011;28:423–9. [DOI] [PubMed]
- 34.Liu YJ, Yuan Y, Zheng J, Tao YZ, Dong ZG, Wang JH, Wang GY. Signal peptide of potato PinII enhances the expression of Cry1Ac in transgenic tobacco. Acta Biochim Biophys Sin (Shanghai) 2004;36:553–558. doi: 10.1093/abbs/36.8.553. [DOI] [PubMed] [Google Scholar]
- 35.Miguel AS, Faure M, Ravanel P, Raveton M. Biological responses of maize (Zea mays) plants exposed to chlorobenzenes. Case study of monochloro-, 1,4-dichloro- and 1,2,4-trichloro-benzenes. Ecotoxicology. 2012;21:315–24. [DOI] [PubMed]
- 36.Gaines TB. Acute toxicity of pesticides. Toxicol Appl Pharmacol. 1969;14:515–534. doi: 10.1016/0041-008X(69)90013-1. [DOI] [PubMed] [Google Scholar]
- 37.European Union Risk Assessment Report (2003) 1,2,4-Trichlorobenzene. https://publications.jrc.ec.europa.eu/repository/handle/JRC24374. Accessed 15 Feb 2024
- 38.Ito M, Prokop Z, Klvana M, Otsubo Y, Tsuda M, Damborsky J, Nagata Y. Degradation of β-hexachlorocyclohexane by haloalkane dehalogenase LinB from γ -hexachlorocyclohexane-utilizing bacterium Sphingobium sp. MI1205. Arch Microbiol. 2007;188:313–325. doi: 10.1007/s00203-007-0251-8. [DOI] [PubMed] [Google Scholar]
- 39.Pereira RC, Monterroso C, Macías F. Phytotoxicity of hexachlorocyclohexane: effect on germination and early growth of different plant species. Chemosphere. 2010;79:326–333. doi: 10.1016/j.chemosphere.2010.01.035. [DOI] [PubMed] [Google Scholar]
- 40.Matsumura F, Ghiasuddin SM. Evidence for similarities between cyclodiene type insecticides and picrotoxinin in their action mechanisms. J Environ Sci Health B. 1983;18:1–14. doi: 10.1080/03601238309372355. [DOI] [PubMed] [Google Scholar]
- 41.Hood EE, Gelvin SB, Melchers LS, Hoekema A. New Agrobacterium helper plasmids for gene-transfer to plants. Transgenic Res. 1993;2:208–18.
- 42.Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16:735–743. doi: 10.1046/j.1365-313x.1998.00343.x. [DOI] [PubMed] [Google Scholar]
- 43.Edwards K, Johnstone C, Thompson C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991;19:1349. doi: 10.1093/nar/19.6.1349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nagata Y, Futamura A, Miyauchi K, Takagi M. Two different types of dehalogenases, LinA and LinB, involved in γ-hexachlorocyclohexane degradation in Sphingomonas paucimobilis UT26 are localized in the periplasmic space without molecular processing. J Bacteriol. 1999;181:5409–5413. doi: 10.1128/JB.181.17.5409-5413.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.