

Abstract
Eukaryotic cells contain several small, highly abundant, nuclear-localized non-coding RNAs which play important roles in gene expression. These small nuclear RNAs (snRNAs) together with their associated proteins form ribonucleoprotein particles, called snRNPs, which function in splicing of introns from primary genomic transcripts. Through a combination of RNA-RNA and RNA-protein interactions, two of the snRNPs named U1 and U2 recognize the 5’ and 3’ splice sites and the branch site of introns. A complex remodeling of RNA-RNA and protein-based interactions follows, which results in the assembly of a catalytically competent spliceosome, in which the snRNAs and their bound proteins play central roles. During this process, the basepairing interaction between two of the snRNAs, U4 and U6, is disrupted and replaced by extensive basepairing between U2 and U6. U6 itself replaces U1 at the 5’ splice site and U5 forms non-canonical basepairing interactions with the exonic sequences immediately adjacent to the 5’ and 3’ splice sites. Thus, RNA-RNA interactions involving U2, U5 and U6 serve to juxtapose the reacting groups of the first and second steps of splicing. In addition to its structural role, U6 is also thought to participate in formation of the spliceosomal active site. Recent analyses point to an additional role for snRNAs in regulation of various aspects of RNA biogenesis, from transcription to polyadenylation and RNA stability. These snRNP-mediated regulatory roles likely serve to ensure the coordination of the different processes involved in biogenesis of RNAs and point to the central importance of the snRNAs in eukaryotic gene expression.
Keywords: Spliceosome, snRNAs, U6, U2, splicing, ribozyme, catalysis, RNA World, group II introns, U1, U4, U5, U11, U12
Introduction: The challenge of splicing and evolution of eukaryotic snRNAs
A fascinating feature of the modern eukaryotic genes is the nearly ubiquitous presence of intervening sequences or introns, which interrupt the continuity of the information content of genes. Thus, before primary gene transcripts can be used by the cell, introns must be accurately removed or “spliced”[1]. In addition, recent research indicate that introns themselves often harbor regulatory or otherwise functional sequences and their accurate and timely removal is often critical for their cellular function [2]. The intronic sequences in higher eukaryotic genes are much longer than the non-intronic sequences - the exons - and the sequence-based information that specify the intron-exon boundaries are highly complex and poorly understood. Thus, the accurate separation of these two sets of functional sequences that coexist in eukaryotic primary transcripts is a highly challenging task for the eukaryotic gene expression machinery.
Although the modern mammalians have one of the most complex splicing patterns among extant eukaryotes, it is likely that even in primordial eukaryotes splicing was already a highly complex process. Based on currently accepted models of evolution of eukaryotes, introns likely originated from self-splicing ribozymes that dated from pre-cellular life and constituted the majority of the genomes of ancient eukaryotes [3]. Later on, likely in order to prevent genomic instability, the introns lost their self-splicing capacity and instead, the splicing function was delegated to a cellular machine, the spliceosome, which acted in trans to remove introns from primary transcripts. Although the origin and evolution of the early spliceosomes is still largely mysterious, several lines of evidence suggest that they likely evolved from self-splicing introns [4]. This hypothesis is partly based on the fact that the mechanism of intron removal by the spliceosome, performed through two consecutive transesterification reactions resulting in removal of a branched lariat intron, is identical to the splicing reaction performed by a class of extant self-splicing introns called the group II introns [5](Figs. 1 and 2). These introns, which are found in all three kingdoms of life, are RNA-centric catalytic sequences composed of a number of basepaired RNA structures called “domains” (Fig. 2)[6]. Extensive research has elucidated the identify of the catalytically essential sequences in these introns (Fig. 2)[6]. Intriguingly, the RNA components of the spliceosome, the snRNAs, show unmistakable similarities to fragments of the catalytically essential domains of group II introns in sequence, secondary structure and function. Of the five major spliceosomal snRNAs (U1, U2, U4, U5 and U6), three of them (U2, U5 and U6) have clear structural and functional similarities to critical domains of group II introns (Fig. 2). Domain swapping experiments have indicated that isolated domains of group II introns and U5 and U6 snRNA substructures could functionally replace each other, proving their functional equivalence [5,6]. Another set of sequences in group II introns are functionally equivalent to U1 snRNA, despite the lack of structural similarity (Fig. 2). Currently there are no known functional or structural equivalents for the U4 snRNA in group II introns and the evolutionary origin of this snRNA is completely unknown. On the other hand, a number of domains of the group II introns do not have an equivalent among the spliceosomal snRNAs and it is likely that in the spliceosome, these RNA domains are replaced by spliceosomal proteins. While the possibility of convergent evolution cannot be formally ruled out, the above-mentioned similarities strongly suggest that at least a number of snRNAs are evolutionary remnants of primordial self-splicing ribozymes.
Fig. 1.

U6 and U2 snRNAs and the mRNA at the time of first and second steps of splicing. The location of U6, U2 and the U6 intramolecular stemloop is shown. The intron is shown by a thick light blue line connecting the two exons (shown). Position of the 5’ splice site (5’SS), 3’ splice site (3’SS) and branch site are shown. Solid arrows point to the site of the nucleophilic attack during the two steps of splicing. The first step involves a nucleophilic attack by the 2’ hydroxyl group of a specific adenosine residue in the intron, the branch site adenosine (the bulged A), on the 5’ splice site. This leads to a transesterification reaction in which the 2’ oxygen of the branch site adenosine replaces the 3’ oxygen of the last nucleotide of the upstream exon. The result of this reaction is the release of the first exon and the formation of an unusual 2’−5’ linkage between the branch site adenosine and the first nucleotide of the intron (right panel). During the second step, the free 3’ hydroxyl moiety of the newly released exon is activated for a similar nucleophilic attack on the 3’ splice site, resulting in ligation of the two exons and release of the intron as a branched lariat. Base-pairing interactions are shown by short black lines. The location of the 2’−5’ linkage formed after the first step of splicing at the branch site is shown.
Fig. 2.
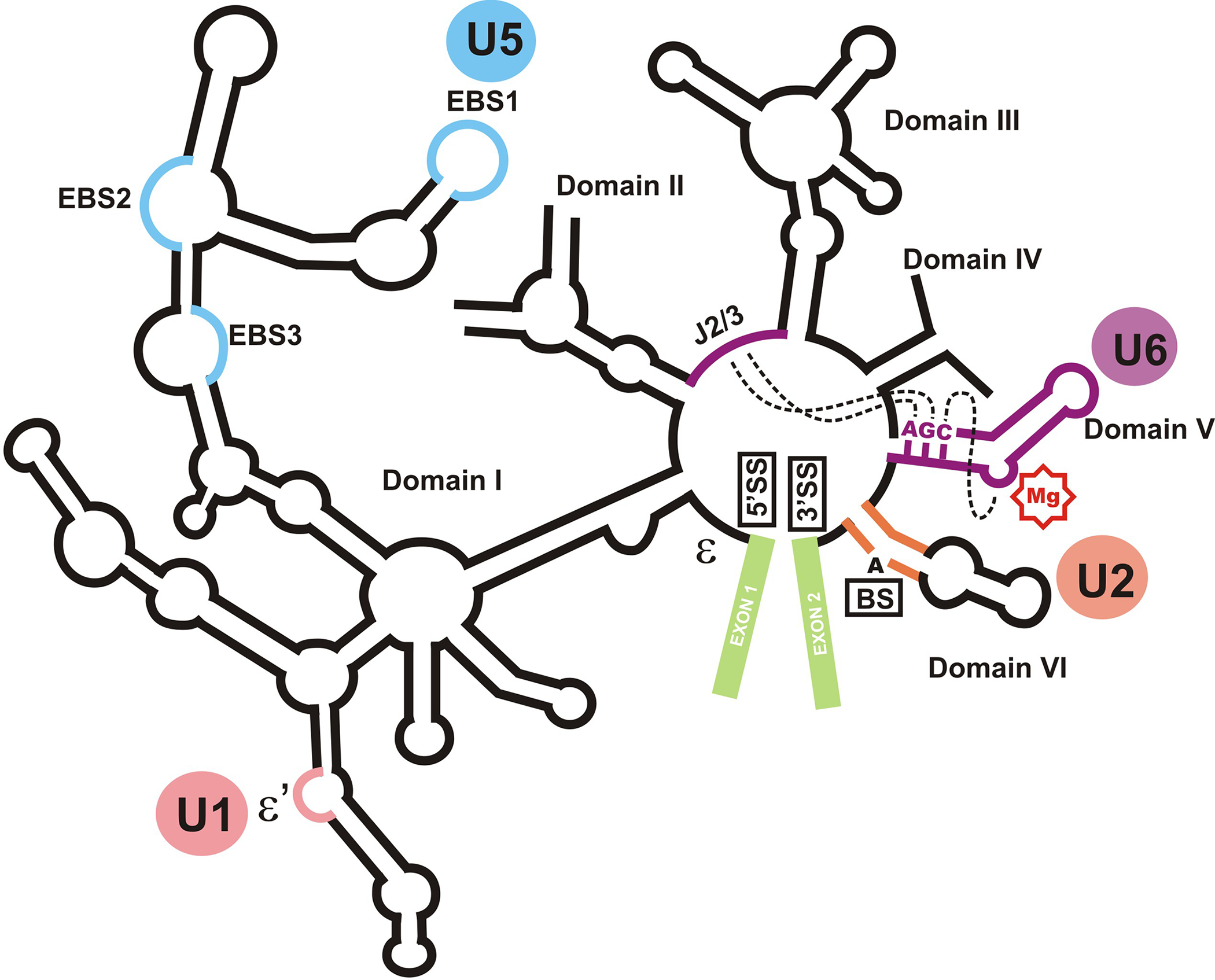
The structural organization of the group II self-splicing intron aI5γ. The location of domains I through VI, the two exons (shown in green), the splice sites and the branch site (5’SS, 3’SS and BS, respectively) are shown. The position of J2/3 and the AGC sequence are indicated. The metal binding site of domain V is shown by a red “Mg” sign. Dashed lines connects regions which are juxtaposed to form the catalytic core. The circles denote the functional equivalent of each domain or subdomain in the spliceosome. ε and ε’ sites, which are involved in an interaction important in recognition of the 5’ splice site are shown.
Roles of snRNAs in the spliceosome
The snRNAs were first discovered in 1970’s as small, highly abundant nuclear RNAs which formed the core of ribonucleoprotein particles which showed strong reactivity with the immune sera from patients with autoimmune disorders [7,8]. Further analysis indicated the presence of sequence complementarity between one of the snRNAs, U1, and the sequences found at the 5’ splice site of primary transcripts, ultimately leading to the discovery of their involvement in splicing [9]. Further research identified a second set of snRNAs, named U11, U12, U4atac and U6atac which are functional counterparts of U1, U2, U4 and U6 snRNAs, respectively, and participate in the formation of a “minor” spliceosome which is responsible for removal of an atypical subset of introns, most of which have alternative consensus sequences at the splice sites and branch site [10]. U5 snRNA is found in both “major” and “minor” spliceosomes. Each of the nine spliceosomal snRNAs are stably associated with a set of proteins, creating the small nuclear ribonucleoprotein (snRNP) particles which form the main functional subunits of the spliceosome. Analysis of the spliceosomal function suggest that snRNPs and several non-snRNP spliceosomal proteins assemble on each intron in a stepwise, elaborate fashion through a large number of conformational rearrangements which start from the recognition of the splice junctions and culminate in splicing catalysis, followed by disassembly and recycling of the spliceosomal components [1,11](Fig. 3).
Fig. 3.
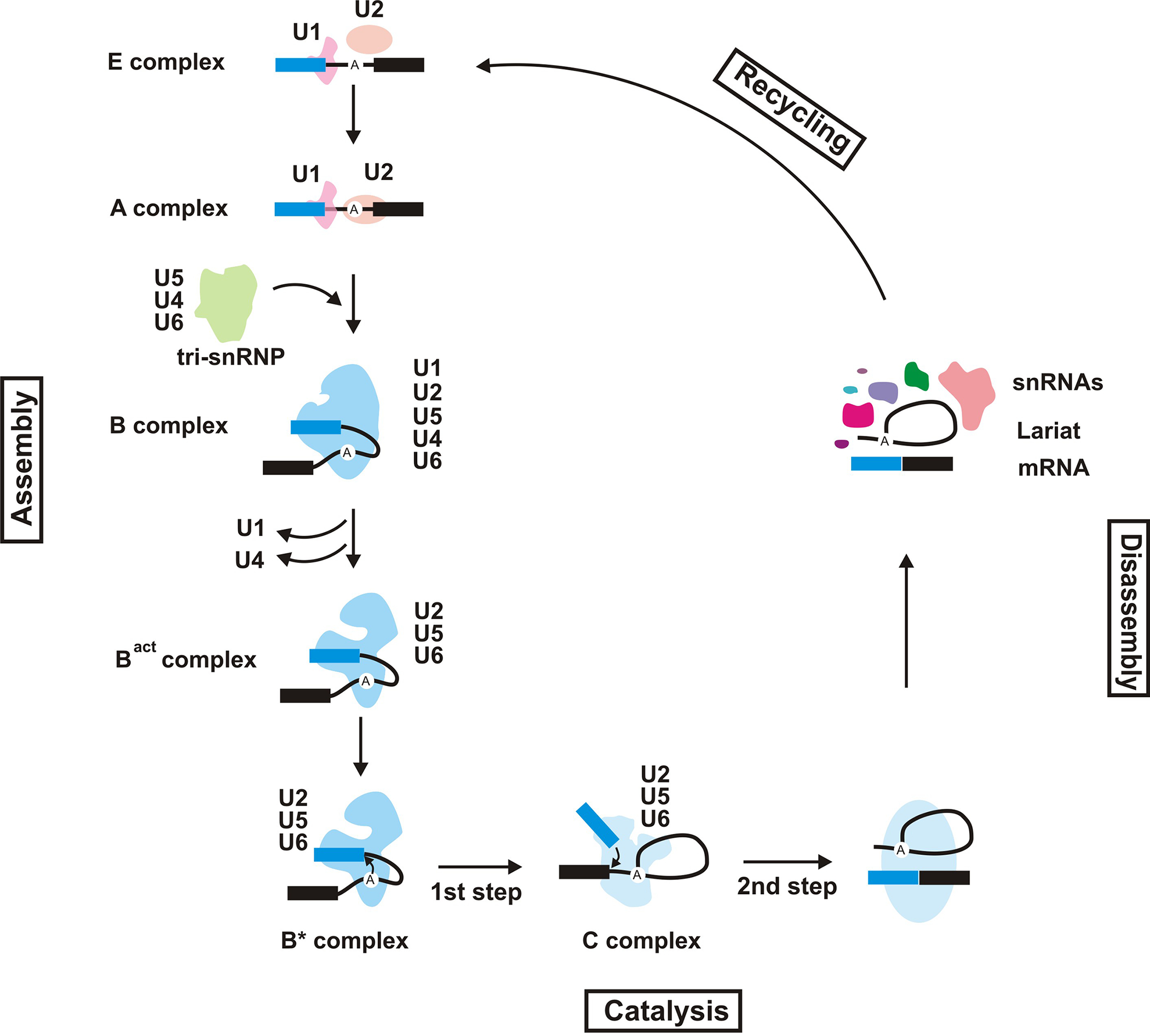
The spliceosomal cycle. The spliceosomal complexes formed during a splicing cycle are shown. The snRNAs present in each splicing complex is indicated. The exons on the pre-mRNA are shown as blue and black rectangles, with the intron drawn as a thin line connecting the two. The position of the branch site adenosine is marked.
Recognition of the 5’ splice site by U1 snRNP
Recognition of introns in primary transcripts is partly mediated by detection of “consensus” sequences found at the junction of introns and exons, the 5’ and 3’ splice sites. These consensus sequences are rather short: 5’AG/GURAGU and 5’YAG/G for 5’ and 3’ splice sites, respectively, where R denotes either a G or an A, and Y denotes a C or a U and forward slash marks the location of the splice site [1,11]. Interestingly, in higher eukaryotes and especially in mammalians, the splice site sequences are highly degenerate and in many cases significantly deviate from the consensus sequence, thus necessitating an elaborate, multi-step recognition mechanism mediated by a combination of RNA-RNA basepairing and RNA-protein interactions [1,11].
The association of U1 snRNP and its functional equivalent in the “minor” spliceosomes, U11, with the 5’ splice site is one of the earliest and arguably most important events in the spliceosomal assembly pathway [11,12](Fig. 3, the E complex). The recognition and binding of 5’ splice site is mediated both by basepairing of a single-stranded sequence at the 5’ end of U1 snRNA to the 5’ splice site and through an intricate web of interactions between the pre-mRNA and U1C, a U1-specific protein [5,11,13]. Interestingly, a high-resolution structure of U1 snRNP indicated the presence of a number of interactions between U1C and the nucleotides at the 5’ end of U1 which basepair to the 5’ splice site, thus providing a structural basis for the dual RNA-protein recognition of the 5’ splice site [12,13]. While this RNA-protein recognition of the 5’ splice site by U1 is functionally critical for the majority of cellular transcripts in vivo, several other proteins also contribute to the selection of the 5’ splice site [12]. Further, at least some primary transcripts can be spliced in the absence of U1 snRNP in vitro, pointing to extensive redundancy in the splicing machinery. Finally, the binding of U1 is not necessarily synonymous with productive splicing, since the binding of U1 to sequences involved in negative regulation of splicing has been documented [12]. Current data suggest that the binding of U1 to such elements is important in splicing regulation and exclusion of pseudo splice sites, underscoring the importance of sequence context in splicing.
The U2 snRNP and recognition of the branch site and 3’ splice site
Once bound to the 5’ splice site in a sequence context which is conducive to splicing, U1 helps initiate the spliceosomal assembly by forming a network of interactions with U2 snRNP that plays a dominant role in recognition of the 3’ splice site and the branch site, another region in the introns which is recognized by the spliceosomes [12,14]. Branch sites, which are typically located ~30 nucleotides from the 3’ end of introns, contain the adenosine which acts as the nucleophile of the first transesterification step of splicing and forms the branched, lariat structure found in the splicing intermediates and post-splicing introns (Figs. 1 and 3). In early spliceosomes, U2 snRNP is loosely associated with the end of the intron through protein-mediated interactions. However, in an ATP-dependent step which involves the displacement of intron-bound proteins and remodeling of basepairing interactions within the U2 snRNA, U2 forms a stable interaction with the branch site and 3’ splice site [11,13,15](Fig. 3, the A complex). This interaction is partly mediated through a base-pairing interaction between U2 and sequences flanking the branch site adenosine (Fig. 4), and is stabilized by several RNA-protein interactions. The basepairing between U2 and the branch site leaves the branch site adenosine in an unpaired, extrahelical conformation necessary for efficient splicing (Fig. 4)[5,6]. In addition, the interaction between U1 and U2 snRNPs generates a loop that brings the 5’ and 3’ splice sites together and helps to “define” the introns and exons [14].
Fig. 4.

The known RNA-RNA interactions at the spliceosomal catalytic core at the time of the first step of splicing. The central domain of human U6 and the 5’ domain of human U2, which contain the sequences necessary for splicing in vivo, are shown. The basepairing interactions between U2 and U6 which form helices I, II and III are shown. The exons in pre-mRNA are drawn as rectangles, with the intron as a thin solid line. The sequence of the branch site of intron is shown. The conserved stemloop I of U5 snRNA is shown at the bottom, with the thin dashed lines marking non-canonical basepairs between the exonic sequences next to the splice sites and the U5 loop I. The ACAGAGA and AGC domains are highlighted in yellow. The dotted, curved red line joining circled nucleotides points to a tertiary interaction detected in activated spliceosomes. The dotted red line connects the binding sites for two functionally-required metal ions (shown by red “Mg” signs) which may be located near each other in activated spliceosomes.
Formation of a catalytically active spliceosome
Three of the snRNPs, U4, U5 and U6, form a ternary complex termed the “tri-snRNP” and collectively integrate into the assembling spliceosomes [1,5](Fig. 3, the B complex). Of all the spliceosomal snRNAs, U6 is the most conserved and contains two invariant domains, the ACAGAGA and AGC boxes, which play a critical functional role in splicing (Fig. 4)[5,16]. Further, it contains an intramolecular stemloop (ISL) which is almost identical to the catalytic domain of group II introns and similar to its group II intron counterpart, binds a functionally required divalent cation, pointing to a critical role in splicing catalysis for this snRNA (Fig. 4, 5)[5,16]. Perhaps in order to prevent it from prematurely forming a catalytically active structure, within the tri-snRNP U6 is kept in an inactive conformation through a basepairing interaction with U4 snRNA that prevents the formation of its functionally critical ISL. Current data do not indicate any additional functions for U4 snRNA except acting as a negative chaperon for U6. Once the basepaired U4/U6 complex joins the spliceosome in association with U5 within the tri-snRNP, the U4/U6 duplex is unwound in a tightly controlled manner and U2 snRNA replaces U4 as the basepairing partner of U6 (Fig. 4)[1,5,16]. In addition, U6 replaces U1 at the 5’ splice site, forming canonical and non-canonical basepairing interactions with this sequence (Fig. 3, Bact complex; Fig. 4). The basepairing interactions between U6 and U2 allow the formation of the U6 ISL and further, serve to juxtapose the branch site, which is bound to the branch-binding sequence in U2, and the 5’ splice site, which is bound by U6 (Fig. 4). At the same time, U5 snRNA forms non-canonical basepairing interactions with the exon sequences immediately adjacent to the splice sites and participates in aligning the exons to ensure their optimal positioning for the second step of splicing (Fig. 4)[1,5,16]. These basepairing rearrangements are accompanied by an extensive rearrangement of protein-protein and protein-RNA interactions, culminating in the formation of catalytically competent spliceosomes (B* and C complexes, Fig. 3). U1 and U4 are not stably associated with fully assembled spliceosomes, thus leaving U2, U5 and U6 snRNAs as the only spliceosomal RNA components required for catalysis (Fig. 4)[5]. Interestingly, as mentioned above, these three snRNAs have clear structural and functional counterparts in self-splicing group II introns, raising the possibility that the catalytic core of the two splicing systems may be closely similar (Fig. 2)[5,6,17–19].
Fig. 5.

Structural and functional similarities between the catalytically crucial domain V of group II introns and U6 snRNA. The central domains of human U6 and 5’ domain of human U2 snRNAs are shown. The pre-mRNA is shown as a lighter green line, with the sequence of the branch site indicated. The ACAGAGA and AGC sequences in U6 and domain V are highlighted in yellow. The sites of phosphorothioate interference, which may point to metal binding sites, are shown with red marks.
Catalysis of splicing: the role of snRNAs
Mutagenesis studies have shown that at least in vitro, the conserved loop of U5, which was previously shown to be the functionally important domain of the molecule (Fig. 4), was in fact dispensable for splicing [1,5]. On the other hand, it has been recently shown that under certain conditions, several positions within the branch binding sequence of U2 snRNA can be mistakenly recognized as the 5’ splice site [20]. These results imply that the branch binding sequence of U2, which is functionally the most critical region of this snRNA, is not essential for spliceosomal catalysis at least under certain conditions. As the rest of U2 seems to mainly fulfill structural roles by forming basepairing interactions with other U2 sequences or with U6 snRNA [5,15], these results suggest that U6 snRNA may be the only RNA that is absolutely crucial for splicing catalysis, at least under the conditions studied so far.
Several additional lines of evidence suggest that U6 may form part of the catalytic domain of the spliceosome [5,16]. U6 is the most conserved of all spliceosomal snRNAs and several point mutations in its two evolutionarily invariant sequences, the ACAGAGA and AGC boxes, lead to a block in splicing, pointing to a critical function for these two sequences (Fig. 4, 5). Crosslinking and mutational complementation analyses have indicated that the first step of splicing occurs in close proximity to the ACAGAGA box, suggesting that this sequence is in immediate vicinity of or even forms part of the spliceosomal active site. Current data suggest that in group II introns, the active site is formed by juxtaposition of AGC triad and the asymmetric internal bulge of domain V along with a short purine-rich sequence (J2/3, Fig. 2) which is considered functionally equivalent to the ACAGAGA box in U6 [19]. Interestingly, U6 contains the equivalent of all these sequences which form the active site in group II introns, and close similarities in phosphorothioate interference patterns between catalytic domain V of group II introns and U6 (Fig. 5) suggest that they may be functionally related [5, 17–19]. Further, hydroxyl radical footprinting and in vivo mutagenesis studies have pointed to the proximity of AGC triad, the ACAGAGA box and the area near the bulged residue in ISL in functional spliceosomes (Fig. 4)[5], perhaps in an arrangement similar or even identical to the one found in group II introns [5,17–19].
Interestingly, analyses on in vitro-transcribed, protein-free U6 and U2 snRNAs indicate that they can efficiently form a basepaired complex in vitro which in many respects resembles the one formed in the activated spliceosomes (Fig. 4)[5,21–23]. Further, it has been shown that the in vitro-assembled human U2/U6 complex can indeed catalyze a two step splicing reaction which closely resembles the one catalyzed by the self-splicing group II introns and the spliceosome [24,25]. Based on the above data, the snRNAs seem to be fully competent to form the majority, if not all, of the spliceosomal active site and to perform catalysis, similar to the self-splicing group II introns, albeit with much lower efficiency.
If we assume that the spliceosome is an RNA catalyst, the snRNAs are unusual ribozymes in many respects, perhaps most importantly they are unusually small compared to other natural ribozymes catalyzing splicing reactions. The larger size of other natural splicing ribozymes is thought to allow them to fold into complex tertiary structures, which in turn enable them to create sophisticated active sites necessary for such complex reactions. It is conceivable that due to their short length, the U6 and U2 snRNAs at best form an inefficient splicing ribozyme, which requires other spliceosomal factors for stable positioning of the active site elements and the reacting groups. While the exact role played by the proteins in the spliceosomal catalytic core is mostly unknown, their possible roles could range from assisting the snRNAs in assuming their functional structure, assisting in or independently coordinating critical metal ions and participating in the positioning of the substrates, to independently forming part of the active site and even direct involvement in catalysis [26].
Beyond splicing: other biological roles of snRNAs
Although the spliceosomal snRNAs (and their minor spliceosomal counterparts U11, U12, U4atac and U6 atac) play major roles in spliceosomal function, recent data suggest additional roles in regulation of gene expression for the snRNP particles. The interaction of U1 and 5’ splice site, in addition to its function in splice site selection, also seems to play a role in stabilization of some messages[12]. Both U1 and U2 snRNPs have been implicated in transcriptional regulation through stimulation of the rate of formation of the first phosphodiester bond at transcription initiation and interaction with a component of the pre-initiation complex, Transcription Factor II H (TFIIH), respectively [27,28]. U1 snRNP seems to also regulate the efficiency of polyadenylation via the interaction of a U1-specific protein, U1A, with a component of the Cleavage and Polyadenylation Stimulating Factor (CPSF) [12]. In addition, it has been reported that binding of U1 to a 5’ splice site-like sequence in the 3’ untranslated region (3’UTR) of some mRNAs inhibits their polyadenylation, leading to degradation of the RNA [12]. Thus, the snRNAs and their bound proteins seem to act in coordination of the various steps in gene expression, in addition to playing the central role in splicing.
Another spliceosomal snRNP, the Spliced Leader (SL) particle, plays a critical role as the splice donor in a non-canonical, trans splicing reaction mainly observed in some protozoa and lower invertebrates [29]. In the trans splicing reaction, SL is treated as a mini exon plus a short intron, with the exonic sequences “spliced” in trans to a 3’ splice site on the primary transcripts of trans splicing organisms. Thus, unlike the other spliceosomal snRNAs, SL is consumed during the trans splicing reaction.
While the majority of snRNAs play a role in splicing, there are other abundant, small nuclear-localized RNAs which play critical roles in other cellular processes. A non-spliceosomal snRNA, U7, functions in 3’ end processing of replication-dependent histone mRNAs, which are not polyadenylated and instead terminate in a conserved stem-loop (SL) structure generated by an endonucleolytic cleavage of the pre-mRNA [30]. Similar to the spliceosomal snRNAs, U7 also forms a snRNP by associating with a set of proteins which together form the so-called Sm ring, and is recruited to histone pre-mRNA primarily through basepairing interactions via its 5’ end with a purine-rich Histone Downstream Element (HDE) which is located in the vicinity of cleavage site. Together with a protein which binds the SL element, U7 recruits a complex that triggers endonucleolytic cleavage between SL and the HDE by the CPSF73 endonuclease, thus forming the mature histone mRNA.
Conclusion
As detailed above, the ability of snRNAs to form strong, specific interactions via basepairing with another RNA is extensively utilized in the spliceosome and during processing of the histone 3’ ends. Basepairing interactions contribute to substrate recognition (U1, U2 and U7), positioning of the branch site in a strained, catalytic bulged conformation (U2), regulation of the activity of another snRNA (U4), and juxtaposition of reactive substrates (U2, U5 and U6). Since RNA-RNA interactions similar to those formed by some of the spliceosomal snRNAs play identical or closely-related roles in group II introns, it is conceivable that the robustness of RNA-RNA interactions has led to their preservation throughout the evolution of the spliceosome from group II-like ancient ribozymes. While RNAs can perform the above-mentioned tasks with ease and even more effectively than proteins, when it comes to catalysis, proteins seem to have an advantage over RNA, at least in the case of natural ribozymes. The evolutionary reason behind the preservation of U6 snRNAs as a constituent of the spliceosomal catalytic core remains an open question.
Another feature of the snRNAs is their participation in multiple sets of basepairing interactions that at times are mutually exclusive and thus, act as switches between different functional states. The presence of such interactions underscores the highly complex evolutionary pressures under which the snRNAs have evolved. In addition, most of the snRNAs bind a number of proteins which play important functional roles. While in many cases, these proteins complement the function of the snRNA to which they bind, emerging evidence suggest that they can impart a completely novel function on the snRNP. Research in the coming years is likely to provide additional instances of multi-functionality of the snRNPs and further elucidate their contribution to the highly complex network of interactions which regulate the eukaryotic gene expression.
Summary
The majority of cellular snRNAs function in splicing, with their ability to form specific basepairing interactions extensively utilized in recognition of functional sequence elements in primary transcripts. Examples include the interactions between U1 and U6 and the 5’ splice site, U2 and the branch site, and U5 and the exonic sequences.
Another set of RNA-RNA interactions occur between snRNAs and plays important roles in regulation of the timing of their folding into their active structure (in the case of the interaction between U4 and U6) or form a structural scaffold for juxtaposition of the reactive groups in the splicing reaction ( formation of the U2/U6 basepaired complex).
In addition to forming RNA-RNA interactions, U6 snRNA seems to play a critical role in catalysis of the splicing reaction. Whether the spliceosomal proteins play a role in catalysis remains to be determined.
Another aspect of the function of snRNAs is forming interactions with a set of proteins which play important roles in spliceosomal assembly, regulation and coordination of splicing and other steps of gene expression.
Biographies
Saba Valadkhan
Saba Valadkhan started her scientific career in Professor James Manley’s laboratory in Columbia University as a graduate student, where she worked on elucidating the structure and function of U6 and U2 snRNAs. In 2004, she joined the Center for RNA Molecular Biology in Case Western Reserve University School of Medicine as an assistant professor and continued her work on the catalytic properties of the in vitro assembled, protein-free complex of U6 and U2 snRNAs and their relationship to spliceosomal catalysis.
Lalith S. Gunawardane
Lalith Gunawardane is currently a postdoctoral scientist in Valadkhan Lab in the Center for RNA Molecular Biology at Case Western Reserve University. As a graduate student in Professor Haruhiko Siomi’s laboratory in Keio University School of Medicine in Tokyo, he worked on the small non-coding RNAs and their interactions with proteins. Since joining Valadkhan Lab, he has continued his work on analysis of function of nuclear non-coding RNAs in mammalian cells.
References
- 1.Will CL, Luhrmann R: Spliceosome structure and function. In The RNA World. Edited by Gesteland RF, Cech TR, Atkins JF. Cold Spring Harbor Laboratory Press; 2006:369–400. [Google Scholar]
- 2.Chorev M, Carmel L (2012) The function of introns. Front Genet, 3:55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Roy SW, Irimia M (2009)Splicing in the eukaryotic ancestor: form, function and dysfunction. Trends Ecol. Evol. (Amst.) 24:447–455. [DOI] [PubMed] [Google Scholar]
- 4.Rodríguez-Trelles F, Tarrío R, Ayala FJ (2006) Origins and evolution of spliceosomal introns. Annu. Rev. Genet 40:47–76. [DOI] [PubMed] [Google Scholar]
- 5.Valadkhan S (2010) Role of the snRNAs in spliceosomal active site. RNA Biol, 7:345–353. [DOI] [PubMed] [Google Scholar]
- 6.Pyle AM, Lambowitz AM: Group II Introns: Ribozymes That Splice RNA and Invade DNA. In The RNA World. Edited by Gesteland RF, Cech TR, Atkins JF. Cold Spring Harbor Laboratory Press; 2006:469–506. [Google Scholar]
- 7.Lerner MR, Steitz JA (1979) Antibodies to small nuclear RNAs complexed with proteins are produced by patients with systemic lupus erythematosus. Proc Natl Acad Sci U S A, 76:5495–5499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zieve G, Penman S (1976) Small RNA species of the HeLa cell: metabolism and subcellular localization. Cell, 8:19–31. [DOI] [PubMed] [Google Scholar]
- 9.Lerner MR, Boyle JA, Mount SM, Wolin SL, Steitz JA (1980) Are snRNPs involved in splicing? Nature, 283:220–224. [DOI] [PubMed] [Google Scholar]
- 10.Will CL, Lührmann R (2005) Splicing of a rare class of introns by the U12-dependent spliceosome. Biol. Chem, 386:713–724. [DOI] [PubMed] [Google Scholar]
- 11.Wahl MC, Will CL, Lührmann R (2009) The spliceosome: design principles of a dynamic RNP machine. Cell, 136:701–718. [DOI] [PubMed] [Google Scholar]
- 12.Buratti E, Baralle D (2010) Novel roles of U1 snRNP in alternative splicing regulation. RNA Biol, 7:412–419. [DOI] [PubMed] [Google Scholar]
- 13.Valadkhan S, Jaladat Y (2010) The spliceosomal proteome: at the heart of the largest cellular ribonucleoprotein machine. Proteomics, 10:4128–4141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shao W, Kim H-S, Cao Y, Xu Y-Z, Query CC (2012) A U1-U2 snRNP Interaction Network During Intron Definition. Mol. Cell. Biol, 32:470–478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Perriman R, Ares M Jr (2010) Invariant U2 snRNA Nucleotides Form a Stem Loop to Recognize the Intron Early in Splicing. Molecular Cell, 38:416–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Valadkhan S (2007) The spliceosome: a ribozyme at heart? Biol. Chem, 388:693–697. [DOI] [PubMed] [Google Scholar]
- 17.Michel F, Costa M, Westhof E (2009) The ribozyme core of group II introns: a structure in want of partners. Trends Biochem. Sci, 34:189–199. [DOI] [PubMed] [Google Scholar]
- 18.Dayie KT, Padgett RA (2008) A glimpse into the active site of a group II intron and maybe the spliceosome, too. RNA, 14:1697–1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Keating KS, Toor N, Perlman PS, Pyle AM (2010) A structural analysis of the group II intron active site and implications for the spliceosome. RNA, 16:1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Smith DJ, Query CC, Konarska MM (2007) trans-splicing to spliceosomal U2 snRNA suggests disruption of branch site-U2 pairing during pre-mRNA splicing. Mol. Cell, 26:883–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Valadkhan S, Manley JL (2000) A tertiary interaction detected in a human U2-U6 snRNA complex assembled in vitro resembles a genetically proven interaction in yeast. RNA, 6:206–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Butcher SE (2011) The spliceosome and its metal ions. Met Ions Life Sci, 9:235–251. [DOI] [PubMed] [Google Scholar]
- 23.Guo Z, Karunatilaka KS, Rueda D (2009) Single-molecule analysis of protein-free U2-U6 snRNAs. Nat. Struct. Mol. Biol, 16:1154–1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jaladat Y, Zhang B, Mohammadi A, Valadkhan S (2011) Splicing of an intervening sequence by protein-free human snRNAs. RNA Biol, 8: 372–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Valadkhan S, Mohammadi A, Jaladat Y, Geisler S (2009) Protein-free small nuclear RNAs catalyze a two-step splicing reaction. Proc. Natl. Acad. Sci. U.S.A, 106:11901–11906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hsieh J, Andrews AJ, Fierke CA (2004) Roles of protein subunits in RNA-protein complexes: lessons from ribonuclease P. Biopolymers, 73:79–89. [DOI] [PubMed] [Google Scholar]
- 27.McKay SL, Johnson TL (2011) An investigation of a role for U2 snRNP spliceosomal components in regulating transcription. PLoS ONE, 6:e16077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kwek KY, Murphy S, Furger A, Thomas B, O’Gorman W, Kimura H, Proudfoot NJ, Akoulitchev A (2002) U1 snRNA associates with TFIIH and regulates transcriptional initiation. Nature Structural & Molecular Biology, 9:800–805. [DOI] [PubMed] [Google Scholar]
- 29.Lasda EL, Blumenthal T (2011) Trans-splicing. Wiley Interdisciplinary Reviews: RNA, 2:417–434. [DOI] [PubMed] [Google Scholar]
- 30.Ideue T, Adachi S, Naganuma T, Tanigawa A, Natsume T, Hirose T (2012) U7 small nuclear ribonucleoprotein represses histone gene transcription in cell cycle-arrested cells. Proc. Natl. Acad. Sci. U.S.A, 109:5693–5698. [DOI] [PMC free article] [PubMed] [Google Scholar]