

Abstract
A fundamental endeavor of the International Histocompatibility and Immunogenetics Workshop (IHIW) was assembling a collection of DNA samples homozygous through the MHC genomic region. This collection proved invaluable for assay development in the histocompatibility and immunogenetics field, for generating the human reference genome, and furthered our understanding of MHC diversity. Defined by their HLA-A, -B, -C and -DRB1 alleles, the combined frequency of the haplotypes from these individuals is ~20% in Europe. Thus, a significant proportion of MHC haplotypes, both common and rare throughout the world, and including many associated with disease, are not yet represented. In this workshop component, we are collecting the next generation of MHC-homozygous samples, to expand, diversify and modernize this critical community resource that has been foundational to the field. We asked laboratories worldwide to identify samples homozygous through all HLA class I and/or HLA class II genes, or through whole-genome SNP genotyping or sequencing, to have extensive homozygosity tracts within the MHC region. The focus is non-Europeans or those having HLA haplotypes less common in Europeans. Through this effort, we have obtained samples from 537 individuals representing 294 distinct haplotypes, as determined by their HLA class I and II alleles, and an additional 50 haplotypes distinct in HLA class I or II alleles. Although we have expanded the diversity, many populations remain underrepresented, particularly from Africa, and we encourage further participation. The data will serve as a resource for investigators seeking to characterize variation across the MHC genomic region for disease and population studies.
Keywords: MHC, reference haplotype, IHIW, sequencing, assembly, long-read, short-read, HLA, homozygous, ethnicity, ancestry
Introduction
The Major Histocompatibility Complex (MHC) was discovered through decisive roles in tissue transplantation experiments1. The functional polymorphism of the MHC thus identified likely arose through driving immunity to a diversity of pathogens2–4. The MHC has also become associated with susceptibility to a multitude of diseases, particularly those mediated by the immune system, and is established as the most polymorphic region of the human genome5–8. In addition to the extremely polymorphic HLA class I and II loci, the 5Mbp genomic region encodes approximately 175 genes of wide-ranging function and varying degrees of polymorphism9–11. Because of the importance for human health, the MHC was one of the first regions of the human genome to be completely characterized12. Although HLA alleles and haplotypes have diversified tremendously in composition and frequency worldwide13–15, the current human genome reference encompasses little of this diversity, restricted almost exclusively to alleles of European origin (Figure 1).
Figure 1. Geographic representation of the MHC region within the human genome reference sequences (hg38/GRCh38).

From left to right: hg38/GRCh38 haplotype ID, IHWG Cell ID, HLA-A~C~B~DRB1 haplotype, frequency of haplotype in major population groups, population having highest frequency. † - standard reference of hg38 (hap1-7 are the alt_refs28), ‡ - additional haplotype from Houwaart et al.29. The haplotype frequencies were obtained from allelefrequencies.net38; shown for each major ancestry group is the highest frequency within that group. Population having highest frequency at the right is obtained from studies having >1,000 subjects. The completed haplotypes are available with GenBank ID numbers OK649231-6.
Analysis of MHC genetic variation for clinical and basic science is hampered by extreme sequence and structural diversity. This diversity is interspersed with long stretches of low sequence complexity, and unusually strong linkage disequilibrium with defined recombination hotspots16–19. These factors, and the importance for disease research, drive the necessity of developing reference sequences that fully and accurately capture the diversity of the human MHC20,21. Acknowledging the obstacles presented for this goal, the International Histocompatibility Working Group (IHWG), coordinated a worldwide effort spanning 1987 to 2006, collecting approximately 100 samples shown to be homozygous for the HLA class I and II genes22–24. High-density SNP analysis showed 65% of these samples are homozygous throughout the 5Mbp MHC genomic region25. The samples, via their immortalized cell lines, have become an invaluable resource for immunogenetic research, encompassing assay development and optimization, clinical, basic and translational science studies. The original samples included 71 distinct HLA-A~C~B~DRB1 haplotypes and cover the major HLA class II structural variants10. The combined frequency of these haplotypes, however, is only an estimated 20% in Europeans, and considerably fewer in other populations25.
The original MHC reference sequence was generated from multiple individuals12. Recognizing that a chimeric reference was inadequate for identification of causal loci, the MHC haplotype project targeted eight of the MHC homozygous individuals identified by the IHWG9. The eight haplotypes were chosen through their implications in susceptibility to specific autoimmune diseases, and their relatively high frequency in European populations26. Two of these haplotypes (PGF and COX; Figure 1), were completed and remain part of the human reference sequence27,28, and five more of them were completed recently to reference quality29. To improve worldwide representation, this IHIW component was established to broaden the depth and breadth of genetic ancestry of the MHC reference haplotypes. The aim is to collect and sequence further samples identified as homozygous in the MHC genomic region, and representing HLA haplotypes that are frequent in non-European ancestries and/or infrequent in Europeans. Through high throughput targeted sequencing and specifically developed bioinformatics pipelines, the project will create fully representative MHC haplotypes for human immunogenetic studies. The collaborative effort of the working group will increase the number of MHC reference sequences 100-fold, with ongoing efforts to identify further haplotypes from underrepresented groups. Upon completion of new haplotype assemblies, the reference sequences will be submitted to GenBank and published for public use29. This expansion of the MHC haplotype database will bolster development of high throughput and accurate methods of accessing polymorphisms of the MHC in association with human disease30.
Methods and Progress
The 18th International HLA & Immunogenetics workshop took place on 11th to 15th May 2022, in Noordwijkerhout, the Netherlands. The component ‘Creating fully representative MHC reference haplotypes’ was organized by the laboratory group of Paul Norman from the University of Colorado USA, with additional input from the groups of Alex Dilthey (Dusseldorf) and Peter Parham (Stanford). We define the ~5Mbp MHC region as spanning human chromosome 6:28510120–33532223 (GRCh38/hg38), encompassing the GPX5 and ZBTB9 genes, and flanked by SNP identifiers rs2108925 and rs449242.
MHC Region Targeted Sequencing
Genomic library preparation for incoming samples was performed as described10,31,32. In brief, 500ng of genomic DNA for each sample was enzymatically fragmented to an average length of 900bp and a universal adaptor added to the end of each fragment, followed by end repair and bead-based purification. To select the desired size of fragmented DNA, we performed bead-based dual size selection, achieving a near uniform size distribution for each sample. To allow identification of each individual sample, unique Illumina barcodes were added to the universal adaptor using PCR. Following post-PCR clean-up, the samples were pooled into groups of 24. The sample pools were enriched for the MHC genomic region using a custom-designed biotinylated DNA probe panel10. The enrichment probes target the MHC by hybridization during a heated incubation, followed by a limited number of PCR cycles10. DNA fragments representing the MHC region were then separated from the remaining genomic DNA using streptavidin beads. The enrichment method was based on the Kapa HyperCap Target Enrichment protocol and kit v2. The resulting MHC enriched DNA was sequenced using an Illumina NovaSeq 6000 instrument, set up with an SP flow cell to produce 2×250 bp reads.
HLA Genotype Calling
HLA alleles were determined from the sequence data using the consensus calls obtained from three algorithms: NGSengine® 2.10.0 (GenDX, Utrecht, the Netherlands), HLA Explore™ 2.0.0 (Omixon Biocomputing Ltd. Budapest, Hungary) and HLA*LA33. We used IMGT database version 3.44.1_934. We determined a consensus call from the three algorithms, as previously described31. Here, 95% of cases agreed across the three methods at two fields of resolution, and any discrepancies were manually inspected for read alignment errors, which we observe to be more frequent for homozygous than heterozygous samples.
Samples and contributors
We began with the 95 HLA homozygous IHWG cell lines previously sequenced and scaffolded10, eight of which are now sequenced to reference standards29. We included an additional 12 samples, purchased from IHWG. The original 95 IHWG samples have stretches of homozygosity through the MHC with a mean of 4.3Mbp25. Because many of the original IHWG samples had been obtained from consanguineous individuals, these homozygous tracts contain long stretches of sequence identity by descent (IBD). We continued by identifying samples through high-resolution HLA genotyping or through whole-genome analysis, as homozygous for HLA class I and class II. For rare haplotypes, we also considered individuals homozygous for HLA class I or class II. The DNA samples had been genotyped locally using standard or high resolution NGS methods, with the remainder imputed from whole-genome SNP data. Due to the rarity of the individuals we sought, a significant resource was the DKMS donor registry having HLA genotype data from millions of individuals35–37. We anticipate the mean homozygous tract length to be shorter for many of the newly-collected individuals, who were randomly selected from the given populations and their homozygosity based on Hardy-Weinberg proportions rather than known IBD. Moreover, we previously identified one from 100 samples that genotyped as homozygous for classical HLA genes to be heterozygous in flanking genes25. For these reasons, we have sought duplicate examples of specific HLA haplotypes where possible.
As a result of this workshop component and other efforts, to date we have collected samples from 15 laboratories, with a total of 537 HLA class I and/or class II homozygous DNA samples (Table 1, Figure 2A). The DNA samples were sequenced and genotyped for HLA class I and II alleles using the targeted approach as described above. Represented in the 537 samples are 294 distinct full MHC region haplotypes, as determined by their HLA class I and II alleles (Supplementary Figure 1A). Of the 294 haplotypes, 262 are represented by one sample each. Also represented by samples homozygous through HLA class I or class II only are an additional 24 haplotypes distinguished by their HLA class I alleles, and 26 by HLA class II alleles (Supplementary Figure 1B–C). Taken together, there are 215 distinct HLA class I haplotypes and 199 distinct HLA class II haplotypes represented in homozygous state in this cohort (Supplementary Figure 1B–C). These estimates of uniqueness are based on the classical HLA genotypes and the number of distinct haplotypes present may increase upon analysis of the remaining genes and flanking regions.
Table 1.
HLA homozygous DNA samples and their origins.
| Laboratory | ||||
|---|---|---|---|---|
| PI | Institute | Country | Origin | Samples |
| South Asia | 1 | |||
| Oceania | 4 | |||
| South America | 1 | |||
| Nicholas Nii-Trebi | University of Ghana | Ghana | West Africa | 12 |
| Gehad ElGhazali | UAE University | UAE | Middle East | 10 |
| Charles Khor | NCGHM Hospital Tokyo | Japan | East Asia | 5 |
| Edinur Hisham Atan | Universiti Sains Malaysia | Malaysia | Southeast Asia | 4 |
| Theofanis Chatzistamatiou | Academy of Athens | Greece | Europe | 18 |
| Christina Voorter | Maastricht University | Netherlands | Europe | 4 |
| Esteban Arrieta-Bolaños | University Hospital Essen | Germany | Central America | 5 |
| Europe (USA) | 4 | |||
| Jorge Oksenberg | UCSF | USA | Europe (USA) | 33 |
| Maria Luiza Petzl-Erler | Federal University of Paraná | Brazil | South America | 14 |
| Ketevan Gendzekhadze | City of Hope, Duarte | USA | South America | 5 |
| Rafael Cita | Pio XII Foundation | Brazil | South America | 13 |
| (total) | 537 |
Figure 2: Geographic origins and representation of HLA homozygous DNA samples analyzed.
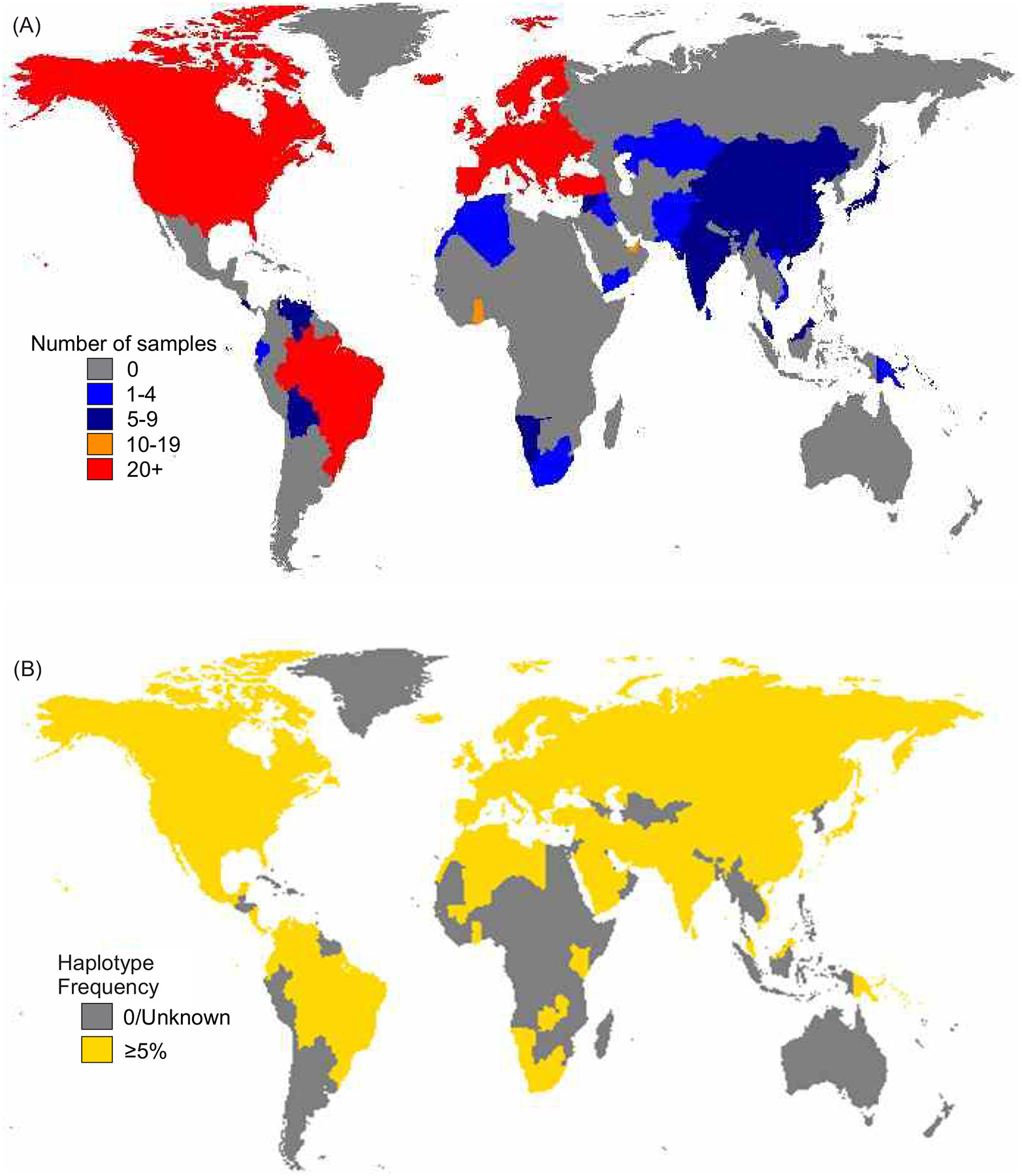
A. Shows the country of origin for the samples included in the study. Colours indicate the number of samples from a given country (grey 0, light blue 1–4, dark blue 5–9, orange10–19, red >20).
B. Shows an estimation of the global representation of the HLA-A~C~B~DRB1 haplotypes included in the study. Countries are coloured yellow if 5% or more of individuals are estimated to carry at least one of the haplotypes. The haplotype frequencies were obtained from allelefrequencies.net38.
To estimate the increase in coverage obtained for European populations, we compared the frequencies of the haplotypes detailed here, with those of the extensive DKMS data set of HLA haplotypes derived from Europeans35. In this database, the combined frequency of the 4-locus haplotypes (HLA-A~C~B~DRB1) that are equivalent to those originally characterized from the IHIW cell lines is 20.1% (Supplementary Figure 1D). This figure is similar to the previous estimate of 20%25, which was based on allefrequencies.net data38. With addition of the haplotypes detailed here, the frequency increases to 35.5%, and includes all 20 of the most frequent HLA-A~C~B~DRB1 haplotypes observed in Europe (Supplementary Figure 1D). Moreover, the combined allele frequency in Europe of the distinct 6-locus haplotypes (HLA-A~C~B~DRB1~DQB1~DPB1) originally characterized from the IHIW cell lines10 is 6.3%, and addition of the haplotypes described here increases the representation to 23.8% (Supplementary Figure 1E). Relative to Europeans however, information on the worldwide distribution of HLA haplotypes is limited. The worldwide distributions of the 4-locus HLA haplotypes present here were therefore determined using the allelefrequencies.net webtool38. Although HLA frequency data is absent for some countries, we estimate many human populations will be represented to a minimum of 5% total haplotype frequency (Figure 2B). Remaining is a lack of representation from certain geographic regions, including Africa, South Asia and Oceania, and we continue to welcome samples originating from these areas ahead of the 19th IHIW to be held in Japan in 2026 (https://ihiw19.org/). Please contact the corresponding author (paul.norman@ucdenver.edu) for more information or to participate in these ongoing efforts.
Conclusion
The efforts of the members and contributors of this 18th International HLA & Immunogenetics Workshop component have amassed an unprecedented collection of HLA homozygous samples from around the globe. The culmination of these efforts will result in the assembly of over 500 representative MHC haplotypes. In tandem with additional ongoing initiatives, such as the pangenome haplotype project39,40, representation of worldwide human diversity by reference haplotypes for the MHC genomic region will be substantially improved. This database will be an invaluable tool for studying the MHC, enabling the identification of genetic variants that directly impact human health and disease. We continue to accept samples identified as homozygous for HLA class I, class II or for both. Although we have begun to shift the balance away from European ancestry haplotypes, other populations remain underrepresented, and are the focus of our ongoing collection efforts.
Supplementary Material
Acknowledgments
We extend thanks to the 18th IHIW meeting organizers, Sebastiaan Heidt, Eric Spierings, and the HLA Working Group the Netherlands (HLA-WN). We are grateful to Wietse Mulder of GenDX and Atilla Bérces of Omixon for the use of their respective software packages. We are also grateful for support from the NIH backed HLA Region Genomics Consortium (HLA-RGC), chaired by Effie Petersdorf. HAE acknowledges support given by Ministry of Higher Education Malaysia (Fundamental Research Grant Scheme: FRGS/1/2020/STG03/USM/03/5).
Funding
This work was supported by NIH/NIAID U01 AI090905
Footnotes
Conflicts of Interest
The authors declare no conflicts of interest.
References
- 1.Bodmer W A historical perspective on HLA. Immunotherapy Advances. 2023;3(1). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hedrick PW, Thomson G. Evidence for balancing selection at HLA. Genetics. 1983;104(3):449–456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Parham P, Ohta T. Population biology of antigen presentation by MHC class I molecules. Science. 1996;272(5258):67–74. [DOI] [PubMed] [Google Scholar]
- 4.Lenz TL, Spirin V, Jordan DM, Sunyaev SR. Excess of Deleterious Mutations around HLA Genes Reveals Evolutionary Cost of Balancing Selection. Mol Biol Evol. 2016;33(10):2555–2564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Trowsdale J, Knight JC. Major histocompatibility complex genomics and human disease. Annu Rev Genomics Hum Genet. 2013;14:301–323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dendrou CA, Petersen J, Rossjohn J, Fugger L. HLA variation and disease. Nat Rev Immunol. 2018;18(5):325–339. [DOI] [PubMed] [Google Scholar]
- 7.Terao C, Raychaudhuri S, Gregersen PK. Recent Advances in Defining the Genetic Basis of Rheumatoid Arthritis. Annu Rev Genomics Hum Genet. 2016;17:273–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Matzaraki V, Kumar V, Wijmenga C, Zhernakova A. The MHC locus and genetic susceptibility to autoimmune and infectious diseases. Genome Biology. 2017;18(1):76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Horton R, Wilming L, Rand V, et al. Gene map of the extended human MHC. Nat Rev Genet. 2004;5(12):889–899. [DOI] [PubMed] [Google Scholar]
- 10.Norman PJ, Norberg SJ, Guethlein LA, et al. Sequences of 95 human MHC haplotypes reveal extreme coding variation in genes other than highly polymorphic HLA class I and II. Genome Research. 2017;27(5):813–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jensen JM, Villesen P, Friborg RM, Mailund T, Besenbacher S, Schierup MH. Assembly and analysis of 100 full MHC haplotypes from the Danish population. Genome Res. 2017;27(9):1597–1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.The-MHC-sequencing-consortium. Complete sequence and gene map of a human major histocompatibility complex. The MHC sequencing consortium. Nature. 1999;401(6756):921–923. [DOI] [PubMed] [Google Scholar]
- 13.Single RM, Meyer D, Mack SJ, et al. Haplotype frequencies and linkage disequilibrium among classical HLA genes. In: JA H, ed. Immunobiology of the Human MHC: Proceedings of the 13th International Histocompatibilty Workshop and Conference. Vol 1. Seattle: IHWG Press; 2007:705–746. [Google Scholar]
- 14.Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojobori T. HLA 1991, Proceedings of the Eleventh International Histocompatibility Workshop and Conference. 1992.
- 15.Prugnolle F, Manica A, Charpentier M, Guégan JF, Guernier V, Balloux F. Pathogen-driven selection and worldwide HLA class I diversity. Curr Biol. 2005;15(11):1022–1027. [DOI] [PubMed] [Google Scholar]
- 16.Kulski JK, Suzuki S, Shiina T. Haplotype Shuffling and Dimorphic Transposable Elements in the Human Extended Major Histocompatibility Complex Class II Region. Front Genet. 2021;12:665899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cullen M, Perfetto SP, Klitz W, Nelson G, Carrington M. High-resolution patterns of meiotic recombination across the human major histocompatibility complex. American journal of human genetics. 2002;71(4):759–776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jeffreys AJ, Kauppi L, Neumann R. Intensely punctate meiotic recombination in the class II region of the major histocompatibility complex. Nat Genet. 2001;29(2):217–222. [DOI] [PubMed] [Google Scholar]
- 19.Huttley GA, Smith MW, Carrington M, O’Brien SJ. A scan for linkage disequilibrium across the human genome. Genetics. 1999;152(4):1711–1722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Petersdorf EW, O’HUigin C. The MHC in the era of next-generation sequencing: Implications for bridging structure with function. Hum Immunol. 2019;80(1):67–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.D’Antonio M, Reyna J, Jakubosky D, et al. Systematic genetic analysis of the MHC region reveals mechanistic underpinnings of HLA type associations with disease. eLife. 2019;8:e48476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yang SY, Milford E, Hammerling U, Dupont B. Description of the Reference Panel of B-Lymphoblastoid Cell Lines for Factors of the HLA system: The B-Cell Line Panel Designed for the Tenth International Histocompatibility Wprkshop. In: Dupont B, ed. Immunobiology of HLA. Histocompatibility Testing. Vol 1. New York: Springer-Verlag; 1987:11–19. [Google Scholar]
- 23.Marsh SGE, Packer R, Heyes JM, et al. The International Histocompatibility Workshop cell panel. In: Charron D, ed. Genetic diversity of HLA. Functional and medical implications. Vol 1. Paris: EDK; 1996:26–28. [Google Scholar]
- 24.Mickelson E, Hurley C, Ng J, et al. 13th IHWS Shared Resources Joint Report. IHWG Cell and Gene Bank and reference cell panels. In: JA H, ed. Immunobiology of the Human MHC: Proceedings of the 13th International Histocompatibilty Workshop and Conference. Vol 1. Seattle: IHWG Press; 2006:523–553. [Google Scholar]
- 25.Norman PJ, Norberg SJ, Nemat-Gorgani N, et al. Very long haplotype tracts characterized at high resolution from HLA homozygous cell lines. Immunogenetics. 2015;67(9):479–485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gonzalez-Galarza FF, McCabe A, Melo Dos Santos EJ, Jones AR, Middleton D. A snapshot of human leukocyte antigen (HLA) diversity using data from the Allele Frequency Net Database. Hum Immunol. 2021;82(7):496–504. [DOI] [PubMed] [Google Scholar]
- 27.Horton R, Gibson R, Coggill P, et al. Variation analysis and gene annotation of eight MHC haplotypes: the MHC Haplotype Project. Immunogenetics. 2008;60(1):1–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nassar LR, Barber GP, Benet-Pagès A, et al. The UCSC Genome Browser database: 2023 update. Nucleic Acids Research. 2022;51(D1):D1188–D1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Houwaart T, Scholz S, Pollock NR, et al. Complete sequences of six major histocompatibility complex haplotypes, including all the major MHC class II structures. HLA. 2023;102(1):28–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dilthey AT. State-of-the-art genome inference in the human MHC. Int J Biochem Cell Biol. 2021;131:105882. [DOI] [PubMed] [Google Scholar]
- 31.Farias TDJ, Brugiapaglia S, Croci S, et al. HLA-DPB1*13:01 associates with enhanced, and KIR2DS4*001 with diminished protection from developing severe COVID-19. HLA. 2024;103(1):e15251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Norman PJ, Hollenbach JA, Nemat-Gorgani N, et al. Defining KIR and HLA Class I Genotypes at Highest Resolution via High-Throughput Sequencing. American journal of human genetics. 2016;99(2):375–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Dilthey AT, Mentzer AJ, Carapito R, et al. HLA*LA-HLA typing from linearly projected graph alignments. Bioinformatics. 2019;35(21):4394–4396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Barker DJ, Maccari G, Georgiou X, et al. The IPD-IMGT/HLA Database. Nucleic Acids Res. 2023;51(D1):D1053–d1060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Seitz S, Lange V, Norman PJ, Sauter J, Schmidt AH. Estimating HLA haplotype frequencies from homozygous individuals - A Technical Report. Int J Immunogenet. 2021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Lange V, Böhme I, Hofmann J, et al. Cost-efficient high-throughput HLA typing by MiSeq amplicon sequencing. BMC Genomics. 2014;15(1):63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schöfl G, Lang K, Quenzel P, et al. 2.7 million samples genotyped for HLA by next generation sequencing: lessons learned. BMC Genomics. 2017;18(1):161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gonzalez-Galarza FF, McCabe A, Santos E, et al. Allele frequency net database (AFND) 2020 update: gold-standard data classification, open access genotype data and new query tools. Nucleic Acids Res. 2020;48(D1):D783–D788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liao WW, Asri M, Ebler J, et al. A draft human pangenome reference. Nature. 2023;617(7960):312–324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ebert P, Audano PA, Zhu Q, et al. Haplotype-resolved diverse human genomes and integrated analysis of structural variation. Science. 2021;372(6537). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.