

Abstract
Populations of anadromous brown trout, also known as sea trout, have suffered recent marked declines in abundance due to multiple factors, including climate change and human activities. While much is known about their freshwater phase, less is known about the species' marine feeding migrations. This situation is hindering the effective management and conservation of anadromous trout in the marine environment. Using a panel of 95 single nucleotide polymorphism markers we developed a genetic baseline, which demonstrated strong regional structuring of genetic diversity in trout populations around the English Channel and adjacent waters. Extensive baseline testing showed this structuring allowed high‐confidence assignment of known‐origin individuals to region of origin. This study presents new data on the movements of anadromous trout in the English Channel and southern North Sea. Assignment of anadromous trout sampled from 12 marine and estuarine localities highlighted contrasting results for these areas. The majority of these fisheries are composed predominately of stocks local to the sampling location. However, there were multiple cases of long‐distance movements of anadromous trout, with several individuals originating from rivers in northeast England being caught in the English Channel and southern North Sea, in some cases more than 1000 km from their natal region. These results have implications for the management of sea trout in inshore waters around the English Channel and southern North Sea.
Keywords: brown trout, genetic stock identification, management, mixed‐stock fishery, SNP
1. INTRODUCTION
Brown trout (Salmo trutta L.) is a ubiquitous fish species found naturally over much of Europe, North Africa and western Asia in a wide range of river types (Kershner et al., 2019). Across this range, brown trout show a great range of morphologies (Ferguson & Prodöhl, 2022; Verspoor et al., 2019) and genetic variants (Bernatchez, 2001; Ferguson, 1989; Ferguson & Taggart, 1991; King et al., 2016; Quéméré et al., 2016; Vilas et al., 2010). These genetic variants can often be highly localized, with distinct patterns of genetic variation between fish inhabiting different parts of a catchment and/or adjacent rivers (Bekkevold et al., 2020; Bouza et al., 1999; Ferguson, 1989; Griffiths et al., 2009; King et al., 2020). These levels of significant genetic separation allow the recognition of distinct populations and reflect both the phylogeographic history of the species (Bernatchez, 2001; Cortey et al., 2009; McKeown et al., 2010) and more recent events that have acted to restrict or eliminate gene flow, for example, the construction of dams and weirs (King et al., 2020; Osmond et al., 2024), leading to the emergence of distinct genetic signatures due to drift and adaptation. In turn, these distinct populations can be used as operational taxonomic units for the assessment of straying (King et al., 2016) in anadromous individuals (hereafter referred to as sea trout) and for tracing the at‐sea movements of fish (Bekkevold et al., 2021; Koljonen et al., 2014; Prodöhl et al., 2017). Both are achieved by assigning sea trout back to their population or region of origin based on similarities between the genotypes of the migratory form (sea trout) and the population genetic signature of resident trout in different candidate rivers/regions of origin.
The English Channel is one of the busiest waterways in Europe for both commercial and recreational fishing, cross‐Channel trade and as a navigation route from the Atlantic to the southern North Sea and the Baltic (Glegg et al., 2015). Along its length several major rivers flow into it, including the Seine and, historically, it forms the route of the palaeo‐Channel River (Lericolais et al., 2003). Thus, many of the rivers of this region have a common history, beginning as tributaries of the much larger ancient Channel River and sharing riverine geologies. Similarly, the trout of this region have a shared history dating from before the last glacial maximum (Bernatchez, 2001; McKeown et al., 2010) and have been affected by rising sea levels after the last glacial maximum, leading to the separation of many former Channel River tributaries into distinct catchments.
More recently, populations of both trout and Atlantic salmon have been severely affected by human‐related activities, including targeted estuarine net fisheries, changes to river navigability and barriers to upstream movement (weirs, dams), point‐source and diffuse pollution, loss of spawning habitat and many stocking and translocation events (Losee et al., 2024; Nevoux et al., 2019). This combination of historic and contemporary factors has shaped the present mosaic of genetic groupings of trout in rivers on both sides of the English Channel and in the southern North Sea (King et al., 2016, 2020; Quéméré et al., 2016). Research has been able to inform on the impact of many of the factors driving population level variation in trout, particularly those acting in the freshwater phase of the trout lifecycle (King et al., 2020; Paris et al., 2015). However, trout –unlike salmon– exhibit a continuum of life history variation from fully resident through freshwater migration to fully anadromous individuals (Ferguson et al., 2019).
There is a long history of studies investigating the marine distribution of different stocks and the mixed‐stock nature of marine fisheries in anadromous salmonids at different spatial scales (Cormack & Skalski, 1992; Tucker et al., 2009). Recently, there has been extensive investigation of the marine distribution of different Atlantic salmon stocks and the mixed‐stock nature of targeted marine fisheries assessed using genetic baselines (Bradbury et al., 2015; Gilbey et al., 2017, 2021); to date, however, there have been only a limited number of similar studies on sea trout (Bekkevold et al., 2021; Koljonen et al., 2014; Prodöhl et al., 2017). Unlike Atlantic salmon, however, anadromous trout are thought to feed more locally to their natal rivers (Jonsson & Jonsson, 2014; Malcolm et al., 2010; Potter et al., 2017), rather than migrating long distances to offshore feeding grounds in the north Atlantic (Gilbey et al., 2017, 2021). Nonetheless, several tagging and tracking studies have reported highly variable degrees of movement, including longer migrations of limited numbers of individuals (Hawley et al., 2024; Kallio‐Nyberg et al., 2002; Malcolm et al., 2010; Potter et al., 2017). Additionally, distinct regional differences in migration patterns have been reported (Potter et al., 2017).
With anadromous salmonids being subject to multiple stressors, both in their freshwater and marine environments, many species have suffered marked declines in abundance over recent decades (ICES, 2013). While management and conservation measures for trout in freshwater, including knowledge of when and where to implement such measures, are now relatively well understood, an understanding of how, when and where to implement protection measures for trout in the marine environment is much less advanced. Similar to Atlantic salmon (Gillson et al., 2022), within the marine environment, stressors of sea trout include aquaculture, coastal developments (i.e. tidal lagoons, inshore and offshore wind farms), and by‐catch in non‐target fisheries (Nevoux et al., 2019; Thorstad et al., 2016). Given the importance of anadromous individuals to the resilience of trout populations (Goodwin et al., 2016), effective conservation and management of such populations requires extensive information on species biology, behaviour, life cycle and the challenges they face at different life history stages (Nevoux et al., 2019; Whelan et al., 2017), including knowledge of when and where sea trout go during their marine migrations (O'Sullivan et al., 2022; Thorstad et al., 2016). Of particular relevance is the incidence of individuals taken as by‐catch in non‐target marine fisheries; again, data on this specific to sea trout are very poor (Elliott et al., 2023).
In this study, we constructed a genetic baseline for trout sampled from 107 rivers around the English Channel, southern Irish Sea and southern North Sea based on 95 single nucleotide polymorphism (SNP) markers. Our objectives were (1) to catalogue the structuring of, and genetic variation between, trout populations in these areas, (2) to assess the scale at which reliable assignment to the baseline could be achieved using leave‐one‐out analyses and genotypes from known‐origin individuals, and (3) to investigate the stock composition of sea trout sampled from multiple marine and estuarine locations along the English Channel, Bristol Channel and southern North Sea coasts of England, France and the Netherlands.
2. MATERIALS AND METHODS
2.1. Study species
Across their native range, brown trout are distributed from North Africa to northern Russia and from Iceland east to the Caspian Sea (Jonsson & Jonsson, 2009). The species is facultatively anadromous and is typified by complex variation in life history, both within and between populations, from fully resident, through partial migration within freshwater systems, to fully anadromous individuals which spend time (ranging from a few days to upwards of 2 years) in the marine environment (Thorstad et al., 2016). Anadromous populations are found from northern Portugal to the White Sea, the Baltic Sea and Iceland (Jonsson & Jonsson, 2009). Anadromous and resident individuals are typically found in the same rivers, often share spawning sites and are fully interfertile (Goodwin et al., 2016), with several studies finding no neutral genetic differences between resident and migratory individuals within the same river (Charles et al., 2005; Goodwin et al., 2016).
The decision to migrate is a threshold trait. A genetically determined propensity to migrate (Lemopoulos et al., 2018) interacts with environmental factors (Nevoux et al., 2019) and physiological condition to ultimately control the decision to migrate or stay resident (Ferguson et al., 2019). The benefits of anadromy generally involve increased feeding opportunities in the marine environment. This leads to a larger body size of anadromous individuals compared to resident trout with a resultant increased fecundity (Goodwin et al., 2016).
For partially migrating species the advantages of anadromy differentially affect the sexes. Female fecundity is strongly dependent on body size (Goodwin et al., 2016; Thériault et al., 2007), while male reproductive success is limited by availability of mates (Thériault et al., 2007). As a consequence of these differences in selective factors affecting sex‐related fecundity, the majority of anadromous trout are female (Le Cren, 1985).
2.2. Sample collection
For baseline construction, adipose finclip or scale samples from juvenile resident trout were obtained from various sources (Table S1). Samples were collected during routine electrofishing surveys in the UK and Ireland by the Environment Agency in England, Inland Fisheries Ireland or, specifically, by the SAMARCH project team (www.samarch.org) and in France by INRAE U3E Unit, Office Français de la Biodiversité, Bretagne Grands Migrateurs, Seinormigr and Fédération Départmentale de Pêche et de Protection du Milieu Aquatique 14, 22, 27, 29, 35, 50, 62, 76 and 80 as part of inventory surveys. Samples from two Danish rivers consisted of mature adults collected on spawning sites by a team from the Technical University of Denmark—details in Bekkevold et al. (2020).
Scale and finclip samples from 398 sea trout were obtained from commercial and recreational fisheries from English, French and Dutch coastal and estuarine areas (Appendix S1 and Figure S1). These collections represent a range of samples caught in targeted commercial salmonid netting activities (i.e. TT and EAN), as by‐catch in commercial fisheries targeting non‐salmonids (i.e. RYE), recreational fisheries (i.e. OUS and MER) or targeted sampling (i.e. KIM and COR) undertaken specifically for the SAMARCH research project (www.samarch.org). Details of these fisheries are given in Appendix S1.
2.3. Molecular methods
Genomic DNA was extracted using the HotSHOT method of Truett et al. (2000) for southern UK and Irish samples, Omega Biotek E.Z.N.A. kits for NE English and Danish samples and NucleoSpin® 96 Tissue kits (Macherey‐Nagel) for French samples. All individuals were genotyped at 95 biallelic single nucleotide polymorphism (SNP) loci (Osmond et al., 2023) on the Fluidigm EP1 Genotyping System using 96.96 Dynamic Genotyping Arrays and scored using the Fluidigm SNP Genotyping analysis software. Genotype plots of each locus were manually inspected for quality of individual genotyping and clustering. Individual points that fell outside of the heterozygote or homozygote genotype clusters were considered to have poor quality data and left uncalled for that locus (Clemento et al., 2011). Individual genotypes with more than five uncalled loci were excluded from subsequent analyses. Each run included two positive (individuals of known genotype) and two negative (no DNA) controls.
2.4. Data quality assurance
Juvenile salmonid populations can sometimes be characterised by large numbers of closely related individuals, i.e. full‐sibs (Goodwin et al., 2016), the presence of which can lead to biases in the inference of population structure (Anderson & Dunham, 2008) and genetic stock identification (Östergren et al., 2020). To assign sibship within each sample of fish we used a maximum‐likelihood method, implemented in COLONY v2.0 (Jones & Wang, 2010). Settings were: high precision medium length run, assuming both male and female polygamy without inbreeding and a conservative 0.5% error rate for both scoring error rate and allelic dropout rate. To check for consistency, analyses were run twice using different random number seeds. Full sibs were trimmed from the data set using Waples and Anderson's (2017) Yank‐2 method—all but two random members of families with three or more individuals were removed.
Linkage disequilibrium (LD) between all pairs of loci within each population was tested using GENEPOP v3.4 (Raymond & Rousset, 1995). Significance was estimated using a Markov chain method using default parameters (1000 de‐memorizations, 100 batches and 1000 iterations). False Discovery Rate (FDR; Benjamini & Hochberg, 1995) was used to correct significance levels for multiple comparisons—https://www.multipletesting.com (Menyhart et al., 2021). Using GenoDive v3.03 (Meirmans, 2020), deviations from Hardy–Weinberg Equilibrium (HWE) for each locus and population was assessed using Nei's (1987) heterozygosity‐based G IS estimator with significance based on 999 permutations.
2.5. Basic measures of genetic diversity
GenoDive v3.03 (Meirmans, 2020) was used to calculate observed (H O) and unbiased expected heterozygosity (H E) and Weir and Cockerham's (1984) estimator of F ST were calculated with significance of F ST values determined using 999 bootstrap replicates.
2.6. Population genetic structure and identification of reporting groups
Depending on location, salmonid fisheries often target mixed stocks of fish with ‘stocks’ comprising multiple, geographically proximate and genetically similar rivers (Moran & Anderson, 2019). To investigate population genetic structuring of trout populations, we performed two analyses. Firstly, we used STRUCTURE v2.3.4 (Pritchard et al., 2000) which implements a Bayesian‐based Markov Chain Monte Carlo (MCMC) model‐based clustering method to jointly delineate K, the number of partitions of the data set and q, the proportion of each individual's genome originating from each of the K partitions. STRUCTURE was run with a burn‐in of 100,000 iterations followed by 250,000 iterations with the number of inferred populations (K) ranging from 1 to 15. Ten independent runs were performed using the admixture model with correlated allele frequencies and not using the population of origin information as a prior. We used the ΔK method of Evanno et al. (2005) to determine the most likely number of clusters. Hierarchical analyses were performed, based on the ΔK results for the full data set, to identify finer‐levels of structure. Where the number of rivers in a hierarchical analysis was less than 15, the maximum K was set at N rivers + 1. POPHELPER v1.0.6 (Francis, 2017) was used to calculate ΔK and to visualize the consensus data after alignment of multiple runs at optimum K values using CLUMPP v1.1.2 (Jakobsson & Rosenberg, 2007).
A neighbour‐joining dendrogram based on Cavalli‐Sforza and Edwards (1967) chord distance (D CE) was used to identify population‐level genetic structure. The dendrogram was constructed and visualized using POPULATIONS v1.2.32 (Langella, 1999) and MEGA v6 (Tamura et al., 2013), respectively. Baseline reporting groups, upon which subsequent assignments would be based, were identified using a combination of the STRUCTURE and neighbour‐joining analyses.
2.7. Genetic stock identification analyses
We employed two widely utilized pieces of assignment software for the mixed stock analyses (MSA) and individual assignment (IA) of sea trout caught in estuarine and marine waters to both individual river and reporting groups as defined in the population structure analyses (see Section 3). cBayes (Neaves et al., 2005) implements the Bayesian procedures of Pella and Masuda (2001). For stock composition estimation, eight 50,000‐iteration Markov Chain Monte Carlo (MCMC) chains were run, with initial values set at 0.9 for each chain for different samples. Means and 95% confidence intervals of the estimated stock contributions were determined from the combined final 1000 iterations from each chain. RUBIAS uses a Bayesian conditional genetic stock identification model to provide mixture proportion estimates and assign individuals to population/stock of origin (Moran & Anderson, 2019). Assignment proportions and their 95% credible intervals were generated using the MCMC method based on 100,000 sweeps following a burn‐in of 10,000 sweeps.
We used two tests to assess the accuracy of assignments to our SNP baseline. Firstly, Leave‐One‐Out (LOO) analysis, as implemented in RUBIAS, was used to assess assignment accuracy and efficiency. Secondly, we assessed the mixed‐stock and individual assignment of 436 individuals of known origin from 25 baseline rivers using both cBayes and RUBIAS. Full details of these tests and their results are given in Appendix S1.
Mixed stock analysis and individual assignment to reporting group for the 12 marine and estuarine derived collections of sea trout were estimated using both cBayes and RUBIAS. Analyses were run using the conditions given above.
Least‐cost migration distances for each marine‐caught sea trout were calculated using the marmap R package (Pante & Simon‐Bouhet, 2013). For the East Anglian and Dutch fishery samples where fish were sampled from multiple locations, we took the approximate midpoint between the extreme sampling locations on each stretch of coastline. For regional level assignments, we calculated the minimum, maximum and average distance that fish could have migrated from a river of origin within a reporting group to the marine sampling location.
3. RESULTS
3.1. Data quality
A total of 4085 individuals were genotyped at 95 SNP loci. Comparison of genotypes from repeated samples gave an error rate of 0.0014% (46 mismatches from 31,920 allele calls). In total, 98 individuals were removed after failing to be genotyped at ≥6 loci. The number of full‐sib families per baseline sample ranged from 0 to 9 (mean families per river = 2.48). The maximum number of individuals in any full‐sib family was 10. In total, 125 full‐sib individuals were removed following analysis with the program COLONY. The final dataset comprised 3067 baseline, 436 known origin and 371 marine‐/estuarine‐caught sea trout.
After FDR correction, 32 pairs of loci (out of a total of 477,755 pairwise comparisons) were in significant linkage across the 107 baseline samples. There were 354 significant deviations from HWE (out of a total of 10,165 baseline sample/locus combinations). As none of these significant results showed any consistent patterns across loci or baseline samples, all loci and samples were retained for further analyses.
3.2. Population genetic structure
Global F ST was 0.109 (p = 0.001). Pairwise F ST values ranged from zero (p = 0.512) between the East Looe and West Looe rivers in southern Cornwall to 0.266 (p = 0.001) between the Horn (Bretagne) and Sow (southeast Ireland) rivers.
The results of the STRUCTURE and neighbour‐joining analyses were in broad agreement with both identifying a high degree of regional structuring within the 107 baseline rivers, with neighbouring rivers being genetically more similar to each other, sometimes over long stretches of coastline. The neighbour‐joining analysis identified 13 geographical structured groups of rivers (Figure 1) with the number of rivers per group ranging from two from Denmark (DENMARK) to 20 from Devon & Cornwall (DEVCORN). STRUCTURE identified K = 2 (ΔK = 122.4) as the most likely partition of the full dataset, splitting the rivers into western and eastern groupings (Figure 2). Subsequent hierarchical analyses identified further subdivision within both the western and eastern groups and broadly recovered the same population groupings as found in the neighbour‐joining analysis (Figure 2). STRUCTURE also highlighted that the distinction between genetic groups tended to be geographically limited, for example, in Britain between the Hampshire Basin and southeast English rivers (Figure 2) and in France between the rivers of Lower and Upper Normandy (Figure 2).
FIGURE 1.
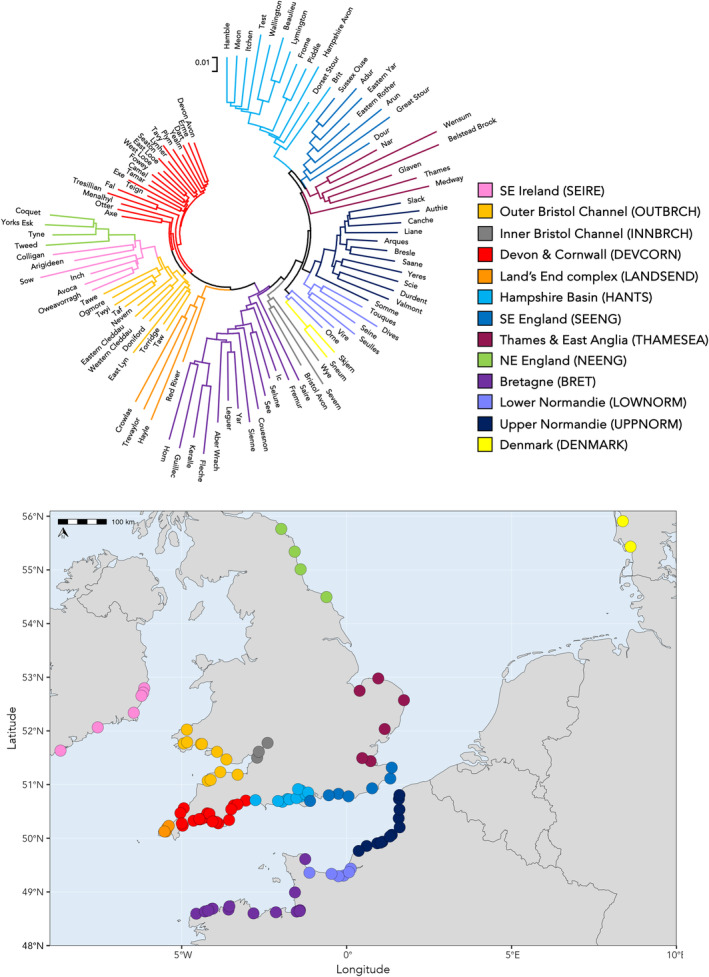
Unrooted neighbour‐joining (NJ) dendrogram, based on Cavalli‐Sforza and Edwards' chord distance (D CE), showing relationships between the 107 resident trout populations sampled for the SNP baseline. Branches are colour coded by reporting group. The map gives the location of the mouth of each sampled river with coloured points giving reporting group membership as determined the NJ dendrogram. Full sample site details are given in Table S1.
FIGURE 2.
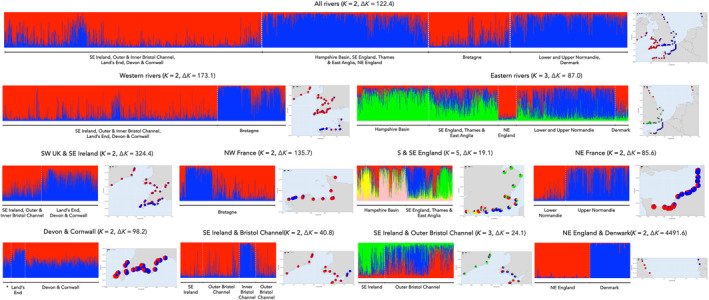
Results of the hierarchical STRUCTURE analysis for the 107 resident trout baseline rivers. Results of each STRUCTURE analysis are shown as bar plots with vertical columns represent the assignment probabilities of individuals to each of the K inferred clusters. For clarity, results are plotted by reporting groups rather than individual rivers. Maps show the location of each sampled river with pie charts giving the population‐level assignment to each genetic cluster. Plots of ΔK values for each analysis are given in Figure S5.
3.3. Baseline testing
Based on the regional structuring identified in the STRUCTURE and neighbour‐joining analyses, we identified 13 groups of rivers (hereafter referred to as reporting groups), with the addition of a group of French hatchery populations, as the basis for the baseline testing and assignment of sea trout. Results of the initial baseline testing are given in detail in Appendix S1. Briefly, LOO analysis found generally high levels (>85%) of assignment accuracy and efficiency to reporting group (Figure S2). Conversely, assignment success to individual rivers was highly variable. For some rivers assignment had very high (>95%) accuracy and efficiency, i.e. SEV, WEN, TYN (Figure S3), however, most rivers demonstrated much lower assignment success. For example, for many of the rivers in the DEVCORN reporting group accuracy and efficiency of assignment to an individual river was below 50% (Figure S3). Mixed‐stock and individual assignment of the known‐origin collections showed similar trends to the LOO analysis, with collections assigning strongly to their region of origin and highly variable success of assignment to river of origin (Figure S4, Tables S2 and S3). There were also clear differences in the ability of RUBIAS and cBayes to correctly assign collections and individual fish to their rivers of origin (Figure S4). Based on these results, here we report only regional mixed‐stock and individual assignments for the 12 marine‐ and estuarine‐caught collections determined using cBayes. However, cBayes MSA and IA results of assignment to river of origin and RUBIAS results for both regional and river MSA and IA are presented in Tables S4 and S5.
3.4. Assignment of marine and estuarine collections
Assignment of the 12 collections of marine and estuarine sampled sea trout showed contrasting patterns of assignment. The four estuarine collections (TT, TAM, PLH and OUS, Figures 3 and 4) showed very little evidence of mixing of fish from different reporting groups, with each collection being dominated by migratory fish from the same reporting group as that to which the sampled estuaries belonged (Figures 3 and 4, Tables S4 and S5). For example, the majority of sea trout sampled in the Taw/Torridge estuary belonged to the Outer Bristol Channel (OUTBRCH) reporting group with a single individual assigning strongly to the DEVCORN reporting group (Figures 3 and 4, Tables S4.09 and S5.09). Likewise, 29 of 30 fish sampled in a recreational sea trout rod fishery in the tidal reaches of the Sussex Ouse, a member of the SE England (SEENG) reporting group, assigned to that reporting group. The remaining individual had strongest assignment to the NE England (NEENG) reporting group (Table S5.12).
FIGURE 3.
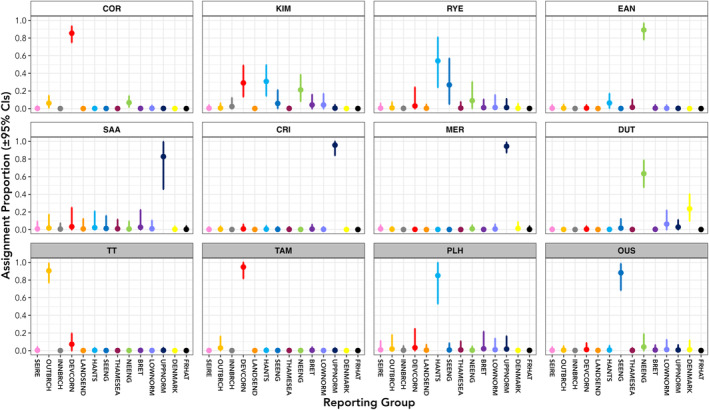
Mean estimated stock composition assigned to reporting group of origin, with 95% confidence intervals, for eight marine (white chart header) and four estuarine (grey chart header) collections of anadromous trout. Reporting regions are colour coded as given in Figure 1. Marine collection abbreviations: COR, southern Cornwall targeted netting; CRI, Criel‐sur‐Mer recreational beach nets; DUT, Dutch commercial fishery by‐catch; EAN, East Anglian drift‐net fishery; KIM, Kimmeridge Bay targeted netting; MER, Mers‐les‐Bains and Le Tréport recreational beach nets; RYE, Rye Harbour commercial net fishery; SAA, Saâne illegal nets. Estuarine collection abbreviations: OUS, Sussex Ouse estuary recreational rod fishery; PLH, Poole Harbour; TAM, River Tamar tidal limit fish trap; TT, Taw/Torridge shared estuary. Reporting group abbreviations: BRET, Bretagne; DENMARK, Denmark; DEVCORN, Devon and Cornwall; FRHAT, French hatchery populations; HANTS, Hampshire Basin; INNBRCH, inner Bristol Channel; LANDSEND, Land's End complex; LOWNORM, Lower Normandie; NEENG, northeast England; OUTBRCH, outer Bristol Channel; SEENG, southeast England; SEIRE, southeast Ireland; THAMESEA, River Thames and East Anglia; UPPNORM, Upper Normandie.
FIGURE 4.
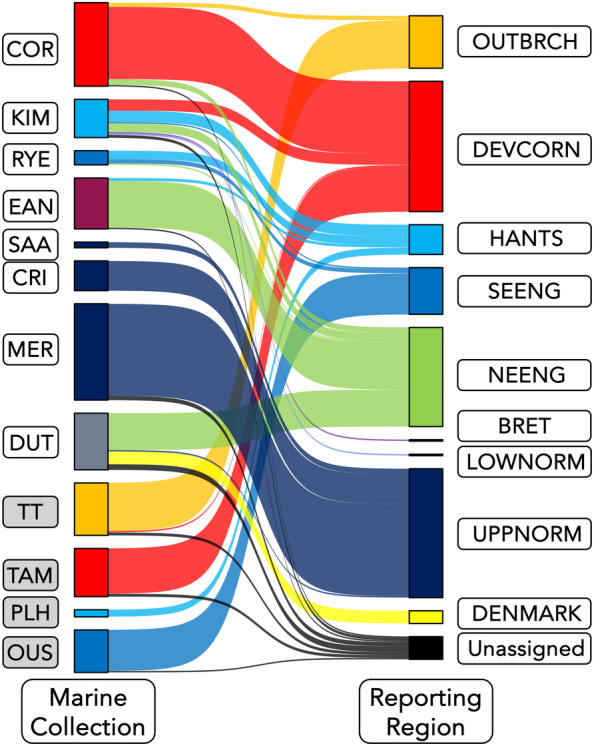
Sankey plot showing individual assignment of marine and estuarine caught anadromous trout to reporting region of origin. Marine and estuarine collections are colour coded by the reporting region they are located in while reporting regions are colour coded as given in Figure 1. Individuals were considered ‘Unassigned’ if the maximum probability of assignment to any reporting group was <0.7. Marine collection abbreviations: COR, southern Cornwall targeted netting; CRI, Criel‐sur‐Mer recreational beach nets; DUT, Dutch commercial fishery by‐catch; EAN, East Anglian drift‐net fishery; KIM, Kimmeridge Bay targeted netting; MER, Mers‐les‐Bains and Le Tréport recreational beach nets; RYE, Rye Harbour commercial net fishery; SAA, Saâne illegal nets. Estuarine collection abbreviations: OUS, Sussex Ouse estuary recreational rod fishery; PLH, Poole Harbour; TAM, River Tamar tidal limit fish trap; TT, Taw/Torridge shared estuary. Reporting group abbreviations: BRET, Bretagne; DENMARK, Denmark; DEVCORN, Devon and Cornwall; HANTS, Hampshire Basin; LOWNORM, Lower Normandie; NEENG, northeast England; OUTBRCH, outer Bristol Channel; SEENG, southeast England; UPPNORM, Upper Normandie.
The marine collections were more variable in their assignments to reporting group (Figures 3 and 4). Similar to the estuarine collections, some of the marine collections showed minimal variation in assignment outside of their expected reporting groups. For instance, sea trout in the collections from SAA, CRI and MER, which were caught in French waters in nets set close to the shore at the mouths of the Saâne, Yères and Bresle rivers, respectively, caught only fish from the Upper Normandie (UPPNORM) reporting group (Figures 3 and 4). Likewise, in southwest England the COR sea trout samples were dominated by fish from the DEVCORN reporting group, with minor contributions from both OUTBRCH and NEENG rivers.
By contrast, the sea trout caught at KIM and RYE in southern England were more variable in their origins. Adult fish from six regions were caught at KIM, originating mainly from the three southern English reporting groups (DEVCORN, Hampshire Basin (HANTS) and SEENG). However, fish from Bretagne (BRET), Lower Normandie (LOWNORM) and NEENG were also sampled here (Figures 3 and 4), while sea trout originating from the HANTS, SEENG and NEENG regions were sampled at RYE.
The two collections from the southern North Sea (EAN and DUT) were dominated by fish originating from the NEENG reporting group, with a significant contribution of trout from Danish rivers to the DUT samples. There were only minor contributions from English Channel reporting groups to these collections, with two fish of HANTS origin caught in the EAN nets and a single UPPNORM sea trout caught in Dutch waters (Figure 4).
3.5. Migration distances
Migration distances between the 12 marine and estuarine collections and the rivers of each reporting group are presented in Table S6. This shows that the majority of sea trout were on average captured in close proximity to their natal rivers. For instance, the average capture distance for HANTS fish caught at KIM was 63.6 km. However, there are instances of very long‐distance movements of sea trout, especially for those originating in NEENG rivers. THE NEENG fish caught at KIM and COR were on average 800 and 965 km from their natal rivers (Table S6).
4. DISCUSSION
Here we present an extensive SNP‐based genetic baseline for trout from English Channel and surrounding rivers, describing extensive, regional‐based genetic structuring that allows high‐confidence assignment of marine‐caught sea trout to their region of origin.
4.1. Trout populations show strong regional genetic structure
The strong regional structuring of the trout populations in rivers screened here reiterates a pattern of distinct genetic groupings spanning sometimes long stretches of coastline and commonly observed in many anadromous salmonid species (Beacham et al., 2020, 2021; Bradbury et al., 2015; Koljonen et al., 2014; Layton et al., 2020; Small et al., 2015). At the broadest scale, populations were split into two distinct eastern and western groups, with the split corresponding approximately with the Isle of Portland on the English coast of the Channel and the Cotentin Peninsula on the French coast. The Cotentin Peninsula and the relatively shallow waters to the north of the peninsula have previously been identified as a significant feature in the genetic structuring of a variety of marine organisms (Dauvin, 2012), including northern French trout populations (Quéméré et al., 2016).
Within each of the two main trout population groupings finer‐scales of genetic structuring were also found. Three genetic groups of trout were identified in rivers entering the Channel on both the English and French Channel coasts. These corresponded with the three main geological zones existing on both sides of the Channel and it is likely that the genetic patterns observed are associated with the geology/water chemistry of the waters in which these fish live. Multiple, interacting factors help determine the chemical composition of river water. Of particular importance is underlying geology, which has a strong influence on pH, conductivity and concentrations of dissolved ions (Jarvie et al., 2002; Liu et al., 2000; Rothwell et al., 2010). Brittany and southern Devon/Cornwall are dominated by Devonian age bedrock with granitic inclusions (e.g. the tors of Dartmoor), resulting in more acidic river water (pH ≤7) with low conductivity. Additionally, the upland areas of Brittany, Devon and Cornwall are dominated by blanket peat bog, reinforcing the acidic nature of river water in the area. Further east along both coasts in Normandy and south and southeast England the geology is dominated by Cretaceous era limestones and chalks, resulting in river water with pH values consistently above 7.
It has been suggested that the geological characteristics, and therefore, chemical characteristics, of river catchments may be an important factor in determining the accuracy of homing through olfactory‐based imprinting during smolting (Keefer & Caudill, 2014), which may help to maintain regional structuring via reduced straying between genetically distinct groups of rivers (Bourret et al., 2013). Additionally, underlying geology has been proposed to be a selective agent in the process of local adaptation in Atlantic salmon (Bourret et al., 2013). The hierarchical genetic structure detected here in English Channel trout also occurs in Atlantic salmon populations inhabiting rivers flowing into the Channel, with these patterns also having been linked to underlying geology (Ikediashi et al., 2018; Perrier et al., 2011). Moreover, the locations of transitions in genetic profiles between groups are coincident in both species, providing stronger evidence that underlying geology is playing a major role in driving local adaptation in trout living along these coasts.
4.2. Consequences of regional structure for assignment to the baseline
The greater success of assignments to regions of origin reflects the metapopulation structure found in many salmonid species that have anadromous life‐history stages (Schtickzelle & Quinn, 2007), with rivers in close proximity connected by gene flow via straying individuals from neighbouring rivers. Straying appears to be an integral part of salmonid life history. For instance, in a Danish fjord system, Källo, Baktoft, Birnie‐Gauvin, et al. (2022) and Källo, Baktoft, Kristensen, et al. (2022) found high levels of straying of anadromous trout across multiple life history stages. Brown trout populations show strong regional genetic structuring (Bekkevold et al., 2020; Koljonen et al., 2014; Prodöhl et al., 2017), especially for rivers in the Channel region (King et al., 2016, 2020; Quéméré et al., 2016); within regional groups, however, there tend to be low levels of differentiation between populations in neighbouring rivers. For reporting groups with the largest sea trout runs (OUTBRCH, DEVCORN, NEENG, LOWNORM and UPPNORM) mean pairwise F ST values were ≤0.04, indicative of little genetic differentiation between rivers within regions. Conversely, mean pairwise F ST values between reporting groups were generally >0.08, supporting the assertion that genetic assignment performs better when there are large genetic distances between baseline stocks (Araujo et al., 2014). Other salmonid fishery stock composition studies utilizing extensive genetic baselines have also found greater assignment success to regional groups of geographically proximate rivers rather than to individual rivers (Bekkevold et al., 2021; Griffiths et al., 2010; Harvey et al., 2019; King et al., 2016; Koljonen et al., 2014; Prodöhl et al., 2017). In some cases, reporting groups have incorporated rivers covering from several hundreds to thousands of kilometres of coastline (Gilbey et al., 2016, 2018; Jeffery et al., 2018; Wennevik et al., 2019).
To minimize biases in estimates of stock composition, a reasonably complete baseline is necessary to capture the genetic signal of the potentially important stocks that may be present in mixtures (Araujo et al., 2014). One advantage for assignment studies is that the metapopulation structure often found in salmonid species (Schtickzelle & Quinn, 2007) reduces the need to sample all rivers potentially contributing to marine catches. It is not always possible, either logistically or financially, to exhaustively sample all sea trout‐producing rivers in a region. Thus, a valid assumption of a regionally based assignment strategy is that samples originating from rivers not included in the baseline will likely be allocated to rivers from the same region, an approach that can reduce overall project costs (Beacham et al., 2020), albeit at the expense of a possible loss of finer resolution.
One of the potential limitations of genetic stock identification studies is the possible influence of unsampled ‘ghost’ reported regions, with the presence of fish derived from such regions likely to result in low individual assignment probabilities (Bradbury et al., 2015). Sixteen sea trout had assignment probabilities below 0.7 (Table S5), with the majority having low assignment to at least three reporting groups. These fish could possibly have originated from rivers in regions such as west Wales, southern Norway or southwest Sweden, which have been shown to be genetically distinct from some of the reporting groups identified here (Bekkevold et al., 2020, 2021; Prodöhl et al., 2017). Alternatively, these low assignment fish could have originated from rivers within our reporting groups. For instance, five of the low assignment sea trout (sampled from COR, TT and TAM) had assignments to only the OUTBRCH and DEVCORN reporting groups. It is clear that at the individual level some fish in these reporting groups are genetically very similar to each other (Figure 2) and in the LOO analysis the highest mis‐assignment of OUTBRCH fish was to the nearby DEVCORN group and vice versa.
4.3. Stock structure of marine and estuarine collections
In the current study, assignment results showed only very limited evidence of stock mixing of sea trout in the four estuarine collections. We can assume that these collections are the result of sampling local fish returning to their natal river prior to spawning. This was confirmed by the IA to river analyses (Tables S5.09–S5.12), which showed that the majority of fish caught in estuaries assigned to rivers flowing into the four estuaries. However, there were some fish that were clearly straying into these estuaries, with, for example, a NEENG fish caught in the recreational rod fishery in the Sussex Ouse (OUS), and three DEVCORN group fish caught in the net fishery in the Taw/Torridge (TT) estuary. Similarly, four of the marine‐caught collections (COR, SAA, CRI and MER) were predominantly sampling fish from local rivers. The main COR sampling sites were in Cawsand Bay, situated at the seaward (southwest) edge of the Tamar estuary, with four major sea trout rivers (LYN, TAM, TAV and PLY) flowing out through the estuary. While few of the fish could reliably be assigned to river of origin, the main river‐level assignments covered an ~80 km stretch of coast within the DEVCORN reporting group from the East and West Looe rivers (25 km to the west of the estuary) to the Dart (~55 km to the east of the estuary). Previous research has shown a degree of straying of sea trout from rivers along this stretch of coast into three of the Tamar estuary rivers (King et al., 2016). Likewise, the three samples of sea trout from the Upper Normandy coast (SAA, CRI, MER) also sampled predominately local fish. The nets in all three locations were recreational nets set from beaches during May to July when, again, fish would be returning to freshwater prior to spawning. Such targeting of local populations is not an uncommon feature of coastal fisheries targeting salmonids species. Fisheries for Atlantic salmon and Arctic charr on the Labrador coast of Canada (Bradbury et al., 2015, 2018; Layton et al., 2020) typically sampled fish from within ~150 km of the capture site. Similarly, net fisheries for sea trout in the Gulf of Finland have been shown to be catching fish predominantly from rivers proximal to the netting areas (Koljonen et al., 2014).
4.4. Southern North Sea collections are dominated by NE English sea trout
The two marine collections from the southern North Sea (EAN and DUT) were dominated by fish from rivers in northeast England i.e. the NEENG reporting group. The sea trout originating from rivers in this region are known to make long marine migrations, predominately migrating south along the east English North Sea coast. For instance, many sea trout tagged in the River Tweed have been caught in drift net fisheries along the East Anglian coast as well as in Dutch, German and Danish waters (Malcolm et al., 2010). This migration pattern has been confirmed using genetic assignment tests (Bekkevold et al., 2021). Thus, the southern North Sea appears to be important feeding grounds for multiple North Sea trout stocks (Bekkevold et al., 2021), with the results presented here providing evidence of sea trout originating from English Channel rivers (both English and French) also utilizing this area.
4.5. Eastwards movements of southern English sea trout
The results for the KIM, RYE, EAN and DUT collections highlight a tendency for some of the sea trout from Channel rivers to move in an easterly direction once entering the marine environment. DEVCORN origin‐fish were caught in Dorset at KIM and HANTS origin sea trout were present in the EAN collections and formed the majority of the fish sampled from RYE. Additionally, an UPPNORM fish was caught in the DUT net fishery. Previous historical tagging studies on sea trout smolts and kelts from the River Axe (DEVCORN reporting group) have shown that although on entering the marine environment the majority migrated west, some of the tag returns were from Hampshire Basin rivers to the east, coastal nets along the Dorset and Hampshire coasts and the southern North Sea (Potter et al., 2017; Solomon, 1994). These fish appeared to be following the dominant west to east current that flows along the northern (English) side of the Channel into the southern North Sea (Dauvin, 2019; Winther & Johannessen, 2006).
4.6. Long‐distance and cross‐channel movements
Some instances of very long‐distance movements of sea trout from rivers in the NEENG reporting group were observed, with sea trout from northeast England being sampled from COR (4 fish), KIM (6 fish), RYE (1 fish). Additionally, a single sea trout caught in the Sussex Ouse recreational rod fishery had a probability (p = 0.68) just below our 0.7 cut‐off of originating from a river in the NEENG reporting group (Table S5.12). Historic tagging studies undertaken on multiple life history stages of River Tweed sea trout have recorded only a single tag recovery from the English Channel (Malcolm et al., 2010). For the NEENG origin fish caught at Cawsand Bay, this represents a migration distance of ~1000 km (Table S6).
There were only two confirmed instances of cross‐Channel movements of sea trout with individuals sampled at KIM originating from the BRET and LOWNORM reporting groups. Such cross‐Channel movements do appear to be uncommon with only three tag recoveries from the northern French coast of sea trout tagged in southern English rivers (Potter et al., 2017). This finding is in contrast with the situation in the Irish Sea where frequent movements of trout from eastern Irish rivers into British coastal waters and vice versa were reported (Prodöhl et al., 2017).
4.7. Bycatch threats to sea trout during marine sojourns
In the marine environment sea trout exhibit a mainly piscivorous diet, with species such as sprat (Sprattus sprattus), sand eels (Ammodytes spp.) and herring (Clupea harengus) being dominant components of the diet (Knutsen et al., 2001; Poiesz et al., 2020; Roche et al., 2017). There are extensive commercial fisheries for two of these species (sprat and herring) in the southern North Sea and English Channel (Dauvin, 2019; Knijn et al., 1993) and it is likely that there is widespread bycatch of sea trout in these fisheries, although bycatch levels appear to be under‐recorded (Elliott et al., 2023). Additionally, it is likely that there will be bycatch in fisheries for fish species that have overlapping prey spectra with sea trout. For instance, our samples from RYE were caught in a net fishery that targets sea bass (Dicentrarchus labrax), which, like sea trout, are known to also feed on sprat and sand eel (Kelley, 1987; Spitz et al., 2013).
4.8. Management implications
The results presented here have implication for the management of sea trout in inshore waters around the English Channel and southern North Sea. Currently, for the UK, there is an extensive body of national and regional legislation designed to protect migratory salmonids from exploitation in inshore fisheries (Sumner, 2015); measures include protection from incidental capture in non‐target fisheries and total netting bans in estuarine areas. However, some of these measures lack consistency across different regions. For instance, net headline—the recommended depth below which nets should be set—varies between 1.5 and 3 m in different Inshore Fisheries & Conservation Authority regions along the southern English coast (Sumner, 2015).
Marine protected areas (MPAs) offer one route to safeguard sea trout during their marine migrations. Such areas offer protection within the designated region to both resident fish species and also species that transit through them (Breen et al., 2015). At present, however, evidence that MPAs are effective for the conservation of highly mobile species such as sea trout is limited (Breen et al., 2015). Nevertheless, to determine the efficacy of MPAs, to regulate fisheries and contribute to policy we require knowledge of where and when individuals are at sea (O'Sullivan et al., 2022). Genetic assignment studies, such as that presented here can help identify both fish movements and fisheries pressure on species, thereby providing evidence crucial to the designation and meaningful placement of MPAs (Jeffery et al., 2022).
Effective conservation of sea trout stocks in the marine environment therefore must include measures to minimize the risk of incidental capture. Based on inter‐river connectivity, as determined from population genetic data and prioritization analyses, a number of potential MPAs for English Channel sea trout have recently been proposed, (M. Vanhove, R. A. King, L. Meslier, A.‐L. Besnard, J. Stevens and S. Launey, unpublished data). Scenarios took into account factors, such as fishing density and other human effects on the marine environment, resulting in proposed protection areas along the south Devon and Cornish coasts, northern Brittany, Lower Normandy, the area between Dorset/Hampshire and the Cotentin Peninsula and the eastern Channel between Kent/Sussex and Upper Normandy (M. Vanhove, R. A. King, L. Meslier, A.‐L. Besnard, J. Stevens and S. Launey, unpublished data). Interestingly, two of these areas (Dorset/Hampshire and Kent/Sussex) are where we found the highest levels of stock mixing in our marine sea trout samples, strengthening the evidence that these areas should be designated as protected areas for sea trout in the English Channel.
FUNDING INFORMATION
This research was funded by the European Union Interreg France (Channel) England programme project ‘Salmonid Management Around the Channel’ (SAMARCH) with additional funding from the Missing Salmon Alliance. DRO was supported by a GW4 NERC Doctoral Training Programme PhD studentship as part of the FRESH programme.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflicts of interest.
Supporting information
Appendix S1
Figures S1–S5
Table S1
Table S2
Table S3
Table S4
Table S5
Table S6
ACKNOWLEDGEMENTS
We thank Simon Toms, Lawrence Talks, Willie Roche and the staff of the Environment Agency, Devon & Severn, Southern and Cornwall Inshore Fisheries Conservation Authorities, Inland Fisheries Ireland, INRAE U3E Unit, Office Français de la Biodiversité, Bretagne Grands Migrateurs, Seinormigr, FDPPMA14, 22, 27, 29, 35, 50, 62, 76 & 80 and André Breukelaar (Rijkswaterstaat) for co‐ordinating and supporting sample collection.
King, R. A. , Ellis, C. D. , Bekkevold, D. , Ensing, D. , Lecointre, T. , Osmond, D. R. , Piper, A. , Roberts, D. E. , Launey, S. , & Stevens, J. R. (2024). Leveraging the genetic diversity of trout in the rivers of the British Isles and northern France to understand the movements of sea trout (Salmo trutta L.) around the English Channel. Evolutionary Applications, 17, e13759. 10.1111/eva.13759
Contributor Information
R. Andrew King, Email: r.a.king@exeter.ac.uk.
Jamie R. Stevens, Email: j.r.stevens@exeter.ac.uk.
DATA AVAILABILITY STATEMENT
Data for this study are available at: https://doi.org/10.5061/dryad.1ns1rn92w.
REFERENCES
- Anderson, E. C. , & Dunham, K. K. (2008). The influence of family groups on inferences made with the program Structure. Molecular Ecology Resources, 8(6), 1219–1229. 10.1111/j.1755-0998.2008.02355.x [DOI] [PubMed] [Google Scholar]
- Araujo, H. A. , Candy, J. R. , Beacham, T. D. , White, B. , & Wallace, C. (2014). Advantages and challenges of genetic stock identification in fish stocks with low genetic resolution. Transactions of the American Fisheries Society, 143(2), 479–488. 10.1080/00028487.2013.855258 [DOI] [Google Scholar]
- Beacham, T. D. , Wallace, C. , Jonsen, K. , McIntosh, B. , Candy, J. R. , Rondeau, E. B. , Moore, J.‐S. , Bernatchez, L. , & Withler, R. E. (2020). Accurate estimation of conservation unit contribution to coho salmon mixed‐stock fisheries in British Columbia, Canada, using direct DNA sequencing for single nucleotide polymorphisms. Canadian Journal of Fisheries and Aquatic Sciences, 77(8), 1302–1315. 10.1139/cjfas-2019-0339 [DOI] [Google Scholar]
- Beacham, T. D. , Wallace, C. , Jonsen, K. , Sutherland, B. G. , Gummer, C. , & Rondeau, E. B. (2021). Estimation of conservation unit and population contribution to Chinook salmon mixed‐stock fisheries in British Columbia, Canada, using direct DNA sequencing for single nucleotide polymorphisms. Canadian Journal of Fisheries and Aquatic Sciences, 78(10), 1422–1434. 10.1139/cjfas-2020-0462 [DOI] [Google Scholar]
- Bekkevold, D. , Hojesjo, J. , Nielsen, E. E. , Aldven, D. , Als, T. D. , Sodeland, M. , Kent, M. P. , Lien, S. , & Hansen, M. M. (2020). Northern European Salmo trutta (L.) populations are genetically divergent across geographical regions and environmental gradients. Evolutionary Applications, 13(2), 400–416. 10.1111/eva.12877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bekkevold, D. , Piper, A. , Campbell, R. , Rippon, P. , Wright, R. M. , Crundwell, C. , Wysujack, K. , Stevens, J. R. , King, R. A. , Aarestrup, K. , & Maltby, A. (2021). Genetic stock identification of sea trout (Salmo trutta L.) along the British North Sea Coast shows prevalent long‐distance migration. ICES Journal of Marine Science, 78(3), 952–966. 10.1093/icesjms/fsaa240 [DOI] [Google Scholar]
- Benjamini, Y. , & Hochberg, Y. (1995). Controlling the false discovery rate – A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B: Statistical Methodology, 57(1), 289–300. 10.1111/j.2517-6161.1995.tb02031.x [DOI] [Google Scholar]
- Bernatchez, L. (2001). The evolutionary history of brown trout (Salmo trutta L.) inferred from phylogeographic, nested clade, and mismatch analyses of mitochondrial DNA variation. Evolution, 55(2), 351–379. 10.1111/j.0014-3820.2001.tb01300.x [DOI] [PubMed] [Google Scholar]
- Bourret, V. , Dionne, M. , Kent, M. P. , Lien, S. , & Bernatchez, L. (2013). Landscape genomics in Atlantic salmon (Salmo salar): Searching for gene‐environment interactions driving local adaptation. Evolution, 67(12), 3469–3487. 10.1111/evo.12139 [DOI] [PubMed] [Google Scholar]
- Bouza, C. , Arias, J. , Castro, J. , Sánchez, L. , & Martínez, P. (1999). Genetic structure of brown trout, Salmo trutta L., at the southern limit of the distribution range of the anadromous form. Molecular Ecology, 8(12), 1991–2001. 10.1046/j.1365-294x.1999.00794.x [DOI] [PubMed] [Google Scholar]
- Bradbury, I. R. , Hamilton, L. C. , Rafferty, S. , Meerburg, D. , Poole, R. , Dempson, J. B. , Robertson, M. J. , Reddin, D. G. , Bourret, V. , Dionne, M. , Chaput, G. , Sheehan, T. F. , King, T. L. , Candy, J. R. , & Bernatchez, L. (2015). Genetic evidence of local exploitation of Atlantic salmon in a coastal subsistence fishery in the Northwest Atlantic. Canadian Journal of Fisheries and Aquatic Sciences, 72(1), 83–95. 10.1139/cjfas-2014-0058 [DOI] [Google Scholar]
- Bradbury, I. R. , Wringe, B. F. , Watson, B. , Paterson, I. , Horne, J. , Beiko, R. , Lehnert, S. J. , Clément, M. , Anderson, E. C. , Jeffery, N. W. , Duffy, S. , Sylvester, E. , Robertson, M. , & Bentzen, P. (2018). Genotyping‐by‐sequencing of genome‐wide microsatellite loci reveals fine‐scale harvest composition in a coastal Atlantic salmon fishery. Evolutionary Applications, 11(6), 918–930. 10.1111/eva.12606 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breen, P. , Posen, P. , & Righton, D. (2015). Temperate marine protected areas and highly mobile fish: A review. Ocean and Coastal Management, 105, 75–83. 10.1016/j.ocecoaman.2014.12.021 [DOI] [Google Scholar]
- Cavalli‐Sforza, L. L. , & Edwards, A. W. F. (1967). Phylogenetic analysis models and estimation procedures. American Journal of Human Genetics, 19(3P1), 233–257. [PMC free article] [PubMed] [Google Scholar]
- Charles, K. , Guyomard, R. , Hoyheim, B. , Ombredane, D. , & Baglinière, J. L. (2005). Lack of genetic differentiation between anadromous and resident sympatric brown trout (Salmo trutta) in a Normandy population. Aquatic Living Resources, 18(1), 65–69. 10.1051/alr:2005006 [DOI] [Google Scholar]
- Clemento, A. J. , Abadia‐Cardoso, A. , Starks, H. A. , & Garza, J. C. (2011). Discovery and characterization of single nucleotide polymorphisms in Chinook salmon, Oncorhynchus tshawytscha . Molecular Ecology Resources, 11, 50–66. 10.1111/j.1755-0998.2010.02972.x [DOI] [PubMed] [Google Scholar]
- Cormack, R. M. , & Skalski, J. R. (1992). Analysis of coded wire tag returns from commercial catches. Canadian Journal of Fisheries and Aquatic Sciences, 49(9), 1816–1825. 10.1139/f92-201 [DOI] [Google Scholar]
- Cortey, M. , Vera, M. , Pla, C. , & García‐Marín, J. L. (2009). Northern and southern expansions of Atlantic brown trout (Salmo trutta) populations during the Pleistocene. Biological Journal of the Linnean Society, 97(4), 904–917. 10.1111/j.1095-8312.2009.01220.x [DOI] [Google Scholar]
- Dauvin, J.‐C. (2012). Are the eastern and western basins of the English Channel two separate ecosystems? Marine Pollution Bulletin, 64(3), 463–471. 10.1016/j.marpolbul.2011.12.010 [DOI] [PubMed] [Google Scholar]
- Dauvin, J.‐C. (2019). The English Channel: La Manche. In Sheppard C. (Ed.), World Sea: An environmental evaluation (2nd ed., pp. 153–188). Academic Press. [Google Scholar]
- Elliott, S. A. M. , Acou, A. , Beaulaton, L. , Guitton, J. , Reveillac, E. , & Rivot, E. (2023). Modelling the distribution of rare and data‐poor diadromous fish at sea for protected area management. Progress in Oceanography, 210, 102924. 10.1016/j.pocean.2022.102924 [DOI] [Google Scholar]
- Evanno, G. , Regnaut, S. , & Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Molecular Ecology, 14(8), 2611–2620. 10.1111/j.1365-294X.2005.02553.x [DOI] [PubMed] [Google Scholar]
- Ferguson, A. (1989). Genetic differences among brown trout, Salmo trutta, stocks and their importance for the conservation and management of the species. Freshwater Biology, 21(1), 35–46. 10.1111/j.1365-2427.1989.tb01346.x [DOI] [Google Scholar]
- Ferguson, A. , & Prodöhl, P. A. (2022). Identifying and conserving sympatric diversity in trout of the genus Salmo, with particular reference to Lough Melvin, Ireland. Ecology of Freshwater Fish, 31(2), 177–207. 10.1111/eff.12651 [DOI] [Google Scholar]
- Ferguson, A. , Reed, T. E. , Cross, T. F. , McGinnity, P. , & Prodöhl, P. A. (2019). Anadromy, potamodromy and residency in brown trout Salmo trutta: the role of genes and the environment. Journal of Fish Biology, 95, 692–718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferguson, A. , & Taggart, J. B. (1991). Genetic differentiation among the sympatric brown trout (Salmo trutta) populations of Lough Melvin, Ireland. Biological Journal of the Linnean Society, 43(3), 221–237. 10.1111/j.1095-8312.1991.tb00595.x [DOI] [Google Scholar]
- Francis, R. M. (2017). POPHELPER: An R package and web app to analyse and visualize population structure. Molecular Ecology Resources, 17(1), 27–32. 10.1111/1755-0998.12509 [DOI] [PubMed] [Google Scholar]
- Gilbey, J. , Cauwelier, E. , Coulson, M. W. , Stradmeyer, L. , Sampayo, J. N. , Armstrong, A. , Verspoor, E. , Corrigan, L. , Shelley, J. , & Middlemas, S. (2016). Accuracy of assignment of Atlantic Salmon (Salmo salar L.) to rivers and regions in Scotland and northeast England based on single nucleotide polymorphism (SNP) markers. PLoS One, 11(10), e0164327. 10.1371/journal.pone.0164327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbey, J. , Coughlan, J. , Wennevik, V. , Prodöhl, P. , Stevens, J. R. , de Leaniz, C. G. , Ensing, D. , Cauwelier, E. , Cherbonnel, C. , Consuegra, S. , Coulson, M. W. , Cross, T. F. , Crozier, W. , Dillane, E. , Ellis, J. S. , García‐Vázquez, E. , Griffiths, A. M. , Gudjonsson, S. , Hindar, K. , … Verspoor, E. (2018). A microsatellite baseline for genetic stock identification of European Atlantic salmon (Salmo salar L.). ICES Journal of Marine Science, 75(2), 662–674. 10.1093/icesjms/fsx184 [DOI] [Google Scholar]
- Gilbey, J. , Utne, K. R. , Wennevik, V. , Beck, A. C. , Kausrud, K. , Hindar, K. , de Leaniz, C. G. , Cherbonnel, C. , Coughlan, J. , Cross, T. F. , Dillane, E. , Ensing, D. , García‐Vázquez, E. , Hole, L. R. , Holm, M. , Holst, J. C. , Jacobsen, J. A. , Jensen, A. J. , Karlsson, S. , … Verspoor, E. (2021). The early marine distribution of Atlantic salmon in the north‐east Atlantic: A genetically informed stock‐specific synthesis. Fish and Fisheries, 22(6), 1274–1306. 10.1111/faf.12587 [DOI] [Google Scholar]
- Gilbey, J. , Wennevik, V. , Bradbury, I. R. , Fiske, P. , Hansen, L. P. , Jacobsen, J. A. , & Potter, T. (2017). Genetic stock identification of Atlantic salmon caught in the Faroese fishery. Fisheries Research, 187, 110–119. 10.1016/j.fishres.2016.11.020 [DOI] [Google Scholar]
- Gillson, J. P. , Bašić, T. , Davison, P. , Riley, W. D. , Talks, L. , Walker, A. M. , & Russell, I. C. (2022). A review of marine stressors impacting Atlantic salmon Salmo salar, with an assessment of the major threats to English stocks. Reviews in Fish Biology and Fisheries, 32(3), 879–919. 10.1007/s11160-022-09714-x [DOI] [Google Scholar]
- Glegg, G. , Jefferson, R. , & Fletcher, S. (2015). Marine governance in the English Channel (La Manche): Linking science and management. Marine Pollution Bulletin, 95(2), 707–718. 10.1016/j.marpolbul.2015.02.020 [DOI] [PubMed] [Google Scholar]
- Goodwin, J. C. A. , King, R. A. , Jones, J. I. , Ibbotson, A. , & Stevens, J. R. (2016). A small number of anadromous females drive reproduction in a brown trout (Salmo trutta) population in an English chalk stream. Freshwater Biology, 61(7), 1075–1089. 10.1111/fwb.12768 [DOI] [Google Scholar]
- Griffiths, A. M. , Koizumi, I. , Bright, D. , & Stevens, J. R. (2009). A case of isolation by distance and short‐term temporal stability of population structure in brown trout (Salmo trutta) within the River Dart, southwest England. Evolutionary Applications, 2(4), 537–554. 10.1111/j.1752-4571.2009.00092.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths, A. M. , Machado‐Schiaffino, G. , Dillane, E. , Coughlan, J. , Horreo, J. L. , Bowkett, A. E. , Minting, P. , Toms, S. , Roche, W. , Gargan, P. , McGinnity, P. , Cross, T. , Bright, D. , Garcia‐Vazquez, E. , & Stevens, J. R. (2010). Genetic stock identification of Atlantic salmon (Salmo salar) populations in the southern part of the European range. BMC Genetics, 11, 27. 10.1186/1471-2156-11-31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harvey, A. C. , Quintela, M. , Glover, K. A. , Karlsen, O. , Nilsen, R. , Skaala, O. , Sægrov, H. , Kålås, S. , Knutar, S. , & Wennevik, V. (2019). Inferring Atlantic salmon post‐smolt migration patterns using genetic assignment. Royal Society Open Science, 6(10), 10. 10.1098/rsos.190426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawley, K. L. , Urke, H. A. , Kristensen, T. , & Haugen, T. O. (2024). Balancing risks and rewards of alternate strategies in the seaward extent, duration and timing of fjord use in contemporary anadromy of brown trout (Salmo trutta). BMC Ecology and Evolution, 24, 27. 10.1186/s12862-023-02179-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- ICES . (2013). Report of the workshop on sea trout. ICES CM 2013/SSGEF: 15. International Council for the Exploration of the Sea. [Google Scholar]
- Ikediashi, C. , Paris, J. R. , King, R. A. , Beaumont, W. R. C. , Ibbotson, A. , & Stevens, J. R. (2018). Atlantic salmon Salmo salar in the chalk streams of England are genetically unique. Journal of Fish Biology, 92(3), 621–641. 10.1111/jfb.13538 [DOI] [PubMed] [Google Scholar]
- Jakobsson, M. , & Rosenberg, N. A. (2007). CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics, 23(14), 1801–1806. 10.1093/bioinformatics/btm233 [DOI] [PubMed] [Google Scholar]
- Jarvie, H. P. , Oguchi, T. , & Neal, C. (2002). Exploring the linkages between river water chemistry and watershed characteristics using GIS‐based catchment and locality analyses. Regional Environmental Change, 3(1–3), 36–50. 10.1007/s10113-001-0036-6 [DOI] [Google Scholar]
- Jeffery, N. W. , Lehnert, S. J. , Kess, T. , Layton, K. K. S. , Wringe, B. F. , & Stanley, R. R. E. (2022). Application of omics tools in designing and monitoring marine protected areas for a sustainable blue economy. Frontiers in Genetics, 13, 886494. 10.3389/fgene.2022.886494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeffery, N. W. , Wringe, B. F. , McBride, M. C. , Hamilton, L. C. , Stanley, R. R. E. , Bernatchez, L. , Kent, M. , Clément, M. , Gilbey, J. , Sheehan, T. F. , Bentzen, P. , & Bradbury, I. R. (2018). Range‐wide regional assignment of Atlantic salmon (Salmo salar) using genome wide single‐nucleotide polymorphisms. Fisheries Research, 206, 163–175. 10.1016/j.fishres.2018.05.017 [DOI] [Google Scholar]
- Jones, O. R. , & Wang, J. L. (2010). COLONY: A program for parentage and sibship inference from multilocus genotype data. Molecular Ecology Resources, 10(3), 551–555. 10.1111/j.1755-0998.2009.02787.x [DOI] [PubMed] [Google Scholar]
- Jonsson, B. , & Jonsson, N. (2009). A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta, with particular reference to water temperature and flow. Journal of Fish Biology, 75(10), 2381–2447. 10.1111/j.1095-8649.2009.02380.x [DOI] [PubMed] [Google Scholar]
- Jonsson, B. , & Jonsson, N. (2014). Naturally and hatchery produced European trout Salmo trutta: Do their marine survival and dispersal differ? Journal of Coastal Conservation, 18(2), 79–87. 10.1007/s11852-012-0224-1 [DOI] [Google Scholar]
- Kallio‐Nyberg, I. , Saura, A. , & Ahlfors, P. (2002). Sea migration pattern of two sea trout (Salmo trutta) stocks released into the Gulf of Finland. Annales Zoologici Fennici, 39(3), 221–235. [Google Scholar]
- Källo, K. , Baktoft, H. , Birnie‐Gauvin, K. , & Aarestrup, K. (2022). Variability in straying behaviour among repeat spawning anadromous brown trout (Salmo trutta) followed over several years. ICES Journal of Marine Science, 79(9), 2453–2460. 10.1093/icesjms/fsac183 [DOI] [Google Scholar]
- Källo, K. , Baktoft, H. , Kristensen, M. L. , Birnie‐Gauvin, K. , & Aarestrup, K. (2022). High prevalence of straying in a wild brown trout (Salmo trutta) population in a fjord system. ICES Journal of Marine Science, 79(5), 1539–1547. 10.1093/icesjms/fsac079 [DOI] [Google Scholar]
- Keefer, M. L. , & Caudill, C. C. (2014). Homing and straying by anadromous salmonids: a review of mechanisms and rates. Reviews in Fish Biology and Fisheries, 24(1), 333–368. 10.1007/s11160-013-9334-6 [DOI] [Google Scholar]
- Kelley, D. F. (1987). Food of sea bass in UK waters. Journal of the Marine Biological Association of the United Kingdom, 67, 275–286. [Google Scholar]
- Kershner, J. L. , Williams, J. E. , Gresswell, R. E. , & Lobón‐Cerviá, J. (2019). Trout and char of the world. American Fisheries Society. [Google Scholar]
- King, R. A. , Hillman, R. , Elsmere, P. , Stockley, B. , & Stevens, J. R. (2016). Investigating patterns of straying and mixed stock exploitation of sea trout, Salmo trutta in rivers sharing an estuary in south‐west England. Fisheries Management and Ecology, 23(5), 376–389. 10.1111/fme.12181 [DOI] [Google Scholar]
- King, R. A. , Stockley, B. , & Stevens, J. R. (2020). Small coastal streams – Critical reservoirs of genetic diversity for trout (Salmo trutta L.) in the face of increasing anthropogenic stressors. Ecology and Evolution, 10(12), 5651–5669. 10.1002/ece3.6306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knijn, R. J. , Boon, T. W. , Heessen, H. J. L. , & Hislop, J. R. G. (1993). Atlas of North Sea fishes (ICES Cooperative Research Report No. 194). Retrieved from Copenhagen, Denmark.
- Knutsen, J. A. , Knutsen, H. , Gjosaeter, J. , & Jonsson, B. (2001). Food of anadromous brown trout at sea. Journal of Fish Biology, 59(3), 533–543. 10.1006/jfbi.2001.1662 [DOI] [Google Scholar]
- Koljonen, M.‐L. , Gross, R. , & Koskiniemi, J. (2014). Wild Estonian and Russian sea trout (Salmo trutta L.) in Finnish coastal sea trout catches: Results of genetic mixed‐stock analysis. Hereditas, 151, 177–195. [DOI] [PubMed] [Google Scholar]
- Langella, O. (1999). Populations v1.2.28 . http://bioinformatics.org/populations/
- Layton, K. K. S. , Dempson, B. , Snelgrove, P. V. R. , Duffy, S. J. , Messmer, A. M. , Paterson, I. G. , Jeffery, N. W. , Kess, T. , Horne, J. B. , Salisbury, S. J. , Ruzzante, D. E. , Bentzen, P. , Côté, D. , Nugent, C. M. , Ferguson, M. M. , Leong, J. S. , Koop, B. F. , & Bradbury, I. R. (2020). Resolving fine‐scale population structure and fishery exploitation using sequenced microsatellites in a northern fish. Evolutionary Applications, 13(5), 1055–1068. 10.1111/eva.12922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Cren, E. D. (1985). The biology of sea trout. Atlantic Salmon Trust. [Google Scholar]
- Lemopoulos, A. , Uusi‐Heikkilä, S. , Huusko, A. , Vasemägi, A. , & Vainikka, A. (2018). Comparison of migratory and resident populations of brown trout reveals candidate genes for migration tendency. Genome Biology and Evolution, 10(6), 1493–1503. 10.1093/gbe/evy102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lericolais, G. , Auffret, J. P. , & Bourillet, J. F. (2003). The Quaternary Channel River: Seismic stratigraphy of its palaeo‐valleys and deeps. Journal of Quaternary Science, 18(3–4), 245–260. 10.1002/jqs.759 [DOI] [Google Scholar]
- Liu, Z. J. , Weller, D. E. , Correll, D. L. , & Jordan, T. E. (2000). Effects of land cover and geology on stream chemistry in watersheds of Chesapeake Bay. Journal of the American Water Resources Association, 36(6), 1349–1365. 10.1111/j.1752-1688.2000.tb05731.x [DOI] [Google Scholar]
- Losee, J. P. , Palm, D. , Claiborne, A. , Madel, G. , Persson, L. , Quinn, T. P. , Brodin, T. , & Hellström, G. (2024). Anadromous trout from opposite sides of the globe: biology, ocean ecology, and management of anadromous brown and cutthroat trout. Reviews in Fish Biology and Fisheries, 34(1), 461–490. 10.1007/s11160-023-09824-0 [DOI] [Google Scholar]
- Malcolm, I. A. , Godfrey, J. , & Youngson, A. F. (2010). Review of migratory routes and behaviour of Atlantic salmon, sea trout and European eel in Scotland's coastal environment: Implications for the development of marine renewables . https://data.marine.gov.scot/dataset/review‐migratory‐routes‐and‐behaviour‐atlantic‐salmon‐sea‐trout‐and‐european‐eel‐scotland's
- McKeown, N. J. , Hynes, R. A. , Duguid, R. A. , Ferguson, A. , & Prodöhl, P. A. (2010). Phylogeographic structure of brown trout Salmo trutta in Britain and Ireland: glacial refugia, postglacial colonization and origins of sympatric populations. Journal of Fish Biology, 76(2), 319–347. 10.1111/j.1095-8649.2009.02490.x [DOI] [PubMed] [Google Scholar]
- Meirmans, P. G. (2020). GENODIVE version 3.0: Easy‐to‐use software for the analysis of genetic data of diploids and polyploids. Molecular Ecology Resources, 20(4), 1126–1131. 10.1111/1755-0998.13145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menyhart, O. , Weltz, B. , & Gyorffy, B. (2021). MultipleTesting.com: A tool for life science researchers for multiple hypothesis testing correction. PLoS One, 16(6), e0245824. 10.1371/journal.pone.0245824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran, B. M. , & Anderson, E. C. (2019). Bayesian inference from the conditional genetic stock identification model. Canadian Journal of Fisheries and Aquatic Sciences, 76(4), 551–560. 10.1139/cjfas-2018-0016 [DOI] [Google Scholar]
- Neaves, P. I. , Wallace, C. G. , Candy, J. R. , & Beacham, T. D. (2005). cBayes: Computer program for mixed‐stock analysis of allelic data, version v5.01 . http://www.pac.dfo‐mpo.gc.ca/sci/mgl/Cbayes_e.htm
- Nei, N. (1987). Molecular evolutionary genetics. Columbia University Press. [Google Scholar]
- Nevoux, M. , Finstad, B. , Davidsen, J. G. , Finaly, R. , Josset, Q. , Poole, R. , Höjesjö, J. , Aarestrup, K. , Persson, L. , Tolvanen, O. , & Jonsson, B. (2019). Environmental influences on life history strategies in partially anadromous brown trout (Salmo trutta, Salmonidae). Fish and Fisheries, 20, 1051–1082. [Google Scholar]
- Osmond, D. R. , King, R. A. , Russo, I.‐R. , Bruford, M. W. , & Stevens, J. R. (2024). Living in a post‐industrial landscape: repeated patterns of genetic divergence in brown trout (Salmo trutta L.) across the British Isles. Diversity and Distributions, 30, e13854. 10.1111/ddi.13854 [DOI] [Google Scholar]
- Osmond, D. R. , King, R. A. , Stockley, B. , Launey, S. , & Stevens, J. R. (2023). A low‐density single nucleotide polymorphism panel for brown trout (Salmo trutta L.) suitable for exploring genetic diversity at a range of spatial scales. Journal of Fish Biology, 102(1), 258–270. 10.1111/jfb.15258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Östergren, J. , Palm, S. , Gilbey, J. , & Dannewitz, J. (2020). Close relatives in population samples: Evaluation of the consequences for genetic stock identification. Molecular Ecology Resources, 20(2), 498–510. 10.1111/1755-0998.13131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Sullivan, R. J. , Ozerov, M. , Bolstad, G. H. , Gilbey, J. , Jacobsen, J. A. , Erkinaro, J. , Rikardsen, A. H. , Hindar, K. , & Aykanat, T. (2022). Genetic stock identification reveals greater use of an oceanic feeding ground around the Faroe Islands by multi‐sea winter Atlantic salmon, with variation in use across reporting groups. ICES Journal of Marine Science, 79(9), 2442–2452. 10.1093/icesjms/fsac182 [DOI] [Google Scholar]
- Pante, E. , & Simon‐Bouhet, B. (2013). marmap: A package for importing, plotting and analyzing bathymetric and topographic data in R. PLoS One, 8(9), e73051. 10.1371/journal.pone.0073051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paris, J. R. , King, R. A. , & Stevens, J. R. (2015). Human mining activity across the ages determines the genetic structure of modern brown trout (Salmo trutta L.) populations. Evolutionary Applications, 8(6), 573–585. 10.1111/eva.12266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pella, J. , & Masuda, M. (2001). Bayesian methods for analysis of stock mixtures from genetic characters. Fishery Bulletin, 99(1), 151–167. [Google Scholar]
- Perrier, C. , Guyomard, R. , Bagliniere, J. L. , & Evanno, G. (2011). Determinants of hierarchical genetic structure in Atlantic salmon populations: environmental factors vs anthropogenic influences. Molecular Ecology, 20(20), 4231–4245. 10.1111/j.1365-294X.2011.05266.x [DOI] [PubMed] [Google Scholar]
- Poiesz, S. S. H. , Witte, J. I. J. , & van der Veer, H. W. (2020). Only a few key prey species fuel a temperate coastal fish food web. Marine Ecology Progress Series, 653, 153–166. 10.3354/meps13472 [DOI] [Google Scholar]
- Potter, E. , Campbell, R. , Sumner, K. , & Marshall, S. (2017). Marine migrations and distribution of sea trout from rivers in Great Britain. In Harris G. (Ed.), Sea trout: Science and Management (pp. 205–227). Matador. [Google Scholar]
- Pritchard, J. K. , Stephens, M. , & Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics, 155(2), 945–959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prodöhl, P. A. , Antoniacomi, A. , Bradley, C. , Carlsson, J. , Carvalho, G. R. , Coughlan, J. , Coyne, J. , Cross, M. C. , Cross, M. E. , Davies, C. A. , Dillane, E. , Gargan, P. , Hynes, R. , McGinnity, P. , Milner, N. , Reed, T. , Roche, W. , Taylor, M. , Tysklind, & Cross, T. F. (2017). Population genetics and genetic stock identification of anadromous Salmo trutta from the Irish Sea and adjacent areas, using microsatellite DNA loci. In Harris G. (Ed.), Sea trout: Science and Management (pp. 69–95). Matador. [Google Scholar]
- Quéméré, E. , Bagliniere, J. L. , Roussel, J. M. , Evanno, G. , McGinnity, P. , & Launey, S. (2016). Seascape and its effect on migratory life‐history strategy influences gene flow among coastal brown trout (Salmo trutta) populations in the English Channel. Journal of Biogeography, 43(3), 498–509. 10.1111/jbi.12632 [DOI] [Google Scholar]
- Raymond, M. , & Rousset, F. (1995). GENEPOP (version‐1.2) – Population genetics software for exact tests and ecumenicism. Journal of Heredity, 86(3), 248–249. 10.1093/oxfordjournals.jhered.a111573 [DOI] [Google Scholar]
- Roche, W. , Milner, N. , Davies, C. , Shephard, S. , King, J. , Coyne, J. , Gargan, P. , & Hughes, R. (2017). Feeding ecology of sea trout in the Irish Sea. In Harris G. (Ed.), Sea trout: Science and Management (pp. 371–395). Matador. [Google Scholar]
- Rothwell, J. J. , Dise, N. B. , Taylor, K. G. , Allott, T. E. H. , Scholefield, P. , Davies, H. , & Neal, C. (2010). A spatial and seasonal assessment of river water chemistry across North West England. Science of the Total Environment, 408(4), 841–855. 10.1016/j.scitotenv.2009.10.041 [DOI] [PubMed] [Google Scholar]
- Schtickzelle, N. , & Quinn, T. P. (2007). A metapopulation perspective for salmon and other anadromous fish. Fish and Fisheries, 8(4), 297–314. 10.1111/j.1467-2979.2007.00256.x [DOI] [Google Scholar]
- Small, M. P. , Olive, S. D. R. , Seeb, L. W. , Seeb, J. E. , Pascal, C. E. , Warheit, K. I. , & Templin, W. (2015). Chum salmon genetic diversity in the northeastern Pacific Ocean assessed with single nucleotide polymorphisms (SNPs): applications to fishery management. North American Journal of Fisheries Management, 35(5), 974–987. 10.1080/02755947.2015.1055014 [DOI] [Google Scholar]
- Solomon, D. J. (1994). Sea trout investigations – Phase 1 final report . R&D Note 318. National Rivers Authority.
- Spitz, J. , Chouvelon, T. , Cardinaud, M. , Kostecki, C. , & Lorance, P. (2013). Prey preferences of adult sea bass Dicentrarchus labrax in the northeastern Atlantic: implications for bycatch of common dolphin Delphinus delphis . ICES Journal of Marine Science, 70(2), 452–461. 10.1093/icesjms/fss200 [DOI] [Google Scholar]
- Sumner, K. (2015). Review of protection measures for Atlantic salmon and sea trout in inshore waters . Environment Agency, UK.
- Tamura, K. , Stecher, G. , Peterson, D. , Filipski, A. , & Kumar, S. (2013). MEGA6: Molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution, 30(12), 2725–2729. 10.1093/molbev/mst197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thériault, V. , Bernatchez, L. , & Dodson, J. J. (2007). Mating system and individual reproductive success of sympatric anadromous and resident brook charr, Salvelinus fontinalis, under natural conditions. Behavioral Ecology and Sociobiology, 62(1), 51–65. 10.1007/s00265-007-0437-8 [DOI] [Google Scholar]
- Thorstad, E. B. , Todd, C. D. , Uglem, I. , Bjorn, P. A. , Gargan, P. G. , Vollset, K. W. , Halttunen, E. , Kålås, S. , Berg, M. , & Finstad, B. (2016). Marine life of the sea trout. Marine Biology, 163(3), 19. 10.1007/s00227-016-2820-3 [DOI] [Google Scholar]
- Truett, G. E. , Heeger, P. , Mynatt, R. L. , Truett, A. A. , Walker, J. A. , & Warman, M. L. (2000). Preparation of PCR‐quality mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). BioTechniques, 29(1), 52. 10.2144/00291bm09 [DOI] [PubMed] [Google Scholar]
- Tucker, S. , Trudel, M. , Welch, D. W. , Candy, J. R. , Morris, J. F. T. , Thiess, M. E. , Wallace, C. , Teel, D. J. , Crawford, W. , Farley, E. V., Jr. , & Beacham, T. D. (2009). Seasonal stock‐specific migrations of juvenile sockeye salmon along the West Coast of North America: Implications for growth. Transactions of the American Fisheries Society, 138(6), 1458–1480. 10.1577/t08-211.1 [DOI] [Google Scholar]
- Verspoor, E. , Coulson, M. W. , Greer, R. B. , & Knox, D. (2019). Unique sympatric quartet of limnetic, benthic, profundal and piscivorous brown trout populations resolved by 3D sampling and focused molecular marker selection. Freshwater Biology, 64(1), 121–137. 10.1111/fwb.13199 [DOI] [Google Scholar]
- Vilas, R. , Bouza, C. , Castro, J. , López, A. , & Martínez, P. (2010). Management units of brown trout from Galicia (NW: Spain) based on spatial genetic structure analysis. Conservation Genetics, 11(3), 897–906. 10.1007/s10592-009-9934-9 [DOI] [Google Scholar]
- Waples, R. S. , & Anderson, E. C. (2017). Purging putative siblings from population genetic data sets: A cautionary view. Molecular Ecology, 26(5), 1211–1224. 10.1111/mec.14022 [DOI] [PubMed] [Google Scholar]
- Weir, B. S. , & Cockerham, C. C. (1984). Estimating F‐statistics for the analysis of population structure. Evolution, 38(6), 1358–1370. 10.2307/2408641 [DOI] [PubMed] [Google Scholar]
- Wennevik, V. , Quintela, M. , Skaala, O. , Verspoor, E. , Prusov, S. , & Glover, K. A. (2019). Population genetic analysis reveals a geographically limited transition zone between two genetically distinct Atlantic salmon lineages in Norway. Ecology and Evolution, 9(12), 6901–6921. 10.1002/ece3.5258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whelan, K. , Harris, G. , Milner, N. , Potter, E. , Llewelyn, I. , & Callagher, C. (2017). Looking ahead: Management priorities for sea trout. In Harris G. (Ed.), Sea trout: Science and Management (pp. 555–585). Matador. [Google Scholar]
- Winther, N. G. , & Johannessen, J. A. (2006). North Sea circulation: Atlantic inflow and its destination. Journal of Geophysical Research‐Oceans, 111(C12), 12. 10.1029/2005jc003310 [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1
Figures S1–S5
Table S1
Table S2
Table S3
Table S4
Table S5
Table S6
Data Availability Statement
Data for this study are available at: https://doi.org/10.5061/dryad.1ns1rn92w.