

Abstract
Background
The coagulopathy of traumatic brain injury (TBI) remains poorly understood. Contradictory descriptions highlight the distinction between systemic and local coagulation, with descriptions of systemic hypercoagulability despite intracranial hypocoagulopathy. This perplexing coagulation profile has been hypothesized to be due to tissue factor release. The objective of this study was to assess the coagulation profile of TBI patients undergoing neurosurgical procedures. We hypothesize that dura violation is associated with higher tissue factor and conversion to a hypercoagulable profile and unique metabolomic and proteomic phenotype.
Methods
This is a prospective, observational cohort study of all adult TBI patients at an urban, level-1 trauma center who underwent a neurosurgical procedure from 2019 to 2021. Whole blood samples were collected before and then one hour following dura violation. Citrated rapid and tissue plasminogen activator (tPA) thrombelastography (TEG) were performed, in addition to measurement of tissue factory activity, metabolomics, and proteomics.
Results
Overall, 57 patients were included. The majority (61%) were male, the median age was 52 years, 70% presented after blunt trauma, and the median Glasgow Coma Score was 7. Compared to pre-dura violation, post-dura violation blood demonstrated systemic hypercoagulability, with a significant increase in clot strength (maximum amplitude of 74.4 mm versus 63.5 mm, p<0.0001) and a significant decrease in fibrinolysis (LY30 on tPA-challenge TEG of 1.4% versus 2.6%, p =0.04). There were no statistically significant differences in tissue factor. Metabolomics revealed notable increases in metabolites involved in late glycolysis, cysteine and one carbon metabolites, and metabolites involved in endothelial dysfunction/arginine metabolism/responses to hypoxia. Proteomics revealed notable increase in proteins related to platelet activation and fibrinolysis inhibition.
Conclusion
A systemic hypercoagulability is observed in TBI patients, characterized by increased clot strength and decreased fibrinolysis and a unique metabolomic and proteomics phenotype independent of tissue factor levels.
Level of Evidence:
n/a (basic science)
Keywords: traumatic brain injury, trauma induced coagulopathy, metabolomics, proteomics, thrombelastography
Social media summary:
The coagulopathy of TBI remains poorly understood, but Coleman et al show that TBI and dura violation is associated with robust hypercoagulability and unique metabolomic phenotype without tissue factor increase. #surgscience @JuliaColemanMD @OhioStateSurg @CuDeptSurg @mitchelljayc
Introduction
The coagulopathy of traumatic brain injury (TBI) remains poorly understood. While coagulopathy occurs in nearly half of patients who incur a head injury, obstacles remain in predicting these patients and understanding the derangements in hemostatic capacity after TBI, in particular if also accompanied by polytrauma or hemorrhagic shock1. Contradictory descriptions in the literature highlight the distinction between systemic and local coagulation environment, with characterization of hypocoagulopathy and progression of intracranial hemorrhage to hypercoagulability due to systemic immune response, inflammation, and platelet hyperactivity2–4. This latter pathologic hypercoagulability has been linked to higher rates of macrothrombotic morbidity including deep venous thrombosis and microthrombotic morbidity like acute lung and kidney injury5–7.
The mechanisms underlying the hypercoagulability of TBI remain elusive, however previous work suggest a role of complement, platelet activation, fibrinolysis inhibition, endotheliopathy, and prothrombotic microparticles, among other players8. The principal process of TBI-related hypercoagulability is thought to be driven by release of tissue factor from the dura, or another neuronal source, into the general circulation9–11. These higher levels of tissue factor are hypothesized to drive increased thrombin generation and alter platelet responsiveness following cortical injury. Despite these theories, which ultimately suggest that the coagulopathy of TBI is an expression of disseminated intravascular coagulopathy (DIC), data to support tissue factor release after brain injury resulting in consumptive coagulopathy has not been proven and as such, a gap in the literature remains. Evolving omics technology have enhanced our understanding of the mechanisms underlying trauma-induced coagulopathy, but significant work remains in the field of TBI specifically, despite suggestion that neurometabolites and proinflammatory proteins may play a significant role in the pathology of cerebral injury12,13 and interplay with coagulation.
The aim of this study is to better characterize coagulopathy of brain injury in trauma patients, specifically by evaluating hemostatic capacity before and after dura violation in the procedural setting with viscoelastic hemostatic assays, tissue factor activity, metabolomics, and proteomics14. Given the data describing tissue factor release after TBI, we hypothesize that dura violation is associated with higher circulating levels of tissue factor, conversion to a hypercoagulable profile, and a unique metabolomic and proteomic phenotype.
Methods
In order to examine the coagulopathy of TBI, we conducted a prospective, observational cohort study from 2019 to 2021 at an academic Level-1 trauma center. The study was approved by the local Institutional Review Board (IRB#17–2093) and performed under waiver of consent. This study was compliant with STROBE guidelines (checklist in Supplemental Digital Content, SDC 1).
Patient and sample collection
Inclusion criteria were patients admitted to the surgical trauma intensive care unit (ICU) with traumatic brain injuries (including subarachnoid, intraparenchymal or epidural bleeds) requiring neurosurgical intervention. At this institution, there is no dedicated neuro ICU and as such, all patients with ICU needs and TBI are admitted to the surgical ICU, where they are cared for collaboratively by surgical critical care intensivists and neurosurgery. Once a patient was admitted to the ICU with a TBI, whether isolated or as part of a polytrauma complex, a 24/7 professional research assistant pager was notified for enrollment at time of neurosurgical procedures. Patients were excluded if minors (< 18 years old) or pregnant. When a patient was in the operating room (after immediate transport from the emergency department or ICU), whole blood was drawn before dura violation and then one hour thereafter (given the half life of tissue factor is approximately one hour) in citrated vacuum tubes (35 mL, 3.2% sodium citrate).
Sample processing
Citrated rapid (CR-) thrombelastography (TEG) and tissue plasminogen activator (tPA-) challenged TEG (ThromboTherapeutics Inc., Walpole, MA) were performed with the TEG 5000 Thrombelastography Hemostasis Analyzer (Haemonetics, Niles, IL) according to manufacturer instructions. CR-TEG results in the following: activating clotting time (ACT, time elapsed from initiation of test until onset of clot formation in seconds), angle (rate of clot strength increase in degrees), maximum amplitude (MA; maximal clot strength achieved in millimeters), and percent clot lysis 30 minutes after reaching MA (LY30; lysis 30 minutes after MA in %). The tPA-Challenged TEG, which has been validated in previous studies and detects fibrinolysis with a greater sensitivity than citrated rapid TEG15–17, uses 75 ng/mL of exogenous tPA to assess degree of tPA-provoked fibrinolysis. The degree of fibrinolytic responsiveness is categorized as normal, “tPA-sensitive”, or “tPA-resistant,” the latter being linked to increased mortality in trauma patients17,18. The remaining blood was spun down to platelet-poor plasma (1000 g at 4oC for 15 minutes, then 12600 g at 4oC for six minutes) for storage in −80oC until time of omic analysis and tissue factor measurements.
Metabolomics analyses
For metabolomics, metabolites were extracted from plasma samples for mass spectrometric analysis using previously described methods19. Briefly, metabolites were extracted using a methanol, acetonitrile, and water (5:3:1) solution. Extracts were centrifuged to pellet insoluble fractions. Supernatants were transferred to autosampler vials and loaded into a Vanquish UHLPC coupled to a Q-Exactive mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) with a Kinetex C18 column (Phenomenex). Raw data was processed using Maven (Princeton University, Princeton, NJ, USA) equipped with the KEGG database.
Proteomics analyses
For proteomics, analyses were performed as previously described20. Briefly, plasma samples were digested in the S-Trap 96-well plate (Protifi, Huntington, NY) following the manufacturer’s procedure. The Evosep One system (Evosep, Odense, Denmark) was used to separate peptides on a Pepsep columnpacked with ReproSil C18 1.9 um, 120A resin, and the system was coupled to the timsTOF Pro mass spectrometer (Bruker Daltonics, Bremen, Germany) via the nano-electrospray ion source (Captive Spray, Bruker Daltonics). The mass spectrometer was operated in diaPASEF mode, and we used a project-specific library generated from 24 high-pH reverse-phase peptide fractions acquired with PASEF technology. For tissue factor analysis, plasma tissue factory activity (CY-QUANT MV-TF activity) was measured through ELISA (BioCytex) per manufacturer instruction.
Statistical analysis
All statistical analyses were performed using in R (version 4.1.0; 2021-05-18). Wilcoxon signed rank test was used for pairwise comparison (for pre- and post-dura violation) for the thrombelastography and tissue factor results. For the omic results, row-wise, log transformation normalization was performed with normalization to constant sums, and then again pairwise comparison was performed (for pre- and post-dura violation), in addition to multivariate analysis for the metabolomic data including PCA, PLS-DA, hierarchical clustering analysis as has previously been described21. Finally, in order to assess any correlation of the omic data to measurements of hemostatic capacity, proteome and metabolome wide correlations to TEG measurements were performed. Raw omics data was correlated using the Spearman method. Omics data that was mean centered and divided by the square root of the standard deviation for each analyte achieved a normal distribution. Pearson correlations were calculated for the normalized data. Top significant correlates were selected for linear regression and results that showed a significant p-value of the predicted slope were plotted. Statistical significance was considered at p < 0.05.
Results
Overall, 57 patients were included, 42 of whom underwent craniotomy, four underwent extraventricular drain placement, and 10 underwent intracranial bolt (Table 1). The majority (61%) were male, the median age was 52 years old, and 70% presented after blunt trauma. The median Glasgow Coma Score was 7 (3–14 interquartile range [IQR]). One patient was taking warfarin for atrial fibrillation, and two patients were taking clopidogrel. The majority of patients did not present in shock, with only 7% (4) presenting with systolic blood pressure < 90 mm Hg. None of the patients required massive transfusion, and while 9% (5) received packed red blood cells (PRBC) and 7% (4) received fresh frozen plasma (FFP) in the first hour, none received platelets, cryoprecipitate, or tranexamic acid. Three patients (5%) developed deep venous thrombosis. The mortality rate was 24% (14).
Table 1. Demographic and clinical characteristics of study population.
Presented as medians and interquartile ranges or % (n) as appropriate.
| Demographics | |
| Sex, % male (n) | 68% (39) |
| Age (years) | 52 (34–61) |
| BMI | 26 (23–30) |
| Injury Characteristics | |
| Blunt, % (n) | 70% (40) |
| GCS | 7 (3–14) |
| Physiologic Characteristics | |
| SBP (mm Hg) | 124 (111–148) |
| Base deficit (meq/L) | 5.7 (3.2–9.6) |
| Lactate (mmol/L) | 2.7 (2.2–4.9) |
| Hematology/coagulation assays | |
| Hemoglobin (g/dL) | 14.2 (12.8–15.0) |
| Hematocrit (%) | 42.7 (38.6–45.0) |
| Platelets (10^9/L) | 250 (214–302) |
| INR | 1.12 (1.05–1.27) |
| PTT (sec) | 27.1 (24.8–31.6) |
| Clinical Outcomes | |
| Mortality, % (n) | 24% (14) |
| >/= 1U PRBC in 1st hour | 9% (5) |
| >/= 1U FFP in 1st hour | 7% (4) |
| Ventilator days | 3 (0–8) |
| ICU LOS (days) | 6 (3–14) |
| Hospital LOS (days) | 11 (5–26) |
BMI=Body Mass Index, GCS=Glasgow Coma Score, SBP=systolic blood pressure, INR=International Normalized Ratio, PTT=partial thromboplastin time, RBC=red blood cell, ICU = intensive care unit, LOS=length of stay
Thrombelastography
Compared to pre-dura violation, post-dura violation blood samples demonstrated a systemic hypercoagulability characterized by increased clot strength and decreased fibrinolysis (SDC 2). After dura violation, there was a significant increase in clot strength (median maximum amplitude of 74.4 mm [70.2–77.7 interquartile range, IQR] versus 63.5 mm [58.5–68.8 IQR], p<0.0001). Similarly, after dura violation, there was a significant decrease in fibrinolysis, specifically sensitivity to tPA (LY30 on tPA-challenge TEG of 1.4% (0.4–3.4 IQR] versus 2.6% [0.7–7.3 IQR], p = 0.04), with conversion to tPA resistant phenotype (LY30 < 1.8%) in the post-dura violation group. There were no differences in time to clot formation or rate of clot propagation (SDC 2).
Metabolomics
Metabolomic analyses were performed on samples from pre- and post-dura violation (Figure 1A). In total, a total of 131 metabolites were identified (SDC 3) with significant changes in the metabolomic profile pre- and post-dura violation (SDC 2, Figure 1). Key increases in serine biosynthesis and carnitine and fatty acid metabolism and decreases in glycolysis, TCA cycle, GSH homeostasis, urea cycle, sulfur metabolism, indole and tryptophane, and glycerophospholipid biosynthesis were observed.
Figure 1.
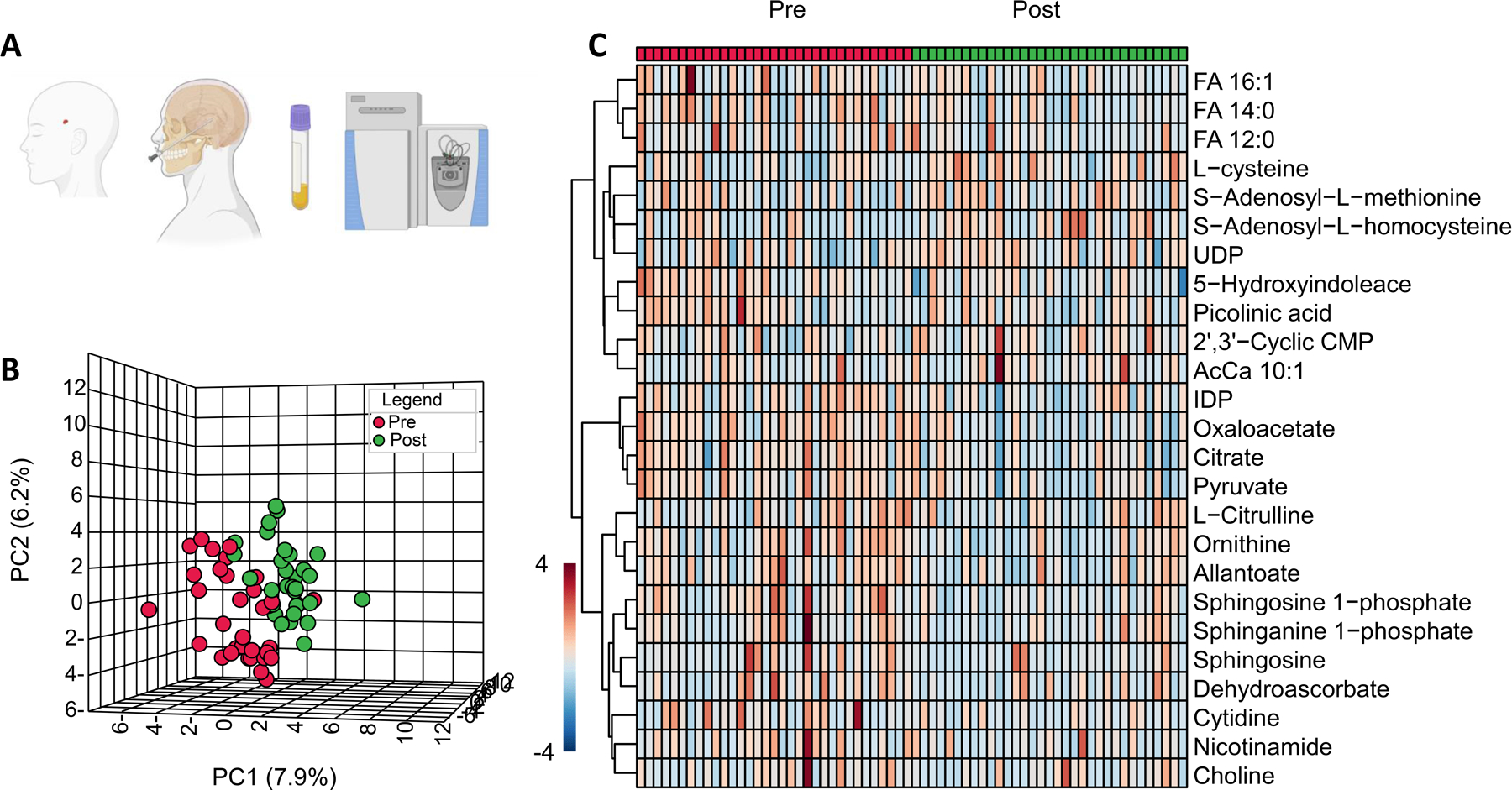
Depiction of samples drawn pre- and post-dura violation (A), partial least square discriminant analysis (B), and hierarchical clustering map of the top 25 significant changes (C).
Compared to pre-dura violation (SDC 2), there were significant increases in amino acid L-cysteine (amino acid), nucleotide 5–6-dihydrothymine, serine biosynthesis metabolite 5, adenosyl-L-homocysteine, arginine and proline metabolite tissue creatinine, and carnitine and fatty acid metabolite 0-decenoyl-L-carnitine. In contrast, there were also significant decreases in a myriad of metabolites, including several amino acids (L-histidine, L-lysine, L-tyrosine), nucleotides (IDP, allantoate, nicotinamide, glycolytic metabolite pyruvate, TCA cycle metabolite oxaloacetate, GSH metabolite dehydroascorbate, serine biosynthesis metabolite 5-adenosyl-L-methionine, urea cycle metabolites (ornithine, L-citrulline), arginine and proline metabolites (4-acetamidobutanoate, guanidinoacetate), sulfur metabolite 3-sulfocatechol, indole and tryptophan metabolite (5-hydroxyindoleacetate, g-oxalocrotonate), glycerophospholipid biosynthesis metabolites (sphingosine, sphingosine-1-phosphate, acetylcholine, sphinganine-1-phosphate), and essential fatty acid metabolites (dodecanoic acid, tetradecanoic acid).
To make better sense of the metabolomic data, partial least square discriminant analysis was performed to separate pre- vs post-dura violation samples (Figure 1B). Results informed the hierarchical clustering map of the top 25 significant changes in Figure 1C. Specifically, top significant metabolites are noted upon normalization of post-dura violation data to baseline values in the volcano plot in Figure 2A. Most significant changes were noted for metabolites involved in late glycolysis/entrance to the Krebs cycle in mitochondria (pyruvate, oxaloacetate, citrate), cysteine and one carbon metabolites (S-adenosyl-methionine – SAM and S-adenosyl-homocysteine – SAH), and metabolites involved in endothelial dysfunction/arginine metabolism/responses to hypoxia (ornithine, sphingosine 1-phosphate – Figure 2B).
Figure 2.
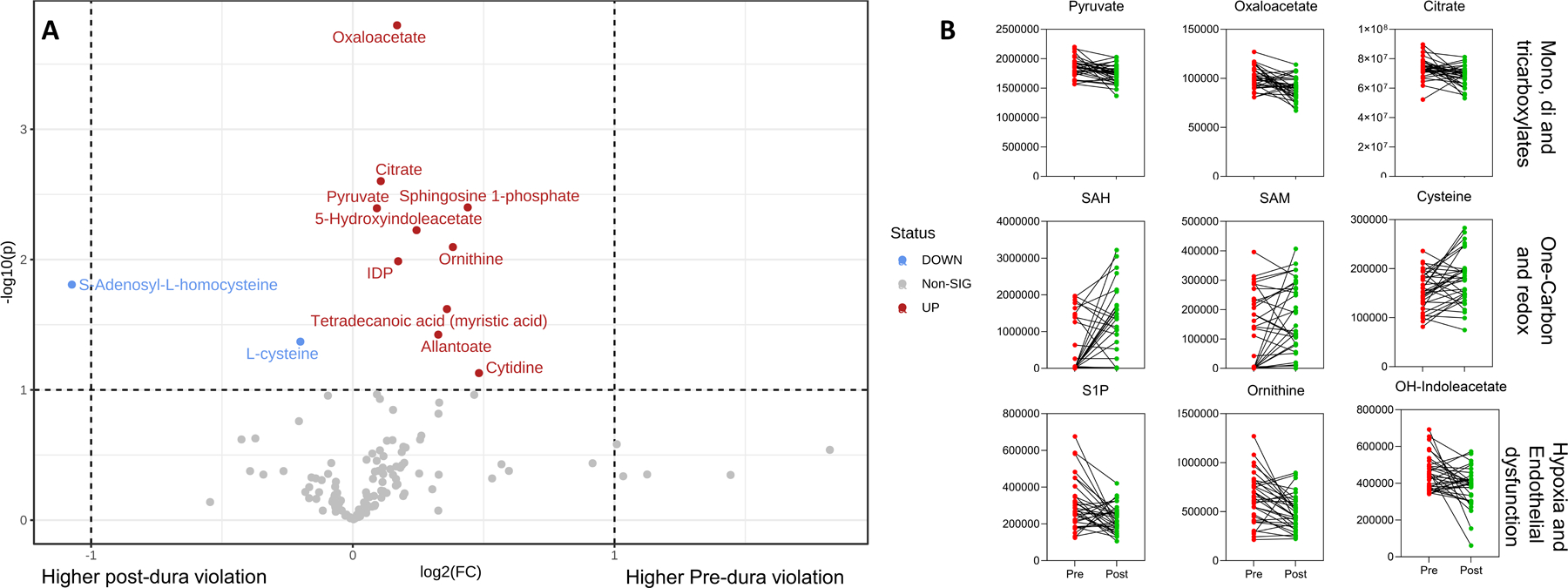
Top significant metabolites noted upon normalization of post-dura violation data to baseline values in volcano plot (A), with selected most significant changes (B).
On correlation analysis with the thrombelastographic evidence, several metabolites significantly positively correlated with maximum amplitude (Figure 3), specifically several fatty acids (12:0, p=0.63 and 22:6, p=0.61, p<0.001 for both).
Figure 3. Metabolome wide correlates to TEG MA, LY30, R parameters.
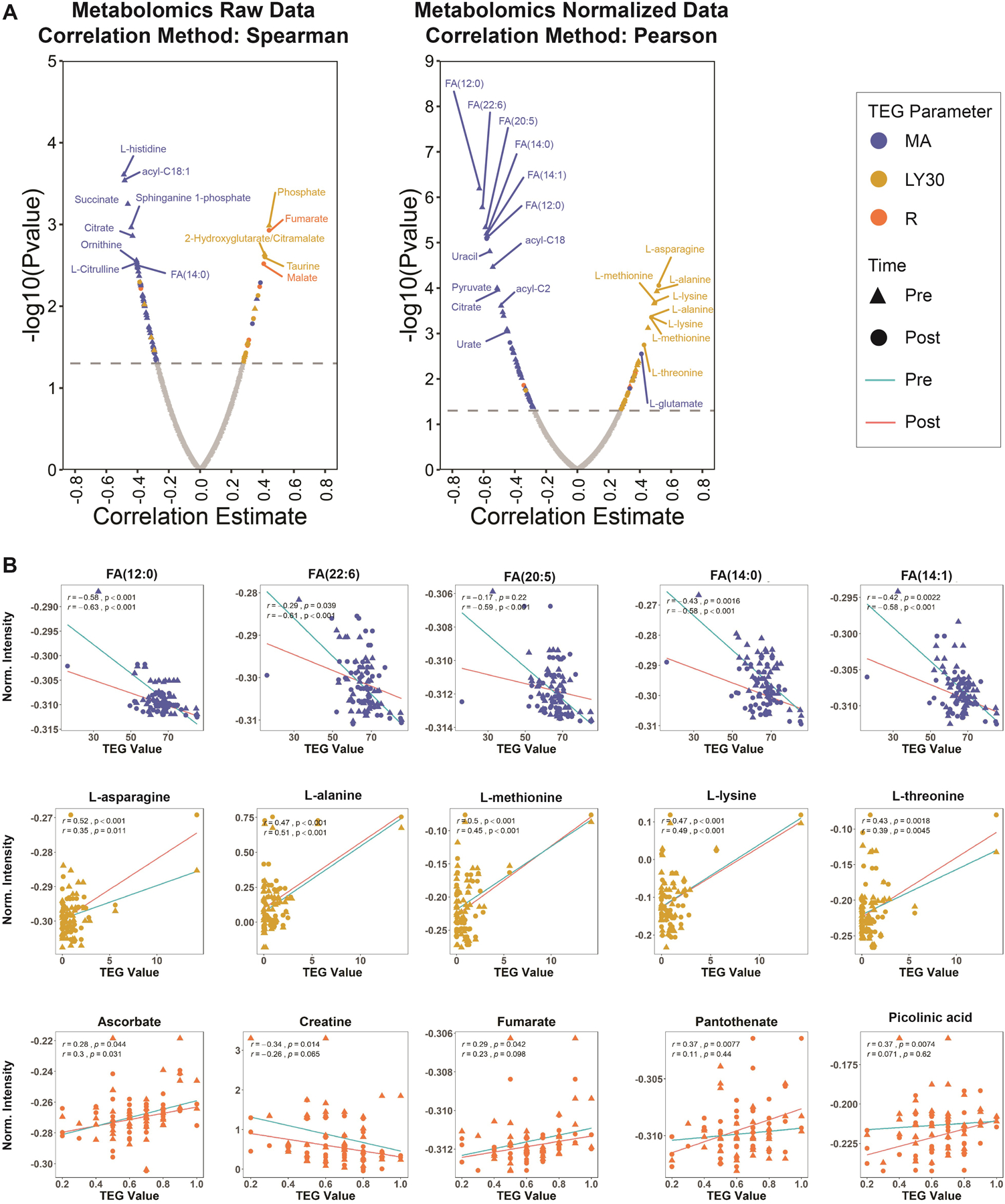
A) Raw metabolomics data was correlated with separate thrombelastography (TEG) parameters independently for ‘pre’ and ‘post’ time points using the Spearman method. Significant correlates (p-value < 0.05) are colored while insignificant hits remain grey. Coloring of significant hits were dictated by the respective TEG parameter as described in the legend. Blue corresponded to maximum amplitude (MA), gold corresponded to LY30 (fibrinolysis 30 minutes after MA), and orange corresponded to R (reaction time, time to clot formation). The correlation results from each time point were plotted together and differentiated by shape. Such that, ‘pre’ was represented with a triangle and ‘post’ was represented with a circle. B) Top correlates were selected for further analysis by linear regression. The line of best fit was colored by time point, teal for ‘pre’ and red for ‘post’.
Proteomics
To expand on the metabolomics analyses and provide additional mechanistic insights on the TBI-associated coagulopathy, proteomic analyses were performed on samples from pre- and post-dura violation. In total, a total of 233 proteins were identified (SDC 4) which demonstrated significant changes in the proteomic profile pre- and post-dura violation, with key changes in a variety of proteins related to platelet degranulation and activation, intracellular calcium signaling, and fibrinolysis (SDC 2). Compared to pre-dura violation, there were significant increases in proteins responsible for platelet activation (including retinoic acid receptor responder protein 2, amyloid A1 metalloproteinase inhibitor-1, platelet glycoprotein Ib and V, clusterin, protease C1 inhibitor, CD109, alpha-1-antichymotryspin, inter-alpha-trypsin inhibitor heavy chain H3 and H4, kininogen-1, sulfhydryl oxidase 1, complement factor D, coagulation factor VII and X, serotransferrin, pro low-density lipoprotein receptor-related protein 1, fibrinogen alpha and beta chain) and fibrinolysis inhibition (protease C1 inhibitor, alpha-2-antiplasmin) and decreases in proteins responsible for fibrinolysis promotion (kallikrein, tetranectin, plasminogen). However, there were also significant decreases (whether due to consumption or decreases in release into circulation) in platelet activating and antifibrinolytic proteins including selenoprotein P, heat shock protein beta-1, hyaluronidase-1, thrombospandin-1, superoxide dismutase, coagulation factor V, XI, and XIII, platelet factor 4, platelet-activating factor acetylhydrolase, beta-2- and alpha-1B-glycoprotein, multimerin-1, platelet basic protein, fermitin, galectin-3-binding protein, alpha-1-acid-glycoprotein-1, carboxypeptidase B2, and vitronectin. While there were significant increases in several procoagulant, antifibrinolytic proteins, there were also increases in some proteins with an anticoagulant effect (such as antithrombin and tissue factor pathway inhibitor), highlighting the complexity of these protein profiles and the importance of interpretation of net effect.
On correlation analysis with thrombelastography, several proteins significantly positively correlated with maximum amplitude (Figure 4), specifically apolipoprotein C-1 (r=0.63), platelet activating factor acetylhydrolase 1b subunit (p=0.66), fibrinogen beta chain (r=0.63), gamma chain (r=0.60) and alpha chain (r=0.62), matrix metallopeptidase 9 (r=0.60), complement component C9 (r=0.66), and fatty acid-binding protein 2 (r=0.68) (p<0.001 for all).
Figure 4. Proteome wide correlates to TEG MA, LY30, R parameters.
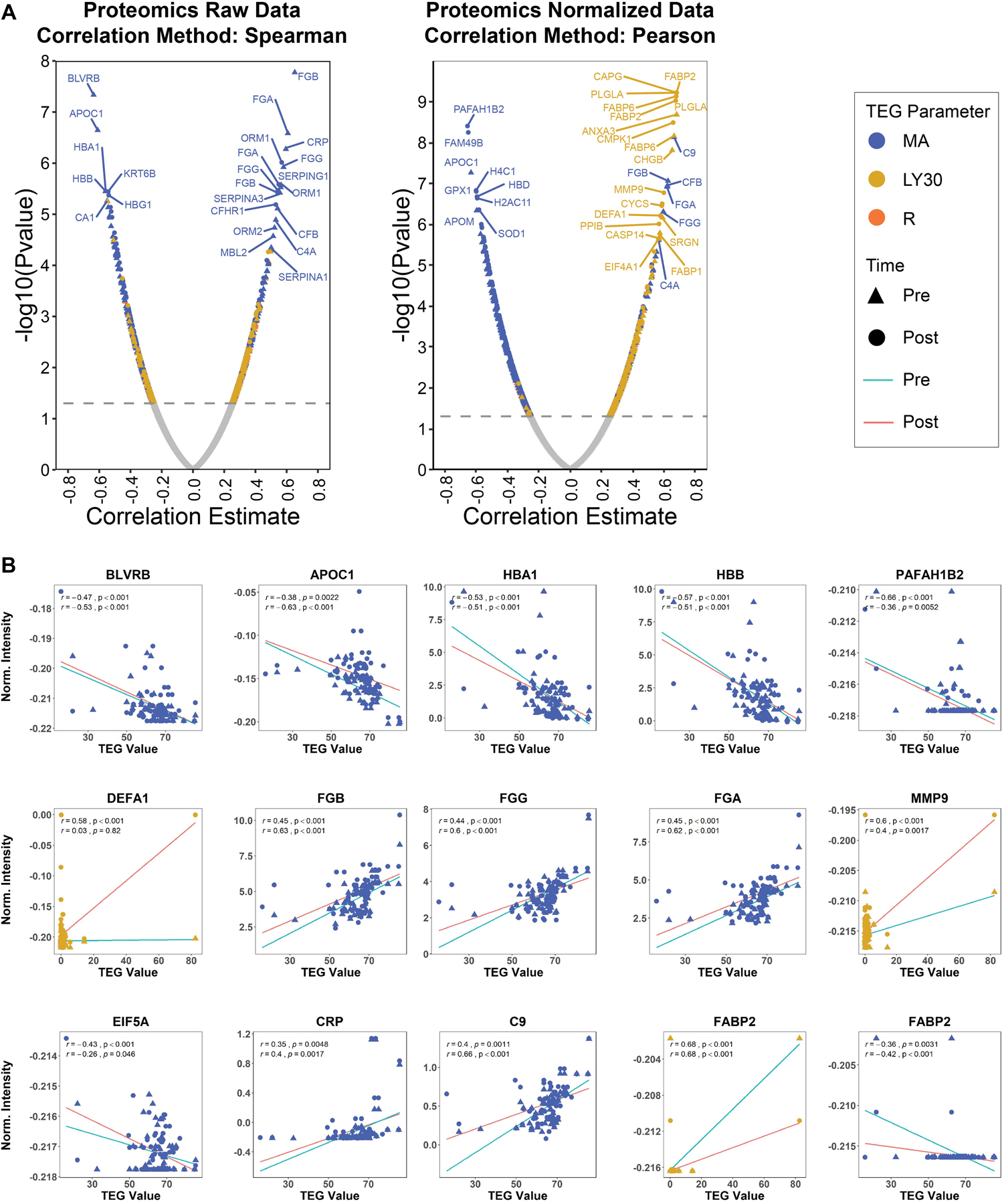
A) Raw proteomics data was correlated with separate TEG parameters independently for ‘pre’ and ‘post’ time points using the Spearman method. Significant correlates (p-value < 0.05) are colored while insignificant hits remain grey. Coloring of significant hits were dictated by the respective thrombelastography (TEG) parameter as described in the legend. Blue corresponded to maximum amplitude (MA), gold corresponded to LY30 (fibrinolysis 30 minutes after MA), and orange corresponded to R (reaction time, time to clot formation). The correlation results from each time point were plotted together and differentiated by shape. Such that, ‘pre’ was represented with a triangle and ‘post’ was represented with a circle. B) Top correlates were selected for further analysis by linear regression. The line of best fit was colored by time point, teal for ‘pre’ and red for ‘post’.
Tissue factor activity
There were no statistically significant differences in tissue factor activity pre-dura violation versus post-dura violation (median 5.1 fM [1.5–30.6 IQR) versus 5.2 fM [2.7–18.0], p=0.98).
Discussion
After dura violation, a systemic hypercoagulability is observed in TBI patients, characterized by increased clot strength, decreased fibrinolysis, and increased resistance to tPA. Additionally, TBI and dura violation causes a unique metabolomic and proteomic phenotype. Systemic metabolism is affected by substrate-specific metabolic derangements, with increases in serine biosynthesis and carnitine and fatty acid metabolism and decreases in glycolysis, TCA cycle, GSH homeostasis, urea cycle, sulfur metabolism, indole and tryptophane, and glycerophospholipid biosynthesis. Further, the proteomic profile shifts to one with significant representation of proteins which promote platelet activation and diminish fibrinolysis. These robust omic changes are not accompanied by changes in circulating tissue factor. These findings highlight the importance of consideration of tissue specific versus systemic hemostatic profiles, in which we describe patients with intracranial hemorrhage yet who develop a robust systemic hypercoagulability. This work adds to a growing body of literature developing deeper understanding of the coagulopathy and omics of TBI, challenging the paradigm that tissue factor drives TBI-related hypercoagulability.
In this prospective study, TBI after dura violation was associated with a conversion to a hypercoagulable state with increased clot strength. This contributes to previous works on the coagulopathy of TBI, which has been riddled with contradictory reports, highlighting clinical equipoise in understanding the hemostatic capacity derangements after this injury. While some reports have described hypocoagulopathy in TBI, other reports have found a hypercoagulability associated with head injury2–4. It is worth mentioning that when hypocoagulopathic phenotypes of trauma-induced coagulopathy (TIC) in TBI are observed, it is often in patients taking anticoagulant or antiplatelet medications pre-injury22. Moreover, TBI patients with a described hypocoagulopathy often present with a mixed picture of TBI with hemorrhagic shock or specific polytrauma injury complexes23,24. Our work describes a hypercoagulable coagulopathy of TBI, which adds to the ongoing mixed phenotypes described. Further, as is well known with TIC, it is a dynamic phenotype which changes over time; the reports of coagulopathy related to TBI likely have conflicting findings because TBI-related coagulopathy is also dynamic, with an accelerated onset as early as 12 hours after injury and as delayed as five days after injury25. This time course has been classically described as an early picture of hypocoagulopathy and hyperfibrinolysis, which drives early intracranial hemorrhage progression, with subsequent conversion to a hypercoagulable, antifibrinolytic profile26. The results of our work, however, highlight that derangement to hypercoagulability may occur within hours of injury and/or neurosurgical procedures, driven not by tissue factor release as previously thought and described in the DIC literature27.
Extensive literature examining the mechanism of TBI-related hypercoagulability suggests that inflammation plays a crucial role and may be mediated by activation of complement systems, endothelial shedding, protein C activation, von Willebrand factor (vWF) activity, brain-derived extracellular vesicles and extracellular vesicles from endothelial cells and platelets26; these pro-inflammatory and procoagulant proteins may be more pronounced after dura violation and neurosurgical procedures, leading to aberrant platelet activity. Our findings of significant increases in clot strength on thrombelastography after dura violation highlight the role of platelets in promoting the pathologic hypercoagulability of TBI, which has been suggested in previous works8. This elevation of maximum amplitude is similar in the numbers reports in previous works of trauma patients that has been linked to increased risk of thrombotic complications (MA 71.9 mm in patients with thromboses versus 68.5 mm in patients without thromboses, compared to 74.4 mm post-dura violation versus 63.5 mm pre-dura violation in our patients)28, suggesting these changes observed in MA in our data have clinical significance. The increased platelet-mediated clot strength in our patients may be an important player in mitigating the locally hypocoagulopathic environment of hemorrhagic cerebrum, as evidenced by prior reports of higher maximum clot firmness (another viscoelastic metric of clot strength) in TBI survivors versus non-survivors29 and poorer outcomes in TBI patients with thrombocytopenia30.
Beyond the cellular contributions to the pathologic hypercoagulability of TBI, our data highlights an important transition to fibrinolytic shutdown and tPA resistance. There has been controversy in the literature as to whether TBI drives a hyperfibrinolytic or antifibrinolytic phenotype31,32. In a prospective study of severely injured patients, Samuels et al delineate differential coagulation phenotypes based on injury complexes24; the authors ultimately found no evidence that TBI contributes to hyperfibrinolysis, suggesting that fibrinolytic shutdown may be more prevalent in patients with head injury, as found after dura violation in our population. Additional work by Cohen et al also did not find hyperfibrinolysis in TBI patients without concomitant hypoperfusion and shock33. In another prospective study of over 1,000 TBI patients, Meizoso et al found that fibrinolysis shutdown was more common in TBI patients, occurring in up to one fourth of TBI patients, and when controlling for sex, injury mechanism, injury severity and shock, TBI independently decreased lysis at 30 minutes34. As described in our work, this TBI-related fibrinolytic shutdown occurs as early as one hour after injury, perhaps as a reactive, compensatory mechanism. This association between fibrinolytic shutdown and TBI may ultimately be driven by tPA resistance and elevated PAI-1 levels35. The process of an initial thrombin and fibrin burst following by conversion to fibrinolytic shutdown in TBI is further suggested by a gradual decrease in fibrinogen and D-dimer over the first three hours following brain injury36, a distinct process from the tissue factor-drive DIC.
Previous metabolomic analyses of injured patients or animal models of traumatic injury have focused on the phenotypes provoked by hemorrhagic shock versus tissue injury37, but a paucity of data has explored TBI, in particular in tandem with coagulation data. This literature gap is important, as multi-omics data extends relevance into the care of trauma patients and precision medicine; previous descriptions of omics highlight its importance for trauma surgeons in identifying diagnostic and therapeutic markers to assess response to interventions and inform clinically-decision making based on dynamic cellular and subcellular level processes38. Our data describes increases in serine biosynthesis and fatty acid metabolism and consumption of critical metabolites in the glycolysis and TCA pathway, amongst others. This consumptive picture of the TCA cycle provides a compelling mechanistic explanation for why neuron-specific enolase, which converts 2-phosphoglycerate to phosphoenolpyruvate in glycolysis, serves as a marker of TBI which may be seen concomitantly with dura violation39. A drastic increase in homocysteine was observed after dura violation in the setting of TBI, which bears clinical significance for several reasons. Homocysteine catabolism can be dysregulated by injury and stress, leading to neurotoxic effects. Tchantchou et al found that hyperhomocysteinemic rats had increased oxidative stress, upregulated expression of hypercoagulable proteins, exacerbated blood-brain barrier dysfunction, infiltration of inflammatory cells into the cortex, and aggravated anxiety-related behaviors40. Not only is homocysteine tied to neurotoxicity, but is also associated with hypercoagulability via reduced antithrombin, endotheliopathy propagation, platelet dysfunction and hyperactivity, and diminished plasmin activity and fibrinolysis41.
As has been previously described in injury-associated hemorrhagic shock models42, TBI promoted lipolysis and beta-oxidation of lipids, as suggested by an increase in fatty acid- acyl-carnitine O-decenoyl-L-carnitine (acyl-C10:1) and decreases in essential fatty acids dodecanoic acid (lauric acid) and tetradecanoic acid (myristic acid). Fatty acids have been established as important potential metabolite markers of severity of TBI, with medium-chain fatty acids, octanoic and decanoic acids, being highly associated with severity of injury12. Not only do these changes in fatty acids have implications in neurotrauma, but fatty acids are also associated with prothrombotic effects43.
In addition to increased lipolysis metabolites, our data revealed decreases in citrulline, arginine, and proline metabolites, which has been described in previous TBI metabolomic investigations44. These same changes have been observed in critically injured patients and those in hemorrhagic shock37 and linked to adverse clinical outcomes45. These reductions in arginine ultimately lead to reduction in nitric oxide necessary to maintain cerebral perfusion. Further, ornithine, a crucial part of the arginine and nitric oxide pathway, was significantly decreased in TBI patients after dura violation, as seen in previous TBI literature46. Ornithine is a substrate for ornithine decarboxylase for polyamine synthesis, with polyamines serving as markers of TBI and mediating fibrinolysis47. Beyond fibrinolysis, ornithine has been described as a marker of endotheliopathy, competing with arginine as a substrate with nitric oxide synthase; the end result in the setting of TBI is that ornithine induces oxidative stress and decreases blood-brain barrier integrity48. Creatinine, another important metabolite in the arginine and proline pathway, significantly increased after dura violation in our population; in a previous study aimed to identify predictors of coagulopathy in TBI and develop machine learning (ML) models to predict the risk of coagulopathy, Yang et al found that creatinine was a core component in contributing to prediction of coagulopathy in TBI patients1.
To complement the metabolomic data which characterizes a shift towards platelet hyperactivity and fibrinolysis shutdown, our proteomic data demonstrated key increases in proteins related to platelet activation, degranulation, and aggregation which correlated to thrombelastographic measurements of platelet activity. Examining the mechanisms and natural history of TBI has been recently empowered by proteomic data, which has further characterized the neuroinflammatory response to TBI49. Our data describes significant changes in several proteins related to platelet activation and activation with positive correlations to measurements of clot strength, including apolipoproteins, which are also known to be involved in the neuroinflammatory response to brain injury13. Other proteomic work describes significantly increased levels of proteins related to neutrophil degranulation and mobilization, which in sum with our work highlight a picture of pan-cellular hyperactivity and recruitment49. The proteomic data additionally showed key increases in antiplasmin and fibrinogen, which correlated to hypercoagulability on thrombelastography. Fibrinogen is known to increase immediately after TBI before a precipitous decline and is believed to participate in destruction of the blood-brain barrier and neuroinflammation36. The release of antiplasmin as a result of TBI may be a compensatory mechanism to mitigate intracerebral hemorrhage progression, as previous proteomic work has identified an inversed relationship between antiplasmin and progressive hemorrhagic injury50.
Limitations to this study include a reliance on steady-state measurements for the omic observations; therefore, conclusions on aberrant metabolic fluxes through glycolysis, Krebs cycle, purine oxidation, and fatty acid/amino acid oxidation, in addition to protein cascades, will require further validation with flux analysis studies based on incubation with stable isotope-label tracers. Another limitation is the lack of control patients, specifically TBI patients who do not undergo neurosurgical procedures, however this study focused on our specific cohort as we are most interested in patients who had dura violation, which we believe this is part of the mechanism driving TBI-related systemic hypercoagulability. Lastly, this TBI cohort is heterogeneous in the neurosurgical intervention (a minority underwent EVD or bolt instead of craniotomy), which may affect our interpretation of how dura violation impacts coagulation.
In conclusion, in this prospective study of TBI patients undergoing neurosurgical procedures, there is a robust conversion to a hypercoagulable phenotype, accompanied with a drastic shift in the metabolomic and proteomic profile. These changes happen early and rapidly after injury and procedures, independent of tissue factor, and highlight cause for caution in resuscitation of TBI patients with procoagulant agents. Further, these data demonstrate that while a patient may be locally hypocoagulable in the setting of a traumatic brain hemorrhage, this does not necessarily signify a systemic coagulation concordance; thus, while TBI patients may have an organ-specific hypocoagulable, hyperfibrinolytic profile, systemic hypercoagulability and fibrinolytic shutdown predominate. As such, these data contextualize why early venous thromboembolism prophylaxis is reasonable early in many TBI patients. Ultimately, these data highlight the systemic hypercoagulability of TBI after dura violation, with procoagulant metabolites leading to increased clot strength, decreased clot breakdown, and tPA resistance which represents a unique and previously undescribed phenotype after trauma unrelated to tissue factor as once was previously thought. Further, these data highlight the potential role for omics as a tool to identify early metrics of pathologic hypercoagulability in TBI and guide precision medicine to mitigate thrombotic morbidity. Additional work is merited to further delineate the relationship between these observations.
Supplementary Material
Supplemental Digital Content 1. STROBE guidelines.
Supplemental Digital Content 2. Thrombelastography, metabolomics, and proteomics pre- and post-dura violation. Presented as medians and interquartile ranges.
Supplemental Digital Content 3. Changes in metabolomic profiles pre- and post-dura violation. Presented as medians and interquartile ranges.
Supplemental Digital Content 4. Changes in proteomic profiles pre- and post-dura violation. Presented as medians and interquartile ranges.
Sources of support:
Research reported in this publication was supported by the National Institute of General Medical Sciences of the National Institutes of Health (T32 GM008315 and P50 GM049222). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health or other sponsors of the project.
Footnotes
Meeting Presentation: This study was presented at the 36th annual meeting of the Eastern Association for the Surgery of Trauma, January 17-21, 2023, in Lake Buena Vista, FL.
References
- 1.Yang F, Peng C, Peng L, Wang J, Li Y, Li W. A Machine Learning Approach for the Prediction of Traumatic Brain Injury Induced Coagulopathy. Frontiers in Medicine. 2021;8:792689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hubbard WB, Dong JF, Cruz MA, Rumbaut RE. Links between thrombosis and inflammation in traumatic brain injury. Thrombosis Research. 2021;198:62–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Di Battista AP, Rizoli SB, Lejnieks B, Min A, Shiu MY, Peng HT, et al. Sympathoadrenal Activation is Associated with Acute Traumatic Coagulopathy and Endotheliopathy in Isolated Brain Injury. Shock. 2016;46(3 Suppl 1):96–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lee TH, Hampton DA, Diggs BS, McCully SP, Kutcher M, Redick BJ, et al. Traumatic brain injury is not associated with coagulopathy out of proportion to injury in other body regions. The Journal of Trauma and Acute Care Surgery. 2014;77(1):67–72. [DOI] [PubMed] [Google Scholar]
- 5.Abdelmalik PA, Boorman DW, Tracy J, Jallo J, Rincon F. Acute Traumatic Coagulopathy Accompanying Isolated Traumatic Brain Injury is Associated with Worse Long-Term Functional and Cognitive Outcomes. Neurocritical Care. 2016;24(3):361–70. [DOI] [PubMed] [Google Scholar]
- 6.Yasui H, Donahue DL, Walsh M, Castellino FJ, Ploplis VA. Early coagulation events induce acute lung injury in a rat model of blunt traumatic brain injury. Am J Physiol Lung Cell Mol Physiol. 2016;311(1):L74–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Labbadia R, Diomedi Camassei F, Antonucci L, Guzzo I, Muda OA, et al. Reversible glomerular damage in disseminated intravascular coagulation. Pediatr Transplant. 2022;26(1):e14147. [DOI] [PubMed] [Google Scholar]
- 8.Fletcher JJ, Kade AM, Sheehan KM, Wilson TJ. A case-cohort study with propensity score matching to evaluate the effects of mannitol on venous thromboembolism. Journal of Clinical Neuroscience. 2014;21(8):1323–8. [DOI] [PubMed] [Google Scholar]
- 9.Halpern CH, Reilly PM, Turtz AR, Stein SC. Traumatic coagulopathy: the effect of brain injury. Journal of Neurotrauma. 2008;25(8):997–1001. [DOI] [PubMed] [Google Scholar]
- 10.Hubbard WB, Sim MMS, Saatman KE, Sullivan PG, Wood JP. Tissue factor release following traumatic brain injury drives thrombin generation. Res Pract Thromb Haemost. 2022;6(4):e12734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Suehiro E, Fujiyama Y, Kiyohira M, Motoki Y, Nojima J, Suzuki M. Probability of Soluble Tissue Factor Release Lead to the Elevation of D-dimer as a Biomarker for Traumatic Brain Injury. Neurologia Medico-Chirurgica. 2019;59(2):63–67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Posti JP, Dickens AM, Orešič M, Hyötyläinen T, Tenovuo O. Metabolomics Profiling As a Diagnostic Tool in Severe Traumatic Brain Injury. Front Neurol. 2017;8:398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lindblad C, Pin E, Just D, Nimer FA, Nilsson P, Bellander B, et al. Fluid proteomics of CSF and serum reveal important neuroinflammatory proteins in blood-brain barrier disruption and outcome prediction following severe traumatic brain injury: a prospective, observational study. Critical Care. 2021;25(1):103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Huibregtse ME, Bazarian JJ, Shultz SR, Kawata K. The biological significance and clinical utility of emerging blood biomarkers for traumatic brain injury. Neurosci Biobehav Rev. 2021;130:433–447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Genét GF, Ostrowski SR, Sørensen AM, Johansson PI. Detection of tPA-induced hyperfibrinolysis in whole blood by RapidTEG, KaolinTEG, and functional fibrinogen TEG in healthy individuals. Clinical and Applied Thrombosis/Hemostasis. 2012;18(6):638–44. [DOI] [PubMed] [Google Scholar]
- 16.Kupesiz A, Rajpurkar M, Warrier I, Hollon W, Tosun O, Lusher J, et al. Tissue plasminogen activator induced fibrinolysis: standardization of method using thromboelastography. Blood Coagulation & Fibrinolysis. 2010;21(4):320–4. [DOI] [PubMed] [Google Scholar]
- 17.Moore HB, Moore EE, Chapman MP, Huebner BR, Einersen PM, Oushy S, et al. Viscoelastic Tissue Plasminogen Activator Challenge Predicts Massive Transfusion in 15 Minutes. Journal of the American College of Surgeons. 2017;225(1):138–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Moore HB, Moore EE, Huebner BR, Dzieciatkowska M, Stettler GR, Nunns GR, et al. Fibrinolysis shutdown is associated with a fivefold increase in mortality in trauma patients lacking hypersensitivity to tissue plasminogen activator. The Journal of Trauma and Acute Care Surgery. 2017;83(6):1014–1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nemkov T, Reisz JA, Gehrke S, Hansen KC, D’Alessandro A. High-Throughput Metabolomics: Isocratic and Gradient Mass Spectrometry-Based Methods. Methods in Molecular Biology. 2019;1978:13–26. [DOI] [PubMed] [Google Scholar]
- 20.LaCroix IS, Cohen M, Moore EE, Dzieciatkowska M, Nemkob T, Schaid TR, et al. Omics Markers of Red Blood Cell Transfusion in Trauma. Int J Mol Sci. 2022;23(22):13815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.D’Alessandro A, Nemkov T, Moore HB, Moore EE, Wither M, Nydam T, et al. Metabolomics of trauma-associated death: shared and fluid-specific features of human plasma vs lymph. Blood Transfus. 2016;14(2):185–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Böhm JK, Güting H, Thorn S, Schäfer N, Rambach V, Schӧchl H, et al. Global Characterisation of Coagulopathy in Isolated Traumatic Brain Injury (iTBI): A CENTER-TBI Analysis. Neurocritical Care. 2021;35(1):184–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Martin G, Shah D, Elson N, Boudreau R, Hanseman D, Pritts TA, et al. Relationship of Coagulopathy and Platelet Dysfunction to Transfusion Needs After Traumatic Brain Injury. Neurocritical Care. 2018;28(3):330–337. [DOI] [PubMed] [Google Scholar]
- 24.Samuels JM, Moore EE, Silliman CC, Banerjee A, Cohen MJ, Ghasabyan A, et al. Severe traumatic brain injury is associated with a unique coagulopathy phenotype. The Journal of Trauma and Acute Care Surgery. 2019;86(4):686–693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lustenberger T, Talving P, Kobayashi L, Inaba K, Lam L, Plurad D, et al. Time course of coagulopathy in isolated severe traumatic brain injury. Injury. 2010;41(9):924–8. [DOI] [PubMed] [Google Scholar]
- 26.Fletcher-Sandersjöö A, Thelin EP, Maegele M, Svensson M, Bellander BM. Time Course of Hemostatic Disruptions After Traumatic Brain Injury: A Systematic Review of the Literature. Neurocritical Care. 2021;34(2):635–656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wada T, Shiraishi A, Gando S, Yamakawa K, Fujishima S, Saitoh D, et al. Disseminated intravascular coagulation immediately after trauma predicts a poor prognosis in severely injured patients. Scientific Reports. 2021;11(1):11031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Coleman JRKA, Moore EE, Moore HB, Gonzalez E, Majercik S, Cohen MJ, White T, Pieracci FM. It’s sooner than you think: blunt solid organ injury patients are already hypercoagulable upon hospital admission - results of a bi-institutional, prospective study. American Journal of Surgery. 2019;218(6):1065–1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schöchl H, Solomon C, Traintinger S, Nienaber U, Tacacs-Tolnai A, Windhofer C, et al. Thromboelastometric (ROTEM) findings in patients suffering from isolated severe traumatic brain injury. Journal of Neurotrauma. 2011;28(10):2033–41. [DOI] [PubMed] [Google Scholar]
- 30.Lillemäe K, Luostarinen T, Reinikainen M, Bendel S, Laitio R, Hoppu S, et al. Early thrombocytopenia is associated with an increased risk of mortality in patients with traumatic brain injury treated in the intensive care unit: a Finnish Intensive Care Consortium study. Acta Neurochir. 2022;164(10):2731–2740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fair KA, Farrell DH, McCully BH, Rick EA, Dewey EN, Hilliard C, et al. Fibrinolytic Activation in Patients with Progressive Intracranial Hemorrhage after Traumatic Brain Injury. Journal of Neurotrauma. 2021;38(8):960–966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hayakawa M, Maekawa K, Kushimoto S, Kato H, Sasaki J, Ogura H, et al. Hyperfibrinolysis in severe isolated traumatic brain injury may occur without tissue hypoperfusion: a retrospective observational multicentre study. Critical Care. 2017;21(1):222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cohen MJ, Brohi K, Ganter MT, Manley GT, Mackersie RC, Pittet JF. Early coagulopathy after traumatic brain injury: the role of hypoperfusion and the protein C pathway. The Journal of Trauma. 2007;63(6):1254–61. [DOI] [PubMed] [Google Scholar]
- 34.Meizoso JP, Dudaryk R, Mulder MB, Ray JJ, Karcutskie CA, Eidelson SA, et al. Increased Risk Of Fibrinolysis Shutdown Among Severely Injured Trauma Patients Receiving Tranexamic Acid. The Journal of Trauma and Acute Care Surgery. 2018;84(3):426–432. [DOI] [PubMed] [Google Scholar]
- 35.Nakae R, Murai Y, Wada T, Fujiki Y, Kanaya T, Takayama Y, et al. Hyperfibrinolysis and fibrinolysis shutdown in patients with traumatic brain injury. Scientific Reports. 2022;12(1):19107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nakae R, Murai Y, Takayama Y, Namatama K, Matsumoto Y, Kanaya T, et al. Neurointensive Care of Traumatic Brain Injury Patients Based on Coagulation and Fibrinolytic Parameter Monitoring. Neurologia Medico-Chirurgica. 2022;62(12):535–541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Slaughter AL, Nunns GR, D’Alessandro A, Banerjee A, Hansen KC, Moore EE, et al. The Metabolopathy of Tissue Injury, Hemorrhagic Shock, and Resuscitation in a Rat Model. Shock. 2018;49(5):580–590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jayaraman SP, Anand RJ, DeAntonio JH, Mangino M, Aboutanos MB, Kasirajan V, et al. Metabolomics and Precision Medicine in Trauma: The State of the Field. Shock. 2018;50(1):5–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kawata K, Liu CY, Merkel SF, Ramirez SH, Tierney RT, Langford D. Blood biomarkers for brain injury: What are we measuring? Neurosci Biobehav Rev. 2016;68:460–473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tchantchou F, Goodfellow M, Li F, Ramsue L, Miller C, Puche A, et al. Hyperhomocysteinemia-Induced Oxidative Stress Exacerbates Cortical Traumatic Brain Injury Outcomes in Rats. Cell Mol Neurobiol. 2021;41(3):487–503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dionisio N, Jardín I, Salido GM, Rosado JA. Homocysteine, intracellular signaling and thrombotic disorders. Current Medicinal Chemistry. 2010;17(27):3109–19. [DOI] [PubMed] [Google Scholar]
- 42.Peltz ED, D’Alessandro A, Moore EE, Chin T, Silliman CC, Sauaia A, et al. Pathologic metabolism: an exploratory study of the plasma metabolome of critical injury. The Journal of Trauma and Acute Care Surgery. 2015;78(4):742–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.de Lange Z, Kahler B, Smuts CM, Pieters M. Plasma phospholipid fatty acids are associated with altered fibrin clot properties in a population-based setting. Prostaglandins Leukot Essent Fatty Acids. 2019;143:1–7. [DOI] [PubMed] [Google Scholar]
- 44.Zheng F, Xia ZA, Zeng YF, Luo J, Sun P, Cui H, et al. Plasma metabolomics profiles in rats with acute traumatic brain injury. PloS one. 2017;12(8):e0182025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Schaid TR Jr., Cohen MJ, D’Alessandro A, Silliman CC, Moore EE, Sauaia A, et al. Trauma Induces Intravascular Hemolysis, Exacerbated by RBC Transfusion and Associated with Disrupted Arginine-Nitric Oxide Metabolism. Shock. 2022;59(1):12–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zheng F, Zhou YT, Li PF, Hu E, Li T, Tang T, et al. Metabolomics Analysis of Hippocampus and Cortex in a Rat Model of Traumatic Brain Injury in the Subacute Phase. Front Neurosci. 2020;14:876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zahedi K, Huttinger F, Morrison R, Murray-Stewart T, Casero RA, Strauss KI. Polyamine catabolism is enhanced after traumatic brain injury. Journal of Neurotrauma. 2010;27(3):515–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Walter FR, Harazin A, Tóth AE, Veszelka S, Santa-Maria AR, Barna L, et al. Blood-brain barrier dysfunction in L-ornithine induced acute pancreatitis in rats and the direct effect of L-ornithine on cultured brain endothelial cells. Fluids Barriers CNS. 2022;19(1):16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Shultz SR, Shah AD, Huang C, Dill LK, Schittenhelm RB, Morganti-Kossman MC, et al. Temporal proteomics of human cerebrospinal fluid after severe traumatic brain injury. Journal of Neuroinflammation. 2022;19(1):291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Karri J, Cardenas JC, Matijevic N, Wang Y, Choi S, Zhu L, et al. Early Fibrinolysis Associated with Hemorrhagic Progression Following Traumatic Brain Injury. Shock. 2017;48(6):644–650. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Digital Content 1. STROBE guidelines.
Supplemental Digital Content 2. Thrombelastography, metabolomics, and proteomics pre- and post-dura violation. Presented as medians and interquartile ranges.
Supplemental Digital Content 3. Changes in metabolomic profiles pre- and post-dura violation. Presented as medians and interquartile ranges.
Supplemental Digital Content 4. Changes in proteomic profiles pre- and post-dura violation. Presented as medians and interquartile ranges.