

Abstract
Type I diabetes is an autoimmune disease mediated by T-cell destruction of β cells in pancreatic islets. Currently, there is no known cure, and treatment consists of daily insulin injections. Genome-wide association studies and twin studies have indicated a strong genetic heritability for type I diabetes and implicated several genes. As most strongly associated variants are noncoding, there is still a lack of identification of functional and, therefore, likely causal variants. Given that many of these genetic variants reside in enhancer elements, we have tested 121 CD4+ T-cell enhancer variants associated with T1D. We found four to be functional through massively parallel reporter assays. Three of the enhancer variants weaken activity, while the fourth strengthens activity. We link these to their cognate genes using 3D genome architecture or eQTL data and validate them using CRISPR editing. Validated target genes include CLEC16A and SOCS1. While these genes have been previously implicated in type 1 diabetes and other autoimmune diseases, we show that enhancers controlling their expression harbor functional variants. These variants, therefore, may act as causal type 1 diabetic variants.
Keywords: type 1 diabetes, non-coding variants, enhancer elements, GWAS, 3D genome architecture, massively parallel reporter assay (MPRA)
Introduction
Type I diabetes mellitus (T1D) is categorized as an autoimmune disease caused by T-cell-mediated loss of β cells in pancreatic islets of Langerhans that leads to insulin deficiency. Autoreactive T cells are known to be key mediators of β-cell destruction (1). T1D is most commonly present in childhood or adolescence; however, it can appear at any age. The genetic factor comprises ~80% of an individual’s risk for developing the disease. In the past decade, a series of genome-wide association studies (GWAS) have identified more than 50 genetic loci for conferring T1D risk (2–4). As most of the underlying single nucleotide polymorphisms (SNPs) associated with T1D, as well as those in linkage disequilibrium (LD), are in noncoding sequences, interpreting their role in T1D is challenging.
The development of massively parallel reporter assays (MPRAs) has tremendously increased our capacity to test regulatory sequences carrying genetic variation for any potential effect (5, 6). To determine differences in allelic enhancer activity, MPRA can be utilized to test thousands of regulatory alleles in a single reporter library (7). Previously, it had been applied to characterize cancer-associated variants in cancerous cell lines, blood cell disease traits, and heart disease-associated variants (8–10).
In the current study, we employ MPRAs to quantify the relative effect of T1D variants located in regulatory sequences identified in activated CD4+ T cells. We tested 121 enhancers and 242 alleles, of which four variants showed significant differential enhancer activity. Three of these variants are in the CLEC16A gene locus, and one lies in the BCL2L15 gene locus. Target genes for these functionally validated T1D enhancers are assigned by promoter capture Hi-C and expression quantitative trait loci (eQTL), a subset of which we validate using CRISPR inhibition (CRISPRi) enhancer repression.
Methods
Prioritization and functional analysis of variants
We downloaded the NHGRI GWAS Catalog (https://www.ebi.ac.uk/gwas/docs/file-downloads), and all coordinates are hg19. We identified 11 T1D trait SNPs associated (p < 1.0 × 10−5) with the disease, respectively. Next, we identified proxy SNPs in linkage disequilibrium (LD) with a GWAS SNP based on the 1,000 Genomes Project of CEU ancestry by utilizing the SNAP web server (http://archive.broadinstitute.org/mpg/snap/ldsearch.php). We overlapped the set of T1D-associated SNPs with our previously described H3K4me1 ChIP-seq data, a putative enhancer mark, from primary human CD4+ T cells (11) and identified 121 noncoding variants in enhancer regions.
Design and synthesis of MPRA library
For the multiplex enhancer SNP validation assay, we leveraged synthetic oligonucleotide array synthesis and adopted self-transcribing active regulatory region sequencing (STARR-seq) (12) (https://www.addgene.org/71509/). An oligonucleotide library was synthesized containing the 230 nucleotide genomic regions as previously described (13).
Activated CD4+ T-cell culture
MPRA experiments were performed in activated CD4+ T cells. Human CD4+ T cells were obtained from Precision for Medicine, grown at 37°C and 5% CO2 in TexMACSTM (Miltenyi-Biotech, Gaithersburg, MD, USA) medium with 10% human serum (Sigma, Saint Louis, MO, USA) and 1% penicillin–streptomycin (Gibco, Grand Island, NY, USA). Naive CD4+ T cells were activated with bead-bound anti-CD3-biotin and anti-CD28-biotin, and expansion of activated CD4+ cells was carried out by adding IL-2 at 10 ng/mL (Miltenyi Systems) to the cultures at 48 h. Cells were electroporated with a Neon transfection kit and device (Invitrogen, Bend, OR, USA) with the following transfection parameters—pulse voltage: 2,100, pulse width: 20, and pulse number: 1.
MPRA output library construction
MPRA library preparation was performed as previously described (13). Briefly, after cell collection, total RNA from rinsed cell pellets was prepared using the QIAGEN RNeasy kit. Poly-A RNA was isolated from 50 μg of total RNA by µMACS mRNA isolation kit (Miltenyi Biotech). RNA was then treated with turboDNase (4 U) for 30 min at 37°C (Invitrogen). DNase-treated poly-A RNA was purified using the RNeasy kit. Plasmid-specific cDNA was synthesized using Superscript III (Life Technologies, Austin, TX, USA) incubated for 1.5 h at 55°C and inactivated at 80°C for 15 min. Following synthesis, cDNA was treated with RNaseA (Sigma) at 37°C for 30 min. cDNA was purified using AMPure beads in a 1.5:1 bead:cDNA ratio and then amplified and indexed for sequencing using a two-stage PCR as described previously ((12). The cDNA sample from each replicate was used as an input into first-round gene-specific PCR reaction, and KAPA hi-fidelity polymerase (KAPA Biosystem, Wilmington, MA, USA). All libraries were sequenced on the NextSeq 550 (Illumina) performing 1 × 75 cycles.
MPRA data normalization and analysis
Sequencing raw reads from RNA and plasmid libraries are checked for adapter sequences and low-quality reads (q score < 20) using FASTQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and trimmed using Trim Galore (https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/). Trimmed reads were mapped to the amplicon library using the Bowtie2 aligner (14). The mapped reads were quantified against all tested sequences using the “featureCounts” function from the Rsubread package (15). We employed quantitative allele-specific analysis of reads (QuASAR)-MPRA (16) to define allele-specific activity from MPRA read counts. Allelic enhancer activity p-values are adjusted with the Benjamini–Hochberg method to control for multiple testing. Significant allelic enhancer activity was defined at an FDR < 10%.
T1D enhancer SNP transcription factor binding site analyses
We utilized motifbreakR in R to determine the effect of significant enhancer variants from MPRA on TF binding (19). motifbreakR uses the position weight matrices (PWM) from multiple databases such as Hocomocos, Jaspar, SwissRegulon, CisBP, and hPDI. motifbreakR calculates a p-value for alternate and reference allele versions of the TF PWM and then determines binding difference and significance. For the final results, we only use significant binding changes with a strong effect predicted and filtered TFs for expression in activated CD4+ T cells using the DICE database (17) (Supplementary Table S2).
CRISPR inhibition
Guide RNAs (gRNAs) targeting enhancers were designed with Benchling (https://benchling.com) using a 200-bp window around the enhancer variant (Supplementary Table S3). The two best-scoring gRNAs were synthesized from Synthego, Redwood City, CA, USA. Reconstituted gRNAs were pooled for each enhancer and electroporated into Jurkat-CRISPRi cells (see below). Nontargeting control (NTC) guides as negative controls were obtained from ThermoFisher, Bend, OR, USA (TrueGuide™ sgRNA Negative Control, nontargeting 1, Catalog No. A35526).
Jurkat cells were transduced with the CRISPRi Lentivirus hEF1a-Blast-dCas9-SALL1-SDS3 from Horizon (Catalog ID: VCAS12247). Cells were cultured in RPMI 1640 with 10% FBS and 15 µg/mL of blasticidin for 2 weeks. Single-cell clones were isolated postflow sorting. Single clones were expanded in the presence of antibiotics and confirmed for Cas9 expression using qPCR. These single-cell clones were cryopreserved. Cryopreserved Jurkat CRISPRi cells were initially cultured in RPMI 1640 supplemented with 10% FBS, 1× normocin (InvivoGen). After 3 days of culture, 15 µg/mL blasticidin was added to the culture medium. Cells were maintained and fed every other day. Cells were collected at a density of 2 × 106 per CRISPRi target or NTC, washed with PBS, and resuspended in 45 µL of buffer T (Neon Transfection Kit). Two 15-µM gRNAs per target enhancer were combined at a 1:1 ratio to a final volume of 6 µL. In total, 5 µL of combined gRNA was then added to a tube of cells and mixed well for each CRISPRi-targeted enhancer. The NTC gRNA was diluted to 15 µM, according to the ThermoFisher protocol. Next, cells were mixed with 5 µL of NTC gRNA in the same manner as the enhancer targets. Using the Neon Electroporation system, 10 µL of cell/gRNA suspension was electroporated at 1,350 V, 10 ms, for three pulses. Cells were placed in 190 µL of rescue medium in a flat bottom 96-well plate and allowed to culture for 3 days postelectroporation. Cells were then removed from the plate, washed with PBS, and frozen until RNA isolation was performed.
RNA was isolated using the Qiagen RNeasy Mini kit with DNase treatment. RNA was eluted in 30 µL of RNase-free water. Concentration was tested using the Qubit RNA High Sensitivity assay. After concentration was tested, cDNA was synthesized using the iScript cDNA kit, using 75 ng as a standard following the manufacturer’s protocol. Post-cDNA synthesis, qPCR was performed using TaqMan probes (ThermoFisher) for pcHi-C/eQTL target genes and TBP as the internal control gene (Supplementary Table S3). Analysis of qPCR data from targeted enhancers and NTC was performed using the TaqMan delta-delta Ct method in Excel and plotted in GraphPad Prism.
Statistical tests and visualizations
Any additional statistical tests were performed in the R environment (https://www.r-project.org/). Additional graphs and figures were prepared using R packages, GraphPad Prism, BioRender.com, and Abode Illustrator.
Result
Differential regulatory activity of T1D enhancer variants
For non-coding variants associated with diseases, such as T1D, to be causal, the variants must have a functional effect. To determine if noncoding, T1D-associated variants have a gene regulatory effect, we first identified SNPs in linkage disequilibrium using 1,000 Genomes data, and then determined if they reside at candidate enhancer elements in activated CD4+ T cells using previously generated chromatin maps of H3K4me1 (11) (Figure 1A), which is known to mark enhancers (18). We identified 121 T1D regulatory variants (Supplementary Table S1). To determine an allelic difference in enhancer activity due to the T1D variant, the 121 variants (242 alleles) were tested by MPRA in activated CD4+ T cells (Figure 1A). After adjusting for multiple corrections with Benjamini–Hochberg (BH) at a 10% False discovery rate (FDR), we were able to identify four variants having differential allelic enhancer activity in activated CD4+ T cells (Figures 1B, C). Of these, three variants showed stronger activity for the reference allele, indicating that the alternate allele is weakening enhancer activity, while rs12599402 has stronger activity for the alternate allele (Figure 1). Three significant MPRA variants (rs12599402, rs7203150, and rs9746695) are from the CLEC16A locus at chromosome 16p13.13 and lie within intronic enhancers within the gene. The fourth variant, rs2358995, is an intronic enhancer variant in the BCL2L15 and AP4B1-AS1 genes, an antisense lncRNA on the opposite strand at chromosome 1p13.2. Collectively, these data show that T1D-associated noncoding variants can act in a functional manner to alter enhancer activity.
Figure 1.

Identification of functional noncoding T1D variants. (A) Identification of T1D enhancer variants from activated CD4+ T cells and MPRA design. (B) Scatter plot of reference versus alternate MPRA allelic activity for enhancer variants. (C) Bar plot of MPRA activity for each significant enhancer allele.
Transcription factor binding analysis of functional enhancer variants
The most likely mechanism for enhancer variants to have a functional effect is by altering transcription factor (TF) binding. We used motifbreakR to predict if each variant is likely to alter TF binding based on the alternate allele change to the corresponding motif (19). After filtering motifs for TF expression in activated CD4+ T cells, each variant is predicted to alter multiple motifs (Figures 2A, B; Supplementary Table S2). For example, rs9746695 alters motifs for ZEB1, SMAD2, and TCFs. rs12599402 is predicted to change the binding of several factors, including ESRRA, RFXANK, a regulator of HLA class II promoters, and LEF1. The rs2358995 variant impacts SMAD2, GATA3, and FOXJ motifs, whereas rs7203150 is associated with changes in HOXB3 and MYB motifs. Many of these TFs, such as TCF12, LEF1, SMAD2, and GATA3, are known to regulate SOCS1 and CLEC16A by also binding their promoters (20). The homotypic binding of TFs at a gene’s promoter and enhancers is a common feature of gene regulation (21, 22), indicating the above factors may be key regulators of SOCS1 and CLEC16A expression.
Figure 2.

T1D enhancer SNP-associated transcription factor network. (A) Top-scoring TF motifs altered by the T1D enhancer variants. (B) Example motifs for each variant shown. (C) Protein–protein interactions for TFs associated with each T1D enhancer variant. The hub colors correspond to the variant colors in (A, B).
To understand how such diverse TFs could play a role in T1D, we asked if there were known relationships between these factors. Protein–protein interactions indicate that many of these TFs do interact (Figure 2C). The implication is that individual variants in different enhancers may impact the same complex or similar complexes of TFs by altering the binding of interacting TFs. Furthermore, KEGG pathway analysis of the TFs associated with the functional variants indicates that these factors play a coordinated role in regulating “adherens junction”, “Th17 differentiation”, “human T-cell leukemia virus 1 infection”, and others (Supplementary Figure S1A).
T1D enhancer variant target gene identification
To gain insight into how enhancer variants contribute to T1D etiology or pathogenesis, we utilized promoter capture Hi-C data from activated CD4+ T cells to identify target genes for each enhancer variant (23), as well as existing eQTL data. rs9746695 and rs7203150 enhancer variants are known eQTL variants for CLEC16A expression (Open Targets database; 24). After filtering for expression in activated CD4+ T cells of promoter-capture Hi-C data-interacting genes, the rs12599402, rs9746695, and rs7203150 intronic enhancers of CLEC16A, C-type lectin domain containing 16A—a known T1D susceptibility gene (25), have long-range distal interactions over 250 kb with the SOCS1 and RMI2 promoters (Figure 3A; Supplementary Figure S1B). SOCS1, a suppressor of cytokine signaling 1, acts as a negative regulator of interleukin-2 (IL-2), interleukin-3 (IL-3), and interferon-gamma (IFN-γ) signaling. RMI2, RecQ-mediated genome instability 2, is essential for genome stability (26). rs9746695 is an eQTL variant for RMI2 expression in whole blood (Open Targets database). The RMI2 isoform, RMI2-004, was reported as a T1D risk gene. RMI has been shown to gain chromatin accessibility and higher transcriptional output in response to IFN-α expression in islets of T1D individuals (27). All three variants are also eQTL for DEXI (Open Targets database), the promoter of which is next to CLEC16A. DEXI, a dexamethasone-induced protein, may play a role in controlling inflammatory responses (28). The rs2358995 variant within the BCL2L15, B-cell lymphoma 2-like 15, gene is a previously reported eQTL for BCL2L15 expression in LCL (29) and transverse colon (30), and for PTPN22. The enhancer also shows an interaction with the PTPN22 gene body, supporting the idea that it may also regulate PTPN22, but the variant is too close to the PTPN22 promoter to distinguish a significant interaction with the promoter from a false positive proximity ligation (Figure 3B). PTPN22 encodes a tyrosine phosphatase that functions as a key regulator of immune homeostasis through regulations of TCR and BCR signaling.
Figure 3.

Promoter captures Hi-C enhancer target genes. (A) Browser shot of CLEC16A-SOCS1 locus showing enhancer variants (orange lines), H3K4me1 ChIP-seq data from activated CD4+ T cells (red), and activated CD4+ T-cell pcHi-C data with variant interactions in blue. (B) Browser shot of BCL2L15 locus with data as described in (A).
CRISPR validation of target gene interactions and enhancer activity
To confirm the target gene interactions as well as enhancer activity of the region, we established a dead Cas9 (dCas9) CRISPRi Jurkat T-cell line and targeted the CD4+ T-cell enhancers, which are also open chromatin regions in Jurkat cells (Figure 4A). We tested the effects of enhancer repression on target expression using qPCR as compared to nontargeting control (NTC) guides. We targeted the three CLEC16A intronic enhancer variant regions and tested the effects on CLEC16A and SOCS1 expression. We found significant downregulation of SOCS1 associated with editing in the rs12599402 (pval = 0.0009) and rs9746695 (pval = 0.0024) enhancers. While the rs7203150 enhancer repression was not significant (pval = 0.0699), SOCS1 expression trended downward (Figures 4B, C). A significant decrease in expression of CLEC16A was only associated with editing the rs9746695 enhancer (pval = 0.0002) (Figure 4D). All three variants significantly downregulate DEXI expression (Supplementary Figure S2). We next targeted the rs2358995 enhancer and tested the effect on BCL2L15 and PTPN22. BCL2L15 was significantly downregulated (pval = 0.0002), but no significant change was found for PTPN22 (pval = 0.6938) (Figure 4E).
Figure 4.
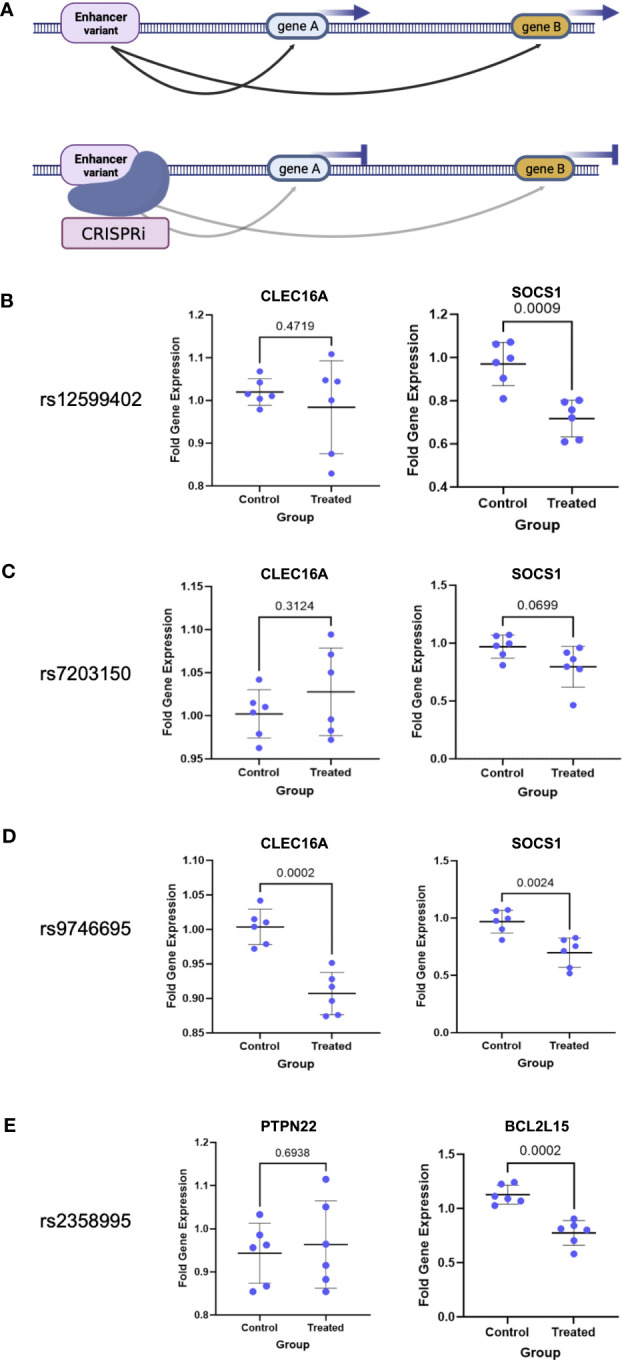
CRISPRi validation of enhancer–target gene interactions. (A) Schematic of enhancer interaction and CRISPRi effect on a true target gene. (B–E) Enhancer target gene expression level after enhancer (treated) repression by CRISPRi or using nontargeting guides (control). p-values are indicated above the line in each comparison.
Discussion
Genome-wide association studies have proven to be a compelling approach to identifying a genetic basis for T1D, as well as many common human diseases and traits; however, few genes have been found to be causal, although T1D-associated coding variants have been found for several genes (3, 31, 32). As most significant variants lie in a noncoding region of the genome, epigenomic technologies to identify candidate cis-regulatory elements, such as enhancers, have breathed new life into GWAS data. SNPs identified from GWAS can affect the function of transcriptional enhancers by altering transcription factor binding and, in turn, target gene expression levels through interactions with distant gene promoters via looping in 3D space. This warrants a critical examination of the genetic variation in cis-regulatory elements and a further need to determine their target genes if we are to understand the phenotypic consequences of functional regulatory variants. Such an understanding of variants associated with T1D may provide better insights into the etiology and potentiate novel therapeutic interventions.
In recent years, massively parallel reporter assays have become a powerful approach to dissecting the functional effects of noncoding variants associated with various diseases (7, 33–37). Our MPRA study of T1D-associated variants has pinpointed four enhancer variants from the CLEC16A and BCL2L15 loci as potentially causal. However, these enhancer variants do not necessarily regulate just the nearest neighboring gene and can regulate more than a single gene.
The rs9746695 (chr16:11207894, T/C) and rs7203150 (chr16:11207722, T/C) intronic enhancer variants of CLEC16A are known eQTL variants for CLEC16A expression, and promoter capture Hi-C indicates interactions with SOCS1 and RMI2. We tested the effect of this enhancer on the expression of CLEC16A and SOCS1 by CRISPRi and found that targeting the rs9746695 region of the enhancer reduced the expression of each gene, but targeting the rs7203150 region of the enhancer had no effect. It is unclear how variants within the same enhancer could have such differing results, other than blocking distinctly different TF-binding sites. One possibility is inefficient targeting by rs7203150 gRNAs. Other technical aspects such as transfection efficiency, could play a role, as not all cells take up the gRNA, therefore, all gene expression is measured in a background of unedited cells. rs9746695 is also a known eQTL variant controlling RMI2 expression. This indicates that the enhancer controls the expression of at least three genes: CLEC16A, SOCS1, and RMI2. rs12599402 (chr16:11189888, T/C) in an intronic enhancer ~18 kb away also interacts with SOCS1 and RMI2 but is not a known CLEC16A eQTL variant. Our CRISPRi validation confirmed enhancer activity and control of SOCS1 expression, but it does not appear to control CLEC16A. RMI2 was not tested. All three variants are also eQTL variants for DEXI, and our CRISPRi experiments confirm that the enhancers are controlling its expression. These functional variants, therefore, can impact the expression of multiple genes, and the expression of a single gene may be impacted by multiple T1D risk variants and distinct enhancer elements.
CLEC16A has been identified as a susceptibility gene for type 1 diabetes, multiple sclerosis, and adrenal dysfunction, with a clear role in T cells yet to be determined. CLEC16A is a membrane-associated endosomal protein and one of the important regulators of mitophagy. Its role in the pancreas is better studied; however, the function of mitophagy and related pathways in T1D biology is limited (38). A role in autophagy in T-cell survival and proliferation has been linked to the development of autoimmunity in related autoimmune disorders (39–42).
Similar to CLEC16A, DEXI has also been implicated as a T1D gene. However, recent studies in NOD mice found that DEXI knockout mice did not have an increased susceptibility to diabetes in the way that CLEC16A knockdown mice did (43).
The SOCS1 gene has been implicated in multiple autoimmune disorders, though variants in the gene have not been identified as causal. SOCS1 is a suppressor of cytokine signaling, and as such, regulates multiple cytokines involved in immune response, including IL-2 and IFN-γ. SOCS1 operates through the inhibition of the catalytic activity of JAK/STAT pathway members and TYK2. This in turn restricts the overall cellular communications that occur via cytokine signaling. Many of the cytokines regulated by SOCS1 are implicated in inflammation and inflammatory responses (44). Therefore, misregulation of SOCS1 via enhancer variants may promote a proinflammatory response. Such a proinflammatory response may prove a key driver of T1D pathogenic T cells. SOCS1 is capable of protecting against viral-mediated induction of T1D through the suppression of IFN-γ (45). Similarly, reduction of SOCS1 expression can lead to the expansion of pathogenic T cells in circulation (46).
Little is known about the function of RMI2 in T cells. However, a decrease in RMI2 expression in other systems was shown to lead to the downregulation of RUNX2 (47), which is an important repressor of Tfh-cell differentiation (42). Skewing in the population of circulating Tfh cells or their precursors is highly associated with T1D (48). It is feasible that the downregulation of RMI2 via the rs9746695 T1D enhancer risk allele leads to increased Tfh-cell formation.
The rs2358995 (chr1:114429515, A/C) BCL2L15 intronic enhancer variant is an eQTL for both BCL2L15 and PTPN22. We tested the effect of CRISPRi on expression by targeting the enhancers but did not see a significant change in PTPN22 expression. Even though the enhancer forms a long-range interaction with the PTPN22 gene body, there is no interaction with the gene promoter.
We validated an impact on BCL2L15 expression. The BCL2L15 gene is a proapoptotic member of the BCL2 family, which could impact T-cell proliferation and differentiation. It is not well studied, but a recent study showed that the knockdown of BCL2L15 leads to the downregulation of STAT1 and STAT3 (49), which, if translated to CD4+ T cells, would have important implications for their function and differentiation.
PTPN22 is strongly associated with type 1 diabetes, systemic lupus erythematosus, and rheumatoid arthritis. A coding variant within this gene was first linked to T1D and may even be causal (50, 51). It is noted as one of the most important non-HLA genetic risk factors in the predisposition to multiple autoimmune diseases. The lymphoid-specific protein is a molecular adapter protein associated with the negative regulation of TCR signaling, which may result in the survival of autoreactive T cells (51). We showed that the PTPN22 eQTL variant rs2358995 does indeed reduce enhancer activity. Reduced expression of PTPN22 through this enhancer variant may also predispose individuals to T1D.
Additionally, we found that the candidate TFs whose binding is predicted to be altered at T1D-associated enhancer variants have known interactions and play a role in shared pathways. For example, TCF1, LEF1, and ESRRA play important roles in CD4+ T-cell proliferation and differentiation (52, 53). The relationships between the candidate TFs would explain how different variants can alter binding for distinct TFs but result in a common phenotype, T1D. Altering TF binding to cis-regulatory elements provides a mechanism for how T1D-associated variants alter enhancer activity.
In summary, using MPRA in activated primary human CD4+ T cells, we identified four functional T1D variants residing in intronic enhancers of CLEC16A and BCL2L15 genes. We utilized 3D genomic and eQTL data to assign the putative target genes such as SOCS1 and CLEC16A and validated enhancer activity and target genes for variants containing distal enhancers using CRISPRi in Jurkat T cells. The validation identifies target genes that are well-known factors in driving the proinflammatory state of pathogenic T cells that drive ß-cell destruction. Dysregulated T-cell signaling due to functional regulatory variants also has the potential to skew T-cell differentiation, another key contribution to the etiology of T1D. We only validated a limited set of target genes, and future studies in primary T cells, including the use of base editors, could provide additional insight. We further predicted the effect of enhancer variants on TF binding, which indicated that shared pathways or regulatory networks may be at play in T1D. Collectively, these data prioritize these variants as candidate causal variants for T1D.
Data availability statement
The sequencing data generated by this study are available from the NCBI Short Read Archive (SRA) under accession PRJNA1120596.
Ethics statement
Ethical approval was not required for the studies involving humans because we used cells from a commercial biorepository that collects cells under their own IRB: Precision for Medicine Institutional Review Board (IRB) https://www.precisionformedicine.com/specialty-lab-services/biospecimens/. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from a commercial biorepository under IRB oversight. Written informed consent to participate in this study was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and the institutional requirements.
Author contributions
AM: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. AJ: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. EW: Writing – review & editing, Data curation, Formal analysis, Investigation. ND: Writing – review & editing, Data curation, Formal analysis. MG: Investigation, Writing – review & editing. DH: Writing – review & editing, Investigation. RDH: Conceptualization, Funding acquisition, Supervision, Writing – original draft, Writing – review & editing.
Funding Statement
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Institute of Diabetes and Digestive and Kidney Diseases (R01DK103667).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1387253/full#supplementary-material
References
- 1. Pugliese A. Autoreactive T cells in type 1 diabetes. J Clin Invest. (2017) 127:2881–91. doi: 10.1172/JCI94549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Noble JA, Erlich HA. Genetics of type 1 diabetes. Cold Spring Harb Perspect Med. (2012) 2:a007732. doi: 10.1101/cshperspect.a007732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Onengut-Gumuscu S, Chen WM, Burren O, Cooper NJ, Quinlan AR, Mychaleckyj JC, et al. Fine mapping of type 1 diabetes susceptibility loci and evidence for colocalization of causal variants with lymphoid gene enhancers. Nat Genet. (2015) 47:381–6. doi: 10.1038/ng.3245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Shapiro MR, Thirawatananond P, Peters L, Sharp RC, Ogundare S, Posgai AL, et al. De-coding genetic risk variants in type 1 diabetes. Immunol Cell Biol. (2021) 99:496–508. doi: 10.1111/imcb.12438 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Melnikov A, Murugan A, Zhang X, Tesileanu T, Wang L, Rogov P, et al. Systematic dissection and optimization of inducible enhancers in human cells using a massively parallel reporter assay. Nat Biotechnol. (2012) 30:271–7. doi: 10.1038/nbt.2137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Patwardhan RP, Lee C, Litvin O, Young DL, Pe’er D, Shendure J. High-resolution analysis of DNA regulatory elements by synthetic saturation mutagenesis. Nat Biotechnol. (2009) 27:1173–5. doi: 10.1038/nbt.1589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Tewhey R, Kotliar D, Park DS, Liu B, Winnicki S, Reilly SK, et al. Direct identification of hundreds of expression-modulating variants using a multiplexed reporter assay. Cell. (2016) 165:1519–29. doi: 10.1016/j.cell.2016.04.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Ulirsch JC, Nandakumar SK, Wang L, Giani FC, Zhang X, Rogov P, et al. Systematic functional dissection of common genetic variation affecting red blood cell traits. Cell. (2016) 165:1530–45. doi: 10.1016/j.cell.2016.04.048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Choi J, Zhang T, Vu A, Ablain J, Makowski MM, Colli LM, et al. Massively parallel reporter assays of melanoma risk variants identify MX2 as a gene promoting melanoma. Nat Commun. (2020) 11:2718. doi: 10.1038/s41467-020-16590-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Richter F, Morton SU, Kim SW, Kitaygorodsky A, Wasson LK, Chen KM, et al. Genomic analyses implicate noncoding de novo variants in congenital heart disease. Nat Genet. (2020) 52:769–77. doi: 10.1038/s41588-020-0652-z [DOI] [Google Scholar]
- 11. Hawkins RD, Larjo A, Tripathi SK, Wagner U, Luu Y, Lonnberg T, et al. Global Chromatin State Analysis Reveals Lineage-Specific Enhancers during the Initiation of Human T helper 1 and T helper 2 Cell Polarization. Immunity. (2013) 38:1271–84. doi: 10.1016/j.immuni.2013.05.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Arnold CD, Gerlach D, Stelzer C, Boryn LM, Rath M, Stark A. Genome-wide quantitative enhancer activity maps identified by STARR-seq. Science. (2013) 339:1074–7. doi: 10.1126/science.1232542 [DOI] [PubMed] [Google Scholar]
- 13. Doni Jayavelu N, Jajodia A, Mishra A, Hawkins RD. Candidate silencer elements for the human and mouse genomes. Nat Commun. (2020) 11:1061. doi: 10.1038/s41467-020-14853-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. (2012) 9:357–9. doi: 10.1038/nmeth.1923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Liao Y, Smyth GK, Shi W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. (2019) 47:e47. doi: 10.1093/nar/gkz114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Kalita CA, Moyerbrailean GA, Brown C, Wen X, Luca F, Pique-Regi R. QuASAR-MPRA: accurate allele-specific analysis for massively parallel reporter assays. Bioinformatics. (2018) 34:787–94. doi: 10.1093/bioinformatics/btx598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Schmiedel BJ, Singh D, Madrigal A, Valdovino-Gonzalez AG, White BM, Zapardiel-Gonzalo J, et al. Impact of genetic polymorphisms on human immune cell gene expression. Cell. (2018) 175:1701–1715 e1716. doi: 10.1016/j.cell.2018.10.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Heintzman ND, Hon GC, Hawkins RD, Kheradpour P, Stark A, Harp LF, et al. Histone modifications at human enhancers reflect global cell-type-specific gene expression. Nature. (2009) 459:108–12. doi: 10.1038/nature07829 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Coetzee SG, Coetzee GA, Hazelett DJ. motifbreakR: an R/Bioconductor package for predicting variant effects at transcription factor binding sites. Bioinformatics. (2015) 31:3847–9. doi: 10.1093/bioinformatics/btv470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Liska O, Bohar B, Hidas A, Korcsmaros T, Papp B, Fazekas D, et al. TFLink: an integrated gateway to access transcription factor-target gene interactions for multiple species. Database (Oxford). (2022) 2022. doi: 10.1093/database/baac083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Gotea V, Visel A, Westlund JM, Nobrega MA, Pennacchio LA, Ovcharenko I. Homotypic clusters of transcription factor binding sites are a key component of human promoters and enhancers. Genome Res. (2010) 20:565–77. doi: 10.1101/gr.104471.109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ezer D, Zabet NR, Adryan B. Homotypic clusters of transcription factor binding sites: A model system for understanding the physical mechanics of gene expression. Comput Struct Biotechnol J. (2014) 10:63–9. doi: 10.1016/j.csbj.2014.07.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Javierre BM, Burren OS, Wilder SP, Kreuzhuber R, Hill SM, Sewitz S, et al. Lineage-specific genome architecture links enhancers and non-coding disease variants to target gene promoters. Cell. (2016) 167:1369–1384 e1319. doi: 10.1016/j.cell.2016.09.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Koscielny G, An P, Carvalho-Silva D, Cham JA, Fumis L, Gasparyan R, et al. Open Targets: a platform for therapeutic target identification and validation. Nucleic Acids Res. (2017) 45:D985–94. doi: 10.1093/nar/gkw1055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Pandey R, Bakay M, Hakonarson H. CLEC16A-an emerging master regulator of autoimmunity and neurodegeneration. Int J Mol Sci. (2023) 24(9): 8224. doi: 10.3390/ijms24098224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Hudson DF, Amor DJ, Boys A, Butler K, Williams L, Zhang T, et al. Loss of RMI2 increases genome instability and causes a bloom-like syndrome. PloS Genet. (2016) 12:e1006483. doi: 10.1371/journal.pgen.1006483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Colli ML, Ramos-Rodriguez M, Nakayasu ES, Alvelos MI, Lopes M, Hill JLE, et al. An integrated multi-omics approach identifies the landscape of interferon-alpha-mediated responses of human pancreatic beta cells. Nat Commun. (2020) 11:2584. doi: 10.1038/s41467-020-16327-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Dos Santos RS, Marroqui L, Velayos T, Olazagoitia-Garmendia A, Jauregi-Miguel A, Castellanos-Rubio A, et al. DEXI, a candidate gene for type 1 diabetes, modulates rat and human pancreatic beta cell inflammation via regulation of the type I IFN/STAT signalling pathway. Diabetologia. (2019) 62:459–72. doi: 10.1007/s00125-018-4782-0 [DOI] [PubMed] [Google Scholar]
- 29. Kerimov N, Hayhurst JD, Peikova K, Manning JR, Walter P, Kolberg L, et al. A compendium of uniformly processed human gene expression and splicing quantitative trait loci. Nat Genet. (2021) 53:1290–9. doi: 10.1038/s41588-021-00924-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Momozawa Y, Dmitrieva J, Theatre E, Deffontaine V, Rahmouni S, Charloteaux B, et al. IBD risk loci are enriched in multigenic regulatory modules encompassing putative causative genes. Nat Commun. (2018) 9:2427. doi: 10.1038/s41467-018-04365-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Rodriguez A, Alfaro JM, Balthazar V, Pineda Trujillo N. Association analysis of PTPN22, CTLA4 and IFIH1 genes with type 1 diabetes in Colombian families. J Diabetes. (2015) 7:402–10. doi: 10.1111/1753-0407.12192 [DOI] [PubMed] [Google Scholar]
- 32. Robertson CC, Inshaw JRJ, Onengut-Gumuscu S, Chen WM, Santa Cruz DF, Yang H, et al. Fine-mapping, trans-ancestral and genomic analyses identify causal variants, cells, genes and drug targets for type 1 diabetes. Nat Genet. (2021) 53:962–71. doi: 10.1038/s41588-021-00880-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Kircher M, Xiong C, Martin B, Schubach M, Inoue F, Bell RJA, et al. Saturation mutagenesis of twenty disease-associated regulatory elements at single base-pair resolution. Nat Commun. (2019) 10:3583. doi: 10.1038/s41467-019-11526-w [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Bourges C, Groff AF, Burren OS, Gerhardinger C, Mattioli K, Hutchinson A, et al. Resolving mechanisms of immune-mediated disease in primary CD4 T cells. EMBO Mol Med. (2020) 12:e12112. doi: 10.15252/emmm.202012112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Lu X, Chen X, Forney C, Donmez O, Miller D, Parameswaran S, et al. Global discovery of lupus genetic risk variant allelic enhancer activity. Nat Commun. (2021) 12:1611. doi: 10.1038/s41467-021-21854-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. McQuerry JA, McLaird M, Hartin SN, Means JC, Johnston J, Pastinen T, et al. Massively parallel identification of functionally consequential noncoding genetic variants in undiagnosed rare disease patients. Sci Rep. (2022) 12:7576. doi: 10.1038/s41598-022-11589-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Mouri K, Guo MH, de Boer CG, Lissner MM, Harten IA, Newby GA, et al. Prioritization of autoimmune disease-associated genetic variants that perturb regulatory element activity in T cells. Nat Genet. (2022) 54:603–12. doi: 10.1038/s41588-022-01056-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Soleimanpour SA, Gupta A, Bakay M, Ferrari AM, Groff DN, Fadista J, et al. The diabetes susceptibility gene Clec16a regulates mitophagy. Cell. (2014) 157:1577–90. doi: 10.1016/j.cell.2014.05.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Schmid D, Pypaert M, Munz C. Antigen-loading compartments for major histocompatibility complex class II molecules continuously receive input from autophagosomes. Immunity. (2007) 26:79–92. doi: 10.1016/j.immuni.2006.10.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Gonzalez CD, Lee MS, Marchetti P, Pietropaolo M, Towns R, Vaccaro MI, et al. The emerging role of autophagy in the pathophysiology of diabetes mellitus. Autophagy. (2011) 7:2–11. doi: 10.4161/auto.7.1.13044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Gros F, Arnold J, Page N, Decossas M, Korganow AS, Martin T, et al. Macroautophagy is deregulated in murine and human lupus T lymphocytes. Autophagy. (2012) 8:1113–23. doi: 10.4161/auto.20275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Jeong J, Choi YJ, Lee HK. The role of autophagy in the function of CD4(+) T cells and the development of chronic inflammatory diseases. Front Pharmacol. (2022) 13:860146. doi: 10.3389/fphar.2022.860146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Nieves-Bonilla JM, Kiaf B, Schuster C, Kissler S. The type 1 diabetes candidate gene Dexi does not affect disease risk in the nonobese diabetic mouse model. Genes Immun. (2020) 21:71–7. doi: 10.1038/s41435-019-0083-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Pandey R, Bakay M, Hakonarson H. SOCS-JAK-STAT inhibitors and SOCS mimetics as treatment options for autoimmune uveitis, psoriasis, lupus, and autoimmune encephalitis. Front Immunol. (2023) 14:1271102. doi: 10.3389/fimmu.2023.1271102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Barral AM, Thomas HE, Ling EM, Darwiche R, Rodrigo E, Christen U, et al. SOCS-1 protects from virally-induced CD8 T cell mediated type 1 diabetes. J Autoimmun. (2006) 27:166–73. doi: 10.1016/j.jaut.2006.08.002 [DOI] [PubMed] [Google Scholar]
- 46. Luo Y, Vermeer MH, de Haan S, Kinderman P, de Gruijl FR, van Hall T, et al. Socs1-knockout in skin-resident CD4(+) T cells in a protracted contact-allergic reaction results in an autonomous skin inflammation with features of early-stage mycosis fungoides. Biochem Biophys Rep. (2023) 35:101535. doi: 10.1016/j.bbrep.2023.101535 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Fu LY, Zhu T, Zhou X, Yu R, He Z, Zhang P, et al. ChIP-Hub provides an integrative platform for exploring plant regulome. Nat Commun. (2022) 13:3413. doi: 10.1038/s41467-022-30770-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Scherm MG, Ott VB, Daniel C. Follicular helper T cells in autoimmunity. Curr Diabetes Rep. (2016) 16:75. doi: 10.1007/s11892-016-0770-2 [DOI] [PubMed] [Google Scholar]
- 49. Yang D, Liu A, Wu Y, Li B, Nan S, Yin R, et al. BCL2L15 depletion inhibits endometrial receptivity via the STAT1 signaling pathway. Genes (Basel). (2020) 11(7): 816. doi: 10.3390/genes11070816 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Bottini N, Musumeci L, Alonso A, Rahmouni S, Nika K, Rostamkhani M, et al. A functional variant of lymphoid tyrosine phosphatase is associated with type I diabetes. Nat Genet. (2004) 36:337–8. doi: 10.1038/ng1323 [DOI] [PubMed] [Google Scholar]
- 51. Bottini N, Vang T, Cucca F, Mustelin T. Role of PTPN22 in type 1 diabetes and other autoimmune diseases. Semin Immunol. (2006) 18:207–13. doi: 10.1016/j.smim.2006.03.008 [DOI] [PubMed] [Google Scholar]
- 52. Michalek RD, Gerriets VA, Nichols AG, Inoue M, Kazmin D, Chang CY, et al. Estrogen-related receptor-alpha is a metabolic regulator of effector T-cell activation and differentiation. Proc Natl Acad Sci U.S.A. (2011) 108:18348–53. doi: 10.1073/pnas.1108856108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Yang BH, Wang K, Wan S, Liang Y, Yuan X, Dong Y, et al. TCF1 and LEF1 control treg competitive survival and tfr development to prevent autoimmune diseases. Cell Rep. (2019) 27:3629–3645 e3626. doi: 10.1016/j.celrep.2019.05.061 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The sequencing data generated by this study are available from the NCBI Short Read Archive (SRA) under accession PRJNA1120596.