

Abstract
Background:
Elucidating biological mechanisms contributing to bipolar disorder (BD) is key to improved diagnosis and treatment development. With converging evidence implicating the metabotropic glutamate receptor 5 (mGlu5) in the pathology of BD, here, we therefore test the hypothesis that recently identified deficits in mGlu5 are associated with functional brain differences during emotion processing in BD.
Methods:
Positron emission tomography (PET) with [18F]FPEB was used to measure mGlu5 receptor availability and functional imaging (fMRI) was performed while participants completed an emotion processing task. Data were analyzed from 62 individuals (33±12 years, 45% female) who completed both PET and fMRI, including individuals with BD (n=18), major depressive disorder (MDD: n=20), and psychiatrically healthy comparisons (HC: n=25).
Results:
Consistent with some prior reports, the BD group displayed greater activation during fear processing relative to MDD and HC, notably in right lateralized frontal and parietal brain regions. In BD, (but not MDD or HC) lower prefrontal mGlu5 availability was associated with greater activation in bilateral pre/postcentral gyri and cuneus during fear processing. Furthermore, greater prefrontal mGlu5-related brain activity in BD was associated with difficulties in psychomotor function (r≥0.904, p≤0.005) and attention (r≥0.809, p≤0.028).
Limitations:
The modest sample size is the primary limitation.
Conclusions:
Deficits in prefrontal mGlu5 in BD were linked to increased cortical activation during fear processing, which in turn was associated with impulsivity and attentional difficulties. These data further implicate an mGlu5-related mechanism unique to BD. More generally these data suggest integrating PET and fMRI can provide novel mechanistic insights.
Keywords: Bipolar disorder, Metabotropic glutamate receptor 5, Positron emission tomography, Functional magnetic resonance imaging, Dorsolateral prefrontal cortex, Orbitofrontal cortex
INTRODUCTION
The early and accurate diagnosis of bipolar disorder (BD) represents a significant challenge in psychiatry. Despite mania/hypomania being the defining diagnostic feature, individuals with BD spend up to three times as much time in a depressed state relative to mania/hypomania and are more likely to seek treatment and present to clinicians in a depressed mood state.(Hirschfeld et al., 2005; Judd et al., 2002; Kupka et al., 2007) Therefore, it is unfortunately not surprising that BD is most often incorrectly diagnosed as major depressive disorder (MDD).(Young, 2009) Misdiagnosis can often lead to inappropriate treatment selection, with standard antidepressant therapies often having limited efficacy for bipolar depression.(Cheniaux and Nardi, 2019; Singh and Rajput, 2006) Critically, misdiagnosis and delay of appropriate treatment is associated with increased risk of recurrence and chronicity of episodes, greater number of emergency room visits and hospital admissions, and increased suicide attempts.(Hong et al., 2016; Keramatian et al., 2022; Peters et al., 2016; Shen et al., 2018) Taken together, this highlights our limited understanding of overlapping and unique neurobiological mechanisms of BD and MDD and the ongoing need for research aimed at identifying objective biomarkers, a crucial step towards the ultimate goal of improved diagnosis and the discovery of novel targets for developing safer and more effective treatments.
Neuroimaging studies could help the identification of biomarkers that differentiate BD from MDD and other psychiatric disorders. Indeed, numerous research groups have reported prominent brain structural, functional, and molecular abnormalities in BD.(Blond et al., 2012; Holmes et al., 2023; Ino et al., 2022; Schumer et al., 2023; Strakowski et al., 2012) Findings are particularly robust within corticolimbic circuitry that is known to play an essential role in executive function and emotional processing,(Friedman and Robbins, 2022) which appears to subserve emotional dysregulation and cognitive difficulties, prominent features of BD.(Bi et al., 2022; Strakowski et al., 2012) For instance, relative to psychiatrically healthy individuals, individuals with BD display altered corticolimbic circuitry engagement and connectivity during tasks assessing emotional processing, with findings oftentimes being mood-state specific.(Bi et al., 2022; Liu et al., 2012; Strakowski et al., 2012) Research directly comparing BD and MDD indicates some similarities in emotional processing common to a depressive mood-state but also highlight emotional processing differences between BD depression and MDD that may be associated with stimuli valence and distinct processing pathways [for reviews, see (Gong et al., 2020; Han et al., 2019; Siegel-Ramsay et al., 2022)]. Specifically, an implicit emotion face-processing paradigm has been used in conjunction with functional imaging by our research group and others to successfully elicit engagement of functional brain networks relevant to BD and has yielded significant differences between BD, MDD, and healthy control (HC) groups in corticolimbic activation and functional connectivity.(Goldman et al., 2022; Johnston et al., 2017; Kerestes et al., 2012; Liu et al., 2012; Liu et al., 2014) The underlying molecular etiology of these differences in brain network engagement and functional connectivity remain unknown and may reflect unique neurobiological mechanisms of BD.
The metabotropic glutamate receptor 5 (mGlu5) is a predominately neuronal, postsynaptic receptor, and substantial preclinical evidence highlights the necessity of mGlu5 signaling in mediating types of synaptic plasticity subserving learning and memory, particularly fear learning.(Asch et al., 2023; Rodrigues et al., 2002) Further, more recent evidence from clinical investigations points to mGlu5 as potentially contributing to prefrontal disfunction in stress and trauma-related psychiatric disorders, including BD and MDD,(Asch et al., 2023; Esterlis et al., 2022; Esterlis et al., 2018; Holmes et al., 2017; Holmes et al., 2023) and the potential utility of mGlu5 receptor availability as a BD “trait biomarker”.(Holmes et al., 2023) Specifically, using positron emission tomography (PET) with the mGlu5 receptor specific radiotracer, [18F]FPEB, we found lower mGlu5 receptor availability in prefrontal regions, particularly dorsolateral prefrontal cortex (dlPFC) and OFC, among individuals with BD, irrespective of mood state, relative to both individuals with MDD during depression and healthy controls.(Holmes et al., 2023) Here, we performed analyses in a subset of participants who completed both fMRI and PET in order to further investigate the potential functional consequence and clinical relevance of the previously observed lower mGlu5 receptor availability in dlPFC and OFC in BD. To address this overarching goal, we completed the following: (1) assessed whether we were able to demonstrate differences similar to those previously reported in brain activation during the implicit processing of fearful emotion between BD, MDD, and a healthy comparison (HC) group, (2) examine relationships between functional brain responses and prefrontal mGlu5 receptor availability and how such relationships might be a function of diagnosis, and (3) explore possible relationships between clinical symptoms and functional activation of brain regions relevant to the primary hypotheses.
METHODS AND MATERIALS
Study participants
Recruitment methods for this study included flyers, word of mouth, community outreach, and referrals from local clinics/hospitals. To determine eligibility, participants underwent physical and neurological examination to rule out major medical or neurological illnesses with potential central nervous system effects. Screening involved electrocardiography, complete blood counts, serum chemistries, thyroid function test, liver function test, urinalysis and urine toxicology screening, and urine pregnancy tests (for individuals assigned “female at birth”). Diagnoses were confirmed using the Structured Clinical Interview for DSM-5.(First et al., 2015) Exclusion criteria were lifetime history of psychotic disorders; diagnosis of alcohol or substance use disorder (with the exception of nicotine and cannabis use); positive urine toxicology [testing for amphetamine, buprenorphine, cocaine, 2-ethylidene-1,5-dimethyl-3,3-diphenylpyrrolidine (EDDP), 3,4-Methylenedioxymethamphetamine (MDMA), methadone, methamphetamine, morphine, oxazepam, oxycodone, phencyclidine (PCP), Dextropropoxyphene (PPX), and secobarbital; iScreen™ Urine Dx test, Abbott Rapid Dx North America] or pregnancy tests before any scan; history of loss of consciousness for more than 5 minutes; significant medical condition; and contraindications to MRI/PET scans. Exclusion criteria were the same for the control group, except for the addition of no current or history of any DSM diagnosis, except for nicotine or cannabis use. The Yale University Human Investigation Committee and the Radioactive Drug Research Committee approved the study, and all participants provided written informed consent before inclusion in the study.
For this study, we analyzed data from a total of N=62 individuals (Table 1), including participants with bipolar disorder (BD: n=18; 32.5±10.7 years; 55.6% female), major depressive disorder (MDD: n=20; 34.6±11.1 years; 45.0% female), as well as individuals with no history of psychiatric illness, who served as the healthy comparison group (HC: n=24; 33.0±13.0 years; 37.5% female).
Table 1.
Description of demographic features, clinical characteristics, and neuroimaging parameters of the study sample (N=62).
| Demographics | HC (n=24) | BD (n= 18) | MDD (n=20) | Statistical Test (F or χ2, p-value) |
|---|---|---|---|---|
| Age, years (mean±SD) | 33.0±13.7 | 32.5±10.7 | 34.6±11.1 | 0.135, 0.874 |
| Sex (n, %female) | 9, 37.5% | 10, 55.6% | 9,45.0% | 1.354, 0.508 |
| Tobacco (n, %users) | 5, 21.7% | 5, 27.8% | 4, 21.1% | 0.287, 0.866 |
| Cannabis (n, %users) | 8, 33.3% | 7, 38.9% | 7, 35.0% | 0.142, 0.932 |
| Clinical Characteristics | ||||
| Depression, HAMD (mean±SD) | 0.6±0.9a,b | 8.3±5.7b | 13.7±6.4 | 42.280, <0.001 |
| Impulsivity, BIS (mean±SD) | 49.5±7.0a,b | 71.1±8.8 | 68.3±11.7 | 35.493, <0.001 |
| Psychotropic medication (n, %users) | --- | 4, 22.2% | 8, 40.0% | 1.386, 0.239 |
| Neuroimaging Parameters | ||||
| Time between scans, days (mean±SD) | 4.0±4.6 | 2.3±2.4 | 4.0±4.5 | 1.122, 0.332 |
| fMRI motion, %censored (mean±SD) | 2.4±3.1 | 2.1±2.9 | 1.5±2.3 | 0.500, 0.609 |
| fMRI motion, FD (mm/TR) (mean±SD) | 0.2±0.05 | 0.2±0.06 | 0.2±0.06 | 0.842, 0.436 |
| FPEB injected dose, MBq (mean±SD) | 157.0±35.7 | 153.9±35.5 | 170.9±14.1 | 1.854, 0.166 |
| FPEB injected mass, μg (mean±SD) | 0.4±0.2 | 0.4±0.2 | 0.3±0.2 | 0.340, 0.713 |
| Plasma free fraction, fp (mean±SD) | 0.05±0.008 | 0.04±0.007 | 0.05±0.010 | 1.677, 0.196 |
| FPEB injection time, hh:mm (mean±SD) | 12:53±1:22 | 12:45±0:59 | 13:04±0:58 | 0.315, 0.731 |
Significantly different (p<0.05uncorr) from BD
Significantly different (p<0.05uncorr) from MDD
Abbreviations: Barratt Impulsiveness Scale (BIS); bipolar disorder (BD); framewise displacement (FD); functional magnetic resonance imaging (fMRI); Hamilton Depression Scale (HAMD); Major Depressive Disorder (MDD); Mega Becquerel (MBq); psychiatrically healthy control (HC); repetition time (TR); standard deviation (SD).
Clinical assessments
Depression symptoms were assessed using the Hamilton Depression (HAMD) 17 Item scale.(Hamilton, 1960) Of participants with BD (BD1 n=15; BD2 n=3), nine were euthymic (HAMD≤7) and 9 were mild (HAMD 8–16; n=7) to moderately (HAMD 17–23, n=2) depressed on the study day. Of MDD participants, all met criteria for past MDD, and on the study day, five were euthymic, seven were mildly (n=4) to moderately (n=3) depressed, and eight we severely depressed (HAMD≥24). The Barratt Impulsiveness Scale version 11 (BIS) was used to evaluate impulsivity.(Stanford et al., 2009) A subset of study participants (n=12 HC, n=7 BD, n=10 MDD) completed the CogState testing battery (https://www.cogstate.com/)-specifically, the Detection Test (DET), Identification Test (IDN), and International Shopping List Delayed Recall ISLR)-were used to assess psychomotor function, attention, and verbal learning/memory, respectively.(Fredrickson et al., 2010; Thompson et al., 2011)
Magnetic resonance imaging
The emotional face task
Faces from the Ekman series (Ekman, 1976) depicting fear, happiness, or a neutral affect were shown to participants via Eprime software on a computer attached to a projector (Psychology Software Tools, Pittsburgh, PA). Participants were instructed to make indicate if the displayed face was male or female by button press. Each face was presented for 2 s with 4–12 s inter-stimulus-interval, during which a cross-hair fixation point was displayed. Each run comprised 10 gray scale face stimuli (5 females, 5 males) with each actor exhibiting all three of the expressions, yielding a total of 30 stimuli in each run. The order of stimuli presentation was counterbalanced for facial expression, gender, identity, as well as the duration of the inter-stimulus-interval. Participants completed four runs of the task. Details of this emotional face gender-labeling task have been previously reported, (Kerestes et al., 2012; Liu et al., 2012) and additional information is provided in the Supplement.
MRI data processing and analysis
The methods for structural and functional MRI data acquisition have been previously reported (for details, see Supplement).(Kerestes et al., 2012; Liu et al., 2012) Processing and analysis of fMRI were performed using SPM 12 (http://fil.ion.ucl.ac.uk/spm/). The three task stimulus conditions (fear, happy, neutral) were modeled separately in an event-related design that included 6-parameter motion-correction regressors and a regressor for framewise displacement (FD)-based motion-artifact censoring.(Jones et al., 2022) For FD-based censoring, we used a >0.5mm/TR and 5% data-loss threshold. The resulting censored dataset included 98% of the original frames (i.e., an average of 2% being censored).
Implicit processing of fearful emotional expressions was assessed as the BOLD response to fearful faces relative to unmodelled inter-stimulus intervals (i.e., FEAR > fixation), hereafter referred to as the “FEAR BOLD.” To investigate the main effect of group/diagnosis, we performed both ROI and voxel-wise analyses of the FEAR response. Additionally, relationships with dlPFC or OFC mGlu5 receptor availability (as measured by PET, see below) were examined using standard linear modeling: i.e., ANCOVAs with the PET parameter (volume of distribution, VT) as a covariate. Brain-wide (voxel-wise) analyses were conducted using a cluster-extent based thresholding with a primary/voxel threshold of p<0.005 and a cluster-level threshold of puncor<0.05. For post hoc testing following significant F-test results, parameter estimates (β weights) for significant clusters identified in brain-wide analyses were extracted using the MarsBaR toolbox (version 0.45, https://marsbar-toolbox.github.io/). MarsBaR was also used to extract individual subject β-weights from the right and left dlPFC and OFC ROIs (Figure S1).
Positron emission tomography (PET) with [18F]FPEB
All study participants underwent PET imaging with the mGlu5 receptor-specific radiotracer [18F]FPEB. Radiotracer was synthesized onsite (Park et al., 2015) and administered intravenously as a bolus plus constant infusion. This administration schedule has been demonstrated to achieve a state of equilibrium by 60 min after the start of injection.(Park et al., 2015; Sullivan et al., 2013) Emission data were acquired from 90–120 min after the start of injection on the high-resolution research tomograph (HRRT; Siemens, Erlangen, Germany). A 6-min transmission scan was obtained for attenuation correction, and head motion was tracked using the Polaris Vicra optical tracking system (Vicra; NDI System). Dynamic scan data were reconstructed with corrections using the ordered subset expectation maximization-based MOLAR algorithm. There is not a valid reference region for mGlu5,(Patel et al., 2007) thus, volume of distribution (VT), defined as the ratio of radioligand concentration in ROI to the concentration in plasma at equilibrium, was used as the outcome measure. VT was estimated by the equilibrium analysis method (Sullivan et al., 2013) and a venous plasma input function (Park et al., 2015) as previously reported and described in greater detail in the Supplemental Materials. Given reports of circadian variation in mGlu5 receptor availability as measured by PET,(DeLorenzo et al., 2016; Elmenhorst et al., 2016) all injections were performed between 10:48 and 15:42 hours (average time of 12:54 hours ± 68 minutes), with no differences in the average injection time between groups (Table 1). Further, in the current dataset, there were no apparent relationships between time of injection and regional VT values (Figure S2).
Statistical analysis
Analysis of variance (ANOVA) with Least Significant Difference (LSD) post hoc tests or Chi2 tests were used to assess differences between demographic and radiotracer characteristics across groups. Primary hypotheses were tested using ROI and whole-brain analyses. For this study, dorsolateral prefrontal cortex (dlPFC) and orbitofrontal cortex (OFC), as defined using the Anatomical Automatic Labeling (AAL) atlas,(Rolls et al., 2020) were selected as the primary regions of interest (ROIs), and FEAR BOLD β-weights were extracted using the MarsBaR toolbox (version 0.45, https://marsbar-toolbox.github.io/, Figure S1A). Group differences in the FEAR BOLD were assessed using either a mixed effects general linear model, with ROI (dlPFC, OFC) and side (left, right) as within subject measures and diagnosis (HC, BD, MDD) as a fixed effect, or a multivariate ANOVA (MANOVA) with SPM cluster β-weights as dependent variables and diagnosis as a fixed effect. Pairwise post hoc tests were corrected for multiple comparisons (Bonferroni’s family-wise error). To evaluate associations between FEAR BOLD β-weights and clinical and cognitive measures, we computed Pearson’s r. Findings were considered significant at a two-tailed p<0.05 threshold.
Statistical tests were performed using IBM SPSS Statistics, version 29.0.1.0 (IBM Corp., Armonk, NY) and Prism GraphPad, version 10.0.2 (GraphPad Software, Boston, Massachusetts) was used for data visualization. Brain-wide (voxel-wise) analyses were conducted in SPM with a primary/voxel threshold of p<0.005 and a cluster-level threshold of puncor<0.05.
RESULTS
Among the study participants, there were no significant differences between groups in age, male:female ratio, tobacco, or cannabis users (Table 1). For individuals in the clinical groups, there were no differences in overall psychiatric medication use, with a full medication list provided in Table S1. Scan parameters for both fMRI and PET were not statistically different between groups; specifically, there were no group-wise differences in %censoring, FD, injected PET tracer dose or mass (Table 1).
Group differences in implicit processing of fearful affect
In our initial assessment of the functional brain responses during the fearful facial emotion processing task (i.e., FEAR BOLD), we took a region of interest (ROI)-based approach (Figure S1), with OFC and dlPFC, as they are defined in our PET analyses (Figure S1A), as ROIs. Using a mixed effects general linear model (Figure S1B), there were main effects of both ROI (OFC>dlPFC; F1,59=11.47, p=0.001) and hemisphere (right>left; F1,59=64.05, p<0.001). There was also a significant group*hemisphere interaction (F2,59=4.07, p=0.022) such that right lateralized brain responses in the BD group were significantly greater than those observed in HC group (pFWE=0.047).
The brain-wide analysis of the FEAR BOLD revealed overall group differences (Figure 1A) and identified five clusters where there were significant main effects of diagnosis (voxel threshold p<0.005, cluster threshold p<0.05). Statistics and anatomical location of each cluster are provided in Table 2. Notably, Cluster 1, which included voxels in the right inferior frontal gyrus, insula posterior OFC and superior temporal pole, remained significant after multiple comparison correction (puncorr<0.001; pFWE=0.014), as did Cluster 2 (puncorr<0.001; pFWE=0.001), with voxels located in the right supramarginal and the inferior and superior parietal gyri.
Figure 1. Regions of hyperactivation during implicit processing of fearful affect in BD.

[A] Significant clusters (voxel p<0.005, cluster p<0.05) identified in a voxel-wise ANOVA testing for an overall main effect of group are visualized in common space in the axial orientation. Each of the five clusters is indicated by an arrowhead in a color that coordinates with colored text on Table 2 and [B] bar graph x-axis labels. Bar graph displays extracted individual subject β-weights (line at group mean) and pairwise group comparisons for each cluster [post hoc tests with Bonferroni’s (family-wise error, FWE) correction for multiple comparisons]: * pFWE <0.05; ** pFWE <0.01; *** pFWE <0.001 relative to BD.
Table 2.
Significant clusters (voxel p<0.005, cluster p<0.05) identified in a voxel-wise test of group differences in the FEAR BOLD (β), and significant correlations between individual subject FEAR BOLD and measures of impulsiveness and verbal memory, where applicable.
| Cluster | P FWE | P uncorr | K E | Peak Voxel | Anatomical Designation (AAL) | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| 1 | 0.014 | <0.001 | 516 | 50 | 20 | −10 | R. IFG, Insula, Posterior OFC, Superior temporal pole |
| 2 | 0.001 | <0.001 | 783 | 54 | −34 | 42 | R. Supramarginal, Inferior & Superior parietal gyri |
| 3 | 0.325 | 0.009 | 232 | −62 | −38 | 24 | L. Superior temporal & Supramarginal gyri |
| 4 | 0.610 | 0.022 | 170 | −46 | −26 | 34 | L. Inferior parietal & Supramarginal gyri |
| ISLR | r=−0.907, p=0.005 | ||||||
| 5 | 0.632 | 0.024 | 166 | 60 | −12 | 22 | R. Pre/post central gyri, Rolandic operc., Supramarginal gyrus |
| BIS | r=0.486, p=0.041 | ||||||
| ISLR | r=−0.805, p=0.015 | ||||||
Note: Cluster numbers and text color correspond those used in Figure 1.
Abbreviations: automated anatomical atlas version 3 (AAL); Barratt Impulsiveness Scale version 11 (BIS); blood oxygenation level dependent (BOLD); cluster extent (KE); family-wise error (FWE); inferior frontal gyrus (IFG); International Shopping List Delayed Recall (ISLR); left (L); operculum (operc.); orbitofrontal cortex (OFC); right (R); uncorrected (uncorr).
To determine which group(s) were driving this overall main effect, we performed a MANOVA with the mean β-weights extracted from each of the five significant clusters as dependent variables, and diagnosis as a fixed factor (Figure 1B). For all five clusters, the FEAR BOLD in the BD group was significantly greater than that observed in the HC group (pFWE≤0.006). and relative to the MDD group across all five clusters (pFWE≤0.033). The MDD group tended to show greater FEAR BOLD as compared with HC within Cluster 1 (right inferior frontal gyrus, insula posterior OFC and superior temporal pole), but this did not remain significant following multiple comparison correction (puncorr=0.028; pFWE=0.084). For the remaining four clusters, MDD was not different from HC, with or without multiple comparison correction. The results of this analysis held up after correcting for medication status to account for the potential impact of psychotropic medication (Table S2).
Correlations between prefrontal mGlu5 receptor availability and brain activation
Results of brain-wide ANCOVA analyses testing for relationships between dlPFC or OFC mGlu5 availability (i.e., [18F]FPEB VT, see Table S3) and FEAR BOLD are shown in Figure 2. For the analysis of correlations with dlPFC mGlu5 receptor availability, no correlations, positive or negative, were observed in the HC or MDD groups. In the BD group, there were no significant positive correlations, however, we observed significant negative correlations in four clusters of voxels (Figure 2A and 2C, Table 2A): Cluster 1 (puncorr=0.035; pFWE=0.632) was located in left pre/postcentral gyri; Cluster 2 (puncorr=0.027; pFWE=0.530) in the right pre/postcentral gyri; and Clusters 3 and 4 (both puncorr=0.037; pFWE=0.602) in the right cuneus, calcarine, and superior occipital gyrus and left cuneus, superior and middle occipital gyri, respectively. For the OFC ANCOVA (Figure 2B and 2D), four clusters sharing the same peak voxels as those from the dlPFC analysis (Table 2B) were also found to be negatively correlated with brain activation within the BD group. As with dlPFC, no correlations for OFC mGlu5 availability were observed in the HC and MDD groups.
Figure 2. Negative correlations between prefrontal mGlu5 receptor availability and FEAR BOLD response in BD.

Significant clusters (voxel p<0.005, cluster p<0.05) identified in a voxel-wise ANCOVA testing for correlations between [A] dlPFC and [B] OFC mGlu5 receptor availability ([18F]FPEB VT) and brain activation during implicit processing of fearful facial expressions (fMRI FEAR BOLD β) among individuals with BD are visualized in common space in the axial orientation. Each of the significant clusters is indicated by an arrowhead in a color that coordinates with colored text on Table 3. Negative relationships between [18F]FPEB VT values and ANCOVA-identified cluster mean FEAR BOLD β weights are shown for [C] dlPFC and [D] OFC.
Correlations with clinical features: exploratory analyses
Given that our finding of group differences in FEAR BOLD were driven by hyperactivation in BD relative to the two other groups and that correlations between FEAR BOLD and prefrontal mGlu5 receptor availability were specific to the BD group, our correlation analyses focused on the BD group.
Hyperactivation in BD is associated with impulsiveness and verbal memory impairment
We first tested for associations between clusters of significant hyperactivation in BD (Figure 1) and clinical features. There was a significant positive correlation for β-weights extracted from the cluster in right pre/postcentral gyrus, Rolandic operculum, and supramarginal gyrus, with impulsiveness (r=0.486 p=0.041, Figure 3A), and verbal memory (r=−0.850, p=0.015, Figure 3B), such that higher measures of impulsiveness and worse performance on the delayed recall task was observed in individuals with the greatest brain activation in this brain region during passive processing of fearful expression. There was additionally a negative relationship between verbal memory and β-weights extracted from the cluster located in left inferior parietal and supramarginal gyri (r=−0.907, p=0.005, Figure 3B).
Figure 3. Correlations with clinical features in BD.
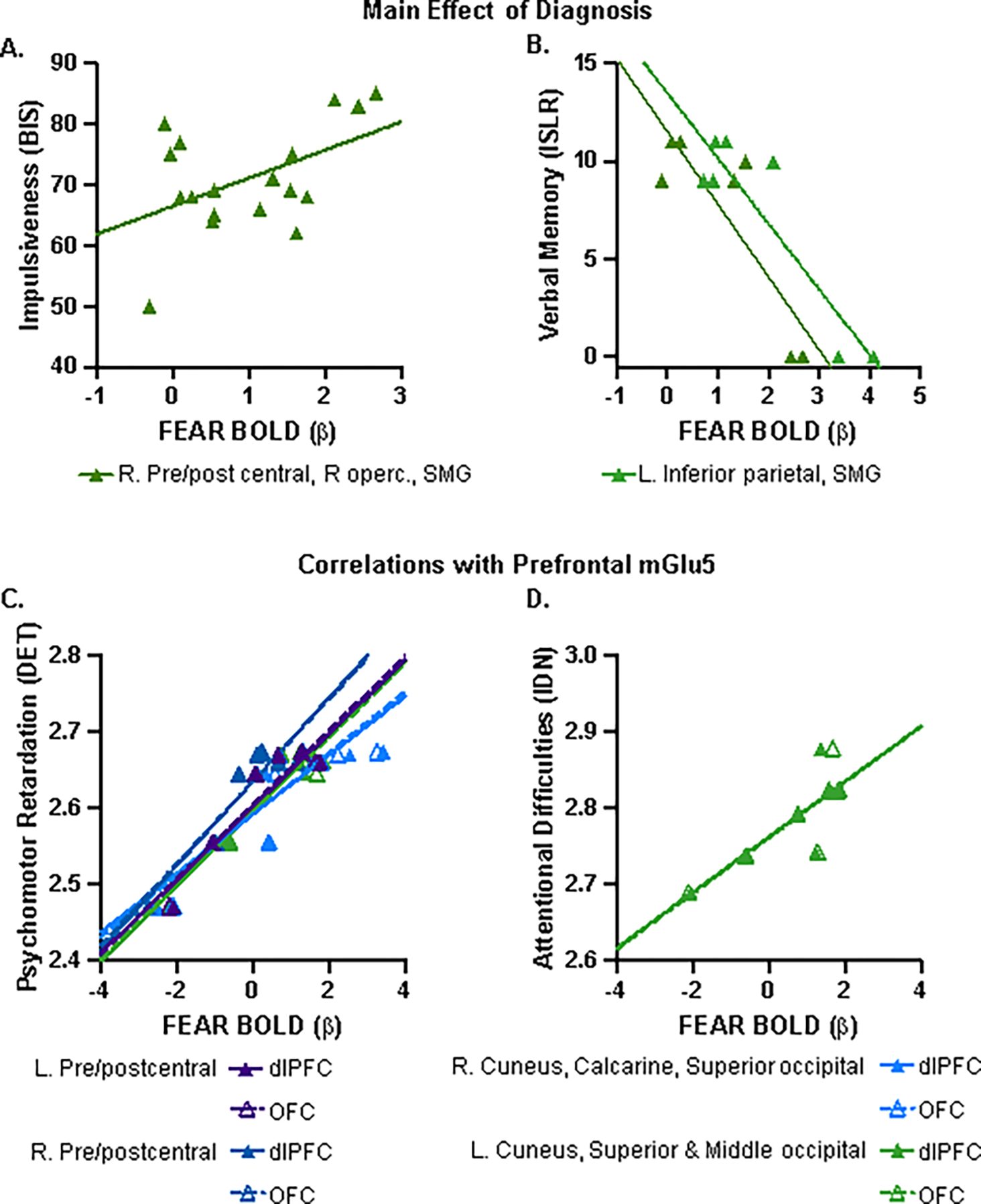
Significant positive correlation between clusters of hyper-activation in the BD group (FEAR BOLD β) and [A] impulsiveness as measured by the Barratt Impulsiveness Scale (BIS), and [B] negative correlations with verbal memory as assessed by the International Shopping List Delayed Recall (ISLR) task. Exact Pearson’s r and associated p-values are provided in Table 3. Significant correlations between dlPFC and OFC mGlu5-related FEAR BOLD and [C] psychomotor function, as measured by the Detection Test (DET), and [D] attentional difficulties assessed by the Identification Test (IDN). [E] Exact Pearson’s r and associated p-values are provided in the table. Abbreviations: dorsolateral prefrontal cortex (dlPFC); left (L); orbitofrontal cortex (OFC); right (R); supramarginal gyrus (SMG); Rolandic operculum (R operc.)
Prefrontal mGlu5-correlated brain activation in BD is associated with psychomotor function and attentional difficulties
We proceeded with exploratory analyses testing for potential relationships between dlPFC and OFC mGlu5 receptor-related brain activity, clinical measures, and cognitive function in the BD group. There were no significant correlations with depression symptom severity or impulsiveness (data not shown). However, for both the dlPFC and OFC derived clusters, we observed significant correlations with psychomotor function (Figure 3C) such that greater brain activation during fearful emotional processing was associated with greater psychomotor retardation, with Pearson’s correlations ranging from r=0.904 to r=0.955 (p=0.003 to p<0.001). Additionally, FEAR BOLD of the left cuneus, superior and middle occipital gyri clusters derived from both dlPFC and OFC were associated with greater attentional difficulties (Figure 3D; r=0.809–0.835, p=0.028–0.019). Exact Pearson’s r and associated p-values are provided in Table 3.
Table 3.
Significant clusters (voxel p<0.005, cluster p<0.05) identified in a voxel-wise test of correlations between [A] dlPFC and [B] OFC [18F]FPEB VT values and the FEAR response (β); and significant correlations between individual subject FEAR responses and performance on cognitive tasks, where applicable.
| Cluster | A. dIPFC VT covariate | B. OFC VT covariate | Peak voxel | Anatomical Designation (AAL) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| p FWE | P uncorr | k E | p FWE | p uncorr | k E | x | y | z | ||
| 1 | 0.632 | 0.035 | 219 | 0.754 | 0.050 | 186 | −36 | −14 | 36 | L. Pre/postcentral gyri |
| DET | r=0.925 p=0.003 | r=0.936 p=0.002 | ||||||||
| 2 | 0.530 | 0.027 | 247 | 0.690 | 0.041 | 203 | 52 | −6 | 36 | R. Pre/postcentral gyri |
| DET | r=0.924 p=0.003 | r=0.932 p=0.002 | ||||||||
| 3 | 0.602 | 0.033 | 227 | 0.644 | 0.037 | 215 | 16 | −82 | 22 | R. Cuneus, Calcarine, Superior occipital gyrus |
| DET | r=0.904 p=0.005 | r=0.919 p=0.003 | ||||||||
| 4 | 0.602 | 0.033 | 227 | 0.600 | 0.032 | 227 | −18 | −84 | 16 | L. Cuneus, Superior & Middle occipital gyri |
| DET | r=0.955 p<0.001 | r=0.948 p=0.001 | ||||||||
| IDN | r=0.809 p=0.028 | r=0.835 p=0.019 | ||||||||
Abbreviations: automated anatomical atlas version 3 (AAL); cluster extent (KE); Detection Test (DET); dorsolateral prefrontal cortex (dlPFC); family-wise error (FWE); Identification Test (IDN); inferior frontal gyrus (IFG); left (L); orbitofrontal cortex (OFC); right (R); uncorrected (uncorr).
DISCUSSION
To our knowledge, this is the first study to examine task fMRI and [18F]FPEB PET in the same individuals with bipolar disorder (BD) or major depressive disorder (MDD), and as compared with psychiatrically healthy controls (HC). Consistent with the study hypothesis and previous research,(Gong et al., 2020; Han et al., 2019; Liu et al., 2012; Siegel-Ramsay et al., 2022; Strakowski et al., 2012) individuals with BD displayed altered brain function during a fearful facial expression processing task (i.e., the FEAR BOLD) relative to the MDD and HC groups, including elevated activation of the a priori ROIs dlPFC and OFC. In the BD group, we further observed negative associations between prefrontal mGlu5 and functional brain circuitry subserving fearful facial expression processing, with no such relationship apparent within either the HC or MDD group. Exploratory analyses additionally revealed relationships between prefrontal-mGlu5-related fear processing in individuals with BD and their performance on tests of verbal memory, impulsiveness, and related cognitive domains.
We first assessed whether the fearful emotional processing task used here produced similar differences in brain activation to those previously reported using both ROI and voxel-wise analyses. For the ROI approach, dlPFC and OFC were selected based on our recent report of lower mGlu5 receptor availability in a larger sample (n=27) of individuals with BD,(Holmes et al., 2023) their well-defined roles in executive function and emotional regulation, (Friedman and Robbins, 2022) as well as being specifically implicated in threat detection and the cognitive regulation of subjective fear.(Kroes et al., 2019; Shih and Chang, 2021) Indeed, we observed differential activation in our a priori frontal ROIs, more specifically right lateralized responses in the BD group were significantly greater than those observed in HC group. The brain-wide analysis revealed further frontal and parietal regions of enhanced activation among BD participants relative to MDD and HC, including supramarginal gyrus, Rolandic operculum, inferior frontal gyrus, and posterior OFC. While we did not observe differences in These findings are in agreement with general observations of differences in emotional brain circuitry function and engagement between BD and MDD,(Gong et al., 2020; Han et al., 2019; Liu et al., 2012; Siegel-Ramsay et al., 2022; Strakowski et al., 2012) and are additionally consistent with the notion that mGlu5 deficits in these prefrontal regions could be contributing to brain dysfunction, especially pertaining to emotional processing in BD.
Another finding of note is the observed positive correlation between hyperactivation of the right supramarginal gyrus and Rolandic operculum and a measure of impulsivity in BD, such that higher self-reported impulsiveness was observed in the individuals with greater FEAR BOLD. This right lateralized brain region has previously been implicated in trait impulsivity among typically developing youth,(Inuggi et al., 2014) and the right supramarginal gyrus, in particular, has been found to be related with attentional impulsivity(Besteher et al., 2019) and “distractibility”(Demeter et al., 2011) in healthy adults. In addition, the Rolandic operculum and supramarginal gyrus have long been recognized for their roles in emotion recognition and regulation, especially as it pertains to distinguishing emotions of self-versus-other.(Adolphs, 2002; Lamm et al., 2019) Finally, a recent meta-analysis investigating neural correlates of emotional processing identified right-lateralized hyperactivation of this region in studies of BD across mood states.(Förster et al., 2023)
While not part of the primary analyses, we tested for associations between OFC or dlPFC mGlu5 receptor availability and FEAR BOLD signals extracted from the 5 Clusters identified by the brain-wide analysis. When looking across all three groups, we observed no significant correlations (N=62, r= −0.091 to −0.189, p=0.481 to 0.142). We also looked within each group independently (Table S4), resulting in a single significant correlation between dlPFC mGlu5 availability and Cluster 2 (right supramarginal gyrus, inferior and superior parietal gyri) in the HC group (r= −0.438, p=0.032). These (null) results are perhaps not surprising: regional mGlu5 receptor availability may not be strongly associated with local activation, but rather associated with regulation of downstream target region activation. Therefore, to address this question of downstream functional consequences of mGlu5 receptor availability and the potential interaction with, or impact of psychiatric diagnosis, we performed a brain-wide analysis to test for relationships between mGlu5 receptor availability in our prefrontal a priori ROIs and FEAR BOLD responses. In the BD group, we observed significant negative correlations such that lower prefrontal mGlu5 receptor availability was associated with greater FEAR BOLD in bilateral pre and postcentral gyri, and bilateral cuneus and superior occipital gyrus. The precentral gyrus (primary motor cortex) and postcentral gyrus (primary somatosensory cortex) have long been recognized as integral to sensory motor function; however, there is mounting evidence implicating additional roles in emotional processing and emotion regulation.(Adolphs, 2002; Adolphs et al., 2000; Damasio et al., 2000; Kropf et al., 2019) Of particular relevance, the somatosensory cortex has been identified within the context of fear learning and retrieval of fearful memories.(Damasio et al., 2000) The observed relationships between the FEAR BOLD in pre and postcentral gyri and our measure of psychomotor function are, therefore, congruent with these roles in both sensory motor function and emotional processing. Bilateral cuneus, the other region whose FEAR BOLD was found to negatively correlate with prefrontal mGlu5 in BD, is part of the default mode network (DMN), a brain circuit typically “active” in a resting state and inactive or inhibited during a task,(Greicius et al., 2003) including deactivation during an emotional faces task, not dissimilar to the one used here, in healthy adults.(Sreenivas et al., 2012) Evidence also suggests the DMN may help mediate shifts in attentional states to focus on internal versus external stimuli.(Buckner et al., 2008; Scheibner et al., 2017; Weissman et al., 2006) Presently, we found the bilateral cuneus FEAR BOLD to be associated with psychomotor retardation, and FEAR BOLD in left cuneus, superior and middle occipital gyri were additionally associated with attentional difficulties in BD. We propose these data, along with the negative nature of the observed correlations (i.e., lower prefrontal mGlu5 in BD relative to HC and MDD corresponding with hyperactivation during the emotional processing task) collectively support a hypothetical mechanism by which deficient prefrontal mGlu5 receptor availability (and perhaps, by proxy, receptor signaling and function) in BD contributes to inefficient top-down control of lower-order processing networks, including disinhibition of DMN, which in turn contributes to alterations in function across cognitive domains important for emotional regulation and impulse control.
Interestingly, in the HC and MDD groups, there were no significant correlations between dlPFC or OFC mGlu5 and the FEAR BOLD anywhere in the brain. While one might hypothesize the involvement or influence of mGlu5 receptor availability would be “transdiagnostic”, we frequently observe diagnosis-specific roles for mGlu5.(Asch et al., 2022; Baldassarri et al., 2023; Davis et al., 2019a; Esterlis et al., 2022; Holmes et al., 2023) For example, we have reported upregulation of mGlu5 receptor in individuals with PTSD but not MDD, and variability in associations between mGlu5 availability and symptoms among people with PTSD vs MDD, including with suicidality.(Davis et al., 2019b) Furthermore, differential relationships between mGlu5 availability and cognitive processes, most specifically, attention, among individuals with PTSD versus individuals with MDD were shown.(Esterlis et al., 2022) Most recently, we conducted the first study to examine mGlu5 involvement in depression among individuals with BD versus MDD. We showed that in BD depression and euthymia, mGlu5 levels are significantly lower than in individuals who are controls or have MDD, but no differences between MDD with depression vs controls. Furthermore, we showed differential associations between mGlu5 availability and mood symptoms in those with MDD but not with BD during depression.(Holmes et al., 2023) Therefore, the role of mGlu5 receptor availability is complex and appears to be context, or in this case, diagnosis, dependent. Studies such as these, aimed at understanding the unique contribution of mGlu5 receptor to BD pathology and symptomatology, are critical toward the development of targeted, mechanistically informed therapeutic approaches.
The present study is not without limitations, with a number warranting specific discussion. First, and most importantly, the study sample size, although not uncommon for PET, particularly in clinical populations, is limited and smaller than typically desirable for fMRI studies. Our comparatively small sample size significantly limits statistical power. As such, the current results and interpretations should be viewed critically and with caution and larger replication studies are warranted. Second, there are a number of demographic and clinical variables, such as socioeconomic status, illness duration, number of major depressive or manic episodes, et cetera, that we were not able to statistically account for and could be contributing to, or partially mediating the presented findings. Third, the BD sample is limited in mood symptom severity due to recruitment of individuals across mood states and exclusion of individuals with severe medically uncontrolled symptoms due to safety reasons. Therefore, we are not able to address any questions regarding mood state as a potential mediator of the observed relationships between prefrontal mGlu5 and brain function, and additional research in larger samples that capture a broader range of mood symptoms is needed. Alternatively, because we included participants across a limited range of mood states, the findings here can be interpreted as reflecting a BD trait marker, rather than a state-dependent phenotype. Forth, 22.2% of the BD group and 40.0% of the MDD group reported using some type of psychotropic medication. While not excluding medicated individuals is arguably of greater ecological validity, it does introduce a potential confound given potential effects on individual neurochemistry. It will be of interest to perform a systematic investigation of the potential influence of past or present medication use on mGlu5 availability and brain function and the possible down-stream impact on mood and cognitive symptoms. Finally, we limited our covariate analyses of mGlu5 measures in dlPFC and OFC and therefore cannot comment on the possible contribution of regions outside our a priori selected ROIs. Certainly, other regions, particularly subcortical regions such as amygdala, hippocampus, and striatum, express mGlu5 receptor and have been implicated in the pathobiology of BD,(Asch et al., 2023; Blond et al., 2012; Esterlis et al., 2018; Strakowski et al., 2012) and will be important to consider in our ongoing and future work.
To summarize, we show evidence that combining molecular and functional imaging modalities provides novel insights regarding differential mechanistic neurobiological underpinnings of BD. Specifically, this is the first study to combine analyses of task fMRI and [18F]FPEB PET performed in the same individuals with BD and MDD, offering unique insight into the relationships between alterations in brain function and prefrontal mGlu5 and probe the influence of psychiatric diagnosis. Indeed, our findings demonstrate relationships specific to BD between prefrontal mGlu5 availability and brain activation during emotional processing. Further, our data suggest a mechanism by which deficient prefrontal mGlu5 in BD contributes to pathological alterations in emotional processing and impulsiveness, and highlights the potential of targeting mGlu5 as a strategy for improving mood and cognitive outcomes in BD. Moreover, based on the present findings and constant with our previous report,(Holmes et al., 2023) mGlu5 dysregulation may represent a trait marker for BD, which could aid with early differential diagnosis from MDD. While these initial findings are promising, additional work is required to confirm the nature of the observed findings (in terms of trait vs. state), to evaluate the effect of glutamate/mGlu5 receptor modulation in BD relative to MDD, and to examine mGlu5 receptor availability in BD individuals across a broader range of symptom severity and mood states. Ultimately, we hope this work inspires future continued research efforts aimed at characterizing unique molecular therapeutic targets and identifying the specific individuals (or endophenotypes) who would most benefit.
Supplementary Material
HIGHLIGHTS.
This study combines analyses of brain function (fMRI) and mGlu5 receptor availability (PET imaging with [18F]FPEB) performed in the same individuals with bipolar disorder (BD) and major depressive disorder (MDD).
Findings provide evidence of relationships between prefrontal mGlu5 availability and brain activation during emotional processing that is specific to BD.
We propose a potential mechanism by which deficient prefrontal mGlu5 in BD contributes to pathological alterations in emotional processing and impulsiveness.
Studies aimed at understanding the unique contribution of mGlu5 receptor to BD pathology and symptomatology, are critical toward the development of targeted, mechanistically informed therapeutic approaches.
Acknowledgments
The authors would like to acknowledge the expert staff at the Yale PET Center and Magnetic Resonance Research Center. We thank the individuals who took part in this study.
Funding
This work was funded by the Dana Foundation (Esterlis), The Brain & Behavior Research Foundation (NARSAD, Esterlis), the National Institutes of Mental Health (R01MH104459, R01MH116657, Esterlis; T32MH014276-47, Asch), and the National Center for Advancing Translational Science (KL2TR001862, Asch).
Abbreviations:
- [18F]FPEB
18F-3-fluoro-5-[(pyridin-3-yl)ethynyl]benzonitrile
- BIS
Barratt Impulsiveness Scale version 11
- BD
bipolar disorder
- BOLD
blood-oxygen-level-dependent
- K E
Cluster extent
- DET
Detection Test
- dlPFC
dorsolateral prefrontal cortex
- FWE
family-wise error
- FD
framewise displacement
- fMRI
functional magnetic resonance imaging
- HAMD
Hamilton Depression Rating Scale, 17-Item
- IDN
Identification Test
- ISLR
International Shopping List Delayed Recall Task
- MDD
major depressive disorder
- MBq
mega Becquerel
- mGlu5
metabotropic glutamate receptor 5
- OFC
orbitofrontal cortex
- PET
positron emission tomography
- HC
psychiatrically healthy comparison
- ROI
region of interest
- TR
repetition time
- V T
volume of distribution
Footnotes
Declaration of Competing Interest
The authors have no conflicts of interest to disclose.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- Adolphs R, 2002. Neural systems for recognizing emotion. Current opinion in neurobiology 12, 169–177. [DOI] [PubMed] [Google Scholar]
- Adolphs R, Damasio H, Tranel D, Cooper G, Damasio AR, 2000. A role for somatosensory cortices in the visual recognition of emotion as revealed by three-dimensional lesion mapping. Journal of neuroscience 20, 2683–2690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asch RH, Hillmer AT, Baldassarri SR, Esterlis I, 2022. The metabotropic glutamate receptor 5 as a biomarker for psychiatric disorders, International Review of Neurobiology. Academic Press. [DOI] [PubMed] [Google Scholar]
- Asch RH, Hillmer AT, Baldassarri SR, Esterlis I, 2023. Chapter Six - The metabotropic glutamate receptor 5 as a biomarker for psychiatric disorders, In: Knackstedt LA, Schwendt M (Eds.), International Review of Neurobiology. Academic Press, pp. 265–310. [DOI] [PubMed] [Google Scholar]
- Baldassarri SR, Asch R, Hillmer AT, Pietrzak RH, DellaGioia N, Esterlis I, Davis MT, 2023. Nicotine Use and Metabotropic Glutamate Receptor 5 (mGluR5) in Individuals with Major Depressive and Post Traumatic Stress Disorders. Chronic Stress. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Besteher B, Gaser C, Nenadić I, 2019. Brain structure and trait impulsivity: A comparative VBM study contrasting neural correlates of traditional and alternative concepts in healthy subjects. Neuropsychologia 131, 139–147. [DOI] [PubMed] [Google Scholar]
- Bi B, Che D, Bai Y, 2022. Neural network of bipolar disorder: Toward integration of neuroimaging and neurocircuit-based treatment strategies. Translational psychiatry 12, 143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blond BN, Fredericks CA, Blumberg HP, 2012. Functional neuroanatomy of bipolar disorder: structure, function, and connectivity in an amygdala–anterior paralimbic neural system. Bipolar disorders 14, 340–355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckner RL, Andrews-Hanna JR, Schacter DL, 2008. The Brain’s Default Network. Annals of the New York Academy of Sciences 1124, 1–38. [DOI] [PubMed] [Google Scholar]
- Cheniaux E, Nardi AE, 2019. Evaluating the efficacy and safety of antidepressants in patients with bipolar disorder. Expert Opinion on Drug Safety 18, 893–913. [DOI] [PubMed] [Google Scholar]
- Damasio AR, Grabowski TJ, Bechara A, Damasio H, Ponto LL, Parvizi J, Hichwa RD, 2000. Subcortical and cortical brain activity during the feeling of self-generated emotions. Nature neuroscience 3, 1049–1056. [DOI] [PubMed] [Google Scholar]
- Davis M, Hillmer A, Holmes S, Pietrzak R, DellaGioia N, Nabulsi N, Matuskey D, Angarita G, Carson R, Krystal J, Esterlis I, 2019a. In vivo evidence for dysregulation of mGluR5 as a biomarker of suicidal ideation. Proc Natl Acad Sci U S A 116, 11490–11495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis MT, Hillmer A, Holmes SE, Pietrzak RH, DellaGioia N, Nabulsi N, Matuskey D, Angarita GA, Carson RE, Krystal JH, 2019b. In vivo evidence for dysregulation of mGluR5 as a biomarker of suicidal ideation. Proceedings of the National Academy of Sciences 116, 11490–11495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeLorenzo C, Gallezot J-D, Gardus J, Yang J, Planeta B, Nabulsi N, Ogden RT, Labaree DC, Huang YH, Mann JJ, Gasparini F, Lin X, Javitch JA, Parsey RV, Carson RE, Esterlis I, 2016. In vivo variation in same-day estimates of metabotropic glutamate receptor subtype 5 binding using [11C]ABP688 and [18F]FPEB. Journal of Cerebral Blood Flow & Metabolism 37, 2716–2727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demeter E, Hernandez-Garcia L, Sarter M, Lustig C, 2011. Challenges to attention: a continuous arterial spin labeling (ASL) study of the effects of distraction on sustained attention. Neuroimage 54, 1518–1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekman P, 1976. Pictures of facial affect. Consulting Psychologists Press. [Google Scholar]
- Elmenhorst D, Mertens K, Kroll T, Oskamp A, Ermert J, Elmenhorst E, Wedekind F, Beer S, Coenen H, Bauer A, 2016. Circadian variation of metabotropic glutamate receptor 5 availability in the rat brain. J Sleep Res Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- Esterlis I, DeBonee S, Cool R, Holmes S, Baldassari SR, Maruff P, Pietrzak RH, Davis MT, 2022. Differential Role of mGluR5 in Cognitive Processes in Posttraumatic Stress Disorder and Major Depression. Chronic Stress 6, 24705470221105804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esterlis I, Holmes SE, Sharma P, Krystal JH, DeLorenzo C, 2018. Metabotropic glutamatergic receptor 5 and stress disorders: Knowledge gained from receptor imaging studies. Biological psychiatry 84, 95–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- First M, Williams J, Karg R, Spitzer R, 2015. Structured clinical interview for DSM-5—Research version (SCID-5 for DSM-5, research version; SCID-5-RV). Arlington, VA: American Psychiatric Association, 1–94. [Google Scholar]
- Förster K, Maliske LZ, Schurz M, Henneberg PM, Dannlowski U, Kanske P, 2023. How do bipolar disease states affect positive and negative emotion processing? Insights from a meta-analysis on the neural fingerprints of emotional processing. Bipolar disorders. [DOI] [PubMed] [Google Scholar]
- Fredrickson J, Maruff P, Woodward M, Moore L, Fredrickson A, Sach J, Darby D, 2010. Evaluation of the usability of a brief computerized cognitive screening test in older people for epidemiological studies. Neuroepidemiology 34, 65–75. [DOI] [PubMed] [Google Scholar]
- Friedman NP, Robbins TW, 2022. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacology 47, 72–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldman DA, Sankar A, Rich A, Kim JA, Pittman B, Constable RT, Scheinost D, Blumberg HP, 2022. A graph theory neuroimaging approach to distinguish the depression of bipolar disorder from major depressive disorder in adolescents and young adults. Journal of Affective Disorders 319, 15–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong J, Wang J, Qiu S, Chen P, Luo Z, Wang J, Huang L, Wang Y, 2020. Common and distinct patterns of intrinsic brain activity alterations in major depression and bipolar disorder: voxel-based meta-analysis. Translational psychiatry 10, 353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greicius MD, Krasnow B, Reiss AL, Menon V, 2003. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis. Proceedings of the national academy of sciences 100, 253–258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton M, 1960. A rating scale for depression. J Neurol Neurosurg Psychiatry 23, 56–62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han K-M, De Berardis D, Fornaro M, Kim Y-K, 2019. Differentiating between bipolar and unipolar depression in functional and structural MRI studies. Progress in Neuro-Psychopharmacology and Biological Psychiatry 91, 20–27. [DOI] [PubMed] [Google Scholar]
- Hirschfeld RM, Cass AR, Holt DC, Carlson CA, 2005. Screening for bipolar disorder in patients treated for depression in a family medicine clinic. The Journal of the American board of family practice 18, 233–239. [DOI] [PubMed] [Google Scholar]
- Holmes S, Girgenti M, Davis M, Pietrzak R, DellaGioia N, Nabulsi N, Matuskey D, Southwick S, Duman R, Carson R, Krystal J, Esterlis I, Group TSBS, 2017. Altered metabotropic glutamate receptor 5 markers in PTSD: in-vivo and postmortem evidence PNAS 114, 8390–8839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes SE, Asch RH, Davis MT, DellaGioia N, Pashankar N, Gallezot JD, Nabulsi N, Matuskey D, Sanacora G, Carson RE, Blumberg HP, Esterlis I, 2023. Differences in Quantification of the Metabotropic Glutamate Receptor 5 Across Bipolar Disorder and Major Depressive Disorder. Biol Psychiatry 93, 1099–1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong W, Zhang C, Xing MJ, Wu ZG, Wang ZW, Chen J, Yuan CM, Su Y-S, Hu YY, Cao L, 2016. Contribution of long duration of undiagnosed bipolar disorder to high frequency of relapse: A naturalistic study in China. Comprehensive Psychiatry 70, 77–81. [DOI] [PubMed] [Google Scholar]
- Ino H, Honda S, Yamada K, Horita N, Tsugawa S, Yoshida K, Noda Y, Meyer JH, Mimura M, Nakajima S, 2022. Glutamatergic Neurometabolite Levels in Bipolar Disorder: A Systematic Review and Meta-analysis of Proton Magnetic Resonance Spectroscopy Studies. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging. [DOI] [PubMed] [Google Scholar]
- Inuggi A, Sanz-Arigita E, González-Salinas C, Valero-García AV, García-Santos JM, Fuentes LJ, 2014. Brain functional connectivity changes in children that differ in impulsivity temperamental trait. Front Behav Neurosci 8, 156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston JAY, Wang F, Liu J, Blond BN, Wallace A, Liu J, Spencer L, Cox Lippard ET, Purves KL, Landeros-Weisenberger A, Hermes E, Pittman B, Zhang S, King R, Martin A, Oquendo MA, Blumberg HP, 2017. Multimodal Neuroimaging of Frontolimbic Structure and Function Associated With Suicide Attempts in Adolescents and Young Adults With Bipolar Disorder. The American journal of psychiatry 174, 667–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones MS, Zhu Z, Bajracharya A, Luor A, Peelle JE, 2022. A Multi-Dataset Evaluation of Frame Censoring for Motion Correction in Task-Based fMRI. Apert Neuro 2, 1–25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Judd LL, Akiskal HS, Schettler PJ, Endicott J, Maser J, Solomon DA, Leon AC, Rice JA, Keller MB, 2002. The long-term natural history of the weekly symptomatic status of bipolar I disorder. Archives of general psychiatry 59, 530–537. [DOI] [PubMed] [Google Scholar]
- Keramatian K, Pinto JV, Schaffer A, Sharma V, Beaulieu S, Parikh SV, Yatham LN, 2022. Clinical and demographic factors associated with delayed diagnosis of bipolar disorder: Data from Health Outcomes and Patient Evaluations in Bipolar Disorder (HOPE-BD) study. Journal of Affective Disorders 296, 506–513. [DOI] [PubMed] [Google Scholar]
- Kerestes R, Bhagwagar Z, Nathan PJ, Meda SA, Ladouceur CD, Maloney K, Matuskey D, Ruf B, Saricicek A, Wang F, Pearlson GD, Phillips ML, Blumberg HP, 2012. Prefrontal cortical response to emotional faces in individuals with major depressive disorder in remission. Psychiatry Research: Neuroimaging 202, 30–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroes MCW, Dunsmoor JE, Hakimi M, Oosterwaal S, collaboration NP, Meager MR, Phelps EA, 2019. Patients with dorsolateral prefrontal cortex lesions are capable of discriminatory threat learning but appear impaired in cognitive regulation of subjective fear. Social Cognitive and Affective Neuroscience 14, 601–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kropf E, Syan SK, Minuzzi L, Frey BN, 2019. From anatomy to function: the role of the somatosensory cortex in emotional regulation. Braz J Psychiatry 41, 261–269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kupka R, Altshuler L, Nolen W, Suppes T, Luckenbaugh D, Leverich G, Frye M, Keck PJ, McElroy S, Grunze H, Post R, 2007. Three times more days depressed than manic or hypomanic in both bipolar I and bipolar II disorder. Bipolar disorders 9, 531–535. [DOI] [PubMed] [Google Scholar]
- Lamm C, Rütgen M, Wagner IC, 2019. Imaging empathy and prosocial emotions. Neuroscience letters 693, 49–53. [DOI] [PubMed] [Google Scholar]
- Liu J, Blond BN, van Dyck LI, Spencer L, Wang F, Blumberg HP, 2012. Trait and state corticostriatal dysfunction in bipolar disorder during emotional face processing. Bipolar disorders 14, 432–441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu TY, Chen YS, Su TP, Hsieh JC, Chen LF, 2014. Abnormal early gamma responses to emotional faces differentiate unipolar from bipolar disorder patients. Biomed Res Int 2014, 906104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park E, Sullivan JM, Planeta B, Gallezot J-D, Lim K, Lin S-F, Ropchan J, McCarthy TJ, Ding Y-S, Morris ED, 2015. Test–retest reproducibility of the metabotropic glutamate receptor 5 ligand [18F] FPEB with bolus plus constant infusion in humans. European Journal of Nuclear Medicine and Molecular Imaging 42, 1530–1541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patel S, Hamill TG, Connolly B, Jagoda E, Li W, Gibson RE, 2007. Species differences in mGluR5 binding sites in mammalian central nervous system determined using in vitro binding with [18F] F-PEB. Nuclear medicine and biology 34, 1009–1017. [DOI] [PubMed] [Google Scholar]
- Peters AT, West AE, Eisner L, Baek J, Deckersbach T, 2016. The Burden of Repeated Mood Episodes in Bipolar I Disorder: Results From the National Epidemiological Survey on Alcohol and Related Conditions. J Nerv Ment Dis 204, 87–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigues SM, Bauer EP, Farb CR, Schafe GE, LeDoux JE, 2002. The group I metabotropic glutamate receptor mGluR5 is required for fear memory formation and long-term potentiation in the lateral amygdala. Journal of Neuroscience 22, 5219–5229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolls ET, Huang C-C, Lin C-P, Feng J, Joliot M, 2020. Automated anatomical labelling atlas 3. Neuroimage 206, 116189. [DOI] [PubMed] [Google Scholar]
- Scheibner HJ, Bogler C, Gleich T, Haynes J-D, Bermpohl F, 2017. Internal and external attention and the default mode network. NeuroImage 148, 381–389. [DOI] [PubMed] [Google Scholar]
- Schumer MC, Chase HW, Rozovsky R, Eickhoff SB, Phillips ML, 2023. Prefrontal, parietal, and limbic condition-dependent differences in bipolar disorder: a large-scale meta-analysis of functional neuroimaging studies. Molecular psychiatry, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen H, Zhang L, Xu C, Zhu J, Chen M, Fang Y, 2018. Analysis of Misdiagnosis of Bipolar Disorder in An Outpatient Setting. Shanghai Arch Psychiatry 30, 93–101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shih C-W, Chang C. h., 2021. Medial or lateral orbitofrontal cortex activation during fear extinction differentially regulates fear renewal. Behavioural Brain Research 412, 113412. [DOI] [PubMed] [Google Scholar]
- Siegel-Ramsay JE, Bertocci MA, Wu B, Phillips ML, Strakowski SM, Almeida JRC, 2022. Distinguishing between depression in bipolar disorder and unipolar depression using magnetic resonance imaging: a systematic review. Bipolar disorders 24, 474–498. [DOI] [PubMed] [Google Scholar]
- Singh T, Rajput M, 2006. Misdiagnosis of bipolar disorder. Psychiatry (Edgmont) 3, 57–63. [PMC free article] [PubMed] [Google Scholar]
- Sreenivas S, Boehm SG, Linden DEJ, 2012. Emotional faces and the default mode network. Neuroscience Letters 506, 229–234. [DOI] [PubMed] [Google Scholar]
- Stanford MS, Mathias CW, Dougherty DM, Lake SL, Anderson NE, Patton JH, 2009. Fifty years of the Barratt Impulsiveness Scale: An update and review. Personality and individual differences 47, 385–395. [Google Scholar]
- Strakowski SM, Adler CM, Almeida J, Altshuler LL, Blumberg HP, Chang KD, DelBello MP, Frangou S, McIntosh A, Phillips ML, Sussman JE, Townsend JD, 2012. The functional neuroanatomy of bipolar disorder: a consensus model. Bipolar disorders 14, 313–325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sullivan JM, Lim K, Labaree D, Lin S. f., McCarthy TJ, Seibyl JP, Tamagnan G, Huang Y, Carson RE, Ding Y-S, 2013. Kinetic analysis of the metabotropic glutamate subtype 5 tracer [18F] FPEB in bolus and bolus-plus-constant-infusion studies in humans. Journal of Cerebral Blood Flow & Metabolism 33, 532–541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson TAC, Wilson PH, Snyder PJ, Pietrzak RH, Darby D, Maruff P, Buschke H, 2011. Sensitivity and Test–Retest Reliability of the International Shopping List Test in Assessing Verbal Learning and Memory in Mild Alzheimer’s Disease. Archives of Clinical Neuropsychology 26, 412–424. [DOI] [PubMed] [Google Scholar]
- Weissman DH, Roberts KC, Visscher KM, Woldorff MG, 2006. The neural bases of momentary lapses in attention. Nat Neurosci 9, 971–978. [DOI] [PubMed] [Google Scholar]
- Young AH, 2009. Bipolar disorder: diagnostic conundrums and associated comorbidities. J Clin Psychiatry 70, e26–e26. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.