

Latin America has more than twice as many vascular plant species as Africa south of the Sahara plus Madagascar, in a smaller area.
Abstract
We compare the numbers of vascular plant species in the three major tropical areas. The Afrotropical Region (Africa south of the Sahara Desert plus Madagascar), roughly equal in size to the Latin American Region (Mexico southward), has only 56,451 recorded species (about 170 being added annually), as compared with 118,308 recorded species (about 750 being added annually) in Latin America. Southeast Asia, only a quarter the size of the other two tropical areas, has approximately 50,000 recorded species, with an average of 364 being added annually. Thus, Tropical Asia is likely to be proportionately richest in plant diversity, and for biodiversity in general, for its size. In the animal groups we reviewed, the patterns of species diversity were mostly similar except for mammals and butterflies. Judged from these relationships, Latin America may be home to at least a third of global biodiversity.
Detailed studies over the past several decades have made it possible to compare accurately the numbers of species of indigenous vascular plants in Africa, Latin America, and Southeast Asia, and the results are unexpected. In summary, the Afrotropical Region (Africa south of the Sahara with Madagascar), with an area of 22,657,000 km2, is home to 56,451 plant species. These were enumerated accurately following the development of the regularly updated African Plant Database (1) and the Madagascar Catalogue (2), both regional nomenclature repositories with standardized taxonomy. For Latin America (Mexico south, including the West Indies), with 19,197,000 km2, a similar effort (3) has recorded 118,308 known species. These registers are kept updated; the numbers provided here are current. Thus, Latin America is home to more than twice as many plant species as the Afrotropical Region in an area only about 85% of its size. Tropical Africa is comparatively even poorer in species, since 9582 of the species recorded in the Afrotropical Region occur only in Madagascar (587,000 km2), and 16,405 of the species occur only in Southern Africa (2,704,000 km2), with 41 species shared uniquely between Madagascar and Southern Africa. There are also 4330 plant species shared uniquely between Tropical and Southern Africa. These figures demonstrate the well-known floristic richness of Southern Africa. If we subtract the plant species found only in Madagascar and Southern Africa from the total, Tropical Africa, with an area of 19,366,000 km2, is home to only 30,423 plant species in an area essentially the same size as Latin America, which has some 3.8 times as many species of plants—the difference between the two areas is growing rapidly due to the higher rate of discovery of new species in Latin America. Although Latin America is ecologically more diverse than mainland tropical Africa, the latter includes rainforest, coastal forest, extensive dry plains, and the mountains of East and Southern Africa. There is no area in Africa, however, nearly as rich in plant species as the Andes—a true major area of endemism that remains relatively poorly known biologically [see also (4)].
A recent review of floras that are currently being prepared for Southeast Asia (excluding Bangladesh, India, and Bhutan but including from Myanmar and Thailand west to the Philippines and New Guinea), with an area of 5,708,000 km2, shows that this region is home to approximately 50,000 indigenous species of vascular plants (5, 6). In contrast, the entire Afrotropical Region, with an area almost four times as large, has only 56,451 recorded species and is much better known than its Asian counterpart.
Before analyzing the reasons for these remarkable disparities, it will be useful to compare the figures for vascular plants (1, 3, 5–7) with those for selected groups of animals, ones that are well known enough to make possible meaningful comparisons. To obtain the most accurate figures possible for these groups, we consulted the available literature (8–19) and then discussed each group with leading global experts. By doing so, we have been able to present the most up-to-date figures for each of them that are available anywhere.
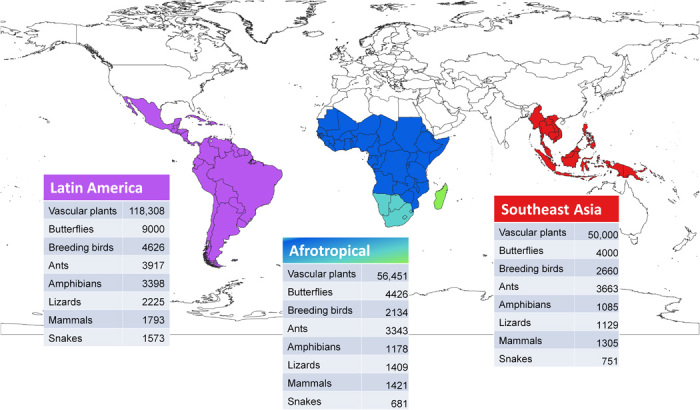
Current estimates of the species present in each area mentioned are summarized in Table 1 and illustrated in Fig. 1. The number of Afrotropical species that occur only in Madagascar (i.e., are not shared with either tropical or Southern Africa) is listed separately. In general, the regional abundances of species of breeding birds, lizards, and snakes are roughly in line with those for vascular plants. However, there are some notable differences between the patterns we reviewed for the groups listed.
Table 1. Estimated total numbers of described species for main taxonomic groups across selected tropical regions (1, 3, 5–19).
| Taxonomic group | Latin America* | Afrotropical Region† |
Afrotrop spp. only in
Madagascar |
Percentage
Mad/Afrotrop‡ |
Southeast Asia |
| Vascular plants | 118,308 | 56,451 | 9582 | 17% | 50,000 |
| Butterflies | 9000 | 4426 | 320 | 7% | 4000 |
| Breeding birds | 4626 | 2134 | 263 | 12% | 2660 |
| Ants | 3917 | 3343 | 867 | 26% | 3663 |
| Amphibians | 3398 | 1178 | 341 | 29% | 1085 |
| Lizards | 2225 | 1409 | 337 | 24% | 1129 |
| Mammals | 1793 | 1421 | 246 | 17% | 1305 |
| Snakes | 1573 | 681 | 109 | 16% | 751 |
*Mexico southward through all of South America, including the West Indies.
†That is, including Madagascar.
‡Percentage contribution of Madagascar to Afrotropical Region species total, rounded to the nearest percentage.
Fig. 1. Map of the world showing the three tropical major regions with a summary of the biodiversity richness for each, with the three floristically distinctive subunits of the Afrotropical Region: Tropical Africa (dark blue), Southern Africa (pale blue), and Madagascar (green).
For Madagascar, there are fewer species of butterflies recorded than would be expected from the number in the Afrotropical region, but the ratio for moths resembles that in other groups (28,529 regionally, 4681 in Madagascar = 17.2%). There are many more species of amphibians in the Neotropics than one would estimate from their numbers in other tropical regions. Significantly, there are relatively more African mammals compared with their Neotropical counterparts. While rodents resemble other groups in this respect [870 species (Latin America) versus 430 species (Afrotropical region)], primates (144 versus 192) and, especially, hoofed animals, Cetartiodactyla (27 versus 89), present unusual distribution patterns.
Part of the difference in mammalian species diversity between Latin America and Tropical Africa (cf. Cetartiodactyla) certainly can be attributed to the extermination of most of the large mammals of the Neotropical region by the humans who arrived there some 10,000 years ago (20). In Africa, the entire course of evolution of hominids over the past 6 to 8 million years took place in the presence of large mammals, many of which survive to the present. In addition, North American mammals originally invaded South America following the elevation of the Panama land bridge some 3.5 million years ago, with uplands developing about 2.2 million years ago (21). North American mammals invaded South America in far greater numbers than Eurasian ones invaded Africa, possibly another factor in reducing the diversity of the surviving South American mammals. In contrast, the reason that fewer ant species are recorded in the Neotropics than would be expected may well be that they remain understudied there.
For every group of organisms, except butterflies, Madagascar has remarkably many species relative to the rest of the Afrotropical Region, of which it constitutes just over 1/40th the size. Considering that there are probably at least 3000 additional species of vascular plants to be discovered and named in Madagascar (2), Madagascar’s contribution to the vascular plant flora of the Afrotropical Region is probably actually more than 20%.
Considering that a rapidly growing 118,308 species of vascular plants are known from Latin America and that the global total now is about 384,000 species (22), it seems likely that a third of more or all existing vascular plant species occur in Latin America and that the same may hold true for biodiversity in general. How many species that would imply depends on estimates for the total existing number of eukaryotes and so cannot be made with any degree of precision at this point.
It is important now to turn to a consideration of how well known vascular plants are in each of the three tropical regions we have analyzed [Table 2 (1–3, 7, 23, 24) and Fig. 2]. On average, 173 newly described species have been added to the flora of the Afrotropical Region each year since 2008, with 54 of these from Madagascar [Table 2 (1, 7)]. An average of 749 new species has been described annually from Latin America for the past 25 years, with no demonstrable tendency to level off [Table 2 (23)]. The number of newly described plant species added for Southeast Asia (5,708,000 km2) has been growing at an average of 364/year for the past 12 years, more than twice as many as for the entire Afrotropical Region (22,657,000 km2), which is almost four times as large. This is a clear indication of how very rich in plant species and relatively poorly known Southeast Asia remains [see also (6)].
Table 2. Number of newly described species of vascular plants for Latin America, Southeast Asia, and specified portions of Africa, by year, 2008–2019 (1–3, 7, 23, 24).
| Year | Latin America | Southeast Asia |
Afrotropical
Region |
Tropical Africa | Southern Africa | Madagascar |
| 2008 | 697 | 298 | 190 | 94 | 46 | 50 |
| 2009 | 704 | 291 | 223 | 116 | 54 | 53 |
| 2010 | 807 | 291 | 172 | 103 | 38 | 31 |
| 2011 | 647 | 246 | 155 | 63 | 65 | 27 |
| 2012 | 633 | 401 | 134 | 65 | 27 | 42 |
| 2013 | 729 | 317 | 166 | 77 | 44 | 45 |
| 2014 | 707 | 374 | 153 | 40 | 41 | 72 |
| 2015 | 873 | 453 | 212 | 88 | 47 | 77 |
| 2016 | 833 | 404 | 178 | 71 | 22 | 85 |
| 2017 | 917 | 448 | 152 | 42 | 31 | 79 |
| 2018 | 728 | 450 | 126 | 56 | 34 | 55 |
| 2019 | 714 | 395 | 169 | 122 | 21 | 36 |
| 12-year mean | 749 | 364 | 173 | 78 | 41 | 54 |
Fig. 2. Plant species from the Andes Mountains, East Africa, Madagascar, and Malaysia.

(A) Chuquiraga jussieui at the base of volcano Chimborazo, Ecuador. Photo credit: Flora of The World. (B) Vachellia tortilis (Acacia tortilis), grazed by giraffes at the foot of Mt. Kilimanjaro. Photo credit: David C. Western. (C) Alluaudia procera spiny thicket, Berenty, Madagascar. The family Didieriaceae is highly characteristic of Madagascar. Moringa drouhardii on the left, and noxious introduced Opuntia stricta in the foreground. Photo credit: Flora of the World. (D) Rafflesia cantleyi, a unique parasitic plant, northern Malaysia rainforest. Photo credit: Charles Davis.
In recent years, the publication of new species of vascular plants by scientists living in Latin American countries has increased markedly. These increases have become possible because of freely available botanical online resources (e.g., Tropicos, The International Plant Name Index, and Biodiversity Heritage Library), low publication costs, and rapid electronic publication. At the same time, traditional studies of tropical plants have continued in Europe, North America, and Asia, as they have been increasing locally within the tropical countries themselves.
Since Latin America is somewhat smaller than the Afrotropical Region, the reasons for its far greater species richness deserve analysis. Comparing continental areas of rainforest, forest occupies about 5,500,000 km2 in the Amazon Basin, with additional extensive areas in the Chocó of Northwest South America, the Mata Atlȃntica of Brazil (originally 1,500,000 km2, but with only about 7% of the original area remaining), and especially the extraordinarily rich forests and highland vegetation of the Andes [e.g., (25), for butterflies]. In contrast, all of the rainforests in Africa occupy only about 3,900,000 km2. Most of the 5,700,000 km2 area of Southeast Asia was originally rainforest, so that species number comparisons with Africa seem reasonable. Overall, considering that, with respect to their biodiversity, tropical forests are the richest habitats on Earth, their regional representation obviously has an effect on the overall numbers of species found in the major regions considered here.
Although Latin America is somewhat smaller than the Afrotropical Region, it is home to more than twice as many species of vascular plants and of several groups of animals as well. A review of the geological and climatic history of Africa and South America provides some reasons for this major discrepancy (26). When these continents separated from their original positions as parts of Gondwanaland, about 88 million years ago, their floras, judged from the fossil record, were similar. The continents moved apart gradually, so that by the start of the Tertiary Era, 66 million years ago, they were only about 800 km apart, with islands scattered between them. Africa and Eurasia became relatively close by the middle Miocene Period, about 17 to 18 million years ago, with biotic exchange between them still limited but increasing from that time onward (27).
At the end of the Cretaceous Period, Africa and South America were low-lying and level, with moist subtropical forests extending from coast to coast. To the north, across the Tethys Sea, the plants and animals of Eurasia and North America were markedly different from their tropical counterparts. By the Eocene Period (56 to 34 million years ago), the plants and animals of the two southern continents had become significantly different (28), and they have continued to diversify further to the present. Subsequently, with the formation of the Rift Valley system of East Africa, the eastern and southern regions of the continent were gradually uplifted to their present heights, some of the mountains (e.g., Mt. Kilimanjaro, 5900 m) within the past 1 million years. The mountains of South Africa, which is topographically quite rugged, range up to 3450 m in height. Mostly subsea 17 million years ago, East Africa rose relatively rapidly to its present elevation, especially during the most recent 5 to 6 million years. As this process continued, the African mountains were attaining their present heights, and the contours of the modern Rift Valley were taking shape (29).
This extensive orogeny profoundly affected regional climates and hence the nature of local ecosystems (26). In Sub-Saharan Africa, savannas and dry woodlands replaced many of the moist forests that had originally occupied about half the total area (30). Although still biologically rich, the drier habitats have proportionately many fewer species than do tropical moist forests. As they expanded, they greatly reduced Africa’s moist forests and left many of them as separate patches [e.g., (31)]. The separation of South America from Antarctica about 55 million years ago, eventually leading to the formation of ice sheets in the south, strongly cooled the Benguela Current, running up the west coast of Africa, and caused the spread of arid climates there too. The original rainforest, now much smaller and fragmented, also became much poorer in species of plants and animals than it had been originally (26).
The events that led to the formation of the modern biota of Africa were quite distinct from those that took place in Latin America and in Southeast Asia. In Latin America, the elevation of the Andes over the past 6 to 10 million years has protected most of South America from spreading aridity, with the effects of the cold, northward-flowing Humboldt Current mainly confined to the relatively narrow strip of level lands west of the Andes. In Southeast Asia, the predominant insularity and moisture have preserved the original biological richness, which has also been accentuated by extensive regional orogeny. Because of the factors discussed, the cool periods during Pleistocene affected Africa much more severely than they did the other two major tropical regions. The well-known and extensive biological interchange between North and South America following the establishment of their Pleistocene connection and of the nearly continuous mountains now linking them has enriched the biota of the Andes greatly. Rapid speciation in these mountains, partly based on the northern immigrants, has significantly added to the overall biological richness of most groups of South American organisms (27, 28, 32).
UNRESOLVED QUESTIONS
The destruction of ecosystems is taking place so rapidly throughout the tropics that it will be difficult to obtain additional information bearing on the matters reviewed here. With the global population currently 7.8 billion, projected to increase to nearly 10 billion over the next 30 years, and our current consumption of natural productivity having reached around 175% of what is available sustainably (www.footprintnetwork.org), the future for most natural ecosystems looks dim. In Sub-Saharan Africa, the situation is especially dire; the 1.2 billion people there are estimated to double by mid-century and quadruple by the end of this century. Few predict that there will be many areas of tropical forest left by the end of the century. Therefore, to find answers to the obvious questions of how many species are there, how their ecosystems function, and so forth, we will need to do the best we can as soon as we can. Thus far, we have named no more than 10% of tropical plants and animals, and at current rates of discovery, most are likely to be long gone before we become aware of their existence. A division of effort between strengthening our knowledge of well-known groups, similar to those reviewed here, to reveal general patterns, and gaining some knowledge of the lesser-known groups by carefully designed sampling approaches, is apparently the best that we can do. But it is well worth the effort to do so, for its own sake and to provide the best possible basis for conservation.
Acknowledgments
We are grateful to the following people for the invaluable discussion on data for selected groups of animals that allow for comparison with our data with plants: A. Bauer (reptiles) and B. Fisher (ants); lepidopteran data were discussed with S. Collins, J. Dobson, J. D. Holloway, D. Lees, R. K. Rollins, R. Vane-Wright, and M. Williams. We are also grateful to D. Middleton, National Parks Board, Singapore, for the valuable discussions on Southeast Asia and to S. Miller, U.S. National Museum, Smithsonian Institution, for additional discussions. We thank C. Davidson and S. Christoph (Flora of The World), C. Davis, and D. C. Western for permitting use of their photographs. Funding: No grants were received for any aspect of this study. Author contributions: P.H.R., R.E.G., and P.B.P. contributed equally to the article, C.C. to the Afrotropical analysis, C.U.U. to the Latin American analysis, and C.N.J. to the analysis of vertebrates. Competing interests: The authors declare that they have no competing interests. Data and materials availability: Data used in the analyses are publicly available in the online databases cited in the main text.
REFERENCES AND NOTES
- 1.African Plant Database (APD), Conservatoire et Jardin botaniques de la Ville de Genève and South African National Biodiversity Institute, Pretoria (2020); www.ville-ge.ch/musinfo/bd/cjb/africa/.
- 2.P. B. Phillipson, S. Andriambololonera, P. P. Lowry II, N. Manjato, N. Rabarimanarivo, M. Rakotonirina, N. Ravololomanana, G. E. Schatz. The Madagascar Catalogue, Progress to Date and Prospects for the Future (Abstracts, XXI AETFAT Congress, Nairobi, Kenya 2017), pp. 241.
- 3.C. Ulloa Ulloa, P. Acevedo-Rodríguez, S. Beck, M. J. Belgrano, R. Bernal, P. E. Berry, L. Brako, M. Celis, G. Davidse, R. C. Forzza, S. Robbert Gradstein, O. Hokche, B. León, S. León-Yánez, R. E. Magill, D. A. Neill, M. Nee, P. H. Raven, H. Stimmel, M. T. Strong, J. L. Villaseñor, J. L. Zarucchi, F. O. Zuloaga, P. M. Jørgensen, Vascular Plants of the Americas (VPA) Website, Tropicos, Botanical Information System at the Missouri Botanical Garden, St. Louis, Missouri, USA (2020); www.tropicos.org/Project/VPA.
- 4.Brummitt N., Araújo A. C., Harris T., Areas of plant diversity—What do we know? Plants People Planet 2020, (2020). [Google Scholar]
- 5.Middleton D. J., Armstrong K., Baba Y., Balslev H., Chayamarit K., Chung R. C. K., Conn B. J., Fernando E. S., Fujikawa K., Kiew R., Luu H. T., Aung M. M., Newman M. F., Tagane S., Tanaka N., Thomas D. C., Tran T. B., Utteridge T. M. A., van Welzen P. C., Widyatmoko D., Yahara T., Wong K. M., Progress on Southeast Asia’s Flora projects. Gardens Bull. Singap. 71, 267–319 (2019). [Google Scholar]
- 6.Joyce E. M., Thiele K. R., Silk F. J. W., Crayn D. M., Checklist of the vascular flora of the Sunda-Sahul Convergence Zone. Biodivers. Data J. 8, e51094 (2020). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Madagascar Catalogue, Catalogue of the Plants of Madagascar. Missouri Botanical Garden, St. Louis, U.S.A. & Antananarivo, Madagascar (2020); www.tropicos.org/Project/Madagascar.
- 8.P. Uetz, P. Freed, J. Hosĕk, Eds., The Reptile Database 2019 (2020); www.reptile-database.org.
- 9.Vane-Wright R. I., Ecological and behavioural origins of diversity in butterflies. Symposia of the Royal Entomological Society of London 9, 56–70 (1978). [Google Scholar]
- 10.R. I. Vane-Wright, Butterflies (Natural History Museum, London, 2015), pp. 128. [Google Scholar]
- 11.Holloway J. D., A numerical investigation of the biogeography of the butterfly fauna of India, and its relation to continental drift. Biol. J. Linn. Soc. 1, 373–385 (1969). [Google Scholar]
- 12.J. D. Holloway, The biogeography of Indian butterflies, in Ecology and Biogeography in India, W. Junk, Ed. (The Hague, 1974), pp. 473–499. [Google Scholar]
- 13.Holloway J. D., Jardine N., Two approaches to zoogeography: A study based on the distribution of butterflies, birds and bats in the Indo-Australian area. Proc. Linn. Soc. Lond. 179, 153–188 (1968). [Google Scholar]
- 14.G. Lamas, La sistemática sobre mariposas (Lepidoptera: Hesperioidea y Papilionoidea) en el mundo: Estado actual y perspectivas futuras, in Contribuciones Taxonómicas en Órdenes de Insectos Hiperdiversos, J. E. Llorente, A. Lanteri, Eds. (México DF, Universidad Nacional Autónoma de México, 2008). pp. 57–70, 1 tab. [Google Scholar]
- 15.AntWeb. Version 8.12.7. California Academy of Sciences (2020); www.antweb.org/.
- 16.IUCN, The IUCN Red List of Threatened Species (2018); https://www.iucnredlist.org.
- 17.BirdLife International and Handbook of the Birds of the World, Bird species distribution maps of the world. Version 2018.1. (2018); http://datazone.birdlife.org/species/requestdis.
- 18.Goodman S. M., Benstead J. P., Updated estimates of biotic diversity and endemism for Madagascar. Oryx 39, 73–77 (2005). [Google Scholar]
- 19.S. M. Goodman, M. J. Raherilalao, A. P. Raselimanana, V. Roarimalala, Chapter 18, Advances over the past 30 years on the land vertebrates occurring in the terrestrial protected areas of Madagascar, in The Terrestrial Protected Areas of Madagascar: Their History, Description, and Biota, S. M. Goodman, M. J. Raherilalao, S. Wohlauser, Eds. (Western and Southern Madagascar – Synthesis, 2018), vol. 3, pp. 1679–1705. [Google Scholar]
- 20.Janzen D. H., Martin P. S., Neotropical anachronisms: The fruits the gomphotheres ate. Science 215, 19–27 (1982). [DOI] [PubMed] [Google Scholar]
- 21.Graham A., The role of land bridges, ancient environments, and migrations in the assembly the North American flora. J. Syst. Evol. 56, 405–429 (2018). [Google Scholar]
- 22.Nic Lughadha E., Govaerts R., Belyaeva I., Black N., Lindon H., Allkin R., Nicolson N., Counting Counts: Revised estimates of numbers of accepted species of flowering plants, seed plants, vascular plants and land plants with a review of other recent estimates. Phytotaxa 272, 82–88 (2016). [Google Scholar]
- 23.Ulloa Ulloa C., Acevedo-Rodríguez P., Beck S., Belgrano M. J., Bernal R., Berry P. E., Brako L., Celis M., Davidse G., Forzza R. C., Gradstein S. R., Hokche O., León B., León-Yánez S., Magill R. E., Neill D. A., Nee M., Raven P. H., Stimmel H., Strong M. T., Villaseñor J. L., Zarucchi J. L., Zuloaga F. O., Jørgensen P. M., An integrated assessment of the vascular plant species of the Americas. Science 358, 1614–1617 (2017). [DOI] [PubMed] [Google Scholar]
- 24.International Plant Names Index (IPNI), Kew Science (2020); www.ipni.org.
- 25.R. K. Robbins, P. A. Opler, Butterfly diversity and a preliminary comparison with bird and mammal diversity, in Biodiversity II: Understanding and Protecting our Biological Resources, M. L. Reaka-Kudla, D. E. Wilson, E. O. Wilson, Eds. (Joseph Henry Press, National Academy of Sciences, Washington, D.C. 1997), pp. 69–82. [Google Scholar]
- 26.D. I. Axelrod, P. H. Raven, Late Cretaceous and Tertiary vegetation of Africa, in Biogeography and Ecology of Southern Africa M. J. A. Werger, Ed. (W. Junk, The Hague, with D. I. Axelrod, 1978), pp. 77–130. [Google Scholar]
- 27.P. H. Raven, Plate tectonics and southern hemisphere biogeography, in Tropical Botany, K. Larsen, L. B. Holm-Nielsen, Eds. (Academic Press, 1979), pp. 3–24. [Google Scholar]
- 28.Raven P. H., Axelrod D. I., History of the flora and fauna in Latin America. Am. Sci. 63, 420–429 (1975). [Google Scholar]
- 29.Wichura H., Jacobs L. L., Lin A., Polcyn M. J., Manthi F. K., Winkler D. A., Strecker M. R., Clemens M., A 17-My-old whale constrains onset of uplift and climate change in East Africa. Proc. Natl.l Acad. Sci. U.S.A. 112, 3910–3915 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Osborne C. P., Charles-Dominique T., Stevens N., Bond W. J., Midgley G., Lehmann C. E. R., Human impacts in African savannas are mediated by plant functional traits. New Phytol. 220, 10–24 (2018). [DOI] [PubMed] [Google Scholar]
- 31.Demenou B. B., Piñeiro R., Hardy O. J., Origin and history of the Dahomey Gap separating West and Central African rain forests: Insights from the phylogeography of the legume tree Distemonanthus benthamianus. J. Biogeogr. 43, 1020–1031 (2016). [Google Scholar]
- 32.Couvreur T. L. P., Odd man out: why are there fewer plant species in African rain forests? Plant Syst. Evol. 301, 1299–1313 (2014). [Google Scholar]